
Submitted:
24 January 2026
Posted:
26 January 2026
You are already at the latest version
Abstract
The microscopic identification of insect derived processed animal proteins (PAPs) is essential for feed control within the European Union, where light microscopy remains the official method for detecting prohibited animal proteins. This study synthesizes current morphological knowledge for the insect species authorized as feed materials Hermetia illucens, Musca domestica, Tenebrio molitor, Alphitobius diaperinus, Acheta domesticus, Gryllodes sigillatus, Gryllus assimilis and Bombyx mori and provides a comprehensive framework for their recognition in processed feed. Diagnostic traits were compiled from reference laboratory material, the scientific literature and the EURL AP micrograph collection, with emphasis on cuticular structures, setae, denticles, spiracles, tracheal elements, and species specific features such as spine like sensilla in H. illucens, gin trap structures in T. molitor, and distinctive antennal or cercal fragments in Orthoptera. Dipteran PAPs are characterized by unsclerotized cuticle, denticle bands, and spiracular morphology, whereas Coleopteran fragments display stronger sclerotization, larger mouthparts, and urogomphi. Orthopteran meals exhibit the greatest structural diversity due to the use of nymphs and adults, yielding leg, wing, antennal, and cercal fragments. Across taxa, tracheal and muscle fibre structures provide reliable confirmation of insect origin but lack species specificity. The study highlights persistent gaps in reference material, particularly for Musca domestica, and underscores the need for enhanced taxonomic training and the integration of automated image recognition tools and complementary molecular methods for species level confirmation. The identification key and morphological guidance presented here aim to support routine laboratory diagnostics and strengthen regulatory compliance in feed monitoring.
Keywords:
EU
; feed
; insects
; microscopical identification
Introduction
Bovine spongiform encephalopathy (BSE), a prion disease transmitted through feed, is controlled in the European Union (EU) through banning of the use of processed animal proteins (PAPs) in ruminant diets. To strengthen feed safety, the EU implemented the General Food Law (Regulation (EC) No 178/2002) and the Hygiene Package (Regulations (EC) No 852/2004 and 183/2005). Within this regulatory framework, insect producers are responsible for ensuring product safety and full compliance with EU legislation.
Following recommendations from the European Food Safety Authority (EFSA), the use of insects in animal feed was introduced by Regulation (EU) 2017/893. This regulation amended Regulation (EC) No 999/2001 and Regulation (EU) No 142/2011, marking the first official step toward the inclusion of insect-derived PAPs in aquaculture feed. The authorization initially covered seven species: the black soldier fly (Hermetia illucens), the common housefly (Musca domestica), the yellow mealworm (Tenebrio molitor), the lesser mealworm (Alphitobius diaperinus), the house cricket (Acheta domesticus), the banded cricket (Gryllodes sigillatus), and the field cricket (Gryllus assimilis). In 2021, Commission Regulation (EU) 2021/1925 expanded the list of authorized species to include the silkworm (Bombyx mori). Later that year, Commission Regulation (EU) 2021/1372 partially lifted the feed ban by permitting the use of insect PAPs, poultry PAPs, and pig PAPs in poultry and pig feed, while maintaining the prohibition on intra-species recycling.
Currently, two official analytical methods are recognized in the EU for detecting animal proteins in feed: light microscopy and real-time polymerase chain reaction (PCR) (European Commission 2009, 2013, 2020, 2022). Light microscopy, routinely applied since 2004, enables the detection of morphological markers of animal-derived PAPs, including insect material, after an extraction step, with a detection limit of approximately 0.1%. Its effectiveness is linked to its simplicity, the modest level of expertise required, and the absence of specialized equipment. When sufficient fragments are present, light microscopy can even support identification at higher taxonomic levels, such as order and, in some cases, family (Veys and Baeten, 2018).
The first work describing key diagnostic features of insect fragments under light microscopy was conducted by Ottoboni et al. (2017), focusing on H. illucens, B. mori, and T. molitor. Veys and Baeten (2018) expanded this knowledge by characterizing morphological traits of four legally authorized insect PAPs from three orders: Diptera (H. illucens), Coleoptera (T. molitor and A. diaperinus), and Orthoptera (G. assimilis). Weiner and Kwiatek (2022) further examined samples containing PAPs from H. illucens and T. molitor. More recently, Marien et al. (2024) proposed a specific real-time PCR method for the detection of B. mori and performed light microscopy observations on industrial feed samples containing B. mori, and pure insect meal samples of H. illucens and T. molitor.
The identification of fly larvae is extensively documented, mainly due to their medico-legal relevance rather than feed-science applications (Grzywacz et al., 2017; Barros et al., 2019; Walczak and Grzywacz, 2024). Grzywacz et al. (2017) developed species-level identification keys for Muscidae. Sultana et al. (2021) provided detailed descriptions, taxonomic keys, and illustrations for 17 Gryllidae species of Pakistan, bringing existing information up to date. Their work highlighted diagnostic differences between morphologically similar species and presented a taxonomic key for the species occurring in Sindh, including A. domesticus, Acheta hispanicus, Gryllus (Gryllus) bimaculatus, Gryllus (Gryllus) campestris, Gryllus septentrionalis, G. sigillatus, and Gryllodes supplicans. However, such tools are still lacking in the context of feed authentication and control. Their development would clearly support the accurate identification of PAPs from authorized species and help distinguish them from non-authorized ones, as previously emphasized by Veys and Baeten (2018).
In this work, we further investigated the morphological features of fragments from pure insect samples and, based on the available bibliography, provide a first draft of an identification key for the authorized species.
Taxonomic Overview of Authorized Insects in Feed in EU
Currently, eight insect species belonging to four different taxonomic orders are authorized for use in feed within the European Union: Diptera (flies), Coleoptera (beetles), Orthoptera (crickets), and Lepidoptera (moths) (Table 1).
Diagnostic Morphological Features of Insects Authorized for Use in Feed
The most reliable features used by laboratories to identify insect material in processed feed are chitinous structures that withstand rendering and heat treatment. These include the cuticle, setae, tracheal elements, muscle fibres, spiracles, and mouthparts. Among these, cuticle structure is often the most informative diagnostic trait. All species share the presence of a tegument, although its degree of sclerotization varies markedly; from the soft cuticle of Diptera larvae to the more rigid and highly chitinized exoskeleton characteristic of the larval, pupal, nymphal, and adult stages of Coleoptera, Orthoptera, and Lepidoptera. Setae are present in all taxa, but their abundance, morphology, and function differ by species and developmental stage. Spine-like sensilla are a distinctive morphological feature of H. illucens larvae and are not consistently reported in the other authorized insect species used in feed; their presence therefore represents a species-specific trait characteristic of H. illucens. All insects considered in this study, across all active life stages (larvae, nymphs, pupae, adults), possess a tracheal system for respiration, with external openings (spiracles) that remain identifiable after processing. All developmental stages; larvae, pupae, nymphs, and adults, breathe through spiracles connected to the tracheal system, and mandibles are present in all these stages, making them common yet still useful diagnostic elements. Tracheoles appear as small, transparent, spiral-shaped tubes, and muscle fibres are universal identifiable features shared by all species, indicating only the presence of insect material but not allowing species-level discrimination. Muscle fragments typically appear as yellow or transparent rectangular pieces, sometimes showing visible zig-zag striation, which confirms the presence of animal tissue. The origin of muscle fibres from insects can be confirmed by the observation of associated tracheal elements.
The primary morphological distinctions among species include the presence or absence of legs, wings, and visible head structures such as antennae. Fly larvae, such as H. illucens and M. domestica, are typical maggots, lacking legs and possessing an indistinct head capsule or mouth hooks. While entirely absent in fly larvae, legs are present in all other life stages of the remaining species. In contrast, larvae of T. molitor and A. diaperinus have well-defined mouthparts (mandibles) and abdominal appendages. A. diaperinus is generally darker and more strongly sclerotized than T. molitor.
Crickets possess strongly developed hind legs for jumping in both nymph and adult stages. Sultana et al. (2021) described the legs of A. domesticus as yellowish with a few brown spots and numerous hairs, and noted that the posterior tibia bears eleven spines on its basal side; the cerci are well developed and pointed. In Gryllus species, the legs are blackish with brown spots, the posterior femora are relatively short and thick, and the posterior tibia is armed with six spines on each margin. G. sigillatus is characterized by reduced wings.
The following key is based on micrographs from the internal laboratory reference collection, the EURL-AP micrograph archive, and published literature (Table S1; Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6 and Figure 7). Larvae of M. domestica were not available in our collection, and although fragment descriptions in processed feed exist for most authorized insect species, information for M. domestica larvae remains limited.
Identification Key for EU—Authorized Insect PAPs
1. Initial separation based on presence of adult/nymph structures
1A. No wings, no compound eyes, no segmented antenna fragments, no cerci → Larval/Pupal pathway → go to 2
1B. Wings, compound eyes, segmented antenna fragments, cerci, articulated leg fragments present → Adult/Nymph pathway (Orthoptera) → go to 6
2. Larval / Pupal Pathway
2A. Absence of legs or claws
→ Diptera larvae → go to 3
2B. Thoracic legs, pseudolegs, mandibles, claws, urogomphi, or gin-traps present
→ Coleoptera or Lepidoptera larvae/pupae → go to 4
3. Diptera Larvae
3A. Hermetia illucens (Black soldier fly)
- Honeycomb-like reticulated cuticle (4–6-sided cells, thick walls, broad lumen)
- Dense small setae; long yellow-brown bristles
- Spine-like sensilla present
- Linear, unidirectional denticles (spinose bands)
- Dorsal posterior spiracles with multiple radial openings
- Anal opening bordered by short, thick spine-like setae
- Colour: grey-cream → dark brown
→ Diagnostic conclusion: Honeycomb cuticle + spine-like sensilla + linear denticles = H. illucens (Figure 1)
3B. Musca domestica (Housefly)
- Smooth cuticle
- Fine, sparse setae
- No spine-like sensilla, scale-like spine
- Posterior spiracles kidney-shaped with complete peritreme enclosing three M-shaped sinuous slits
- Pale, soft fragments
→ Diagnostic conclusion: Smooth cuticle + M-shaped spiracles = M. domestica
4. Coleoptera and Lepidoptera Larvae / Pupae
4A. Tenebrio molitor (Yellow mealworm)
- Soft to amber, lightly sclerotized cuticle
- Greyish-yellow → greyish-amber
- Irregular light spots and scattered dark dots (some with very short bristles)
- Gin-trap fragments present (pupal origin)
- Mandibles, thoracic legs recognizable by their claws, urogomphi, anal spine
→ Diagnostic conclusion: Gin traps + soft amber cuticle with spotted pattern = T. molitor (Figure 3)
4B. Alphitobius diaperinus (Lesser mealworm)
- Moderately sclerotized, darker cuticle
- Uniform amber–brown pigmentation
- Sparse short setae
- Mandibles, thoracic legs recognizable by their claws, urogomphi, anal spine
- Gin-traps absent
→ Diagnostic conclusion: Darker, more sclerotized cuticle + absence of gin traps = A. diaperinus (Figure 4)
4C. Bombyx mori (Silkworm) – pupal fragments
- Yellow to dark brown cuticle
- Cuticular fragments display a pattern that partially resembles that of H. illucens
- Smooth surface with few setae
- Pupal leg fragments (leg sheaths)
- Spiracles present
→ Diagnostic conclusion: Yellow→brown smooth cuticle + pupal leg fragments = B. mori (Figure 2)
6. Adult / Nymph Pathway (Orthoptera)
(Identified by articulated legs, claws, antennae, cerci, wing fragments, compound eyes)
Shared Orthopteran structures
- Pigmented, sclerotized cuticular fragments with setae attached or isolated
- Tibial spines, femoral and tarsal fragments
- Sclerotized mandibles
- Wing fragments (membranous or sclerotized)
- Compound eye fragments
- Cercal fragments
- Segmented antennal flagellomeres
- Claws (often reddish-tipped)
- Muscle tissue, tracheal system fragments, muscle with tracheae
6A. Acheta domesticus (House cricket)
- Pale, lightly sclerotized cuticle
- Segmented antennal flagellomeres
- Cerci, Claws
- Leg fragments
Legs yellowish with a few brown spots; posterior tibia armed with eleven spines on the basal side.
→ Diagnostic conclusion: Pale cuticle + typical cricket appendages = A. domesticus (Figure 5)
6B. Gryllodes sigillatus (Banded cricket)
- Light cuticle (slightly darker than A. domesticus)
- Segmented antennal flagellomeres
- wings reduced → Less Wing fragments
- Cerci, Claws
- Leg fragments,
→ Diagnostic conclusion: Light cuticle + cricket appendages = G. sigillatus (Figure 6)
6C. Gryllus assimilis (Field cricket)
- Darker, more strongly sclerotized cuticle
- Wing fragments
- Robust segmented antennal flagellomeres
- Cerci, Claws
- Leg fragments, mandibles, muscle + tracheae
- In Gryllus species, the legs are blackish with brown spots, the posterior femora are relatively short and thick, and the posterior tibia is armed with six spines on each margin.
- Honeycomb-like cuticle may occur
→ Diagnostic conclusion: Dark cuticle + differentiated appendages + honeycombed fragments = G. assimilis. (Figure 7)
Diptera meal consists primarily of larval instars and pre-pupal stages (Veys and Baeten, 2018). The larvae are apodal (Barros et al., 2018). As a consequence of the holometabolous development of Diptera, these stages are only weakly differentiated, and therefore relatively few particles can be confidently identified as being of insect origin (Veys and Baeten, 2018). Nevertheless, the predominance of unsclerotized cuticle fragments, together with the presence of denticles arranged in spinose bands and long setae, provides reliable diagnostic criteria for recognizing Diptera larvae (Veys and Baeten, 2018; Szpila, 2009).
H. illucens material ranges in colour from grey-cream to dark brown (Barros et al., 2019; Weiner and Kwiatek, 2022; Marien et al., 2024). The cuticle of H. illucens is densely covered with setae (Oliveira et al., 2016; Ottoboni et al., 2017; Veys and Baeten, 2018), which under light microscopy may appear either attached to cuticular fragments or as isolated particles (Veys and Baeten, 2018).
Spine-like sensilla are distinctively present in H. illucens larvae and are not consistently reported in other authorized insect species used in feed, making them a species-specific trait of H. illucens. Long, yellow to yellow-brown bristles are also frequently observed (Ottoboni et al., 2017; Veys and Baeten, 2018; Weiner and Kwiatek, 2022). Small, linearly organized groups of denticles oriented in the same direction, corresponding to the spinose bands of the larval segments, commonly occur in H. illucens PAPs (Veys and Baeten, 2018).Under a light microscope, H. illucens can be recognized by its distinctive cuticular structures, which display irregular, cell-like patterns composed of four-, five-, or six-sided units with thick walls surrounding a lighter central lumen, giving the cuticle a characteristic honeycomb-like appearance; in some fragments, a central darker dot may also be visible (Ottoboni et al., 2017; Veys and Baeten, 2018; Weiner and Kwiatek, 2022; Rebora et al., 2023). Weiner and Kwiatek (2022) reported that some dark-brown cuticular fragments of H. illucens can resemble rape husks, complicating morphological classification. However, rape husks typically exhibit a darker central region, whereas the corresponding structures in H. illucens fragments show a less pronounced central darkening (Weiner and Kwiatek, 2022). Ottoboni et al. (2017) also reported the presence of pyramidal cuticular structures, although their precise characterization in H. illucens remained challenging, and further investigation was recommended to better define these elements.
Figure 1.
H. illucens under light microscopy: 1–3 Honeycomb-like cuticle; 4–11 Cuticular fragments with bristles; 12,15,17 Cuticular fragments with bristles and spiracle; 13–15 Spine-like sensilla; 19 Setae; 20 Fragments of the tracheal system; 21-24: Spine-like sensilla, bristles and setae.
Figure 1.
H. illucens under light microscopy: 1–3 Honeycomb-like cuticle; 4–11 Cuticular fragments with bristles; 12,15,17 Cuticular fragments with bristles and spiracle; 13–15 Spine-like sensilla; 19 Setae; 20 Fragments of the tracheal system; 21-24: Spine-like sensilla, bristles and setae.

The antennae of H. illucens are inserted anterolaterally on a moderately prominent ring-shaped socket with a distinct articulatory membrane (Fabian et al., 2025). Dorsal posterior spiracles, composed of multiple openings arranged radially on the ecdysial surface (Oliveira et al., 2016), may also be detected, as well as the anal opening, whose edges bear short, thick, spine-like setae (Oliveira et al., 2016; Barros et al., 2019; EURL-AP micrograph collection). Posterior spiracles are considered the most reliable diagnostic structures for identifying fly larvae. Their morphology is highly distinctive, and the number and arrangement of spiracular openings can vary between species and even across developmental stages (Duncan et al., 2010; Raś et al., 2018).
M. domestica larvae, characterized by their typical maggot-like form, lack the heavy sclerotization observed in H. illucens. For M. domestica, the morphology of the posterior spiracle is a key diagnostic feature (Apasrawirote et al., 2022). The spiracles are kidney-shaped, and a complete peritreme enclosing three distinctly sinuous, M-shaped slits is characteristic of this species (Grzywacz et al., 2017; Apasrawirote et al., 2022). Under high magnification, the cuticle may display minute bands of cuticular spines or spinules. Cortinhas et al. (2020) described the cephalic collar spines as composed of numerous scale-like structures, distinguishing them from those of other muscid species. They also noted spines on the anterior portion of the first thoracic segment. After segmentation between the cephalic region and the first thoracic segment, the ventrally concentrated spines become more slender (Cortinhas et al., 2020). These morphological traits may suggest the presence of similar spines in M. domestica. Despite these well-defined traits in intact larvae, the current literature provides very limited morphological descriptions of M. domestica fragments as they occur in processed feed. This gap is also reflected in the EURL-AP micrograph collection, where such fragmentary remains are not represented. As a result, the identification of house fly fragments remains challenging and is currently insufficiently supported by reference material.
B. mori, commercially processed and used at the pupal stage, exhibit cuticular fragments with a pattern that partially resembles that of H. illucens, with coloration ranging from yellow to dark brown (Figure 2; Harris, 1979; Ottoboni et al., 2017; Marien et al., 2024). Ottoboni et al. (2017) reported the absence of setae among PAP fragments derived from B. mori; however, Marien et al. (2024) documented their presence in this species. Similar structures are also observed in the laboratory reference sample (Figure 2). Kumar et al. (1999) described cremastral setae on the terminal abdominal segment of B. mori, which might explain those observations.
Figure 2.
B. mori meal: 1–5: Cuticle fragments (specimens 2 and 3 showing oval spiracles); 6,8: Seta; 7: Spiracle.
Figure 2.
B. mori meal: 1–5: Cuticle fragments (specimens 2 and 3 showing oval spiracles); 6,8: Seta; 7: Spiracle.
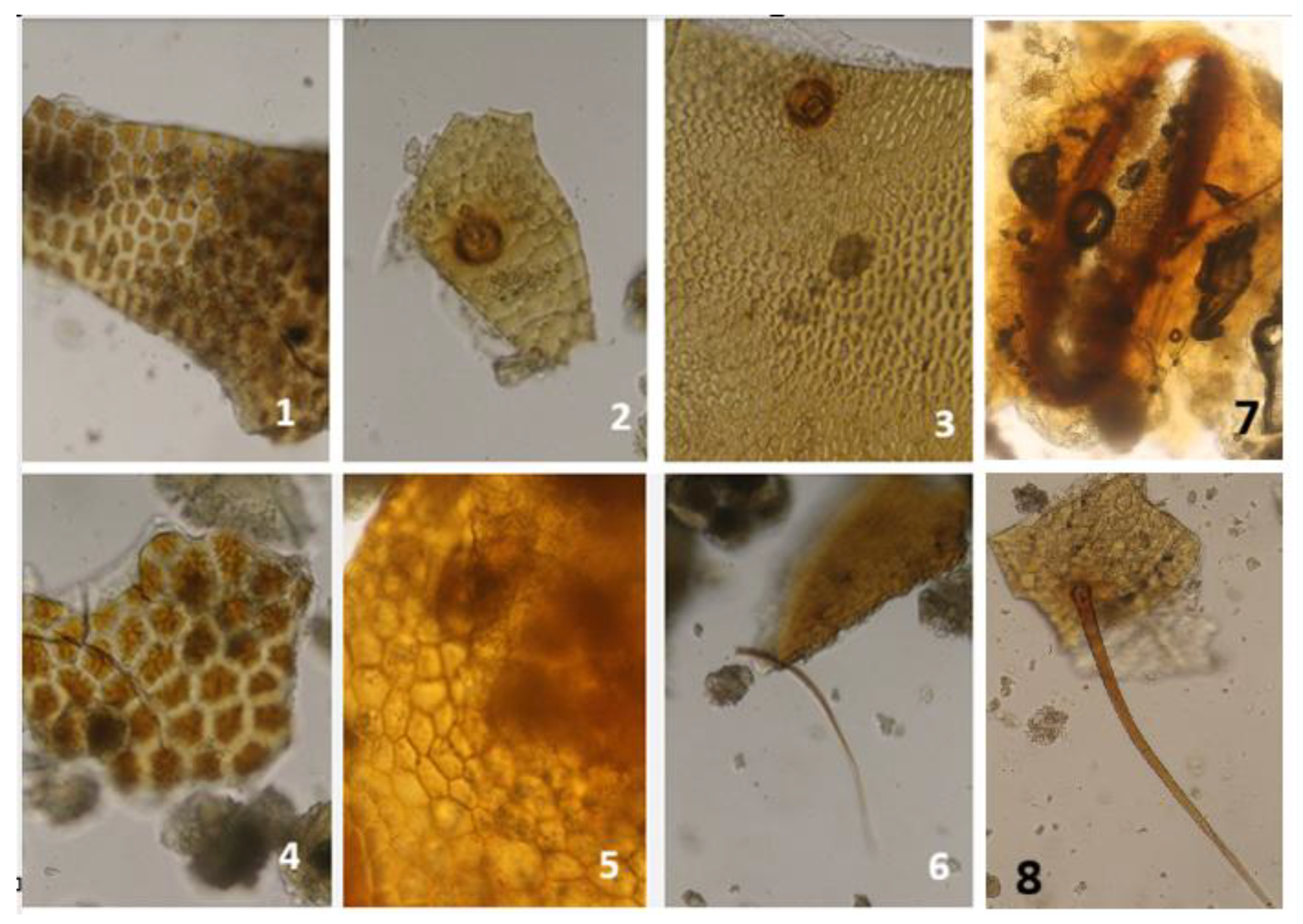
Fragments of the exoskeleton of T. molitor are described as bright greyish-yellow to deep greyish-amber-brown in colour, often displaying irregular light spots and sporadic black dots (Ottoboni et al., 2017; Weiner and Kwiatek, 2022). Some of these dark dots bear very short, dark bristles (Weiner and Kwiatek, 2022). Rare dark-pigmented dots were also noted by Ottoboni et al. (2017), who described them as brownish; however, no bristles were observed in that study. The author further reported that T. molitor cuticular fragments can be difficult to recognize because these dots are not consistently present. Under light microscopy, the bases of sensilla trichoidea (hair-like) and sensilla chaetica (bristle-like) appear as small puncture-like features marking the points where setae were originally attached (Harris, 1979; Marien et al., 2024).
Figure 3.
Fragments from T. molitor: 1 Urogomphi and anal spine fragments; 2 Muscle tissue; 3 Mandibular fragments; 4–10 Cuticular fragments; 11,14 Head capsule fragments; 12 Bristles; 13,15 Cuticle with tracheal structures; 16 Spiracle.
Figure 3.
Fragments from T. molitor: 1 Urogomphi and anal spine fragments; 2 Muscle tissue; 3 Mandibular fragments; 4–10 Cuticular fragments; 11,14 Head capsule fragments; 12 Bristles; 13,15 Cuticle with tracheal structures; 16 Spiracle.

Pupae of T. molitor are characterized by the presence of gin-trap structures (EURL-AP micrograph collection; Hinton, 1946; Wilson, 1971). No published literature was found regarding the presence of gin-traps in A. diaperinus. Mouthpart fragments, including mandibles, urogomphi, and leg fragments identifiable by their claws, are frequently observed. T. molitor possesses ten pairs of spiracles throughout all developmental stages (Raś et al., 2018). Immediately behind the occluding apparatus of each spiracle lies the tracheal vestibule, a spherical three-dimensional structure from which multiple tracheae originate and extend throughout the insect body (Raś et al., 2018). Each spiracle consists of a peritreme that encircles the atrial orifice (Raś et al., 2018).
Fragments of T. molitor can be distinguished from those of A. diaperinus mainly by their larger overall size; structures such as head capsules and mandibles therefore appear proportionally larger within feed material. T. molitor cuticle fragments also tend to exhibit a smoother surface. In contrast to A. diaperinus, T. molitor fragments lack a clearly defined surface pattern or an organized arrangement of bristles (Ottoboni et al., 2017). Veys and Baeten (2018) reported no clear distinction between fragments of T. molitor and A. diaperinus. Fragments of urogomphi are expected in both species, as documented in the EURL-AP micrograph collection for T. molitor and described for the larval stages of A. diaperinus in the literature (Figure 4; Chernaki et al., 2001). The presence of urogomphi in both sexes (Esquivel et al., 2012), which serve as reliable indicators of late-instar larvae, remains a definitive diagnostic feature for confirming Tenebrionid larval origin.
Figure 4.
Fragments from A. diaperinus: 1 Mandibular fragment; 2–6 Moderately sclerotized cuticle and thoracic leg segments; 7–8 Claws; 9,14–15 Spiracles; 10 Cuticle with sparse short setae; 11,12,16 Additional cuticle fragments; 13 Spiracle with tracheal system fragments.
Figure 4.
Fragments from A. diaperinus: 1 Mandibular fragment; 2–6 Moderately sclerotized cuticle and thoracic leg segments; 7–8 Claws; 9,14–15 Spiracles; 10 Cuticle with sparse short setae; 11,12,16 Additional cuticle fragments; 13 Spiracle with tracheal system fragments.

After the second instar, Coleopteran larvae and pupae show limited morphological variation (Park et al., 2014), yet they remain more morphologically differentiated than dipteran larvae, possessing developed mouthparts, legs, and a strongly sclerotized cuticle, which results in a higher proportion of rigid fragments in processed feed (Veys and Baeten, 2018). The larvae of both Coleopteran species lack denticles, in contrast to the denticle-bearing larvae of H. illucens (Veys and Baeten, 2018). The respiratory system of holometabolous insects consists of an internal network of cuticle-lined tracheae and tracheoles (Lowe et al., 2013; Iwan et al., 2015). During microscopic examination of insect meal, tracheolar fragments are more frequently observed in dipteran larvae such as H. illucens than in Coleopteran larvae (Veys and Baeten, 2018).
A wide diversity of particles can be observed under light microscopy in Orthopteran-derived PAP, as the meal is prepared from nymphs or imagos. These developmental stages possess legs, a well-developed head with eyes, mouthparts, and antennae, and, in the final nymphal instars, even wing pads are present (Veys and Baeten, 2018). The resulting particles include pigmented and sclerotized cuticular fragments; leg-derived structures such as tibial spines and femoral or tarsal pieces; sclerotized mandibular fragments; membranous or sclerotized wing fragments; eye material; cercal fragments; segmented antennal flagellomeres; honeycomb-like cuticle (notably in G. assimilis); cuticle bearing setae or isolated setae; claws; muscle tissue; tracheal system fragments; and muscle fibres associated with tracheae (Figure 5, Figure 6 and Figure 7; EURL-AP micrograph collection; Veys and Baeten, 2018).
Figure 5.
Fragments from A. domesticus. 1–3: cuticular fragments bearing setae; 4–5: segmented, cylindrical antennal flagellomeres; 6: tracheal structure; 7–8: tarsal claws; panel 9: isolated setae.
Figure 5.
Fragments from A. domesticus. 1–3: cuticular fragments bearing setae; 4–5: segmented, cylindrical antennal flagellomeres; 6: tracheal structure; 7–8: tarsal claws; panel 9: isolated setae.

Veys and Baeten (2018) reported the presence of distinct short, segmented fragments originating from antennae or leg parts in G. assimilis, a feature not observed in PAPs produced from H. illucens, T. molitor, or A. diaperinus.
The hind tibiae of G. assimilis are characterized by highly sclerotized, robust spines accompanied by a dense covering of coarse setae. Wing fragments show dense, reticulated, leathery venation. The cerci are covered in long sensory hairs, each arising from a distinctive cup-shaped base (trichobothria) visible under high magnification. Adults of G. assimilis share many general traits with A. domesticus, but they are typically darker and exhibit a denser brown pubescence, giving them a noticeably “hairier” appearance under magnification. The mandibles of G. assimilis appear dark brown to black.
Small, non-functional wing fragments with simplified venation, together with sclerotized brownish mandibles, are highly diagnostic for G. sigillatus. The hind tibiae bear comb-like fixed spines that are noticeably smaller and more slender than those of the larger G. assimilis. The abdominal cerci are long and covered in sensory hairs. The cuticular hairs of G. sigillatus tend to be lighter in colour, in contrast to the dark brown to black bristles typical of G. assimilis. As a result, fragmented material from G. assimilis often contains a higher density of dark, coarse setae, whereas A. domesticus exhibits lighter and finer hairs.
Figure 6.
Fragments from G. sigillatus. 1: cuticular fragment; 2: muscle tissue; 3–9: cuticular fragments with setae; 10–12: segmented antennal flagellomeres; 13: cuticle with isolated setae; 14–15: claws.
Figure 6.
Fragments from G. sigillatus. 1: cuticular fragment; 2: muscle tissue; 3–9: cuticular fragments with setae; 10–12: segmented antennal flagellomeres; 13: cuticle with isolated setae; 14–15: claws.

Figure 7.
Fragments from G. assimilis: 1 segmented antennal flagellomere; 2–4: cuticular fragments; 5: muscle tissue with trachea; 6: segmented antennal flagellomere; 7: honeycomb-like cuticle; 8: antennal flagellomere; 9, 15–16: claws; 10–12: cuticle; 13: trachea; 14: isolated setae.
Figure 7.
Fragments from G. assimilis: 1 segmented antennal flagellomere; 2–4: cuticular fragments; 5: muscle tissue with trachea; 6: segmented antennal flagellomere; 7: honeycomb-like cuticle; 8: antennal flagellomere; 9, 15–16: claws; 10–12: cuticle; 13: trachea; 14: isolated setae.

The EURL-AP micrograph collection covers only five insect species currently authorized as feed materials in the EU: H. illucens, T. molitor, A. diaperinus, A. domesticus, and G. assimilis. In H. illucens, the most frequently cited morphological structures include cuticle, anal segment, spine-like sensilla, tracheal system, muscle tissue, and spiracles. In T. molitor, reported fragments comprise cuticle, leg and claw pieces, tracheal system, urogomphi, antennae, anal spine, muscle, spiracle, gin trap, and labrum. For A. diaperinus, characteristic elements include cuticle, setae, legs, anal spine, tracheal system, muscle fibres, and mandibles. Different diagnostic patterns were photographed for the cricket species; G. assimilis is characterized by cuticle, antennae, muscle fibres, mandibles, and wing fragments, whereas A. domesticus typically presents cuticle, cercus fragments, setae, claws, tracheal system, eye fragments, muscle fibres, and wing fragments. Despite its importance for laboratory analysis of insect processed-protein detection in feed in accordance with EU Regulation 152/2009, the collection still lacks several authorized species.
Structural details of cuticular fragments, setae or trichoid sensilla, and tracheolar structures, together with characteristic patterns of muscle fibres, have been shown to provide robust identification criteria for determining the insect origin of particles (Simpson and Douglas, 2013; Veys and Baeten, 2018; Weiner and Kwiatek, 2022). Trichoid sensilla, or setae, are common in all insect-derived PAPs. They vary in quantity, colour, shape, and size, and may appear either still attached to cuticular fragments or as isolated elements within the sample (Simpson and Douglas, 2013; Veys and Baeten, 2018). According to Veys and Baeten (2018), these setae are entirely or nearly unpolarized under polarized light microscopy, a feature that allows them to be distinguished from plant trichomes, commonly present in feed materials such as wheat bran or gluten, which exhibit strong polarization (Veys and Baeten, 2018).
Fragments of insect tracheae and tracheoles may occur on larger cuticular pieces as well as within muscle fibres, confirming the insect origin of the muscle tissue (Veys and Baeten, 2018; Weiner and Kwiatek, 2022). In the study by Veys and Baeten (2018), these structures were reported predominantly in PAP derived from G. assimilis, whereas they were far less frequently observed in material from H. illucens, T. molitor, or A. diaperinus. Weiner and Kwiatek (2022) also noted tracheolar fragments, together with muscle-fibre elements in H. illucens and T. molitor. At higher magnification, the authors observed a spiral, transverse thickening characteristic of tracheoles. Veys and Baeten (2018) recommended searching for tracheal structures in cases where the identification of exoskeletal remains is uncertain.
Insect muscle fibres often display a characteristic zig-zag sarcomeric striation, a feature that distinguishes them from the muscle fibres found in terrestrial or fish PAPs (Veys and Baeten, 2018; Weiner and Kwiatek, 2022). Although this striation reliably confirms that the material is of insect origin, it does not allow identification at lower taxonomic levels (Weiner and Kwiatek, 2022). These fragments therefore serve only as indicators of the presence of insect material and are not species-specific. In addition, the presence of tracheal structures within cuticular fragments and muscle fibres supports the classification of the material as insect-derived rather than originating from other animal taxa (Veys and Baeten, 2018).
Veys and Baeten (2018) also discussed the differences between insect PAPs originating from nearly identical instars, either larval or nymphal stages, and contamination by insect pests, in which imagos are usually predominant. In such cases, remains of coleopteran elytra, membranous hindwings, compound eyes, antennae fragments, and other adult structures will inevitably be detected.
More specific methods for determining authorized insect species are recommended as a second step in feed monitoring, such as DNA-based approaches (van Raamsdonk et al., 2017; Weiner and Kwiatek, 2022). Molecular studies targeting sequences of T. molitor, H. illucens, and B. mori have been conducted by Debode et al. (2017) and Marien et al. (2018, 2025). It is important to emphasize, however, that DNA-based methods and other analytical techniques used in feed control should only be applied as complementary tools after light microscopy has confirmed the presence of PAP fragments. This approach continues the effective strategy that has been legally adopted for decades for the detection of PAPs in feed (Fumière et al., 2009; Veys et al., 2012; Lecrenier et al., 2016; Ottoboni et al., 2017; Veys and Baeten, 2018). Recently, Kaisin et al. (2025) proposed integrating automated recognition software with optical microscopy to reduce human error and accelerate identification.
Conclusion
In the EU, light microscopy, currently the official method for detecting prohibited animal proteins, including insect processed animal proteins, remains a robust and accessible analytical tool. It offers high sensitivity and requires only basic laboratory skills and equipment. Light microscopy is effective in identifying insect PAPs, particularly cuticular fragments and intact structures such as mandibles, legs, or antennal segments, even after milling and drying. However, it does not provide species-level identification and cannot distinguish authorized from non-authorized insect species. For this reason, the continued integration of microscopy with complementary methods will be essential to meet future regulatory and analytical requirements.
There is now a widely recognized need for more specialized training to support taxonomic discrimination of insect species, beyond the standard training currently provided to control laboratories. Integrating automated recognition software with optical microscopy to reduce human error and accelerate identification is also crucial for addressing the limitations of light microscopy.
This work provides guidance for identifying particles of authorized insects in feed. Although key aspects of microscopic identification have been addressed, careful interpretation remains essential. The proposed identification key is useful when fragments are abundant and species-specific, but its diagnostic value decreases when only small, non-distinct structures such as tracheoles or muscle fibres are present. Overall, microscopic detection of insect particles is far more effective when supported by a comprehensive photographic database to assist technicians in routine analysis.
Methodology
Slides were prepared according to the specifications outlined in Section 2.1.2.1.4.4 of Annex VI to Regulation (EC) No 152/2009, as well as the EURL-AP Standard Operating Procedure (SOP) for staining reagents. Pure meals of Hermetia illucens, Tenebrio molitor, Alphitobius diaperinus, Acheta domesticus, Gryllodes sigillatus, and Gryllus assimilis were used for the preparation of reference slides. Laboratory reference material of Bombyx mori was acquired as an industrial ready-made meal. Material from Musca domestica remains difficult to obtain; therefore, morphological features for this species were extracted from the available scientific literature. Reference material observation and preparation were carried out in the Feed and Food Microscopy Laboratory at AGROLAB Alimentalia (Italy). Characteristic features of insects were determined by microscopic images using the Zeiss Axiovert 25 Inverted Phase Contrast Microscope. Observations were conducted at magnifications of 10X, 20X, and 32X, each paired with a 10X ocular lens. A 0.4 N.A. condenser equipped with a stage and phase slider was employed during examination. Photographic documentation of the slides included an additional 2.5X magnification.
Microsoft Copilot was used to assist with the structural refinement of the identification key; all scientific interpretations and taxonomic decisions were made by the authors.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgments
The author expresses sincere gratitude to Dr. Giulio Lora, Site Manager at AGROLAB Alimentalia S.r.l., Italy, for granting access to microscopy tools and internal reference materials essential to this work. Special thanks are extended to Dr. Arianna Salvador, Lab Leader of Microbiology and Molecular Biology, for permitting the use of the food and feed microscopy laboratory for image capture and slide observation. Appreciation is also due to Dr. Enrico Goldin, Head of CRM Food, and his team for kindly providing samples and reference material analyzed in this study. The author used Microsoft Copilot to support English language editing and structural improvement.
References
- Regulation (EC) No 999/2001 of 22 May 2001 laying down rules for the prevention, control and eradication of certain transmissible spongiform encephalopathies. Official Journal of the European Communities, L 147, 1–40.
- Regulation (EC) No 178/2002 of 28 January 2002 laying down the general principles and requirements of food law, establishing the European Food Safety Authority and laying down procedures in matters of food safety. Official Journal of the European Communities, L 31, 1–24.
- Regulation (EC) No 852/2004 of 29 April 2004 on the hygiene of foodstuffs. Official Journal of the European Union, L 139, 1–54.
- Regulation (EC) No 183/2005 of 12 January 2005 laying down requirements for feed hygiene. Official Journal of the European Union, L 35, 1–22.
- Regulation (EC) No 152/2009 of 27 January 2009 laying down the methods of sampling and analysis for the official control of feed. Official Journal of the European Union, L 54, 1–130.
- Regulation (EC) No 1069/2009 of 21 October 2009 laying down health rules as regards animal by-products and derived products not intended for human consumption and repealing Regulation (EC) No 1774/2002. Official Journal of the European Union, L 300, 1–33.
- Commission Regulation (EU) No 142/2011 of 25 February 2011 implementing Regulation (EC) No 1069/2009 of the European Parliament and of the Council laying down health rules as regards animal by-products and derived products not intended for human consumption and implementing Council Directive 97/78/EC regarding certain samples and items exempt from veterinary checks at the border under that Directive. Official Journal of the European Union, L 54, 1–254.
- Regulation (EU) No 51/2013 of 16 January 2013 amending Regulation (EC) No 152/2009 as regards the methods of analysis for the determination of constituents of animal origin for the official control of feed. Official Journal of the European Union, L 20, 33–43.
- Regulation (EU) 2017/893 of 24 May 2017 amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council and Annexes X, XIV and XV to Commission Regulation (EU) No 142/2011 as regards the provisions on processed animal protein. Official Journal of the European Union, L 138, 92–116.
- Regulation (EU) 2020/1560 of 26 October 2020 amending Annex VI to Regulation (EC) No 152/2009 laying down the methods of analysis for the determination of constituents of animal origin for the official control of feed. Official Journal of the European Union, L 357, 17–23.
- Implementing Regulation (EU) 2021/404 of 24 March 2021 establishing the lists of third countries, territories or zones thereof from which entry into the Union of animals, germinal products and products of animal origin is authorised in accordance with Regulation (EU) 2016/429 of the European Parliament and of the Council. Official Journal of the European Union, L 114, 1–237.
- Implementing Regulation (EU) 2022/893 of 7 June 2022 amending Annex VI to Regulation (EC) No 152/2009 as regards the methods of analysis for the detection of constituents of terrestrial invertebrates for the official control of feed. Official Journal of the European Union, L 155, 24–35.
- Apasrawirote, D.; Boonchai, P.; Muneesawang, P.; et al. Assessment of deep convolutional neural network models for species identification of forensically important fly maggots based on images of posterior spiracles. Sci. Rep. 2022, 12, 4753. [CrossRef]
- Barros, L.M.; Gutjahr, A.L.N.; Ferreira-Keppler, R.L.; Martins, R.T. Morphological description of the immature stages of Hermetia illucens (Linnaeus, 1758) (Diptera: Stratiomyidae). Microsc. Res. Tech. 2019, 82(3), 178–189. [CrossRef]
- Chernaki, A.M.; Almeida, L.M. Morphology of the immature stages and adult of Alphitobius diaperinus. Rev. Bras. Zool. 2001, 18(2), 351–363. [CrossRef]
- Cortinhas LB, Martins Mendonça P, Braga MV, Queiroz MMC. Ultrastructure of the Immature Stages of Musca domestica (Diptera: Muscidae: Muscinae). J Med Entomol. 2020 Nov 13;57(6):1712-1721. [CrossRef]
- Debode, F.; Marien, A.; Gérard, A.; et al. Development of real-time PCR tests for the detection of Tenebrio molitor in food and feed. Food Addit. Contam. Part A 2017, 34(8), 1421–1426. [CrossRef]
- EFSA. Scientific opinion on a risk profile related to production and consumption of insects as food and feed. EFSA J. 2015, 13, 4257.
- Esquivel, J.F.; Crippen, T.L.; Ward, L.A. Improved visualization of Alphitobius diaperinus—Part I: Morphological features for sex determination. Psyche 2012, 328478. [CrossRef]
- EURL-AP Micrograph Collection. Animal structures. Available online: https://www.eurl.craw.eu/publications/micrograph-collection/animal-structures/ (accessed on 24 January 2026).
- Fabian, B.; Schneeberg, K.; Löwe, S.; et al. Transformations of head structures during larval development of Hermetia illucens. J. Morphol. 2025, 286, e70048. [CrossRef]
- Fumière, O.; Veys, P.; Boix, A.; et al. Methods of detection and identification of PAPs. Biotechnol. Agron. Soc. Environ. 2009, 13, 59–70.
- Grzywacz, A.; Hall, J.R.; Pape, T.; Szpila, K. Muscidae of forensic importance. Int. J. Legal Med. 2017, 131(3), 855–866. [CrossRef]
- Harris, R. A Glossary of Surface Sculpturing; California Department of Food and Agriculture: Sacramento, CA, USA, 1979.
- Hinton, H.E. The gin-traps of beetle pupae. Trans. R. Entomol. Soc. Lond. 1946, 97, 473–496.
- Lecrenier, M.C.; Marbaix, H.; Dieu, M.; et al. Identification of bovine blood biomarkers. Food Chem. 2016, 213, 417–424.
- Marien, A.; Debode, F.; Aerts, C.; et al. Detection of Hermetia illucens by real-time PCR. J. Insects Food Feed 2018, 4(2), 91–96. [CrossRef]
- Marien, A.; Dubois, B.; Anselmo, A.; et al. Authentication of insect-based products. Insects 2025, 16(7), 729. [CrossRef]
- Marien, A.; Dubois, B.; Anselmo, A.; et al. Detection of Bombyx mori in feed by real-time PCR. Agriculture 2024, 14, 1996. [CrossRef]
- Oliveira, F.R.; Doelle, K.; Smith, R.P. External morphology of Hermetia illucens. Ann. Res. Rev. Biol. 2016, 9(5), 1–10.
- Ottoboni, M.; Tretola, M.; Cheli, F.; et al. Light microscopy with differential staining for insect PAPs. Food Addit. Contam. Part A 2017, 34(8), 1377–1383. [CrossRef]
- Park, J.B.; Choi, W.H.; Kim, S.H.; et al. Developmental characteristics of Tenebrio molitor. Int. J. Ind. Entomol. 2014, 28(1), 5–9.
- Raś, M.; Iwan, D.; Kamiński, M.J. Tracheal system in holometabolous insects. J. Anat. 2018, 232, 997–1015. [CrossRef]
- Rebora, M.; Salerno, G.; Piersanti, S.; et al. The armoured cuticle of Hermetia illucens. Sci. Rep. 2023, 13, 22101. [CrossRef]
- Simpson, S.J.; Douglas, A.E. The Insects: Structure and Function, 5th ed.; Cambridge University Press: Cambridge, UK, 2013.
- Sultana, R.; Sanam, S.; Kumar, S.; et al. A review of Gryllidae with new species records. ZooKeys 2021, 1078, 1–33. [CrossRef]
- Szpila, K. Key for identification of blowfly larvae. In Current Concepts in Forensic Entomology; Springer: Dordrecht, The Netherlands, 2009.
- van Raamsdonk, L.W.D.; van der Fels-Klerx, H.J.; de Jong, J. New feed ingredients: The insect opportunity. Food Addit. Contam. Part A 2017, 34(8), 1384–1397. [CrossRef]
- Veys, P.; Baeten, V. Protocol for isolation of insect PAPs. Food Control 2018, 92, 496–504. [CrossRef]
- Walczak, K.; Grzywacz, A. Identification key to early instar Muscidae. Forensic Sci. Int. 2024, 360, 112028. [CrossRef]
- Weiner, A.; Kwiatek, K. Microscopic detection of insects in poultry feeds. J. Vet. Res. 2022, 66(1), 93–100. [CrossRef]
- Wilson, M.C.L. Morphology and mechanism of pupal gin-traps. J. Stored Prod. Res. 1971, 7(1), 21–30. [CrossRef]
- Zdeňková, K.; Čermáková, E.; Vejl, P.; Čermáková, A.; Vašek, J. Analytical methods for identification of edible and feed insects. Foods 2025, 14(11), 2002. [CrossRef]

Table 1.
The specific species and their respective taxonomic orders and families are as follows:.
| Common Name | Scientific Name | Order | Family |
| Black soldier fly | Hermetia illucens | Diptera | Stratiomyidae |
| Common housefly | Musca domestica | Diptera | Muscidae |
| Yellow mealworm | Tenebrio molitor | Coleoptera | Tenebrionidae |
| Lesser mealworm | Alphitobius diaperinus | Coleoptera | Tenebrionidae |
| House cricket | Acheta domesticus | Orthoptera | Gryllidae |
| Banded cricket | Gryllodes sigillatus | Orthoptera | Gryllidae |
| Field cricket | Gryllus assimilis | Orthoptera | Gryllidae |
| Silkworm | Bombyx mori | Lepidoptera | Bombycidae |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



