
Submitted:
23 January 2026
Posted:
23 January 2026
You are already at the latest version
Abstract
Sub-Saharan Africa is undergoing a rapid nutritional and epidemiological transition, characterized by a rising incidence of Type 2 Diabetes (T2D) among lean populations. While the "Westernization" of the gut microbiome is well-documented globally, the specific ecological shifts driving metabolic dysfunction in the African context remain under-synthesized. This systematic review aims to catalog diet-driven taxonomic alterations and evaluate their mechanistic link to T2D. A systematic search was conducted following PRISMA guidelines using PubMed, Scopus, and Google Scholar (2015–2026). Studies were included if they compared the gut microbiomes of urban vs. rural African populations and reported metabolic outcomes. Quality was assessed using the Newcastle-Ottawa Scale. Twelve studies meeting inclusion criteria were identified, representing a pooled cohort of N=6,097 participants across 22 countries. The analysis revealed a consistent "rural signature" marked by the enrichment of fiber-degrading Treponema and Prevotella ("VANISH" taxa). Urbanization was associated with the significant depletion of these guilds and a compensatory bloom of Bacteroides and Bifidobacterium. This taxonomic shift was statistically correlated with elevated fasting blood glucose, HbA1c, and diabetes prevalence in urban cohorts. Mechanistically, the loss of Treponema likely compromises SCFA-mediated insulin signaling, while the rise of mucin-degrading Bacteroides may drive metabolic endotoxemia. Urbanization in Africa is associated with the loss of key ancestral microbes that protect metabolic health. Given that commercial probiotics often contain taxa already enriched in urban Africans (Bifidobacterium), we propose that indigenous fermented foods (e.g., Ogi, Kunu), which provide a Lactobacillus-rich acidic niche, represent a more ecologically appropriate therapeutic strategy to restore metabolic homeostasis. All identified associations are observational, and causal inferences cannot be established.
Keywords:
gut microbiome
; Type 2 Diabetes
; nutritional transition
; dietary fiber
; Sub-Saharan Africa
; indigenous fermented foods
1. Introduction: The Silent Epidemic
The current epidemiology of Type 2 Diabetes (T2D) in Sub-Saharan Africa (SSA) is characterized by a rapid transition from a historically rare condition, affecting less than 1% of the population prior to 1985, to a major public health crisis, with an estimated 24 million adults living with the disease as of 2021[1,2]. While urbanization and lifestyle changes are driving localized increases in adiposity [3,4], a distinct epidemiological disconnect exists regarding the clinical phenotype of African patients compared to those in the West. Unlike high-income countries where over 80% of diabetes cases are inextricably linked to overweight or obesity and insulin resistance, evidence suggests that the majority of diabetes patients in the general SSA population are young, lean, and characterized primarily by beta-cell secretory dysfunction rather than the insulin resistance typically observed in Western cohorts [5]. Despite this phenotypic variation, the region faces a severe projected escalation in non-communicable diseases. Current projections estimate a 134% surge in cases by 2045, the steepest rise globally [2], compounded by a diagnostic gap where 60% of individuals remain undiagnosed [1].
1.1. Loss of Diversity’ and the ‘Westernization’ of the Microbiome
The concept of the ‘Westernization’ of the microbiome refers to a distinct shift in the composition and function of the human gut microbiota associated with industrialization and urbanization [6]. This phenomenon is characterised by a significant ‘loss of diversity,’ defined as a reduction in the richness and evenness of microbial species residing in the gut [7].
This transition manifests in several key ecological alterations:
- Depletion of Specialist Taxa: The process involves the disappearance of specific bacterial clades often termed ‘VANISH’ (Volatile and/or Associated Negatively with Industrialized Societies of Humans) taxa [7,9]. These include fibre-degrading species such as Treponema, Succinivibrio, and Xylanibacter [10,11,12].
- Rise of Generalists and Pathobionts: As specialist species disappear, there is a bloom of “BloSSUM” (Bloom or Selected in Societies of Urbanization/Modernization) taxa, such as Bacteroides and Enterobacteriaceae, which thrive on host-derived nutrients (e.g., mucins) and are often resilient to antibiotics and bile acids [7,13].
1.2. The ‘Old Friends’ and ‘Missing Microbes’ Hypotheses: Urbanization as an Ecological Transition in Africa
Building on the concept of microbiome Westernization, the ‘Missing Microbes’ and ‘Old Friends’ hypotheses provide a useful concept for understanding how rapid urbanization may influence metabolic and immune health in African populations. The ‘Missing Microbes’ hypothesis proposes that cumulative features of modern life; including widespread antibiotic use, caesarean delivery, and reduced microbial exposure in early life; have led to the progressive loss of ancient, co-evolved members of the human gut microbiota [14]. The related ‘Old Friends’ hypothesis extends this concept by suggesting that regular exposure to these microbes is required for proper immune regulation; in their absence, immune responses may become dysregulated, contributing to inflammatory and metabolic diseases [7,15].
In the African context, these hypotheses carry particular relevance as the continent undergoes a rapid epidemiological and nutritional transition [11]. African populations, especially those living in rural or traditional settings such as hunter-gatherer communities, currently harbour some of the most diverse gut microbiomes globally, characterized by the presence of taxa commonly described as “Old Friends,” including Prevotella, Treponema, and Xylanibacter [11]. These taxa are largely absent from populations in high-income countries, where industrialization-associated microbial loss is already well established.
Unlike Western settings, where the loss of ancestral microbes has largely occurred historically, microbial erosion in Africa is unfolding contemporaneously. Urbanization and the adoption of Westernized dietary patterns are associated with a measurable decline in fibre-adapted microbial taxa and overall microbial diversity [11]. Comparative studies demonstrate that rural African populations retain microbial communities associated with high-fibre diets and reduced inflammatory markers, whereas these features are diminished in urban African cohorts [6,11]. In this sense, urbanization represents an active ecological transition within the gut microbiome, rather than a completed state.
This microbial shift has been linked to parallel increases in non-communicable diseases (NCDs), including asthma, obesity, and Type 2 Diabetes, reflecting health trajectories previously observed in industrialized regions [16]. Within this framework, the “missing microbes” are not merely theoretical losses but may constitute a functional deficit in immune and metabolic education, particularly when microbial disruption occurs during early life [11].
Viewed ecologically, urbanization can be conceptualized as a process analogous to biodiversity loss within the human gut ecosystem. The rapid transition from traditional to industrialized living environments creates a mismatch between the human genome and its long-standing microbial partners, destabilizing co-evolved metabolic and immunoregulatory networks [15]. This process is characterized by the progressive loss of ancestral taxa that function as ecological keystones, including Treponema and Oxalobacter, which provide critical services such as complex fibre fermentation and modulation of host inflammatory responses [12,14,17]. As these functional guilds decline, ecological niches within the gut may be occupied by inflammation-associated pathobionts, contributing to a dysbiotic microbiome state [13]. Such industrialized microbiome configurations have been associated with reduced resilience and impaired capacity to support host metabolic homeostasis, potentially contributing to the rising burden of obesity, autoimmune conditions, and chronic inflammatory diseases observed during epidemiological transition [14].
1.3. Rationale and the “African Paradox”
While the theoretical frameworks of “Westernization” and “Missing Microbes” are well-established globally, critical gaps remain in applying these models to Sub-Saharan Africa. First, the majority of existing microbiome-diabetes literature is derived from Western cohorts where Type 2 Diabetes is inextricably linked to obesity and insulin resistance [5]. This creates a phenotypic discordance, as the African diabetic phenotype is frequently characterized by lean body mass and primary beta-cell failure rather than adiposity-driven insulin resistance [2,5]. Consequently, microbial markers of “dysbiosis” identified in Western studies (e.g., low Akkermansia) may not be biologically relevant to the African context.
Second, global reviews often aggregate “non-Western” populations into a single category, obscuring the specific ecological distinctiveness of the African gut. For example, the loss of Treponema—a taxon abundant in rural Africa but effectively extinct in the West—is rarely addressed in standard diabetes literature, yet it may represent a crucial, region-specific metabolic regulator [11]. Finally, there is a therapeutic gap: current probiotic interventions are based on Western gut profiles (e.g., supplementing Bifidobacterium). Without a systematic understanding of the specific taxa lost during African urbanization, therapeutic strategies risk being ecologically redundant or ineffective.
Therefore, this systematic review aims to bridge this gap by:
- I.
- cataloguing the specific gut microbiome shifts associated with urbanization in African populations;
- II.
- evaluating the strength of the evidence linking “rural” taxa extinction to metabolic dysfunction;
- III.
- critically assessing the potential of indigenous fermented foods (Ogi and Kunu) to restore these lost functional guilds. This review follows the PRISMA 2020 (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) guidelines
2. Materials and Methods
2.1. Search Strategy and Data Sources
This systematic review was conducted in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines [47]. A comprehensive search was performed on PubMed, Google Scholar, and Scopus for articles published between January 1, 2015, and January 20, 2026. Earlier studies were included to capture foundational rural–urban microbiome contrasts, while emphasis was placed on studies published after 2020. Google Scholar was used as a supplementary source to identify recently published studies not yet indexed in PubMed or Scopus. The search strategy employed two distinct concept clusters to capture both dysbiosis and potential therapeutic interventions:
- (Microbiome Cluster): (“gut microbiome” OR “gut microbiota”) AND (“Africa” OR “Sub-Saharan”) AND (“urbanization” OR “rural” OR “modernization”)
- (Metabolic Cluster): (“diabetes” OR “glucose” OR “insulin”) AND (“gut microbiome”) AND (“Africa”)
- (Therapeutic Cluster): (“fermented food” OR “Ogi” OR “Kunu”) AND (“microbiome” OR “probiotic”) Reference lists of eligible articles were hand-searched to identify additional relevant studies (“snowballing”).
To ensure comprehensiveness, the search process was refined by: (1) restricting results to peer-reviewed articles published in the English language; (2) manually screening the reference lists of all eligible articles to identify relevant studies not indexed in standard databases (“snowballing”); and (3) utilizing Google Scholar to capture grey literature and recent preprints. Duplicate records were removed prior to screening
2.2. Eligibility Criteria
Studies were selected based on the PICO (Population, Intervention, Comparison, Outcome) framework:
- Population: Human participants residing in Sub-Saharan Africa (e.g., Nigeria, Ghana, South Africa, Tanzania) or African migrants in Western nations.
- Exposure: Urbanization, defined as residence in urban centers compared to rural/traditional settings, or consumption of Westernized vs. traditional diets.
- Comparators: Rural, hunter-gatherer, or traditional agriculturalist populations.
- Outcomes: Quantitative data on gut microbiome composition (16S rRNA or Metagenomic sequencing) AND/OR metabolic markers (Fasting Blood Glucose, HbA1c, Insulin, BMI).
- Exclusion: Reviews, animal studies, editorials, and studies published in non-English languages were excluded.
2.3. Study Selection and Data Extraction
Initial screening of titles and abstracts was conducted to exclude irrelevant records. Full-text articles of potentially eligible studies were retrieved and assessed against inclusion criteria. Data were extracted into a structured matrix including: Author/Year, Country, Sample Size (N), Sequencing Method (16S vs. Shotgun), Key Bacterial Taxa (enriched/depleted), and Metabolic Outcomes.
2.4. Quality Assessment
The methodological quality of included studies was assessed using a modified Newcastle-Ottawa Scale (NOS) adapted for cross-sectional microbiome studies. Three primary domains were evaluated:
- Selection Bias: Was the sample representative of the target population? (e.g., random sampling vs. small convenience cohorts).
- Comparability: Did the study control for key confounders? (e.g., age, sex, BMI, or diet).
- Outcome Assessment: Was the sequencing methodology valid and reproducible? (e.g., 16S/Metagenomics vs. unclear methods).
Based on these domains, studies were stratified into three quality tiers:
- Good: Low risk of bias in both Selection and Comparability domains.
- Fair: High risk of bias in one domain (typically small sample size or lack of multivariate adjustment).
- Poor: High risk of bias in multiple domains or insufficient reporting of methods. Full risk of bias ratings for each study are detailed in Supplementary Table S1.
2.5. Data Synthesis
Due to the heterogeneity of sequencing platforms (different 16S variable regions vs. metagenomics) and reporting formats, a quantitative meta-analysis was not feasible. Instead, a narrative synthesis was conducted. Microbiome results were grouped taxonomically to identify consistent “signature” taxa associated with urbanization. Metabolic outcomes were synthesized to evaluate the strength of the association between specific microbial enterotypes and Type 2 Diabetes prevalence.
3. Results
3.1. Study Characteristics
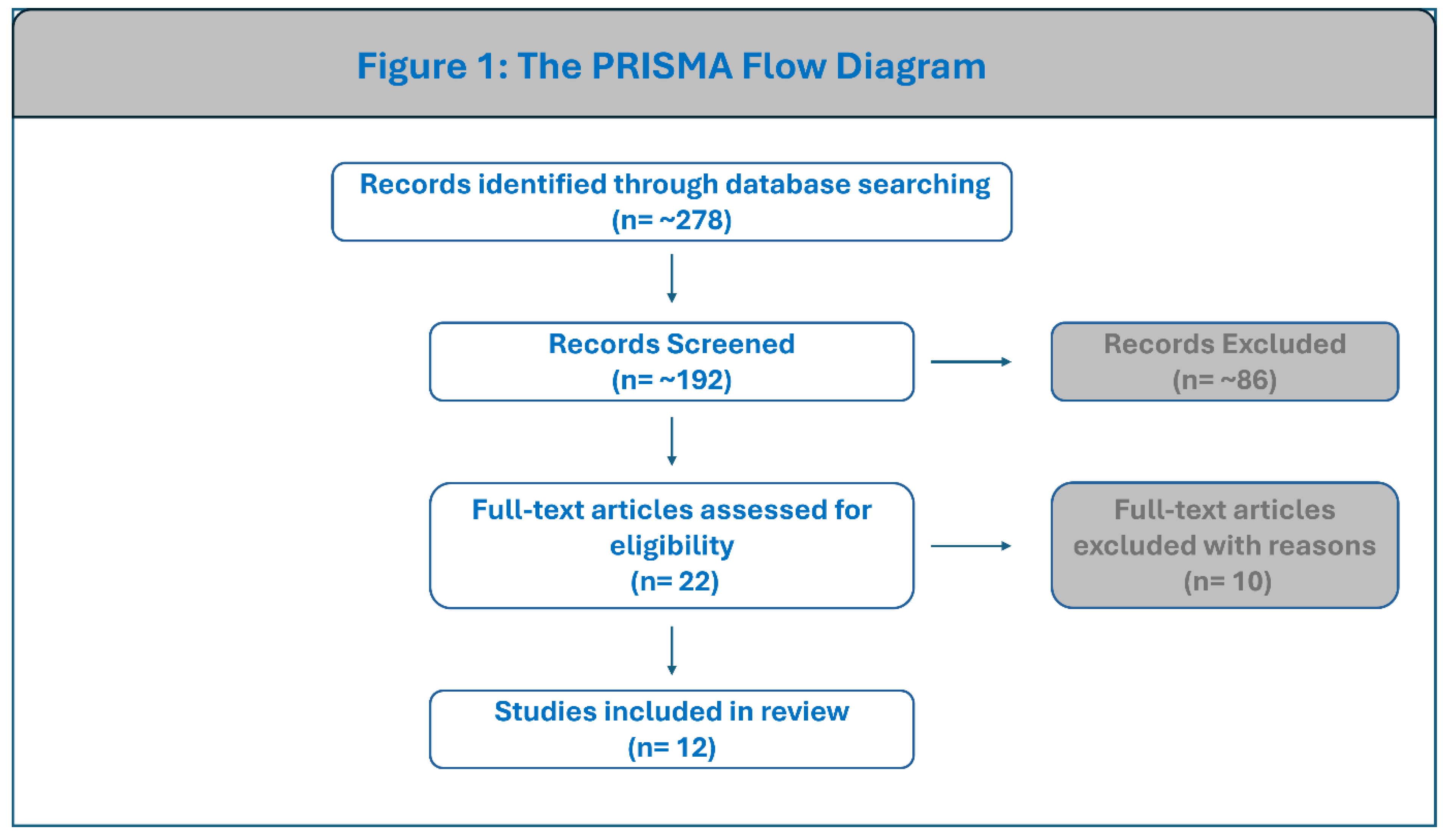
A total of 12 studies published between 2017 and 2025 were included, representing a pooled cohort of approximately 6,100 individuals across 22 countries, including Nigeria, Ghana, South Africa, Tanzania, Zambia, Burkina Faso, and Cameroon, as well as comparator populations in Europe, the Middle East, and the Americas. The majority of these studies (n=9) utilized 16S rRNA gene sequencing to characterize the microbiome, while a subset of recent large-scale studies (n=3) employed shotgun metagenomics for higher-resolution profiling. The limited number of eligible studies reflects the current scarcity of comparative rural–urban microbiome research in African populations rather than restrictive inclusion criteria. Detailed characteristics of the 12 included studies are summarized in Table 1.
Figure 1.
PRISMA flow diagram of study selection. The flowchart illustrates the sequential screening process, from identification of 278 records to the final inclusion of 12 studies. Parts of this figure were drawn using pictures from Servier Medical Art. Servier Medical Art by Servier is licensed under a Creative Commons Attribution 3.0 Unported License (https://creativecommons.org/licenses/by/3.0/).
Figure 1.
PRISMA flow diagram of study selection. The flowchart illustrates the sequential screening process, from identification of 278 records to the final inclusion of 12 studies. Parts of this figure were drawn using pictures from Servier Medical Art. Servier Medical Art by Servier is licensed under a Creative Commons Attribution 3.0 Unported License (https://creativecommons.org/licenses/by/3.0/).
Methodological Heterogeneity and Robustness
Significant heterogeneity was observed in sequencing methodologies, with nine studies utilizing 16S rRNA amplicon sequencing and three recent large-scale studies employing shotgun metagenomics . Despite this variation in resolution, the core ecological findings were remarkably consistent across platforms. The depletion of Treponema and Prevotella in urban cohorts was identified with equal statistical significance in both low-resolution 16S studies (e.g., Angelakis et al. [18]) and high-resolution metagenomic datasets (e.g., Maghini et al. [17]), suggesting that the “urbanization signature” is a robust biological signal rather than a methodological artifact.
The methodological quality of the included studies was generally good, though three studies were rated as ‘Fair’ due to small sample sizes (see Supplementary Table S1).
3.2. The “Urbanization Signature” (The Microbiome Data)
Based on the systematic review of the 12 identified studies, there is a distinct dichotomy in the bacterial genera associated with rural versus urban populations, as well as a complex narrative regarding microbial diversity.
3.2.1. Rural-Enriched Genera: The Prevalence of Prevotella and Treponema
Across the studies, the most consistent signature of rural, traditional, or non-industrialized populations is the enrichment of bacteria capable of degrading complex polysaccharides and fiber. The genus Prevotella appears in almost every study as the dominant taxon in rural cohorts, including populations in Burkina Faso, Ghana, Tanzania, and Namibia [18,19,20,21].
Specifically, the genus Treponema (often identified as Treponema succinifaciens) serves as a distinct biomarker for traditional lifestyles. It was reported as significantly enriched or exclusively present in rural/traditional populations in 9 out of the 12 studies analyzed [18,19,20,21,22,23,24,25,26]. These bacteria are frequently described as “old friends” or “VANISH” (Volatile and/or Associated Negatively with Industrialized Societies of Humans) taxa, which are functionally adapted to high-fiber diets but are lost rapidly upon urbanization or migration [19,21]. Alongside Treponema and Prevotella, the genus Succinivibrio was also frequently cited as a rural marker, particularly in studies from Tanzania, Ghana, and Cameroon [18,19,20,24].
3.2.2. Urban-Enriched Genera: The Rise of Bacteroides and Bifidobacterium
In contrast, urban and westernized populations consistently displayed an enrichment of “BloSSUM” (Bloom or Selected in Societies of Urbanization/Modernization) taxa (Figure 2). The genus Bacteroides was repeatedly identified as a hallmark of urban living, found in higher abundance in urban cohorts from Ghana, South Africa, Tanzania, and comparative Western populations in the USA and Italy [19,23,25,26].
Similarly, the genus Bifidobacterium was consistently enriched in urban populations. It was identified as a key urban discriminator in multiple large-scale studies, including those by Maghini et al. [26], Pyuza et al. [19] and Angelakis et al. [18] often correlating with the consumption of dairy products and processed foods common in industrialized settings [19,23,26]. In the South African cohorts, Bifidobacterium was specifically associated with the urban Soweto group compared to the rural Bushbuckridge group [27]. Other genera frequently associated with the urban environment included Blautia, Faecalibacterium, and members of the Enterobacteriaceae family, such as Escherichia and Shigella [19,21].
3.2.3. Trends in Gut Microbiome Diversity
The “loss of diversity” trend—where urbanization supports the hypothesis that a reduction in microbial richness—was the dominant narrative across most studies, though notable exceptions highlight the complexity of the African context.
The majority of the analyzed papers reported significantly higher alpha diversity (species richness and evenness) in rural populations compared to their urban or westernized counterparts. For instance, rural Ghanaians showed significantly greater diversity than urban Ghanaians and migrants in Amsterdam [19,21]. Similarly, rural Tanzanians exhibited higher diversity than urban residents, a trait linked to the consumption of diverse, fiber-rich rural diets [19]. Angelakis et al. [18] noted that traditional rural populations had significantly richer microbiomes compared to urban French and Saudi cohorts, suggesting that modernization depletes the “ancestral” human microbiota [18].
However, this trend was not universal. Two studies reported the opposite pattern, where rural populations had lower diversity than urban ones. Afolayan et al. [22] found that the nomadic Fulani of Nigeria had significantly lower species richness compared to the semi-urban Jarawa [22]. The authors hypothesized that the Fulani’s specific, narrower diet (heavily reliant on dairy and specific grains) and isolation might limit microbial diversification compared to the more varied diet of the urbanized Jarawa. Similarly, Hodges et al [46]. found reduced alpha diversity in rural Zambian controls compared to urban cohorts. These findings suggest that the “loss of diversity” model is not absolute and may depend on specific cultural dietary restrictions, environmental exposures, or the transitional nature of the specific “rural” communities studied [7,19,22]. A detailed summary of the specific taxonomic alterations observed in each included study is provided in Table 2.
3.3. The Metabolic Link: Urbanization and Diabetes Prevalence
Based on data from Verhaar et al. [21] Ecklu-Mensah et al. [25] and Pyuza et al. [19] there is a consistent and statistically significant trend where urbanization and migration to Western environments are associated with a higher prevalence of Type 2 Diabetes (T2D) and elevated blood glucose levels.
3.3.1. Epidemiological Gradients of Hyperglycemia
In the study by Verhaar et al. [21] which examined Ghanaian populations across a migration gradient, the prevalence of T2D progressively increased from the rural setting to urban and migrant settings. Specifically, diabetes prevalence was lowest in rural Ghana (6.6%), nearly doubled in urban Ghana (11.7%), and was highest among Ghanaian migrants living in Amsterdam (16.5%). This trend was mirrored by HbA1c levels, which were significantly higher in the urban and migrant groups (41.9 and 40.0 mmol/mol, respectively) compared to the rural group (33.2 mmol/mol) [21].
Ecklu-Mensah et al. [25] observed a similar disparity when comparing populations across the epidemiological transition. They reported that diabetes prevalence was significantly lower in low-to-middle-income countries compared to high-income countries like the United States [25]. For instance, the prevalence of diabetes in Ghanaian women was 6.8% compared to 19.9% in women from the US, while South African women had a prevalence of only 3.5%. A similar pattern was observed in men, where diabetes prevalence was as low as 1.0% in Ghana compared to 17.5% in the US [25].
Even in cohorts screened for chronic disease, urbanization impacted glycemic control. Pyuza et al. [19] studied a cohort in Tanzania, excluding individuals with known chronic diseases to focus on healthy adults. Despite this exclusion, they found a statistically significant difference in glycemic control between locations: participants living in urban Moshi had significantly higher random blood sugar levels (5.30 mmol/L) compared to those living in rural Moshi (4.95 mmol/L) [19].
3.3.2. Bacterial Correlates of Metabolic Dysfunction
Regarding the statistical links between specific bacteria and metabolic health, these studies identified direct associations between the loss of traditional “rural” microbes and adverse metabolic outcomes. Verhaar et al. [21] classified a specific set of bacteria as “VANISH” taxa—microbes such as Treponema, Succinivibrio, and Prevotella that are abundant in rural populations but disappear in urbanized groups [21]. They found a direct statistical link where participants lacking these “VANISH” microbes had a significantly higher prevalence of diabetes, higher HbA1c levels, and higher body mass index (BMI) compared to individuals who retained them [21].
Ecklu-Mensah et al. [25] also found significant associations between microbial composition and adiposity. They observed that the Prevotella enterotype, which dominates in African populations, was statistically correlated with a lower BMI compared to the Bacteroides enterotype found in Western populations [25]. Furthermore, they identified specific taxa such as Christensenella and Oscillospira that were significantly enriched in non-obese individuals, whereas obese individuals showed an enrichment of Lachnospira and Dialister [25]. Additionally, Maghini et al. [17] explicitly noted that individuals carrying Treponema succinifaciens had a significantly lower hip circumference, a key anthropometric marker for obesity and metabolic risk, further supporting the link between these non-industrialized bacterial strains and metabolic health [17].
3.3.3. Strength of Association and Effect Magnitude
While a quantitative meta-analysis was precluded by data heterogeneity, the narrative synthesis reveals a high magnitude of effect regarding urbanization and metabolic risk. The association between urban living and hyperglycemia was not merely statistically significant but clinically profound. Verhaar et al. reported a near-doubling of diabetes prevalence (6.6% to 11.7%) between rural and urban Ghanaians, with migrants exhibiting a further increase to 16.5% [21]. Similarly, Ecklu-Mensah et al. observed an approximate three-fold disparity in diabetes prevalence between Ghanaian men (1.0%) and African American men (17.5%) [25]. These distinct “step-wise” increases in disease burden across the rural-urban-migrant gradient provide strong epidemiological evidence for a dose-dependent relationship between westernized lifestyle exposure and metabolic dysfunction.
4. Discussion
4.1. Confirmation of the “Invisible Extinction” in Africa
The systematic review of these 12 African studies provides robust empirical support for Blaser’s “disappearing microbiota” hypothesis, identifying the depletion of Treponema species—particularly Treponema succinifaciens—as a definitive biomarker of urbanization [14]. Consistent with the global theory that industrialization precipitates the loss of “VANISH” (Volatile and/or Associated Negatively with Industrialized Societies of Humans) taxa, the African data demonstrates that Treponema and Prevotella are ubiquitous in rural cohorts—such as the Xhosa in South Africa and rural Ghanaians—but are rapidly replaced by “BloSSUM” taxa like Bacteroides and Bifidobacterium in urban centers [6,21]. This shift is supported by experimental literature for the abandonment of fiber-rich traditional diets and increased antibiotic exposure, mirroring the ecological collapse observed in Western populations [17,21].
However, the African microbiome landscape reveals critical regional nuances that complicate the binary simplicity of the global model. Unlike the uniform “high diversity in rural vs. low diversity in urban” pattern often cited in global literature, specific African populations defy this trend. Notably, the nomadic Fulani of Nigeria exhibit significantly lower microbial richness compared to their semi-urban neighbors, a deviation driven by a specialized, restrictive dairy-centric diet and pathogen exposure rather than industrialization [7,22]. Furthermore, recent metagenomic profiling suggests that Treponema extinction is not immediate in all urban African contexts; the taxon persists in densely populated informal settlements like Nairobi, potentially due to high rates of circular migration between rural and urban areas which may facilitate microbial re-seeding [17]. Despite these localized exceptions, the overarching evidence may contribute to the fact that urbanization in Africa is driving a rapid, identifiable extinction of ancestral heritage taxa, a loss that is statistically associated with the rising prevalence of Type 2 Diabetes across the continent [21].
4.2. Mechanisms of Metabolic Dysfunction
Based on the synthesized mechanism papers, this taxonomic shift from a rural profile enriched with Treponema and Prevotella to an urban profile dominated by Bacteroides likely results in a “double hit” to metabolic health: a reduction in glucose-stimulated insulin secretion and a concurrent rise in systemic insulin resistance.
First, the depletion of Treponema and Prevotella critically diminishes the gut’s capacity to produce specific short-chain fatty acids (SCFAs) that regulate incretin hormones. Prevotella species and Treponema succinifaciens are specialized carbohydrate fermenters that produce significant quantities of succinate and acetate, which are key precursors for the production of propionate through microbial cross-feeding [28,29]. These metabolites function as ligands for the G-protein coupled receptors FFAR2 (GPR43) and FFAR3 (GPR41) located on intestinal L-cells; activation of these receptors directly stimulates the secretion of Glucagon-like peptide-1 (GLP-1) and Peptide YY (PYY) [30,31]. Because GLP-1 is essential for enhancing glucose-dependent insulin secretion from pancreatic β-cells, the loss of these “VANISH” taxa is consistent with a blunted incretin response and impaired glycemic control [31,32].
Simultaneously, the urban-associated rise in Bacteroides and Enterobacteriaceae is actively associated with metabolic endotoxemia. While Bacteroides are metabolically flexible, in the context of a low-fiber urban diet, they often degrade host mucins to survive, thereby thinning the protective colonic mucus layer and compromising intestinal barrier integrity [33,34]. This “leaky gut” facilitates the translocation of Lipopolysaccharide (LPS), a potent endotoxin found in the outer membranes of the proliferating Enterobacteriaceae (e.g., Klebsiella and Escherichia), into the systemic circulation [35,36]. Elevated circulating LPS binds to Toll-like receptor 4 (TLR4) on immune and metabolic cells, triggering the NF-κB inflammatory pathway and releasing cytokines such as TNF-α and IL-6, which directly inhibit insulin receptor signaling and cause systemic insulin resistance [33,37].
Critically, this suggests that the metabolic risk in urbanizing Africa is not merely acute but cumulative. If these microbial shifts occur early in life during the rural-to-urban transition, the chronic loss of SCFA-mediated metabolic buffering may render the next generation of urban Africans refractory to standard dietary interventions, accelerating the onset of T2D in progressively younger cohorts.
4.3. The Ecological Mismatch: Commercial Probiotics vs. Indigenous Therapeutics
These findings highlight a critical flaw in current therapeutic approaches: a fundamental “ecological mismatch” between Western interventions and African biology. Our results confirm that Bifidobacterium is a “BloSSUM” taxon already enriched in urban African populations due to dietary shifts [38]. Consequently, administering standard commercial probiotics, which typically rely on Bifidobacterium strains, is ecologically redundant—akin to watering a flooded garden.
In contrast, indigenous fermented foods like Ogi (cereal gruel) and Kunu (millet beverage) offer a superior, ecologically aligned intervention. Unlike monoculture pills, these foods serve as reservoirs for a complex consortium of Lactobacillus (reclassified as Limosilactobacillus, Lactiplantibacillus), Leuconostoc, Pediococcus, and Weissella species [38,39]. When consumed, these microorganisms drive lactic acid fermentation, producing organic acids and bacteriocins that lower luminal pH and inhibit pathogens [42,43]. Crucially, strains such as Lactobacillus plantarum, ubiquitous in Ogi, have been shown to upregulate tight junction proteins (e.g., zonula occludens-1), directly counteracting the urban-associated ‘leaky gut’ and reducing the translocation of inflammatory LPS [55].
Historically, fermentation in Africa functioned as an evolutionary “predigestion” strategy, enabling early humans to extract calories and nutrients from chemically defended plant resources, a process theorized to have supported human brain expansion [38]. This survival mechanism evolved into deep cultural traditions where specific foods are mandated for life stages; for instance, cereal-based gruels like Ogi in Nigeria and Uji in Kenya are universally established as the primary weaning foods for infants and the preferred sustenance for the elderly and convalescents due to their high digestibility and nutrient density [39,41,47,48]. In the domain of ethnomedicine and maternal care, the Ethiopian beverages Borde and Shameta, alongside the Nigerian drink Kunu, are traditionally prescribed to lactating women to stimulate milk production and restore strength postpartum [40,49,50]. Furthermore, specific fermented products serve distinct therapeutic roles: the raw liquor of Ogi is administered to treat diarrhea, fermented milk Nunu is consumed to induce relaxation and manage hypertension, and Ugba is applied topically to treat wounds [40,49,51]. Beyond health, these foods anchor social cohesion; Mursik (smoke-flavored fermented milk) is obligatory at Kalenjin weddings and athletic celebrations in Kenya, while alcoholic beverages like Pito and Burukutu facilitate conflict resolution and communal labor in West Africa [48,49,50].
We propose that this fermentation-friendly, acidic niche functions as a form of “ecological engineering,” mimicking the ancestral gut environment necessary to support the re-establishment of missing rural taxa like Treponema. Thus, Ogi and Kunu function not merely as nutritional supplements, but as ecological drivers capable of restoring the physiological conditions required to recover African indigenous biodiversity (Figure 3).
4.4. Environmental and Socioeconomic Confounders
Beyond diet and antibiotic exposure, urbanization introduces complex environmental and socioeconomic shifts that independently influence the microbiome-diabetes axis. Improvements in sanitation (WASH), while critical for hygiene, physically isolate individuals from environmental microbial reservoirs and soil-transmitted helminths; the loss of these immunomodulatory parasites is correlated with reduced gut diversity and increased T2D risk [14,44,52]. Socioeconomic status (SES) also drives variation distinct from nutrition; in Tanzania, housing quality (e.g., flooring material) was a stronger predictor of microbiota composition than dietary intake [19], while household crowding in Indigenous populations has been linked to maintained species richness via horizontal transmission [34]. Furthermore, emerging evidence suggests that urban pollutants, including PM2.5 and chemical toxins, deplete beneficial taxa and elevate pro-inflammatory biomarkers associated with insulin resistance [53]. These environmental pressures ultimately interact with host genetics, where ethnicity-specific metabolic phenotypes may modulate susceptibility to dysbiosis [11,54].
4.5. Limitations
The interpretation of these findings requires caution due to several methodological and demographic limitations. A primary constraint is the cross-sectional design of most included studies, which captures only a snapshot of the microbiome. While this allows for the identification of associations, it precludes the determination of causality or the observation of the temporal sequence of “VANISH” taxon loss during the rural-to-urban transition.
Geographically, the data is heavily skewed toward West and South Africa (e.g., Ghana, Nigeria, South Africa), with sparser representation from Central or East Africa. Given the vast ethnic, genetic, and dietary diversity across the Sub-Saharan region, findings derived from these specific hubs may not be fully generalizable to populations with distinct agro-ecological practices in other parts of the continent.
Furthermore, complex socioeconomic confounders remain largely unaddressed. Urbanization is a multifaceted transition involving not only dietary changes but also shifts in socioeconomic status (SES), sanitation access, and healthcare utilization. Most reviewed studies did not control for household income or antibiotic accessibility, variables known to independently influence both microbiome assembly and metabolic risk.
Finally, technical limitations exist regarding taxonomic resolution and dietary data. The reliance on 16S rRNA sequencing in the majority of studies [19,26] limits functional insight compared to metagenomics [24,44]. Additionally, very few studies explicitly tracked dietary intake rigorously; while some adjusted for broad dietary patterns, others failed to capture specific micronutrient or fiber intake data [18,21,45]. To advance the field, future research must prioritize longitudinal clinical trials that control for these SES confounders and specifically test the efficacy of Ogi and Kunu in restoring gut diversity.
5. Conclusion and Future Directions
5.1. Conclusion and Public Health Implications
This systematic review provides the first comprehensive synthesis of the “invisible extinction” currently unfolding within the African gut microbiome. The evidence from N=6,097 participants across 22 countries reveals a consistent, urbanization-induced ecological collapse: the depletion of ancestral, fiber-degrading specialists (Treponema, Prevotella) and the compensatory bloom of mucin-degrading generalists (Bacteroides). This taxonomic shift is not merely an ecological curiosity but a critical driver of metabolic dysfunction, statistically linked to the rising incidence of Type 2 Diabetes in lean African populations. The loss of SCFA-producing “VANISH” taxa, coupled with the rise of endotoxemia-driving Enterobacteriaceae, creates a mechanistic predisposition to hyperglycemia.
Critically, these findings challenge the utility of importing Western dietary guidelines. Public health policy in Sub-Saharan Africa must pivot from generic “low-calorie” recommendations to specific “microbiome-preservation” strategies. National health agendas should explicitly advocate for the protection of heritage diets and the integration of indigenous fermented foods like Ogi and Kunu into standard nutritional guidelines, framing them not as “alternative” options but as essential, culturally accessible tools for metabolic resilience.
5.2. Recommendations for Future Research and Capacity Building
To move beyond association and establish causality, the field must pivot toward four strategic priorities:
- Longitudinal Migration Cohorts: Most current data is cross-sectional. Future studies should follow rural-to-urban migrants in real-time (e.g., over 12–24 months) to pinpoint exactly when the extinction of Treponema occurs and identify the specific dietary tipping points that trigger this loss.
- From 16S to Metagenomics: Given that 75% of reviewed studies relied on 16S rRNA sequencing, our functional understanding is limited. Future efforts must prioritize deep shotgun metagenomics to resolve strain-level diversity and map the specific gene clusters (e.g., CAZymes) responsible for fiber degradation.
- Clinical Trials of Indigenous Therapeutics: We urgently call for Randomized Controlled Trials (RCTs) to evaluate the efficacy of Ogi and Kunu in pre-diabetic urban Africans. Specifically, trials should assess whether daily supplementation can re-seed “VANISH” taxa, reduce systemic endotoxemia (LPS), and improve insulin sensitivity.
- Capacity Building and Funding: The current reliance on Western-led studies creates a persistent data bias. Addressing this gap requires a structural shift in global science funding. We call for equitable partnership models that prioritize direct investment in African-led research consortia and local sequencing infrastructure. Strengthening intra-African collaboration is essential to ensure that the future of African precision medicine is defined by African researchers.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Funding
No funds, grants, or other support were received.
Conflicts of Interest
The authors have no conflicts of interest to declare that are relevant to the content of this article.
References
- Soh, R., Ngaha Damndja, W., & Njintang Yanou, N. (2024). Functional foods of sub-Saharan Africa and their implications in the management of type 2 diabetes: A review. Food Sci Nutr, 12(1), 24–34. [CrossRef]
- Ekpor, E., Akyirem, S., & Adade Duodu, P. (2024). Prevalence of metabolic dysfunction-associated fatty liver disease and its association with glycemic control in persons with type 2 diabetes in Africa: A systematic review and meta-analysis. PLOS Glob Public Health, 4(5), e0002835. [CrossRef]
- Namazi, N., Moghaddam, S. S., Esmaeili, S., Peimani, M., Tehrani, Y. S., Bandarian, F., Shobeiri, P., Nasli-Esfahani, E., Malekpour, M. R., Rezaei, N., Rezaei, N., Arjmand, B., Larijani, B., & Farzadfar, F. (2024). Burden of type 2 diabetes mellitus and its risk factors in North Africa and the Middle East, 1990-2019: findings from the Global Burden of Disease study 2019. BMC Public Health, 24(1), 98. [CrossRef]
- Olamoyegun, M. A., Alare, K., Afolabi, S. A., Aderinto, N., & Adeyemi, T. (2024). A systematic review and meta-analysis of the prevalence and risk factors of type 2 diabetes mellitus in Nigeria. Clinical Diabetes and Endocrinology, 10(1). [CrossRef]
- Magodoro, I. M., Castle, A. C., Tshuma, N., Goedecke, J. H., Sewpaul, R., Manasa, J., Manne-Goehler, J., Ntusi, N. A., Nyirenda, M. J., & Siedner, M. J. (2024). Associations of HIV and prevalent type 2 diabetes mellitus in the context of obesity in South Africa. J Multimorb Comorb, 14, 26335565241293691. [CrossRef]
- Ramaboli, M. C., Ocvirk, S., Khan Mirzaei, M., Eberhart, B. L., Valdivia-Garcia, M., Metwaly, A., Neuhaus, K., Barker, G., Ru, J., Nesengani, L. T., Mahdi-Joest, D., Wilson, A. S., Joni, S. K., Layman, D. C., Zheng, J., Mandal, R., Chen, Q., Perez, M. R., Fortuin, S.,…O’Keefe, S. J. D. (2024). Diet changes due to urbanization in South Africa are linked to microbiome and metabolome signatures of Westernization and colorectal cancer. Nat Commun, 15(1), 3379. [CrossRef]
- Daunizeau, C., Franck, M., Boutin, A., Ruel, M., Poliakova, N., Ayotte, P., & Bélanger, R. (2025). The gut microbiota of Indigenous populations in the context of dietary westernization: a systematic review and meta-analysis. Frontiers in Nutrition, 12. [CrossRef]
- Davidson, P. (2025). Gut Microbiota, Migration, and Heritage. In Routledge Handbook of Food and Cultural Heritage (pp. 387–398). [CrossRef]
- Harrison, L. C., Allnutt, T. R., Hanieh, S., Roth-Schulze, A. J., Ngui, K. M., Stone, N. L., Bandala-Sanchez, E., Backshell, L., Gurruwiwi, G., Gondarra, V., Couper, J. J., Craig, M. E., Davis, E. A., Huynh, T., Soldatos, G., Wentworth, J. M., Vuillermin, P., Penno, M. A. S., Biggs, B. A., & Consortium, E. S. G. (2025). Indigenous infants in remote Australia retain an ancestral gut microbiome despite encroaching Westernization. Nat Commun, 16(1), 9904. [CrossRef]
- Alencar, R. M., Martinez, J. G., Machado, V. N., Alzate, J. F., Ortiz-Ojeda, C. P., Matias, R. R., Benzaquem, D. C., Santos, M. C. F., Assuncao, E. N., Lira, E. C., Astolfi-Filho, S., Hrbek, T., Farias, I. P., & Fantin, C. (2024). Preliminary profile of the gut microbiota from amerindians in the Brazilian amazon experiencing a process of transition to urbanization. Braz J Microbiol, 55(3), 2345–2354. [CrossRef]
- Moyo, G. T., Tepekule, B., Katsidzira, L., Blaser, M. J., & Metcalf, C. J. E. (2025). Getting ahead of human-associated microbial decline in Africa: the urgency of sampling in light of epidemiological transition. Trends Microbiol, 33(11), 1173–1184. [CrossRef]
- Palladino, G., Marangi, M., Scicchitano, D., Turroni, S., Rampelli, S., & Candela, M. (2025). Gut microbiome structure in asylum seekers newly arrived in Italy from Africa. Sci Rep, 15(1), 40596. [CrossRef]
- Zhang, C., Yu, Y., Yue, L., Chen, Y., Chen, Y., Liu, Y., Guo, C., Su, Q., & Xiang, Z. (2025). Gut microbiota profiles of sympatric snub-nosed monkeys and macaques in Qinghai-Tibetan Plateau show influence of phylogeny over diet. Commun Biol, 8(1), 95. [CrossRef]
- Blaser, M. J., & Dominguez-Bello, M. G. (2025). The Invisible Extinction. Annu Rev Microbiol, 79(1), 311–334. [CrossRef]
- Zhang, G., & Le Souef, P. (2024). The influence of modern living conditions on the human microbiome and potential therapeutic opportunities for allergy prevention. World Allergy Organ J, 17(1), 100857. [CrossRef]
- Vinogradova, E., Nurislam, M., Madiyar, N., Zharkyn, J., Rakhilya, A., Aliya, S., Makhabbat, B., Samat, K., & Almagul, K. (2024). Impact of urbanization on gut microbiome mosaics across geographic and dietary contexts. mSystems, 9(10), 00585–00524.
- Maghini, D. G., Oduaran, O. H., Olubayo, L. A. I., Cook, J. A., Smyth, N., Mathema, T., Belger, C. W., Agongo, G., Boua, P. R., Choma, S. S. R., Gomez-Olive, F. X., Kisiangani, I., Mashaba, G. R., Micklesfield, L., Mohamed, S. F., Nonterah, E. A., Norris, S., Sorgho, H., Tollman, S.,…Hazelhurst, S. (2025). Expanding the human gut microbiome atlas of Africa. Nature, 638(8051), 718–728. [CrossRef]
- Angelakis, E., Bachar, D., Yasir, M., Musso, D., Djossou, F., Gaborit, B., Brah, S., Diallo, A., Ndombe, G. M., Mediannikov, O., Robert, C., Azhar, E. I., Bibi, F., Nsana, N. S., Parra, H. J., Akiana, J., Sokhna, C., Davoust, B., Dutour, A., & Raoult, D. (2019). Treponema species enrich the gut microbiota of traditional rural populations but are absent from urban individuals. New Microbes New Infect, 27, 14–21. [CrossRef]
- Pyuza, J. J., van Dorst, M., Barnett, D., Stam, K., Manurung, M., Wammes, L., Konig, M., Kruize, Y., Andongolile, N., Ngowi, A., Shao, E. R., Kullaya, V. I., Mremi, A., Hogendoorn, P. C. W., Msuya, S. E., Jochems, S. P., Penders, J., Yazdanbakhsh, M., & de Steenhuijsen Piters, W. A. A. (2025). Tanzanian gut microbiota profiles linked to high but rapidly waning yellow fever antibody titers. NPJ Biofilms Microbiomes, 11(1), 110. [CrossRef]
- Truter, M., Koopman, J. E., Jordaan, K., Tsamkxao, L. O., Cowan, D. A., Underdown, S. J., Ramond, J. B., & Rifkin, R. F. (2024). Documenting the diversity of the Namibian Ju|’hoansi intestinal microbiome. Cell Rep, 43(2), 113690. [CrossRef]
- Verhaar, B. J. H., van der Linden, E. L., Hayfron-Benjamin, C. F., Owusu-Dabo, E., Darko, S. N., Twumasi-Ankrah, S., Henneman, P., Beune, E., Meeks, K. A. C., Nieuwdorp, M., Herrema, H., van den Born, B. H., & Agyemang, C. (2025). Gut microbiota shift in Ghanaian individuals along the migration axis: the RODAM-Pros cohort. Gut Microbes, 17(1), 2471960. [CrossRef]
- Afolayan, A. O., Ayeni, F. A., Moissl-Eichinger, C., Gorkiewicz, G., Halwachs, B., & Högenauer, C. (2019). Impact of a nomadic pastoral lifestyle on the gut microbiome in the Fulani living in Nigeria. Frontiers in Microbiology, 10, 2138.
- De Filippo, C., Di Paola, M., Ramazzotti, M., Albanese, D., Pieraccini, G., Banci, E., Miglietta, F., Cavalieri, D., & Lionetti, P. (2017). Diet, Environments, and Gut Microbiota. A Preliminary Investigation in Children Living in Rural and Urban Burkina Faso and Italy. Front Microbiol, 8, 1979. [CrossRef]
- Rampelli, S., Gallois, S., D’Amico, F., Turroni, S., Fabbrini, M., Scicchitano, D., Candela, M., & Henry, A. (2024). The gut microbiome of Baka forager-horticulturalists from Cameroon is optimized for wild plant foods. iScience, 27(3), 109211. [CrossRef]
- Ecklu-Mensah, G., Choo-Kang, C., Maseng, M. G., Donato, S., Bovet, P., Viswanathan, B., Bedu-Addo, K., Plange-Rhule, J., Oti Boateng, P., Forrester, T. E., Williams, M., Lambert, E. V., Rae, D., Sinyanya, N., Luke, A., Layden, B. T., O’Keefe, S., Gilbert, J. A., & Dugas, L. R. (2023). Gut microbiota and fecal short chain fatty acids differ with adiposity and country of origin: the METS-microbiome study. Nat Commun, 14(1), 5160. [CrossRef]
- Oduaran, O. H., Tamburini, F. B., Sahibdeen, V., Brewster, R., Gomez-Olive, F. X., Kahn, K., Norris, S. A., Tollman, S. M., Twine, R., Wade, A. N., Wagner, R. G., Lombard, Z., Bhatt, A. S., & Hazelhurst, S. (2020). Gut microbiome profiling of a rural and urban South African cohort reveals biomarkers of a population in lifestyle transition. BMC Microbiol, 20(1), 330. [CrossRef]
- Gomollon, F., Dignass, A., Annese, V., Tilg, H., Van Assche, G., Lindsay, J. O., Peyrin-Biroulet, L., Cullen, G. J., Daperno, M., Kucharzik, T., Rieder, F., Almer, S., Armuzzi, A., Harbord, M., Langhorst, J., Sans, M., Chowers, Y., Fiorino, G., Juillerat, P.,…Ecco. (2017). 3rd European Evidence-based Consensus on the Diagnosis and Management of Crohn’s Disease 2016: Part 1: Diagnosis and Medical Management. J Crohns Colitis, 11(1), 3–25. [CrossRef]
- Adamberg, S., & Adamberg, K. (2024). Prevotella enterotype associates with diets supporting acidic faecal pH and production of propionic acid by microbiota. Heliyon, 10(9), e31134.
- Tannock, G. W. (2023). Understanding the gut microbiota by considering human evolution: A story of fire, cereals, cooking, molecular genetics, and possibly the “old friends” hypothesis. Microbiology and Molecular Biology Reviews, 87(4), e00044-23.
- Christiansen, C. B., Gabe, M. B. N., Svendsen, B., Dragsted, L. O., Rosenkilde, M. M., & Holst, J. J. (2018). The impact of short-chain fatty acids on GLP-1 and PYY secretion from the isolated perfused rat colon. American Journal of Physiology-Gastrointestinal and Liver Physiology, 315(1), G53–G65.
- Zeng, Y., Wu, Y., Zhang, Q., & Xiao, X. (2023). Crosstalk between glucagon-like peptide 1 and gut microbiota in metabolic diseases. mBio, 15, e02032-23.
- Arukha, A. P., Nayak, S., & Swain, D. M. (2025). Effect of Akkermansia muciniphila on GLP-1 and insulin secretion. Nutrients, 17(2516).
- Maciejewska-Markiewicz, D., Prowans, P., & Stachowska, E. (2024). Impact of intestinal barrier dysfunction and sodium butyrate on type 2 diabetes mellitus: A narrative review. Nutrients, 17(1753).
- Silk, K., Neth, B., & Harrison, U. (2024). Advancing microbiome research in Māori populations: insights from recent literature exploring the gut–microbiome–immune axis in metabolic health. GeroScience, 47, 1–22.
- Di Vincenzo, F., Del Gaudio, A., Petito, V., Lopetuso, L. R., & Scaldaferri, F. (2024). Gut microbiota, intestinal permeability, and systemic inflammation: a narrative review. Internal and Emergency Medicine, 19, 275–293.
- Hecht, A. L., Harling, L. C., Friedman, E. S., Tanes, C., Lee, J., Firrman, J., Hao, F., Tu, V., Liu, L., Patterson, A. D., Bittinger, K., Goulian, M., & Wu, G. D. (2024). Dietary carbohydrates regulate intestinal colonization and dissemination of Klebsiella pneumoniae. The Journal of Clinical Investigation, 134(9), e174726.
- Xia, T., Hu, Y., Xu, H., et al. (2024). Achyranthes bidentata polysaccharide improves type 2 diabetes mellitus by regulating the gut microbiota/SCFAs/GLP-1/cAMP signaling pathway. International Journal of Biological Macromolecules, 270, 132256.
- José, G. P. M., Juliano, D. D. L., Gilberto, V. d. M. P., & Ramesh, C. R. (2024). Trending Topics on Fermented Foods. Springer. [CrossRef]
- Adesemoye, E. T., Sanni, A. I., Spano, G., Capozzi, V., & Fragasso, M. (2025). Lactic Acid Bacteria Diversity in Fermented Foods as Potential Bio-Resources Contributing to Alleviate Malnutrition in Developing Countries: Nigeria as a Case Study. Fermentation, 11(2). [CrossRef]
- Obafemi, Y. D., Ajayi, A. A., Adebayo, H. A., Oyewole, O. A., & Olumuyiwa, E. O. (2024). The role of indigenous Nigerian fermented agrifoods in enhancing good health and well˗being. Discover Food, 4(1). [CrossRef]
- Houngbedji, M., Jespersen, J. S., Wilfrid Padonou, S., & Jespersen, L. (2025). Cereal-based fermented foods as microbiota-directed products for improved child nutrition and health in sub-Saharan Africa. Crit Rev Food Sci Nutr, 65(18), 3422–3443. [CrossRef]
- Lys, I. M. (2024). The Role of Lactic Fermentation in Ensuring the Safety and Extending the Shelf Life of African Indigenous Vegetables and Its Economic Potential. Applied Research, 4(1). [CrossRef]
- Ndudi, W., Edo, G. I., Samuel, P. O., Jikah, A. N., Opiti, R. A., Ainyanbhor, I. E., Essaghah, A. E. A., Ekokotu, H. A., Oghroro, E. A. E., & Agbo, J. J. (2024). Traditional fermented foods of Nigeria: microbiological safety and health benefits. Journal of Food Measurement and Characterization, 18(6), 4246–4271. [CrossRef]
- Ngwese, M. M., Adegbite, B. R., Zinsou, J. F., Fitzstevens, J. L., Schmidt, V. T., Moure, P. A. N., Maloum, M. N., Tyakht, A. V., Huus, K. E., Youngblut, N. D., Kremsner, P. G., Adegnika, A. A., & Ley, R. E. (2025). Infection with gut parasites correlates with gut microbiome diversity across human populations in Africa. Gut Microbes, 17(1), 2587966. [CrossRef]
- Hodges, P., Mweetwa, M., Kayamba, V., O’Keefe, S., Ocvirk, S., Neuhaus, K., Kelly, P. (2025). P1321 Inflammatory bowel disease and the rural to urban microbiome transition in Zambia. J Crohns Colitis, 19(1), i2382–i2384. [CrossRef]
- Page MJ, McKenzie JE, Bossuyt PM, Boutron I, Hoffmann TC, Mulrow CD, et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ. 2021;372:n71. [CrossRef]
- Banwo, K., Adebo, O. A., & Falade, T. D. O. (2024). Examples of lactic-fermented foods of the African continent. In G. Vinderola, A. C. Ouwehand, S. Salminen, & A. von Wright (Eds.), Lactic acid bacteria: Microbiological and functional aspects (pp. 312–327). CRC Press.
- Khayeka-Wandabwa, C., Choge, J. K., Linnemann, A. R., & Schoustra, S. (2024). Linking fermented foods to microbial composition and valorisation: Blueprint for Kenya. Food Reviews International, 40(10), 3424–3444. [CrossRef]
- Adesulu-Dahunsi, A. T., Adesulu, A. O., Fagbemi, A. S., & Ezeamagu, C. O. (2025). African cereal based fermented beverages as functional drinks with potential impact on human gut microbiota and health implications. Discover Food, 5, 106. [CrossRef]
- Hawaz, H., Bottari, B., Scazzina, F., & Carini, E. (2025). Eastern African traditional fermented foods and beverages: Advancements, challenges, and perspectives on food technology, nutrition, and safety. Comprehensive Reviews in Food Science and Food Safety, 24(2), e70137. [CrossRef]
- Ashaolu, T. J. (2024). Fermented fruits and legumes in Africa: Production factors and health benefits. International Journal of Food Science and Technology, 59(10), 6866–6874. [CrossRef]
- Zhao, F., Ding, Y., Mozaffarian, D., & Zuo, T. (2025). Dietary urbanization and the human gut microbiome in the pathogenesis of non-communicable diseases. Cell Metabolism, 37. [CrossRef]
- Nguyen, H. K., & Isaksson, C. (2024). Exploring vertebrate gut microbial diversity in the urbanization era. Microbiota and Host, 2024, e240003. [CrossRef]
- Gong, J. Y., Sajjadi, S. F., Motala, A. A., Shaw, J. E., & Magliano, D. J. (2025). Variation in type 2 diabetes prevalence across different populations: The key drivers. Diabetologia, 68, 2327–2339. [CrossRef]
- Wang, J., Ji, H., Wang, S., Liu, H., Zhang, W., Zhang, D., & Wang, Y. (2018). Probiotic Lactobacillus plantarum promotes intestinal barrier function by strengthening the epithelium and modulating gut microbiota. Frontiers in Microbiology, 9, 1953. [CrossRef]
Figure 2.
Ecological transition of the gut microbiome in Sub-Saharan Africa. The schematic illustrates the fundamental shift from a high-diversity “rural” microbiome (left), characterized by the abundance of fiber-degrading Prevotella and Treponema, to a low-diversity “urban” microbiome (right) dominated by Bacteroides and Enterobacteriaceae. This “Westernization” effect is driven by the reduction of microbiota-accessible carbohydrates (MACs) and increased antibiotic exposure in urban centers.Parts of this figure were drawn using pictures from Servier Medical Art. Servier Medical Art by Servier is licensed under a Creative Commons Attribution 3.0 Unported License (https://creativecommons.org/licenses/by/3.0/.
Figure 2.
Ecological transition of the gut microbiome in Sub-Saharan Africa. The schematic illustrates the fundamental shift from a high-diversity “rural” microbiome (left), characterized by the abundance of fiber-degrading Prevotella and Treponema, to a low-diversity “urban” microbiome (right) dominated by Bacteroides and Enterobacteriaceae. This “Westernization” effect is driven by the reduction of microbiota-accessible carbohydrates (MACs) and increased antibiotic exposure in urban centers.Parts of this figure were drawn using pictures from Servier Medical Art. Servier Medical Art by Servier is licensed under a Creative Commons Attribution 3.0 Unported License (https://creativecommons.org/licenses/by/3.0/.
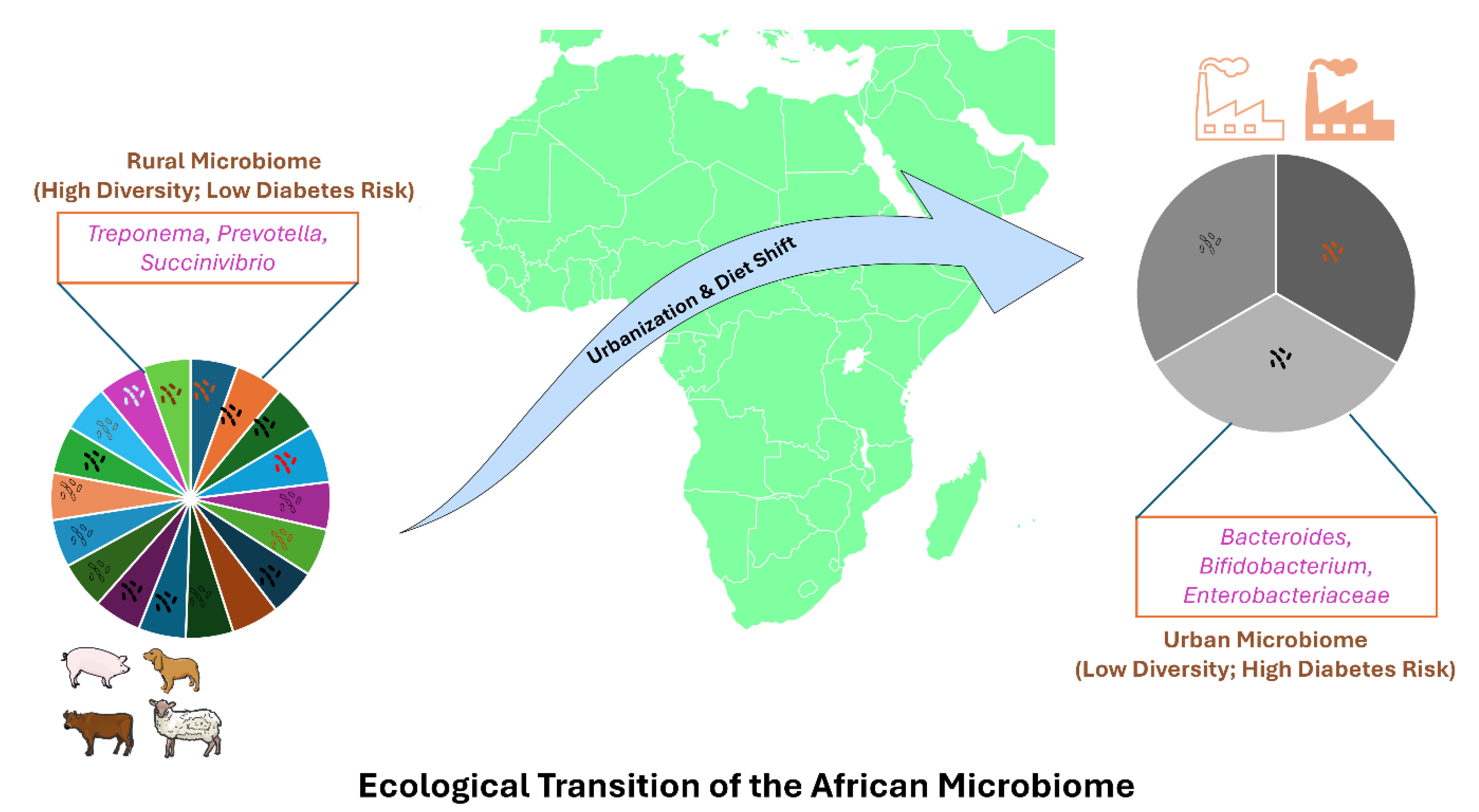
Figure 3.
Therapeutic potential of African indigenous fermented foods.
Traditional fermentation serves as a biotechnology to restore gut health. Foods such as Ogi (cereal gruel), Kunu (millet beverage), and Nunu (fermented milk) act as reservoirs for beneficial lactic acid bacteria (L. plantarum, L. fermentum).
Parts of this figure were drawn using pictures from Servier Medical Art. Servier Medical Art by Servier is licensed under a Creative Commons Attribution 3.0 Unported License (https://creativecommons.org/licenses/by/3.0/).
Figure 3.
Therapeutic potential of African indigenous fermented foods.
Traditional fermentation serves as a biotechnology to restore gut health. Foods such as Ogi (cereal gruel), Kunu (millet beverage), and Nunu (fermented milk) act as reservoirs for beneficial lactic acid bacteria (L. plantarum, L. fermentum).
Parts of this figure were drawn using pictures from Servier Medical Art. Servier Medical Art by Servier is licensed under a Creative Commons Attribution 3.0 Unported License (https://creativecommons.org/licenses/by/3.0/).
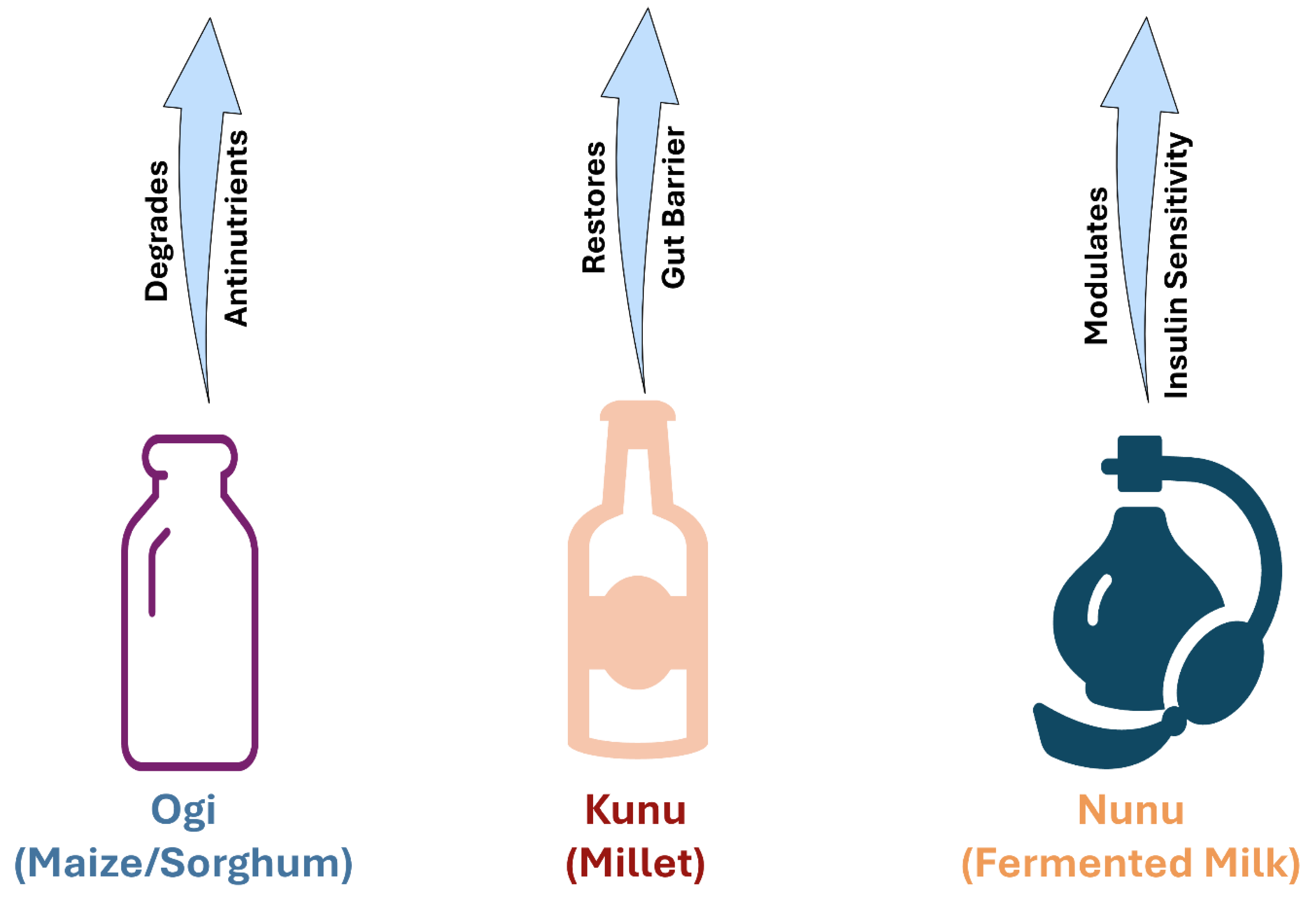

Table 1.
Summary Table of Study Characteristics.
| Characteristic | Count/Details |
| Total Studies | 12 |
| Total Participants | 6,097 |
| Countries Represented | 22 (Algeria, Burkina Faso, Cameroon, Congo, France, French Guiana, French Polynesia, Gabon, Ghana, Italy, Jamaica, Kenya, Namibia, Netherlands, Nigeria, Saudi Arabia, Senegal, Seychelles, South Africa, Tanzania, USA, Zambia) |
| Sequencing Method |
16S rRNA (9 studies): Afolayan, Angelakis, De Filippo, Ecklu-Mensah, Hodges, Oduaran, Pyuza, Truter, Verhaar. Shotgun Metagenomics (3 studies): Maghini, Ngwese, Rampelli. |
Table 2.
Summary of significant taxonomic alterations reported in included studies. Taxa listed were found to be significantly differentially abundant ($p < 0.05$) between rural/traditional and urban/industrialized cohorts.
Table 2.
Summary of significant taxonomic alterations reported in included studies. Taxa listed were found to be significantly differentially abundant ($p < 0.05$) between rural/traditional and urban/industrialized cohorts.
| Study (Author, Year) | Country | Comparison | Rural / Traditional Enriched Taxa (“VANISH” / Old Friends) | Urban / Industrialized Enriched Taxa (“BloSSUM” / Pathobionts) |
| Afolayan et al. (2019) | Nigeria | Nomadic Fulani vs. Semi-Urban Jarawa | Prevotella | Bacteroides |
| Angelakis et al. (2019) | 10 Countries* | Rural vs. Urban (Multi-country) | Treponema, Prevotella | Bifidobacterium, Bacteroides, Streptococcus |
| De Filippo et al. (2017) | Burkina Faso | Rural Mossi vs. Urban/Italian | Prevotella, Treponema, Xylanibacter | Bacteroides, Enterobacteriaceae |
| Ecklu-Mensah et al. (2023) | Ghana, SA | Rural vs. Urban vs. US/UK | Prevotella (Enterotype 2) | Bacteroides (Enterotype 1), Lachnospira |
| Hodges et al. (2025) | Zambia | Rural vs. Urban | Prevotella, High Alpha Diversity | Bacteroides, Low Alpha Diversity |
| Maghini et al. (2025) | South Africa | Rural Bushbuckridge vs. Urban Soweto | Treponema succinifaciens | Bifidobacterium, Bacteroides |
| Ngwese et al. (2025) | Gabon | Rural vs. Urban | High Alpha Diversity, Prevotella | Low Alpha Diversity |
| Oduaran et al. (2020) | South Africa | Rural vs. Urban Women | Prevotella | Bifidobacterium, Nuella |
| Pyuza et al. (2025) | Tanzania | Rural vs. Urban Moshi | Treponema, Succinivibrio, Prevotella | Bacteroides, Bifidobacterium, Escherichia |
| Rampelli et al. (2024) | Cameroon | Hunter-Gatherer vs. Urban | Treponema, Succinivibrio | Bacteroides |
| Truter et al. (2024) | Namibia | San Hunter-Gatherers vs. Bantu | Treponema, Prevotella | Bacteroides |
| Verhaar et al. (2025) | Ghana | Rural vs. Urban vs. Migrants (AMS) | Treponema, Succinivibrio, Prevotella | Bacteroides, Blautia, Faecalibacterium |
Note: Angelakis et al. included populations from Senegal, Nigeria, Gabon, Kenya, Tanzania, and others.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



