Submitted:
28 January 2026
Posted:
29 January 2026
You are already at the latest version
Abstract
High-resolution action spectra of photosynthetic Emerson enhancement were determined using a unique, custom-built three-beam oxygen electrode system in the Joliot configuration, featuring a bare platinum cathode. This specialized apparatus utilized three independent light beams—including one modulated at 10 Hz to precisely quantify real-time changes in the quantum yield of Photosystem II resulting from the simultaneous excitation of Photosystem I. This methodology provides a direct and physiologically relevant analysis of radiant energy distribution between the two photosystems in intact algal cells, achieving a level of precision that remains a benchmark in the field. Comprehensive spectra were obtained for key representatives of five major algal divisions: Rhodophyta, Chlorophyta, Dinophyta, Cryptophyta, and Cyanobacteria. The results demonstrate that enhancement spectra vary systematically according to the excitation balance between the photosystems. Furthermore, photoadaptation studies in Glenodinium sp. and Rhodomonas D3 revealed taxon-specific responses in energy distribution under high irradiance. By integrating these high-resolution measurements with contemporary photosynthetic theory, this work clarifies fundamental biophysical mechanisms of spectral photoadaptation and provides critical insights into the ecological zonation of algal species in coastal aquatic environments.
Keywords:
photosynthesis action spectra
; emerson enhancement effect
; photosystem II quantum yield
; oxygen electrode (Joliot configuration)
; spectral photoadaptation
; algal bio-energetics
1. Introduction
Oxygenic photosynthesis relies on the cooperation of two functionally distinct reaction centers, Photosystem I (PSI) and Photosystem II (PSII), which operate in series to convert light energy into chemical energy [1]. In a classic discovery, Emerson [2] showed that in algae, such as Porphyridium and Chlorella, illumination with a single wavelength produced a markedly lower photosynthetic rate than simultaneous illumination with two wavelengths, revealing a synergistic effect between the two photosystems, later termed the Emerson enhancement effect.
The distribution of absorbed energy between PSI and PSII depends on both the spectral composition of light and the photo adaptive state of the algal cells [3]. However, direct measurement of this distribution remains technically challenging, particularly in intact organisms under physiological conditions. Conventional methods based on chlorophyll fluorescence or low-temperature spectroscopy often lack the temporal and spectral resolution necessary to disentangle PSI and PSII contributions in vivo.
Photoadaptation in algae involves structural and functional adjustments that optimize photosynthetic efficiency across varying irradiances. These include changes in photosystem size and number, the PSI:PSII stoichiometry, and the spectral distribution of excitation energy [4,5]. Under low light, algal cells typically enhance the transfer of green radiation to PSII relative to blue radiation, a pattern reflected in the excitation spectra of chlorophyll a fluorescence [6,7]. These fluorescence spectra closely correlate with the action spectra of PSII oxygen evolution [8,9], providing indirect insight into photosystem-specific energy transfer.
Spectral photoadaptation has been documented in natural phytoplankton assemblages [10,11,12,13]. In some species, such as the cryptomonad Rhodomonas salina, low-light acclimation increases blue-light utilization [14]), suggesting coupled spectral responses of PSI and PSII [15]. Many algae grown under high irradiance show higher PSI/PSII ratios, larger PSI absorption cross-sections, and higher rates of cyclic photophosphorylation than low-light cultures [16,17,18,19,20,21,22]. In addition to algae, Emerson enhancement has been observed in higher plants, particularly under broad-spectrum irradiance typical of natural light fields [23,24,25].
Earlier enhancement studies often relied on complex or indirect measurements, such as fluorescence or multiple-electrode setups, or frozen specimens, which made it difficult to control the sample’s physiological state [26,27]. The development of the modulated oxygen electrode system [28,29,30,31] enabled more rapid and precise enhancement measurements by directly monitoring oxygen evolution at high temporal resolution. The present study enhances this methodology by using a single Joliot-type platinum electrode system with three independently controlled light beams, one modulated and two continuous, to measure high-resolution Emerson enhancement spectra in intact marine algae representing the major algal pigment systems. This approach enables precise determination of the spectral distribution of excitation energy between PSI and PSII under near-physiological conditions.
Rational
The theoretical and practical foundations of this study derive from the work on intensity-modulated light by [30,31,32]. In this context, light modulation enables differentiation between the effects of the modulated light and those of other light sources on the organism under study. A lock-in amplifier detects extremely weak modulated signals buried in large amounts of noise and other signals. Modulating the light intensity is the essential first step that "labels" the signal so the amplifier can find it. Wang and Myers (1976b) [32] defined AC as the modulated signal of photosynthetic oxygen evolution produced by a modulated “measuring” light beam of intensity Iₘ. In their equation 8, the quantum yield of this modulated radiation (AC/Iₘ), when added to a continuous “background” light beam of intensity Ib, is proportional to U, the fraction of open PSII reaction centers (Table 1).
A linear relationship between photosynthetic rate and light intensity (that is, between AC and Iₘ) requires a constant U, meaning that the wavelength scanning by the modulated measuring beam must not alter the redox state of the PSII reaction centers. When the two radiant beams have identical spectra, the AC amperometric signal reflects Ub, the steady-state open state of PSII reaction centers, as set by the background illumination.
In the present study, a scanning monochromatic beam of variable wavelength (Is) was added to the two beams to extend this model. This beam should not influence AC so long as it is not intense enough to change U. When the scanning beam is absorbed predominantly by pigments associated with PSII, U decreases because the supply of electrons to PSII increases without a corresponding rise in demand. This results in a decline in AC.
Conversely, when the scanning beam is absorbed primarily by pigments associated with PSI, U increases because the demand for electrons from PSII rises without additional electron supply, thereby increasing AC. Finally, when the spectral compositions of the scanning and measuring beams match, the relative electron distribution between the two photosystems remains unchanged, and U and AC remain constant. Thus, by monitoring the variation in AC with the wavelength of the scanning beam, the present experimental system enables the direct determination of action spectra for the quantum distribution of excitation energy between PSI and PSII, relative to a defined reference wavelength.
2. Results
A traditional criterion for classifying algae across different divisions is their characteristic complement of photosynthetic and accessory pigments, which together determine the whole-cell absorption spectrum (Table 2). The biliprotein-containing red algae (Porphyridium cruentum, Porphyra perforata, and Phycodrys sp.) and the cyanobacterium (Oscillatoria (Trichodesmium) thiebautii) in the present study share a similar complement of light-harvesting pigments, including phycoerythrin, phycocyanin, allophycocyanin, chlorophyll a, and simple carotenoids. The enhancement spectra of these taxa (Figure 1; Table 3) showed strong, well-defined signals. Enhancement maxima coincided with the principal absorption peaks of PSI chlorophyll a (435-440 nm and 678-680 nm) and its associated accessory pigments, whereas minima occurred at wavelengths corresponding to radiation primarily absorbed by PSII chlorophyll a (675-677 nm) and by its associated biliproteins and carotenoids in the green region of the spectrum (Table 2; [9]). The pigments complement in each algae can be expected to contribute significantly to whole-cell light absorption in the algal species examined (Table 2). The pigment assignments correspond to the observed enhancement maxima (peak wavelengths ± 2 nm) identified in the present study. These spectral attributions are based on previously characterized absorption features of known pigments, modified from [33], and supplemented with unpublished spectrophotometric data and representative literature sources cited below.
The enhancement spectrum of Porphyridium cruentum (Figure 1) closely resembled its PSI fractional absorption spectrum in this strain [34]. Distinct maxima or shoulders were evident near 436 and 680 nm (chlorophyll a), 477 and 495 nm (carotenoids), and 625 nm (chlorophyll a and phycocyanin). A pronounced minimum near 550 nm corresponded to B-phycoerythrin, a pigment associated with PSII. Enhancement spectra of Porphyra perforata and Phycodrys sp. were broadly similar to each other and closely matched previously published spectra for PSI activity and Emerson enhancement in red algae [27,35,36,37].
The enhancement spectrum of the red cyanobacterium Oscillatoria (Trichodesmium) thiebautii also resembled that of the red algae. The spectral region above 550 nm showed excellent agreement with the PSI activity spectrum [31] for the phycocyanin-rich cyanobacterium Anacystis nidulans. Shoulder features and peaks in the 480-520 nm region suggested an association between carotenoids and PSI light-harvesting.
The green macrophyte Ulva sp. exhibited a pigment complement typical of chlorophyll a-b algal systems. The action spectra of the two photosystems in such taxa are known to differ primarily in the spectral regions corresponding to PSII-associated chlorophyll b absorption [29,38,39] (Figure 1). As expected for green algae, the enhancement spectrum of Ulva showed pronounced minima at approximately 475 nm and 650 nm, coinciding with the in vivo absorption peaks of chlorophyll b [40], and maxima near 420, 436, 630, and 681 nm, corresponding to chlorophyll a absorption (Table 2). A subsidiary maximum near 520 nm likely resulted from energy transfer from carotenoids to PSI. The overall shape of this spectrum closely matched previously reported enhancement spectra for Ulva, other chlorophytes, and higher plants [35,38,41,42] , as well as the fluorescence excitation spectrum of the chlorophyll-protein complex I from green algae [43,44].
The cryptomonad Rhodomonas D3, like other red-colored cryptophytes [45,46], contains type I phycoerythrin and chlorophyll c as prominent light-harvesting pigments [26,33,47]. The dominant carotenoids are alloxanthin (approximately 70% of total carotenoids), with monadoxanthin (12%), crocoxanthin (9%), and α-carotene (8%). These pigments have di- or tri-peaked chromophores typical of α- and β-carotenoid structures.
The Rhodomonas D3 enhancement spectra (Figure 1) were obtained from cultures acclimated to low, medium, and high irradiance, as described in the Methods section. In all cultures, the principal enhancement peak was centered at 680 nm, corresponding to PSI chlorophyll a absorption. However, the 440-nm chlorophyll a peak was most pronounced in low-light cells, diminished in medium-light cells, and nearly absent under high-light conditions. In the green spectral region (525-600 nm), dominated by phycoerythrin absorption, the spectra showed clear minima at low and medium light intensities, consistent with the PSII association of this pigment.
The three Rhodomonas D3 spectra displayed minima at between 460 and 475 nm and at 495 nm, regions where absorption was dominated by PSII chlorophyll c and carotenoids (notably alloxanthin (Table 2)). Maxima between 500 and 515 nm, particularly prominent in the low-light spectrum, suggested participation of one or more carotenoids in PSI excitation, consistent with patterns observed in the red and blue-green algae examined (Figure 1). The enhancement spectrum of low-light Rhodomonas D3 closely resembled the PSI/PSII low-temperature fluorescence excitation spectrum reported for cryptomonads by Snyder and Biggins (1987).
The pigment system of Glenodinium sp. (now Heterocapsa pygmeae) was characteristic of photosynthetic dinoflagellates, containing chlorophyll a, chlorophyll c₂, and the carotenoid peridinin as the primary light-harvesting pigments, along with smaller amounts of β-carotene and diadinoxanthin [56,58] (Table 2). Enhancement spectra were determined for low-light and medium-light cultures. The minima at approximately 470 nm and 635 nm were slightly red-shifted relative to the in vivo absorption peaks of chlorophyll c. The magnitude of this shift was greater in Glenodinium than in Rhodomonas and corresponded to known differences in the chlorophyll c absorption maxima of the respective chlorophyll a-c protein complexes [53,59].
Both enhancement spectra of Glenodinium sp. (Figure 1) showed prominent activity maxima near 495 nm, close to the in vivo absorption maxima of β-carotene and diadinoxanthin (Table 2). These features implicate one or both of these carotenoids in PSI light harvesting. The 495-nm peak was considerably stronger in medium-light cells than in low-light cells, correlating with twice as high cellular ratios of (β-carotene + diadinoxanthin) to peridinin (0.362 versus 0.156; [9]).

Table 2.
Pigments contributing to whole-cell absorption in the studied species.
| Genus | In vivo absorption maxima (nm) | Pigment |
|---|---|---|
|
Porphyra perforata abci (Rhodophyta) |
436, 677 493, 546, 565 617 |
Chlorophyll a R-phycoerythrin R-phycocyanin |
|
Phycodrys dc (Rhodophyta) |
438, 625, 680 498, 540, 566 625 |
Chlorophyll a R-phycoerythrin T-phycocyanin |
|
Rhodomonas bdfij (Cryptophyta) |
435, 638, 676 460, 632 545, 565 460, 495 |
Chlorophyll a Chlorophyll c2 Phycoerythrin type I Alloxanthin |
|
Glenodinium p (Pyrrophyta) |
437, 590, 622, 676 462, 590, 632 525 (broad) 497 |
Chlorophyll a Chlorophyll c2 Peridinin β-Carotene and minor xanthophylls |
|
Ulva abqr (Chlorophyta) |
437, 678 475, 652 475 |
Chlorophyll a Chlorophyll b Lutein and other carotenoids |
a [48]Haxo and Blinks (1950); b [16]French and Fork (1961); c [49]Haxo et al. (1955); d [50]O’Carra and O’hEocha (1976); e resembles Selesseria decipiens [48]Haxo and Blinks (1950); r [51]Haxo and Fork (1959), like Rhodomonas lens; f [45]Chapman (1966); i [46]Pennington el al. (1985); J [52]MacColl et al. (1976); l [53]Ingram and Hiller (1983); m [54]Mann and Myers (1968); n [55]Goedheer (1973); p [56]Prézelin et al. (1976); q [35]Fork (1963); r [57]Vidaver (1966).
The minima observed in the green spectral region, centered at approximately 526 nanometers in medium-light cells and 550 nanometers in low-light cells, coincided with the broad absorption band of peridinin, the principal light-harvesting pigment associated with PSII in dinoflagellates [56]. Pronounced enhancement maxima near 436 and 680 nanometers indicate that a substantial proportion of chlorophyll a in this species functions in PSI light-harvesting.
Comparable measurements with the diatom Chaetoceros gracilis and the phycocyanin-containing cryptomonad Chroomonas sp. (data not shown) produced nearly flat enhancement spectra across both the visible and far-red regions. The apparent absence of discernible enhancement signals in these taxa suggests a balanced distribution of excitation energy between PSI and PSII at all measured wavelengths, consistent with the interpretation proposed for diatoms in [54].
3. Discussion
Low-temperature fluorescence emission at liquid nitrogen temperature (77 K; -196 °C) has long been used as an alternative method for studying the distribution of excitation energy between the two photosystems [60,61]. However, these measurements do not preserve sample viability, and assigning specific pigments to the photosystems under cryogenic conditions may not always be valid [16](Larkum and Barrett 1983, section VI.D).
The enhancement method presented here, therefore, complements and refines low-temperature studies by preserving physiological integrity and enabling in vivo determination of energy-transfer relationships. This approach may also aid the interpretation of prior spectroscopic investigations, such as [7,39,62,63,64,65,66,67,68].
For instance, differences between pigment-photosystem assignments observed at low temperature and those derived from enhancement spectra under physiological conditions can reveal the dynamic redistribution of energy pathways in living algal cells. The present measurements also help interpret field data. In our previous research, the submerged aquatic plant Najas Marina L. ceased growth at a much higher light intensity in river field studies [69] than in laboratory studies using neutral-density filters [70]. Spectrally imbalanced attenuation of light, which disrupts the balance between PSI and PSII excitation and reduces photosynthetic efficiency, could explain this observation and may account for significant variability in the response of such plants to growth at depth, since different colors of light attenuate at different rates in natural waters. Spectrally distinct attenuation rates produce spectra that deviate further from white light with depth than those of light attenuated by neutral-density filters [71].
The close correspondence between the enhancement spectra obtained in this study and those previously reported for well-characterized algal pigment systems validates the experimental design. This agreement instills strong confidence in the interpretation of photoadaptive features observed in the less-studied pigment systems of dinoflagellates and cryptomonads. The facility and precision of the current methodology make it particularly well suited to examining, using Emerson enhancement spectra, the actual distribution of light-harvesting components between the two photosystems and to confirming information from less direct measurements, such as low-temperature spectra and biochemical analyses of the photosystems. Additionally, it enables analysis of how this balance is regulated by environmental and physiological factors, including nutrient availability, growth phase, and the spectral composition of incoming light.
3.1. Rhodophytes and Cyanobacteria
Photosynthetic enhancement in the photosynthetic systems of red algae and cyanobacteria is driven by the excitation balance between PSI and PSII. Therefore, the strongest enhancement peaks were expected in biliprotein-containing algae, whose photosystem pigments have distinct, minimally overlapping absorption spectra [52]. This separation creates broad wavelength regions in which excitation is dominated by one photosystem rather than the other. Because photosynthetic enhancement in the photosynthetic systems of red algae and cyanobacteria is driven by the excitation balance between PSI and PSII, the strongest enhancement peaks were expected in biliprotein-containing algae, whose photosystem pigments have distinct, minimally overlapping absorption spectra [52]. This separation creates broad wavelength regions in which excitation is dominated by one photosystem.
Indeed, in the three red algal species and the cyanobacterium examined, prominent enhancement maxima were observed at the chlorophyll a absorption peaks near both ends of the visible spectrum, and enhancement minima occurred in the central spectral regions dominated by phycoerythrin absorption. The broad enhancement shoulders between 590 and 650 nanometers were of particular interest. These features confirm that a portion of the phycocyanin pool contributes to PSI excitation, as previously proposed for Anacystis nidulans [31] and Cyanidium caldarium [39].
3.2. Chlorophytes
The enhancement spectrum for Ulva sp. closely matched previously reported chlorophyte spectra. Prior research has shown that enhancement amplitude correlates strongly with the PSI/PSII activity ratio and the redox state of PSII primary acceptors, which are reduced by PSII and reoxidized by PSI. Reviews by Barber (2008) and Govindjee et al. (2012) [72,73] support this interpretation of the spectra obtained in our study.
3.3. Cryptomonads
French and Fork (1961) [26] did not observe Emerson enhancement in Rhodomonas D3, making our earlier study [74] the first to document enhancement spectra for any cryptomonad. Earlier work [51] showed that, unlike other biliprotein-containing algae, much of the light absorbed by chlorophyll a in Rhodomonas D3 was used to excite PSII, with activity peaks at 436 and 675 nm. We estimated this contribution to be less than half that of biliproteins in cryptomonads [33,47], suggesting that a substantial portion of chlorophyll a remains associated with PSI rather than with PSII.
Chlorophyll a was proposed [62] to serve as the exclusive light-harvesting pigment for PSI in cryptomonads. If correct, enhancement spectra should show maxima at chlorophyll a absorption wavelengths and minima at those of chlorophyll c and phycoerythrin. Our spectra support this prediction and indicate energy transfer from minor carotenoids to PSI, as observed elsewhere in cryptomonads [63,75] and other algae [55,76].
Comparisons of enhancement spectra in cultures acclimated to low and high irradiance reveal a dual role for the carotenoid alloxanthin. Under low light, alloxanthin transfers more than half of the absorbed energy to PSII chlorophyll a, whose absorption is red-shifted relative to PSI [14,77]. This red-shift is reflected in minima near 460 and 495 nm. Under high light, Rhodomonas D3 exhibits nearly a fourfold increase in the alloxanthin-to-chlorophyll a ratio, acting primarily as a blue-light screen [47] rather than redistributing energy to PSI. Consequently, high-light cells reduce the blue radiation available to both photosystems by enhancing absorption by photosynthetically inactive alloxanthin. These findings have since been confirmed by observations that alloxanthin participates in non-photochemical quenching in Rhodomonas salina [77] and in cyanobacteria [78].
3.4. Dinoflagellates
The current findings for Glenodinium provide actual photosynthesis data with higher spectral resolution that corroborate earlier observations. The pigment associations in Glenodinium in the present enhancement spectra align with earlier physiological ([56]Prézelin et al. 1976) and biochemical [59,79](Prézelin and Alberte 1978; Boczar et al. 1980) research. The action spectrum of the peridinin-rich dinoflagellate Gonyaulax polyedra closely resembled that of the fucoxanthin-containing brown alga Endarachne sp. [26](French and Fork, 1961), suggesting that the association of both chlorophyll c and peridinin with PSII prevails in all dinoflagellates and brown algae.
3.5. Comparative Interpretation Between Species
Enhancement spectra can be compared across the representative taxa examined, including Rhodomonas D3 (Cryptophyta), Glenodinium sp. (Dinophyta), Ulva sp. (Chlorophyta), Porphyra perforata and Porphyridium cruentum (Rhodophyta), and Oscillatoria (Trichodesmium) thiebautii (Cyanobacteria). The present data reveal both conserved and divergent strategies in the distribution and utilization of excitation energy between the two photosystems. These differences reflect distinct pigment compositions and photoadaptive capacities that have evolved in response to the optical environments typical of their natural habitats.
Across the examined taxa, enhancement maxima were observed near 436 and 680 nanometers, wavelengths corresponding to chlorophyll a absorption peaks characteristic of PSI. However, the spectral regions between 480 and 550 nanometers, dominated by carotenoid and biliprotein absorption, showed minima with pronounced species-specific variability, indicating that accessory pigments in algae of different taxa can play different roles in modulating energy transfer and photoprotection.
In biliprotein-rich red algae and cyanobacteria, the clear spectral segregation of PSI and PSII pigments yields strong enhancement patterns with well-defined maxima and minima. This pattern is consistent with efficient partitioning of radiant energy between the two photosystems and with minimal spectral overlap between chlorophyll a and biliprotein absorption bands. The broad enhancement shoulders observed between 590 and 650 nanometers may indicate a partial contribution of phycocyanin to PSI excitation, consistent with earlier interpretations by [31,39].
In contrast, the chlorophyte Ulva sp., which can be considered representative of other chlorophyll a–b pigment systems, including higher plants, shows more moderate enhancement amplitudes, reflecting the close spectral overlap between chlorophyll a and b. These spectra align with well-established correlations between the magnitude of the enhancement and the PSI-to-PSII activity ratio [72,73], indicating that chlorophytes modulate photosystem excitation primarily through changes in the functional balance of the two photosystems’ reaction centers rather than through distinct pigment-specific pathways.
Among the more spectrally complex taxa, the cryptomonad Rhodomonas D3 and the dinoflagellate Glenodinium sp. illustrate contrasting strategies for managing excitation energy under variable irradiance. In Rhodomonas D3, enhanced alloxanthin absorption under high-light conditions acts as a dynamic optical screen, selectively attenuating blue and green radiation and thereby reducing apparent excess excitation on both photosystems. This specialized adaptation, distinct among algae, has been confirmed by Lawrenz & Richardson (2017) and likely represents a photoprotective mechanism rather than a redistribution of excitation toward PSI. In Glenodinium sp., however, increased irradiance enhances the effectiveness of green radiation in driving PSI activity, implying a partial reallocation of excitation energy mediated by carotenoids such as peridinin and diadinoxanthin. This spectral plasticity is consistent with Glenodinium's adaptation to life under high-light conditions [82,83].
The pronounced enhancement in the far-red regions of the spectra of Oscillatoria, Rhodomonas, and Ulva aligns with and is better interpreted in light of older and recent reports of increased photosynthetic efficiency under far-red light in algae and higher plants [56,84,85]. These reports indicate that the ability of different species to modulate pigment composition and allocation in response to irradiance likely underlies these effects. Under high-light conditions, Rhodomonas D3 increased the ratio of the major light-harvesting carotenoid (alloxanthin) to chlorophyll a severalfold. In contrast, Glenodinium sp. maintained a stable or slightly decreased ratio of peridinin to chlorophyll a.
The present results confirm the photosynthetic contributions of chlorophyll a to PSI and peridinin to PSII, and support the view that photo-adaptive responses among dinoflagellates may vary considerably between species [13,86]. Inconsistencies among pigment composition, photochemical activity, and irradiance response in dinoflagellates have been widely documented [87]. As observed for alloxanthin in medium- and high-light Rhodomonas D3 cultures, the photosynthetic enhancement spectra presented here reveal photosynthetically inactive carotenoids that function primarily in light screening. No such evidence was detected in medium-light Glenodinium sp., suggesting that inactive carotenoids did not accumulate under these conditions. However, our enhancement spectra from red tide populations of the dinoflagellate Prorocentrum micans[88] indicated the presence of such inactive carotenoids. Since then, photoprotective carotenoids and related compounds have been identified across a wide range of algal taxa [89,90,91], underscoring the generality of these mechanisms in mitigating excess light stress in aquatic phototrophs.
4. Materials and Methods
4.1. Measurement of Enhancement Spectra
Photosynthetic rate measurements were performed using the automated action-spectrum apparatus (Figure 2), an upgrade of the apparatus described in detail in [33]. The system measures PSII activity in microalgae by placing a thin layer of algal cells directly on a recessed bare platinum oxygen electrode of the Joliot configuration [28], held in place by a semipermeable 33 pm thick Visking dialysis membrane. With macroalgae, a raised electrode was used, so that the retaining dialysis membrane held the algal blade in close contact with the platinum surface. This setup produced an amperometric signal proportional to the rate of photosynthetic oxygen production. The sample was simultaneously irradiated by a continuous background light beam and a beam from a 150 W xenon arc lamp, which passed through a microcomputer-controlled, motor-driven monochromator and was chopped at 10 Hz by a rotating disc. The reduced image of the grating was focused on the surface of the algae [33]. The computer controlled the light level at each wavelength by rotating a calibrated circular-graduated UV-VIS-NIR neutral-density disk (Oriel Corp., model 28650) using feedback from a calibrated silicon photodiode (EG&G model UV-444BQ). Final quantum corrections were made using a thermopile (Kettering Scientific Research, Model 68) to keep the monochromatic, modulated beam’s quantum flux incident on the electrode mount constant across all wavelengths. The monochromator light was scanned across the visible spectrum, and the amperometric signal from the electrode was extracted and amplified using a lock-in amplifier. A second motor, also under computer control, rotated a circular, variable-density neutral filter in response to feedback from a silicon photodiode monitoring the light reaching the electrode, thereby maintaining a constant quantum flux across all wavelengths. The computer automatically acquired the amperometric signal and recorded its amplitudes as a function of wavelength.
The following instrumental and procedural modifications were implemented to enable measurement of photosynthetic enhancement:
Three radiant beams, defined in Table 3, were directed at the algal sample on the electrode:
- Background beam: A broadband continuous light source was applied at an intensity Ib sufficient to elicit a strong yet linear oxygen-evolution response. Its spectral distribution (referred to as radiation II) was preferentially absorbed by accessory pigments associated with PSII and overlapped with wavelengths corresponding to regions of maximal quantum yield in the PSII action spectrum of each alga.
- Measuring beam: This modulated beam of intensity Im had the same spectral characteristics as the higher-intensity background beam (radiation II). A 10 Herz modulation frequency produced the highest-amplitude alternating-current oxygen-evolution signal, consistent with findings from later independent studies [92].
- Scanning beam: A weak, continuous, monochromatic beam with a five-nanometer half-bandwidth was generated by the computer-controlled monochromator and used to scan the spectral range (radiation I). The instrument automatically maintained a constant quantum flux to the sample across all wavelengths.
Careful adjustments to the intensities of the background and measuring beams established experimental conditions in which the photosynthetic system reached a steady state with a constant proportion of open PSII reaction centers (U). Under these conditions, the modulated oxygen-evolution signal (AC) was linearly proportional to the intensity of the modulated measuring beam (Iₘ), indicating that the PSII quantum yield remained constant and unaffected despite small fluctuations in the background beam intensity or by the addition of a scanning beam of the same spectral composition (radiation II).
The intensities of the three radiant beams were calibrated as follows: After aligning the algal mount, the sample was first exposed to a weak modulated measuring beam of radiation II. A weak, continuous background beam of a radiation II spectrum was then introduced, and the modulated oxygen signal (AC) was monitored as the background intensity (Ib) was incrementally increased until AC reached its maximum. This procedure was repeated across a range of Iₘ and Ib intensities to obtain a stable, low-noise AC signal with a high Ib/Iₘ ratio. The combined intensities of the two beams were kept within the linear region of the AC versus I curve, which was determined separately for each sample.
Once intensities for a stable signal were established, the third scanning beam was added. Its wavelength was initially set to the spectral peak that produced the maximal AC signal without reducing its amplitude (that is, without altering U). The scanning beam was then shifted to 695 nanometers, a wavelength predominantly absorbed by pigments associated with PSI (radiation I). If adding this beam produced a measurable increase in the AC signal (representing enhancement), the intensity of the scanning beam was gradually reduced to the lowest level at which enhancement could still be detected, ensuring that Is remained smaller than (Ib + Im).
The modulated oxygen-evolution signal was continuously recorded as the scanning beam was swept across the range from 380 to 720 nanometers. Each complete spectral scan required approximately one hour. The method enabled measurement of small enhancement changes superimposed on large modulated signals. The modulated oxygen-evolution signal was continuously acquired by the computer as the scanning beam was swept across the 380-720 nm range. Each complete spectral scan required approximately one hour. The method thereby enabled measurement of small enhancement changes superimposed on large modulated signals.
4.2. Algal Material
Studies of macroalgae were conducted using specimen mounts similar to those described in [33. Porphyra perforata J. Agardh (Rhodophyta) and Ulva sp. (probably U. californica Wille, 1899; Chlorophyta) were collected from rocky intertidal habitats and maintained under low irradiance (approximately 3 µmol photons m⁻² s⁻¹) in flowing seawater aquaria at approximately 18 °C. Phycodrys sp. (strain JS-SC4#119; Rhodophyta) was maintained under similar light conditions at approximately 12 °C.
The cyanobacterium Oscillatoria (Trichodesmium) aff. thiebautii (Gomont ex Gomont) Geitler (Qingdao isolate from the South China Sea; Cyanobacteria) was cultured at 20-23 °C in Provasoli’s enriched seawater medium [94]. Cultures were vigorously shaken and continuously illuminated at 20 µmol photons m⁻² s⁻¹ using cool white fluorescent lamps. The in vivo spectral properties and pigment composition of this isolate, grown under identical conditions, were previously characterized [95].
Table 3.
Measurement of light intensities in µmol photons·m-²·s-¹ and optical filter combinations used to determine the Emerson enhancement spectra, which are presented in Figure 1. The table lists the experimental parameters used to measure the Emerson enhancement spectra shown in Figure 1. It specifies the intensities of the background, measuring, and scanning light beams, as well as the combinations of optical filters that define their spectral distributions. .
Table 3.
Measurement of light intensities in µmol photons·m-²·s-¹ and optical filter combinations used to determine the Emerson enhancement spectra, which are presented in Figure 1. The table lists the experimental parameters used to measure the Emerson enhancement spectra shown in Figure 1. It specifies the intensities of the background, measuring, and scanning light beams, as well as the combinations of optical filters that define their spectral distributions. .
| Species | n | Background Beam | Measuring Beam | Scanning Beam |
|---|---|---|---|---|
| Porphyridium cruentum | 2 | 150.5 1+3 | 10.5 2+5 | 10.5 7 |
| (7.0) | (0.5) | (0.5) | ||
| Porphyra perforata | 1 | 60.5 1+3 | 2.5 2+3 | 6.5 7 |
| (3.0) | (0.12) | (0.3) | ||
| Phycodrys sp. | 1 | 60.5 1+3 | 5.5 2+3 | 3.5 7 |
| (3.0) | (0.25) | (0.15) | ||
|
Oscillatoria (trichodesmium) thiebautii |
3 | 5.5 3 | 15.5 2+3 | 2.5 7 |
| (0.25) | (0.7) | (0.12) | ||
| Ulva sp. | 3 | 40.0 1+4 | 4.5 2+4 | 20.5 6 |
| (1.6) | (0.15) | (0.8) |
Beam filter definitions1 Calflex-C heat-reflecting filter; 2 2 cm of water; 3 Corning CS4-64 green filter; 4 Balzers interference filter 460 nm, 8 nm bandwidth; 5 Balzers interference filter 555 nm, 8 nm bandwidth; 6 at 460 nm; 7 at 555 nm.
Rhodomonas D3 (Provasoli isolate, UTEX #2419; Cryptophyta) was grown in D medium [93] at 18 °C in batch cultures, manually agitated daily. Cultures were acclimated to low light (20 µmol photons m⁻² s⁻¹), medium light (210 µmol photons m⁻² s⁻¹), and high light (370 µmol photons m⁻² s⁻¹) under cool white fluorescent illumination.
Glenodinium sp. (Bernhard isolate, UTEX #2421; now Heterocapsa pygmeae Loeblich, Schmidt, and Sherley; Dinophyta, [94]) was grown in GPM medium [95] at irradiances of 27 µmol photons m⁻² s⁻¹ (low light) and 270 µmol photons m⁻² s⁻¹ (medium light), under conditions similar to those described by [56]. Other culture conditions matched those described above for Rhodomonas D3.
Porphyridium cruentum (Agardh) Nägeli (UTEX #161; Rhodophyta) was maintained in batch culture at 17 °C in GPM medium [95]. Flasks were shaken vigorously and continuously illuminated at 20 µmol photons m⁻² s⁻¹.
With the exception of the cyanobacterial genus Oscillatoria (Trichodesmium), which is ecologically important and contains an atypical form of phycoerythrin [96,97]. The selected species contained conventional pigment composition and spectral absorption properties. The major absorption features of these taxa have been summarized previously [33] and in the references therein, the major absorption features of Porphyridium cruentum were described in [98].
5. Conclusions
The investigation demonstrates that Emerson enhancement spectra provide a superior, non-invasive method for examining in vivo energy transfer, thereby preserving physiological integrity when cryogenic techniques are inadequate. By validating this approach against characterized algal systems, the research shows that diverse taxonomic groups employ both conserved and divergent strategies to regulate excitation energy. For instance, algae with biliproteins rely on distinct spectral segregation, whereas chlorophytes manage energy via functional equilibrium within overlapping spectra. Moreover, photosynthetically less well-understood taxa, such as cryptomonads and dinoflagellates, exhibit considerable spectral plasticity; they can either reallocate energy between photosystems or use pigments like alloxanthin as photoprotective screens, depending on incident light intensity. These discoveries clarify why spectrally imbalanced illumination in natural aquatic environments can diminish photosynthetic efficiency and impede growth compared with controlled laboratory conditions. Ultimately, these findings corroborate the idea that an organism’s capacity to modulate pigment composition and energy allocation is an evolutionary adaptation that equips a species to meet the specific optical challenges of its natural milieu.
Author Contributions
Conceptualization, M.A. and A.N.; Writing—original draft preparation, A.N.; writing—review and editing, A.N. and M.A.; Both authors have read and agreed to the published version of the manuscript.
Funding
This research received funding from the Foundation for Ocean Research. La Jolla, California, from the resources of the Scripps Institution of Oceanography- Marine Biology Research Department, and the Food Chain Research Group. AN was supported by Scholarships from the Gershon Meyerbaum Fund for Oceanography, the Hebrew University of Jerusalem, and the Graduate Department of the University of California, San Diego (UCSD). Further research funds were received from grants to OHH and FTH from the US Department of Energy (DOE) and the National Science Foundation (NSF).
Data Availability Statement
Original spectral data can be obtained from [9].
Acknowledgments
This paper is dedicated to my late mentors, Francis T. Haxo and Osmund Holm-Hansen, whose guidance, scientific vision, and confidence in my abilities were instrumental in shaping this research. Rocky Booth helped design the system, and Tim Nichols developed and built an interface between a Commodore 64 computer and a custom experimental spectrum-measuring system constructed for this research. The current study expands the foundational work in [74], with explicit written permission for reproduction and extension obtained from the publisher of the scientific journal Photosynthetica, under the auspices of the Institute of Experimental Botany at the Czech Academy of Sciences in Prague, Czechia. This manuscript significantly updates and enriches the original research by incorporating 40 additional references. These references reflect key advancements and broader contextual insights in the field, underscoring the evolving nature of our understanding and the ongoing relevance of this investigation.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Govindjee, G. On the Evolution of the Concept of Two Light Reactions and Two Photosystems for Oxygenic Photosynthesis: A Personal Perspective. Photosynthetica 2023, 61, 37–47. [CrossRef]
- Emerson, R.; Rabinowitch, E. Red Drop and Role of Auxiliary Pigments in Photosynthesis. Plant Physiology 1960, 35, 477–485.
- Evans, J.R. A Quantitative Analysis of Light Distribution between the Two Photosystems, Considering Variation in Both the Relative Amounts of the Chlorophyll-Protein Complexes and the Spectral Quality of Light. Photobiochemistry and Photobiophysics 1986, 10, 135–147.
- Falkowski, P.; Owens, T.G. Light-Shade Adaptation. Plant Physiol. 1980, 66, 592–595.
- Malkin, S.; Herbert, S.K.; Fork, D.C. Light Distribution, Transfer and Utilization in the Marine Red Alga Porphyra perforata: From Photoacoustic Energy-Storage Measurements. Biochimica et Biophysica Acta (BBA) - Bioenergetics 1990, 1016, 177–189. [CrossRef]
- Gallagher, J.C.; Wood, A.M.; Alberte, R.S. Ecotypic Differentiation in the Marine Diatom Skeletonema costatum: Influence of Light Intensity on the Photosynthetic Apparatus. Mar. Biol. 1984, 82, 121–134. [CrossRef]
- Stadnichuk, I.N.; Novikova, T.M.; Miniuk, G.S.; Boichenko, V.A.; Bolychevtseva, Yu.V.; Gusev, E.S.; Lukashev, E.P. Phycoerythrin Association with Photosystem II in the Cryptophyte Alga Rhodomonas salina: Biochemistry Moscow 2020, 85, 679–688. [CrossRef]
- Neori, A.; Vernet, M.; Holm-Hansen, O.; Haxo, F.T. Comparison of Chlorophyll Far-Red and Red Fluorescence Excitation Spectra with Photosynthetic Oxygen Action Spectra for Photosystem II in Algae. Marine Ecology Progress Series 1988, 44, 297–302.
- Neori, A. Excitation Spectra of Chlorophyll a Fluorescence in vivo : Their Correlation with Photosynthetic Action Spectra and Their Use in the Study of Algal Photoadaptation, University of California, San Diego, 1986.
- Mitchell, B. g.; Kiefer, D. a. Determination of Absorption and Fluorescence Excitation Spectra for Phytoplankton. In Marine Phytoplankton and Productivity; American Geophysical Union (AGU), 1984; pp. 157–169 ISBN 978-1-118-66951-8.
- Neori, A. The Use of Fluorescence Excitation Spectra of Chlorophyll a in Phytoplankton to Estimate Photoadaptation and Its Relation to Photosynthesis Action Spectra. In Proceedings of the Group for Aquatic Productivity GAP II International Workshop, Haifa, Israel; 1984.
- Falkowski, P.G.; LaRoche, J. Acclimation to Spectral Irradiance in Algae. Journal of Phycology 1991, 27, 8–14. [CrossRef]
- Iglesias-Prieto, R.; Trench, R.K. Acclimation and Adaptation to Irradiance in Symbiotic Dinoflagellates. II. Response of Chlorophyll–Protein Complexes to Different Photon-Flux Densities. Marine Biology 1997, 130, 23–33. [CrossRef]
- Lawrenz, E.; Richardson, T.L. Differential Effects of Changes in Spectral Irradiance on Photoacclimation, Primary Productivity and Growth in Rhodomonas salina (Cryptophyceae) and Skeletonema costatum (Bacillariophyceae) in Simulated Blackwater Environments. Journal of Phycology 2017, 53, 1241–1254. [CrossRef]
- Richardson, K.; Beardall, J.; Raven, J.A. Adaptation of Unicellular Algae to Irradiance: An Analysis of Strategies. New Phytologist 1983, 93, 157–191. [CrossRef]
- Larkum, A.W.D.; Barrett, J. Light-Harvesting Processes in Algae. In Advances in Botanical Research; Woolhouse, H.W., Ed.; Academic Press, 1983; Vol. 10, pp. 1–219.
- Fujita, Y.; Murakami, A. Regulation of Electron Transport Composition in Cyanobacterial Photosynthetic System: Stoichiometry among Photosystem I and II Complexes and Their Light-Harvesting Antennae and Cytochrome B6/f Complex. Plant and Cell Physiology 1987, 28, 1547–1553. [CrossRef]
- Machalek, K.M.; Davison, I.R.; Falkowski, P.G. Thermal Acclimation and Photoacclimation of Photosynthesis in the Brown Alga Laminaria saccharina. Plant, Cell & Environment 1996, 19, 1005–1016. [CrossRef]
- Satoh, A.; Kurano, N.; Senger, H.; Miyachi, S. Regulation of Energy Balance in Photosystems in Response to Changes in CO2 Concentrations and Light Intensities during Growth in Extremely-High-CO2 -Tolerant Green Microalgae. Plant Cell Physiol 2002, 43, 440–451. [CrossRef]
- Ueno, Y.; Aikawa, S.; Kondo, A.; Akimoto, S. Adaptation of Light-Harvesting Functions of Unicellular Green Algae to Different Light Qualities. Photosynth Res 2019, 139, 145–154. [CrossRef]
- Santabarbara, S.; Villafiorita Monteleone, F.; Remelli, W.; Rizzo, F.; Menin, B.; Casazza, A.P. Comparative Excitation-Emission Dependence of the FV/FM Ratio in Model Green Algae and Cyanobacterial Strains. Physiologia Plantarum 2019, 166, 351–364. [CrossRef]
- Dubinsky, Z.; Falkowski, P.G.; Wyman, K. Light Harvesting and Utilization by Phytoplankton. Plant Cell Physiol. 1986, 27(7), 1335–1349.
- Dring, M.J.; Lüning, K. Emerson Enhancement Effect and Quantum Yield of Photosynthesis for Marine Macroalgae in Stimulated Underwater Light Fields. Mar. Biol. 1985, 87, 109–117.
- Jerlov, N.G. Marine Optics; Elsevier Oceanography Series; 2nd ed.; Elsevies: Amsterdam, 1976;
- McCree, K.J. Significance of Enhancement for Calculations Based on the Action Spectrum for Photosynthesis 1. Plant Physiology 1972, 49, 704–706. [CrossRef]
- French, C.S.; Fork, D.C. Computer Solutions for Photosynthesis Rates from a Two Pigment Model. Biophysical Journal 1961, 1, 669–681. [CrossRef]
- Ried, A.; Hessenberg, B.; Metzler, H.; Ziegler, R. Distribution of Excitation Energy among Photosystem I and Photosystem II in Red Algae. I. Action Spectra of Light Reactions I and II. Biochimica et Biophysica Acta (BBA) - Bioenergetics 1977, 459, 175–186. [CrossRef]
- Joliot, P.; Joliot, A. A Polarographic Method for Detection of Oxygen Production and Reduction of Hill Reagent by Isolated Chloroplasts. Biochimica et Biophysica Acta (BBA) - Bioenergetics 1968, 153, 625–634. [CrossRef]
- Joliot, P.; Joliot, A.; Kok, B. Analysis of the Interactions between the Two Photosystems in Isolated Chloroplasts. Biochimica et Biophysica Acta (BBA) - Bioenergetics 1968, 153, 635–652. [CrossRef]
- Wang, R.T.; Myers, J. On the Distribution of Excitation Energy to Two Photoreactions of Photosynthesis. Photochemistry and Photobiology 1976, 23, 405–410. [CrossRef]
- Wang, R.T.; Stevens, C.L.R.; Myers, J. Action Spectra for Photoreactions I and II of Photosynthesis in the Blue-Green Alga Anacystis nidulans. Photochemistry and Photobiology 1977, 25, 103–108. [CrossRef]
- Wang, T.; Myers, J. Simultaneous Measurment of Action Spectra for Photoreactions I and II of Photosynthesis. Photochemistry and Photobiology. 1976, 23, 411–414.
- Neori, A.; Vernet, M.; Holm-Hansen, O.; Haxo, F.T. Relationship between Action Spectra for Chlorophyll a Fluorescence and Photosynthetic O2 Evolution in Algae. J Plankton Res 1986, 8, 537–548. [CrossRef]
- Ley, A.C.; Butler, W.L. Energy Transfer from Photosystem II to Photosystem I in Porphyridium cruentum. Biochimica et Biophysica Acta (BBA) - Bioenergetics 1977, 462, 290–294. [CrossRef]
- Fork, D. c. Observations on the Function of Chl a and Accessory Pigments in Photosynthesis. In Photosynthetic Mechanisms of Green Plants; Kok, B., Jagendorf, A.T., Eds.; Nat. Acad. Sci. Nat. Res. Council: Washington D.C., 1963; pp. 352–361.
- Ludlow, C.J.; Park, R.B. Action Spectra for Photosystems I and II in Formaldehyde Fixed Algae 1. Plant Physiology 1969, 44, 540–543. [CrossRef]
- Hamada, E. a. M.; Dowiadar, S.M.A.; Punnet, T. Influence of Spectral Range and Carbon and Nitrogen Sources on Oxygen Evolution and Emerson Enhancement in Chlamydomonas reinhardtii. Biologia Plantarum 2003, 3, 389–397. [CrossRef]
- Ried, A. Improved Action Spectra of Light Reaction I And II. In Proceedings of the Photosynthesis, two centuries after its discovery by Joseph Priestley; Forti, G., Avron, M., Melandri, A., Eds.; Springer Netherlands: Dordrecht, 1972; pp. 763–772.
- Kramer, H.J.M.; Westerhuis, W.H.J.; Amesz, J. Low Temperature Spectroscopy of Intact Algae. Physiol. vég. (Paris) 1985, 23, 535–543.
- Kok, B.; Gott, W. Activation Spectra of 700 Mμ Absorption Change in Photosynthesis. Plant Physiology 1960, 35, 802. [CrossRef]
- Joliot, P. Cinétiques Des Réactions Liées a l’émission d’oxygène Photosynthétique. Biochimica et Biophysica Acta (BBA) - Biophysics including Photosynthesis 1965, 102, 116–134. [CrossRef]
- Myers, J.; Graham, J.-R. Enhancement in Chlorella. Plant Physiol. 1963, 38, 1–5. [CrossRef]
- Itagaki, T.; Nakayama, K.; Okada, M. Chlorophyll-Protein Complexes Associated with Photosystem I Isolated from the Green Alga, Bryopsis maxima. Plant Cell Physiol 1986, 27, 1241–1247. [CrossRef]
- Laisk, A.; Oja, V.; Eichelmann, H.; Dall’Osto, L. Action Spectra of Photosystems II and I and Quantum Yield of Photosynthesis in Leaves in State 1. Biochimica et Biophysica Acta (BBA) - Bioenergetics 2014, 1837, 315–325. [CrossRef]
- Chapman, D.J. Studies on the Carotenoids of the Flagellate Cryptophyceae, and the Chloroplast Pigments of the Endosymbiotic Algae in Cyanophora paradoxa and Glaucocystis nostochinearum. Ph.D. Dissertation, University of California, San Diego, Calif .: San Diego, 1966.
- Pennington, F.C.; Haxo, F.T.; Borch, G.; Liaaen-Jensen, S. Carotenoids of Cryptophyceae. Biochemical Systematics and Ecology 1985, 13, 215–219. [CrossRef]
- Vernet, M.; Neori, A.; Haxo, F.T. Role of Carotenoid Pigments in Photosynthesis of Cryptophyceae. In Proceedings of the Transactions American Geophysical Union; Washington D.C., 1985; Vol. 66 (51), p. 1302.
- Haxo, F.T.; Blinks, L.R. Photosynthetic Action Spectra of Marine Algae. J Gen Physiol 1950, 33, 389–422. [CrossRef]
- Haxo, F.; O’hEocha, C.O.; Norris, P. Comparative Studies of Chromatographically Separated Phycoerythrins and Phycocyanins. Archives of Biochemistry and Biophysics 1955, 54, 162–173. [CrossRef]
- O’Carra, P.; O’hEocha, C. Algal Biliproteins and Phycobilins. In Chemistry and biochemistry of plant pigments; Goodwin, T.W., Ed.; Academic Press: New York, 1965; pp. 328–376 ISBN 978-0-12-289902-7.
- Haxo, F.T.; Fork, D.C. Photoreactive Pigments in Flagellates: Photosynthetically Active Accessory Pigments of Cryptomonads. Nature 1959, 184, 1051–1052. [CrossRef]
- MacColl, R.; Berns, D.S.; Gibbons, O. Characterization of Cryptomonad Phycoerythrin and Phycocyanin. Archives of Biochemistry and Biophysics 1976, 177, 265–275. [CrossRef]
- Ingram, K.; Hiller, R.G. Isolation and Characterization of a Major Chlorophyll Alc2 Light-Harvesting Protein from a Chroomonas Species (Cryptophyceae). Biochimica et Biophysica Acta (BBA) - Bioenergetics 1983, 722, 310–319. [CrossRef]
- Mann, J.E.; Myers, J. Photosynthetic Enhancement in the Diatom Phaeodactylum tricornutum. Plant Physiol 1968, 43, 1991–1995. [CrossRef]
- Goedheer, J.C. Chlorophyll a Forms in Phaeodactylum tricornutum: Comparison with Other Diatoms and Brown Algae. Biochimica et Biophysica Acta (BBA) - Bioenergetics 1973, 314, 191–201. [CrossRef]
- Prézelin, B.B.; Ley, A.C.; Haxo, F.T. Effects of Growth Irradiance on the Photosynthetic Action Spectra of the Marine Dinoflagellate, Glenodinium Sp. Planta 1976, 130, 251–256. [CrossRef]
- Vidaver, W. Separate Action Spectra for the Two Photochemical Systems of Photosynthesis. Plant Physiology 1966, 41, 87–89.
- Johansen, J.E.; Svec, W.A.; Liaaen-Jensen, S.; Haxo, F.T. Carotenoids of the Dinophyceae. Phytochemistry 1974, 13, 2261–2271. [CrossRef]
- Boczar, B.A.; Prézelin, B.B.; Markwell, J.P.; Thornber, J.P. A Chlorophyll c -containing Pigment—Protein Complex from the Marine Dinoflagellate, Glenodinium Sp. FEBS Letters 1980, 120, 243–247. [CrossRef]
- Butler, W.L. On the Primary Nature of Fluorescence Yield Changes Associated with Photosynthesis. Proceedings of the National Academy of Sciences 1972, 69, 3420–3422. [CrossRef]
- Murata, N. The Discovery of State Transitions in Photosynthesis 40 Years Ago. Photosynth Res 2009, 99, 155–160. [CrossRef]
- Lichtlé, C.; Jupin, H.; Duval, J.C. Energy Transfers from Photosystem II to Photosystem I in Cryptomonas rufescens (Cryptophyceae). Biochimica et Biophysica Acta (BBA) - Bioenergetics 1980, 591, 104–112. [CrossRef]
- Bruce, D.; Biggins, J.; Steiner, T.; Thewalt, M. Excitation Energy Transfer in the Cryptophytes. Fluorescence Excitation Spectra and Picosecond Time-Resolved Emission Spectra of Intact Algae at 77 K. Photochemistry and Photobiology 1986, 44, 519–525. [CrossRef]
- Snyder, U.K.; Biggins, J. Excitation-Energy Redistribution in the Cryptomonad Alga Cryptomonas ovata. Biochimica et Biophysica Acta (BBA) - Bioenergetics 1987, 892, 48–55. [CrossRef]
- Krausz, E.; Hughes, J.L.; Smith, P.J.; Pace, R.J.; Årsköld, S.P. Assignment of the Low-Temperature Fluorescence in Oxygen-Evolving Photosystem II. Photosynth Res 2005, 84, 193–199. [CrossRef]
- Mohanty, P.; Allakhverdiev, S.I.; Murata, N. Application of Low Temperatures during Photoinhibition Allows Characterization of Individual Steps in Photodamage and the Repair of Photosystem II. Photosynth Res 2007, 94, 217–224. [CrossRef]
- Romero, E.; van Stokkum, I.H.M.; Novoderezhkin, V.I.; Dekker, J.P.; van Grondelle, R. Two Different Charge Separation Pathways in Photosystem II. Biochemistry 2010, 49, 4300–4307. [CrossRef]
- Luimstra, V.M.; Schuurmans, J.M.; Verschoor, A.M.; Hellingwerf, K.J.; Huisman, J.; Matthijs, H.C.P. Blue Light Reduces Photosynthetic Efficiency of Cyanobacteria through an Imbalance between Photosystems I and II. Photosynth Res 2018, 138, 177–189. [CrossRef]
- Agami, M.; Beer., S.; Waisel, Y. Seasonal Variations in the Growth Capacity of Najas marina L. as a Function of Various Water Depths at the Yarkon Springs, Israel.. Aquatic Botany. 1984, 19, 45–51.
- Agami, M.; Beer, S.; Waisel, Y. Growth and Photosynthesis of Najas marina L. as Affected by Light Intensity. Aquatic Botany 1980, 9, 285–289. [CrossRef]
- Frenette, J.-J.; Massicotte, P.; Lapierre, J.-F. Colorful Niches of Phytoplankton Shaped by the Spatial Connectivity in a Large River Ecosystem: A Riverscape Perspective. PLOS ONE 2012, 7, e35891. [CrossRef]
- Barber, J. Photosynthetic Energy Conversion: Natural and Artificial. Chemical Society Reviews 2009, 38, 185–196. [CrossRef]
- Govindjee; Björn, L.O.; Nickelsen, K. Evolution of the Z-Scheme of Electron Transport in Oxygenic Photosynthesis. In Proceedings of the Photosynthesis Research for Food, Fuel and the Future; Kuang, T., Lu, C., Zhang, L., Eds.; Springer: Berlin, Heidelberg, 2013; pp. 827–833.
- Neori, A.; Haxo, F.T. Effect of Photoadaptation on Action Spectra of Photosynthetic Enhancement in Rhodomonas D3 and in Glenodinium Sp. Photosynthetica (Praha) 1990, 24, 573–585.
- Zhang, Y.-Z.; Li, K.; Qin, B.-Y.; Guo, J.-P.; Zhang, Q.-B.; Zhao, D.-L.; Chen, X.-L.; Gao, J.; Liu, L.-N.; Zhao, L.-S. Structure of Cryptophyte Photosystem II–Light-Harvesting Antennae Supercomplex. Nat Commun 2024, 15, 4999. [CrossRef]
- Siefermann-Harms, D. Carotenoids in Photosynthesis. I. Location in Photosynthetic Membranes and Light-Harvesting Function. Biochimica et Biophysica Acta (BBA) - Reviews on Bioenergetics 1985, 811, 325–355. [CrossRef]
- Kaňa, R.; Kotabová, E.; Sobotka, R.; Prášil, O. Non-Photochemical Quenching in Cryptophyte Alga Rhodomonas Salina Is Located in Chlorophyll a/c Antennae. PLOS ONE 2012, 7, e29700. [CrossRef]
- Staleva, H.; Komenda, J.; Shukla, M.K.; Šlouf, V.; Kaňa, R.; Polívka, T.; Sobotka, R. Mechanism of Photoprotection in the Cyanobacterial Ancestor of Plant Antenna Proteins. Nat Chem Biol 2015, 11, 287–291. [CrossRef]
- Prézelin, B.B.; Alberte, R.S. Photosynthetic Characteristics and Organization of Chlorophyll in Marine Dinoflagellates. Proceedings of the National Academy of Sciences 1978, 75, 1801–1804. [CrossRef]
- Herbert, S.K.; Fork, D.C.; Malkin, S. Photoacoustic Measurements in Vivo of Energy Storage by Cyclic Electron Flow in Algae and Higher Plants 1. Plant Physiology 1990, 94, 926–934. [CrossRef]
- Larkum, A.W.D.; Grossman, A.R.; Raven, J.A. Photosynthesis in Algae: Biochemical and Physiological Mechanisms; Advances in Photosynthesis and Respiration; Springer International Publishing: Cham, 2020; Vol. 45; ISBN 978-3-030-33396-6.
- Johnsen, G.; Sakshaug, E. Biooptical Characteristics of PSII and PSI in 33 Species (13 Pigment Groups) of Marine Phytoplankton, and the Relevance for Pulse-Amplitude-Modulated and Fast-Repetition-Rate Fluorometry. Journal of Phycology 2007, 43, 1236–1251. [CrossRef]
- Johnsen, G.; Sakshaug, E. Bio-Optical Characteristics and Photoadaptive Responses in the Toxic and Bloom-Forming Dinoflagellates Gyrodinium aureolum, Gymnodinium galatheanum, and Two Strains of Prorocentrum minimum. Journal of Phycology 1993, 29, 627–642. [CrossRef]
- Zhen, S.; Bugbee, B. Far-Red Photons Have Equivalent Efficiency to Traditional Photosynthetic Photons: Implications for Redefining Photosynthetically Active Radiation. Plant, Cell & Environment 2020, 43, 1259–1272. [CrossRef]
- Oliveira, C.Y.B.; Abreu, J.L.; Santos, E.P.; Matos, Â.P.; Tribuzi, G.; Oliveira, C.D.L.; Veras, B.O.; Bezerra, R.S.; Müller, M.N.; Gálvez, A.O. Light Induces Peridinin and Docosahexaenoic Acid Accumulation in the Dinoflagellate Durusdinium glynnii. Appl Microbiol Biotechnol 2022, 106, 6263–6276. [CrossRef]
- Millie, D.F.; Kirkpatrick, G.J.; Vinyard, B.T. Relating Photosynthetic Pigments and in Vivo Optical Density Spectra to Irradiance for the Florida Red-Tide Dinoflagellate Gymnodinium breve. Marine Ecology Progress Series 1995, 120, 65–75.
- Warner, M.E.; Berry-Lowe, S. Differential Xanthophyll Cycling and Photochemical Activity in Symbiotic Dinoflagellates in Multiple Locations of Three Species of Caribbean Coral. Journal of Experimental Marine Biology and Ecology 2006, 339, 86–95. [CrossRef]
- Vernet, M.; Neori, A.; Haxo, F.T. Spectral Properties and Photosynthetic Action in Red-Tide Populations of Prorocentrum micans and Gonyaulax polyedra. Mar. Biol. 1989, 103, 365–371. [CrossRef]
- Dionisio-sese, M.L. Aquatic Microalgae As Potential Sources Of UV-Screening Compounds. Philippine Journal of Science 2010, 139, 5–16.
- Rastogi, R.P.; Richa; Sinha, R.P.; Singh, S.P.; Häder, D.-P. Photoprotective Compounds from Marine Organisms. Journal of Industrial Microbiology and Biotechnology 2010, 37, 537–558. [CrossRef]
- López-Rosales, L.; Gallardo-Rodríguez, J.J.; Sánchez-Mirón, A.; Cerón-García, M.D.C.; Belarbi, E.H.; García-Camacho, F.; Molina-Grima, E. Simultaneous Effect of Temperature and Irradiance on Growth and Okadaic Acid Production from the Marine Dinoflagellate Prorocentrum belizeanum. Toxins 2014, 6, 229–253. [CrossRef]
- Martín-Girela, I.; Curt, M.D.; Fernández, J. Flashing Light Effects on CO2 Absorption by Microalgae Grown on a Biofilm Photobioreactor. Algal Research 2017, 25, 421–430. [CrossRef]
- Healey, F.P. The Carotenoids of Four Blue-Green Algae. Journal of Phycology 1968, 4, 126–129. [CrossRef]
- Loeblich, A.R., III; Schmidt, R.J.; Sherley, J.L. Scanning Electron Microscopy of Heterocapsa pygmaea Sp. Nov., and Evidence for Polyploidy as a Speciation Mechanism in Dinoflagellates. Journal of Plankton Research 1981, 3, 67–79. [CrossRef]
- Loeblich, A.R. A Seawater Medium for Dinoflagellates and the Nutrition of Cachonina niei. Journal of Phycology 1975, 11, 80–86. [CrossRef]
- Haxo, F.T.; Lewin, R.A.; Lee, K.W.; Li, M.-R. Fine Structure and Pigments of Oscillatoria(Trichodesmium) Aff. thiebautii (Cyanophyta) in Culture. Phycologia 1987, 26, 443–456. [CrossRef]
- McCarthy, J.J.; Carpenter, E.J. Oscillatoria(Trichodesmium) thiebautii (Cyanophyta) in the Central North Atlantic Ocean. Journal of Phycology 1979, 15, 75–82. [CrossRef]
- Duysens, L. Transfer of Excitation Energy in Photosynthesis, 1952.
Figure 1.
Emerson enhancement spectra of seven algae from the Rhodophyte, Chlorophyte, Dinoflagellate, Cryptomonad, and Cyanobacteria divisions.
Figure 1.
Emerson enhancement spectra of seven algae from the Rhodophyte, Chlorophyte, Dinoflagellate, Cryptomonad, and Cyanobacteria divisions.
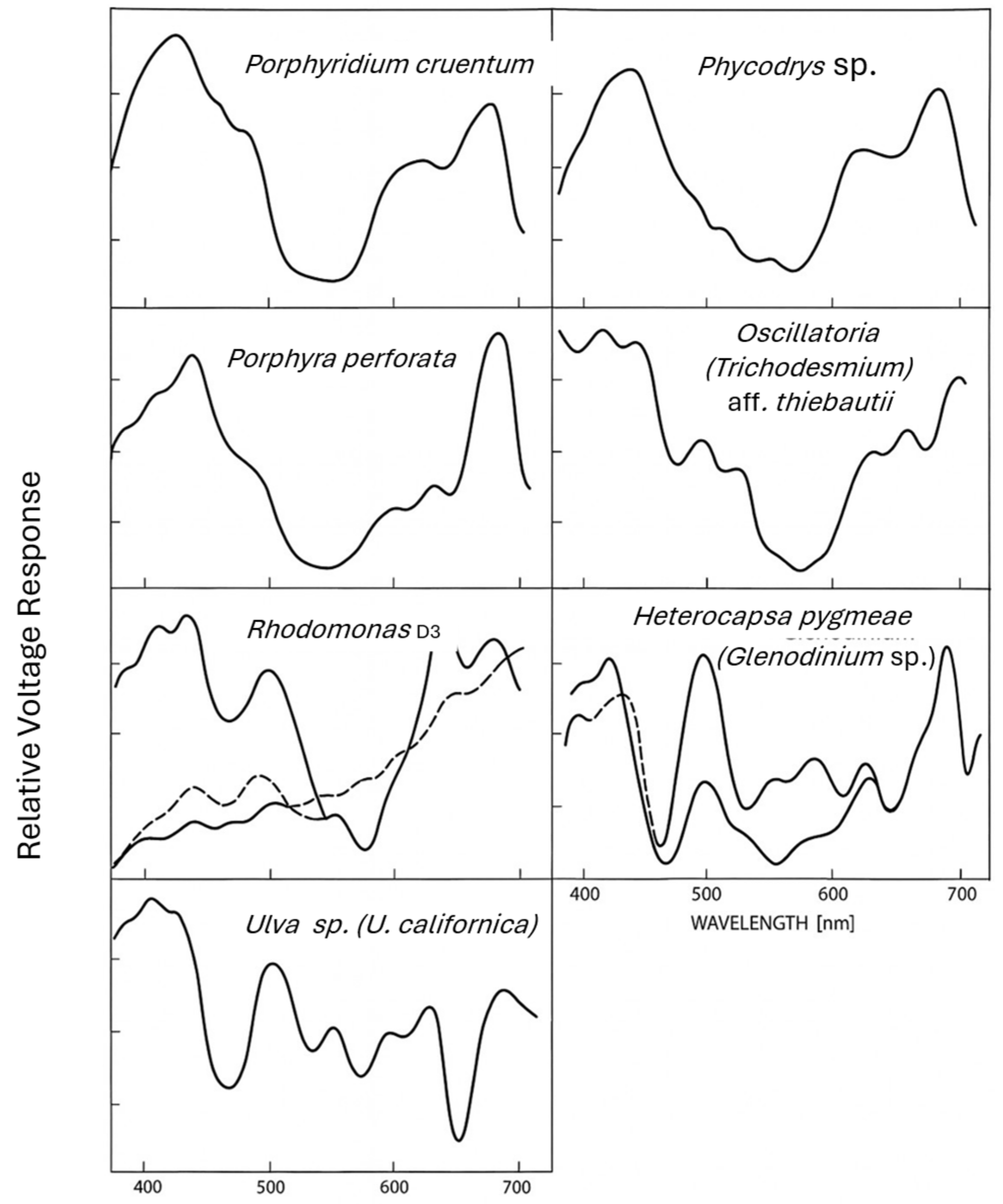
Figure 2.
The custom experimental spectrum measuring system developed for the measurements presented in Figure 1. A. A photograph; B. An outline of the components of the experimental system: 1, a bare platinum oxygen electrode of the Joliot configuration [28,33]; 2, a continuous background light beam; 3, a modulated monochromatic measuring beam light source; 4, a microcomputer-control unit and amplifier; 5, a motor-driven Bausch & Lomb high intensity monochromator; 6, a second motor, also under computer control, rotated a circular, variable-density neutral filter in response to feedback from a silicon photodiode (7), to maintain a constant quantum flux across all wavelengths, through a, beam-chopper wheel (8). The computer automatically acquired the oxygen-evolution signals and recorded their amplitudes as a function of wavelength using an interface to a Commodore 64 computer (9).
Figure 2.
The custom experimental spectrum measuring system developed for the measurements presented in Figure 1. A. A photograph; B. An outline of the components of the experimental system: 1, a bare platinum oxygen electrode of the Joliot configuration [28,33]; 2, a continuous background light beam; 3, a modulated monochromatic measuring beam light source; 4, a microcomputer-control unit and amplifier; 5, a motor-driven Bausch & Lomb high intensity monochromator; 6, a second motor, also under computer control, rotated a circular, variable-density neutral filter in response to feedback from a silicon photodiode (7), to maintain a constant quantum flux across all wavelengths, through a, beam-chopper wheel (8). The computer automatically acquired the oxygen-evolution signals and recorded their amplitudes as a function of wavelength using an interface to a Commodore 64 computer (9).
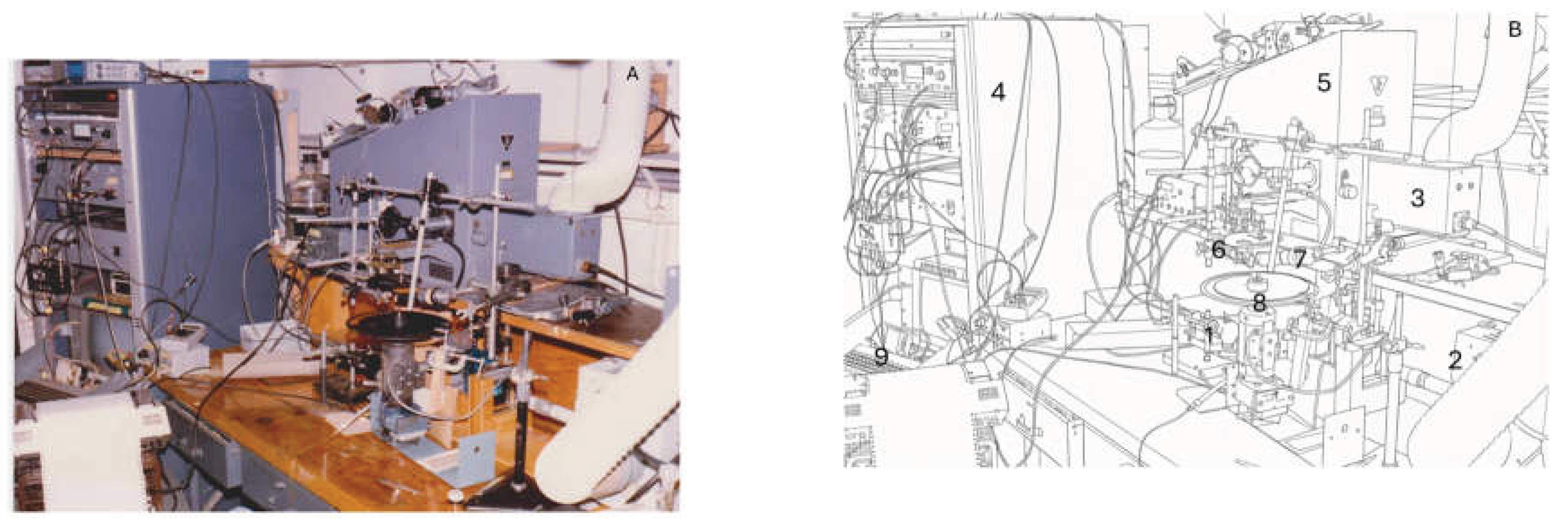
Table 1.
Definitions of the variables used in the study.
| Variable | Definition | Units |
|---|---|---|
| AC | Signal of modulated photosynthetic oxygen evolution resulting from the modulated measuring light beam of intensity Iₘ. | Arbitrary (voltage) |
| Iₘ | Intensity of the modulated measuring radiant beam. | µmol photons·m-²·s-¹ |
| Ib | Intensity of the continuous background radiant beam. | µmol photons·m-²·s-¹ |
| Is | A weak scanning beam | µmol photons·m-²·s-¹ |
| U | Fraction of open Photosystem II reaction centers. | Dimensionless (range: 0-1) |
| Ub | Intensity of a background beam that controls the opening state of Photosystem II reaction centers. | Dimensionless (range: 0-1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



