Submitted:
27 January 2026
Posted:
28 January 2026
You are already at the latest version
Abstract
Toll-like receptors (TLRs) are pattern recognition receptors which links the body’s innate and adaptive immune system, and have recently become central targets in modern vaccine adjuvant design. This review aims to investigate how TLR2, TLR3, TLR4, TLR5, TLR7/8, and TLR9 agonists can enhance antigen presentation and influence T-cell and antibody activity in conventional vaccines, which mostly exhibit low immunogenicity. Computational modeling and simulation studies have already illustrated ligand-receptor binding, signaling pathways, and resulting immune activation across antigen presenting cells, while cases such as MPLA and CpG-ODN, as well as licensed vaccines like Cervarix, Fendrix, and Heplisav-B have already utilized the potential translational application of TLR agonists. But despite these promising progress, inconsistencies between in silico and in vivo findings, species-specific responses, limitations in delivery method, and safety concerns persists. The review essentially highlights the potential of using multi-TLR strategies to support next generation vaccine adjuvant development.
Keywords:
toll-like receptors
; vaccine
; adjuvant
; TLR agonists
; immunogenicity
I. Introduction
1.1. Toll-like Receptors: Mechanisms and Potential as Vaccine Adjuvants
Adjuvants are substances often incorporated into vaccines to enhance and amplify the magnitude and durability of the body’s immune responses to the co-administered antigen in the drug, by specifically activating innate immune pathways that subsequently shape the adaptive immunity [1].
In recent years, Toll-like receptors (TLRs) have emerged as one of the most extensively studied adjuvant targets worldwide. TLRs primarily present a promising capability to mimic pathogen-associated molecular patterns [2,3].
From a mechanistic perspective, TLRs are a family of pattern recognition receptors within the body’s innate immune system which can recognize and detect conserved molecular motifs derived from pathogens. They are a system of proteins that then triggers a cascade of signals which inevitably lead to the activation of transcription factors and result in the production of pro-inflammatory cytokines and the upregulation of co-stimulatory molecules [4,5]. Along with this, TLR agonists are molecules capable of selectively binding to, and activating specific TLRs, thereby mimicking pathogenic signals which then initiate immune responses that ultimately enhance antigen presentation and adaptive immunity [6]. Given these immunostimulatory properties, TLR agonists have progressed as potential candidates for vaccine adjuvant development.
1.2. Innate Immune Signaling as a Determinant of Adaptive Immune Outcomes
The increasing interest in utilizing TLRs as vaccine adjuvants, has exemplified the context and linkage between the innate and adaptive immunity. TLRs simply demonstrate this molecular relationship, as their activation triggers signaling cascades that induce the production of pro-inflammatory cytokines, chemokines, and co-stimulatory molecules which collectively enable effective antigen presentation and lymphocyte priming [4,5]. Dendritic cells also serve as a central link in this process, as their maturation and migration to secondary lymphoid organs enable antigen-specific B and T cell responses, which are essential for long-term immunity [5]. These interactions are particularly relevant to current vaccine design, as the qualitative features of the adaptive response such as antibody isotype, T helper cell polarization, or cytotoxic T-cell activity, are strongly influenced by the innate cues provided during early immune activation [7].
1.3. Molecular Determinants of TLR Activation and Their Immunological Consequences
In immunological research, understanding molecular interactions has increasingly relied on the application of both computational and in silico approaches. Techniques such as molecular docking, molecular dynamics simulations, as well as structure-based modeling and analyses have enabled a detailed characterization of ligand–receptor interaction residues, binding modes, and conformational changes that are often challenging to resolve experimentally [8]. Such approaches not only complement biochemical and immunological studies through systematic comparison, but also provide clear answers to theoretical understandings and facilitate the screening and optimization of novel adjuvant candidates in a cost- and time-efficient manner [8,9].
1.4. Vaccine Development and the Significance of Adjuvants
Globally, vaccines remain as one of the most effective ways to prevent and control certain infectious diseases [10]. Its early success primarily depended largely on live-attenuated or inactive pathogens which could stimulate immune responses. Vaccines train the body’s immune system to recognize them as an intruder without causing the actual illness. In this manner, the body learns to produce the necessary antibodies to inevitably resist the real pathogen when it eventually comes, which can lessen or even prevent the severity [11]. In contrast, modern vaccines have increasingly been developed to use subunit, recombinant, or nucleic acid-based agents, as they incorporate only specific pathogen components rather than the entire organism, thereby generally improving safety. Despite these advantages, these approaches often exhibit limited immunogenicity, particularly when administered alone [11,12]. As such, the inclusion of immune adjuvants are most often necessary in order to enhance and shape antigen-specific immune responses, which essentially prolongs and improves vaccine efficacy as it can help reduce the antigen dose requirement and enable immunity for the wider diversity in a population [11,13].
1.4.1. Progression of Vaccine Adjuvant Development and Innate Immune Understanding
As vaccine design continued to prioritize safety, precision, and efficacy, adjuvant research likewise transitioned from an empiric enhancement towards a more targeted immunomodulation mechanism. Subsequent generations of adjuvants which included emulsions, saponin-based formulations, and nanoparticle systems, were then developed to augment antigen delivery and immunomodulation [14,15]. The characterization of pattern recognition receptors, particularly TLRs, was instrumental in this shift, as it highlighted the importance of molecular pathways that can easily shape the quality of the immune response rather than merely its magnitude [3]. Due to this major conceptual shift, it formally established TLR pathways as potential targets for modulating innate immunity to influence adaptive outcomes, thereby also laying the foundation for contemporary molecular adjuvant platforms, reflecting a shift toward rational design guided by clear mechanistic understandings.
1.4.2. Mechanistic and Clinical Considerations in Next-Generation Vaccine Platforms
Despite the significant conceptual advantages of vaccine formulation and mechanistically guided adjuvant strategies, several challenges continue to complicate their functional translation. Safety remains as a central concern, particularly for TLR agonists and other molecular adjuvants that engage potent innate pathways, as they may induce excessive inflammation or off-target effects when not properly modulated. TLR pathways require careful calibration to prevent excessive inflammation, systemic toxicity, or immunopathology, particularly when agonists are combined or co-formulated with multiple vaccine components [16,17].
Complications also arise from species-dependent receptor expression, ligand-specific signaling biases, or signaling variability, which limits the predictability of successful preclinical findings and create difficulties and inconsistencies in translating computational or in vitro results to clinical outcomes [18,19]. These collective challenges simply justify the need for continuous investigation of mechanistic insights into molecular determinants of TLR activation as well as integrative approaches that bridge molecular analyses, with immunological outcomes as a foundation to support more rational, effective, and feasible adjuvant development.
1.5. Mechanistic and Translational Synthesis of TLR Agonist–Based Adjuvant Research
The lack of understanding and integration between molecular-level binding analyses to functional immunological applications and feasibility motivated the present review to be made, with the aim of providing a comprehensive overview of the molecular binding mechanisms of TLR agonists in the development of immune adjuvants. Drawing on the findings derived from computational modeling and simulation studies, the review summarizes and compares the various classes of TLR agonists, discusses recent advances in elucidating mechanistic ligand–receptor interactions, and highlights key translational challenges hindering clinical implementation, thereby supporting future directions for the rational design of TLR-based adjuvants essential for advancing vaccine efficacy and safety.
II. Molecular and Immunological Basis of Toll-like Receptors and Their Agonists
2.1. Toll-like Receptors (TLRs)
Toll-like receptors (TLRs) are integral membrane pattern recognition receptors (PRRs) and serve as the first line of defense in the innate immune system, enabling the detection of pathogens and initiation of immune responses [20,21]. TLRs serves as a link between innate and adaptive immune responses by recognizing microbial intruders through Pathogen-Associated Molecular Patterns (PAMPs) or inflammation or stress signals derived from the host tissue known as Danger-Associated Molecular Patterns (DAMPs) [20,21,22,23].
- A. Domain Organization and Signaling Mechanisms of Toll-Like Receptors
Figure 1.
Structural Organization of Toll-like Receptors (TLRs). TLRs are single-pass transmembrane proteins belonging to the Interleukin-1 Receptor or Toll-Like Receptor superfamily. Each receptor consists of three major regions: the extracellular leucine-rich repeat (LRR) domain, the transmembrane domain, and the intracellular TIR (Toll-IL-1 receptor) domain. The LRR domain serves as the ligand-binding site, recognizing pathogen-associated molecular patterns (PAMPs) such as bacterial lipoproteins, lipopolysaccharides, viral RNA, or unmethylated CpG DNA. The transmembrane domain anchors the receptor to the plasma membrane or endosomal membrane, facilitating structural integrity and signal relay. Upon ligand binding, the intracellular TIR domain interacts with downstream adaptor proteins including MyD88 and TRIF, which are critical for propagating signal transduction. Within the TIR domain, three conserved amino acid motifs—Box 1, Box 2, and Box 3—mediate these interactions and are indispensable for initiating intracellular signalling cascades [24].
Figure 1.
Structural Organization of Toll-like Receptors (TLRs). TLRs are single-pass transmembrane proteins belonging to the Interleukin-1 Receptor or Toll-Like Receptor superfamily. Each receptor consists of three major regions: the extracellular leucine-rich repeat (LRR) domain, the transmembrane domain, and the intracellular TIR (Toll-IL-1 receptor) domain. The LRR domain serves as the ligand-binding site, recognizing pathogen-associated molecular patterns (PAMPs) such as bacterial lipoproteins, lipopolysaccharides, viral RNA, or unmethylated CpG DNA. The transmembrane domain anchors the receptor to the plasma membrane or endosomal membrane, facilitating structural integrity and signal relay. Upon ligand binding, the intracellular TIR domain interacts with downstream adaptor proteins including MyD88 and TRIF, which are critical for propagating signal transduction. Within the TIR domain, three conserved amino acid motifs—Box 1, Box 2, and Box 3—mediate these interactions and are indispensable for initiating intracellular signalling cascades [24].
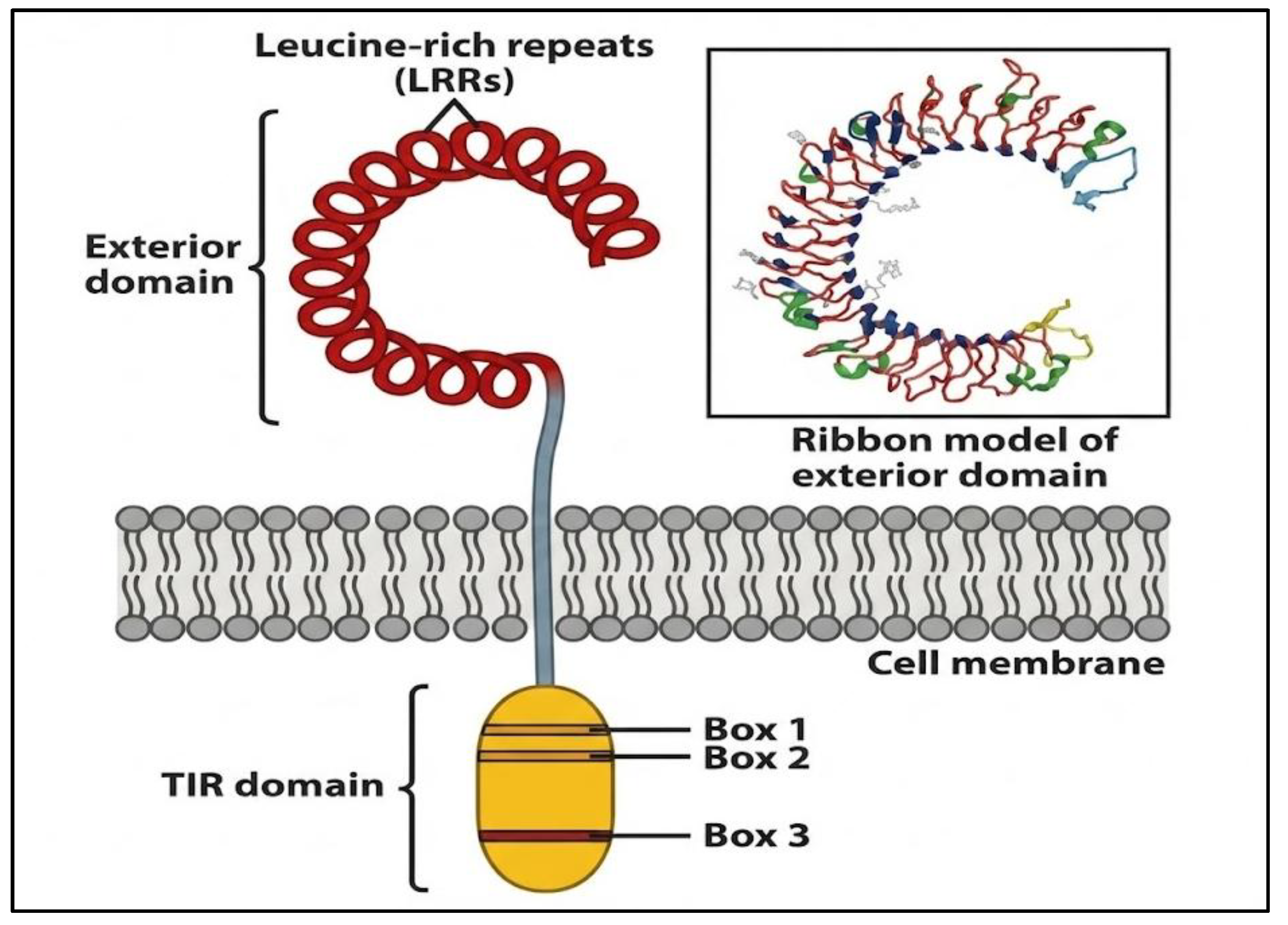
- B. Divergent Signaling Pathways of TLRs: MyD88 vs. TRIF
The Myeloid Differentiation Primary Response 88 (MyD88)-dependent pathway is utilized by all TLRs except TLR3, whereas TLR4 can engage in both MyD88 and TIR-domain-containing Adapter-Inducing Interferon-β (TRIF) [25,26,27]. Ligand binding and TLR dimerization recruit MyD88 to the TIR domain, initiating a kinase cascade in which MyD88 interacts with Interleukin-1 Receptor-Associated Kinases (IRAKs), particularly recruiting IRAK-4 via death domain interactions. IRAK-4 autophosphorylates and activates IRAK-1 or IRAK-2, which then recruits Tumor Necrosis Factor Receptor-Associated Factor 6 (TRAF6) [28]. This complex activates the IκB Kinase (IKK) complex, leading to phosphorylation and degradation of the inhibitory protein IκBα, allowing the transcription factor called Nuclear Factor kappa B (NF-κB) to enter the nucleus and induce proinflammatory cytokines such as Interleukin-6 (IL-6), Tumor Necrosis Factor alpha (TNF-α), and Macrophage Inflammatory Protein-2 (MIP-2). Negative regulators like the Regulator of Calcineurin 1 (RCAN1) limit IκBα phosphorylation to prevent excessive inflammation [27].
In contrast, the TRIF-dependent pathway, triggered by TLR3 and TLR4, recruits TRIF, which activates TANK-Binding Kinase 1 (TBK1) and IKKε, leading to phosphorylation of Interferon Regulatory Factors 3 and 7 (IRF3/7). These factors translocate to the nucleus to induce Type I Interferons (IFN-α/β) and chemokines such as Regulated on Activation, Normal T-cell Expressed and Secreted (RANTES) and Interferon gamma-Induced Protein 10 (IP-10) [25,26,27,28]. Unlike the early-phase MyD88 response, TRIF mediates slower, late-phase NF-κB activation. RCAN1 enhances this branch by promoting IRF7 expression, opposite to its inhibitory role in MyD88 signaling [27]. Mechanistically, these pathways can be antagonistic; for example, in autoimmune diabetes, TRIF signaling induces the immunosuppressive enzyme Indoleamine 2,3-Dioxygenase (IDO), counteracting MyD88-driven pathogenic inflammation [26].
2.2. Toll-like Receptor Agonists
TLR agonist is a natural or synthetic substance that is intended to bind to and activate TLRs [20,22,29]. Despite the nature of TLRs naturally sensing microbial components called PAMPs, researchers use these agonists as immunomodulatory instruments to intentionally activate certain immune pathways [20,23]. Through imitation of the molecular signature of infection, these substances enhance immune preparedness and response specificity against pathogens or abnormal cells [23].
2.2.1. TLR Agonists’ Molecular Activation and Mechanism
The activation of TLRs is initiated when an agonist binds to the extracellular leucine-rich repeat (LRR) domain, promoting the dimerization of two receptor subunits, which functions as a molecular “switch” to convert ligand recognition into intracellular signaling [20,21]. Upon dimerization, the cytoplasmic Toll/Interleukin-1 receptor (TIR) domains recruit specific adaptor proteins, primarily MyD88, which mediates signaling for all TLRs except TLR3, or TRIF, which is utilized by TLR3 and TLR4 [20,21,23]. These interactions subsequently facilitate the assembly of Supramolecular Organizing Centres (SMOCs), such as myddosomes, which serve as massive, organized signaling platforms that amplify and coordinate downstream signal transduction [30].
Through SMOCs, the receptor complex ensures a “digital” or all-or-nothing response, whereby even minimal pathogen detection triggers full activation of the immune signaling cascade, leading to the strong activation of transcription factors including NF-κB and interferon regulatory factors (IRFs) [21,23]. Therefore, these transcription factors regulate the expression of pro-inflammatory cytokines, such as TNF-α and IL-6, and type I interferons, including IFN-α and IFN-β, that orchestrates a rapid and coordinated innate immune response capable of eliminating the pathogen efficiently [21,23,31]
2.3. TLRs in the Immune System
TLRs in the immune system represent an important connecting point between innate and adaptive immunity by translating pathogen recognition into coordinated immune activation. Immune cells such as dendritic cells, macrophages, and B cells, contain TLRs that have a significant role in the immune response as its goal is to have a decisive and rapid response [30].
2.3.1. TLRs on Dendritic Cells (DCs)
Dendritic cells represent a vital component of the immune system as they are the major antigen-presenting cells that detect pathogens and coordinate the activation of T cells, bridging innate and adaptive immunity. DC subtypes are distinguished by different TLRs they produce and display. DCs in humans mostly display TLR1, TLR2, TLR4, TLR8, and TLR9, while plasmacytoid DCs (pDCs) mainly produce and display TLR7 and TLR9. This permits the cells to recognize viral nucleic acids efficiently and to produce type I interferons in large quantities [32].
TLR engagement in dendritic cells drives maturation, characterized by upregulation of co-stimulatory molecules (CD80 and CD86) necessary for T-cell activation. TLR signaling also promotes endosomal acidification, thereby facilitating antigen processing and cross-presentation to cytotoxic T cells, which is a crucial process in antiviral and antitumor immunity [32].
2.3.2. TLRs on Macrophages
Macrophages serve as the main innate immune cells that detect the pathogen components through TLRs and quickly respond to the stimuli by ramping up the phagocytosis and reactive oxygen species (ROS) production among other activities. Furthermore, exposure to some TLR agonists can lead to the establishment of trained immunity which is an epigenetically-driven innate immune memory that enables the macrophages to mount a stronger secondary response to the same or different microbe long after the initial stimulus [30,33].
2.3.3. TLRs on B-Cells
B cells are a type of white blood cell that play a central role in the adaptive immune system by producing antibodies to recognize and neutralize pathogens. They also act as antigen-presenting cells and contribute to immune memory, helping the body respond more effectively to future infections [34]. B lymphocytes employ TLRs to modulate their cell growth and ability to secrete antibodies. In contrast to many immune cells, B lymphocytes exposed to TLR ligand signal predominantly through the MyD88-dependent pathway often results in rapid proliferation and differentiation of the cells. Beyond conventional B-cell responses, a minor group of innate-like B cells utilizes TLRs to detect commensal microbiota-derived cues, thus granting the first line of defense against pathogens like influenza viruses and certain bacteria [35].
2.4. Host-Dependent Modulation of Toll-like Receptor Activity
Biological sex significantly dictates TLR-mediated responses through the interplay of genetic and hormonal factors [36,37,38]. Based on the study conducted in Rome, Italy, the findings state that females generally exhibit more robust TLR activation because the X chromosome carries multiple genes involved in the immune response, including members of the TLR family [37]. Furthermore, sex hormones act as potent modulators; estradiol typically enhances TLR expression and pro-inflammatory cytokine production, whereas androgens like testosterone exert immunosuppressive effects by reducing TLR4 expression in macrophages and other immune cells. This sexual dimorphism is further shaped by the "microgenderome", where sex-specific gut microbiota compositions interact with host hormones to influence the stimulation of pattern recognition receptors [36,37].
The host's age and overall immune status, particularly in the context of chronic conditions like obesity, further refine TLR responsiveness [37,38]. Ageing is associated with immunosenescence, a process that deregulates the immune system as sex hormone concentrations and receptor signalling decline over time. This often leads to a diminished innate response to TLR ligands, which can impair the effectiveness of vaccinations in older populations. A study conducted in Madrid, Spain, presented results that states specific observations in which the chronic low-grade inflammation characteristic of obesity significantly impacts TLR2 and TLR4 expression patterns; specifically, while obese individuals may show elevated fasting TLR2 gene expression, they often exhibit reduced surface expression of TLR4 [36,37]. This reduction in TLR4 surface levels may represent a compensatory mechanism, akin to endotoxin tolerance, where the system attempts to prevent excessive postprandial inflammation triggered by persistent nutrient sensing of excessive lipids or glucose as "danger signals" [38].
2.5. Specific Involvement of TLR Agonists on Vaccine Development
The transition from conventional whole-pathogen vaccines to newer subunit-based formulas has drastically made vaccine developers rely on very effective adjuvants to evoke the desired immune response [23,29]. TLR agonists are one type of adjuvants that enhance immunogenicity by orchestrating different cells of the immune system to react to the presence of the pathogen, resulting in an innate and adaptive immune activation in response to antigen exposure [29].
In specific vaccines like HPV vaccine Cervarix and the hepatitis B vaccine Fendrix, the TLR4 agonist monophosphoryl lipid A (MPL) is used as part of the AS04 adjuvant to enhance immune responses, while the hepatitis B vaccine Heplisav-B incorporates the TLR9 agonist CpG-1018, allowing for a simplified two-dose immunization schedule [29]. TLR5 agonists like flagellin function as potent vaccine adjuvants by maturing lung antigen-presenting cells and upregulating the homing receptor CCR7 to drive their migration to draining lymph nodes. These agonists effectively overcome neonatal immune hyporesponsiveness by correcting endosomal defects and promoting robust antigen cross-presentation, as demonstrated by research on lung mucosal cells conducted at Boston Children's Hospital and Harvard Medical School [39]
In specific cases like outer membrane vesicles (OMVs), a bacterial-derived nanoparticle that displays multiple PAMPs have enabled simultaneous engagement of several TLRs. This "self-adjuvanticity" allows OMVs to generate strong humoral and cell-mediated immune responses without additional adjuvants [35]. In the form of generalized modules for membrane antigens (GMMA), OMVs have emerged as a promising next-generation vaccine platform, as demonstrated by the Phase 1 clinical trial of the GMMA-based Shigella sonnei 1790GAHB vaccine in healthy adults in Europe, which showed an acceptable safety profile and robust immunogenicity following intramuscular administration [35].

Table 1.
Summary of Selected Vaccines Employing Toll-Like Receptor (TLR)–Based Adjuvants and Their Immunological Effects.
Table 1.
Summary of Selected Vaccines Employing Toll-Like Receptor (TLR)–Based Adjuvants and Their Immunological Effects.
| Vaccine | Target Pathogen | TLR Agonist / Adjuvant | Formulation | Key Findings |
| Cervarix | HPV | TLR4 agonist: MPL | AS04 adjuvant | Enhanced immune response |
| Fendrix | Hepatitis B | TLR4 agonist: MPL | AS04 adjuvant | Enhanced immune response |
| Experimental Mucosal Vaccine | DQ-OVA (respiratory model antigen) | TLR5 agonist: Flagellin (FLA-ST) | Intranasal protein subunit vaccine | Strong activation of neonatal lung APCs |
| Heplisav-B | Hepatitis B | TLR9 agonist: CpG-1018 | Subunit vaccine | Enables two-dose schedule |
| 1790GAHB | Shigella sonnei | OMVs (GMMA) with inherent TLR agonists | OMV-based nanoparticle vaccine | Phase 1 trial: safe, strong humoral & cellular responses |
2.6. Nanotechnology and Formulation Strategies in TLR-Based Vaccines
Specific challenges regarding TLR-based vaccines are compounded by species-specific differences in TLR expression—such as variations between murine and human TLR8—which frequently hinder the direct translation of preclinical results to human clinical applications [40,41]. For instance, clinical evaluations of the TLR9 agonists IMO2055 and CPG7909 in patients with non-small-cell lung cancer (NSCLS) and head and neck cancer must be observed with caution due to their significant safety setbacks, highlighting the risks of systemic inflammatory side effects [41].
To enhance therapeutic efficacy while minimizing toxicity, there is a critical need for controlled release and targeted delivery using platforms like nanoparticles, liposomes, hydrogels, and stimuli-responsive polymeric particles [40,41,42]. These systems are essential because small-molecule ligands often have a short in vivo lifespan and suffer from rapid dispersion, causing undesirable systemic dissemination rather than the necessary localized stimulation [40,41]. Lipidation strategies, such as the 18-C fatty acyl chain used in 3M-052, improve bio-localization and bioavailability at the immunization site while reducing systemic spread [40].
III. Systematic Classification of Toll-like Receptors Relevant to Immune Adjuvant Development
3.1. Receptor Family: Pattern Recognition Receptors (PRRs)
TLRs are a major subgroup of pattern recognition receptors (PRRs) centering its role in innate immunity by detecting conserved microbial components and initiating immune signaling pathways. PRRs are known as germline-encoded receptors that are mainly expressed by innate immune cells and have the ability to recognize pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs), enabling rapid immune activation. Among PRRs, TLRs are the most extensively studied due to their well-defined signaling pathways and strong ability to shape adaptive immunity, which is essential for effective vaccine responses [43].
3.1.1. Receptor Subgroup: Toll-like Receptors (TLRs)
TLRs are characterized by extracellular leucine-rich repeat (LRR) domains known for its role in ligand recognition. Based on their cellular localization, TLRs are broadly divided into cell-surface TLRs which are considered critical in enumerating the type of immune response induced and the suitability of each receptor as an adjuvant target [1,30].
3.1.1.1. Structural and Functional Evolution of Toll-like Receptors
TLRs are known to be one of the the most ancient components of the innate immune system, with origins traceable to early metazoans and genes being found across both invertebrates and vertebrates, indicating a deep evolutionary origin of pathogen detection systems [44]. This detection system predates the divergence of major animal lineages, supporting the idea that the TLR–NF-κB signaling module represents one of the earliest immune recognition systems in animals, conserved from sponges and cnidarians to mammals [44].
In invertebrate protostomes (e.g., arthropods and mollusks), TLR homologs frequently exhibit multiple cysteine clusters (mccTLRs) in their extracellular leucine-rich repeat (LRR) domains, a structure thought to reflect an ancestral domain architecture that may have enabled broader or more flexible ligand interactions in simpler immune contexts [45].
In contrast, vertebrate TLRs predominantly possess single cysteine clusters (sccTLRs), a domain configuration associated with more specialized pathogen recognition and signaling roles typical of vertebrate innate immunity; this distinction suggests that the sccTLR form became prominent along the deuterostome lineage leading to vertebrates, potentially as a refinement of TLR function alongside the emergence of adaptive immunity [46].
3.2. TLRs’ Ligand Recognition and Structural Characteristics
The ligand-binding specificity of TLRs is pre-determined in accordance to their LRR ectodomains, forming a structure optimized for distinct molecular patterns. TLR4 requires the accessory protein MD-2 for effective ligand binding, while endosomal TLRs undergo conformational changes upon ligand recognition in acidic compartments. These structural features influence binding affinity and signaling strength, which are key considerations in the rational design of TLR agonists as vaccine adjuvants [47,48].
3.3. Classification of TLRs and Their Agonists
Researchers have further subclassified the group of agonists according to the particular TLR they activate, thus enabling an immunological response of desired nature [20,23,29]. TLR2 agonists such as synthetic Pam3CSK4 or the naturally-occurring MALP-2, imitate bacterial lipopeptides to recruit professional immune cells [32,33]. TLR3 agonists, such as Poly I:C and its safer synthetic derivatives known as Ampligen or ARNAX, act like viral double-stranded RNA, thereby eliciting strong antiviral and T helper 1 (Th1)–type immune responses. TLR4 activation is commonly achieved using a detoxified lipopolysaccharide derivative called monophosphoryl lipid A (MPLA) that provides potent immune stimulation with minimal toxicity. TLR5 recognizes bacterial flagellin, a highly conserved structural protein of bacterial flagella, and its activation promotes robust NF-κB signaling and cytokine production, particularly at mucosal surfaces and has been extensively explored as both a standalone adjuvant and a fusion partner in recombinant vaccines [49]. Other prominent agents include TLR7 or 8 agonists like Imiquimod and Resiquimod (R848) are able to detect viral RNA, while TLR9 agonists known as CpG oligodeoxynucleotides can also detect viral RNA and simulate microbial DNA thereby resulting in elevated levels of protective interferons [33].
3.3.1. Agonist-Specific TLR Signaling and Downstream Immunological Effects
TLRs consist of functional differences that determine why combinations of TLR agonists are increasingly being explored to achieve synergistic immune activation while minimizing adverse effects. TLR4 activation induces robust pro-inflammatory cytokine production but can be limited to having higher reactogenicity, whereas endosomal TLRs such as TLR7/8 and TLR9 tend to promote antiviral and Th1-biased immune responses with improved safety profiles [50].
Table 2.
Functional Framework and Characterization of Toll-Like Receptors: Agonists, Adjuvant Immune Responses, Recognition and Signaling Components.
Table 2.
Functional Framework and Characterization of Toll-Like Receptors: Agonists, Adjuvant Immune Responses, Recognition and Signaling Components.
| TLR | Agonist | Key Signaling Adaptors | Primary Ligands | Functional Role in Adjuvanticity | Dominant Immune Responses |
| TLR2 | Pam3CSK4 (synthetic), MALP-2 (natural) | MyD88-dependent pathway |
Bacterial lipoproteins and lipopeptides | Enhances innate immune activation and antigen presentation | Pro-inflammatory cytokine production |
| TLR3 | Poly I:C, Ampligen, ARNAX | TRIF-dependent pathway | Double-stranded RNA (dsRNA) | Promotes robust antiviral immunity | Strong type I interferon (IFN-α/β) production |
| TLR4 | Monophosphoryl lipid A (MPLA) | MyD88-dependent pathway | Single-stranded RNA (ssRNA) | Potent immune activation; high adjuvant efficacy | Robust pro-inflammatory cytokine production |
| TLR5 | Flagellin (natural or recombinant) | MyD88-dependent pathway |
Bacterial flagellin | Promotes DC maturation and | Pro-inflammatory cytokine production |
| TLR7/8 | Imiquimod, Resiquimod (R848) | MyD88-dependent pathway | Single-stranded RNA (ssRNA) | Enhancement of antiviral immunity | Strong antiviral responses; type I IFN production |
| TLR 9 | CpG oligodeoxynucleotides | MyD88-dependent pathway | Single-stranded RNA (ssRNA) | Promotes cellular and humoral immune responses | Th1-biased immunity; B-cell activation and antibody enhancement |
IV. Spatial and Cellular Distribution of Toll-like Receptors
4.1. Tissue- and Cell-Specific Distribution of Toll-like Receptors
TLRs are distributed across different tissues with regards to their function and localization. TLRs are broadly distributed in barrier tissues (such as skin, lungs, and gastrointestinal tract) and lymphoid organs (including the spleen and lymph nodes), reflecting their roles in early pathogen detection, antiviral immunity, and systemic inflammatory responses [51,52,53]. Beyond these immune cells, TLRs are also functionally exhibited in non-immune compartments, such as epithelial cells, endothelial cells, and even neurons. Their biological importance include aiding in barrier defense, enabling both inflammatory signaling and leukocyte recruitment, and acting on neuroinflammatory and host-pathogen interactions, respectively [54].
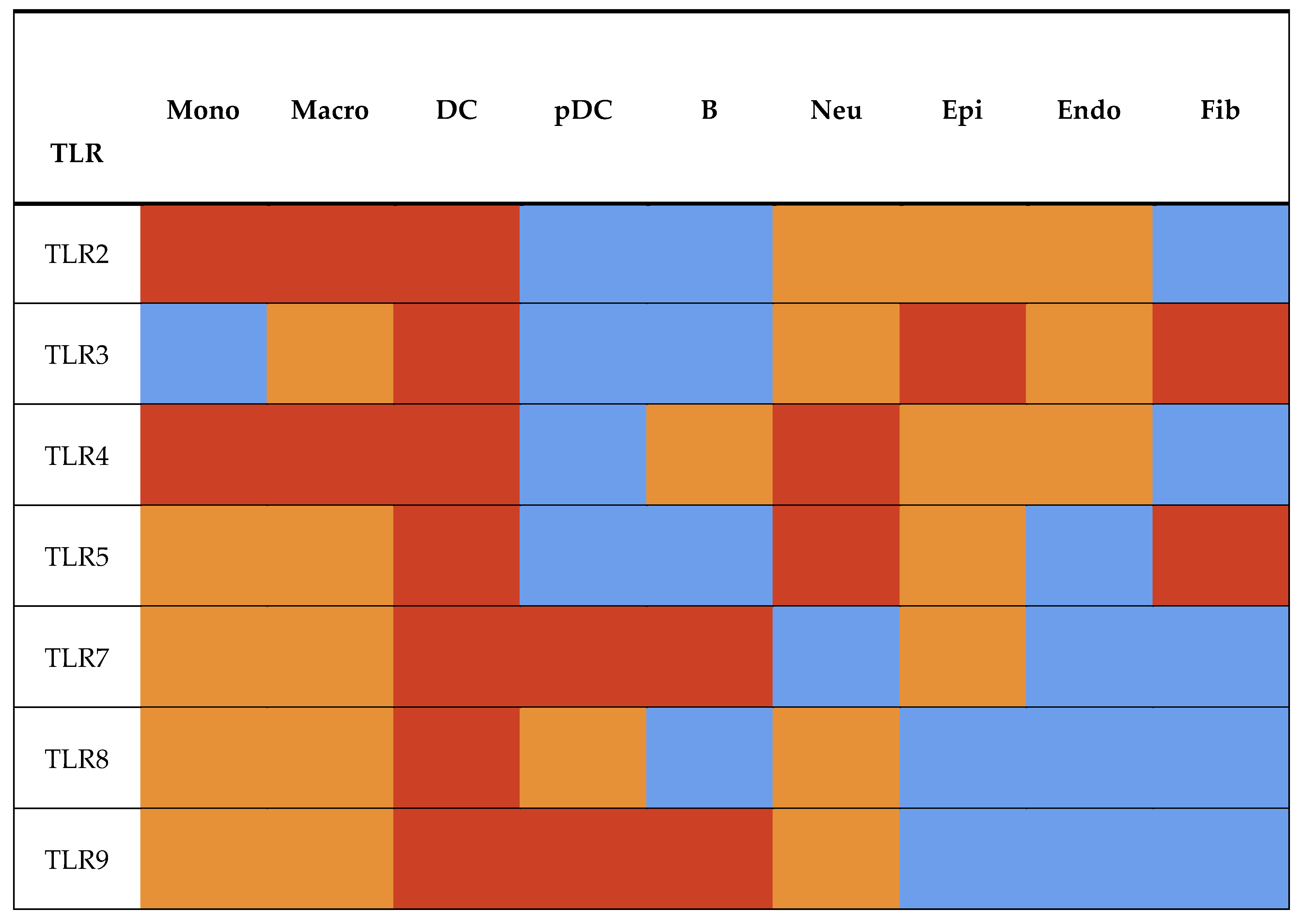
Figure 2.
Heatmap of the Spatial Distribution of TLRs. Cell type abbreviations are as follows: Mono, monocytes; Macro, macrophages; DC, dendritic cells; pDC, plasmacytoid dendritic cells; B, B cells; Neu, neutrophils; Epi, epithelial cells; Endo, endothelial cells; Fib, fibroblasts. Heatmap color intensity represents relative receptor expression, with red indicating high, orange moderate, and blue low expression.
Figure 2.
Heatmap of the Spatial Distribution of TLRs. Cell type abbreviations are as follows: Mono, monocytes; Macro, macrophages; DC, dendritic cells; pDC, plasmacytoid dendritic cells; B, B cells; Neu, neutrophils; Epi, epithelial cells; Endo, endothelial cells; Fib, fibroblasts. Heatmap color intensity represents relative receptor expression, with red indicating high, orange moderate, and blue low expression.
4.1.1. Structural Divergence of mccTLRs and sccTLRs and Its Impact on Tissue-Specific TLR Expression
TLRs are historically classified into two groups based on the physical arrangement of cysteine clusters in their LRR regions: multiple cysteine cluster (mccTLRs), which are primarily ancestral and found in invertebrates, displaying broader but less specialized tissue expression patterns, and single cysteine cluster (sccTLRs), which are predominantly expressed on dendritic cells, macrophages, B cells, and epithelial surfaces of barrier tissues such as the gut and lungs, reflecting their evolved specialization for detecting extracellular and endosomal PAMPs [20,21,30]. Beyond structural variation, this evolutionary divergence is mirrored by distinct patterns of intracellular localization that shape how TLRs encounter and discriminate among microbial ligands. Cell surface TLRs sense microbial exterior components like lipids and proteins at the plasma membrane [30,33,35]. Conversely, endosomal TLRs are isolated inside internal compartments to scan for viral and bacterial nucleic acids [21,23,35].
Two major structural types of TLRs are recognized: single cysteine cluster TLRs (sccTLRs) and multiple cysteine cluster TLRs (mccTLRs). sccTLRs are defined by a single cysteine cluster at the C-terminal end of the leucine-rich repeats adjacent to the plasma membrane, whereas mccTLRs possess two or more cysteine clusters typically including two LRRCTs and often an additional N-terminal cysteine cluster [20,44]. The structural divergence of sccTLRs and mccTLRs showcases an early evolutionary split with mccTLRs likely representing the ancestral form and sccTLRs dominating in vertebrates. The predominance of sccTLRs in vertebrates may have facilitated immune evolution by providing a conserved signaling framework while allowing adaptive diversification in ligand recognition, which enabled vertebrates to efficiently sense and respond to a wide range of pathogens [46].
4.2. TLR Expression in Immune Cells
TLRs are localized within various compartments to recognize distinct microbial components. The innate immune system relies on these receptors contributing to the rapid and early detection of invading pathogens. Among these TLRs, TLR2 is mostly expressed by myeloid immune cells which includes monocytes, macrophages, dendritic cells, and neutrophils, and is also expressed on non-immune cells, especially epithelial and endothelial cells, where it mediates fast inflammation to microbial ligands [55]. TLR3 is expressed on dendritic cell subsets and a large number of non-immune cells, such as epithelial cells, fibroblasts, and endothelial cells, allowing broad detection of viral double-stranded RNA and triggering of the interferon anti-viral response [56].
TLR4 is surface-expressed on macrophages and dendritic cells that selectively identifies lipid A, an active component within the lipopolysaccharide (LPS) of Gram-negative bacteria [53]. TLR5 is expressed in the mucosal compartment including a subset of dendritic cells in the intestine where its activation by bacterial flagellin improves the integrity of the intestine and immune signaling [57]. TLR7 and TLR8 are expressed in the endosomal compartments of monocytes and plasmacytoid dendritic cells, respectively [53]. These receptors are responsible for sensing single-stranded RNA from viral pathogens. TLR9 is expressed on B cells and plasmacytoid dendritic cells that enables the recognition of unmethylated CpG sequences present in the DNA of bacteria and viruses [54].
Table 3.
TLRs’ Structural Classification and Spatial-Functional Organization.
| TLR | Historical Group | Tissue and Organ Distribution | Dominant Cellular Distribution | Cellular Lineage | Function |
| TLR2 | single cysteine cluster (sccTLRs) | Barrier tissues; lymphoid organs | Myeloid cells; barrier-associated non-immune cells | Immune and Non-immune Cells | Senses bacterial lipoproteins, resulting in pro-inflammatory innate signaling |
| TLR3 | single cysteine cluster (sccTLRs) | Barrier and lymphoid tissues | Dendritic cells; stromal cells | Immune and Non-immune cells | Detects dsRNA, leading to antiviral/type I IFN responses |
| TLR4 | single cysteine cluster (sccTLRs) | Systemic organs | Myeloid antigen-presenting cells | Immune and Non-immune cells | Functions in systemic inflammatory responses |
| TLR5 | single cysteine cluster (sccTLRs) | Barrier tissues; mucosal surfaces | Monocytes; dendritic cells; epithelial cells | Immune and non-immune cells | Activation of innate antibacterial immunity |
| TLR7/8 | single cysteine cluster (sccTLRs) | Lymphoid and mucosal tissues | pDCs, B cells; and myeloid cells | Immune cells | Initiates antiviral immune responses |
| TLR9 | single cysteine cluster (sccTLRs) | Hematopoietic and lymphoid tissues | B cells; plasmacytoid DCs | Immune cells | Regulates both the hematopoietic and immune systems |
4.3. Precision Immunization Through TLR Subtype Targeting
The classification of TLRs into cell surface vs endosomal receptors has direct clinical relevance for selecting adjuvants that best match the pathogen type being targeted by a vaccine. Cell surface TLRs predominantly recognize bacterial cell wall components such as lipoproteins and lipopolysaccharides, making agonists targeting these receptors suitable adjuvants for bacterial subunit vaccines [40].
Conversely, endosomal TLRs recognize nucleic acid motifs commonly found in viruses, and agonists for these TLRs have been shown to enhance both humoral and cellular responses in viral vaccine formulations by promoting stronger type I interferon and antigen-presenting cell activation, thereby tailoring the innate signals to the biology of the pathogen and increasing vaccine efficacy [40].
Beyond pathogen-targeted adjuvant choice, host genetic variation in TLR genes has emerged as a potential factor for personalized adjuvant design. Polymorphisms in TLR genes, especially in TLR4, have been associated with differences in vaccine immune responses, suggesting that some individuals may respond suboptimally to certain adjuvanted vaccines due to their inherent TLR genotype, which affects innate signaling thresholds and cytokine production [58]. Recent studies have circulated that the associations between TLR7 and TLR8 polymorphisms and variability in antiviral vaccine responses underscores the role of host TLR genetics in shaping vaccine efficacy and providing a rationale for precision adjuvant strategies that match an individual’s innate sensing profile to the most efficacious TLR agonist adjuvant [59].
V. Translational Applications and Strategies of TLR Agonists in Vaccine Development and Immune Regulation
5.1. Trends in Increasing Use of TLR Agonists in Adjuvant Study
The use of TLR agonists as vaccine and therapeutic adjuvants has increased substantially in recent years with their ability to potentiate innate and adaptive immunity while ensuring disease-specific responses. Numerous clinical trials involving TLR-based adjuvant for infectious diseases and cancer have demonstrated that TLR agonists produce innate immune responses against specific pathogens and malignancies by functioning as highly potent activators of the innate immunity system [60].
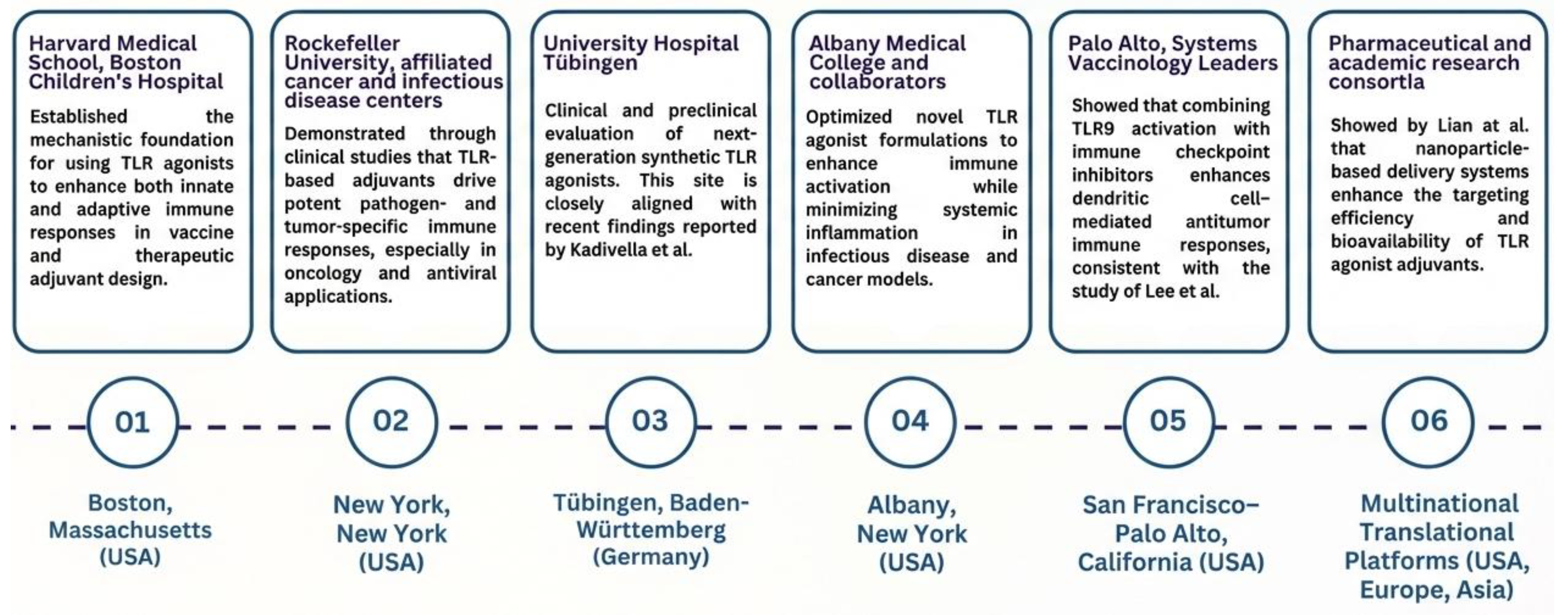
Figure 3.
Emerging Trends in the Application of TLR Agonists as Vaccine and Therapeutic Adjuvants. The horizontal timeline highlights a few major global and national research hubs that has majorly contributed to driving the growing adoption of TLR agonists as vaccine and therapeutic adjuvants. Each node represents a city or state where pivotal studies have advanced both preclinical and clinical applications. Research in Boston, MA (USA) established the mechanistic foundation for using TLR agonists to enhance innate and adaptive immunity. In New York, NY (USA), clinical studies demonstrated pathogen- and tumor-specific immune activation through TLR-based adjuvants. Tübingen (Germany) contributed synthetic TLR4 agonists with improved specificity and reduced toxicity, as discussed by Kadivella and co-authors in 2025 [9]. Preclinical work in Albany, NY (USA) optimized formulations to maximize immune activation while minimizing systemic inflammation. Studies from Seoul, Republic of Korea, particularly the study of Lee alongside co-authors in 2024, showed that combining TLR9 activation with checkpoint inhibitors enhances dendritic cell–mediated antitumor responses [61]. Finally, nanoparticle-based delivery systems developed by Lian et al. improved targeting efficiency and bioavailability of TLR agonists, supporting translational and next-generation immunotherapies. Collectively, these hubs reflect the trend of increasing research activity and innovation in TLR agonist adjuvant development [62].
Figure 3.
Emerging Trends in the Application of TLR Agonists as Vaccine and Therapeutic Adjuvants. The horizontal timeline highlights a few major global and national research hubs that has majorly contributed to driving the growing adoption of TLR agonists as vaccine and therapeutic adjuvants. Each node represents a city or state where pivotal studies have advanced both preclinical and clinical applications. Research in Boston, MA (USA) established the mechanistic foundation for using TLR agonists to enhance innate and adaptive immunity. In New York, NY (USA), clinical studies demonstrated pathogen- and tumor-specific immune activation through TLR-based adjuvants. Tübingen (Germany) contributed synthetic TLR4 agonists with improved specificity and reduced toxicity, as discussed by Kadivella and co-authors in 2025 [9]. Preclinical work in Albany, NY (USA) optimized formulations to maximize immune activation while minimizing systemic inflammation. Studies from Seoul, Republic of Korea, particularly the study of Lee alongside co-authors in 2024, showed that combining TLR9 activation with checkpoint inhibitors enhances dendritic cell–mediated antitumor responses [61]. Finally, nanoparticle-based delivery systems developed by Lian et al. improved targeting efficiency and bioavailability of TLR agonists, supporting translational and next-generation immunotherapies. Collectively, these hubs reflect the trend of increasing research activity and innovation in TLR agonist adjuvant development [62].
5.1.1. Monophosphoryl Lipid A (MPLA) as Clinically Approved TLR4-Based Vaccine Adjuvant
Among the extensively studied immune response enhancers that act upon TLR4 agonists is monophosphoryl lipid A (MPLA), a version of a low toxicity compound activating an immune response. It functions via the TRIF-dependent pathway to facilitate dendritic cell development, improve antigen presentation, and regulate cytokine expression. MPLA has been incorporated into licensed adjuvant systems for vaccines such as AS04 and AS01, causing an increase in the body's cellular and antibody immune response [63,64]. Recent studies by Carter together with co-authors in 2025 identified some key findings related to multiple peptide antigen formulations, demonstrating that when combined with MPLA-containing adjuvant systems, these formulations enhance antigen-specific T-cell responses and broaden immune recognition, supporting their application in the development of multivalent vaccines [63].
5.1.2. Flagellin-Mediated TLR5 Activation Enhancing Antigen-Specific Immunity in Vaccine Models
The recognition of conserved bacterial proteins has made microbial components be considered as promising vaccine adjuvants, particularly bacterial flagellin. Flagellin, the main structural protein of bacterial flagella, acts as a pathogen-associated molecular pattern (PAMP) that engages TLR5 on dendritic and epithelial cells, promoting innate immune activation. Activation of TLR5 initiates MyD88-dependent signaling, resulting in NF-κB activation, pro-inflammatory cytokine release, and improved antigen presentation, which enhances adaptive immune responses [49].
A study in 2025 by Afzal and co-authors shows that recombinant flagellin domains combined with inactivated porcine epidemic diarrhea virus (PEDV) vaccines in mice increased cytokine responses, cellular immunity, and virus-specific antibody titers compared to antigen alone [65]. In mucosal immunization studies, intranasal flagellin co-administered with antigens boosted antigen-specific antibodies and T-cell expansion in a TLR5-dependent manner [66]. Additionally, comparative studies of flagellin from different bacteria, such as Salmonella enterica and Pseudomonas aeruginosa, demonstrated variable but significant adjuvant activity, enhancing cytokine production and IgG responses [67].
5.1.3. TLR7 and TLR8 Agonists as Complementary Adjuvants in TLR4-Based Vaccine Strategies
TLR7 and TLR8 agonists, including synthetic imidazoquinoline derivatives and emerging lipid-modified formulations, have attracted considerable interest for their ability to activate endosomal MyD88-dependent signaling pathways. Engagement of these receptors leads to the activation of NF-κB and IRF, which in return results in an increase in the Th1 and Th17 type of immune response and in the activation of cytotoxic T-cells [18,60,68]. Consequently, ongoing research efforts are focused on optimizing TLR agonist delivery and administration strategies to maximize therapeutic efficacy while minimizing systemic toxicity. Supporting these research efforts, preclinical studies by Crouse and colleagues in 2023 evaluated lipid-modified TLR7/8 agonists in rodent and porcine vaccine models, demonstrating enhanced immunogenicity [69]. Complementing these findings, Jeon and co-authors in 2024 examined formulation and delivery approaches for TLR agonists in cancer vaccine development [60], while Wang and co-authors in 2024 highlighted the translational potential of TLR-based strategies across health and disease contexts [68].
5.1.4. TLR9 Agonists as Clinically Validated Adjuvants Supporting TLR4-Focused Vaccines
TLR9 agonists, particularly CpG oligodeoxynucleotides such as CpG-ODN 1018 are well-established solutions in the development of immune adjuvants [29,70]. CpG-ODN 1018 are generally used in licensed vaccines and authorized for clinical use, such as the hepatitis B vaccine Heplisav-B, where Phase 3 clinical trials in healthy adults aged 18–70 years demonstrated higher seroprotection rates compared with a licensed alum-adjuvanted hepatitis B vaccine, including in older adults, who typically have weaker immune responses [71]. Nonetheless, the emergence of potential immune tolerance with repeated stimulation and the influence of host factors, such as age, on immune responsiveness, reinforce the need for further dosage tuning and formulation optimization [29].
Most TLR agonists in Table 4 were initially evaluated in preclinical models, such as mice or OMV-based platforms. These are important for understanding immune activation, antigen-specific responses, and early safety; however, they are limited by immunostimulatory side effects, inflammation, and systemic toxicity, which are typical of preclinical-only studies [68,69,72,73]. Meanwhile, OMV-based nanoparticle vaccines and preclinical + clinical studies help bridge mechanistic findings in animals to human application. These approaches are crucial for assessing dosing, delivery, and immune responses, but also face limitations such as being less potent in immunocompromised individuals or requiring further validation of safety, dosing, and specificity for disease contexts [60,68].
Table 4.
TLR Agonists in Immunotherapy and Vaccine Design: Targets, Mechanisms, and Applications.
| Target TLR | Representative Agonist(s) |
Primary Application |
Immune Effect | Year and Status |
| TLR3 | ARNAX, Poly ICLC (Hiltonol), HSPPC-96 |
Cancer | Immune modulation enhances checkpoint therapy ; improves antiviral & antibacterial responses | 2018-2024, Experimental |
| TLR4 | MPL | Hepatitis B, Multivalent vaccines |
Enhanced cellular and humoral immune responses |
2025, Licensed |
| TLR5 | Flagellin | Influenza, bacterial vaccines, mucosal vaccines |
Potent innate activation; enhanced antigen presentation | 202 5 Experimental |
| TLR7&8 | Synthetic imidazoquinoline, lipid-modified | Influenza, Cancer |
Strong Th1/Th17 polarization and cytotoxic T-cell activation | 2020-2023, Experimental |
| TLR9 | CpG-ODN 1018 |
Hepatitis B | Higher seroprotection rates than alum-adjuvanted vaccines | 2015, Licensed |
| TLR4 + TLR7&8 | MPLA + lipid-modified TLR 7&8 | SARS-CoV-2, Influenza |
Synergistic, stronger, and sustained immune responses at reduced doses |
2023, Experimental |
| TLR3, TLR4, TLR9 |
Synthetic agonists |
Cancer, Infectious diseases |
Immune modulation; TME alteration enhance checkpoint therapy | 2018-2024, Experimental |
In addition to combination approaches, single-receptor strategies, like activating TLR5 with flagellin, further expand the range of vaccine adjuvant options [57]. These findings help explain how immune activation and antigen presentation occur, although challenges such as inflammation and limited clinical data remain. Moreover, agonists evaluated in Phase 3 clinical trials, such as CpG-ODN 1018 (TLR9) and MPLA (TLR4), demonstrate that the compound is well-characterized, safe, and effective in humans, supporting licensed use. Nonetheless, these advanced trials may encounter potential immune tolerance with repeated stimulation, and responsiveness can be affected by host factors such as age, emphasizing the need for optimized long-term dosing and monitoring [63,71]
5.2. Combination and Emerging TLR Agonist Formulations
The current development is focusing on combination and co-delivery approaches that merge several TLR agonists to take advantage of synergistic immune signaling, including a SARS-CoV-2 receptor-binding domain (RBD) vaccine study reported by Siram and co-authors in 2023, as well as influenza vaccine studies employing TLR7/8-based adjuvants described by Miller alongside his co-authors in 2020 [72,73]. Dual TLR agonist strategies that integrate TLR4 with TLR7/8 are delivered through alum-based or lipid-modified platforms, generating stronger and more sustained immune responses than single-agonist formulations [72,73]. Such methods enhance cellular and humoral immunity at the cost of lowering the required dosage of each agonist, which may lessen toxicity. These approaches are promising for vaccine strategies but also present the need to further study and create advancements for long-term safety, appropriate delivery method, and dosage ratio optimization of the said method [68,72,73].
5.3. Therapeutic and Immunological Applications of Toll-like Receptor Agonists
TLRs are able to regulate specific inflammatory pathways thus making them key drug targets in the treatment of both infectious and immune-related diseases [69,74]. By stimulating the host's immune defenses instead of directly attacking the bacteria, TLR agonists are most effective in overcoming antibiotic resistance. Thus, they provide a different approach to combating multidrug-resistant and hospital-acquired infections [74]. Among antiviral therapies, the TLR7 agonist imiquimod is FDA-approved for the treatment of viral warts. Meanwhile, other TLR agonists are being explored as supplementary therapies for SARS-CoV-2, HIV, and hepatitis B due to their ability to enhance the antiviral interferon responses [18,74].
On the other hand, TLR antagonists are designed for immune hyperactivation diseases such as sepsis, rheumatoid arthritis, and systemic lupus erythematosus (SLE). One particular example is the utilization of eritoran, an agent that is being developed to minimize inflammation-related damage and tissue injury [5,13]. Scientists are also looking into TLR tuning for neurodegenerative disorders, where limiting microglial activation and neuroinflammation may decrease the extent of tissue damage and slow down the development of illnesses like Alzheimer's and Parkinson's disease [68].
5.3.1. Immunotherapy
Oncology researchers are currently developing TLR agonists as immunomodulators that will be able to alter the tumour microenvironment (TME) and increase the immune system's ability to activate cancer. Among these new synthetic agonists is the TLR3 agonist ARNAX, which stimulates dendritic cells and cytotoxic T-cell responses with less systemic inflammation and cytokine-induced toxicity than other agents [63,68]. Besides being used as monotherapies, TLR agonists are also applied in conjunction with immune checkpoint inhibitors since activating TLR3, TLR4, or TLR9 pathways results in increased PD-1/PD-L1 expression and thus making resistant tumours more responsive to checkpoint blockade therapies [68]. Modulators Poly ICLC (Hiltonol) and HSPPC-96 have exhibited the potential to enhance immune cells, particularly in clinical settings after combining with personalized vaccines or radiotherapy, thus helping immune cell infiltration of brain tumours that have been resistant to the immune system [63]. In addition, TLR agonists are also known to have the ability in reprogramming metabolism and remodel the epigenome of innate immune cells via trained immunity, which consequently leads to improved secondary immune responses and long-lasting antitumour effects [74].
VI. Conceptual and Translational Limitations in TLR Agonist–Based Adjuvant Development
While advances in molecular design, computational screening, and adjuvant engineering have substantially expanded the range of candidate immunostimulatory molecules, persistent limitations remain that are not solely technical in nature. In most cases, the assumption that strong receptor binding directly predicts immune potency often fails to account for the complexity of downstream signaling and cellular context [62,75]. Identifying these conceptual gaps is crucial as they contribute to incomplete mechanistic understanding, restrict rational adjuvant optimization, and ultimately hinder efficient clinical translation despite rapid technological progress.
6.1. Evolving Perspectives on the Complexity of Toll-like Receptor Signaling
Current literature indicates that several aspects of TLR biology and agonist-mediated immune modulation remain incompletely understood, which limits the progression of TLR-based adjuvants toward clinical application [18,62]. Beyond molecular interactions, the concept of TLRs and their relationship between agonist design and delivery context being underexplored signifies the main limitation in producing further advancements on the utilization of TLRs in different immunological perspectives, such as vaccine adjuvants progression, targeted therapy utilization etc. Studies investigating TLR agonist delivery strategies, such as nanoparticle encapsulation, lipid-based formulations, or controlled-release systems, remain limited despite their known influence on receptor engagement, tissue distribution, and immune activation profiles [76,77]. Emerging evidence suggests that targeted and controlled delivery approaches may improve both efficacy and safety, yet systematic comparisons across platforms are lacking. In addition, insufficient consideration of inter-species differences in TLR orthologs complicates translational extrapolation from preclinical models to humans [76]. Off-target effects and the potential for immune dysregulation or suppression resulting from chronic TLR stimulation also remain inadequately evaluated [18].
6.1.1. Barriers to Experimental Integration of In Silico Modeling and TLR-Mediated Immune Responses
One major and subsequent limitation is the weak linkage between molecular binding data and functional immunological outcomes. In silico docking and molecular dynamics rapidly identify candidate TLR agonists, yet binding scores or ΔG values are seldom correlated with immune responses such as cytokine production, dendritic cell maturation, or T-cell activation [62,75]. This disconnect arises because TLR signaling is shaped by receptor localization, adaptor usage, and cell-type–specific regulation, which are all factors that cannot be captured by binding metrics alone. The gap is more evident in combinatorial TLR approaches, including TLR4 and TLR7 co-activation, which remain insufficiently explored despite their importance in vaccine adjuvant design [62].
This issue is compounded by limited experimental validation of computational findings. Many predicted ligand–TLR interactions lack confirmation through biophysical or cell-based assays such as surface plasmon resonance, fluorescence polarization, or reporter systems [76,77]. As a result, structure–activity relationships remain weak and translational relevance is reduced. Predictive accuracy is further limited by methodological constraints in docking and simulation studies, including rigid receptor models, simplified solvent representations, and inadequate entropy treatment, which are critical for flexible TLR ectodomains and chemically diverse ligands [62].
6.1.2. Underutilization of Multi-Agonist Approaches in Adjuvant Development
A related conceptual limitation is the limited integration of multi-TLR agonist strategies in vaccine adjuvant design. Most studies continue to emphasize single-agonist systems, even though innate immune activation relies on coordinated signaling across multiple TLR pathways. Research that systematically evaluates combinatorial agonist approaches remains scarce, particularly studies examining pathway cross-talk, signal dominance, dose balance, and temporal coordination required to achieve synergy without triggering dysregulated activation [62,75]. This gap restricts understanding of how convergent TLR signaling shapes immune polarization and response durability, and it slows the development of adjuvant platforms that better reflect physiological immune activation and support translation. Concerns also persist regarding immune exhaustion or tolerance following repeated exposure, excessive innate immune activation, and cytokine-mediated adverse effects [18].
VII. Conclusion and Future Directions
The review examined the role of TLR agonists in immune adjuvant development through which its molecular mechanisms, classifications, and its effects on translational outcomes were extensively discussed. TLRs serve as pattern recognition receptors of the innate immune system by allowing ligand recognition and intracellular signaling through adaptor proteins that detect pathogen-associated molecular patterns and danger-associated molecular patterns, linking both innate and adaptive immunity. Consequently, these signals activate transcription factors, produce pro-inflammatory cytokines and increase co-stimulatory molecule expression which enables optimal immune responses across dendritic cells, macrophages and B cells.
All TLR agonists reviewed, whether natural or synthetic, induce receptor dimerization and trigger immune activation as well as generate distinct immunomodulatory responses which affect both the innate and adaptive immune response, further contributing to the improvement of the immunogenicity of vaccines and reduce antigen dosage requirements. Strategies that have emerged have further demonstrated that dual or multi-TLR agonist formulations, including those delivered through nanoparticle-based and OMV platforms, are able to produce synergistic immune responses while reducing dose toxicity, thereby improving vaccine safety and efficacy profiles. Translational challenges remain visible despite the aforementioned advances, as evidenced by preclinical results which often differ from clinical outcomes due to species-specific TLR responses, host-dependent variability, limited in vivo validation, and inconsistencies between in silico predictions and functional immune responses, as well as in predicting long-term safety and efficacy.
Current studies continue to reveal areas of limited understanding in TLR agonist utilization as vaccine adjuvants, particularly due to the complex nature of TLR signaling networks and the challenge of translating molecular interactions into coordinated immune outcomes across diverse biological and clinical contexts. Thus, future research should prioritize integrating in silico, in vitro, and in vivo approaches such as advanced computational modeling with experimental validation to capture the complexity of interconnected TLR signaling cascades [62], employing multi-omics analyses to identify molecular predictors of immune responses [1] and exploring combined TLR agonist formulations to enhance vaccine efficacy [78]. Additionally, improved molecular docking and simulation techniques should be applied to discover and optimize novel TLR agonists in order to enable the rational design of more effective adjuvants and antiviral agents [79]. The culmination of these recommendations can provide significant benefits to healthcare, particularly in public health, as they provide a valuable framework that supports continued refinement and future advancements in the development and application of TLR agonists as vaccine adjuvants.
Author Contributions
All authors participated in the conceptualization, writing and editing of the manuscript. All authors have read and agreed to the finalized version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
No new data were generated or analyzed in this study. Data sharing is not applicable.
Acknowledgments
The authors would like to forward their gratitude to all the authors of all cited works used in the paper for providing valuable insights that deepened the understanding of vaccine adjuvants, more specifically regarding TLR activation. A gratitude is also being extended to Mr. Brian Harvey Avanceña Villanueva for his invaluable instruction, guidance, and support throughout the preparation of this review paper.
Conflicts of Interest
The authors declare no conflict of interest.
Ethics Statement
Not applicable.
References
- Pulendran, B; S. Arunachalam, P; O’Hagan, DT. Emerging concepts in the science of vaccine adjuvants. Nature reviews Drug discovery 2021, 20(6), 454–75. [Google Scholar] [CrossRef]
- Kayesh, MEH; Kohara, M; Tsukiyama-Kohara, K. TLR agonists as vaccine adjuvants in the prevention of viral infections: an overview. Front Microbiol. 2023, 14, 1249718. [Google Scholar] [CrossRef]
- Ou, BS; Baillet, J; Filsinger Interrante, MV; Adamska, JZ; Zhou, X; Saouaf, OM; et al. Saponin nanoparticle adjuvants incorporating Toll-like receptor agonists drive distinct immune signatures and potent vaccine responses. Sci Adv. 2024, 10(32), eadn7187. [Google Scholar] [CrossRef]
- Zhang, P; Yang, M; Chen, C; Liu, L; Wei, X; Zeng, S. Toll-like receptor 4 (TLR4)/opioid receptor pathway crosstalk and impact on opioid analgesia, immune function, and gastrointestinal motility. Front Immunol. 2020, 11, 1455. [Google Scholar] [CrossRef] [PubMed]
- Luo, L; Lucas, RM; Liu, L; Stow, JL. Signalling, sorting and scaffolding adaptors for Toll-like receptors. J Cell Sci. 2020, 133(5), jcs239194. [Google Scholar] [CrossRef]
- Ko, KH; Cha, SB; Lee, SH; Bae, HS; Ham, CS; Lee, MG; Kim, DH; Han, SH. A novel defined TLR3 agonist as an effective vaccine adjuvant. Frontiers in Immunology 2023, 14, 1075291. [Google Scholar] [CrossRef] [PubMed]
- Maisonneuve, C; Bertholet, S; Philpott, DJ; De Gregorio, E. Unleashing the potential of NOD-and Toll-like agonists as vaccine adjuvants. Proceedings of the National Academy of Sciences 2014, 111(34), 12294–9. [Google Scholar] [CrossRef]
- Ozverel, CS; Erdag, E. Investigation of the molecular interactions of vaccine adjuvants: can a strategic trio of toll-like receptor agonists enhance efficacy in a multifaceted approach? Biomed Biotechnol Res J 2024, 8(1), 27–36. [Google Scholar] [CrossRef]
- Kadivella, M; Varma, VP; Cp, J; Kavela, S; Azam, S; Faisal, SM. Adjuvant activity of a small molecule TLR4 agonist discovered via structure-based virtual screening. Commun Biol. 2025, 8(1), 1382. [Google Scholar] [CrossRef] [PubMed]
- Knisely, JM; Erbelding, E. Vaccines for global health: progress and challenges. J Infect Dis. 2025, 232(1), 25–27. [Google Scholar] [CrossRef]
- Cui, Y; Ho, M; Hu, Y; Shi, Y. Vaccine adjuvants: current status, research and development, licensing, and future opportunities. J Mater Chem B 2024, 12(17), 4118–4137. [Google Scholar] [CrossRef]
- Cao, LM; Yu, YF; Li, ZZ; Zhong, NN; Wang, GR; Xiao, Y; et al. Adjuvants for cancer mRNA vaccines in the era of nanotechnology: strategies, applications, and future directions. J Nanobiotechnology. 2024, 22(1), 308. [Google Scholar] [CrossRef]
- Do, KTH; Willenzon, S; Ristenpart, J; Janssen, A; Volz, A; Sutter, G; et al. The effect of Toll-like receptor agonists on the immunogenicity of MVA-SARS-2-S vaccine after intranasal administration in mice. Front Cell Infect Microbiol. 2023, 13, 1259822. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Tejada, A; Tan, DS; Gin, DY. Development of improved vaccine adjuvants based on the saponin natural product QS-21 through chemical synthesis. Accounts of Chemical Research 2016, 49(9), 1741–56. [Google Scholar] [CrossRef]
- Moni, SS; Abdelwahab, SI; Jabeen, A; Elmobark, ME; Aqaili, D; Gohal, G; Oraibi, B; Farasani, AM; Jerah, AA; Alnajai, MM; Mohammad Alowayni, AM. Advancements in vaccine adjuvants: the journey from alum to nano formulations. Vaccines 2023, 11(11), 1704. [Google Scholar] [CrossRef]
- Zhao, T; Cai, Y; Jiang, Y; He, X; Wei, Y; Yu, Y; Tian, X. Vaccine adjuvants: mechanisms and platforms. Signal transduction and targeted therapy 2023, 8(1), 283. [Google Scholar] [CrossRef] [PubMed]
- Manicassamy, S; Pulendran, B. Modulation of adaptive immunity with Toll-like receptors. In InSeminars in immunology; Academic Press, 1 Aug 2009; Vol. 21, No. 4, pp. 185–193. [Google Scholar] [CrossRef]
- Oboge, H; Riitho, V; Nyamai, M; Omondi, GP; Lacasta, A; Githaka, N; et al. Safety and efficacy of toll-like receptor agonists as therapeutic agents and vaccine adjuvants for infectious diseases in animals: a systematic review. Front Vet Sci. 2024, 11, 1428713. [Google Scholar] [CrossRef] [PubMed]
- Temchura, V; Wagner, JT; Damm, D. Immunogenicity of recombinant lipid-based nanoparticle vaccines: danger signal vs. helping hand. Pharmaceutics 2023, 16(1), 24. [Google Scholar] [CrossRef]
- Behzadi, P.; García-Perdomo, H. A.; Karpiński, T. M. Toll-like receptors: general molecular and structural biology. Journal of Immunology Research 2021, 2021(1), 9914854. [Google Scholar] [CrossRef]
- Xun, Y.; Yang, H.; Kaminska, B.; You, H. Toll-like receptors and toll-like receptor-targeted immunotherapy against glioma. Journal of hematology & oncology 2021, 14(1), 176. Available online: https://link.springer.com/article/10.1186/s13045-021-01191-2.
- Shi, M.; Chen, X.; Ye, K.; Yao, Y.; Li, Y. Application potential of toll-like receptors in cancer immunotherapy: Systematic review. Medicine 2016, 95(25), e3951. [Google Scholar] [CrossRef]
- Luchner, M.; Reinke, S.; Milicic, A. TLR agonists as vaccine adjuvants targeting cancer and infectious diseases. Pharmaceutics 2021, 13(2), 142. [Google Scholar] [CrossRef]
- Tiwari, U.; Das, S.; Tandon, M.; Ramachandran, V.; Saha, R. Vaccines for fungal infections . The National Medical Journal of India 2015, 28(1), 14–19. Available online: https://www.researchgate.net/publication/280561624_Vaccines_for_fungal_infections.
- Ouyang, MZ; Zhou, D; Zhu, Y; Zhang, M; Li, L. The inhibition of MyD88 and TRIF signaling serve equivalent roles in attenuating myocardial deterioration due to acute severe inflammation. Int J Mol Med. 2018, 41(1), 399–408. [Google Scholar] [CrossRef]
- Androulidaki, A; Wachsmuth, L; Polykratis, A; Pasparakis, M. Differential role of MyD88 and TRIF signaling in myeloid cells in the pathogenesis of autoimmune diabetes. PLoS One 2018, 13(3), e0194048. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z; Junkins, RD; Raudonis, R; MacNeil, AJ; McCormick, C; Cheng, Z; Lin, TJ. Regulator of calcineurin 1 differentially regulates TLR-dependent MyD88 and TRIF signaling pathways. PLoS One 2018, 13(5), e0197491. [Google Scholar] [CrossRef] [PubMed]
- Rink, L; Hebel, T; Fukumot, J. Sepsis and signal transduction pathway: cross-talk TLR4/MyD88/TRIF. Am J Biomed Sci. 2015, 3(4), 150–163. [Google Scholar] [CrossRef]
- Yang, J. X.; Tseng, J. C.; Yu, G. Y.; Luo, Y.; Huang, C. Y. F.; Hong, Y. R.; Chuang, T. H. Recent advances in the development of toll-like receptor agonist-based vaccine adjuvants for infectious diseases. Pharmaceutics 2022, 14(2), 423. [Google Scholar] [CrossRef]
- Fitzgerald, K. A.; Kagan, J. C. Toll-like receptors and the control of immunity. Cell 2020, 180(6), 1044–1066. Available online: https://www.cell.com/cell/fulltext/S0092-8674(20)30218-X. [CrossRef]
- Anthoney, N.; Foldi, I.; Hidalgo, A. Toll and Toll-like receptor signalling in development. Development 2018, 145(9), dev156018. [Google Scholar] [CrossRef]
- Seya, T.; Takeda, Y.; Matsumoto, M. A Toll-like receptor 3 (TLR3) agonist ARNAX for therapeutic immunotherapy. Advanced Drug Delivery Reviews 2019, 147, 37–43. [Google Scholar] [CrossRef]
- Owen, A. M.; Fults, J. B.; Patil, N. K.; Hernandez, A.; Bohannon, J. K. TLR agonists as mediators of trained immunity: mechanistic insight and immunotherapeutic potential to combat infection. Frontiers in immunology 2021, 11, 622614. [Google Scholar] [CrossRef]
- Buchta, C. M.; Bishop, G. A. Toll-like receptors and B cells: functions and mechanisms. Immunologic research 2014, 59(1-3), 12–22. [Google Scholar] [CrossRef]
- Mancini, F.; Rossi, O.; Necchi, F.; Micoli, F. OMV vaccines and the role of TLR agonists in immune response. International journal of molecular sciences 2020, 21(12), 4416. [Google Scholar] [CrossRef]
- Fischinger, S; Boudreau, CM; Butler, AL; Streeck, H; Alter, G. Sex differences in vaccine-induced humoral immunity. Semin Immunopathol 2019, 41(2), 239–249. [Google Scholar] [CrossRef] [PubMed]
- Rio, P; Caldarelli, M; Chiantore, M; Ocarino, F; Candelli, M; Gasbarrini, A; et al. Immune cells, gut microbiota, and vaccines: a gender perspective. Cells 2024, 13(6), 526. [Google Scholar] [CrossRef]
- Martínez-García, MÁ; Ojeda-Ojeda, M; Rodríguez-Martín, E; Insenser, M; Moncayo, S; Álvarez-Blasco, F; et al. TLR2 and TLR4 surface and gene expression in white blood cells after fasting and oral glucose, lipid and protein challenges: influence of obesity and sex hormones. Biomolecules 2020, 10(1), 111. [Google Scholar] [CrossRef]
- Sharma, P; Levy, O; Dowling, DJ. The TLR5 agonist flagellin shapes phenotypical and functional activation of lung mucosal antigen-presenting cells in neonatal mice. Front Immunol. 2020, 11, 171. [Google Scholar] [CrossRef] [PubMed]
- Yang, JX; Tseng, JC; Yu, GY; Luo, Y; Huang, CYF; Hong, YR; Chuang, TH. Recent advances in the development of toll-like receptor agonist-based vaccine adjuvants for infectious diseases. Pharmaceutics 2022, 14(2), 423. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S; Ye, J; Wang, H; Sun, M; Zhang, Y; Sang, X; Zhuang, Z. Application of toll-like receptors (TLRs) and their agonists in cancer vaccines and immunotherapy. Front Immunol. 2023, 14, 1227833. [Google Scholar] [CrossRef]
- Díaz-Dinamarca, DA; Salazar, ML; Castillo, BN; Manubens, A; Vasquez, AE; Salazar, F; Becker, MI. Protein-based adjuvants for vaccines as immunomodulators of the innate and adaptive immune response: current knowledge, challenges, and future opportunities. Pharmaceutics 2022, 14(8), 1671. [Google Scholar] [CrossRef]
- Takeuchi, O; Akira, S. Pattern recognition receptors and inflammation. Cell. 2016, 164(6), 1233–1247. [Google Scholar] [CrossRef] [PubMed]
- Brennan, JJ; Gilmore, TD. Evolutionary Origins of Toll-like Receptor Signaling. Mol Biol Evol 2018, 35(7), 1576-1587. Available online: https://academic.oup.com/mbe/article/35/7/1576/4994247. [CrossRef] [PubMed]
- Li, H; Li, X; Zhu, Y; Xu, J; Zhao, Q; Chen, X; et al. Systematic review of structural and immunological features of mollusk toll-like receptors in aquaculture context. Front Immunol 2025, 16, 1671434. Available online: https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2025.1671434/full. [CrossRef]
- Liu, G; Zhang, H; Zhao, C; Zhang, H. Evolutionary History of the Toll-Like Receptor Gene Family across Vertebrates. Genome Biol Evol 2020, 12(1), 3615-3634. Available online: https://pubmed.ncbi.nlm.nih.gov/31800025/. [CrossRef] [PubMed] [PubMed Central]
- Gay, NJ; Symmons, MF; Gangloff, M; Bryant, CE. Assembly and localization of Toll-like receptor signalling complexes. Nat Rev Immunol. 2018, 18(9), 546–558. [Google Scholar] [CrossRef]
- Patra, MC; Choi, S. Recent progress in the development of Toll-like receptor (TLR) agonists. Expert Opin Ther Pat. 2016, 26(6), 719–730. [Google Scholar] [CrossRef]
- Afzal, H; Murtaza, A; Cheng, LT. Structural engineering of flagellin as vaccine adjuvant: quest for the minimal domain of flagellin for TLR5 activation. Mol Biol Rep. 2025, 52(1), 104. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- De Gregorio, E; Caproni, E; Ulmer, JB. Vaccine adjuvants: mode of action. Front Immunol. 2013, 4, 214. [Google Scholar] [CrossRef] [PubMed]
- Pergolizzi, S; Fumia, A; D’Angelo, R; Mangano, A; Lombardo, GP; Giliberti, A; et al. Expression and function of toll-like receptor 2 in vertebrate. Acta Histochemica 2023, 125(3), 152028. [Google Scholar] [CrossRef] [PubMed]
- Zhang, SY; Herman, M; Ciancanelli, MJ; Pérez de Diego, R; Sancho-Shimizu, V; Abel, L; et al. TLR3 immunity to infection in mice and humans. Current Opinion in Immunology 2013, 25(1), 19–33. [Google Scholar] [CrossRef]
- Duan, T; Du, Y; Xing, C; Wang, HY; Wang, RF. Toll-Like Receptor Signaling and Its Role in Cell-Mediated Immunity. Frontiers in Immunology 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Abarca-Merlin, DM; Martínez-Durán, JA; Medina-Pérez, JD; Rodríguez-Santos, G; Alvarez-Arellano, L. From Immunity to Neurogenesis: Toll-like Receptors as Versatile Regulators in the Nervous System. International Journal of Molecular Sciences 2024, 25(11), 5711. [Google Scholar] [CrossRef]
- Mukherjee, S; Karmakar, S; Babu, SPS. TLR2 and TLR4 mediated host immune responses in major infectious diseases: a review. The Brazilian Journal of Infectious Diseases 2016, 20(2), 193–204. [Google Scholar] [CrossRef]
- Chen, Y; Lin, J; Zhao, Y; Ma, X; Yi, H. Toll-like receptor 3 (TLR3) regulation mechanisms and roles in antiviral innate immune responses. Journal of Zhejiang University-SCIENCE B 2021, 22(8), 609–32. [Google Scholar] [CrossRef]
- Lim, JS; Jeon, EJ; Go, HS; Kim, HJ; Kim, KY; Nguyen, TQT; et al. Mucosal TLR5 activation controls healthspan and longevity. Nature Communications 2024, 15(1). [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, P; Falvella, FS; Cheli, S; Perrotta, C; Clementi, E; Radice, S. The role of Toll-like receptor 4 polymorphisms in vaccine immune response. Pharmacogenomics J 2016, 16(1), 96–101. [Google Scholar] [CrossRef] [PubMed]
- Skevaki, C; Pararas, M; Kostelidou, K; Tsakris, A; Routsias, JG. Single nucleotide polymorphisms of Toll-like receptors and susceptibility to infectious diseases. Clin Exp Immunol. 2015, 180(2), 165–177. [Google Scholar] [CrossRef]
- Jeon, D; Hill, E; McNeel, DG. Toll-like receptor agonists as cancer vaccine adjuvants. Human Vaccines & Immunotherapeutics 2023, 20(1). [Google Scholar] [CrossRef]
- Lian, S; Yang, W; Zeng, Y; Tang, R; Wang, K. Targeted nano-drug delivery systems for tumor immunotherapy. Journal of Pharmaceutical Analysis 2025, 101408. [Google Scholar] [CrossRef]
- Liew, C. W.; Polanco, L. F. C.; Manalang, K.; Kurt, R. A.; others. An experimental and computational approach to unraveling interconnected TLR signaling cascades. Informatics in Medicine Unlocked 30 2022, 100939. [Google Scholar] [CrossRef]
- Carter, D; De La Rosa, G; Garçon, N; Moon, HM; Nam, HJ; Skibinski, DAG. The success of toll-like receptor 4 based vaccine adjuvants. Vaccine 2025, 61, 127413. [Google Scholar] [CrossRef]
- Choi, H; Lee, S; Kim, H; Bae, SH; Jo, S; Kim, J; et al. Integration of TLR7/8 agonists into lipid nanoparticles enhances antigen-specific immune responses to N1-methyl-Ψ-modified mRNA-LNP vaccines. Journal of Biological Engineering 2025, 19(1). [Google Scholar] [CrossRef] [PubMed]
- Afzal, H; Hoa, NT; Murtaza, A; Doan, TD; Chung, YC; Ke, GM; et al. Conserved domains of flagellin as adjuvants for an inactivated vaccine against porcine epidemic diarrhea virus. The Veterinary Journal 2025, 313, 106393. [Google Scholar] [CrossRef] [PubMed]
- Li, X; Cao, Y; Mou, M; Li, J; Huang, S; Zhang, E; et al. Enhanced TLR5-dependent migration and activation of antigen-loaded airway dendritic cells by flagellin. Journal of Leukocyte Biology 2023, 113(6), 567–76. [Google Scholar] [CrossRef] [PubMed]
- Pang, S; Liu, M; Wang, L; Shao, M; Zhu, G; Duan, Q. Differential Adjuvant Activity by Flagellins from Escherichia coli, Salmonella enterica Serotype Typhimurium, and Pseudomonas aeruginosa. Vaccines 2024, 12(11), 1212. [Google Scholar] [CrossRef]
- Wang, K; Huang, H; Zhan, Q; Ding, H; Li, Y. Toll-like receptors in health and disease. MedComm 2024, 5(5). [Google Scholar] [CrossRef]
- Crouse, B; Miller, SM; Muelken, P; Hicks, L; Vigliaturo, JR; Marker, CL; et al. A TLR7/8 agonist increases efficacy of anti-fentanyl vaccines in rodent and porcine models. npj Vaccines 2023, 8(1). [Google Scholar] [CrossRef]
- Dowling, DJ. Recent Advances in the Discovery and Delivery of TLR7/8 Agonists as Vaccine Adjuvants. ImmunoHorizons 2018, 2(6), 185–97. [Google Scholar] [CrossRef]
- Janssen, JM; Jackson, S; Heyward, WL; Janssen, RS. Immunogenicity of an investigational hepatitis B vaccine with a toll-like receptor 9 agonist adjuvant (HBsAg-1018) compared with a licensed hepatitis B vaccine in subpopulations of healthy adults 18–70 years of age. Vaccine 2015, 33(31), 3614–8. [Google Scholar] [CrossRef]
- Siram, K; Lathrop, SK; Abdelwahab, WM; Tee, R; Davison, CJ; Partlow, HA; et al. Co-Delivery of Novel Synthetic TLR4 and TLR7/8 Ligands Adsorbed to Aluminum Salts Promotes Th1-Mediated Immunity against Poorly Immunogenic SARS-CoV-2 RBD. Vaccines 2023, 12(1), 21. [Google Scholar] [CrossRef]
- Miller, SM; Cybulski, V; Whitacre, M; Bess, LS; Livesay, MT; Walsh, L; et al. Novel Lipidated Imidazoquinoline TLR7/8 Adjuvants Elicit Influenza-Specific Th1 Immune Responses and Protect Against Heterologous H3N2 Influenza Challenge in Mice. Frontiers in Immunology 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Kuai, R; Sun, X; Yuan, W; Ochyl, LJ; Xu, Y; Hassani Najafabadi, A; et al. Dual TLR agonist nanodiscs as a strong adjuvant system for vaccines and immunotherapy. Journal of Controlled Release 2018, 282, 131–9. [Google Scholar] [CrossRef] [PubMed]
- Omange, R. W.; Kim, J.; Kolhatkar, P. P.; Plott, D. M.; Van Trump, J. I.; Zhang, L.; O’Donnell, T. A.; Chen, N.–H.; Hosny, A.; Wiest, J. S.; Barry, A. H.; Addiego, E. N.; Mengistu, A.; Odorizzi, P. M.; Cai, Y.; Jacobson, R. M.; Wallin, J. J. AI discovery of TLR agonist-driven phenotypes reveals unique immune responses in people living with HIV. Frontiers in Immunology 16 2025, Article 1541152. [Google Scholar] [CrossRef]
- Manček-Keber, M.; Jerala, R. Postulates for validating TLR4 agonists. European Journal of Immunology 2015, 45(6), 1510–1516. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, A.; Rajoria, S.; George, A. L.; Mittelman, A.; Suriano, R.; Tiwari, R. K. Synthetic toll-like receptor 4 agonist peptides as a novel class of adjuvants. PLOS ONE 2012, 7(3), e30839. [Google Scholar] [CrossRef]
- Nouri-Shirazi, M.; Tamjidi, S.; Nourishirazi, E.; Guinet, E. Combination of TLR8 and TLR4 agonists reduces the degrading effects of nicotine on DC-NK mediated effector T cell generation. International immunopharmacology 2018, 61, 54–63. [Google Scholar] [CrossRef]
- Abiri, A.; Rezaei, M.; Zeighami, M. H.; Vaezpour, Y.; Dehghan, L.; KhorramGhahfarokhi, M. Discovery of new TLR7 agonists by a combination of statistical learning-based QSAR, virtual screening, and molecular dynamics. Informatics in medicine unlocked 2021, 27, 100787. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



