Submitted:
22 January 2026
Posted:
26 January 2026
You are already at the latest version
Abstract
We have previously demonstrated that the endocannabinoid system is dysregulated at synaptic terminals in the cerebral cortex of aged rats, characterized by reduced availability of the neuroprotective endocannabinoid 2-arachidonoylglycerol (2-AG) as a result of im-paired metabolic enzyme activity. This deficit was only partially compensated by canna-binoid receptor (CBR) ligand binding. Given that Δ9-tetrahydrocannabinol (THC) func-tions as a CBR ligand, the present study was designed to determine whether a full-spectrum cannabis extract with high THC content, its THC-free fraction, or pure THC could modulate the age-related dysregulation of 2-AG. Synaptosomes isolated from the cerebral cortex of adult and aged rats were incubated with a full-spectrum extract, a THC-free fraction, or pure THC, together with the corre-sponding radiolabeled substrates to assess 2-AG-metabolizing enzyme activity. Our re-sults demonstrate that the age-related decline in 2-AG bioavailability (a) is exacerbated by either the THC-free fraction or pure THC, primarily due to a significant reduction in 2-AG synthesis, and (b) is partially attenuated through inhibition of 2-AG hydrolysis when the extract contains THC. Consequently, a high-THC full-spectrum extract regulates 2-AG metabolism more effectively than THC alone. These findings support the concept that cannabis phytochemicals act synergistically (the entourage effect) and highlight the therapeutic potential of high-THC extracts for restoring reduced 2-AG levels in the aging brain.
Keywords:
aging
; 2-arachidonoylglycerol metabolism
; cerebral cortex
; synaptosomes
; THC
1. Introduction
The endocannabinoid system (ECS) consists of the endogenous ligands, mainly anandamide (AEA) and 2-arachidonoylglycerol (2-AG) [1,2], the CB1 and CB2 membrane receptors (CB1 and CB2) [3], and the enzymes involved in AEA and 2-AG metabolism [4,5,6,7,8]. Diacylglycerol lipase (DAGL) and lysophosphatidate phosphohydrolase (LPAase) are the primary enzymes responsible for 2-AG synthesis, while monoacylglycerol lipase (MAGL) and serine hydrolase alpha-beta-hydrolase domain 6 (ABHD6) are the main enzymes mediating its hydrolysis at pre-synaptic and post-synaptic level, respectively [6,8,9].
2-AG is a crucial endocannabinoid implicated in various physiological and pathological processes, notably in neuroprotection against neurodegenerative disorders via activation of cannabinoid receptors (CBR) in neuronal synapses [10,11,12]. This endocannabinoid acts as a retrograde messenger regulating the release of various neurotransmitters, which underscores the importance of preserving its tone to prevent neurodegenerative processes. Therefore, the availability of 2-AG at the synaptic level represents a key aspect in the study of mechanisms underlying neurodegeneration. Our previous research demonstrated age-related alterations in the expression of proteins (receptors and enzymes), as well as in the enzymatic activities involved in the synthesis (DAGL) and hydrolysis of 2-AG (mainly MAGL), in synaptosomes (SYN) from aged rats. These findings also revealed a reduction in 2-AG levels from the cerebral cortex of aged rats [13], with significant modulation of 2-AG metabolism by CBR ligands [14].
Cannabis sp. (cannabis) contains a variety of metabolites, including terpenoids, sterols, and flavonoids [15], in addition to phytocannabinoids [16,17], as well as other compounds. This diversity suggests that the therapeutic effects of this plant may arise from the synergistic action of multiple compounds rather than a single active constituent [18,19,20]. In neurodegenerative conditions, cannabis-derived compounds have been shown to exert neuroprotective effects and are currently being evaluated in several clinical studies [21]. In the context of aging, the CBR agonist THC has been reported to improve spatial learning and enhance cognitive performance in aged mice [22]. Based on these findings, together with our previous results demonstrating the modulation of 2-AG metabolism by CB receptor ligands [14], we hypothesize that THC-containing cannabis extracts may enhance the metabolism of the neuroprotective endocannabinoid 2-AG, potentially contributing to the mechanism that underlie the cognitive improvements reported for THC. To corroborate this hypothesis, we conducted experiments to assess the enzymatic activities of DAGL, LPAase, and MAGL in SYN isolated from the CC of both adult and aged rats. The study involved treatment with a full-spectrum cannabis extract containing THC (full-spectrum extract), the THC-free fraction from this extract (THC-free extract), and pure THC (THC). By comparing the effects of these treatments on 2-AG metabolism, we aimed to determine whether THC alone or in combination with other cannabis metabolites could influence the enzymatic regulation of 2-AG, thereby contributing to its neuroprotective potential in the context of aging. The outcomes on 2-AG metabolism in the presence of THC alone or within the full-spectrum extract would help elucidate whether THC exerts a specific effect or if a synergistic interaction with other metabolites exists, potentially counteracting the age-related decline in 2-AG levels [13].
2. Results
2.1. Analysis of the Full-Spectrum Extract and the THC-Free Extract
Phytocannabinoid quantification of the full-spectrum extract by HPLC revealed a THC content of 37.73 mg/g inflorescence, with a minimum content of acidic THC (THCA) and CBN. No detectable amounts of CBD were observed (Limit of detection for CBD = 0.73 µg/mL, corresponding to 0.6 mg CBD/g inflorescence) (data not shown).
When the full-spectrum extract was chromatographed by preparative TLC, analysis of the developed plate allowed to identify a fraction that matched the pure THC standard (Rf = 0.65). This fraction (fraction 3) and the THC-free fractions (fraction 1 corresponding to the origin of the TLC plate; fraction 2 corresponding to the compounds that migrate above the origin but remained below the THC-enriched band and fraction 4 corresponding to the compounds that migrated beyond the THC-containing band) were then separately removed from the plate and analyzed by HPLC and GC-MS. These analyses confirmed the non-detectable levels of THC in the THC-free fractions, while the fraction that matched the pure THC standard contained mainly THC with a purity of 92 % (shown in Supplementary Material). As mentioned in Material and Methods section, THC-free fractions were combined for subsequent assays to investigate the effects of the THC-free extract.
2.2. Effect of the THC-Free Extract, the Full-Spectrum Extract, and THC on MAGL Activity
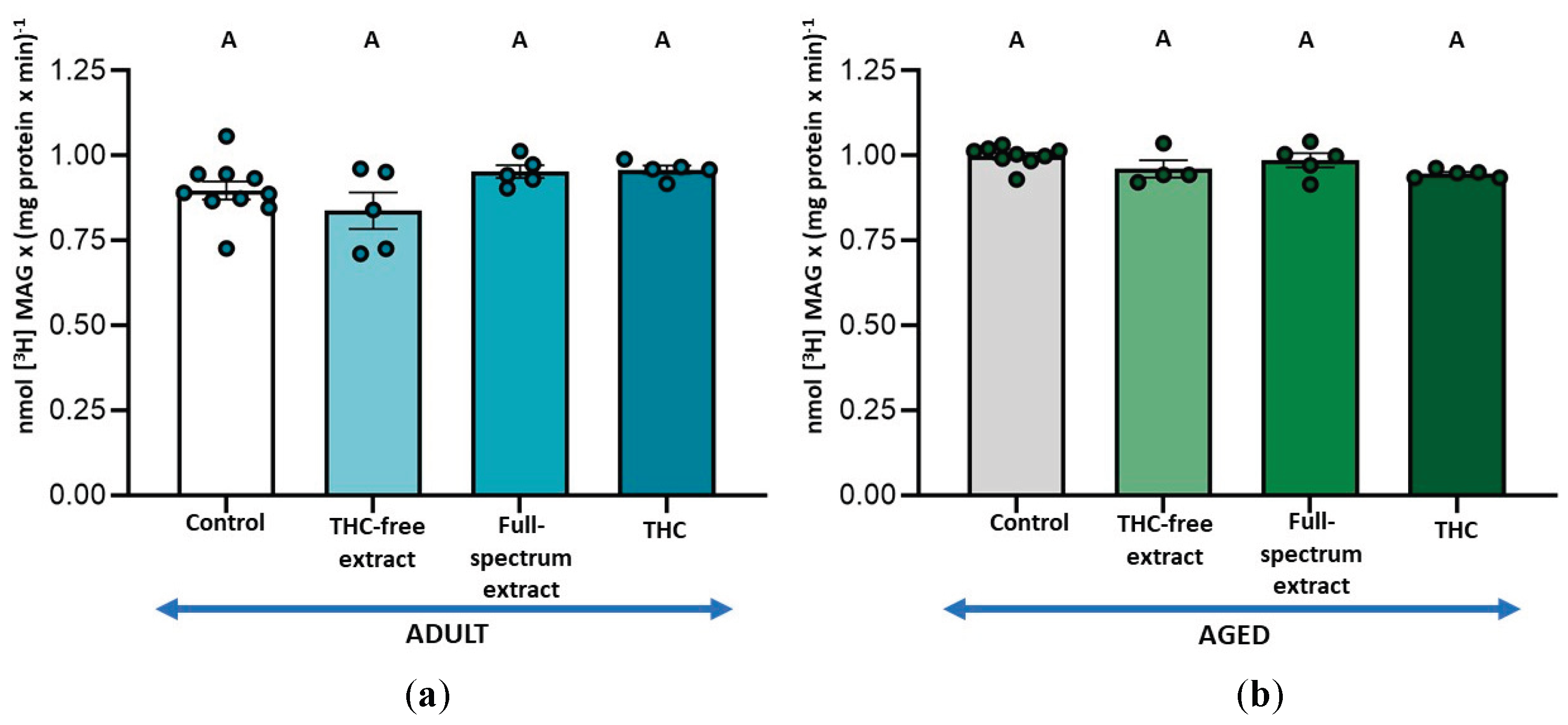
To determine the optimal concentration of THC in the full-spectrum extract that can affect 2-AG metabolism, we selected the measurement of MAGL activity, as it is the simplest assay involved in the availability of this endocannabinoid. The results demonstrated a 7 % inhibition of this activity only in aged SYN and with the amount of full-spectrum extract containing a THC concentration of 1 µM (Figure 1b). Accordingly, 1 µM THC was selected for the subsequent enzyme assays in both, the full-spectrum extract and the pure phytocannabinoid conditions. Additionally, when the effect of the THC-free extract was assayed, it was added in the same amount as the full-spectrum extract containing 1 µM THC; namely, the combined THC-free fractions derived from chromatographing an extract amount equivalent to that containing 1 µM THC were used. Thus, the THC-free extract conditions contained the same components in the same amounts as the 1 µM THC full-spectrum conditions, except for THC.
In adult SYN, no significant changes in MAGL activity were observed under any of the conditions assayed (Figure 2a). Conversely, in aged SYN, both the full-spectrum extract and the THC-free extract led to a similar reduction in MAGL activity, approximately 7.4 %. THC did not result in significant differences in MAGL activity compared to the control or to the different extracts (Figure 2b).
2.3. Effect of the THC-Free Extract, the Full-Spectrum Extract, and THC on DAGL Activity
Assessment of DAGL activity, an enzyme involved in 2-AG synthesis, revealed no significant effects of the treatments in adult SYN. In contrast, in aged SYN, the THC-free extract and the THC resulted in inhibition of DAGL activity by 84 % and 57 %, respectively, with no statistical differences between conditions (Figure 3). DAGL activity assays also revealed glycerol formation, demonstrating the coupling of MAGL activity with DAGL activity (Figure 4). In adult SYN, none of the treatments resulted in significant changes (Figure 4a). In contrast, in aged SYN, both the THC-free extract and THC decreased MAGL activity coupled to DAGL by 22 % and 33 %, respectively, although the difference between these two treatments was not statistically significant (Figure 4b).
2.4. Effect of the THC-Free Extract, the Full-Spectrum Extract, and THC on LPAase Activity
LPAase, another enzyme involved in 2-AG synthesis, was also evaluated (Figure 5). In this context, LPAase activity was unaffected by any of the treatments, both in adult (Figure 5a) and aged (Figure 5b) SYN.
Figure 5.
(a) Effect of the THC-free extract, the full-spectrum extract, and THC on lysophosphatidate phosphohydrolase (LPAase) activity in synaptosomes from adult rat cerebral cortex and; (b) aged rat cerebral cortex. The substrate [3H]LPA was added exogenously and the assay was conducted according to the conditions specified for LPAase activity as was described in the Materials and Methods section. Results are reported as nmol of MAG per mg of protein per minute and are shown as mean ± standard error of the mean (SEM), with individual data points representing the sample size of each group (n varies across columns). Data were analyzed by one-way ANOVA (a (overall model): F = 2.58, p = 0.0804; b (overall model): F = 2.67, p = 0.0769). Similar letters indicate no statistically significant differences between conditions (p ≥ 0.05). Graphic was performed using GraphPad Prism 9.5.1 (https://www.graphpad.com).
Figure 5.
(a) Effect of the THC-free extract, the full-spectrum extract, and THC on lysophosphatidate phosphohydrolase (LPAase) activity in synaptosomes from adult rat cerebral cortex and; (b) aged rat cerebral cortex. The substrate [3H]LPA was added exogenously and the assay was conducted according to the conditions specified for LPAase activity as was described in the Materials and Methods section. Results are reported as nmol of MAG per mg of protein per minute and are shown as mean ± standard error of the mean (SEM), with individual data points representing the sample size of each group (n varies across columns). Data were analyzed by one-way ANOVA (a (overall model): F = 2.58, p = 0.0804; b (overall model): F = 2.67, p = 0.0769). Similar letters indicate no statistically significant differences between conditions (p ≥ 0.05). Graphic was performed using GraphPad Prism 9.5.1 (https://www.graphpad.com).
2.5. Effect of the THC-Free Extract, the Full-Spectrum Extract, and THC on LPA-PLA1 Activity
Given that the LPAase substrate used in the assay was labeled with [3H]-oleic acid at the 1-position, and that the assay was conducted in the presence of a MAGL inhibitor, the release of [3H]-oleic acid specifically reflects the activity of the LPA-PLA1 enzyme (Figure 6). Notably, in adult SYN, there was a significant increase in enzymatic activity induced by the THC-free extract (22 %), the full-spectrum extract (37 %), and THC (44 %), with the difference between the first and last treatments being statistically significant. In aged SYN, only THC elicited a significant increase in enzymatic activity (29 %), restoring it to levels observed in the adult control.
3. Discussion
Aging is a complex and multifactorial biological process that leads to a progressive decline in the physiological functions of all organ systems, with the nervous system being particularly susceptible. At the cerebral level, this process is characterized by synaptic dysfunction, a reduction in neurogenesis, and a diminished responsiveness of glial cells [39,40,41]. In some cases, this physiological aging may constitute an antecedent stage to the pathological aging that is characteristic of neurodegenerative disorders. Extensive research has elucidated the integral roles of the ECS in modulating crucial physiological processes, among them the cognitive decline, a hallmark of normal aging. 2-AG is a bioactive lipid whose function is precisely controlled by its tissue levels, which are critically dependent on the finely tuned balance between its synthesis and degradation. Disruption of this balance can lead to dysregulation of 2-AG activity and affect its physiological functions. The synaptic terminal is a vulnerable neuronal structure where aging disrupts the ECS, particularly 2-AG metabolism [13]. With respect to synaptic disfunction occurring in aging, recent studies have demonstrated that low doses of THC (1-3 mg/kg/day) significantly improve spatial learning and enhance cognitive performance in aged mice. However, a 1:1 combination of THC and CBD does not produce these effects [22]. Based on this, and on our previous findings, which demonstrated a reduction in the availability of the endocannabinoid 2-AG in synaptic terminals of the CC from aged rats, and its partial reversal by CBR ligands [13,14], we hypothesized that cannabis extracts with a specific phytocannabinoid composition could modulate the endogenous metabolism of this neuroprotective endocannabinoid.
To address this hypothesis, we commenced our study with an extract obtained from a plant containing 37.72 mg of THC/g inflorescence. The effects of the THC-free extract, the full-spectrum extract, and THC were assessed on the synthesis (via DAGL and LPAase) and hydrolysis (via MAGL) of 2-AG. Through a dose-dependent analysis of MAGL activity, we identified 1 µM THC as an appropriate concentration for subsequent experiments. The THC concentration range assayed in this analysis was based on studies on synaptic terminals that assess the effects of this phytocannabinoid on catecholamine uptake, release [42] and synthesis [43]; calcium uptake [44]; and cAMP levels [45], among other parameters. Although the magnitude of MAGL inhibition at 1 µM THC was modest (7%), this value was deemed biologically relevant because 2-AG is a retrograde signaling molecule present at low basal levels, and even small changes in its enzymatic regulation can significantly affect synaptic endocannabinoid tone, where local signaling is tightly controlled. The selected THC concentration also modulated DAGL activity but did not alter LPAase activity in aged SYN. However, the possibility that different THC concentrations could impact LPAase activity cannot be excluded. As only full-spectrum and THC-free extracts —but not pure THC— diminished MAGL activity, our findings in aged SYN, indicate that this effect of cannabis is independent of THC (Figure 2b). This suggests that one or more metabolites present in the extract, other than THC, may mediate the observed inhibition.
Reports about MAGL in wild-type COS cells demonstrated that certain herbal extracts with diverse phytocannabinoid compositions —unlike the pure cannabinoids— were capable of inhibiting MAGL activity [46]. As mentioned above, cannabis produces a diverse varied spectrum of bioactive compounds, including cannabinoids and terpenoids. β-caryophyllene, a sesquiterpene with pharmacological properties, including antioxidant and anti-inflammatory activities [47], has been shown to inhibit MAGL in an analgesia model [48]. This terpene has often been described as a ligand for CB2 [49]. In this concern, MAGL activity has been previously shown to be modified by CBR ligands [14]. Although this sesquiterpene was not detected among the compounds identified in the THC-free fraction used in our assays, the caryophyllene oxide present in the fraction could potentially be a metabolite derived from β-caryophyllene oxidation during GC analysis (as shown in the Supplementary Material). Additional biomolecules identified in the THC-free extract included amyrins (shown in Supplementary Material). The triterpenes α- and β-amyrins have also been documented to inhibit the hydrolysis of 2-AG [50]. These compounds may contribute to the inhibition of MAGL, thereby enhancing the availability of the neuroprotective 2-AG.
With respect to 2-AG synthesis, the co-presence of THC with other cannabis metabolites nullifies the inhibitory effect that each exerts individually on DAGL activity in aged SYN, (Figure 3). Given that DAGL and MAGL operate via a shared mechanism of action —both enzymes catalyze the hydrolysis of a fatty acid esterified to glycerol— and both enzymes are members of the lipase-3 family of serine hydrolases [46], it can be hypothesized that the inhibitory effect attributed either to β-caryophyllene or amyrins on MAGL activity may also extend to DAGL activity. Although non-THC compounds present in the THC-free extract, inhibit both MAGL and DAGL activity, THC only inhibit DAGL. This could reflect a different mechanism of action of this THC and the non-THC compounds (eg. amyrins) on DAGL inhibition.
The fact that THC and non-THC compounds inhibition of DAGL is abolished in the presence of the cannabis extract, may appear contradictory. However, this apparent inconsistency highlights the complexity of the enzymatic network and the possibility of interactions among phytochemicals within the extract. The 'entourage effect' caused by the interactions among the different constituents from cannabis plant have been documented in cell-based and animal models. This concept proposes that terpenoids and phytocannabinoids as well as other constituents can individually modulate the activity of one another, thereby enhancing or modifying their overall effects and ultimately leading to increased or novel therapeutic outcomes that are not achieved with their isolated components [18,19,20]. In this concern, the effect of THC on CB1 and CB2 receptors have been reported to range from antagonistic to agonistic, depending on factors such as the presence of other ligands that interact with the same targets [51]. Amyrins has also been shown to exhibit potent biological activity by binding to the CB1 with an affinity 200-300 times greater than that of THC [52]. Thus, it could be hypothesized that terpenes such as amyrins, by binding to the CB1 receptor, may modulate the effect of THC on this receptor, and possibly explain why the presence of both compounds in the full-spectrum cannabis extract leads to different effects compared to when each compound is separated. In this regard, DAGL activity has been previously shown to be modified by CBR ligands [14].
It is noteworthy that, although two DAGL isoforms have been identified, we have previously shown that, in synaptosomal CC fractions, DAGL-β is present in both adult and aged rats, whereas DAGL-α is confined to membrane fractions and is undetectable in synaptosomal CC fractions from either age group [13].
MAGL activity was assessed not only with an exogenous substrate but also with MAG produced during DAGL assay, exhibiting different patterns in aged SYN: 1) the inhibition exerted by the THC-free extract in MAGL coupled to DAGL was four times higher than those observed with the exogenous substrate; 2) while in the MAGL coupled to DAGL assay THC inhibits glycerol production, in MAGL assay this effect was not observed, and 3) in MAGL coupled to DAGL, the presence of THC in the extract restored its own inhibitory effect. This suggests that the observed inhibition in MAGL coupled to DAGL activity is, at least in part, due to the experimental conditions affecting DAGL, as the inhibition of this activity by both the THC-free extract and THC reduces the availability of the substrate for MAGL.
As mentioned previously, LPAase activity was unaffected under the experimental conditions assayed. Previous investigations by our group in 2-AG metabolism in various models —such as amyloidosis induced in synaptic terminals by the oligomeric form of beta-amyloid peptide [36] and the effect of light on retinal photoreceptor cells [53] demonstrated that, whereas DAGL and MAGL activities were significantly altered, LPAase activity remained unchanged.
Upon analyzing the results related to the enzymes involved in 2-AG metabolism under the different experimental conditions, it becomes evident that the metabolic processing of 2-AG is not significantly altered in SYN derived from adult rats. This observation implies that the endogenous tone of 2-AG is adequately maintained to support synaptic function under physiological conditions, while in aged organisms this tone is reported to be mainly reduced [13,54]. This could explain the fact that THC-containing cannabis extracts effects are shown to be positive in aged organisms and not in younger ones [55], and these results are consistent with that observed in the present study.
2-AG not only functions as a bioactive lipid but also serves as a precursor for the synthesis of other lipids, such as arachidonic acid. The release of arachidonic acid could be regulated by the enzymatic activities of MAGL and LPA-phospholipase A (LPA-PLA). In our study, the production of fatty acid from LPA, when LPAase activity was assessed in the presence of a MAGL inhibitor, further indicated the hydrolysis of LPA by a LPA-PLA. This enzymatic activity, which was diminished in aged SYN compared to adult ones, was enhanced by THC in both age groups, restoring the activity in aged SYN to levels observed in control adults. These findings, along with the observation that LPAase activity remained unaltered under these conditions, suggest a reduced availability of the substrate (LPA) for the synthesis of the endocannabinoid 2-AG when THC was present. While our experimental conditions focus on evaluating LPA-PLA1, Yaginuma et al. (2022) demonstrated that LPA-PLA activity hydrolyzes esterified fatty acids at both the sn-1 and sn-2 positions of lysolipids with comparable efficiency [56]. Thus, the increased LPA-PLA activity induced by THC may indicate that this phytocannabinoid facilitates the release of arachidonic acid, which then serves as a substrate for the production of pro-inflammatory molecules. In this respect, it has been reported that THC-induced arachidonic acid release occurs through a receptor-mediated process involving the activation of one or more phospholipases [57] and that the enzymes implicated in eicosanoid-mediated cannabinoid actions include MAGL, phospholipase A2, cyclooxygenases, lipoxygenases, and additional related enzymes. [58].
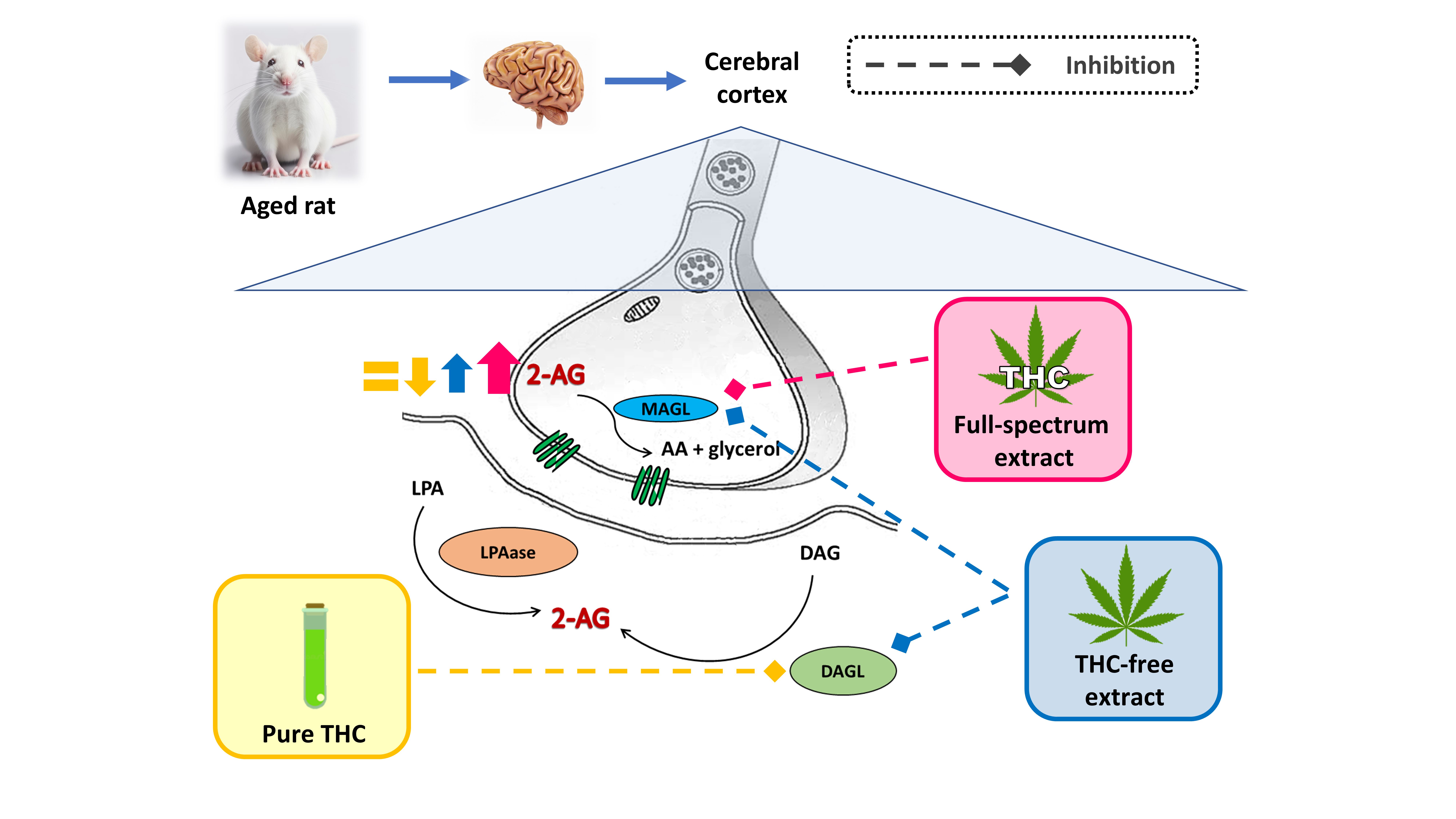
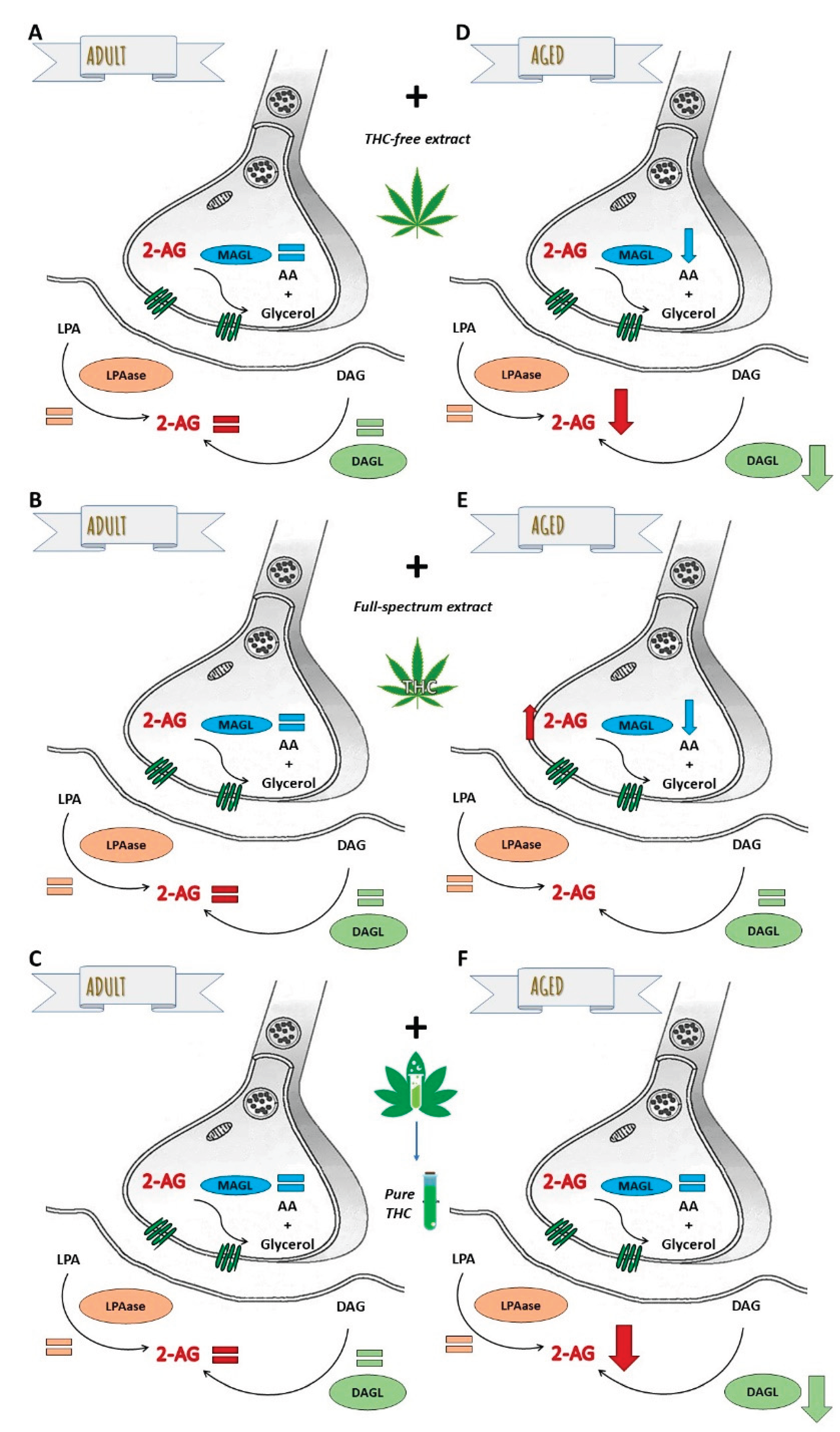
The principal findings of this study, summarized in Figure 7, reveal the following about 2-AG availability: 1) in adult SYN, remains unaffected by the THC-free extract, the full-spectrum extract, or THC (Figure 7 A-C); 2) in aged SYN, is decreased by both the THC-free extract and THC, predominantly via the suppression of DAGL activity (Figure 7 D and F); however, it is increased in the presence of the full-spectrum extract with high THC content by a decrease in MAGL activity (Figure 7 E). The reduction in 2-AG bioavailability induced by the THC-free extract and THC, which is reversed and also increased by the full-spectrum extract, suggests a beneficial interaction between the non-cannabinoid components of the extract and THC. This observation indicates that 2-AG availability could be enhanced by modulating its metabolism through a full-spectrum extract containing, at least, THC as phytocannabinoid. The enzymes involved in 2-AG metabolism represent promising therapeutic targets for mitigating synaptic dysfunction and protecting the nervous system from damage associated with the aging process, however if the observed effects of cannabis extracts are mediated by receptors, or if they result directly from modulation of enzymatic activities, are objectives to be elucidated in future studies.
In this study, we employed in vitro assays in isolated synaptosomes, providing a controlled and well-defined model to explore endocannabinoid metabolism at the synaptic level. This approach offers a solid framework that can be further expanded in future studies to determine whether cannabis extracts also modulate stimulus-evoked 2-AG release and to validate these findings in vivo, thereby extending their physiological relevance.
4. Materials and Methods
4.1. Materials
Cannabis sp. flowers were provided by the non-profit organization Asociación Civil Cultivo mi Medicina, in Bahía Blanca, Buenos Aires, Argentina. Standard solutions of cannabinol (CBN), cannabidiol (CBD), and (-)-Δ9-tetrahydrocannabinol ((-)-Δ9-THC) (1 mg/mL in methanol) were obtained from Cerilliant Corporation (Round Rock, TX, USA). [2-3H]Glycerol (2 Ci/mmol) was sourced from Perkin Elmer (Boston, MA, USA). Lysophosphatidic acid, 1-oleoyl [oleoyl-9,10-3H(N)] (54 Ci/mmol) was acquired from American Radiolabeled Chemicals, Inc. (Saint Louis, MO, USA). Additional reagents, including oleoyl-L-α-lysophosphatidic acid, N-ethylmaleimide (NEM), bovine serum albumin (BSA), 2,5-diphenyloxazole (PPO), and 1,4-bis(5-phenyl-2-oxazolyl) benzene (bis-MSD), were purchased from Sigma-Aldrich (St. Louis, MO, USA). All other chemicals utilized in this study were of the highest available purity.
4.2. Preparation of the Cannabis Extract
Female inflorescences of cannabis were kept at -20 °C in light-protected, nitrogen-filled tubes. Before extraction, acidic cannabinoids decarboxylation was achieved by preheating the inflorescences in an oven at 115 °C for 40 minutes [23]. Cannabis extraction was subsequently performed employing absolute ethanol (EtOH) (5 % w/v) as extraction solvent, as described by Romano and Hazekamp [24], with minor modifications.
4.3. Preparation of the THC-Free Cannabis Extract
EtOH was evaporated from the cannabis extract under a stream of nitrogen to obtain the resin. This resin (1 mg) was separated by preparative thin-layer chromatography (TLC) using a plastic-backed plate coated with silica gel 60 (0.2 mm thickness, Merck), with dimensions of 12.5 cm by 20 cm. The mobile phase consisted of a hexane:ethyl acetate mixture (90:10). A THC standard was spotted along both edges of the plate. One side was developed using iodine vapors, while the other side was developed with a p-anisaldehyde solution (5 mL p-anisaldehyde, 5 mL concentrated H2SO4, 1 mL glacial acetic acid, and 90 mL EtOH) followed by heating with a heat gun. The TLC plate was fractionated to isolate THC from other compounds. The THC fraction and the THC-free fractions were scraped separately, and then washed with EtOH, filtered, and evaporated under a nitrogen stream. The THC-free fractions were subsequently combined to obtain a THC-free extract.
4.4. Identification and Quantification of Phytocannabinoids
Ethanolic extract was analyzed using High-Performance Liquid Chromatography with Diode-Array Detection (HPLC-DAD). Prior to quantification, the resin was re-suspended in methanol.
The HPLC system used was a Thermo Scientific Dionex Ultimate 3000 equipped with a Diode-Array Detector. Samples were detected at a wavelength of 220 nm following separation on a Phenomenex Luna C8 column (250 × 4.6 mm, 5 μm). The mobile phase consisted of acetonitrile and water in an 80:20 ratio, with a flow rate of 1 mL/min. The HPLC method was adapted from the protocol described by Raharjo and Verpoort [25].
To identify and/or quantify phytocannabinoid peaks in the chromatograms, CBD, CBN, and (-)-Δ9-THC standard solutions were used.
THC fraction and THC-free fractions were analyzed by both HPLC (as described above) and gas chromatography mass spectrometry (GC-MS). For GC-MS analysis, a Hewlett Packard CG-6890 with an MS-5972A mass selective detector was employed, using an HP5-MS capillary column (30 m × 250 μm × 0.25 μm). Mass spectra were obtained at 70 eV in the m/z range of 35 to 700 a.m.u, with helium as the carrier gas at a flow rate of 1.0 mL/min. Identification of chromatographic peaks was based on comparison with the MS library, with global match score for THC exceeding 900. Results from these analyses are shown in Supplementary Material.
4.5. Preparation of Synaptosomes
Adult (4 months old, 250–300 g) and aged (28 months old, 450–500 g) Wistar rats (50% male and 50% female) were used in this study. Animals were maintained under controlled environmental conditions and housed two per cage in standard polycarbonate cages, with ad libitum access to chow and water. Rats were obtained from the Laboratory of Experimental Animals (Laboratorio de Animales de Experimentación, LAE), Faculty of Veterinary Sciences, National University of La Plata (La Plata, Argentina), and had been maintained for several years in the institutional animal facility.
Sample size was calculated a priori using the Snedecor and Cochran (1989) [26] formula, based on preliminary data [13,14] to estimate the pooled standard deviation (s) and the expected difference between groups (d). Calculations assumed a significance level of α=0.05 and statistical power of 1−β=0.8. Estimated sample sizes ranged between 1.08 and 1.7 animals per group, depending on the enzyme analyzed. Accordingly, experiments were performed with a minimum of three biological replicates per group, each consisting of a pool of two animals. Due to the unpredictable mortality rates of aged animals, extra animals were included beyond the minimum required. When these animals survived, they were used for additional repetitions to confirm observed trends, within the constraints of variable animal availability. Both adult rats and aged rats were first sedated with carbon dioxide gas and subsequently euthanized by decapitation. Immediately following euthanasia, the cerebral cortices were dissected and used to isolate SYN as previously described [13,27]. The isolated SYN were stored at -80 °C until further use. All experimental procedures were conducted following the approval of the Institutional Animal Care and Use Committee of the Universidad Nacional del Sur (IACUC, protocol number 228-2023-CDBByF Res: 609/23) and in conformity with the guidelines outlined in the Care and Use of Laboratory Animals (National Institutes of Health).
4.6. Preparation of Radiolabeled Substrates
[2-3H]Glycerolphosphatidylcholine (PC) and [2-3H]Triacylglycerol (TAG) were synthesized from bovine retinas that were incubated with [2-3H]glycerol, as described by Pasquaré de Garcia and Giusto [28]. 1,2-Diacyl-sn-glycerol (DAG) and monoacyl-sn-glycerol (MAG) were generated by the action of phospholipase C from Clostridium welchii (Grade B, Calbiochem, Los Angeles, CA) on [2-3H]PC [29], and through the hydrolysis of [2-3H]TAG with pancreatic lipase, respectively [13].
Lipids were extracted according to the method outlined by Folch et al. [30] and then separated by one-dimensional TLC, following the procedures detailed in prior publications [13,29,31]. The specific activities of the substrates employed in the enzymatic assays, as determined using Gas Liquid Chromatography, were 0.17 µCi/µmol for [3H]DAG and 1.15 µCi/µmol for [3H]MAG.
4.7. DAGL Activity Assay
The enzymatic assay was carried out using [3H]DAG, which was prepared by sonicating [3H]DAG plus unlabeled DAG, with equal molar amounts of lysoPtdCho in a 50 mM MOPS buffer (pH 7.4) containing 0.25 % fatty acid-free bovine serum albumin (BSA), resulting in a final volume of 100 µL [31]. The assay for DAGL was started by adding DAG suspensions (600 µM, 11,500 DPM) to SYN (100 µg of protein).
4.8. LPAase Activity Assay
The incubation mixtures consisted of unlabeled 1-oleoyl LysoPtdOH and 1-oleoyl [oleoyl-9,10-3H(N)]-LysoPtdOH (20 µM, 6,000 DPM), 100 mM Tris-HCl (pH 7.4), 1.2 mM dithiothreitol (DTT), 2 mM ethylenediaminetetraacetic acid (EDTA), and SYN (50 µg protein) in a total volume of 100 µL [32]. Prior to substrate addition, SYN were pre-incubated with 2 µM KML29, a selective MAGL inhibitor [33], for 30 minutes; and N-ethylmaleimide (NEM) for 10 minutes to specifically assess NEM-insensitive LAPase activity.
4.9. MAGL Activity Assay
This activity was evaluated by incubating SYN (50 µg protein) with [3H]-MAG (20 µM, 5,000 DPM) in a Tris-HCl buffer (50 mM, pH 7.5) containing 1 mM EDTA, in a total volume of 100 µL, as described by Baker and Chang [34] and Blankman et al. [35].
For each enzymatic assay, SYN were co-incubated for 20 minutes with the respective substrate and either the full-spectrum extract, the THC-free extract, THC, or their vehicle (ethanol), which served as the control. All assays were conducted at 37 °C and the reactions were stopped by the addition of chloroform:methanol (2:1, v/v) or chloroform:methanol (1:1, v/v). The conditions for the enzymatic assays were previously optimized and reported in earlier publications [13,14,36]. Lipid products were then extracted following the method of Folch et al. [30] and separated using previously described techniques with minor modifications, and glycerol was recovered from the aqueous phase [13,37]. The radiolabeled products were quantified after adding 0.25 mL of water and 5 mL of a 0.5 % mixture of 98 % PPO and 2 % bis-MSD in toluene/Arkopal N-100 (4:1, v/v).
Blank controls were prepared in the same manner as the enzymatic assays, except that the SYN proteins were previously denatured by adding chloroform:methanol (2:1, v/v).
Protein concentration was measured using the DC Protein Assay (#500-0116, BIO-RAD, Hercules, CA, USA), following the method established by Lowry et al. [38].
4.10. Statistical Analysis
A minimum of three experiments were conducted, utilizing a pool of two animals per experiment. Each experiment included at least three replicates per condition, with each experiment treated as an individual sample (n). Potential outliers were visually inspected using Q–Q plots. Prior to each statistical analysis, the assumption of normality was verified using the Shapiro–Wilk (p>0.05) test. Data were analyzed using one- or two-way ANOVA, as appropriate. When no significant interaction between age and THC treatment was found, a one-way ANOVA was applied for each age group. In all cases, Fisher’s LSD post hoc test was used, as indicated in the respective figures. For all column figures, bars represent mean ± standard error of the mean (SEM), with individual data points shown (n). Statistical analysis was performed with Infostat version 2018 program (http://www.infostat.com.ar).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: GC-MS chromatogram of Fraction 1; Table S1: Identified compounds in Fraction 1 extract by GC-MS analysis; Figure S2: GC-MS chromatogram of Fraction 2; Table S2: Identified compounds in Fraction 2 extract by GC-MS analysis; Figure S3: GC-MS chromatogram of Fraction 3; Table S3: Identified compounds in Fraction 3 extract by GC-MS analysis; Figure S4: GC-MS chromatogram of Fraction 4. Table S4. Identified compounds in Fraction 4 extract by GC-MS analysis.
Author Contributions
Conceptualization, S.R.S., A.C.P. and S.J.P.; Methodology, S.R.S., A.C.P. and S.J.P.; Investigation, S.R.S. and A.C.P.; Resources (THC-free extract preparation and GC/MS analysis), F.A.M. and A.P.M.; Data curation (HPLC quantification), P.G.M.; Formal analysis, S.R.S., A.C.P. and S.J.P.; Writing—original draft preparation, S.J.P.; Writing—review and editing, all authors; Supervision, S.J.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by funds granted by the Ministerio Nacional de Ciencia, Tecnología e Innovación (Proyectos de Investigación y Desarrollo en Cannabis 3A), the Agencia Nacional de Promoción Científica y Tecnológica (2017-1089), the Consejo Nacional de Investigaciones Científicas y Técnicas (PIP 11220200100707), and the Secretaría General de Ciencia y Tecnología, Universidad Nacional del Sur (24/B250), Argentina, to Prof. S.J. Pasquaré.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Animal Care and Use Committee (IACUC) of the Universidad Nacional del Sur, Argentina (protocol number 228-2023-CDBByF, Res. 609/23), and adhered to the standards set forth in the Guide for the Care and Use of Laboratory Animals by the Institute for Laboratory Animal Research (ILAR) of the National Academy of Sciences (Bethesda, MD, USA).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated during and/or analysed during the current study can be available from the corresponding author on request.
Acknowledgments
The authors would like to acknowledge to the non-profit organization Asociación Civil Cultivo mi Medicina, Bahía Blanca, Buenos Aires, Argentina for providing the Cannabis sp. flowers used in this study.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| 2-AG | 2-arachidonoylglycerol |
| AEA | anandamide |
| BSA | bovine serum albumin |
| CBR | cannabinoid receptors |
| CB1 | cannabinoid receptor 1 |
| CB2 | cannabinoid receptor 2 |
| CC | cerebral cortex |
| DAG | diacylglycerol |
| DAGL | diacylglycerol lipase |
| DTT | dithiotreitol |
| ECS | endocannabinoid system |
| LPA | lysophosphatidic acid |
| LPAase | lysophosphatidate phosphohydrolase |
| LPA-PLA1 | lysophosphatidate phospholipase A1 |
| LPC | lysophosphatidilcholine |
| MAG | monoacylglycerol |
| MAGL | monoacylglycerol lipase |
| ABHD6 | serine hydrolase alpha-beta-hydrolase domain 6 |
| NEM | N-ethylmaleimide |
| SYN | synaptosomes |
| THC | ∆9-tetrahydrocannabinol |
References
- Devane, W. A.; Hanuš, L.; Breuer, A.; Pertwee, R. G.; Stevenson, L. A. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef]
- Sugiura, T.; Kishimoto, S.; Oka, S.; Gokoh, M. Biochemistry, pharmacology and physiology of 2-arachidonoylglycerol, an endogenous cannabinoid receptor ligand. Prog. Lipid Res. 2006, 45, 405–446. [Google Scholar] [CrossRef]
- Howlett, A. C. The cannabinoid receptors. Prostaglandins Other Lipid Mediat. 2002, 68-69, 619–631. [Google Scholar] [CrossRef]
- Maurelli, S.; Bisogno, T.; De Petrocellis, L.; Di Luccia, A.; Marino, G.; et al. Two novel classes of neuroactive fatty acid amides are substrates for mouse neuroblastoma “anandamide amidohydrolase”. FEBS Lett. 1995, 377, 82–86. [Google Scholar] [CrossRef]
- Okamoto, Y.; Wang, J.; Morishita, J.; Ueda, N. Biosynthetic pathways of the endocannabinoid anandamide. Chem. Biodivers. 2007, 4, 1842–1857. [Google Scholar] [CrossRef] [PubMed]
- Gulyas, A. I.; Cravatt, B. F.; Bracey, M. H.; Dinh, T. P.; Piomelli, D.; et al. Segregation of two endocannabinoid-hydrolyzing enzymes into pre- and postsynaptic compartments in the rat hippocampus, cerebellum and amygdala. Eur. J. Neurosci. 2004, 20, 441–458. [Google Scholar] [CrossRef] [PubMed]
- Prescott, S. M.; Majerus, P. W. Characterization of 1,2-diacylglycerol hydrolysis in human platelets. Demonstration of an arachidonoyl-monoacylglycerol intermediate. J. Biol. Chem. 1983, 258, 764–769. [Google Scholar] [CrossRef]
- Dinh, T. P.; Kathuria, S.; Piomelli, D. RNA interference suggests a primary role for monoacylglycerol lipase in the degradation of the endocannabinoid 2-arachidonoylglycerol. Mol. Pharmacol. 2004, 66, 1260–1264. [Google Scholar] [CrossRef] [PubMed]
- Nakane, S.; Oka, S.; Arai, S.; Waku, K.; Ishimab, Y.; et al. 2-Arachidonoyl-sn-glycero-3-phosphate, an arachidonic acid-containing lysophosphatidic acid: occurrence and rapid enzymatic conversion to 2-arachidonoyl-sn-glycerol, a cannabinoid receptor ligand, in rat brain. Arch. Biochem. Biophys. 2002, 402, 51–58. [Google Scholar] [CrossRef]
- Bahr, B. A.; Karanian, D. A.; Makanji, S. S.; Makriyannis, A. Targeting the endocannabinoid system in treating brain disorders. Expert Opin. Investig. Drugs 2006, 15, 351–365. [Google Scholar] [CrossRef]
- Freund, T. F.; Katona, I.; Piomelli, D. Role of endogenous cannabinoids in synaptic signaling. Physiol. Rev. 2003, 83, 1017–1066. [Google Scholar] [CrossRef]
- Mulder, J.; Zilberter, M.; Pasquaré, S. J.; Alpár, A.; Schulte, G.; et al. Molecular reorganization of endocannabinoid signalling in Alzheimer’s disease. Brain 2011, 134, 1041–1060. [Google Scholar] [CrossRef]
- Pascual, A. C.; Gaveglio, V. L.; Giusto, N. M.; Pasquaré, S. J. Aging modifies the enzymatic activities involved in 2-arachidonoylglycerol metabolism. BioFactors 2013, 39, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Pascual, A. C.; Gaveglio, V. L.; Giusto, N. M; Pasquaré, S. J. Cannabinoid receptor-dependent metabolism of 2-arachidonoylglycerol during aging. Exp. Gerontol. 2014, 55, 134–142. [Google Scholar] [CrossRef]
- de Meijer, E. The Chemical Phenotypes (Chemotypes) of Cannabis. In Handbook of Cannabis, online ed.; Pertwee, R., Ed.; Oxford University Press: Oxford, UK, 2015; pp. 89–110. [Google Scholar]
- Aizpurua-Olaizola, O.; Soydaner, U.; Öztürk, E.; Schibano, D.; Simsiret, Y.; et al. Evolution of the Cannabinoid and Terpene Content during the Growth of Cannabis sativa Plants from Different Chemotypes. J. Nat. Prod. 2016, 79, 324–331. [Google Scholar] [CrossRef]
- Hanuš, L. O.; Meyer, S. M.; Muñoz, E.; Taglialatela-Scafati, O.; Appendino, G.; et al. Phytocannabinoids: a unified critical inventory. Nat. Prod. Rep. 2016, 33, 1357–1392. [Google Scholar] [CrossRef]
- Russo, E. B. Taming THC: potential cannabis synergy and phytocannabinoid-terpenoid entourage effects. Br. J. Pharmacol. 2011, 163, 1344–1364. [Google Scholar] [CrossRef] [PubMed]
- Blasco-Benito, S.; Seijo-Vila, M.; Caro-Villalobos, M.; Tundidor, I.; Andradas, C.; et al. Appraising the “entourage effect”: Antitumor action of a pure cannabinoid versus a botanical drug preparation in preclinical models of breast cancer. Biochem. Pharmacol. 2018, 157, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Nallathambi, R.; Mazuz, M.; Namdar, D.; Shik, M.; Namintze, D.; et al. Identification of Synergistic Interaction Between Cannabis-Derived Compounds for Cytotoxic Activity in Colorectal Cancer Cell Lines and Colon Polyps That Induces Apoptosis-Related Cell Death and Distinct Gene Expression. Cannabis Cannabinoid Res. 2018, 3, 120–135. [Google Scholar] [CrossRef]
- Stasiłowicz-Krzemién, A.; Nogalska, W.; Maszewska, Z.; Maleszka, M.; Dobrón, M.; et al. The Use of Compounds Derived from Cannabis sativa in the Treatment of Epilepsy, Painful Conditions, and Neuropsychiatric and Neurodegenerative Disorders. Int. J. Mol. Sci. 2024, 25, 5749. [Google Scholar] [CrossRef]
- Nidadavolu, P.; Bilkei-Gorzo, A.; Krämer, M.; Schürmann, B.; Palmisano, M.; et al. Efficacy of Δ9 -Tetrahydrocannabinol (THC) Alone or in Combination With a 1:1 Ratio of Cannabidiol (CBD) in Reversing the Spatial Learning Deficits in Old Mice. Front. Aging Neurosci. 2021, 13, 718850. [Google Scholar] [CrossRef]
- Casiraghi, A.; Roda, G.; Casagni, E.; Cristina, C.; Musazzi, U. M.; et al. Extraction Method and Analysis of Cannabinoids in Cannabis Olive Oil Preparations. Planta Med. 2018, 84, 242–249. [Google Scholar] [CrossRef]
- Romano, L. L.; Hazekamp, A. Cannabis Oil: chemical evaluation of an upcoming cannabis-based medicine. Cannabinoids 2013, 7, 1–11. [Google Scholar]
- Raharjo, T. J.; Verpoorte, R. Methods for the analysis of cannabinoids in biological materials: a review. Phytochem. Anal. 2004, 15, 79–94. [Google Scholar] [CrossRef]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 8th ed.; Wiley-Blackwell: Ames, IA, USA, 1989. [Google Scholar]
- Cotman, C. W. Isolation of synaptosomal and synaptic plasma membrane fractions. Methods Enzymol. 1974, 31, 445–452. [Google Scholar]
- Pasquaré de García, S. J.; Giusto, N. M. Phosphatidate phosphatase activity in isolated rod outer segment from bovine retina. Biochim. Biophys. Acta 1986, 875, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Gaveglio, V. L.; Pasquaré, S. J.; Giusto, N. M. Metabolic pathways for the degradation of phosphatidic acid in isolated nuclei from cerebellar cells. Arch. Biochem. Biophys. 2011, 507, 271–80. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G. H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Rosenberger, T. A.; Farooqui, A. A.; Horrocks, L. A. Bovine brain diacylglycerol lipase: substrate specificity and activation by cyclic AMP-dependent protein kinase. Lipids 2007, 42, 187–195. [Google Scholar] [CrossRef]
- Fleming, I. N.; Yeaman, S. J. Subcellular distribution of N-ethylmaleimide-sensitive and -insensitive phosphatidic acid phosphohydrolase in rat brain. Biochim. Biophys. Acta 1995, 1254, 161–168. [Google Scholar] [CrossRef]
- Chang, J. W.; Niphakis, M. J.; Lum, K. M.; Cognetta, A. B.; Wanget, C.; et al. Highly selective inhibitors of monoacylglycerol lipase bearing a reactive group that is bioisosteric with endocannabinoid substrates. Chem. Biol. 2012, 19, 579–588. [Google Scholar] [CrossRef]
- Baker, R. R.; Chang, H. A metabolic path for the degradation of lysophosphatidic acid, an inhibitor of lysophosphatidylcholine lysophospholipase, in neuronal nuclei of cerebral cortex. Biochim. Biophys. Acta 2000, 1483, 58–68. [Google Scholar] [CrossRef]
- Blankman, J. L.; Simon, G. M.; Cravatt, B. F. A comprehensive profile of brain enzymes that hydrolyze the endocannabinoid 2-arachidonoylglycerol. Chem. Biol. 2007, 14, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Pascual, A. C.; Gaveglio, V. L.; Giusto, N. M.; Pasquaré, S. J. 2-Arachidonoylglycerol metabolism is differently modulated by oligomeric and fibrillar conformations of amyloid beta in synaptic terminals. Neuroscience 2017, 362, 168–180. [Google Scholar] [CrossRef]
- Giusto, N. M.; Bazán, N. G. Phospholipids and acylglycerols biosynthesis and 14CO2 production from [14C] glycerol in the bovine retina: the effects of incubation time, oxygen and glucose. Exp. Eye Res. 1979, 29, 155–168. [Google Scholar] [CrossRef]
- Lowry, O. H.; Rosebrough, N.; Farr Lewwis, A.; Randall, R. J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Salat, D. H. The declining infrastructure of the aging brain. Brain Connect. 2011, 1, 279–293. [Google Scholar] [CrossRef]
- Stranahan, A. M.; Mattson, M. P. Recruiting adaptive cellular stress responses for successful brain ageing. Nat. Rev. Neurosci. 2012, 13, 209–216. [Google Scholar] [CrossRef]
- Grimm, A.; Eckert, A. Brain aging and neurodegeneration: from a mitochondrial point of view. J. Neurochem. 2017, 143, 418–431. [Google Scholar] [CrossRef] [PubMed]
- Poddar, M. K.; Dewey, W. L. Effects of cannabinoids on catecholamine uptake and release in hypothalamic and striatal synaptosomes. J. Pharmacol. Exp. Ther. 1980, 214, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A. S. Effect of ∆9-tetrahydrocannabinol on the synthesis of dopamine and norepinephrine in mouse brain synaptosomes. J. Pharmacol. Exp. Ther. 1982, 221, 97–103. [Google Scholar] [CrossRef]
- Harris, R. A.; Stokes, J. A. Cannabinoids inhibit calcium uptake by brain synaptosomes. J. Neurosci. 1982, 2, 443–447. [Google Scholar] [CrossRef]
- Little, P. J.; Martin, B. R. The effects of ∆9-tetrahydrocannabinol and other cannabinoids on cAMP accumulation in synaptosomes. Life Sciences 1991, 48, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- De Petrocellis, L.; Ligresti, A.; Moriello, A. S.; Allarà, M.; Bisogno, T.; et al. Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Br. J. Pharmacol. 2011, 163, 1479–1494. [Google Scholar] [CrossRef]
- Machado, K da C.; Islam, M. T.; Ali, E. S.; Rouf, R.; Shaikh, J. U.; et al. A systematic review on the neuroprotective perspectives of beta-caryophyllene. Phytother. Res. 2018, 32, 2376–2388. [Google Scholar] [CrossRef]
- Klawitter, J.; Weissenborn, W.; Gvon, I.; Walz, M.; Klawitteret, J.; et al. β-Caryophyllene Inhibits Monoacylglycerol Lipase Activity and Increases 2-Arachidonoyl Glycerol Levels In Vivo: A New Mechanism of Endocannabinoid-Mediated Analgesia. Mol. Pharmacol. 2024, 105, 75–83. [Google Scholar] [CrossRef]
- Gertsch, J.; Leonti, M.; Raduner, S.; Racz, I.; Chen, J-Z.; et al. Beta-caryophyllene is a dietary cannabinoid. Proc. Natl. Acad. Sci. 2008, 105, 9099–9104. [Google Scholar] [CrossRef]
- Chicca, A.; Marazzi, J.; Gertsch, J.; et al. The antinociceptive triterpene β-amyrin inhibits 2-arachidonoylglycerol (2-AG) hydrolysis without directly targeting cannabinoid receptors. Br. J. Pharmacol. 2012, 167, 1596–1608. [Google Scholar] [CrossRef]
- André, R.; Gomes, A. P.; Pereira-Leite, C.; Marques Da Costa, A.; Monteiro Rodriguez, L.; et al. The Entourage Effect in Cannabis Medicinal Products: A Comprehensive Review. Pharmaceuticals (Basel) 2024, 17, 1543. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, S.; Kathryn, A. B.; Paszcuk, A. F.; Passos, G. F.; Silva, E. S.; et al. Activation of cannabinoid receptors by the pentacyclic triterpene α, β-amyrin inhibits inflammatory and neuropathic persistent pain in mice. Pain 2011, 152, 1872–1887. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, E. C.; Gaveglio, V. L.; Pasquaré, S. J. The Endocannabinoid System Is Present in Rod Outer Segments from Retina and Is Modulated by Light. Mol. Neurobiol. 2019, 56, 7284–7295. [Google Scholar] [CrossRef] [PubMed]
- Piyanova, A.; Lomazzo, E.; Bindila, L.; Lerner, R.; Albayram, O.; et al. Age-related changes in the endocannabinoid system in the mouse hippocampus. Mech. Ageing Dev. 2015, 150, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Bilkei-Gorzo, A.; Albayram, B.; Draffehn, A.; Michel, K.; Piyanova, A.; et al. A chronic low dose of Δ9-tetrahydrocannabinol (THC) restores cognitive function in old mice. Nat. Med. 2017, 23, 782–787. [Google Scholar] [CrossRef]
- Yaginuma, S.; Kawana, H.; Aoki, J. Current Knowledge on Mammalian Phospholipase A1, Brief History, Structures, Biochemical and Pathophysiological Roles. Molecules 2022, 27, 2487. [Google Scholar] [CrossRef]
- Burstein, S.; Budrow, J.; Debatis, M.; Hunter, S. A.; Subramanian, A. Phospholipase participation in cannabinoid-induced release of free arachidonic acid. Biochem. Pharmacol. 1994, 48, 1253–1264. [Google Scholar] [CrossRef]
- Burstein, S. H. Eicosanoid mediation of cannabinoid actions. Bioorg. Med. Chem. 2019, 27, 2718–2728. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(a) Effect of a full-spectrum extract in different THC concentrations on monoacylglycerol lipase (MAGL) activity in synaptosomes from adult rat cerebral cortex and; (b) aged rat cerebral cortex. The substrate [3H] MAG was added exogenously under the assay conditions detailed in the Materials and Methods section. Incubation conditions were optimized for MAGL activity. Results are expressed as nmol of glycerol per mg of protein per minute and are shown as mean ± standard error of the mean (SEM), with individual data points representing the sample size of each group (n varies across columns). Data were analyzed by one-way ANOVA (a (overall model): F = 1.34, p = 0.2802; b (overall model): F = 2.49, p = 0.0473), followed by Fisher’s LSD post-test in b. Identical letters in A and B indicate no statistically significant differences (p ≥ 0.05), whereas different letters in b signify statistically significant differences between conditions (p < 0.05). Graphic was performed using GraphPad Prism 9.5.1 (https://www.graphpad.com).
Figure 1.
(a) Effect of a full-spectrum extract in different THC concentrations on monoacylglycerol lipase (MAGL) activity in synaptosomes from adult rat cerebral cortex and; (b) aged rat cerebral cortex. The substrate [3H] MAG was added exogenously under the assay conditions detailed in the Materials and Methods section. Incubation conditions were optimized for MAGL activity. Results are expressed as nmol of glycerol per mg of protein per minute and are shown as mean ± standard error of the mean (SEM), with individual data points representing the sample size of each group (n varies across columns). Data were analyzed by one-way ANOVA (a (overall model): F = 1.34, p = 0.2802; b (overall model): F = 2.49, p = 0.0473), followed by Fisher’s LSD post-test in b. Identical letters in A and B indicate no statistically significant differences (p ≥ 0.05), whereas different letters in b signify statistically significant differences between conditions (p < 0.05). Graphic was performed using GraphPad Prism 9.5.1 (https://www.graphpad.com).
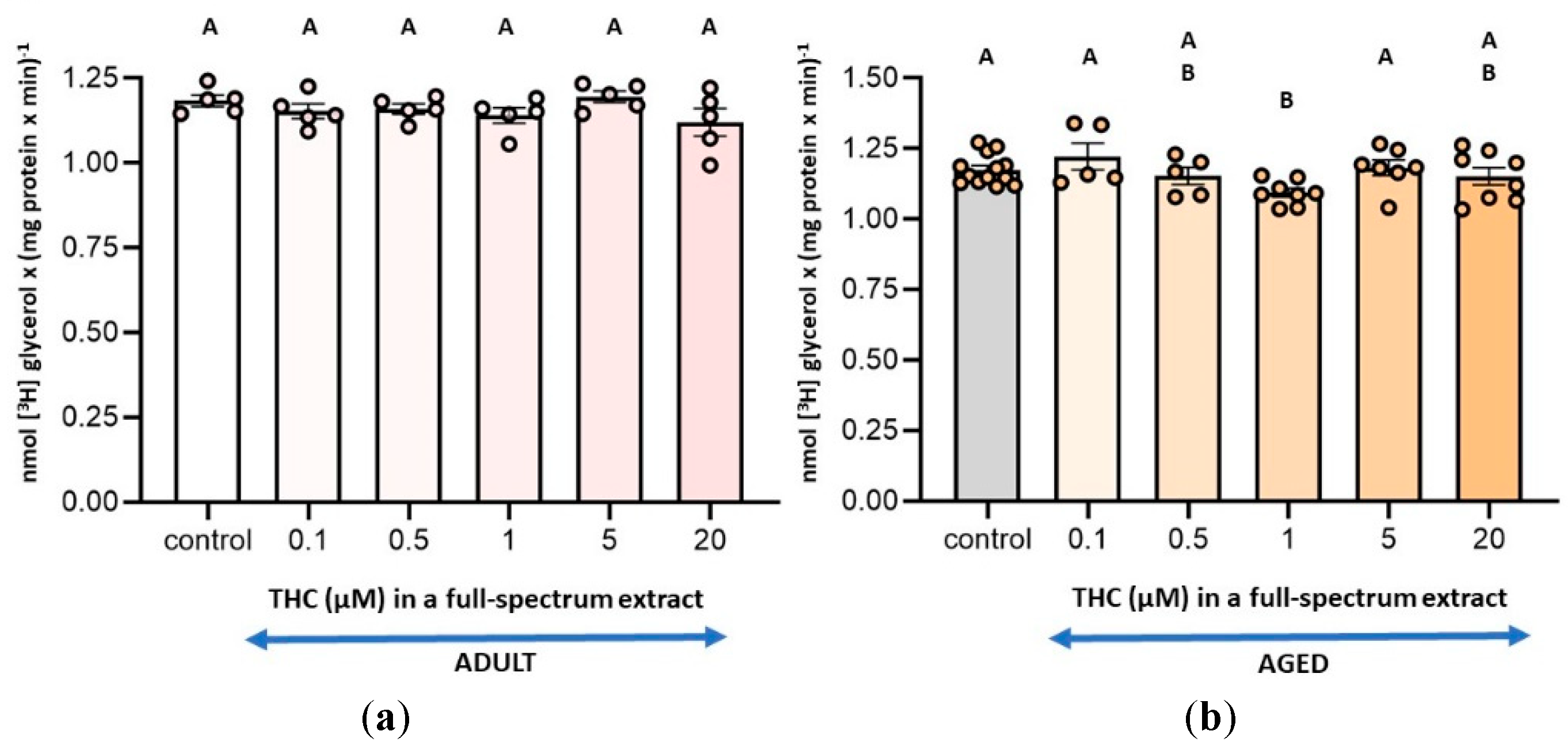
Figure 2.
(a) Effect of the THC-free extract, the full-spectrum extract, and THC on monoacylglycerol lipase (MAGL) activity in synaptosomes from adult rat cerebral cortex; and (b) aged rat cerebral cortex. The assay conditions are outlined in Figure 1 and detailed in the Materials and Methods section. Results are expressed as nmol of glycerol per mg of protein per minute and are shown as mean ± standard error of the mean (SEM), with individual data points representing the sample size of each group (n varies across columns). Data were analyzed by one-way ANOVA (a (overall model): F = 0.95, p = 0.4242; B (overall model): F = 5.90, p = 0.0015), followed by Fisher’s LSD post-test in b. Identical letters in A and B indicate no statistically significant differences (p ≥ 0.05), whereas different letters in b denote statistically significant differences between conditions (p < 0.05). Graphic was performed using GraphPad Prism 9.5.1 (https://www.graphpad.com).
Figure 2.
(a) Effect of the THC-free extract, the full-spectrum extract, and THC on monoacylglycerol lipase (MAGL) activity in synaptosomes from adult rat cerebral cortex; and (b) aged rat cerebral cortex. The assay conditions are outlined in Figure 1 and detailed in the Materials and Methods section. Results are expressed as nmol of glycerol per mg of protein per minute and are shown as mean ± standard error of the mean (SEM), with individual data points representing the sample size of each group (n varies across columns). Data were analyzed by one-way ANOVA (a (overall model): F = 0.95, p = 0.4242; B (overall model): F = 5.90, p = 0.0015), followed by Fisher’s LSD post-test in b. Identical letters in A and B indicate no statistically significant differences (p ≥ 0.05), whereas different letters in b denote statistically significant differences between conditions (p < 0.05). Graphic was performed using GraphPad Prism 9.5.1 (https://www.graphpad.com).
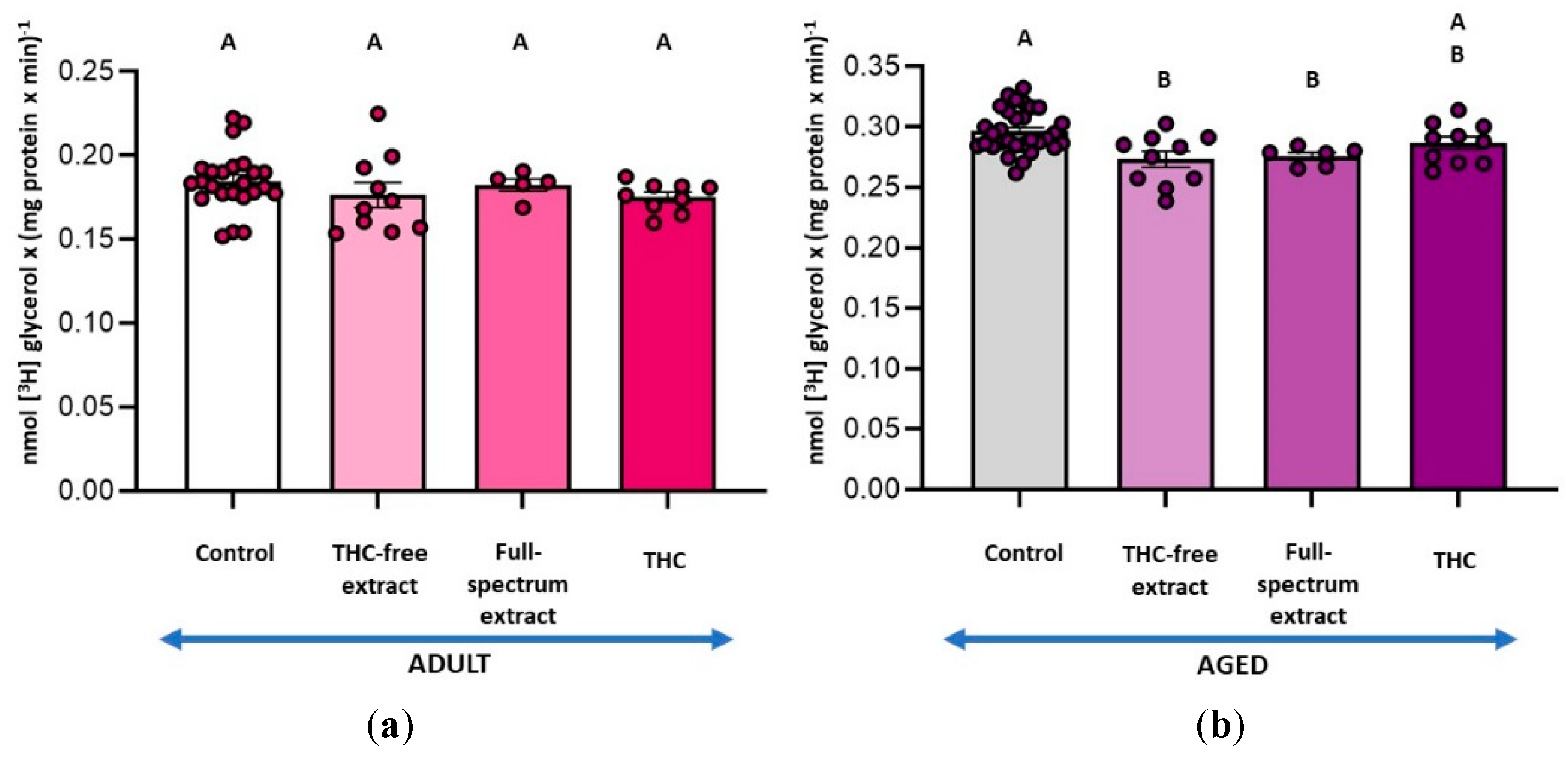
Figure 3.
Effect of the THC-free extract, the full-spectrum extract, and THC on diacylglycerol lipase (DAGL) activity in synaptosomes from Adult and Aged rat cerebral cortex. The substrate [3H]DAG was added exogenously and the assay was conducted according to the conditions specified for DAGL activity as was described in the Materials and Methods section. Results are expressed as nmol of MAG per mg of protein per minute and are shown as mean ± standard error of the mean (SEM), with individual data points representing the sample size of each group (n varies across columns). Data were analyzed by two-way ANOVA (overall model: F = 11.32, p = 0.0001), followed by Fisher’s LSD post-test. Statistical differences between conditions are denoted by different letters (p < 0.05). Graphic was performed using GraphPad Prism 9.5.1 (https://www.graphpad.com).
Figure 3.
Effect of the THC-free extract, the full-spectrum extract, and THC on diacylglycerol lipase (DAGL) activity in synaptosomes from Adult and Aged rat cerebral cortex. The substrate [3H]DAG was added exogenously and the assay was conducted according to the conditions specified for DAGL activity as was described in the Materials and Methods section. Results are expressed as nmol of MAG per mg of protein per minute and are shown as mean ± standard error of the mean (SEM), with individual data points representing the sample size of each group (n varies across columns). Data were analyzed by two-way ANOVA (overall model: F = 11.32, p = 0.0001), followed by Fisher’s LSD post-test. Statistical differences between conditions are denoted by different letters (p < 0.05). Graphic was performed using GraphPad Prism 9.5.1 (https://www.graphpad.com).
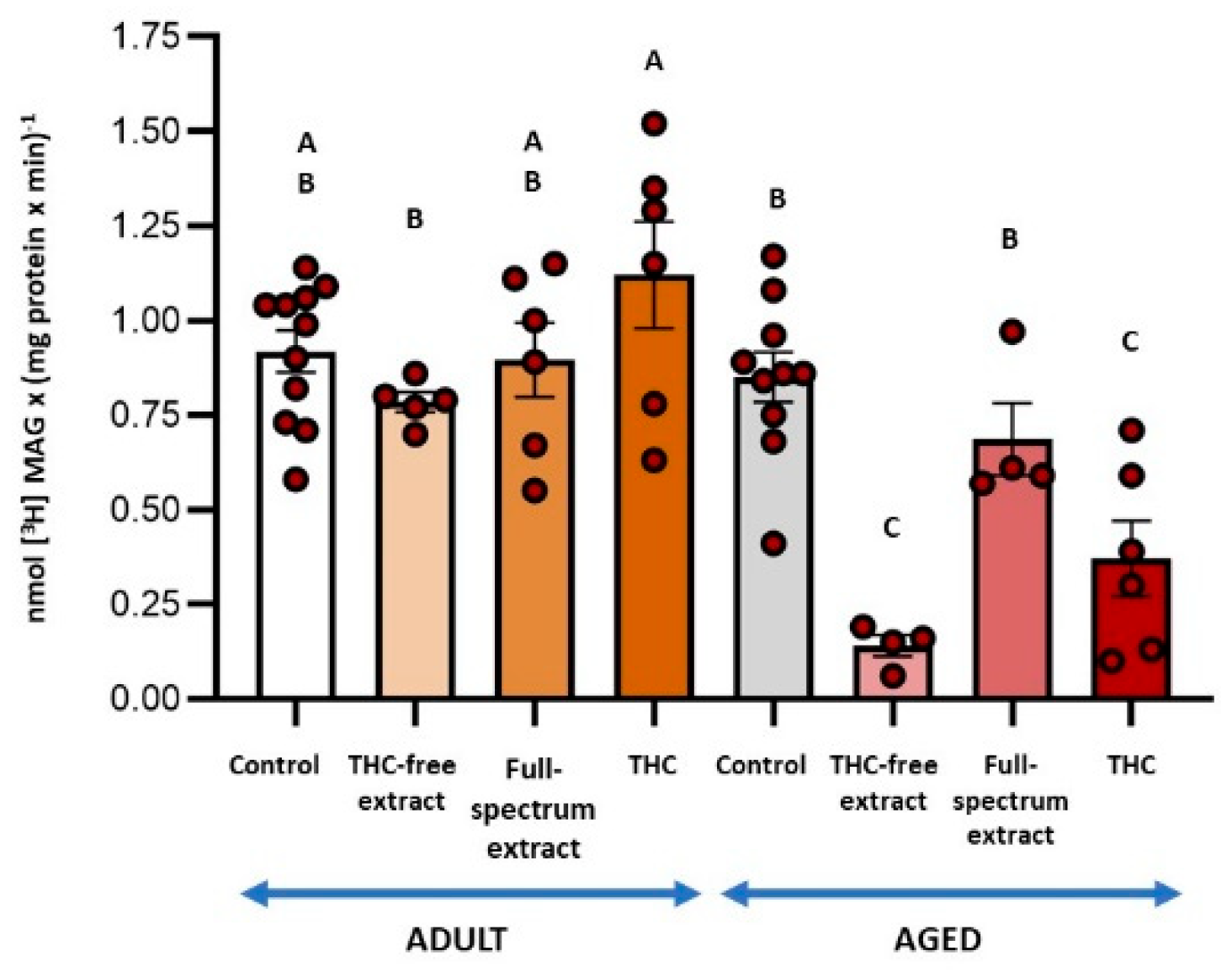
Figure 4.
(a) Effect of the THC-free extract, the full-spectrum extract, and THC on monoacylglycerol lipase (MAGL) activity coupled to diacylglycerol lipase (DAGL) in synaptosomes from adult rat cerebral cortex and; (b) aged rat cerebral cortex. MAGL activity was measured in conjunction with DAGL activity. The substrate [3H]MAG was endogenously produced from the hydrolysis of [3H]DAG by DAGL. Incubation conditions were as detailed for DAGL activity in the Materials and Methods section. Results are reported as nmol of glycerol per mg of protein per minute and are shown as mean ± standard error of the mean (SEM), with individual data points representing the sample size of each group (n varies across columns). Data were analyzed by one-way ANOVA (a (overall model): F = 1.28, p = 0.2932; b (overall model): F = 11.55, p = 0.0001), followed by Fisher’s LSD post-test in b. Similar letters in A and B denote no statistically significant differences (p ≥ 0.05), whereas different letters in b indicate statistically significant differences between conditions (p < 0.05). Graphic was performed using GraphPad Prism 9.5.1 (https://www.graphpad.com).
Figure 4.
(a) Effect of the THC-free extract, the full-spectrum extract, and THC on monoacylglycerol lipase (MAGL) activity coupled to diacylglycerol lipase (DAGL) in synaptosomes from adult rat cerebral cortex and; (b) aged rat cerebral cortex. MAGL activity was measured in conjunction with DAGL activity. The substrate [3H]MAG was endogenously produced from the hydrolysis of [3H]DAG by DAGL. Incubation conditions were as detailed for DAGL activity in the Materials and Methods section. Results are reported as nmol of glycerol per mg of protein per minute and are shown as mean ± standard error of the mean (SEM), with individual data points representing the sample size of each group (n varies across columns). Data were analyzed by one-way ANOVA (a (overall model): F = 1.28, p = 0.2932; b (overall model): F = 11.55, p = 0.0001), followed by Fisher’s LSD post-test in b. Similar letters in A and B denote no statistically significant differences (p ≥ 0.05), whereas different letters in b indicate statistically significant differences between conditions (p < 0.05). Graphic was performed using GraphPad Prism 9.5.1 (https://www.graphpad.com).
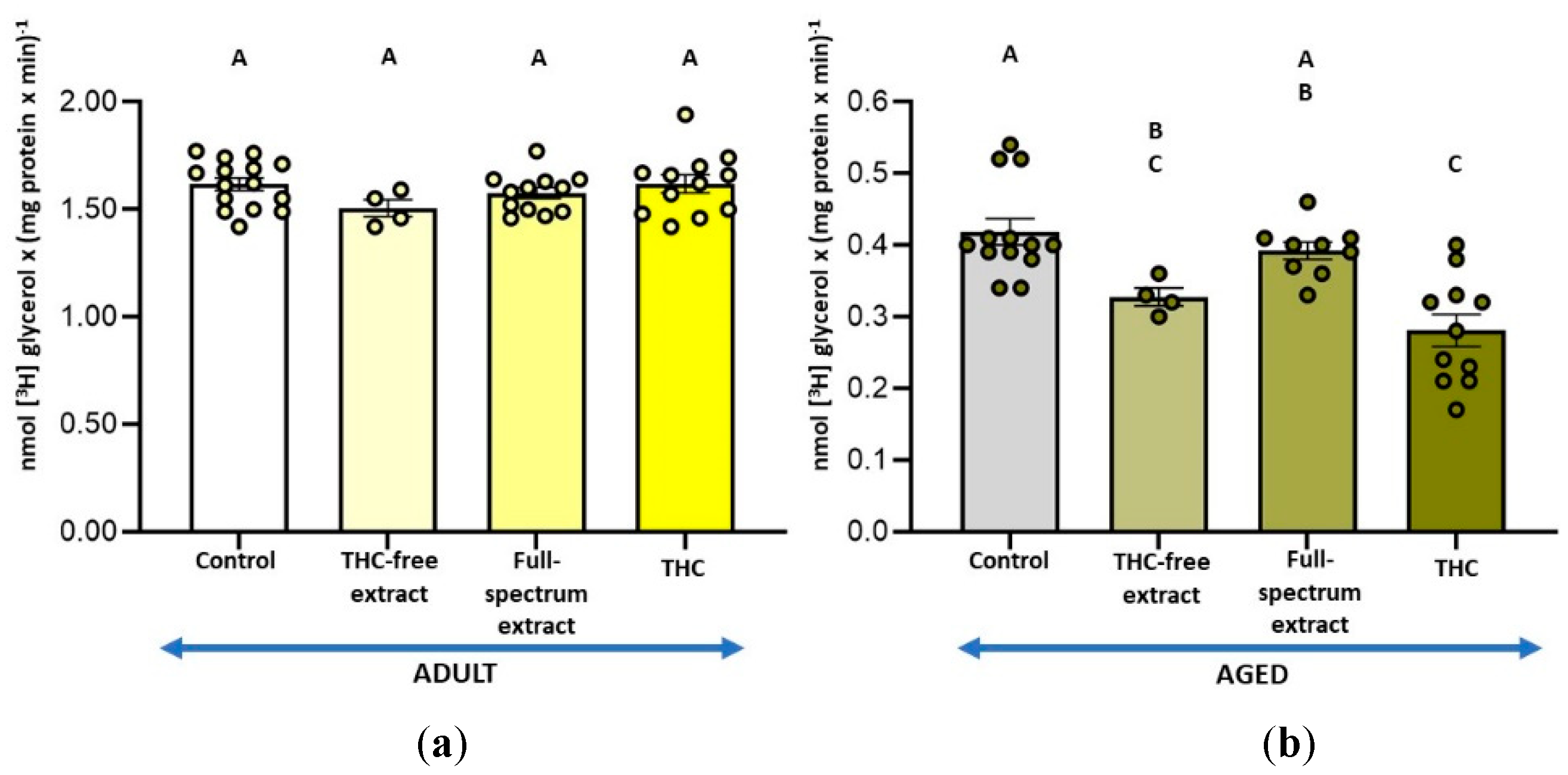
Figure 6.
Effect of the THC-free extract, the full-spectrum extract, and THC on Lysophosphatidate phospholipase A1 activity in synaptosomes from Adult and Aged rat cerebral cortex. Incubation conditions were consistent with those outlined for LPAase activity and specified in the Materials and Methods section. Results are presented as nmol of oleic acid per mg of protein per minute and are shown as mean ± standard error of the mean (SEM), with individual data points representing the sample size of each group (n varies across columns). Data were analyzed by two-way ANOVA (overall model: F = 15.94, p = 0.0001), followed by Fisher’s LSD post-test. Different letters denote statistically significant differences between conditions (p < 0.05). Graphic was performed using GraphPad Prism 9.5.1 (https://www.graphpad.com).
Figure 6.
Effect of the THC-free extract, the full-spectrum extract, and THC on Lysophosphatidate phospholipase A1 activity in synaptosomes from Adult and Aged rat cerebral cortex. Incubation conditions were consistent with those outlined for LPAase activity and specified in the Materials and Methods section. Results are presented as nmol of oleic acid per mg of protein per minute and are shown as mean ± standard error of the mean (SEM), with individual data points representing the sample size of each group (n varies across columns). Data were analyzed by two-way ANOVA (overall model: F = 15.94, p = 0.0001), followed by Fisher’s LSD post-test. Different letters denote statistically significant differences between conditions (p < 0.05). Graphic was performed using GraphPad Prism 9.5.1 (https://www.graphpad.com).
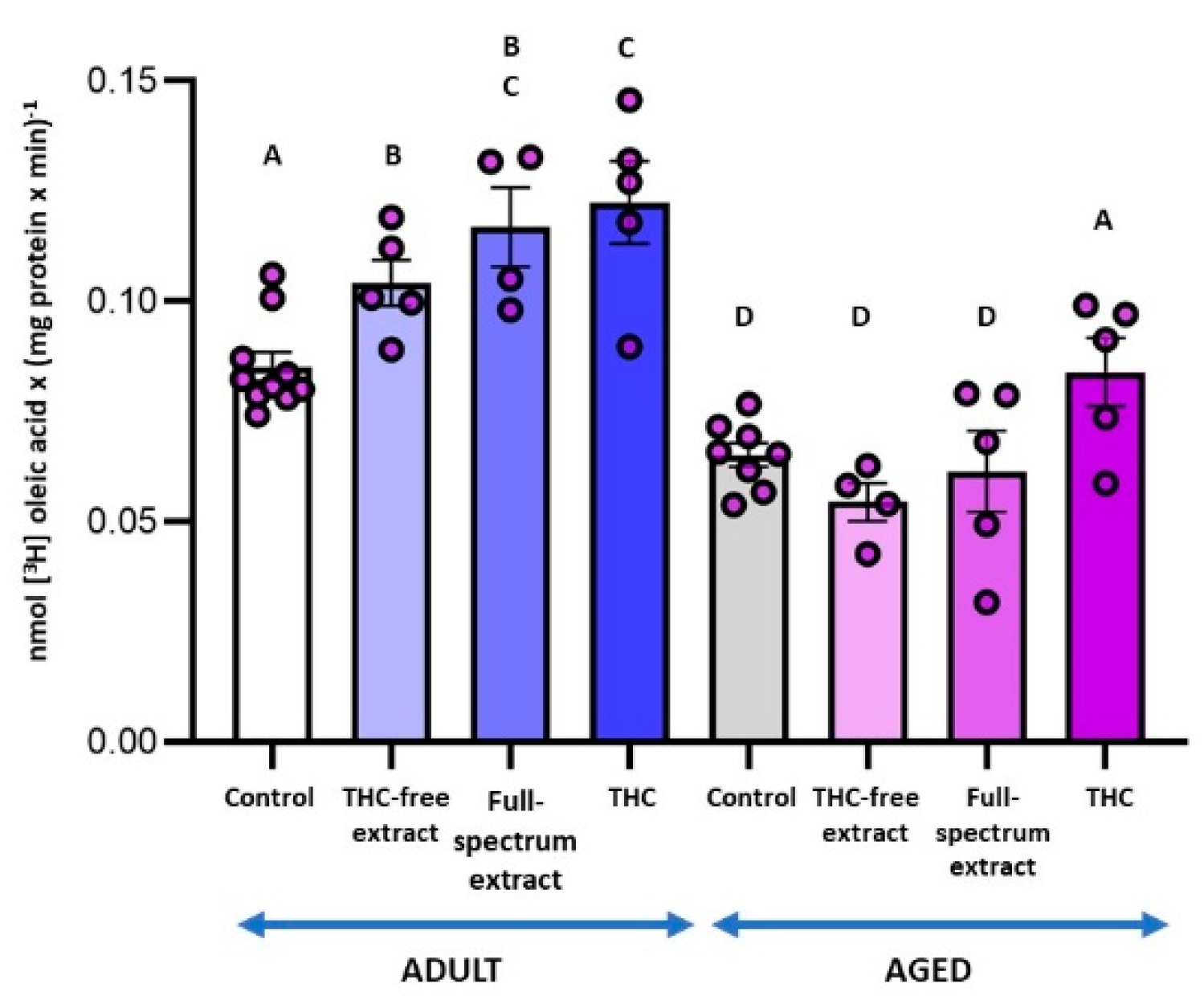
Figure 7.
Differential modulation of 2-AG metabolism in synaptic terminals by THC-free extract, full-spectrum extract, and THC. The relative size of the arrows reflects the magnitude of changes in enzymatic activities related to the synthesis (LPAase and DAGL) and hydrolysis (MAGL) of 2-AG, as well as its synaptic availability in aged synaptic terminals under the experimental conditions, relative to untreated aged synaptic terminals. Graphic was performed using Corel draw 2021.
Figure 7.
Differential modulation of 2-AG metabolism in synaptic terminals by THC-free extract, full-spectrum extract, and THC. The relative size of the arrows reflects the magnitude of changes in enzymatic activities related to the synthesis (LPAase and DAGL) and hydrolysis (MAGL) of 2-AG, as well as its synaptic availability in aged synaptic terminals under the experimental conditions, relative to untreated aged synaptic terminals. Graphic was performed using Corel draw 2021.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



