
Submitted:
26 January 2026
Posted:
28 January 2026
You are already at the latest version
Abstract
Protected national parks continue to face increased pressure from the expansion of human-wildlife interface zones, where habitat encroachment promotes human-wildlife contact and zoonotic disease transmission. Gastrointestinal parasites (GIPs) are a significant health issue in wild Asian elephants (Elephas maximus), affecting their fitness, survival, and potential for cross-species transmission. This study aimed to investigate the prevalence of GIPs among wild elephants at Khao Sip Ha Chan National Park in eastern Thailand. Direct smear, formalin-ethyl acetate sedimentation, flotation, and McMaster egg per gram (EPG) counting were used to examine 135 fecal samples from three populations. The findings showed that two nematodes (Strongyle-type and Strongyloides spp.) and two trematodes (Paramphistomum spp. and Fasciola spp.) were identified. All samples were positive for at least one parasite species, and 84.4% were affected by mixed infections. The prevalence of Stron-gyle-type and Strongyloides spp. varied significantly among the studied populations, but Paramphistomum spp. had a moderate prevalence, and Fasciola spp. had a low prevalence. The high parasite burden could be explained by environmental factors, host density, and movement patterns influencing parasite transmission, and these factors should be further investigated. These findings provide crucial baseline data and underscore the need for integrated parasite surveillance, alongside long-term conservation and future research.
Keywords:
helminth
; prevalence
; wild asian elephants
; mixed infections
; habitat connectivity
1. Introduction
The Eastern Forest Complex (EFCOM) represents one of Thailand’s most ecologically significant conservation networks, comprising a mosaic of interconnected protected areas across seven eastern provinces: Chanthaburi, Chachoengsao, Chon Buri, Rayong, Trat, Prachin Buri, and Sa Kaeo. This landscape includes key biodiversity hotspots such as Khao Ang Rue Nai Wildlife Sanctuary, Khao Soi Dao Wildlife Sanctuary, Khao Khitchakut, Khao Chamao–Khao Wong, and Khao Sip Ha Chan National Parks (Figure 1). Ranging in elevation from 80 to 1600 m above sea level, the EFCOM supports diverse forest types—including moist evergreen, dry evergreen, and mixed deciduous forests—and plays a critical role in sustaining Thailand’s biodiversity. The complex covers more than 2,240 km² and harbors key wildlife species, including the endangered wild Asian elephant (Elephas maximus), gaur (Bos gaurus), banteng (Bos javanicus), sambar deer (Rusa unicolor), and various mesocarnivores and avian species. According to the 2021 population survey conducted by the Chachoengsao Wildlife Research Station, the EFCOM supports approximately 463 wild Asian elephants, highlighting its critical role as one of Thailand’s key strongholds for wild elephant conservation. Within this ecological network, Khao Sip Ha Chan National Park serves as a critical foraging area and movement corridor for wild elephants. Its proximity to agricultural lands and livestock areas creates dynamic interfaces between wild and domestic animals. Such interface zones facilitate wildlife movement but simultaneously increase opportunities for cross-species transmission of pathogens, including gastrointestinal parasites (GIPs). The park, recognized as one of Thailand’s most crucial wild elephant habitats, remains vulnerable to anthropogenic pressures, including agricultural expansion, encroachment, and livestock grazing. Human-driven landscape modification and resource sharing—such as waterholes, salt licks, and crop fields—have enhanced habitat connectivity and the potential for pathogen transmission across wildlife–livestock–human interfaces [1].
Despite the ecological and cultural significance of wild Asian elephants, data on GIPs infections in free-ranging populations remain limited. Most available studies have focused on captive elephants, offering valuable insights into parasite diversity and associated health impacts [2,3,4]. For example, captive elephants in Nepal exhibited GIP prevalence rates of up to 95.2% across 17 parasite species [5], while a study in Chitwan National Park reported a prevalence of 47.6%, dominated by nematodes [6]. In Thailand, molecular detection of nematodes, including Strongyloides, Oesophagostomum aculeatum, and Ancylostoma spp., was reported in wild elephants at the Salak Phra Wildlife Sanctuary in western Thailand [7]. However, the epidemiology of GIPs in wild populations remains poorly characterized, despite their potential to influence health, reproduction, and population viability. Understanding the status of parasite infections in wild elephants is critical, as it informs conservation health strategies, aids early detection of emerging zoonotic threats, and supports One Health approaches to mitigate disease risks at the wildlife–livestock–human interface [1,8].
Wild and captive Asian elephants are highly susceptible to GIPs in their natural habitats and under human care [3]. These infections can cause weight loss, reduced fitness, and deterioration in overall health, ultimately threatening population viability [4]. Documented GIPs in elephants include trematodes such as liver flukes (Fasciola spp.) [4,6], cestodes such as tapeworms (Anoplocephala spp.) [9,10], and numerous strongyle-type nematodes, including Cyathostomidae, Ancylostomidae, Murshidia, Quilonia, Bathmostomum, Grammocephalus, and Equinurbia [6,9,11,12,13]. Among these, nematodes are the most frequently recorded, with heavy infections causing tissue damage, impaired health, and increased morbidity and mortality [14]. In captive settings, such as Kerala, India, gastrointestinal nematode infections have been associated with recurrent clinical illness, including colic, diarrhea, and dependent edema [3,15,16]. In Myanmar’s timber elephants, roundworms and liver flukes were identified as direct causes of 8% of recorded deaths and as contributing factors in an additional 13% of fatalities due to generalized weakness [17]. Beyond elephants, cross-host GIPs infections have been widely documented. In interface habitats shared by wild ungulates, livestock, and non-human primates, six zoonotic gastrointestinal helminths— Trichuris spp., Trichostrongylus spp., Oesophagostomum spp., and Strongyloides—were detected across multiple host species [18]. Their results show that livestock may harbor greater parasite richness than sympatric wild ruminants, thereby increasing the risk of spillovers. Similarly, in regions with high livestock presence, anthropogenic land-use changes have been shown to increase parasite diversity in wildlife, likely through spillovers from domestic animals [19]. These findings demonstrate that in areas where wildlife and livestock coexist, parasite transmission between species is both likely and well documented. This provides a clear rationale for examining GIPs in wild elephants in the national park.
This study aimed to investigate the prevalence and diversity of GIPs in wild Asian elephants (Elephas maximus) within a national park in eastern Thailand. Understanding parasite burdens in this ecologically vital landscape is essential for developing effective strategies for wild elephant conservation and health management. By generating baseline data on parasite infections, this research contributes to integrated One Health frameworks that link wildlife health, ecosystem stability, and community well-being.
2. Materials and Methods
2.1. Study Site and Population
Khao Sip Ha Chan National Park is located in Chanthaburi Province, eastern Thailand (approx. 12°55′23.2″N, 101°45′2.9″E) and spans undulating plains, lowlands, and mountain ranges, playing a crucial role in the conservation of wild elephants. The province spans approximately 6,338 km² and includes over 2,075 km² of conservation forest areas. The province features a tropical monsoon climate, with an average annual temperature of 27.4 °C and a mean rainfall of 3,299.6 mm across approximately 188 rainy days. The elevations range from 100 m in valleys to peaks exceeding 900 m above sea level. This landscape supports evergreen forests, mixed deciduous forests, and grassland mosaics, providing critical habitats for elephants and other large mammals.
Within this ecological matrix, Khao Sip Ha Chan National Park forms a critical conservation area of the EFCOM. The national park is ecologically connected to the Khao Soi Dao Wildlife Sanctuary in the north and the Khao Ang Rue Nai Wildlife Sanctuary in the west, forming an essential corridor for wild elephant movement and foraging. For this study, sampling was concentrated in the Phawa and Khun Song Subdistricts of the Kaeng Hang Maeo District, which represent critical zones of wild elephant activity and frequent human–elephant interactions. Based on the range of areas, three populations were distinguished: Population A (60–70 individuals) inhabits the forested landscapes surrounding Khao Sip Ha Chan National Park in Khun Song Subdistrict. The terrain is dominated by evergreen and mixed deciduous forest, with wild elephants regularly foraging along forest–agriculture edges. Population B (15–20 individuals) ranges around the Phawa Reservoir (Khlong Phawa Yai Reservoir), also in Phawa Subdistrict. This area is characterized by riparian vegetation, secondary forests, and permanent water resources that attract wild elephants year-round, and Population C (20–30 individuals) occurs along the boundary adjoining the Khao Ang Rue Nai Wildlife Sanctuary in Phawa Subdistrict. The sites of sample collection for the three wild elephant populations are distributed along the interface between protected forest areas and adjacent human settlements at Khoa Sip Ha Chan National Park (Figure 2). Notably, several sampling locations are located near domestic livestock-grazing areas. This spatial overlap suggests that wild elephants and domestic livestock may utilize shared ecological resources—such as communal grazing fields, waterholes, and forest edges—thereby increasing the risk of cross-species parasite transmission. Such mixed-use zones serve as potential transmission hotspots, where helminth eggs or larvae may persist in the environment due to fecal contamination, shared water sources, or mechanical vectors. This area forms a transitional habitat between intact protected forest and agricultural land, where wild elephants frequently move across ecological and human-dominated landscapes.
2.2. Fecal Sample Collection
Fresh fecal samples were collected from wild Asian elephants at Khao Sip Ha Chan National Park, Eastern Thailand, between June and August 2024 (rainy season), representing three populations. Wild elephants were tracked on foot, with direct observation using binoculars from a safe distance and sign-tracking based on feeding evidence [20,21]. Dung piles were located along wild elephant foraging routes by tracking signs. Only freshly deposited feces were collected to maintain high sample quality for parasitological analysis. These features included a thin mucous layer covering the dung piles, high dung moisture content, and the absence of insect activity, all of which are good indicators that a sample has been undisturbed and recently excreted [22]. Then, the researcher characterized and graded the collected samples to further confirm their freshness and consistency. All fecal samples were collected using sterile gloves to prevent contamination during handling. Freshly deposited dung piles were carefully examined, and selected material from the inner portion of each bolus was extracted to minimize contamination from soil nematodes or environmental debris. Approximately 100–200 g of feces was placed directly into labelled zip-locked bags. The geographic coordinates of each sampling location were recorded. All samples were stored immediately in a cool box and transported under chilled conditions (within 24 h) to the parasitology laboratory at the Faculty of Veterinary Technology, Kasetsart University, Bangkok. Upon arrival, samples were refrigerated at 4–6 °C and processed within 7 days following established parasitological handling protocols [23].
2.3. Sample Analysis
To comprehensively detect GIPs in wild Asian elephants, four complementary laboratory techniques were employed, following standardized parasitological protocols described in [5]. The direct smear method served as a preliminary screening tool, in which a small amount of fecal material was mixed with saline on a microscope slide and examined under light microscopy for rapid qualitative identification of helminth eggs and larvae. To enhance detection sensitivity, both flotation and formalin–ethyl acetate centrifugal sedimentation techniques were used. Flotation using a saturated salt solution allowed lighter nematode eggs to float to the surface for microscopic examination. In contrast, the sedimentation technique concentrated heavier trematode eggs by centrifugation after mixing feces with formalin and ethyl acetate. In addition to these qualitative methods, parasite burden was quantified using the McMaster egg per gram (EPG) technique, a widely used parasitological method. Fecal suspensions prepared with a flotation solution were introduced into a calibrated McMaster counting chamber. After settling, eggs were counted microscopically and multiplied by a standard conversion factor (×50) to estimate EPG values. This metric provided insights into the intensity of infection across individual wild elephants and populations.
2.4. Statistical Analysis
The prevalence of parasitic infections, the number of positive samples as a percentage of the total number of samples, was calculated and presented as a percentage, followed by a 95% confidence interval (CI) of infected individuals within each of the three wild elephant populations (A, B, and C). The differences in parasitic prevalence across the three distinct populations were compared using Fisher’s exact test, which is appropriate for analyzing proportions when sample sizes are small. To evaluate parasite burden, we used EPG values obtained using the McMaster technique for Strongylus spp. and Strongyloides spp. egg counts. Before selecting statistical tests, we assessed the distribution of EPG data using two steps: visual inspection via histograms and a formal Shapiro-Wilk test for normality. Both approaches indicated that the data did not follow a normal distribution. As a result, we applied non-parametric methods for subsequent analyses. EPG data were described using medians and interquartile ranges (IQRs) to reflect the central tendency and spread. Differences in parasite burden across the three populations were tested using the Kruskal-Wallis test, followed by Dunn’s test to further examine pairwise differences. All statistical analyses were conducted using Stata version 17.0 (StataCorp, Texas, USA), with a p-value < 0.05 considered statistically significant.
3. Results
A total of 135 fecal samples of wild Asian elephants (Elephas maximus) were collected from inside and outside conservation areas, representing three distinct populations: Population A (n=83), Population B (n=13), and Population C (n=39) (Figure 2).
The findings showed that all samples yielded at least one ovum or larva of GIP species, including nematodes and trematodes. The ova stage, two types of nematode eggs, Strongyle-type and Strongyloides spp., and two types of trematode eggs, Paramphistomum spp. and Fasciola spp., were identified (Figure 3). Additionally, during the larval stage, two morphologically distinct nematode larvae were identified: Strongyloides spp. larvae and non-Strongyloides larvae. These larval morphs represent variation in species identification and developmental stages of parasitic nematodes in wild Asian elephants (Figure 4).
3.1. Prevalence of GIPs
All fecal samples tested positive for at least one GIP species, with a high overall prevalence of mixed infections, 84.44% (114/135) (Table 1). The prevalence of nematodes varied consistently between the populations studied (p<0.05). Strongyle-type eggs were the most prevalent, at 100% (13/13) in Population B and 94.9% (37/39) in Population C, which are significantly higher than in Population A, 68.7% (57/83) (p = 0.001). Strongyloides spp. eggs were much more frequent in Population C (87.2%, 37/39) than in Population B (53.8%, 7/13) and Population A (42.2%, 35/83) (p < 0.001).
The prevalence of trematodes among the three populations did not significantly differ (p>0.05). However, Paramphistomum spp. was present at moderate levels (46.1%-67.5%) across all populations. While Fasciola spp. eggs were detected in a few samples, 5.2% (7/135), which 6% (5/83) in Population A, 15.4% (2/13) in Population B, and were absent in Population C.
There were significant differences in larval prevalence between populations (p < 0.001). Population C revealed the highest prevalence of Strongyloides spp. larvae (87.2%; 34/39), followed by 61.5% (8/13) of Population B, which were significantly greater than Population A (39.8%; 33/83). Likewise, non-Strongyloides larvae were detected in all samples collected from Populations B (13/13) and C (39/39), which were significantly higher than those of Population A (72.3%; 60/83) (p < 0.001).
Distribution of Strongyle-type and Strongyloides spp. egg counts among the three populations were presented in Figure 5. A comparison of GIPs loads and distribution across three populations revealed significant differences in Strongyle-type and Strongyloides spp. eggs count (p < 0.001). The highest parasite infection intensity was observed in Population C, characterized by a broader distribution of egg counts and higher median values for both Strongyle-type eggs and Strongyloides spp., at 400 EPG (IQR: 50-800 EPG) and 300 EPG (IQR:100-600 EPG), respectively. The median EPG values recorded in Population C were significantly higher than those of Population A and B (p < 0.01).
The finding indicated that low overall parasite burdens were observed in Populations A and B. The median egg counts for Strongyle-type in Population A and Population B were 50 EPG, but Population A had a wider range of infections (IQR: 0-300 EPG), while Population B had a more concentrated range (IQR: 50-100 EPG). Likewise, the median egg counts of Strongyloides spp. were 50 EPG (IQR: 0-150 EPG) in Population A and 0 EPG (IQR: 0-50 EPG) in Population B. However, there was no significant difference when comparing the median egg counts of both Strongyle-type and Strongyloides spp. (p = 1.000 and p = 0.512, respectively).
3.2. Mixed Infection
There was a high prevalence of mixed infections across all populations (84.4%; 114/135), with 100% (39/39) in Population C, followed by Population A (78.3%, 65/83) and Population B (76.9%, 10/13). Distribution of single and mixed infections, which are combinations of two, three, and four types of GIPs among three populations were presented in Figure 6. The most common mixed infection was a combination of nematodes and trematode including Strongyle-type eggs, Strongyloides spp., and Paramphistomum spp., which were prevalent in Population C (53.8%, 21/39), Population A (36.1%, 30/83), and Population B (23.1%, 3/13), respectively. The finding also indicated that mixed infection, a combination of Strongyle-type and Strongyloides spp., was significantly higher in Population C (46.1%, 18/39, p = 0.005) than in Populations A and B (18.1%, 15/83, and 30.8%, 4/13, respectively). However, other patterns of mixed infection showed no statistically significant differences between populations (p > 0.05). Interestingly, the combination of Strongyle-type and Paramphistomum spp. presented only in Population A (1.2%, 1/83).
4. Discussion
In this study, we investigated the prevalence of GIPs in three wild Asian elephant populations at Khao Sip Ha Chan National Park, eastern Thailand. Four helminth types of ova stages: two nematodes (Strongyle-type and Strongyloides spp.) and two trematodes (Paramphistomum spp. and Fasciola spp.) were identified. The results showed a high prevalence of infection across all populations studied and statistically significant differences in parasite burden and infection intensity between the populations. Strongyle-type and Strongyloides spp. were the most frequently detected. Interestingly, Strongyle-type eggs were present in 100% samples from Population B, 94.9% from Population C, and 68.7% from Population A (p < 0.001), indicating that these parasites are widely distributed among wild elephant populations. Similarly, Strongyloides spp. were markedly more prevalent in Population C (87.2%) than in Population A (42.2%) and in Population B (53.8%) (p < 0.001), highlighting substantial differences in parasite burden between the populations. These results align with previous studies conducted in India and Sri Lanka, which found that the most common nematode detected in wild elephants was Strongylus [10,24,25,26,27,28]. A previous study in Sri Lanka revealed a markedly higher prevalence of strongyles in wild elephants than in captive ones [28]. Additionally, a lower prevalence of strongyle type was observed in private and temple elephants compared to those managed by the forest department, likely due to differences in management practices, habitat utilization, and exposure to infective stages.
Several ecological and environmental factors, including habitat conditions, exposure to contaminated environments, elephant density, and movement patterns, were identified as potential drivers of gastrointestinal parasite infections. Wild elephants, due to their unrestricted movement and foraging behaviors, were more frequently exposed to infective stages of parasites in contaminated grazing areas and water sources [9,24,29]. In comparison, captive elephants were routinely dewormed and fed controlled diets, thus limiting exposure to parasitic diseases [15,28]. The significantly higher prevalence of Strongyle-type and Strongyloides spp. eggs in fecal samples of wild Asian elephants in the present study suggested a more favorable environment for parasite transmission, potentially due to greater habitat fragmentation, higher humidity, and increased contact with infected dung and water sources. In addition, studies in Myanmar and Malaysia demonstrated that the changes in nematode presence are significantly influenced by environmental determinants, seasonality, and host immunity [16,30], further substantiating that the dynamics of these factors shape infection dynamics in this study. Differences in parasite burden between wild and captive elephant populations further emphasize the role of management strategies in controlling parasitic infections. It has been reported that wild elephants tend to have much higher strongyle infection rates than captive and semi-captive elephants, attributed to greater exposure to the environment, free-range foraging, and limited medical intervention [9,31]. Some captive elephants carry GIPs despite these protective measures, but have lower parasite loads than wild elephants [9].
In the present study, two trematode species (Paramphistomum spp. and Fasciola spp.) have moderate prevalence estimates of 46.1%-67.5% in Paramphistomum spp. and a low prevalence rate in Fasciola spp. (0-15.4%). No significant difference among the three populations (p = 0.165) suggested a relatively even distribution of the study area. The absence of Fasciola spp. in Population C is noteworthy, as previous studies have reported considerable variation in trematode prevalence among elephant populations. For instance, a high prevalence of Fasciola spp. infection was reported in wild elephants in India [4] and Sumatran elephants [16,32]. In Nepal, both captive and wild elephants were found to be infected with Fasciola spp., indicating its widespread distribution among Asian elephants [6,33,34]. Given these previous findings, the low prevalence of Fasciola spp. observed in this study might reflect regional differences in parasite transmission dynamics. The relatively low prevalence of Fasciola spp. observed in this study might be attributed to differences in environmental conditions, host exposure, and the availability of intermediate snail hosts, which are essential for the parasite's life cycle. Fasciola spp. require specific freshwater snails as intermediate hosts to complete their development [35]. The absence of Fasciola spp. in Population C indicated that these wild elephants have restricted access to water sources that would support snail populations or display foraging patterns that reduce their risk of exposure to infection. Additionally, the movement patterns and habitat use of wild elephants may further influence infection rates, with populations with greater access to diverse landscapes potentially experiencing lower parasite burdens.
Rumen flukes (Paramphistomum spp.) are common GIPs of large herbivores, including elephants. These trematodes primarily inhabit the forestomach compartments, where they can cause subclinical infections or severe gastrointestinal disturbances [36]. The moderate prevalence reported in this study is consistent with previous studies conducted in Sri Lanka, where Paramphistomum spp. infection has commonly been reported in wild elephants [4,9]. In Thailand, previous studies have reported a high prevalence of Paramphistomum spp. infection rates in cattle with a detected prevalence above 90% [37,38]. The infection of Paramphistomum spp. in wild elephants indicates a common route of parasite transmission between domestic and wild herbivores, which could be further promoted by shared grazing pastures and water sources.
Furthermore, several studies have reported mixed gastrointestinal parasitic infections in wild Asian elephants across their distribution [4,16,30]. This study found a remarkably high prevalence of mixed infections among wild elephant populations in Chanthaburi, Thailand: 100% in Population C, 78.3% in Population A, and 76.9% in Population B. The presence and mix of infections among multiple helminth species within these wild elephant populations are consistent with reported research in Sri Lanka, where mixed infections were found at 47.1%, significantly greater than single infection (21.2%) [9]. Similarly, research conducted in South Wayanad, India, showed that the presence of multiple parasitic species was significantly higher in continuous habitats, likely due to higher biodiversity and greater freedom of movement among elephant populations, which facilitates parasite transmission [10,16]. Increased exposure to diverse grazing areas and contaminated environments has been suggested as a key factor contributing to mixed infections in wild elephants. In Chanthaburi, wild elephants frequently venture outside conservation forest boundaries into agricultural lands and human settlements, where they share grazing areas and water sources with livestock such as cattle and buffalo. This overlapping habitat could further promote parasite transmission between species, increasing the likelihood of mixed infections.
The most common mixed infection detected in this study was Strongyle-type eggs, Strongyloides spp., and Paramphistomum spp., which are more prevalent in Population C (53.8%), Population A (36.14%), and Population B (23.07%). The higher parasite burden in Population C suggested that environmental factors, including humidity, soil conditions, and host density, may facilitate parasite transmission. Similar to findings from studies conducted in Sri Lanka and India, wild elephants exhibited a greater tendency to multiple parasitic infections than their captive counterparts, possibly due to increased exposure to infective parasite stages in their natural habitats [2,24].
Mixed infections of Strongyle type eggs and Strongyloides spp. have been widely reported in wild elephants [25,39]. The frequent co-occurrence of these nematodes suggests shared transmission routes, likely through contaminated grazing areas, water sources, or soil. Many studies reported the occurrence of strongyles and Strongyloides spp. infections in areas where wild elephants can access shaded, humid environments conducive to larval maturation [30]. Moreover, elephants with high parasite loads often experience reduced immunity, making them more susceptible to additional infections [40]. In this present study, however, one parasite species might help other species colonize the host, making mixed infections more likely. This pattern has been recorded in both wild and semi-captive elephants, wherein mixed infections observed in elephants traversing fragmented landscapes were more severe than those observed in elephants in continuous habitats [10,16].
This study highlights the high prevalence and diversity of GIPs among wild Asian elephants in Chanthaburi Province, Thailand. The detection of both nematode and trematode species, particularly Strongylus spp. eggs and Strongyloides spp., which are the most dominant—indicates that wild elephants are consistently exposed to infective stages in their natural environment. Notably, the high rate of mixed infections in Population C suggests that ecological factors such as humidity, habitat fragmentation, host density, and proximity to domestic livestock significantly influence parasite transmission dynamics. The survey area encompasses buffer zones, communal grazing fields, water sources, and wetlands frequently used by both wild elephants and domestic livestock, such as cattle and buffalo. This landscape overlap raises concerns about cross-species transmission of parasites, particularly for shared helminths such as Paramphistomum spp. and Fasciola spp. Although the latter was found at low prevalence, likely due to ecological constraints (e.g., limited availability of aquatic intermediate hosts), its zoonotic potential remains important.
The presence of ruminants near forest edges may serve as reservoirs, contaminating soil and water bodies with parasite eggs and larvae, thereby increasing the risk of infection for wildlife. These findings align with global research indicating that parasites with multi-host capacity can spill over across species in shared habitats [41,42]. For example, cross-infections of the trematodes, Fasciola spp., at the livestock-wildlife interfaces have been reported in Uganda [43]. In Thailand, similar spillover risks were observed in the Huai Kha Khaeng Wildlife Sanctuary, a World Heritage Site located in western Thailand along the border with Myanmar, where both wild banteng and domestic cattle harbored strongyle-type and Fasciola spp. eggs [44]. Environmental conditions, such as rainfall, humidity, and soil moisture, further amplify the risk of parasite transmission. These factors have been shown to enhance parasite survival and development in multi-host systems [45]. Despite formal restrictions, informal livestock grazing within forest buffer zones remains common in Chanthaburi Province, potentially facilitating the transmission of parasitic diseases.
In this study, the presence of zoonotic parasites, such as Strongyloides spp. and Fasciola spp., has implications that extend beyond conservation, affecting One Health concerns for both humans and livestock. We recommend implementing longitudinal, molecular-based surveillance to identify parasite species and their sources of infection. Spatial modeling of transmission hotspots and land-use planning that limits shared access to critical habitats will be essential. Collaborative efforts between conservationists, veterinarians, public health agencies, and local communities are urgently needed to mitigate parasitic risks in human-elephant interface zones.
5. Conclusions
Our study is the first full-scale investigation of the prevalence of GIPs in wild Asian elephants (Elephas maximus) in the study area, using fecal examination methods such as direct smear, flotation, sedimentation, and the McMaster technique. Results showed a high and consistent burden of helminth infection, with strongyle-type eggs and Strongyloides spp. predominant, which varied significantly among the distinct populations of wild elephants. These findings underscore the importance of including parasitological surveillance in conservation interventions for wild elephants at high risk of cross-species transmission, particularly in human-elephant interface zones. Finally, the result demonstrates the importance of a One Health strategy that integrates wildlife, livestock, and human communities in addressing parasite-induced health and conservation issues.
Author Contributions
Conceptualization, S.K., S.T., S.S., and P.S.; methodology, S.K., S.T., S.S., R.C., and P.S.; formal analysis, S.T. and T.J.; investigation, S.K., S.T., P.Y., W.B., and P.S.; writing—original draft preparation, S.K. and W.B.; writing—review and editing, S.K., S.T., and P.S.; supervision, S.T., S.S., R.C. and P.S.; project administration, S.K., P.Y.; and P.S. funding acquisition, P.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Faculty of Veterinary Medicine, Kasetsart University, and the Southeast Asia One Health University Network (SEAOHUN) 2024. (TA24-Y2-104).
Institutional Review Board Statement
The animal study protocol was approved by the Ethics and Animal Care and Use Committee of the Faculty of Veterinary Medicine, Kasetsart University, Thailand (approval number: ACKU67-VET-023), and conducted with permission from the Department of National Parks, Wildlife and Plant Conservation, Thailand.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available from the corresponding author upon request.
Acknowledgments
The authors would like to express their sincere gratitude to the head and staff of Khao Sip Ha Chan National Park, Department of National Parks, Wildlife and Plant Conservation (DNP), Thailand, for their kind support, assistance, and facilitation during fieldwork activities.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| GIPs | Gastrointestinal parasites |
| EFCOM | The Eastern Forest Complex |
References
- Vicente, J.; Vercauteren, K.C.; Gortázar, C. Diseases at the Wildlife–Livestock Interface: Research and Perspectives in a Changing World; Springer Nature: Cham, Switzerland, 2021; vol. 3, pp. 91–119. [Google Scholar]
- Fowler, M.E.; Mikota, S.K. Biology, Medicine and Surgery of Elephants; Blackwell Publishing: Oxford, UK, 2006; pp. 159–181. [Google Scholar]
- Chandrasekharan, K.; Radhakrishnan, K.; Cheeran, J.V.; Muraleedharan, K.N.; Prabhakaran, T. Review of the incidence, etiology and control of common diseases of Asian elephants with special reference to Kerala. In Healthcare Management of Captive Asian Elephants; Ajitkumar, G., Anil, K.S., Alex, P.C., Eds.; Kerala Agricultural University Press: Pookot, India, 2009; pp. 92–100. [Google Scholar]
- Ahmed, N.; Doley, S. Prevalence of gastro-intestinal parasitic load of Asian elephants (Elephas maximus) in Unakoti, Tripura. J. Entomol. Zool. Stud. 2017, 5(4), 1514–1515. [Google Scholar]
- Adhikari, B.; Pokhrel, S.; Acharya, A.; Dhakal, D.; Shrestha, A.; Acharya, K.P. Prevalence of intestinal parasites in captive Asian elephants (Elephas maximus) in Central Nepal. J. Vet. Parasitol. 2025, 39(1), 23–30. [Google Scholar]
- Dahal, G.; Sadaula, A.; Gautam, M.; Magar, A.R.; Adhikari, S. Prevalence of gastrointestinal parasites in endangered captive Asian elephants (Elephas maximus) of Chitwan National Park in Nepal. Arch. Agric. Environ. Sci. 2023, 8(3), 290–294. [Google Scholar] [CrossRef]
- Phuphisut, O.; Maipanich, W.; Pubampen, S.; Yindee, M.; Kosoltanapiwat, N.; Nuamtanong, S.; Adisakwattana, P. Molecular identification of the strongyloid nematode Oesophagostomum aculeatum in the Asian wild elephant Elephas maximus. J. Helminthol. 2016, 90(4), 434–440. [Google Scholar] [CrossRef]
- Deak, G.; Germitsch, N.; Rojas, A.; Sazmand, A. Wildlife parasitology: Emerging diseases and neglected parasites. Front. Vet. Sci. 2024, 11, 1439564. [Google Scholar] [CrossRef]
- Abeysekara, N.; Rajapakse, R.J.; Rajakaruna, R.S. Comparative cross-sectional survey on gastrointestinal parasites of captive, semi-captive, and wild elephants of Sri Lanka. J. Threat. Taxa 2018, 10(5), 11583–11594. [Google Scholar] [CrossRef]
- Abhijith, T.V.; Ashokkumar, M.; Dencin, R.T.; George, C. Gastrointestinal parasites of Asian elephants (Elephas maximus L. 1978) in south Wayanad forest division, Kerala, India. J. Parasit. Dis. 2018, 42, 382–390. [Google Scholar] [CrossRef]
- Thurber, M.I.; O’Connell-Rodwell, C.E.; Turner, W.C.; Nambandi, K.; Kinzley, C.; Rodwell, T.C.; Faulkner, C.T.; Felt, S.A.; Bouley, D.M. Effects of rainfall, host demography and musth on strongyle fecal egg counts in African elephants (Loxodonta africana) in Namibia. J. Wildl. Dis. 2011, 47, 172–181. [Google Scholar] [CrossRef]
- McLean, E.R.; Kinsella, J.M.; Chiyo, P.I.; Obanda, V.; Moss, C.J.; Archie, E.A. Genetic identification of five strongyle nematode parasites in wild African elephants (Loxodonta africana). J. Wildl. Dis. 2012, 48, 707–716. [Google Scholar] [CrossRef]
- Mbaya, A.W.; Ogwiji, M.; Kumshe, H.A. Effects of host demography, season and rainfall on the prevalence and parasitic load of gastrointestinal parasites of free-living elephants (Loxodonta africana) of the Chad Basin National Park, Nigeria. Pak. J. Biol. Sci. 2013, 16, 1152–1158. [Google Scholar] [CrossRef] [PubMed]
- Lynsdale, C.L.; Franco dos Santos, D.J.; Hayward, A.D.; Mar, K.U.; Htut, W.; Aung, H.H.; Soe, A.T.; Lummaa, V. A standardised faecal collection protocol for intestinal helminth egg counts in Asian elephants, Elephas maximus. Int. J. Parasitol. Parasites Wildl. 2015, 4(3), 307–315. [Google Scholar] [CrossRef] [PubMed]
- Saseendran, P.C.; Rajendran, S.; Subramanian, H.; Sasikumar, M.; Vivek, G.; Anil, K.S. Incidence of helminthic infection among annually dewormed captive elephants. Zoos’ Print J. 2004, 19, 1422. [Google Scholar] [CrossRef]
- Hing, S.; Othman, N.; Nathan, S.K.S.S.; Fox, M.; Fisher, M.; Goossens, B. First parasitological survey of endangered Bornean elephants (Elephas maximus borneensis). Endanger. Species Res. 2013, 21, 223–230. [Google Scholar] [CrossRef]
- Mar, K.U. The Demography and Life-History Strategies of Timber Elephants of Myanmar. Ph.D. Thesis, University College London, London, UK, 2007. [Google Scholar]
- Obanda, V.; Ndeereh, D.; Mijele, D.; Ngethe, J.; Waititu, K.; Wambua, L.; Warigia, M.; Gakuya, F.; Alasaad, S. Infection dynamics of gastrointestinal helminths in sympatric non-human primates, livestock and wild ruminants in Kenya. PLoS ONE 2019, 14(6), e0217929. [Google Scholar] [CrossRef]
- Chakraborty, D.; Reddy, M.; Tiwari, S.; Umapathy, G. Land use change increases wildlife parasite diversity in Anamalai Hills, Western Ghats, India. Sci. Rep. 2019, 9, 11975. [Google Scholar] [CrossRef]
- Ishwaran, N. Elephant and woody-plant relationships in Gal Oya, Sri Lanka. Biol. Conserv. 1983, 26(3), 255–270. [Google Scholar] [CrossRef]
- Mohapatra, K.K.; Patra, A.K.; Paramanik, D.S. Food and feeding behaviour of Asiatic elephant (Elephas maximus Linn.) in Kuldiha Wildlife Sanctuary, Odisha, India. J. Environ. Biol. 2013, 34(1), 87–92. [Google Scholar]
- Cox, P.; Griffith, M.; Angles, M.; Deere, D.; Ferguson, C. Concentrations of pathogens and indicators in animal feces in the Sydney watershed. Appl. Environ. Microbiol. 2005, 71(10), 5929–5934. [Google Scholar] [CrossRef]
- Nielsen, M.K.; Vidyashankar, A.N.; Andersen, U.V.; DeLisi, K.; Pilegaard, K.; Kaplan, R.M. Effects of fecal collection and storage factors on strongylid egg counts in horses. Vet. Parasitol. 2010, 167(1), 55–61. [Google Scholar] [CrossRef] [PubMed]
- Vidya, T.N.C.; Sukumar, R. The effect of some ecological factors on the intestinal parasite loads of the Asian elephant (Elephas maximus) in southern India. J. Biosci. 2002, 27(5), 521–528. [Google Scholar] [CrossRef]
- Nishanth, B.; Srinivasan, S.R.; Jayathangaraj, M.G.; Sridhar, R. Incidence of endoparasitism in free-ranging elephants of Tamil Nadu State. Tamilnadu J. Vet. Anim. Sci. 2012, 8(6), 332–335. [Google Scholar]
- Vimalraj, P.G.; Jayathangaraj, M.G. Endoparasitic infections in free-ranging Asiatic elephants of Mudumalai and Anamalai Wildlife Sanctuary. J. Parasit. Dis. 2015, 39, 474–476. [Google Scholar] [CrossRef]
- Heinrich, L. Prevalence and Molecular Identification of Helminths in Wild and Captive Sri Lankan Elephants (Elephas maximus).; Research Project, Royal Veterinary College, University of London, London, UK, 2016. [Google Scholar]
- Abeysinghe, K.S.; Perera, A.N.F.; Pastorini, J.; Isler, K.; Mammides, C.; Fernando, P. Gastrointestinal strongyle infections in captive and wild elephants in Sri Lanka. Gajah 2017, 46, 21–27. [Google Scholar]
- Watve, M.G.; Sukumar, R. Parasite abundance and diversity in mammals: Correlates with host ecology. Proc. Natl. Acad. Sci. USA 1995, 92(19), 8945–8949. [Google Scholar] [CrossRef]
- Chel, H.M.; Iwaki, T.; Hmoon, M.M.; Thaw, Y.N.; Soe, N.C.; Win, S.Y.; Bawm, S.; Htun, L.L.; Win, M.M.; Oo, Z.M.; et al. Morphological and molecular identification of cyathosthomine gastrointestinal nematodes of Murshidia and Quilonia species from Asian elephants in Myanmar. Int. J. Parasitol. Parasites Wildl. 2020, 11, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Chichilichi, B.; Pradhan, C.R.; Babu, L.K.; Sahoo, N.; Panda, M.R.; Mishra, S.K.; Behera, K.; Das, A.; Hembram, A. Incidence of endoparasite infestation in free-ranging and captive Asian elephants of Odisha. Int. J. Livest. Res. 2018, 9, 302–310. [Google Scholar]
- Stremme, C.; Lubis, A.; Wahyu, M. Implementation of regular veterinary care for captive Sumatran elephants (Elephas maximus sumatranus). J. Asian Elephant Spec. Group 2007, 27, 6–14. [Google Scholar]
- Karki, K.; Manandhar, P. Incidence of gastrointestinal helminthes in captive elephants in wildlife reserves of Nepal. Articlesbase: Free Online Articles Directory. 2008. Available online: https://www.scribd.com/document/3722208/Incidence-of-Gastrointestinal-Helminthes-in-Captive-Elephants-in-Wildlife-Reserves-of-Nepa (accessed on 23 October 2025).
- Shahi, M.K.; Gairhe, K.P. Prevalence of helminths in wild Asian elephants and Indian rhinoceros in Chitwan and Bardia National Park, Nepal. Nepal. Vet. J. 2019, 36, 60–74. [Google Scholar] [CrossRef]
- Malatji, D.P.; Mukaratirwa, S.; Kufahakurume, F. Environmental factors influencing the transmission dynamics of Fasciola spp. in large herbivores. Parasitol. Res. 2020, 119, 2891–2903. [Google Scholar]
- Chaoudhary, V.; Hasnani, J.J.; Khyalia, M.K.; Pandey, S.; Chauhan, V.D.; Pandya, S.S.; Patel, P.V. Morphological and histological identification of Paramphistomum cervi (Trematoda: Paramiphistoma) in the rumen of infected sheep. Vet. World 2015, 8(1), 125–129. [Google Scholar] [CrossRef]
- Sakpuaram, T.; Sattabongkot, J.; Boonmars, T. Epidemiology of Paramphistomum infections in livestock in Thailand. Asian J. Vet. Parasitol. 2022, 31(3), 291–301. [Google Scholar]
- Kaewnoi, D.; Wiriyaprom, R.; Indoung, S.; Ngasaman, R. Gastrointestinal parasite infections in fighting bulls in South Thailand. Vet. World 2020, 13(8), 1544–1549. [Google Scholar] [CrossRef] [PubMed]
- Dharmarajan, G. Epidemiology of Helminth Parasites in Wild and Domestic Herbivores at the Mudumalai Wildlife Sanctuary, Tamil Nadu. Ph.D. Thesis, Tamil Nadu Veterinary and Animal Sciences University (TANUVAS), Chennai, India, 1999. [Google Scholar]
- Fontanarrosa, M.F.; Vezzani, D.; Basabe, J.; Eiras, D.F. An epidemiological study of gastrointestinal parasites of dogs from southern Greater Buenos Aires (Argentina): Age, gender, breed, mixed infections, and seasonal and spatial patterns. Vet. Parasitol. 2006, 136(3–4), 283–295. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E.; Taylor, L.H.; Haydon, D.T. Population biology of multihost pathogens. Science 2001, 292(5519), 1109–1112. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.G.; Plein, M.; Morgan, E.R.; Vesk, P.A. Uncertain links in host–parasite networks: Lessons for parasite transmission in a multi-host system. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372(1719), 20160095. [Google Scholar] [CrossRef]
- Namirembe, D.; Huyse, T.; Madsen, H.; et al. Liver fluke and schistosome cross-infection risk between livestock and wild mammals in Western Uganda, a One Health approach. Int. J. Parasitol. Parasites Wildl. 2024, 25(1), 101022. [Google Scholar] [CrossRef]
- Watwiengkam, N.; Patikae, P.; Thiangthientham, P.; Ruksachat, N.; Simkum, S.; Arunlerk, K.; Purisotayo, T. Prevalence of gastrointestinal parasites in free-ranging bantengs (Bos javanicus) and domestic cattle at a wildlife–livestock interface in Thailand. Trends Sci. 2024, 21(3), 7368. [Google Scholar] [CrossRef]
- Babayani, N.D.; Rose Vineer, H.; Walker, J.G.; Davidson, R.K. Climate and parasite transmission at the livestock–wildlife interface. Front. Vet. Sci. 2022, 8, 816303. [Google Scholar] [CrossRef]
Figure 1.
A map of Thailand highlights major protected areas across the eastern provinces, the Eastern Forest Complex (EFCOM). The red star indicates the study site at Khao Sip Ha Chan National Park, Chanthaburi Province. The map (left) shows the eastern region of Thailand.
Figure 1.
A map of Thailand highlights major protected areas across the eastern provinces, the Eastern Forest Complex (EFCOM). The red star indicates the study site at Khao Sip Ha Chan National Park, Chanthaburi Province. The map (left) shows the eastern region of Thailand.
Figure 2.
Locations of sample collection of three wild elephant populations, including population A (red dots), population B (yellow dots), and population C (blue dots).
Figure 2.
Locations of sample collection of three wild elephant populations, including population A (red dots), population B (yellow dots), and population C (blue dots).
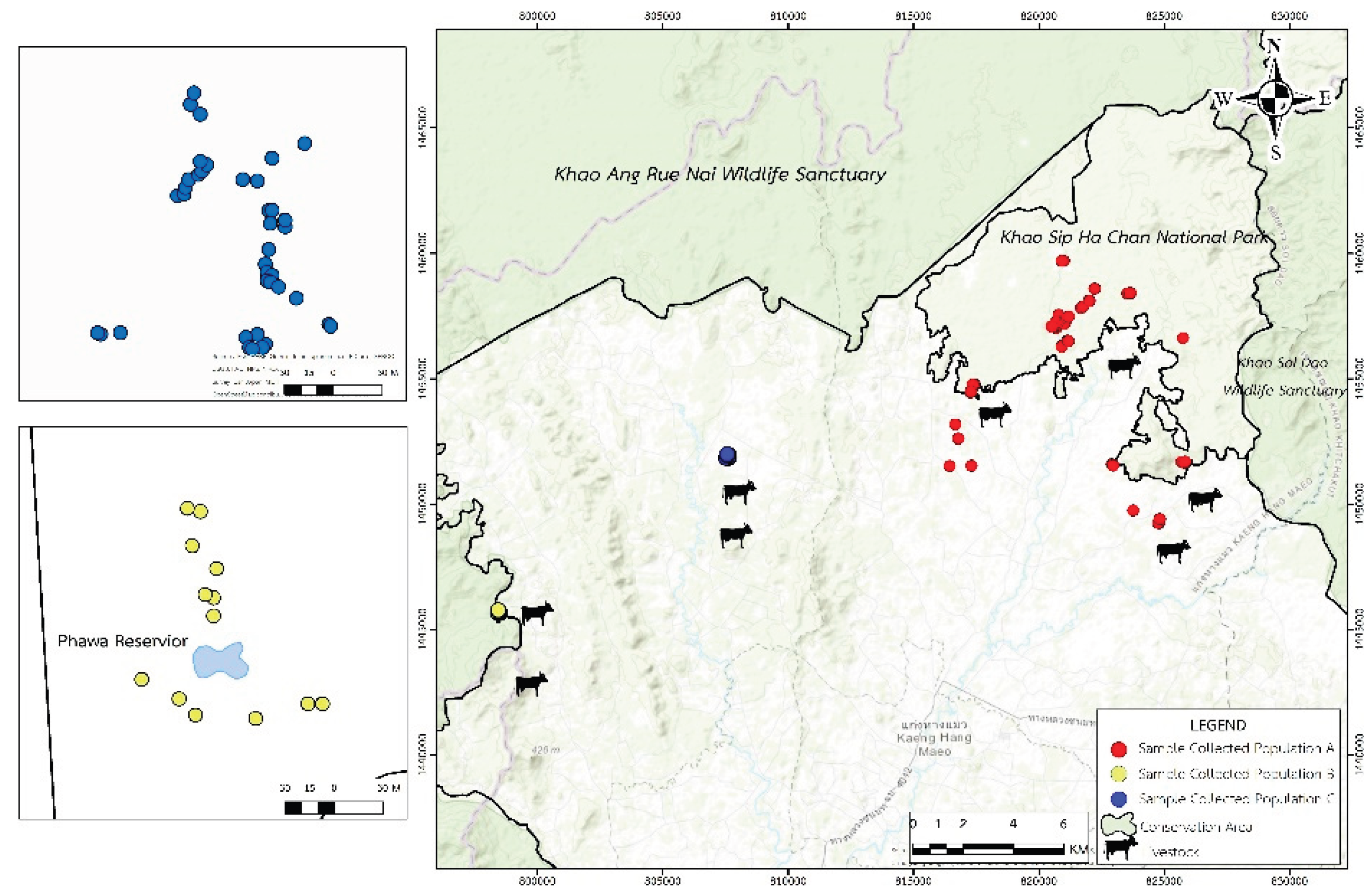
Figure 3.
Photomicrographs of Gastrointestinal Parasites (GIPs) in fecal samples of wild Asian elephants at the national park, as follows: (A, B) Strongyle-type eggs, characterized by thin shells and coarsely granular internal material; (C) Embryonated strongyle-type egg, showing a developing larva within the shell; (D) Embryonated Strongyloides spp. egg, smaller in size with clearly visible larval development; (E, F) Paramphistomum sp. eggs, oval and translucent with loosely packed internal material; (G, H) Fascioloides jacksoni eggs, showing large, oval shapes with smooth shells and granular internal contents. All images captured by light microscopy at 400× magnification. Scale bar in all panels = 50 µm.
Figure 3.
Photomicrographs of Gastrointestinal Parasites (GIPs) in fecal samples of wild Asian elephants at the national park, as follows: (A, B) Strongyle-type eggs, characterized by thin shells and coarsely granular internal material; (C) Embryonated strongyle-type egg, showing a developing larva within the shell; (D) Embryonated Strongyloides spp. egg, smaller in size with clearly visible larval development; (E, F) Paramphistomum sp. eggs, oval and translucent with loosely packed internal material; (G, H) Fascioloides jacksoni eggs, showing large, oval shapes with smooth shells and granular internal contents. All images captured by light microscopy at 400× magnification. Scale bar in all panels = 50 µm.
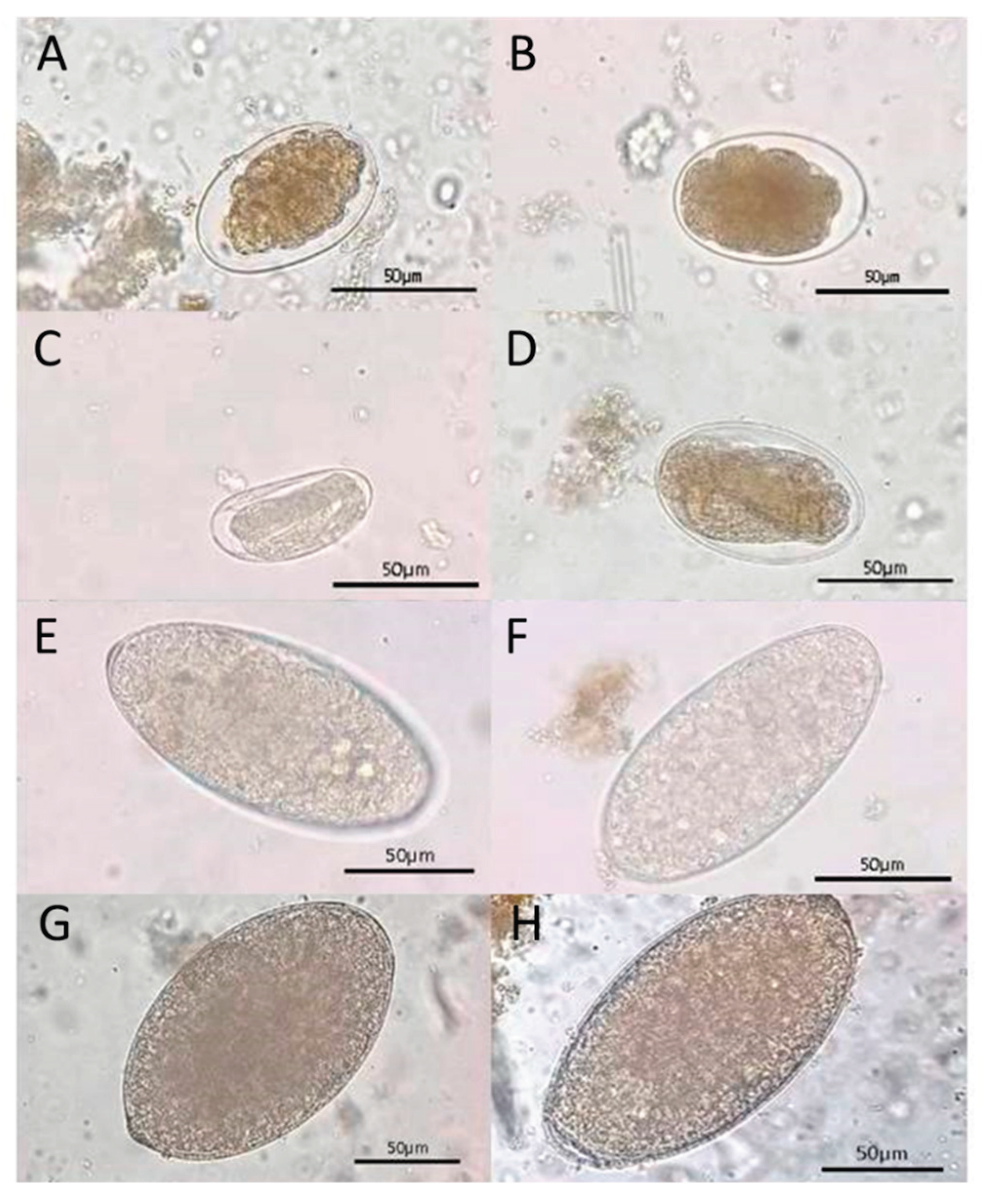
Figure 4.
Photomicrographs of larvae and possible adult nematodes recovered from wild elephant fecal samples, visualized under light microscopy at 400× magnification, as follows: (A) Strongyloides spp. larva, displaying a characteristic coiled posture, thin translucent body, and internal features typical of rhabditiform larvae; (B–F) Non-Strongyloides spp. larvae showing morphological characteristics consistent with strongylid nematodes, including curved or elongated bodies, smooth cuticle, and granular or transparent internal contents. All scale bars = 100 µm.
Figure 4.
Photomicrographs of larvae and possible adult nematodes recovered from wild elephant fecal samples, visualized under light microscopy at 400× magnification, as follows: (A) Strongyloides spp. larva, displaying a characteristic coiled posture, thin translucent body, and internal features typical of rhabditiform larvae; (B–F) Non-Strongyloides spp. larvae showing morphological characteristics consistent with strongylid nematodes, including curved or elongated bodies, smooth cuticle, and granular or transparent internal contents. All scale bars = 100 µm.
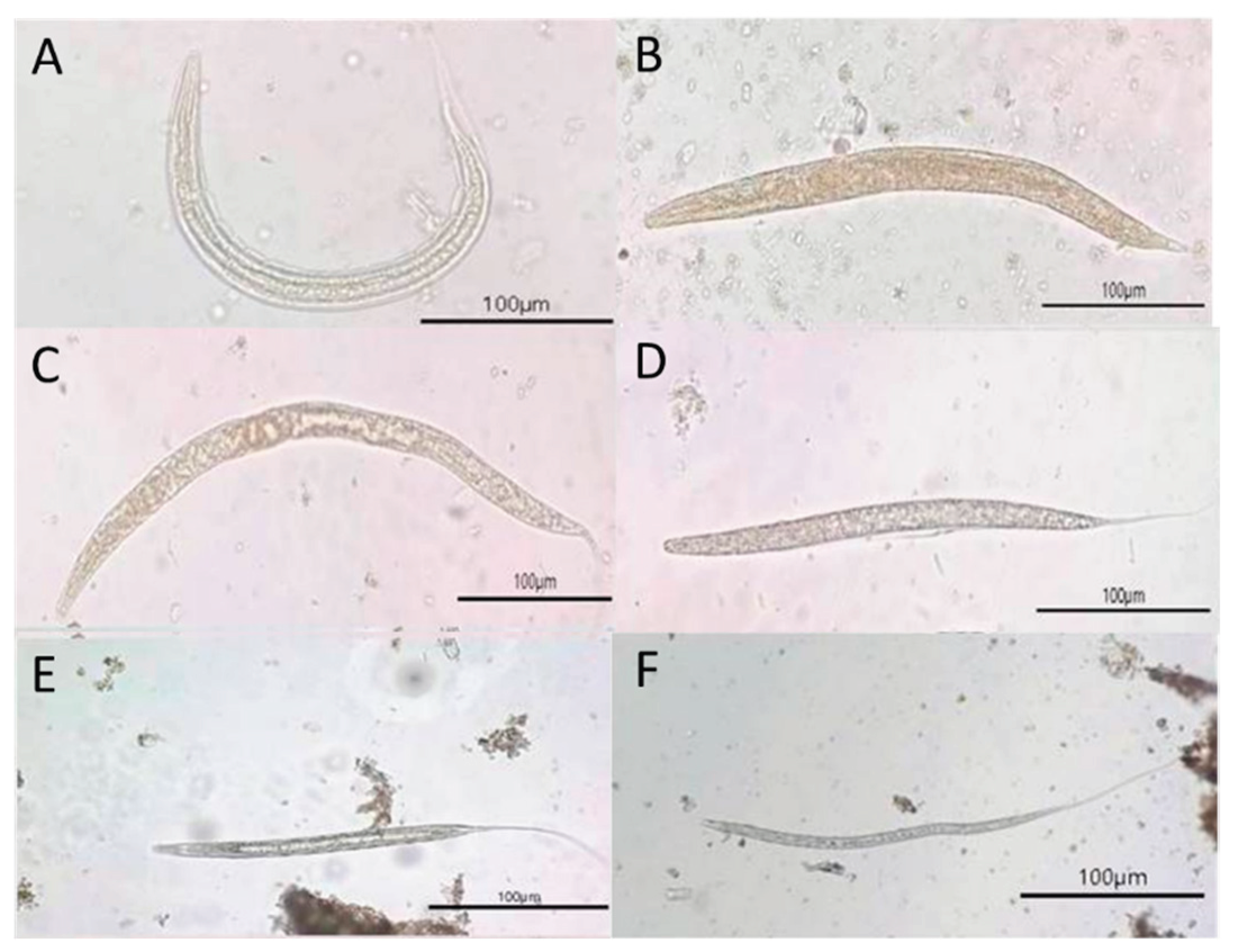
Figure 5.
Distribution of Strongyle-type and Strongyloides spp. egg counts (McMaster method) among three populations of wild Asian elephants.
Figure 5.
Distribution of Strongyle-type and Strongyloides spp. egg counts (McMaster method) among three populations of wild Asian elephants.
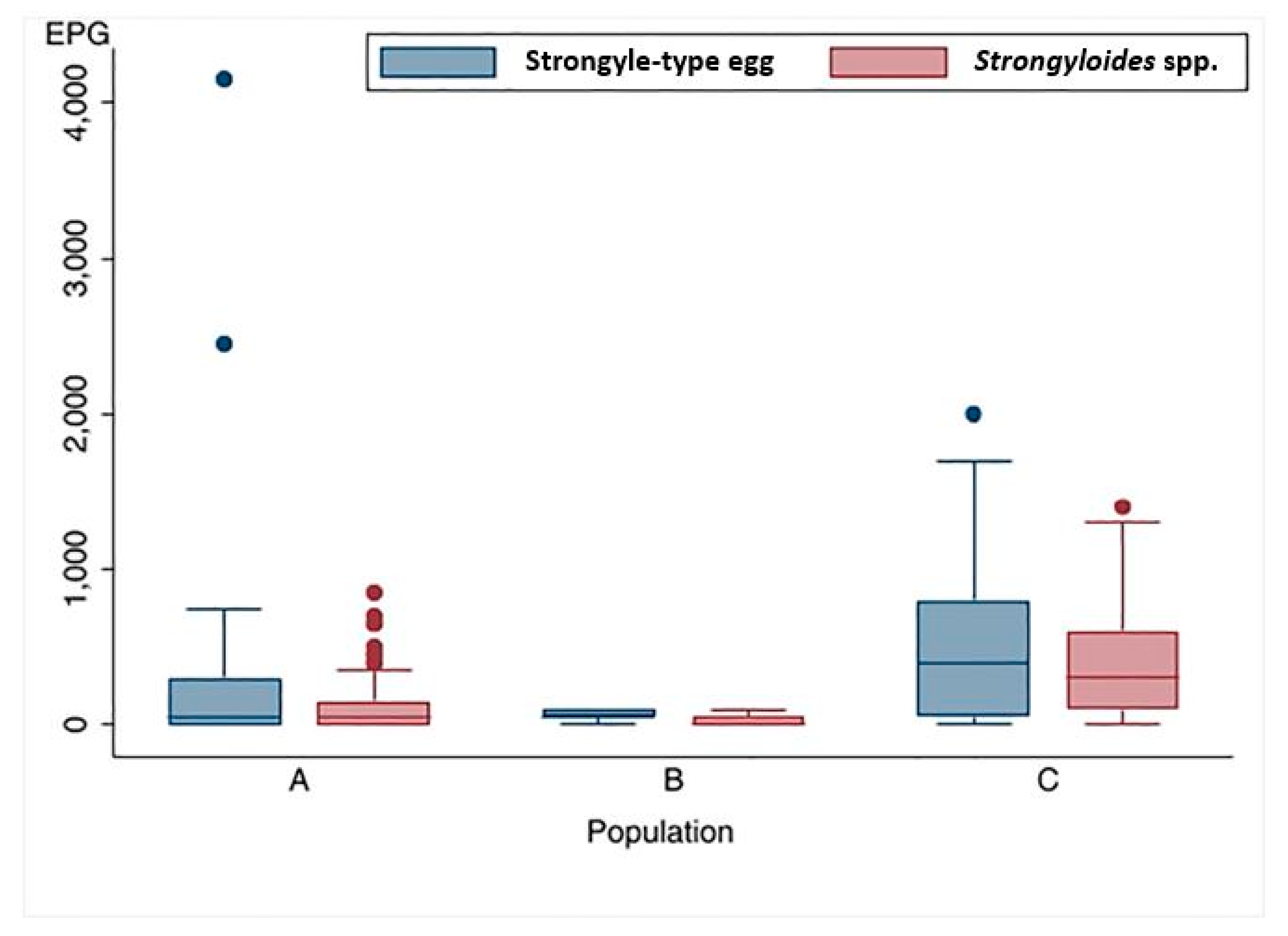
Figure 6.
Distribution of single and mixed infection of GIPs in three populations of Wild Asian elephants.
Figure 6.
Distribution of single and mixed infection of GIPs in three populations of Wild Asian elephants.
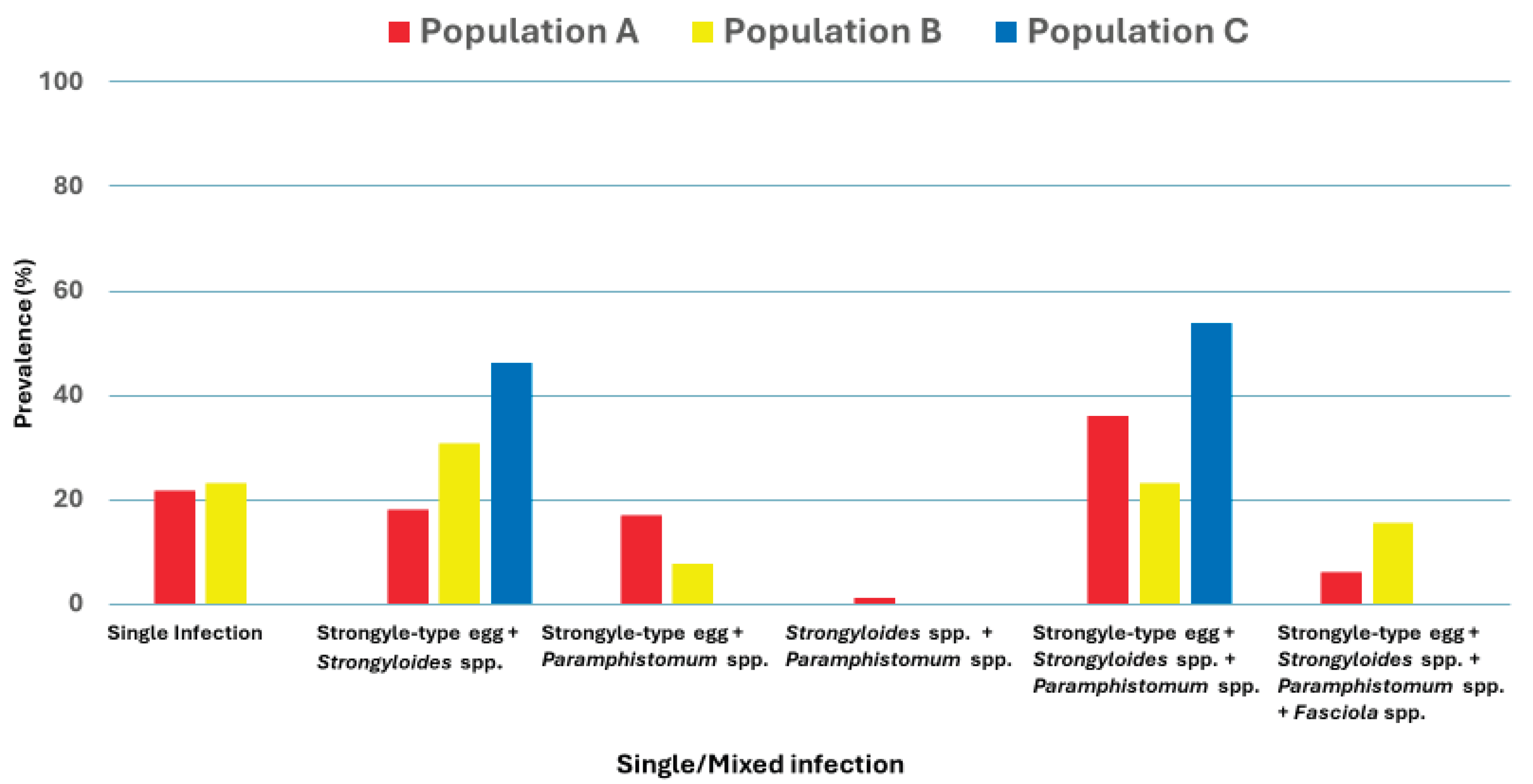

Table 1.
Prevalence of Gastrointestinal Parasites (GIPs) infection in three distinct wild Asian elephant populations.
Table 1.
Prevalence of Gastrointestinal Parasites (GIPs) infection in three distinct wild Asian elephant populations.
| GIPs | Prevalence of GIPs infection | χ2 | p-value | ||||
|---|---|---|---|---|---|---|---|
| Total (N=135) % (95% CI) |
Population A (n=83) % (95% CI) |
Population B (n=13) % (95% CI) |
Population C (n=39) % (95% CI) |
||||
| Egg | Nematode | ||||||
| Strongyle-type | 79.3 (72.4-86.1) | 68.7 (57.5-78.4) | 100 (75.3-100) | 94.9 (82.7-99.4) | 14.84 | 0.001 | |
| Strongyloides spp. | 56.3 (47.9-64.7) | 42.2 (31.4-53.5) | 53.8 (25.1-80.8) | 87.2(72.6-95.7) | 21.88 | < 0.001 | |
| Trematode | |||||||
| Paramphistomum spp. | 61.5 (53.3-69.7) | 67.5 (56.3-77.3) | 46.1 (19.2-74.9) | 53.8 (37.2-70.0) | 3.51 | 0.173 | |
| Fasciola spp. | 5.2 (1.4-8.9) | 6.0 (2.0-13.5) | 15.4 (1.9-45.4) | 0 (0-9.0) | 5.0 | 0.082 | |
| Larval Stage | |||||||
| Strongyloides spp. | 55.6 (47.2-63.9) | 39.8 (29.2-51.1) | 61.5 (31.6-86.1) | 87.2 (72.6-95.7) | 24.37 | < 0.001 | |
| Non-Strongyloides spp. | 83.0 (76.6-89.3) | 72.3 (61.2-81.5) | 100 (75.3-100) | 100 (91.0-100) | 17.37 | < 0.001 | |
| Mixed infection | 84.4 (78.3-90.6) | 78.3 (67.9-86.6) | 76.9 (46.2-95) | 100 (91.0-100) | 10.12 | 0.006 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



