
Submitted:
26 January 2026
Posted:
27 January 2026
You are already at the latest version
Abstract
Muscle protein modification plays a critical role in determining the quality, functional properties, and nutritional value of meat and aquatic products. Over recent decades, non-thermal processing technologies including irradiation, cold plasma, high-pressure processing, ultrasound, and electromagnetic fields have been widely explored in muscle foods. This review aims to critically examine modifications of food proteins subjected to non-thermal processing, with a focus on literature within the last five years. The review first introduces the type and theory of physicochemical modifications of food proteins, which includes protein oxidation, changes in net charge, cross-linking and aggregation. And then the corresponding characterization methods that can be used for food proteins were briefly covered. Finally, the effects of non-thermal processing on muscle proteins were thoroughly discussed. This review will shed light on the complicated mechanism of protein modifications in muscle foods and empower future development of non-thermal processing technologies in muscle foods.
Keywords:
non-thermal processing
; protein oxidation
; protein denaturation
; protein charge
; protein cross-linking
1. Introduction
Muscle foods, such as meat and fish, are highly perishable due to their rich nutrient content and high water activity, which create an ideal environment for microbial growth. Therefore, heat treatment is often required to eliminate harmful microorganisms. However, thermal processing can lead to the degradation of heat-sensitive nutrients such as vitamins and certain amino acids. Moreover, high-temperature processing may result in the formation of potentially hazardous compounds, including heterocyclic amines and polycyclic aromatic hydrocarbons. Excessive oxidation of lipids and proteins is another concern, which have been associated with deterioration in meat quality. In recent decades, non-thermal physical processing technologies has been used to improve the microbial safety, and shelf life of muscle foods without relying on high temperatures [1]. In addition, non-thermal physical processing has been used to improve other quality aspects of muscle foods, such as controlling ice crystals [2,3,4], facilitating marination [5,6,7], tenderizing tough meat [8,9], etc.
In muscle foods, the predominant components (other than water) are protein, and the presence of proteins is often accompanied by oxidizing lipids, reactive sugars, transition metals, etc. These chemical compositions, along with the application of diverse technological processes, make muscle foods a favorable scenario for protein modification [10,11]. While non-thermal processing has been widely applied, its effect on muscle proteins received less attention compared to the effects of bacterial killing/inhibition. Therefore, there is a need to deepen the understanding of the mechanisms, characterization, and consequences of protein modification in muscle foods induced by novel non-thermal physical processing.
The field of non-thermal physical processing is rapidly evolving, and many excellent reviews ([12,13,14,15], to name a few) can be found in the literature about how food proteins are affected by non-thermal physical processing. This review aims to provide an updated overview of the mechanisms and factors governing the physics and chemistry behind food protein modification. The consequences of the selected novel non-thermal processing technologies on muscle proteins will also be covered in detail, with a focus on literature published in the last five years.
2. Theoretical Aspects of Protein Modification in Food System
Many native food proteins are marginally stable, and they are prone to various modifications during storage and processing (Figure 1). A theoretical understanding of protein modification will offer valuable insights about the effects of non-thermal processing on muscle proteins. Here we briefly summarize the main theory in the modification of food proteins.
2.1. Chemical Modifications
For proteins in vivo, various post-translational modifications (PTM) occur, and the PTMs can either activate or inhibit a protein's function, impacting its ability to interact with other molecules or perform its biological role. Common PTMs include phosphorylation, glycosylation, ubiquitination, acetylation, methylation, etc [16]. A relatively new PTM, protein lactylation was discovered on histones in 2019 [17], and Ma et al. [18] recently found that protein lactylation may participate in the meat quality development in pork.
However, many of these active protein modifications are no longer possible or less likely in postmortem muscles when ATP has been used up. During storage and processing, oxidation and the Maillard reaction are two common chemical reactions involving food proteins [19]. Protein oxidation, by a broader definition, refers to all reactions that lead to the removal of electrons from protein. According to Estévez et al. [20], the Maillard reaction is closely linked to protein oxidation. For example, reactive dicarbonyls derived from the Maillard reaction can lead to oxidative deamination of food proteins, and the underlying chemical mechanism has been well discussed [21]. There is a coexistence of proteins, lipids, sugars, transition metals, etc. in many foods, rendering the food proteins susceptible to oxidative stress. In normal viable cells, oxidative stress is well controlled by the dynamic cellular antioxidant system. In contrast, foods generally do not have a dynamic antioxidant system, excessive oxidative stress can occur when the endogenous antioxidants are exhausted. Our previous study showed that during the cold storage of pre-rigor fish muscle, a drastic decrease of intrinsic antioxidants occurred during rigor transformation, accompanied with damage of mitochondria. Moreover, food processing often aggravates protein oxidation, and this has been thoroughly reviewed in meat products [11].
Upon oxidative stress, all amino acids can be oxidized [22]. The thiol group of cysteine residue is highly susceptible to oxidation and loss of thiol groups has been widely used as a marker for protein oxidation in meat [23]. Another marker of protein oxidation is the formation of protein carbonyls, mainly derived from alkaline amino acids, especially lysine [20]. Bao et al. [21] addressed that protein carbonyl is a general term that includes structurally and mechanistically different protein modifications. With extensive oxidation, food proteins can form various intra- and inter-molecular cross-linkages, including disulfide, dityrosine, Schiff-base, and Michael addition products. Excessive intermolecular cross-linking will reduce the protein solubility and contribute to protein aggregation. Other than covalent bonds, the formation of protein aggregates can be induced by hydrophobic interaction and electrostatic interaction. Chi et al. [24] summarized the mechanism and driving forces of protein aggregation in aqueous solution, and the authors pointed out that aggregation depends strongly on factors such as temperature, pH, salt concentration, salt type, surfactant, etc.
2.2. Physical Modifications
2.2.1. Protein Denaturation
Denaturation can be described as a change in the native conformation of a (globular)
protein due to a change in conditions. According to Sanfelice and Temussi [25], native
proteins are just marginally stable as the thermodynamic stability of folded proteins is slightly higher than that of the unfolded ones, and subtle changes in the environment may be sufficient to induce protein denaturation. The most relevant causes for denaturation of food proteins include temperature, pH, solvent composition, interface, or mechanical stress [26].
It is well-known that food proteins undergo thermal denaturation. Although many modern physical processing can be categorized as non-thermal technology, transition local high temperature can occur such as in ultrasound [27]. Compared to thermal denaturation, protein cold denaturation is less well-known. At decreased temperature, protein conformational entropy overtakes the stabilizing hydrophobic effect, resulting in protein unfolding [28].
Balny and Masson [29] pointed out that pressure-induced protein denaturation is a complex phenomenon depending on many factors, including protein structure, pressure, temperature, pH, presence of sugars, salts, etc. Harano et al. [30] indicated that high pressure-induced protein denaturation is driven by an increase of water entropy in the whole system. High pressure leads to a moderately swollen protein structure with a much larger water-accessible surface area as compared to the native protein. Although the translational and rotational motions of water penetrating the protein interior or at the protein surface are largely restricted, translational restriction for water molecules that are sufficiently far from the protein is greatly reduced, therefore the entropy of water in the whole system increases upon pressure denaturation.
It is known that proteins can be denatured by adsorption on interfaces, especially hydrophobic ones [31]. Protein denaturation at interfaces has been extensively studied, and it has been reviewed with regard to different interfaces, including oil-water [32], ice-water [33], air-water [34], etc. As summarized by Walstra and De Roos [34], protein molecules can adsorb onto the interface with different segments, and it is mainly the hydrophobic groups that are adsorbed, and therefore the proteins must change their conformation as most hydrophobic groups are buried in the interior of the native protein. The extent of conformational change depends on the stability of the protein itself, which is affected by temperature, pH, ionic strength, etc. For example, β-casein, an intrinsically disordered protein, unfolds to a considerable extent. Once adsorbed, protein unfolding may continue as the process takes time. The extent of unfolding depends on factors such as available surface area, protein concentration, etc. As the denaturation at the interface is generally irreversible, expansion and subsequent recompression of the interface may lead to significant protein denaturation. Other than these general mechanisms, interfacial areas may have a distinct microenvironment and lead to protein denaturation. One such example can be found in Arsiccio and Pisano [33], where a possible mechanism of ice-induced protein denaturation was discussed.
Gorelov and Morozov [35] used protein monocrystals to study the mechano-chemistry of protein, the ordered packing of molecules allows a uniform distribution of load. It was proposed that extension led to loss of physical contact between globules and stretching the covalent cross-links, and this caused protein denaturation. Proteins experience shear stress during processes such as mixing, centrifugation, pumping, etc. It was demonstrated that high shear force or high shear rate alone only led to slight conformational changes [36]. The same authors suggested that the presence of an air-liquid interface may play a role, and therefore they investigated the combined effects of shear and interface on the denaturation of recombinant human growth hormone (rhGH) and recombinant human deoxyribonuclease (rhDNase). The results showed that aggregation of rhGH only occurred in the presence of air-liquid inerface, and the extent of aggregation increased at higher shear rate. In contrast, rhDNase, a less surface-active protein as compared to rhGH, did not aggregate even at high shear. When surfactant was used, aggregation of rhGH was significantly reduced, likely due to the competitive occupation of the interface by the surfactant. Jaspe and Hagen [37] tested whether a high shear can destabilize horse cytochrome c, a small globular protein. No evidence of protein denaturation was observed even at the highest shear rates. The authors suggested that very high-molecular-weight protein such as multimeric proteins, or high solvent viscosity may result in protein denaturation under high shear.
According to Bennett et al. [38], 3D domain swapping is one mechanism for protein oligomerization (Figure 2). In the process, the folded monomer is subjected to temporary conditions that favor partial unfolding. Then, when conditions were restored, one domain in the open monomer may be replaced by the same domain from another identical protein, and results dimer or higher oligomer, especially at high protein concentration. The swapped “domain” can be a large tertiary globular domain or a small α-helix or β-sheet. One such example was given by Bennett et al. [39], where freezing-thawing led to dimerization of diphtheria toxin. It was suggested that freezing led to a pH drop of the phosphate buffer, which converted the protein into an open monomer, and the open monomers formed domain-swapped dimers when pH returned to normal during thawing.
2.2.2. Protein Net Charge
Some of the amino acid sidechains can carry positive (His, Lys, Arg) or negative charges (Glu, Asp, Cys, Tyr), depending on their pKa values and the pH of the surroundings. Protein charge is largely determined by the amount of charged amino acid residues. The contributions from tightly bound metal, solvent, buffer and co-solvent ions are also significant [40]. According to Gitlin et al. [41], protein charge influences the structure, stability, activity and functionality of proteins. Protein charge can be affected by various chemical modifications induced by physical processing. The most frequent modification is protein oxidation, where a range of charged amino acid residues were affected. Protein carbonylation, one common indicator of protein oxidation, often led to a decrease of positively charged amino acids (Lys, Arg, His). This would alter the net charge, and Yu et al. [42] showed that oxidation can either increase the net negative charge (surrounding pH > pI) or decrease the net positive charge (surrounding pH < pI) of myofibrillar proteins. According to de Graff et al. [43], modifying just one charge of highly-charged proteins with smaller radii of gyration can lead to a great change in protein stability. The relationship between protein oxidation and protein charge in muscle proteins has been thoroughly discussed by [44]. Dignon et al. [45] pointed out that in addition to net charges, the arrangement of charged amino acids (charge patterning) has an important role in determining the propensity of proteins to self-associate, aggregate, or phase separate.
3. Characterization of Common Food Protein Modification
3.1. Sidechain Modification
The most common assays for the determination of protein oxidation in meat have focused on thiol groups and carbonyl groups. A thiol (R-SH) is a carbon-bonded sulfhydryl group and it is highly susceptible to oxidation. Loss of the thiol group is widely used as an indicator of protein oxidation. The predominant method for the detection of thiol groups is a spectrophotometric method based on DTNB [46]. A free thiol reacts with DTNB and forms TNB anion, which absorbs strongly at 412nm. Different values for the extinction coefficient of TNB anion at 412 nm, ranging from 13,600 M-1 cm-1 to 14,150 M-1 cm-1, were reported in the literature [46,47,48,49]. According to Riddles et al. [47], the purified TNB has a value of 14,150 M-1 cm-1 and this value varies depending on factors like pH and buffer system. As an alternative way to quantify thiol groups, a standard curve can be prepared using L-cysteine [50]. Alternative chemicals/methods exist for analyzing thiol groups, such as aromatic disulfide reagents, fluorescent thiol reagents, and thiol reagents that significantly increase mole. This has been thoroughly discussed in a previous review [51].
A carbonyl group (C=O) is a functional group composed of a carbon double-bonded to an oxygen atom. Formation of protein carbonyls has been widely used as a maker for protein oxidation in meat [52]. The original method for carbonyl determination [53] has been adopted and modified to optimize the detection of carbonyls in muscle/meat proteins [54,55,56]. Like many other tissues, meat is a complex, and some of its components may interfere with the measurement of the actual oxidation extent on the protein fraction. Therefore, the method for protein-bound carbonyl determination in meat usually involves protein separation steps such as precipitation of proteins with cold TCA solution. However, oxidation of proteins may result in the release of carbonyls other than those bound to proteins [57], leading to an underestimation of the carbonyl groups. Another challenging aspect is to solubilize the oxidized proteins. Myofibrillar proteins and connective tissue proteins by nature are difficult to dissolve, and oxidation of those proteins results in even lower solubility.
According to Estévez et al. [20], the traditional DNPH-based method measures total protein carbonyls, which are formed by diverse and nonspecific pathways. Protein carbonylation can be induced by free radicals, lipid oxidation, and the Maillard reaction (glycoxidation). Both primary (mainly linked to the radical pathway) and secondary protein carbonyls (mainly linked to lipid oxidation pathway) can be formed in food proteins, and the Maillard reaction-induced protein carbonylation has received less attention and is worth investigating. To gain specificity and information of a mechanistic nature, the analysis of specific protein carbonyls such as α-aminoadipic semialdehyde (AAS) and γ-glutamic semialdehyde (GGS) should be performed.
3.2. Protein Net Charge
Measuring the net charge of a protein is critical for understanding its solubility, stability, and functionality in various foods. One common method for assessing protein net charge is based on electrophoretic mobility, particularly through techniques such as zeta potential analysis. In an electric field, proteins migrate depending on their net charge [58]. This method is sensitive to the ionic strength and pH of the buffer. Another classical approach involves titration-based methods, such as potentiometric titration or electrophoretic titration. These techniques determine how a protein uptakes or releases protons as the pH is gradually changed. Zahler and Shaw [40] pointed out that some of the earliest measurements using pH titrations ignored the tightly bound cations and anions, there might be pH-dependent conformational changes (hence the changes in pKa of residues). As the protein net charge depends on the isoelectric point (pI), Isoelectric focusing gel electrophoresis, which separates proteins based on their pI in a pH gradient, is also widely used for evaluating net charge. And this technique can be coupled with western-blot or MS/MS to further identify the proteins [44].
Polyelectrolyte titration offers easy access to the determination of the surface charge of proteins and other biopolymers. In general, the protein solutions were incubated in excess with an oppositely charged polyelectrolyte, and the residual amount was back-titrated using o-toluidine blue for endpoint detection [59]. By incubating myofibrils with different dyes (positively charged Sarfarin O, neutral bromophenol blue, and negatively charged Orange G), Yu et al. [42] developed a method to estimate the net charges of myofibrils based on the amount of bound dyes. The net charge can also be estimated at the myofilament level based on the determination of Donnan potential [60]. The above-mentioned classical methods like electrophoresis and titration remain useful, while cutting-edge tools such as charge-detection MS [61] and computational predictions [62] are redefining the accuracy and efficiency of charge characterization.
3.3. Structural Characterization
Assessing protein secondary and advanced (tertiary and quaternary) structures is fundamental, as the techno-functionality of protein is tightly coupled to its structure. Various biophysical or spectroscopic methods including Circular dichroism spectroscopy, Fourier-transform infrared spectroscopy, Raman, intrinsic fluorescence, NMR, X-ray crystallography, and cryo-electron microscopy, have been developed to characterize these structural features. A certain technique only works for a special protein system. Collagen has a special triple-helix structure, and therefore, optical rotation can be employed to characterize the structural changes during gelatin gelation [63].
In the field of food science, these techniques can offer more mechanistic insights if experimental results are well interpreted. For example, in the analysis of intrinsic fluorescence, Reshetnyak and Burstein [64] suggested that the total emission of Trp fluorescence can be attributed to four discrete classes of tryptophan residues, designated S, I, II, and III. Classes S and I correspond to residues located in the interior of the protein, while classes II (contact bound water) and III (contact free water) correspond to the tryptophan residues located on the molecular surface (Figure 3). These authors suggested that an increased contribution from class S led to a blue shift of the fluorescence spectrum. Another example is in the determination of secondary structures using the Raman Spectrum of Proteins, where ranges other than the widely used 1600–1700 cm-1 can provide additional structural features, such as disulfide stretch around 500 cm-1 [65]. Zhu et al. [66] quantified the distribution of the disulfide bond conformations (gauche-gauche-gauche, trans-gauche-gauche, and trans-gauche-trans) and thereby obtained mechanistic insights about why garlic juice and allicin can stabilize meringues.
In recent years, computational approaches have dramatically advanced the field. The development of AlphaFold2 and RoseTTAFold has revolutionized tertiary structure prediction from sequence data. These AI-driven models have achieved near-experimental accuracy in many cases and are now widely used to complement experimental methods. One thing worth pointing out is that in food systems, proteins would seldom exist in the state of thermodynamic equilibrium, while this is often the presumption of many computational approaches.
3.4. Protein Cross-Links and Aggregation
Assessing protein cross-linking and aggregation is essential in studying protein functionality. Cross-linking refers to the formation of covalent bonds between amino acid residues either within a single protein or between different protein molecules, while protein aggregation involves the association of protein molecules into larger assemblies through non-covalent or covalent interactions.
Upon oxidation, muscle proteins can form various cross-linked products and disulfide is one of the main cross-linking type found in meat [23]. Non-disulfide type of cross-linking, including dityrosine and carbonyl-involved cross-link, occurs in meat system as well. Analysis of disulfides is based on the analysis of thiol groups. The detection of disulfides usually involves a series of steps: thiol blocking, disulfide reduction, and detection of newly formed thiols. In Raman spectroscopy, deconvolution of the signal between 490 and 550 cm−1 provides information about the relative abundance of each disulfide conformation: from most to least energetically stable: SSg-g-g (496–513 cm−1), SSt-g-g (514–527 cm−1), and SSt-g-t (527–548 cm−1) [66].
Protein cross-linking via a covalent bond leads to increased molecular weight and therefore, cross-linked proteins migrated to a higher position in polyacrimide gels. By running SDS-PAGE with and without a reducing agent (reduce the disulfide bond), information about the cross-linking type can be obtained. Disulfide cross-linking can be both intra- and inter-molecular. Diagonal PAGE has been used as a tool to distinguish the two types of protein cross-linking [67]. The diagonal PAGE is a 2D gel which separates proteins based on molecular weight first under non-reducing conditions and then under reducing conditions. This technique has been used to study protein cross-linking in high-oxygen packaged beef [68,69]. Some of the myofibrillar proteins (e.g., titin and nebulin) have a very large molecular weight and low solubility. Inter-molecular protein cross-linking leads to even larger and less soluble molecules, thereby making the analysis of oxidized proteins by gel electrophoresis rather difficult. Warren et al. [70] developed a vertical agarose gel electrophoresis system that allows separation of titin and other high molecular weight products. Another way to separate and analyze a large protein complex can be achieved with field-flow fractionation techniques [71].
4. Effects of Non-Thermal Processing on Muscle Proteins
Over the years, non-thermal processing technology has been widely applied in muscle foods, here we will focus on more recent studies about non-thermal processing, and emphasize their effects on muscle protein (Table 1). For a more complete background knowledge on how non-thermal processing affects protein functionality, readers are encouraged to refer to the following excellent reviews (Irradiation: [72,73]; ultrasound: [74,75], HPP: [76,77]; cold plasma: [78,79]. electromagnetic field [80,81]; just name a few examples).
4.1. Ultrasound
Ultrasound technology has gained increasing attention as a non-thermal method for modifying food proteins. It primarily uses high-frequency sound waves (typically 20 kHz–100 kHz) to induce physical and chemical changes in protein structures. Ultrasound generates acoustic cavitation and it can lead to localized high temperature and pressure, shear forces, and and the generation of free radicals. As discussed earlier in section 2, all these three aspects (high temperature/pressure, shear force, free radicals) can result in protein modification, being physical or chemical.
Ultrasound has been used in combination with other processes to enhance mass/heat transfer. For example, ultrasound technology has been used to facilitate protein extraction [82], marination [137], freezing [138], thawing [139], etc. It has also been widely used as a homogenization technique to improve colloidal stability. Gul et al. [140] applied ultrasound and high pressure to improve the stability of sesame paste against phase separation, and the results showed that oxidative stability was also enhanced.
The intense physical conditions generated by ultrasound can lead to chemical modifications of amino acid side chains, particularly those that are reactive or surface-exposed. Sidechain modification has been evidenced by decreased thiol groups [82,96,98] and increased carbonyl groups [86,94] in meat proteins. In some studies, an increase of thiol groups was reported [84,87], which may be due to the newly exposed thiol groups upon protein unfolding. The protein unfolding process may also account for the increased surface hydrophobicity [85,99]. Zeta-potential was found to be increased after ultrasound treatment [99,103]. Cavitation-induced shear and pressure disrupt non-covalent interactions such as hydrogen bonds, hydrophobic interactions, and van der Waals forces, and thereby significantly alter the advanced structure of food proteins. However, there is no general consensus in the literature about how ultrasound affect secondary structure of myofibrillar proteins [83,88,89,93]. Despite of this, particle size was decreased, which may be beneficial for solubility and functionality [92,97]. However, excessive ultrasound intensity or prolonged treatment may lead to irreversible denaturation or loss of functionality [82,86,95].
During ultrasound processing, the enhanced mass/heat transfer and hence shorter processing time should result in limited protein modification, while the cavitation effect generally led to protein modification. Recent developments include the use of multi-frequency ultrasound systems to fine-tune the balance between the two opposing effects on protein modification. In the ultrasound-assisted freezing of shrimps, Xu et al. [138] combined different frequencies of ultrasound and achieved faster freezing and less ultra-structural damage of muscle tissue.
4.2. Electromagnetic Fields
Electric Fields and Magnetic Fields fall under the umbrella of electromagnetism, and both can be applied in static-, alternating-, and pulsed fields. The electromagnetic field has been widely used in meat or meat products. A large proportion of the research focused on the control of ice crystals since the electromagnetic field can interact with the water dipole. For example,the magnetic field is believed to be able to break large water clusters into smaller ones and strengthen the hydrogen bonding between water and proteins (hydration shell). This is critical for protein stability.
On top of the dipole polarization of H2O, Sun et al. [141] argued that ions moving under an electric field is the main factor influencing the properties of NaCl solution during freezing. Therefore, redistribution of ions should be considered in dealing with protein functionality under electromagnetic fields. Other than ions and small polar molecules, the electric field can interact directly with the dipole moments of amino acid residues, forcing them to align with the field and therefore unfold the proteins. These effects are highly dependent on field strength, duration, and protein type.
Unlike Electric Fields, Magnetic Fields do not generate heat or violently rupture bonds. Instead, they influence the spatial arrangement of molecules. A strong magnetic field can force peptide chains to align parallel or perpendicular to the field, promoting ordered structures rather than random aggregation. Magnetic fields promote the unfolding of MP and sarcoplasmic proteins like myoglobin, indicated by a general increase in reactive sulfhydryls and surface hydrophobicity. Combining the magnetic field with additives like sodium tripolyphosphate (STPP) or calcium chloride can further optimize protein functionality. For example, AMF electromagnetic fields and STPP work together to enhance protein phosphorylation and solubility in low-salt systems. Wu et al. [106] suggested that a magnetic field could facilitate the rearrangement of the Mb structure, resulting in the transfer of its internal reactive groups to the external environment. This contributed to hydration and cross-linking between MP. Magnetic field can influence the spin state of electrons, potentially slowing down oxidative reactions involving free radicals.
4.3. High-Pressure Processing (HPP)
High-pressure processing (HPP) fundamentally alters muscle proteins by disrupting the delicate balance of intermolecular forces (hydrogen bonds, hydrophobic interactions, and ionic bonds) that hold them in their native structures. HPP can lead to the depolymerization of muscle filaments, releasing free myosin and G-actin [142]. With increasing pressure, the quaternary and tertiary structures begin to collapse while at very high pressures, secondary structure such as the α-helix often transforms into random coils or β-sheets, which favors protein aggreation [115,118]. Zhang et al. [143] explored the effects of HPP on myofibrillar proteins. Their results showed that the quaternary structure dissociates when treated at moderate pressures (100–200 MPa), the tertiary structure is significantly affected at a pressure level above 200 MPa, and secondary structure changes take place at higher pressures (300–700 MPa). These advanced structural changes lead to morphological changes at the microscopic level. For example, Xue et al. [144] showed that pressure greater than 200 MPa induced dimerization and swelling of myosin molecules extracted from rabbit muscle. Xue et al. [145] further suggested that different subfragments of myosin respond to HPP greater than 200 MPa differently, there was slight changes in light meromyosin as compared to heavy meromyosin.
Similar to myofibrillar proteins, sarcoplasmic proteins (globular proteins are highly sensitive to HPP) will denature with HPP. Myoglobin concentration decreased with increased HPP levels. Deoxymyoglobin and metmyoglobin content were greater at 600 MPa compared with non-pressurized control and other HPP levels [146]. This is the primary driver for changes in meat color and light scattering of HPP meat products.
High-Pressure Processing (HPP) can generate free radicals, especially at higher pressures (e.g., above 400-600 MPa) and temperatures, leading to oxidative stress, particularly in lipid-rich foods such as meat. By disrupting cell membranes and releasing catalytic iron, HPP initiates lipid and protein oxidation. Decrease of total thiols [110,111,114,115] and increase of disulfide bond [112] have been reported.
4.4. Irradiation
In food processing, non-thermal irradiation technology generally includes gamma irradiation, electron beam irradiation and UV irradiation. These technologies have been widely used to destroy bacteria, viruses, or insects that may be present in the food. Irradiation may promote reactions within the food components due to the effect of the electron beam or free radicals from the radiolysis of water [147]. As can be seen in Table 1, irradiation may result in various physicochemical changes in muscle proteins, including denaturation, oxidation, aggregation, and fragmentation, etc. These physicochemical changes, at a mild level of irradiation, often led to improved protein functionality. Irradiation of myofibrillar proteins extracted from various sources has been shown to improve protein functionalities such as gelling, foaming, and emulsifying [120,122,124]. Both the protein and the irradiation source have an impact. As compared with myofibrillar proteins, the extracted myosin was more easily destroyed [121]. Compared with γ- ray irradiation, the electron-beam irradiation had a greater impact on the physicochemical properties of MPS and MS [121]. Patil and Khandekar [148] pointed out that LED technology will be an upcoming non-thermal food preservation method, which utilize ROS to primarily target proteins and lipids.
Dąbrowska-Gralak et al. [149] found that the thermal stability of collagen type-I was not significantly affected upon irradiation at a dose range up to 50 kGy. However, oxidative modification was observed evidenced by the introduction of new oxygen-containing functional groups in collagen. And the authors suggested that humidity played a key role regarding the amount and the rate of oxidative damage. Stanca et al. [150] compared the effects of gamma irradiation on the structure and stability of keratin, collagen, and gelatin. Results showed that irradiation led to an increase in random coil structures of keratin at the expense of β-sheet structures, accompanied by a decreased thermal stability. In contrast, no significant change in structure or stability was found for irradiated collagen and gelatin. The authors suggested that proteins with higher degree of ordered structures are more stable against gamma irradiation.
4.5. Cold Plasma
Plasma is composed of reactive species (ROS/RNS), charged particles, neutral atoms, etc., and it is commonly categorized into non-thermal (cold) and thermal types. The former is particularly suitable for heat-sensitive materials due to its low-temperature operation [151]. The generation of RNS is of particular interest to the red meat industry, as it can replace or reduce the use of nitrate/nitrite in meat products. ROS and RNS readily oxidize susceptible amino acids, particularly those with sulfhydryl (thiol) groups like cysteine and often leads to the conversion of sulfhydryl groups into disulfide bonds, promoting protein cross-linking [152]. At higher intensity of cold plasma treatments, severe protein oxidation can occur, which is often suggested by the formation of protein carbonyls. The decrease of thiol groups and the increase of carbonyl groups in muscle proteins following cold plasma treatments have been widely reported [79,130,135]. Jiang et al. [128] also detected the formation of dityrosine in duck myofibrillar proteins. All these oxidative damages are associated with protein aggregation and loss of solubility, the latter being indicated by increased particle size [125,136]. Cold plasma treatment alters both the secondary and tertiary structures of meat proteins. Typically, a decrease in α-helix content and a concurrent increase in β-sheets and random coils were observed. This transition indicates an unfolding of the rigid protein structure, making it more flexible [153]. The protein unfolding is supported by the observation that the surface hydrophobicity of myofibrillar proteins became larger after the treatments [131,133,154]. In contrast, surface hydrophobicity was found to be reduced after cold plasma treatment [129,136], possibly due to the hydrophobic interaction among unfolded myofibrillar proteins.
Both the sidechain modification and structural changes are vital for the techno-functionality of cold-plasma modified muscle proteins. The effect on solubility is dose-dependent: Mild treatment can increase solubility by exposing hydrophilic groups and causing partial unfolding that facilitates water interaction [155], while severe treatment leads to excessive oxidation and extensive cross-linking, resulting in the formation of large, insoluble aggregates and reduced solubility. Cold plasma has shown promise in improving the gelling properties of meat proteins, particularly in surimi [156,157,158] and myofibrillar protein isolates [126,128].
A moderate "opening up" of the protein structure also allow a better adsorption at oil-water or air-water interfaces, which is beneficial for emulsification and foaming. Despite the potential in enhancing techno-functionality of muscle proteins, cold plasma may negatively impact sensory qualities such as the oxidation of myoglobin to metmyoglobin, leading to surface discoloration in red meats like beef [159].
5. Conclusion
The transition toward non-thermal physical processing represents a significant shift in meat and aquatic research and industry. It is well-known that non-thermal processing technologies, such as irradiation, ultrasound, cold plasma, high-pressure processing (HPP), and electromagnetic fields, offer a pathway to enhance microbial safety and preserve the nutritional integrity of muscle foods. The less well-known (or often ignored) aspect about non-thermal processing is that the molecular landscape of muscle proteins can undergo significant alterations, both chemically (oxidation, the Maillard reaction, etc.) and physically (denaturation, aggregation, etc.).
While excessive processing can lead to detrimental aggregation and loss of solubility, mild to moderate non-thermal treatments often improve gelling, foaming, and emulsifying properties by inducing partial unfolding and exposing reactive functional groups. The phenomenon has been widely reported, yet the threshold for “excessive processing” is far from clear. By applying advanced characterization tools and AI-driven modelling tools, the underlying mechanism for muscle protein functionality will be approached. A deeper theoretical understanding of how non-thermal energy interacts with muscle proteins (myofibrillar and sarcoplasmic proteins, extracellular proteins such as collagen and elastin) will empower the development of the next generation of meat and aquatic products, balancing safety, functionality, and consumer appeal.
Author Contributions
Conceptualization, Y.B. and Y.Z.; writing—original draft preparation, Y.B. and H.G.; writing—review and editing, H.H; L.Z., Y. Z., and Y. Z; visualization, W.X.; project administration, Y.B.; funding acquisition, Y.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Academic Degree and Postgraduate Education Reform Project of Jiangsu Province, grant number (KYCX25_4279).
Acknowledgments
The authors would like to acknowledge the financial support from the National Natural Science Foundation of China (32202227), the China Postdoctoral Science Foundation (2023M731380), and the Senior Talent Program of Jiangsu University (21JDG044). Conflicts of Interest: The authors declare no conflicts of interest.
References
- Li, H.; Li, C.; Shoaib, M.; Zhang, W.; Murugesan, A. Advances in Non-Thermal Processing of Meat and Monitoring Meat Protein Gels Through Vibrational Spectroscopy. Foods 2025, 14, 1929. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shi, J.; Huang, X.; Zou, X.; Li, Z.; Zhang, D.; Zhang, W.; Xu, Y. Effects of pulsed electric field on freeze-thaw quality of Atlantic salmon. Innov. Food Sci. Emerg. Technol. 2020, 65, 102454. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Y.; Zhu, F.; Yang, J.; Ma, X.; Lou, Y.; Li, Y. The effects of freezing under a high-voltage electrostatic field on ice crystals formation, physicochemical indices, and bacterial communities of shrimp (Solenocera melantho). Food Control. 2022, 142, 109238. [Google Scholar] [CrossRef]
- Wei, Q.; Sun, Q.; Dong, X.; Kong, B.; Ji, H.; Liu, S. Effect of static magnetic field-assisted freezing at different temperatures on muscle quality of pacific white shrimp (Litopenaeus vannamei). Food Chem. 2024, 438, 138041. [Google Scholar] [CrossRef]
- Guo, L.; Zhang, X.; Guo, Y.; Chen, Z.; Ma, H. Evaluation of ultrasonic-assisted pickling with different frequencies on NaCl transport, impedance properties, and microstructure in pork. Food Chem. 2024, 430, 137003. [Google Scholar] [CrossRef]
- Li, C.; Shi, J.; Zhai, X.; Yang, Z.; Huang, X.; Li, Z.; Li, Y.; Zou, X. Effects of Pulsed Pressure Curing on Beef Quality. Foods 2023, 12, 656. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, L.; Chen, Z.; Ma, H. Tri-frequency simultaneous ultrasound pickling for the acceleration of the NaCl content and quality improvement of pork (longissimus dorsi). J. Sci. Food Agric. 2024, 104, 6242–6251. [Google Scholar] [CrossRef]
- Jeong, S.-H.; Jung, Y.-M.; Kim, S.; Kim, J.-H.; Yeo, H.; Lee, D.-U. Tenderization of Beef Semitendinosus Muscle by Pulsed Electric Field Treatment with a Direct Contact Chamber and Its Impact on Proteolysis and Physicochemical Properties. Foods 2023, 12, 430. [Google Scholar] [CrossRef]
- Gao, S.; Xu, Z.; Wang, H.; Xu, A.; Huan, C.; Guo, X.; Liu, R.; Wu, P.; Meng, X. Effect of ultrasonic processing on beef tenderness in longissimus lumborum during aging by proteomics analysis. J. Food Compos. Anal. 2024, 131, 106220. [Google Scholar] [CrossRef]
- Bao, Y.; Ertbjerg, P.; Estévez, M.; Yuan, L.; Gao, R. Freezing of meat and aquatic food: Underlying mechanisms and implications on protein oxidation. Compr. Rev. Food. Sci. Food Saf. 2021, 20, 5548–5569. [Google Scholar] [CrossRef] [PubMed]
- Soladoye, O.P.; Juárez, M.L.; Aalhus, J.L.; Shand, P.; Estévez, M. Protein Oxidation in Processed Meat: Mechanisms and Potential Implications on Human Health. Compr. Rev. Food. Sci. Food Saf. 2015, 14, 106–122. [Google Scholar] [CrossRef] [PubMed]
- Indiarto, R.; Irawan, A.N.; Subroto, E. Meat Irradiation: A Comprehensive Review of Its Impact on Food Quality and Safety. Foods 2023, 12, 1845. [Google Scholar] [CrossRef]
- Kang, D.; Zhang, W.; Lorenzo, J.M.; Chen, X. Structural and functional modification of food proteins by high power ultrasound and its application in meat processing. Crit. Rev. Food Sci. Nutr. 2021, 61, 1914–1933. [Google Scholar] [CrossRef]
- Pereira, R.N.; Rodrigues, R.; Avelar, Z.; Leite, A.C.; Leal, R.; Pereira, R.S.; Vicente, A. Electrical Fields in the Processing of Protein-Based Foods. Foods 2024, 13, 577. [Google Scholar] [CrossRef]
- Wang, W.; Yang, P.; Rao, L.; Zhao, L.; Wu, X.; Wang, Y.; Liao, X. Effect of high hydrostatic pressure processing on the structure, functionality, and nutritional properties of food proteins: A review. Compr. Rev. Food. Sci. Food Saf. 2022, 21, 4640–4682. [Google Scholar] [CrossRef]
- Ramazi, S.; Zahiri, J. Post-translational modifications in proteins: resources, tools and prediction methods. Database 2021, 2021, baab012. [Google Scholar] [CrossRef]
- Zhang, D.; Tang, Z.; Huang, H.; Zhou, G.; Cui, C.; Weng, Y.; Liu, W.; Kim, S.; Lee, S.; Perez-Neut, M.; Ding, J.; Czyz, D.; Hu, R.; Ye, Z.; He, M.; Zheng, Y.G.; Shuman, H.A.; Dai, L.; Ren, B.; Roeder, R.G.; Becker, L.; Zhao, Y. Metabolic regulation of gene expression by histone lactylation. Nature 2019, 5747779, 575–580. [Google Scholar] [CrossRef]
- Ma, C.; Zhang, W.; Xing, L. Differences in protein lactylation between pale, soft and exudative and red, firm and non-exudative pork. Meat Sci. 2025, 221, 109736. [Google Scholar] [CrossRef]
- Poojary, M.M.; Lund, M.N. Chemical Stability of Proteins in Foods: Oxidation and the Maillard Reaction. Annu. Rev. Food Sci. Technol. 2022, 13, 35–58. [Google Scholar] [CrossRef] [PubMed]
- Estévez, M.; Díaz-Velasco, S.; Martínez, R. Protein carbonylation in food and nutrition: a concise update. Amino Acids 2022, 54, 559–573. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Gou, H.; Sánchez-Terrón, G.; Zhang, L.; Zamora, R.; Hidalgo, F.J.; Estévez, M. Reactions between dicarbonyls and food proteins: when lipid oxidation and the maillard reaction meet protein oxidation. Food Chem. 2025, 495, 146446. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J.; Dean, R.T. Radical-Mediated Protein Oxidation: From Chemistry to Medicine; Oxford University Press, 1997. [Google Scholar]
- Lund, M.N.; Heinonen, M.; Baron, C.P.; Estévez, M. Protein oxidation in muscle foods: A review. Mol. Nutr. Food Res. 2011, 55, 83–95. [Google Scholar] [CrossRef]
- Chi, E.Y.; Krishnan, S.; Randolph, T.W.; Carpenter, J.F. Physical Stability of Proteins in Aqueous Solution: Mechanism and Driving Forces in Nonnative Protein Aggregation. Pharm. Res. 2003, 20, 1325–1336. [Google Scholar] [CrossRef]
- Sanfelice, D.; Temussi, P.A. Cold denaturation as a tool to measure protein stability. Biophys. Chem. 2016, 208, 4–8. [Google Scholar] [CrossRef]
- Van Boekel, M.A. Kinetic Modeling of Reactions in Foods; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar] [CrossRef]
- Zhang, J.; Toldrá, F.; Kang, D.; Zhou, L.; Wang, J.; Zhang, W.; Hu, Y. Benefits of ultrasonic technology application in meat field and its influential mechanism: a review. Crit. Rev. Food Sci. Nutr. 2025, 65, 5726–5751. [Google Scholar] [CrossRef]
- Dias, C.L.; Ala-Nissila, T.; Wong-ekkabut, J.; Vattulainen, I.; Grant, M.; Karttunen, M. The hydrophobic effect and its role in cold denaturation. Cryobiology 2010, 60, 91–99. [Google Scholar] [CrossRef]
- Balny, C.; Masson, P. Effects of high pressure on proteins. Food Rev. Int. 1993, 9, 611–628. [Google Scholar] [CrossRef]
- Harano, Y.; Yoshidome, T.; Kinoshita, M. Molecular mechanism of pressure denaturation of proteins. J. Chem. Phys. 2008, 129, 145103. [Google Scholar] [CrossRef]
- Brash, J.L.; Horbett, T.A. Proteins at interfaces: An overview. In Proteins at Interfaces II: Fundamentals and Applications; American Chemical Society: Washington, DC, 1995; Volume 602, pp. 1–23. [Google Scholar]
- Zhai, J. l.; Day, L.; Aguilar, M.-I.; Wooster, T.J. Protein folding at emulsion oil/water interfaces. Curr. Opin. Colloid Interface Sci. 2013, 18, 257–271. [Google Scholar] [CrossRef]
- Arsiccio, A.; Pisano, R. The Ice-Water Interface and Protein Stability: A Review. J. Pharm. Sci. 2020, 109, 2116–2130. [Google Scholar] [CrossRef] [PubMed]
- Walstra, P.; De Roos, A.L. Proteins at air-water and oil-water interfaces: Static and dynamic aspects. Food Rev. Int. 1993, 9, 503–525. [Google Scholar] [CrossRef]
- Gorelov, A.V.; Morozov, V.N. Mechanical denaturation of globular protein in the solid state. Biophys. Chem. 1987, 28, 199–205. [Google Scholar] [CrossRef]
- Maa, Y.-F.; Hsu, C.C. Effect of high shear on proteins. Biotechnol. Bioeng. 1996, 51, 458–465. [Google Scholar] [CrossRef]
- Jaspe, J.; Hagen, S.J. Do Protein Molecules Unfold in a Simple Shear Flow? Biophys. J. 2006, 91, 3415–3424. [Google Scholar] [CrossRef]
- Bennett, M.J.; Schlunegger, M.P.; Eisenberg, D. 3D domain swapping: A mechanism for oligomer assembly. Protein Sci. 1995, 4, 2455–2468. [Google Scholar] [CrossRef]
- Bennett, M.J.; Choe, S.; Eisenberg, D. Domain swapping: entangling alliances between proteins. Proc Natl Acad Sci U S A 1994, 91, 3127–3131. [Google Scholar] [CrossRef]
- Zahler, C.T.; Shaw, B.F. What Are We Missing by Not Measuring the Net Charge of Proteins? Chemistry – A European Journal 2019, 25, 7581–7590. [Google Scholar] [CrossRef] [PubMed]
- Gitlin, I.; Carbeck, J.D.; Whitesides, G.M. Why Are Proteins Charged? Networks of Charge–Charge Interactions in Proteins Measured by Charge Ladders and Capillary Electrophoresis. Angew. Chem. Int. Ed. 2006, 45, 3022–3060. [Google Scholar] [CrossRef]
- Yu, Q.; Shi, T.; Xiong, Z.; Yuan, L.; Hong, H.; Gao, R.; Bao, Y. Oxidation affects dye binding of myofibrillar proteins via alteration in net charges mediated by a reduction in isoelectric point. Food Res. Int. 2023, 163, 112204. [Google Scholar] [CrossRef] [PubMed]
- De Graff, A.M.; Hazoglou, M.J.; Dill, K.A. Highly Charged Proteins: The Achilles' Heel of Aging Proteomes. Structure 2016, 24, 329–336. [Google Scholar] [CrossRef]
- Bao, Y.; Boeren, S.; Ertbjerg, P. Myofibrillar protein oxidation affects filament charges, aggregation and water-holding. Meat Sci. 2018, 135, 102–108. [Google Scholar] [CrossRef]
- Dignon, G.L.; Best, R.B.; Mittal, J. Biomolecular Phase Separation: From Molecular Driving Forces to Macroscopic Properties. Annu. Rev. Phys. Chem. 2020, 71, 53–75. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Archives of Biochemistry and Biophysics 1959, 82, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Riddles, P.W.; Blakeley, R.L.; Zerner, B. Ellman's reagent: 5,5′-dithiobis(2-nitrobenzoic acid)—a reexamination. Anal. Biochem. 1979, 94, 75–81. [Google Scholar] [CrossRef]
- Riddles, P.W.; Blakeley, R.L.; Zerner, B. Reassessment of Ellman's reagent. Methods Enzymol. 1983, 91, 49–60. [Google Scholar] [CrossRef]
- Riener, C.K.; Kada, G.; Gruber, H.J. Quick measurement of protein sulfhydryls with Ellman's reagent and with 4,4′-dithiodipyridine. Anal. Bioanal. Chem. 2002, 373, 266–276. [Google Scholar] [CrossRef]
- Møller, R.E.; Stapelfeldt, H.; Skibsted, L.H. Thiol Reactivity in Pressure-Unfolded beta-Lactoglobulin. Antioxidative Properties and Thermal Refolding. J. Agric. Food Chem. 1998, 46, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Hansen, R.E.; Winther, J.R. An introduction to methods for analyzing thiols and disulfides: Reactions, reagents, and practical considerations. Anal. Biochem. 2009, 394, 147–158. [Google Scholar] [CrossRef]
- Estévez, M. Protein carbonyls in meat systems: A review. Meat Sci. 2011, 89, 259–279. [Google Scholar] [CrossRef]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar] [CrossRef] [PubMed]
- Decker, E.A.; Xiong, Y.L.; Calvert, J.T.; Crum, A.D.; Blanchard, S.P. Chemical, physical, and functional properties of oxidized turkey white muscle myofibrillar proteins. J. Agric. Food Chem. 1993, 41, 186–189. [Google Scholar] [CrossRef]
- Fagan, J.M.; Sleczka, B.G.; Sohar, I. Quantitation of oxidative damage to tissue proteins. Int. J. Biochem. Cell Biol. 1999, 31, 751–757. [Google Scholar] [CrossRef]
- Soglia, F.; Petracci, M.; Ertbjerg, P. Novel DNPH-based method for determination of protein carbonylation in muscle and meat. Food Chem. 2016, 197, 670–675. [Google Scholar] [CrossRef]
- Headlam, H.A.; Davies, M.J. Markers of protein oxidation: different oxidants give rise to variable yields of bound and released carbonyl products. Free Radic Biol Med. 2004, 36, 1175–1184. [Google Scholar] [CrossRef]
- Kamble, S.; Agrawal, S.; Cherumukkil, S.; Sharma, V.; Jasra, R.V.; Munshi, P. Revisiting Zeta Potential, the Key Feature of Interfacial Phenomena, with Applications and Recent Advancements. ChemistrySelect 2022, 7, e202103084. [Google Scholar] [CrossRef]
- Horn, D.; Heuck, C.C. Charge determination of proteins with polyelectrolyte titration. J. Biol. Chem. 1983, 258, 1665–1670. [Google Scholar] [CrossRef]
- Elliott, G.F.; Bartels, E.M. Donnan potential measurements in extended hexagonal polyelectrolyte gels such as muscle. Biophys. J. 1982, 38, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Strasser, L.; Füssl, F.; Morgan, T.E.; Carillo, S.; Bones, J. Exploring Charge-Detection Mass Spectrometry on Chromatographic Time Scales. Anal. Chem. 2023, 95, 15118–15124. [Google Scholar] [CrossRef] [PubMed]
- Skvortsov, V.S.; Alekseytchuk, N.N.; Khudyakov, D.V.; Romero Reyes, I.V. pIPredict: A computer tool for prediction of isoelectric points of peptides and proteins. Biochemistry (Moscow) Supplement Series B: Biomedical Chemistry 2015, 9, 296–303. [Google Scholar] [CrossRef]
- Xu, W.; Yu, Q.; Gou, H.; Xu, B.; Hong, H.; Gao, R. Insight into the mechanism of the decrease in mechanical strength and water-holding capacity of gels made from oxidized gelatin. Int. J. Biol. Macromol. 2024, 258, 128842. [Google Scholar] [CrossRef]
- Reshetnyak, Y.K.; Burstein, E.A. Decomposition of protein tryptophan fluorescence spectra into log-normal components. II. The statistical proof of discreteness of tryptophan classes in proteins. Biophys. J. 2001, 81, 1710–1734. [Google Scholar] [CrossRef] [PubMed]
- Adar, F. Interpretation of Raman Spectrum of Proteins. Spectroscopy 2022, 37, 9–13,25. [Google Scholar] [CrossRef]
- Zhu, D.; Sadat, A.; Joye, I.J.; Vega, C.; Rogers, M.A. Scientific gastronomy: On the mechanism by which garlic juice and allicin (thio-2-propene-1-sulfinic acid S-allyl ester) stabilize meringues. Food Chem. 2024, 431, 137121. [Google Scholar] [CrossRef] [PubMed]
- Winger, A.M.; Taylor, N.L.; Heazlewood, J.L.; Day, D.A.; Millar, A.H. Identification of intra- and intermolecular disulphide bonding in the plant mitochondrial proteome by diagonal gel electrophoresis. Proteomics 2007, 7, 4158–4170. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Huff-Lonergan, E.; Sebranek, J.G.; Lonergan, S.M. High-oxygen modified atmosphere packaging system induces lipid and myoglobin oxidation and protein polymerization. Meat Sci. 2010, 85, 759–767. [Google Scholar] [CrossRef]
- Moczkowska, M.; Półtorak, A.; Montowska, M.; Pospiech, E.; Wierzbicka, A. The effect of the packaging system and storage time on myofibrillar protein degradation and oxidation process in relation to beef tenderness. Meat Sci. 2017, 130, 7–15. [Google Scholar] [CrossRef]
- Warren, C.M.; Krzesinski, P.R.; Greaser, M.L. Vertical agarose gel electrophoresis and electroblotting of high-molecular-weight proteins. Electrophoresis 2003, 24, 1695–1702. [Google Scholar] [CrossRef]
- Giordani, S.; Marassi, V.; Placci, A.; Zattoni, A.; Roda, B.; Reschiglian, P. Field-Flow Fractionation in Molecular Biology and Biotechnology. Molecules 2023, 28, 6201. [Google Scholar] [CrossRef]
- Hota, S.; Shams, R.; Dash, K.K.; Pawase, P.A.; Mukarram, S.A.; Kovács, B. A comprehensive review on the potential role of irradiation technique on techno-functional properties of plant-based proteins. J. Agric. Food Res. 2025, 23, 102095. [Google Scholar] [CrossRef]
- Kuan, Y.-H.; Bhat, R.; Patras, A.; Karim, A.A. Radiation processing of food proteins – A review on the recent developments. Trends Food Sci. Technol. 2013, 30, 105–120. [Google Scholar] [CrossRef]
- Chen, W.; Ma, H.; Wang, Y.-Y. Recent advances in modified food proteins by high intensity ultrasound for enhancing functionality: Potential mechanisms, combination with other methods, equipment innovations and future directions. Ultrason. Sonochem. 2022, 85, 105993. [Google Scholar] [CrossRef]
- Su, J.; Cavaco-Paulo, A. Effect of ultrasound on protein functionality. Ultrason. Sonochem. 2021, 76, 105653. [Google Scholar] [CrossRef]
- Levy, R.; Okun, Z.; Shpigelman, A. High-Pressure Homogenization: Principles and Applications Beyond Microbial Inactivation. Food Engineering Reviews 2021, 13, 490–508. [Google Scholar] [CrossRef]
- Messens, W.; Van Camp, J.; Huyghebaert, A. The use of high pressure to modify the functionality of food proteins. Trends Food Sci. Technol. 1997, 8, 107–112. [Google Scholar] [CrossRef]
- Li, B.; Peng, L.; Cao, Y.; Liu, S.; Zhu, Y.; Dou, J.; Yang, Z.; Zhou, C. Insights into Cold Plasma Treatment on the Cereal and Legume Proteins Modification: Principle, Mechanism, and Application. Foods 2024, 13, 1522. [Google Scholar] [CrossRef] [PubMed]
- Olatunde, O.O.; Hewage, A.; Dissanayake, T.; Aluko, R.E.; Karaca, A.C.; Shang, N.; Bandara, N. Cold atmospheric plasma-induced protein modification: Novel nonthermal processing technology to improve protein quality, functionality, and allergenicity reduction. Compr. Rev. Food. Sci. Food Saf. 2023, 22, 2197–2234. [Google Scholar] [CrossRef]
- Han, Z.; Cai, M.-j.; Cheng, J.-H.; Sun, D.-W. Effects of electric fields and electromagnetic wave on food protein structure and functionality: A review. Trends Food Sci. Technol. 2018, 75, 1–9. [Google Scholar] [CrossRef]
- Malik, M.A.; Sheikh, M.A.; Mir, N.A. A review on pulsed electric field modification of proteins: Effect on the functional and structural properties. Food Biosci. 2024, 61, 104636. [Google Scholar] [CrossRef]
- Ni, X.; Chen, C.; Li, R.; Liu, Q.; Duan, C.; Wang, X.; Xu, M. Effects of ultrasonic treatment on the structure and functional characteristics of myofibrillar proteins from black soldier fly. Int. J. Biol. Macromol. 2024, 278, 135057. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Jin, W.; Ma, X.; Wen, H.; Li, Y.; Xu, G.; Xu, P.; Cheng, H. A study on the structure-functionality relationship of Solenaia oleivora protein under high-intensity ultrasonication processing. Food Chem. 2024, 460, 140598. [Google Scholar] [CrossRef]
- Wang, H.; Wang, P.; Shen, Q.; Yang, H.; Xie, H.; Huang, M.; Zhang, J.; Zhao, Q.; Luo, P.; Jin, D.; Wu, J.; Jian, S.; Chen, X. Insight into the effect of ultrasound treatment on the rheological properties of myofibrillar proteins based on the changes in their tertiary structure. Food Res. Int. 2022, 157, 111136. [Google Scholar] [CrossRef]
- Wang, X.; Ni, X.; Duan, C.; Li, R.; Jiang, X. e.; Xu, M.; Yu, R. The Effect of Ultrasound Treatment on the Structural and Functional Properties of Tenebrio molitor Myofibrillar Protein. Foods 2024, 13, 2817. [Google Scholar] [CrossRef]
- Kim, Y.J.; Lee, M.H.; Kim, S.-M.; Kim, B.-K.; Yong, H.I.; Choi, Y.-S. Improvement of structural, physicochemical, and rheological properties of porcine myofibrillar proteins by high-intensity ultrasound treatment for application as Pickering stabilizers. Ultrason. Sonochem. 2023, 92, 106263. [Google Scholar] [CrossRef]
- Li, J.; Dai, Z.; Chen, Z.; Hao, Y.; Wang, S.; Mao, X. Improved gelling and emulsifying properties of myofibrillar protein from frozen shrimp (Litopenaeus vannamei) by high-intensity ultrasound. Food Hydrocolloids 2023, 135, 108188. [Google Scholar] [CrossRef]
- Wang, H.; Yang, H.; Chen, X.; Shen, Q. Structural basis for high-intensity ultrasound treatment in the rheology of myofibrillar protein extracted from White Croaker in relation to their solubility. LWT 2022, 156, 112979. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, J.; Wang, H.; Chen, Q.; Kong, B. High-intensity ultrasound improves the physical stability of myofibrillar protein emulsion at low ionic strength by destroying and suppressing myosin molecular assembly. Ultrason. Sonochem. 2021, 74, 105554. [Google Scholar] [CrossRef]
- Wang, H.; Yang, Z.; Liao, J.; Cheng, L.; Jin, D.; Xu, J.; Wang, Q.; Cheng, K.; Zheng, J.; Yang, H.; Shen, Q. Ultrasonication improved myofibrillar protein-stabilized emulsions: Oil/water interface adsorption behavior and rheological behavior. Int. J. Biol. Macromol. 2025, 308, 142390. [Google Scholar] [CrossRef]
- Yu, C.; Li, S.; Sun, S.; Yan, H.; Zou, H. Modification of emulsifying properties of mussel myofibrillar proteins by high-intensity ultrasonication treatment and the stability of O/W emulsion. Colloids Surf. Physicochem. Eng. Aspects 2022, 641, 128511. [Google Scholar] [CrossRef]
- Dai, Y.; Lu, X.; Li, R.; Li, Y.; Dong, H.; Zhu, D.; Cao, Y.; Zhou, W.; Li, J. Effects of ultrasound treatment on the structure, function properties and in vitro digestion of Sipunculus nudus protein. Int. J. Biol. Macromol. 2024, 277, 134422. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.J.; Kim, H.D.; Ye, Y.J.; Kong, M.; Lim, W.S.; Lee, M.H. Effects of ultrasound-induced structural modifications on the emulsifying properties of Tenebrio molitor proteins. Ultrason. Sonochem. 2025, 117, 107354. [Google Scholar] [CrossRef]
- Liu, B.; Wu, Y.; Liang, Q.-y.; Zheng, H. Effects of high-intensity ultrasound on physicochemical and gel properties of myofibrillar proteins from the bay scallop (Argopecten irradians). Ultrason. Sonochem. 2024, 107, 106935. [Google Scholar] [CrossRef]
- Hong, Z.; Kong, Y.; Guo, R.; Huang, Q. Stabilizing effect of silver carp myofibrillar protein modified by high intensity ultrasound on high internal phase emulsions: Protein denaturation, interfacial adsorption and reconfiguration. Int. J. Biol. Macromol. 2024, 265, 130896. [Google Scholar] [CrossRef]
- Liu, X.; Sun, X.; Wei, Y.; Ma, Y.; Sun, P.; Li, X. Effects of ultrasonic treatment on physico-chemical properties and structure of tuna (Thunnus tonggol) myofibrillar proteins. J. Food Compos. Anal. 2022, 108, 104438. [Google Scholar] [CrossRef]
- Deng, X.; Ma, Y.; Lei, Y.; Zhu, X.; Zhang, L.; Hu, L.; Lu, S.; Guo, X.; Zhang, J. Ultrasonic structural modification of myofibrillar proteins from Coregonus peled improves emulsification properties. Ultrason. Sonochem. 2021, 76, 105659. [Google Scholar] [CrossRef]
- Deng, X.-h.; Ni, X.-x.; Han, J.-h.; Yao, W.-h.; Fang, Y.-j.; Zhu, Q.; Xu, M.-f. High-intensity ultrasound modified the functional properties of Neosalanx taihuensis myofibrillar protein and improved its emulsion stability. Ultrason. Sonochem. 2023, 97, 106458. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Li, W.; Li, G.; Zhang, W.; Chen, H.; Jiang, Y.; Li, D. Effect of high-intensity ultrasound on the physicochemical properties of Tenebrio Molitor Protein. Food Hydrocolloids 2023, 134, 108056. [Google Scholar] [CrossRef]
- He, Y.; Zhang, Z.; Hu, B.; Liu, J.; Yue, Z.; Yu, Q.; Liu, Z.; Zhu, Y. Synergistic effects of pulsed electric field and NaCl on myofibrillar proteins and flavor of marinated pork. Int. J. Biol. Macromol. 2025, 292, 139272. [Google Scholar] [CrossRef]
- Kim, Y.J.; Shin, D.-M.; Oh, E.-J.; Chun, Y.G.; Shin, J.-K.; Choi, Y.-S.; Kim, B.-K. Mechanisms underlying the changes in the structural, physicochemical, and emulsification properties of porcine myofibrillar proteins induced by prolonged pulsed electric field treatment. Food Chem. 2024, 456, 140024. [Google Scholar] [CrossRef]
- Dong, M.; Tian, H.; Xu, Y.; Han, M.; Xu, X. Effects of pulsed electric fields on the conformation and gelation properties of myofibrillar proteins isolated from pale, soft, exudative (PSE)-like chicken breast meat: A molecular dynamics study. Food Chem. 2021, 342, 128306. [Google Scholar] [CrossRef]
- Wang, Q.; Wei, R.; Hu, J.; Luan, Y.; Liu, R.; Ge, Q.; Yu, H.; Wu, M. Moderate pulsed electric field-induced structural unfolding ameliorated the gelling properties of porcine muscle myofibrillar protein. Innov. Food Sci. Emerg. Technol. 2022, 81, 103145. [Google Scholar] [CrossRef]
- Jiang, J.; Yang, K.; Gong, H.; Ma, J.; Hu, X.; Zhou, Y.; Zhang, Y.; Sun, W. The conformational modification of myofibrillar protein by magnetic field improves its emulsification properties. Int. J. Biol. Macromol. 2024, 277, 134114. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wang, L.; Guo, J.; Wu, D.; Wang, X.; Wu, M.; Feng, X.; Ma, J.; Zhang, Y.; Sun, W. Structural changes induced by direct current magnetic field improve water holding capacity of pork myofibrillar protein gels. Food Chem. 2021, 345, 128849. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Feng, X.; Jiang, J.; Jiang, Q.; Ma, J.; Sun, W. Magnetic field-mediated oxidative modification of myoglobin: One effective method for improving the gel properties of myofibrillar protein. Food Chem. 2025, 472, 142899. [Google Scholar] [CrossRef]
- Li, Q.; Xia, L.; Lu, J.; Song, Y.; Yang, M.; Zhou, B.; Lin, L.; Miao, W.; Zheng, B.; Zheng, Z. Synergistic effects of alternating magnetic field and sodium tripolyphosphate on functional properties of myofibrillar proteins in low-salt systems. Int. J. Biol. Macromol. 2025, 316, 144660. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, Y.; Yang, L.; Zhao, Y.; Zhu, M.; Wang, H.; Kang, Z.; Ma, H. Low-frequency alternating magnetic field and CaCl2 influence the physicochemical, conformational and gel characteristics of low-salt myofibrillar protein. Food Chem. X 2024, 22, 101341. [Google Scholar] [CrossRef]
- Lee, M.H.; In Yong, H.; Kim, Y.J.; Choi, Y.-S. High-pressure induced structural modification of porcine myofibrillar protein and its relation to rheological and emulsifying properties. Meat Sci. 2023, 196, 109032. [Google Scholar] [CrossRef]
- Zhang, K.; Li, N.; Li, J.; Wang, Y.; Liu, C.; Liu, Y.; Liu, X.; Zhou, D.; Li, D. Improving the gelation and digestive properties of myofibrillar protein in Litopenaeus vannamei by ultra-high pressure. Food Biosci. 2023, 56, 103402. [Google Scholar] [CrossRef]
- Xu, M.; Ni, X.; Liu, Q.; Chen, C.; Deng, X.; Wang, X.; Yu, R. Ultra-high pressure improved gelation and digestive properties of Tai Lake whitebait myofibrillar protein. Food Chem. X 2024, 21, 101061. [Google Scholar] [CrossRef]
- Liu, Q.; Lin, Z.; Chen, X.; Chen, J.; Wu, J.; Chen, H.; Zeng, X. Characterization of structures and gel properties of ultra-high-pressure treated-myofibrillar protein extracted from mud carp (Cirrhinus molitorella) and quality characteristics of heat-induced sausage products. LWT 2022, 165, 113691. [Google Scholar] [CrossRef]
- Han, K.; Feng, X.; Yang, Y.; Tang, X.; Gao, C. Changes in the physicochemical, structural and emulsifying properties of chicken myofibrillar protein via microfluidization. Innov. Food Sci. Emerg. Technol. 2023, 83, 103236. [Google Scholar] [CrossRef]
- Li, J.; Guo, S.; Zhao, H.; Wang, Z.; Liu, S. Effect of High-Pressure Microjet on the Digestion Characteristics of Penaeus vannamei Myofibrillar Protein. J. Food Sci. 2025, 90, e70328. [Google Scholar] [CrossRef]
- Chen, M.; Wang, L.; Xie, B.; Ma, A.; Hu, K.; Zheng, C.; Xiong, G.; Shi, L.; Ding, A.; Li, X.; Qiao, Y.; Sun, Z.; Wu, W. Effects of High-Pressure Treatments (Ultra-High Hydrostatic Pressure and High-Pressure Homogenization) on Bighead Carp (Aristichthys nobilis) Myofibrillar Protein Native State and Its Hydrolysate. Food Bioprocess Technol. 2022, 15, 2252–2266. [Google Scholar] [CrossRef]
- Li, Q.; Liu, H.; Jiang, S.; Zhang, M.; Shan, K.; Ke, W.; Zhao, D.; Nian, Y.; Li, C. The effects of high pressure treatment on the structural and digestive properties of myoglobin. Food Res. Int. 2022, 156, 111193. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Guo, Z.; Wu, D.; Fei, X.; Ei-Seedi, H.R.; Wang, C. High-pressure homogenization influences the functional properties of protein from oyster (Crassostrea gigas). LWT 2021, 151, 112107. [Google Scholar] [CrossRef]
- Yang, J.; Zhou, S.; Chen, Y.; Jin, J.; Song, J. Effects of high hydrostatic pressure processing on the physicochemical properties, functional characteristics, and antioxidant activity of silkworm pupae protein. Food Chem. 2025, 475, 143320. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Liu, X.; Sang, Y.; Tian, G.; Wang, Z.; Hou, Y. Characterization and emulsifying properties of mantle proteins from scallops (Patinopecten yessoensis) treated by high hydrostatic pressure treatment. LWT 2022, 167, 113865. [Google Scholar] [CrossRef]
- Bai, R.; Han, J.; Ye, X.; Yu, J.; Jiang, S.; Li, Z.; Zhang, L.; Yang, C.; Chen, Y.; Wang, S.; Ding, W. Improvement on gel properties of chicken myofibrillar protein with electron beam irradiation: Based on protein structure, gel quality, water state. Int. J. Biol. Macromol. 2024, 280, 135806. [Google Scholar] [CrossRef]
- Shi, G.; Zhou, M.; Wang, L.; Xiao, Z.; Shi, L.; Jiao, C.; Wu, W.; Li, X.; Wang, J.; Qiao, Y.; Liao, L.; Ding, A.; Xiong, G. The effect of gamma and electron beam irradiation on the structural and physicochemical properties of myofibrillar protein and myosin from grass carp. J. Food Biochem. 2021, 45, e13828. [Google Scholar] [CrossRef]
- Hoon Lee, J.; Kim, Y.-J.; Choi, Y.-J.; Kim, T.-K.; Yoon Cha, J.; Kyung Park, M.; Jung, S.; Choi, Y.-S. Effect of gamma-ray and electron-beam irradiation on the structural changes and functional properties of edible insect proteins from Protaetia brevitarsis larvae. Food Chem. 2024, 434, 137463. [Google Scholar] [CrossRef]
- Qin, J.; Liu, L.; Miao, C.; Lan, B.; Liao, T.; Tian, X.; Wu, Z. Impact of Co-60 γ-ray irradiation on the cross-linking and stability of fish collagen: Structural changes and digestibility. Food Hydrocolloids 2024, 157, 110445. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Lee, J.-H.; Kim, T.-K.; Cha, J.Y.; Jung, S.; Choi, Y.-S. Influence of radiation types and doses on the structure and functionality of myofibrillar protein. LWT 2024, 205, 116503. [Google Scholar] [CrossRef]
- Li, K.; Zhou, Y.; Zhu, C.; Du, M.; Chen, B.; Zhao, D.; Bai, Y. Effects of plasma-activated water on structural and functional properties of PSE-like chicken protein isolate. Current Research in Food Science 2025, 10, 101003. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Rao, W.; Sun, Y.; Zhou, C.; Xia, Q.; He, J.; Pan, D.; Du, L. Structural and gel property changes in chicken myofibrillar protein induced by argon cold plasma-activated water: With a molecular docking perspective. Food Res. Int. 2024, 197, 115271. [Google Scholar] [CrossRef]
- Luo, J.; Xu, W.; Liu, Q.; Zou, Y.; Wang, D.; Zhang, J. Dielectric barrier discharge cold plasma treatment of pork loin: Effects on muscle physicochemical properties and emulsifying properties of pork myofibrillar protein. LWT 2022, 162, 113484. [Google Scholar] [CrossRef]
- Jiang, S.; Ye, X.; Xi, J.; Han, H.; Bai, R.; Yang, C.; Wang, S.; Li, L.; Wang, G.; Nie, H.; Ding, W. Effects of cold plasma and sodium bicarbonate on the rheological, foaming, digestibility, and gelation properties of duck myofibrillar protein. Int. J. Biol. Macromol. 2025, 315, 144649. [Google Scholar] [CrossRef] [PubMed]
- Tan, G.; Ning, Y.; Sun, C.; Bu, Y.; Zhang, X.; Zhu, W.; Li, J.; Li, X. Effects of plasma-activated slightly acidic electrolyzed water on salmon myofibrillar protein: Insights from structure and molecular docking. Food Chem. X 2024, 22, 101389. [Google Scholar] [CrossRef]
- Rao, W.; Ju, S.; Sun, Y.; Xia, Q.; Zhou, C.; He, J.; Wang, W.; Pan, D.; Du, L. Unlocking the molecular modifications of plasma-activated water-induced oxidation through redox proteomics: In the case of duck myofibrillar protein (Anas platyrhynchos). Food Chem. 2024, 458, 140173. [Google Scholar] [CrossRef]
- Cheng, J.-H.; Li, J.; Sun, D.-W. Effects of dielectric barrier discharge cold plasma on structure, surface hydrophobicity and allergenic properties of shrimp tropomyosin. Food Chem. 2023, 409, 135316. [Google Scholar] [CrossRef]
- Olatunde, O.O.; Singh, A.; Shiekh, K.A.; Nuthong, P.; Benjakul, S. Effect of High Voltage Cold Plasma on Oxidation, Physiochemical, and Gelling Properties of Myofibrillar Protein Isolate from Asian Sea Bass (Lates calcarifer). Foods 2021, 10, 326. [Google Scholar] [CrossRef]
- Jiang, W.; Wu, R.A.; Hu, X.; Zhang, R.; Ding, T.; Zhou, J. Changes in physicochemical and conformational properties of myofibrillar proteins isolated from mandarin fish (Siniperca chuatsi) treated by atmospheric pressure plasma jet. CyTA - Journal of Food 2023, 21, 625–633. [Google Scholar] [CrossRef]
- Hatab, S.; Koddy, J.K.; Miao, W.; Tang, L.; Xu, H.; Deng, S.; Zheng, B. Atmospheric cold plasma: a new approach to modify protein and lipid properties of myofibrillar protein isolate from hairtail (Trichiurus lepturus) fish. J. Sci. Food Agric. 2021, 102, 2041–2049. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Peng, S.; Hu, J.; Chen, J.; Zhou, X.; Yang, H. Excellent quality acquisition of myofibrillar protein in red shrimp (Solenocera crassicornis) based on regulating the oxidation degree of atmospheric cold plasma treatment. J. Sci. Food Agric. 2024, 105, 2712–2721. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Yang, C.; Bai, R.; Li, Z.; Zhang, L.; Chen, Y.; Ye, X.; Wang, S.; Jiang, H.; Ding, W. Modifying duck myofibrillar proteins using sodium bicarbonate under cold plasma treatment: Impact on the conformation, emulsification, and rheological properties. Food Hydrocolloids 2024, 150, 109682. [Google Scholar] [CrossRef]
- Guo, L.; Hong, C.; Wang, W.; Zhang, X.; Chen, J.; Chen, Z.; Ashokkumar, M.; Ma, H. Evaluation of low-temperature ultrasonic marination of pork meat at various frequencies on physicochemical properties, myoglobin levels, and volatile compounds. Meat Sci. 2024, 217, 109606. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Bao, Y.; Gou, H.; Xu, B.; Hong, H.; Gao, R. Mitigation of mechanical damage and protein deterioration in giant river prawn (Macrobrachium rosenbergii) by multi-frequency ultrasound-assisted immersion freezing. Food Chem. 2024, 458, 140324. [Google Scholar] [CrossRef]
- Santos, S.P.; Robalo, S.S.; Voss, M.; Casarin, B.C.; Dos Santos, B.A.; de Oliveira Mello, R.; Barin, J.S.; de Menezes, C.R.; Campagnol, P.C.B.; Cichoski, A.J. Ultrasound Thawing Optimization as a Novel Strategy to Improve Quality of Slowly Frozen Chicken Breast. Foods 2025, 14, 3446. [Google Scholar] [CrossRef]
- Gul, O.; Sahin, M.S.; Saricaoglu, F.T.; Atalar, I. Effect of sesame protein isolate modified by high-pressure homogenization, high-intensity ultrasound, and high-pressure processing on the colloidal stability of sesame paste: Determination of physicochemical, rheological, microstructural properties and storage stability. Innov. Food Sci. Emerg. Technol. 2024, 96, 103786. [Google Scholar] [CrossRef]
- Sun, W.; Xu, X.; Zhang, H.; Sun, W.; Xu, C. The Mechanism Analysis of NaCl Solution Ice Formation Suppressed by Electric Field. 2006 IEEE 8th International Conference on Properties & applications of Dielectric Materials, Bali, Indonesia, 2006; pp. 770–773. [Google Scholar]
- Bolumar, T.; Orlien, V.; Sikes, A.; Aganovic, K.; Bak, K.H.; Guyon, C.; Stübler, A.-S.; de Lamballerie, M.; Hertel, C.; Brüggemann, D.A. High-pressure processing of meat: Molecular impacts and industrial applications. Compr. Rev. Food. Sci. Food Saf. 2021, 20, 332–368. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, Y.; Zhou, P.; Zhang, X.; Wang, J. Effects of high pressure modification on conformation and gelation properties of myofibrillar protein. Food Chem. 2017, 217, 678–686. [Google Scholar] [CrossRef]
- Xue, S.; Yang, H.; Wang, H.; Tendu, A.A.; Bai, Y.; Xu, X.; Ma, H.; Zhou, G. High-pressure effects on the molecular aggregation and physicochemical properties of myosin in relation to heat gelation. Food Res. Int. 2017, 99, 413–418. [Google Scholar] [CrossRef]
- Xue, Z.; Wang, M.; Chen, X.; Xu, X.; Zhou, G. Conformational and rheological changes of high-pressure processing treated rabbit myosin subfragments during heating. LWT 2020, 122, 108994. [Google Scholar] [CrossRef]
- Denzer, M.; Furbeck, R.; Sullivan, G.; Danao, M.-G.; Mafi, G.G.; Ramanathan, R. Sarcoplasmic model to study the effects of high-pressure processing on beef color. Meat Sci. 2023, 199, 109127. [Google Scholar] [CrossRef]
- Kim, Y.J.; Cha, J.Y.; Kim, T.K.; Lee, J.H.; Jung, S.; Choi, Y.S. The Effect of Irradiation on Meat Products. Food Sci Anim Resour. 2024, 44, 779–789. [Google Scholar] [CrossRef]
- Patil, S.A.; Khandekar, S.P. LED induced non-thermal preservation of muscle foods: A systematic review. Int. J. Food Microbiol. 2025, 426, 110892. [Google Scholar] [CrossRef]
- Dąbrowska-Gralak, M.; Sadło, J.; Głuszewski, W.; Łyczko, K.; Przybytniak, G.; Lewandowska, H. The combined effect of humidity and electron beam irradiation on collagen type I - implications for collagen-based devices. Mater. Today Commun. 2022, 31, 103255. [Google Scholar] [CrossRef]
- Stanca, M.; Gaidau, C.; Zaharescu, T.; Balan, G.-A.; Matei, I.; Precupas, A.; Leonties, A.R.; Ionita, G. Physico-Chemical Changes Induced by Gamma Irradiation on Some Structural Protein Extracts. Biomolecules 2023, 13, 774. [Google Scholar] [CrossRef]
- Zhao, Y.M.; Patange, A.; Sun, D.W.; Tiwari, B. Plasma-activated water: Physicochemical properties, microbial inactivation mechanisms, factors influencing antimicrobial effectiveness, and applications in the food industry. Compr Rev Food Sci F 2020, 19, 3951–3979. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Khan, H.; Shahab, U.; Rehman, S.; Rafi, Z.; Khan, M.Y.; Ansari, A.; Siddiqui, Z.; Ashraf, J.M.; Abdullah, S.M.; Habib, S.; Uddin, M. Protein oxidation: an overview of metabolism of sulphur containing amino acid, cysteine. Front Biosci (Schol Ed) 2017, 9, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Ekezie, F.C.; Cheng, J.H.; Sun, D.W. Effects of atmospheric pressure plasma jet on the conformation and physicochemical properties of myofibrillar proteins from king prawn (Litopenaeus vannamei). Food Chem. 2019, 276, 147–156. [Google Scholar] [CrossRef]
- Hatab, S.; Koddy, J.K.; Miao, W.; Tang, L.; Xu, H.; Deng, S.; Zheng, B. Atmospheric cold plasma: a new approach to modify protein and lipid properties of myofibrillar protein isolate from hairtail (Trichiurus lepturus) fish. J. Sci. Food Agric. 2022, 102, 2041–2049. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Andrés, J.M.; Álvarez, C.; Cullen, P.J.; Tiwari, B.K. Effect of cold plasma on the techno-functional properties of animal protein food ingredients. Innov. Food Sci. Emerg. Technol. 2019, 58, 102205. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, J.; Chen, J.; Zhao, Y.; Deng, S.; Yang, H. Gelatinous quality and quantitative proteomic analyses of snakehead (Channa argus) surimi treated by atmospheric cold plasma. Food Chem. 2024, 459, 140412. [Google Scholar] [CrossRef]
- Wang, X.; Li, M.; Shi, T.; Xie, Y.; Jin, W.; Yuan, L.; Gao, R. Effect of plasma-activated water rinsing on the gelling properties of myofibrillar protein from Aristichthys nobilis surimi: insights from molecular conformational transitions. Food Chem. 2025, 496, 146726. [Google Scholar] [CrossRef]
- Li, M.; Wang, X.; Shi, T.; Xiong, Z.; Jin, W.; Bao, Y.; Monto, A.R.; Yuan, L.; Gao, R. Mechanism of plasma-activated water promoting the heat-induced aggregation of myofibrillar protein from silver carp (Aristichthys Nobilis). Innov. Food Sci. Emerg. Technol. 2024, 91, 103555. [Google Scholar] [CrossRef]
- Bauer, A.; Ni, Y.; Bauer, S.; Paulsen, P.; Modic, M.; Walsh, J.L.; Smulders, F.J.M. The effects of atmospheric pressure cold plasma treatment on microbiological, physical-chemical and sensory characteristics of vacuum packaged beef loin. Meat Sci. 2017, 128, 77–87. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Illustration of common food protein modifications during storage and processing.
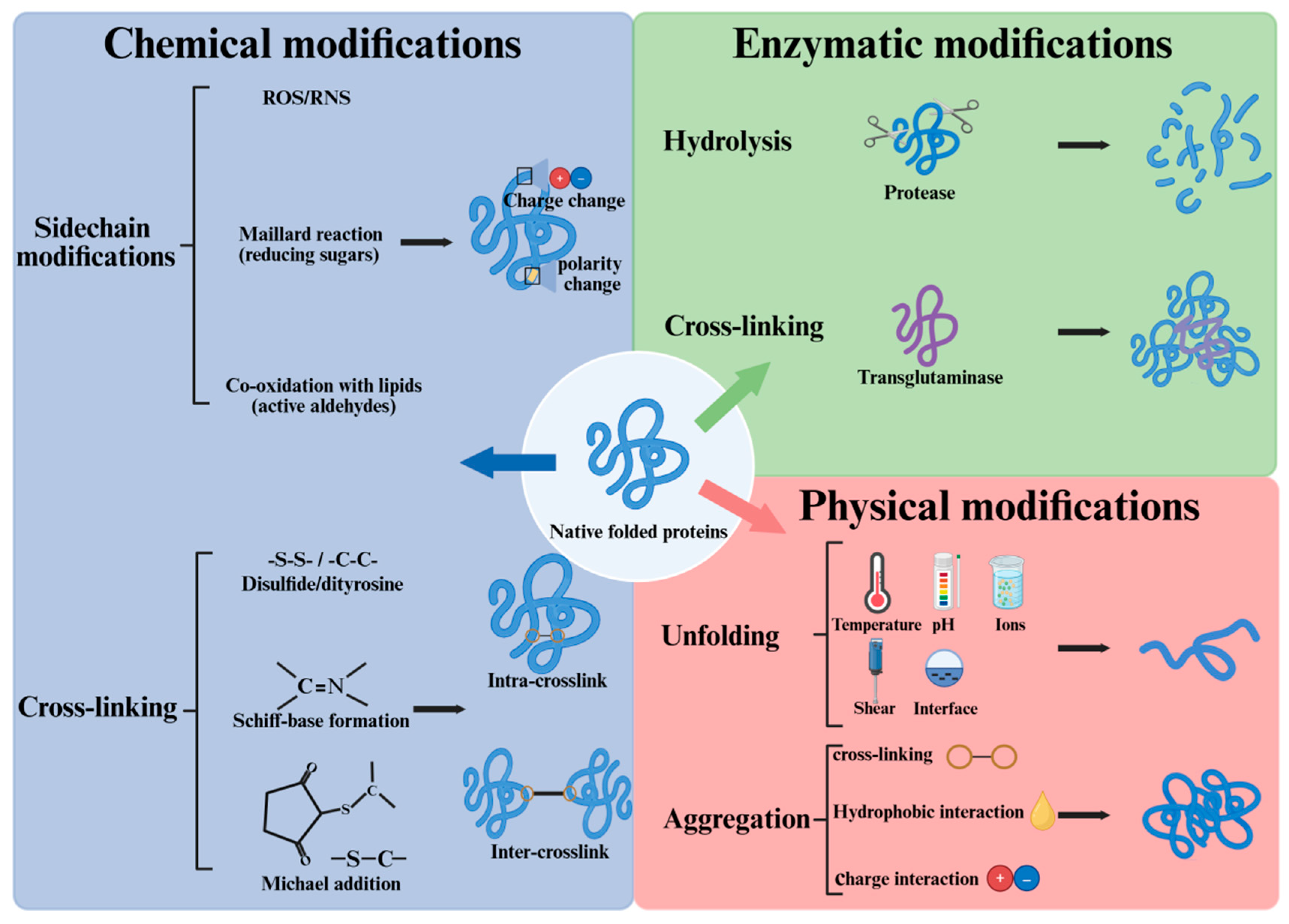
Figure 2.
Illustration of 3D - domain swapping of proteins. Proteins may partially unfold at mildly denaturing conditions, and when the denaturing conditions change, proteins may refold. The refolding process may form domain-swapped dimer or open monomers [38].
Figure 2.
Illustration of 3D - domain swapping of proteins. Proteins may partially unfold at mildly denaturing conditions, and when the denaturing conditions change, proteins may refold. The refolding process may form domain-swapped dimer or open monomers [38].
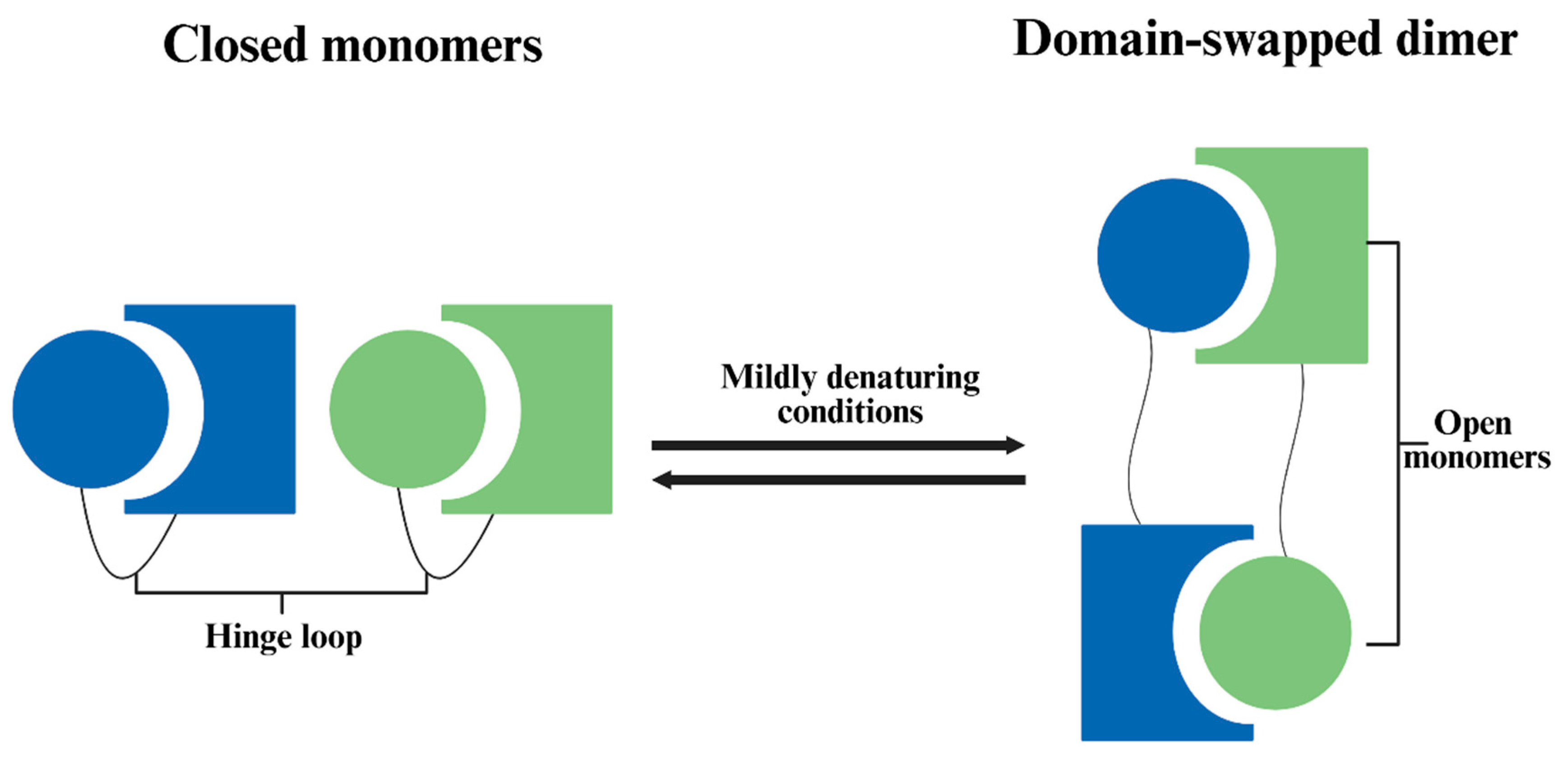
Figure 3.
Interpretation of Trp fluorescence in terms of the micro-environment of Trp residues.
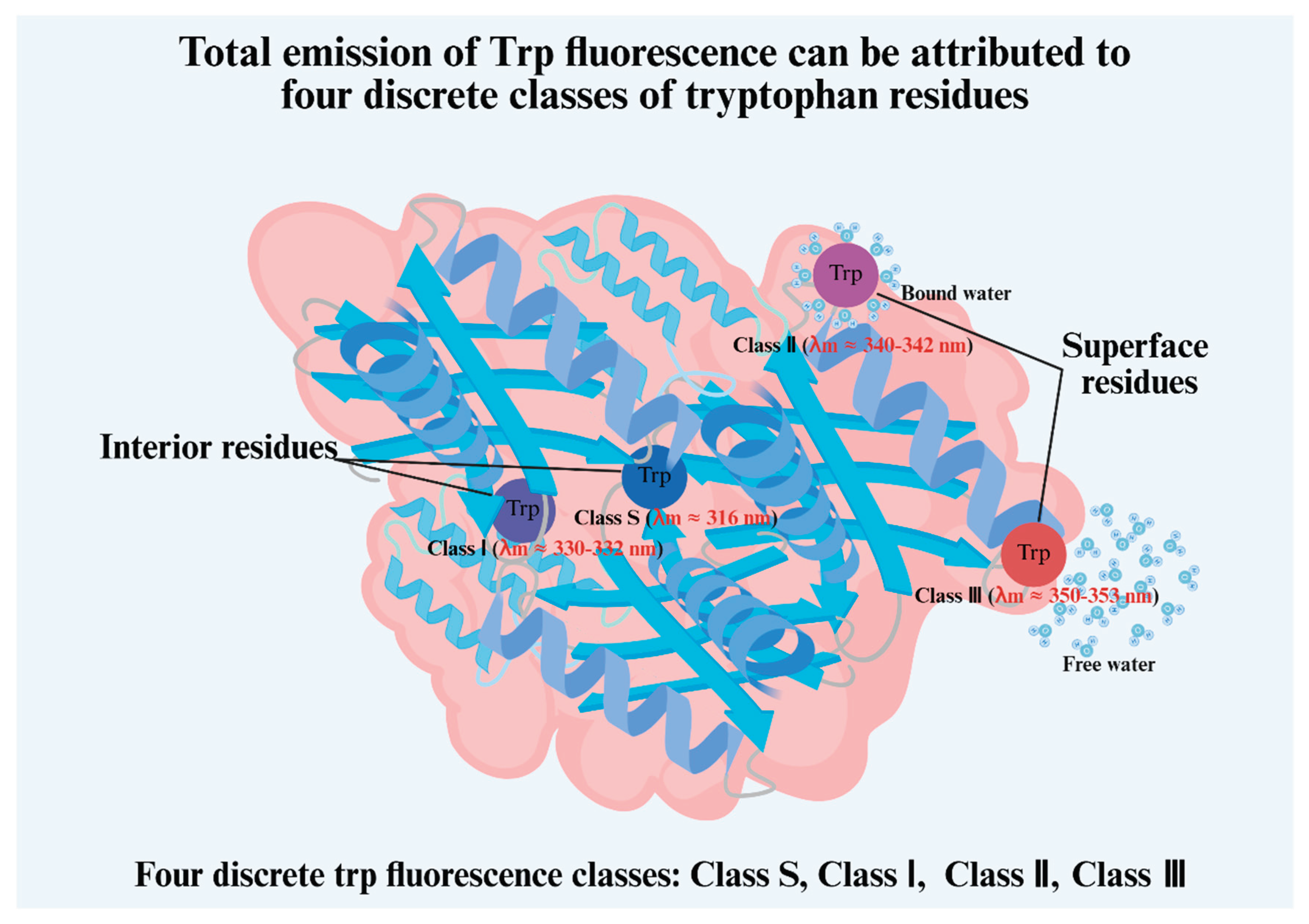

Table 1.
Effects of non-thermal processing on the properties of muscle proteins.
| Technology | Protein | Main effects | Reference |
| Ultrasound | Myofibrillar protein MP from black soldier fly |
↑: Carbonyl content, Surface hydrophobicity ↓: Total thiol group, Particle size, Turbidity, Intrinsic fluorescence intensity |
[82] |
| Protein from Solenaia oleivora | ↑: Surface hydrophobicity, Solubility, Intrinsic fluorescence intensity, Zeta-potential, β-turn, α-helix, ↓: Particle size, Total thiol group, Random coil |
[83] | |
| Myofibrillar protein MP from surimi |
↑: Total thiol group, Surface hydrophobicity | [84] | |
| Myofibrillar protein MP from Tenebrio molitor |
↑: Carbonyl content, Surface hydrophobicity ↓: Total thiol group, Turbidity, Particle size, Intrinsic fluorescence intensity |
[85] | |
| Myofibrillar protein MP from pork |
↑: Carbonyl content, Surface hydrophobicity α-helix, ↓: Turbidity, Particle size, Random coil, β-sheet |
[86] | |
| Myofibrillar protein MP from shrimp |
↑: Surface hydrophobicity, Free sulfhydryl content, Solubility, Zeta-potential ↓: α-helix, Random coil, Particle size |
[87] | |
| Myofibrillar protein MP from fish |
↑: Solubility, Zeta-potential, Random coil, β-turn, β-sheet ↓: Particle size, Intrinsic fluorescence intensity, α-helix |
[88] | |
| Myofibrillar protein MP from pork |
↓: α-helix | [89] | |
| Myofibrillar protein MP from surimi |
↑: Free sulfhydryl content, Solubility, Surface hydrophobicity ↓: Particle size |
[90] | |
| Myofibrillar protein MP from mussel |
↑: Solubility, Zeta-potential, β-turn, Surface hydrophobicity, Free sulfhydryl content ↓: Particle size, Turbidity, Intrinsic fluorescence intensity, β-sheet |
[91] | |
| Myofibrillar protein MP from Sipunculus nudus |
↑: Solubility ↓: Particle size |
[92] | |
| Myofibrillar protein MP from Tenebrio molitor |
↑: Intrinsic fluorescence intensity, Solubility, Zeta-potential, Surface hydrophobicity, α-helix, β-turn ↓: Particle size, β-sheet |
[93] | |
| Myofibrillar protein MP from bay scallop |
↑: Carbonyl content, Intrinsic fluorescence intensity, β-sheet, β-turn ↓: α-helix, Random coil |
[94] | |
| Myofibrillar protein MP from fish |
↑: Solubility, Zeta-potential, β-turn, β-sheet Surface hydrophobicity, Intrinsic fluorescence intensity ↓: Particle size, α-helix, Random coil |
[95] | |
| Myofibrillar protein MP from fish |
↑: Surface hydrophobicity ↓: Total thiol group, Particle size |
[96] | |
| Myofibrillar protein MP from fish |
↑: Carbonyl content, Surface hydrophobicity, Solubility ↓: Particle size, Total thiol group |
[97] | |
| Myofibrillar protein MP from Neosalanx taihuensis |
↑: Intrinsic fluorescence intensity, β-sheet Surface hydrophobicity, Carbonyl content ↓: Total thiol group, α-helix |
[98] | |
| Protein from Tenebrio molitor | ↑: Free sulfhydryl content, Zeta-potential Surface hydrophobicity, α-helix ↓: Particle size, Turbidity, Intrinsic fluorescence intensity, β-turn, β-sheet |
[99] | |
| Pulsed electric field | Myofibrillar protein MP from pork |
↑: Surface hydrophobicity, Carbonyl content, Random coil, β-sheet ↓: Solubility, Total thiol group, Particle size, α-helix, Random coil |
[100] |
| Myofibrillar protein MP from pork |
↑: Intrinsic fluorescence intensity, Solubility, α-helix ↓: Surface hydrophobicity, β-sheet, β-turn, Random coil |
[101] | |
| Myofibrillar protein MP from chicken |
↓: Particle size | [102] | |
| Myofibrillar protein MP from pork |
↓: Total thiol group | [103] | |
| Magnetic field | Myofibrillar protein MP from pork |
↑: Surface hydrophobicity, α-helix, ↓: Intrinsic fluorescence intensity, β-sheet |
[104] |
| Myofibrillar protein MP from pork |
↑: Turbidity | [105] | |
| Myoglobin Myoglobin from equine muscle |
↑: Intrinsic fluorescence intensity ↓: Total thiol group |
[106] | |
| Myofibrillar protein MP from fish |
↑: Solubility, Surface hydrophobicity |
[107] | |
| Myofibrillar protein MP from pork |
↑: Solubility, Surface hydrophobicity, Total thiol group ↓: Turbidity, Intrinsic fluorescence intensity, Particle size |
[108] | |
| High pressure | Myofibrillar protein MP from pork |
↑: Surface hydrophobicity ↓: Particle size, Turbidity |
[109] |
| Myofibrillar protein MP from shrimp |
↑: Carbonyl content, Surface hydrophobicity, β-sheet ↓: α-helix, Intrinsic fluorescence intensity, Total thiol group, |
[110] | |
| Myofibrillar protein MP from fish |
↑: Carbonyl content, Surface hydrophobicity β-sheet, ↓: Total thiol group, Intrinsic fluorescence intensity, Random coil, α-helix, β-turn |
[111] | |
| Myofibrillar protein MP from fish |
↑: Surface hydrophobicity, Particle size, Disulfide bond content ↓: Turbidity, TCA-soluble peptide content |
[112] | |
| Myofibrillar protein MP from chicken |
↑: Surface hydrophobicity, β-sheet, Solubility ↓: α-helix, β-turn, Turbidity |
[113] | |
| Myofibrillar protein MP from shrimp |
↑: Intrinsic fluorescence intensity, Random coil ↓: Particle size, Turbidity, α-helix, Total thiol group, Disulfide-bond content |
[114] | |
| Myofibrillar protein MP from fish |
↑: Zeta-potential, Surface hydrophobicity, β-sheet ↓: Total thiol group, Particle size, α-helix, Random coil |
[115] | |
| Myoglobin Myoglobin from equine muscle |
↓: Intrinsic fluorescence intensity | [116] | |
| Protein from oyster | ↑: Surface hydrophobicity, Solubility, Random coil ↓: Intrinsic fluorescence intensity, α-helix |
[117] | |
| Protein from silkworm pupae |
↑: Turbidity, β-sheet, β-turn ↓: Intrinsic fluorescence intensity, α-helix, Free sulfhydryl content, Surface hydrophobicity |
[118] | |
| Mantle proteins from scallops | ↑: Total thiol group ↓: Solubility, α-helix |
[119] | |
| Irradiation | Myofibrillar protein MP from chicken |
↑: Intrinsic fluorescence intensity, Surface hydrophobicity ↓: Turbidity, Particle size, Total thiol group |
[120] |
| Myofibrillar protein Myosin MP and MS from fish |
↑: Total thiol group ↓: Particle size, Solubility, Ca2+-ATPase Activity |
[121] | |
| Protein from Protaetia brevitarsis | ↑: Solubility, Surface hydrophobicity ↓: Intrinsic fluorescence intensity |
[122] | |
| Collagen Collagen from tilapia skin |
↑: Carbonyl content, Surface hydrophobicity ↓: Free sulfhydryl content, Solubility |
[123] | |
| Myofibrillar protein MP from pork |
↑: α-helix ↓: Solubility, Particle size, Intrinsic fluorescence intensity, β-sheet |
[124] | |
| Cold plasma | Pale, soft and exudative - like chicken protein isolate | ↑: Particle size, Surface hydrophobicity, Zeta-potential, Turbidity, α-helix ↓: Intrinsic fluorescence intensity, Solubility Total thiol group, β-sheet, Random coil |
[125] |
| Myofibrillar protein MP from chicken |
↑: Intrinsic fluorescence intensity, Surface hydrophobicity ↓: β-sheet |
[126] | |
| Myofibrillar protein MP from pork |
↑: Surface hydrophobicity, Carbonyl content ↓: Total thiol group, Particle size |
[127] | |
| Myofibrillar protein MP from duck |
↑: Carbonyl content, Dityrosine content ↓: Total thiol group, Ca2+-ATPase Activity |
[128] | |
| Myofibrillar protein MP from fish |
↑: Intrinsic fluorescence intensity, Solubility, Dityrosine content, Carbonyl content ↓: Particle size, Surface hydrophobicity, Total thiol group |
[129] | |
| Myofibrillar protein MP from duck |
↑: Carbonyl content ↓: Total thiol group |
[130] | |
| Tropomyosin TM from shrimp |
↑: Surface hydrophobicity, β-sheet, Random coil β-turn ↓: Intrinsic fluorescence intensity, α-helix |
[131] | |
| Myofibrillar protein MP from fish |
↑: Carbonyl content ↓: Total thiol group |
[132] | |
| Myofibrillar protein MP from fish |
↑: Turbidity, Surface hydrophobicity, β-sheet ↓: Intrinsic fluorescence intensity, Solubility, Total thiol group, β-turn, Random coil |
[133] | |
| Myofibrillar protein MP from fish |
↑: Surface hydrophobicity, Turbidity ↓: Total thiol group |
[134] | |
| Myofibrillar protein MP from shrimp |
↑: Carbonyl content, Surface hydrophobicity, Turbidity, β-sheet, Random coil ↓: Ca2+-ATPase Activity, Total thiol group, α-helix, β-turn |
[135] | |
| Myofibrillar protein MP from duck |
↑: Turbidity, Particle size, β-turn, Random coil ↓: Solubility, Free sulfhydryl content, Surface hydrophobicity, α-helix, β-sheet |
[136] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



