Submitted:
19 January 2026
Posted:
21 January 2026
You are already at the latest version
Abstract
The Solanaceae family includes some of the most economically and agronomically im-portant crops, such as tomato, potato, pepper and eggplant. Recently, CRISPR/Cas-based genome editing has emerged as a powerful tool for functional genomics and crop improvement, enabling precise and efficient genetic modifications. This review provides an overview of CRISPR/Cas-mediated genome editing technologies and their applications in the major cultivated Solanaceae crops. The use of systems for targeted gene knockout and knock-in approaches is described, together with advances in precision editing strategies such as base editing and prime editing, which allow precise nucleotide substitutions and small sequence changes. The expanding CRISPR toolbox is further explored through alternative Cas proteins, such as Cas12a and Cas13 with distinct targeting features and potential applications. Emerging delivery strategies, including ribonucleoprotein-mediated editing in protoplasts, virus-induced gene editing (VIGE), and de novo induction of meristems, represent promising approaches to generate transgene-free edited plants. In addition, the current status of field trials involving genome-edited Solanaceae crops in Europe is outlined, considering the regulatory landscape and legislative requirements for their release in the environment. Despite regulatory constraints, some ge-nomeedited crops have reached the market, highlighting their potential to contribute to sustainable agriculture and crop improvement.
Keywords:
Solanaceae
; genome editing
; CRISPR/Cas
; protoplasts
; VIGE
; field trials
1. CRISPR/Cas9 Mediated Genome Editing
In the last years genome editing approaches have emerged as a strategy for efficient and targeted genome modifications, especially for crops which have complex genomes and which are difficult to improve through conventional breeding approaches. These techniques allow several kinds of mutations, such as insertions, deletions, replacements, integration of specific sequences of DNA at a desired genomic locus, and site-directed substitutions across the genomes introducing genetic variation precisely and rapidly [1].
Genome editing strategies rely on two elements: sequence-specific nucleases, capable of introducing double-strand-breaks (DSBs) in the genome at or near the site where a DNA sequence modification is desired, and the cell’s DNA repair pathways. These cellular mechanisms can fix the DSBs through either non-homologous end joining (NHEJ) or homology-directed recombination (HDR). NHEJ frequently generates small insertions or deletions (indels) at target sites which can lead to frameshift mutations and ultimately result in loss of gene function. HDR uses a donor template as a source of DNA sequence information to repair the DSB by introducing specific point mutations or inserting/replacing desired sequences into the target site (Figure 1) [2].
The clustered regularly interspaced palindromic repeats (CRISPR)/Cas9 system was discovered in archaea and bacteria as an adaptive defence system that serves as a “genetic memory” of prior invasions and allows to destroy invading viruses and plasmids. The Cas9 protein recognises the target sequence through base pairing between DNA and a short guide RNA (i.e. crRNA interacting with a tracrRNA with structural function) and then cuts the double-stranded DNA, ultimately avoiding the infection (Figure 1) [3]. The transition of the CRISPR/Cas9 system from biological phenomenon to genome engineering tool came through the discovery that the target sequence could be simply reprogrammed by changing 20 nucleotides in the short guide RNA and that its targeting specificity could be combined with the structural properties of the scaffold RNA in a chimeric single guide RNA (gRNA), thus simplifying the engineering system [4]. This way, Cas9 nuclease activity can be supposedly directed to any DNA sequence of the form 5’-N20-NGG-3’, with NGG being the specific protospacer adjacent motif (PAM) recognised by the most widely used Cas9 from Streptococcus pyogenes (SpyCas9).
This system is very versatile and simple to adapt to different purposes, making CRISPR/Cas9 the elected strategy for plant genome editing. Indeed, after its discovery this technique was soon applied to bacteria, yeasts, human cell lines and several model animals, such as fruit flies, nematodes, zebrafishes, mice and rats. Simultaneously, CRISPR experiments have been carried out on plants: it demonstrated its efficacy at first in model plants such as Arabidopsis thaliana and Nicotiana benthamiana [5], then in crop plants including rice, wheat, sorghum, maize, soybean, tomato, sweet orange, grape, tobacco and cotton [6]. Since Solanaceae family includes many of the world’s most important agricultural species (potatoes, tomatoes, eggplants and peppers) and some crop model plants for research (e.g. tomato and tobacco), it is not surprising that several examples of CRISPR/Cas9 applications involved these species. While efficient protocols have been well-established for in vitro regeneration and genetic transformation in tomato and potato, eggplant and bell pepper are still considered as recalcitrant species and are more difficult to be handled. This explains the low number of examples in literature found for CRISPR applications in these two species, if compared to tomato and potato.
1.1. CRISPR/Cas9 for Gene Knockout
The first reports demonstrating the feasibility of gene editing in Solanaceae species regarded tomato (Solanum lycopersicum) and potato (Solanum tuberosum), by knocking out SlAGO7 [7] and StALS [8] genes respectively, both of them conferring recognisable phenotypes to the edited mutants. Later on, many other applications of CRISPR/Cas9-mediated knockout have been reported in Solanaceae crops to improve their performance despite environmental stresses: enhancing disease tolerance through the knockout of either susceptibility genes in tomato [9,10,11,12,13,14] and potato [15,16,17,18,19] or other genes in potato [20,21,22]; reducing the impact of viral diseases in tomato [23,24,25] and potato [26,27]; improving abiotic stress tolerance in tomato [10,28,29,30] and potato [16]. Also genes regulating fruit quality have been targeted in tomato through CRISPR/Cas9 to improve fruit ripening, texture and colouration and the content of bioactive compounds [31,32,33,34,35]. In eggplant (Solanum melongena) DMR6-1 has been knocked out conferring tolerance to Phytophthora spp. [36], and also PPO genes have been disabled to reduce fruit browning after cutting [37] and to highlight pleiotropy in this gene family [38]. Potato tuber quality enhancement has been achieved through CRISPR/Cas9 mediated genome editing of genes involved in starch quality [39], tuber browning [40], steroidal glycoalkaloids biosynthesis [41] and healthy compounds content [42,43,44]. Despite its recalcitrance to genetic transformation and in vitro regeneration, first results have been recently obtained in pepper (Capsicum annuum) by knocking out the functionality of CaAGO7 and CaPPO, conferring respectively a visible curly phenotype or a measurable reduction of enzymatic activity [45], and CaPDS in a hot pepper accession [46].
Considering that domesticated crops display limited genetic diversity, emerging environmental changes and increasing stresses create an urgent need for novel crops that can better stand these conditions. However, it is still challenging to exploit wild species for conventional breeding programs to introgress elite traits in cultivated crops. Multiplexed genome editing approaches (i.e. targeting multiple genes in a single editing event) on genes involved in domestication processes demonstrated the possibility of “reproducing” the genetics events underlying this process and exploiting the wild alleles for crop improvement [47]. Remarkable results have been achieved in tomato by improving the plant architecture, fruit yield and stress resistance thanks to de novo domestication [48,49,50], and similar results were also achieved in another member of Solanaceae family, groundcherry (Physalis pruinosa) [51].
Another interesting application of CRISPR/Cas9 technology aims at targeting cis-regulatory elements (CREs) to change the expression of target genes and fine-tune regulatory networks to improve key traits [52]. Fruit size, inflorescence branching, and plant architecture have been likewise addressed in tomato resulting in bigger fruits with a higher number of locules, more branched inflorescences and indeterminate growth [53,54].
1.2. CRISPR/Cas9 for Gene Knock-In and Allele Swapping
Compared with the multitude of reports on CRISPR/Cas9-mediated knockout experiments, there are significantly fewer reports on the targeted insertion of DNA segments or allele swapping in plants. This is due to the difficulty in obtaining knock-in events because they require HDR and homologous recombination (HR), which are less prevalent and less efficient in plants if compared to NHEJ [55]. Danilo et al. [56] demonstrated the use of HDR to insert some modifications in tomato SlALS1 gene. An HDR-mediated allele replacement was performed on tomato SlALC gene to obtain a longer shelf-life of the fruit [57], and even a targeted recombination between homologous chromosomes was achieved [58]. HR was also exploited in combination with geminiviral replicons to repair the CRISPR/Cas9 mediated DBS and restore the wild-type allele of two tomato genes involved in the carotenoid biosynthesis pathway [59]. The same technique was used to insert a strong promoter upstream of a gene controlling anthocyanin biosynthesis in tomato, achieving ectopic anthocyanins accumulation in the tissues [60]. Moreover, HDR was used in potato to re-establish resistance to Phytophthora infestans in a susceptible variety by replacing a nonsense mutation in all the four alleles of StCCoAOMT [61].
Even if A. tumefaciens mediated transformation proved to be very effective for knockout of genes, this method seemed less efficient for knock-ins or allele swapping. Therefore, other delivery methods have been exploited for HDR applications [62]. For example, a geminiviral multi-replicon system was deployed in tomato to increase the HDR mediated knock-in efficiency and a salt tolerant allele was obtained by inserting a single aminoacidic substitution in HKT1;2 locus [63]. Similarly, herbicide-inhibiting point mutations within the ALS1 locus have been incorporated in potato also using a geminiviral replicon [64].
2. Base Editing and Prime Editing
Further mutagenesis studies showed that the inactivation of a single nuclease domain converts Cas9 into a nickase (nCas9), whereas simultaneous inactivation of both domains preserves its RNA-guided DNA binding ability while abolishing endonuclease activity, generating the so-called “dead Cas9” (dCas9) [65].
Base editing allows to insert targeted point mutations without DSBs or donor DNA templates. It relies on a nCas9 fused to either an adenine or a cytidine deaminase, leading to an A-to-G (adenine base editor, ABE) or a C-to-T substitution (cytosine base editor, CBE), respectively. The Cas-deaminase complex is guided to the target site by the gRNA and is able to induce a deamination on the non-complementary strand, causing the desired nucleotide substitution (Figure 2A) [66]. This technique has been applied at first in tomato targeting SlDELLA or SlETR1, two genes involved in plant hormone signalling [67], SlALS gene [68], SlAGO7 [69] and three genes important for carotenoid accumulation [70]; a transient system has also been used for this purpose in tomato to perform base editing of SlHWS [71]. A good efficiency was achieved also in potato on StGBSS genes involved in starch synthesis [72,73].
Similarly to base editing, prime editing also uses a nCas9 but fused with a reverse transcriptase. The prime editing guide RNA (pegRNA) includes a sgRNA targeting the specific genomic site, a reverse transcript template (RTT) encoding the desired edit, and a primer binding site (PBS) to initiate the reverse transcription process. The protein complex binds the target DNA and induces a single strand break where the PBS hybridizes to start reverse transcription, and eventually to copy the chosen mutation into the genomic DNA (Figure 2B) [74]. This system expands the possibilities for crop improvement by causing substantially every kind of mutation in the desired target site in the genome: indeed it enables the introduction of indels and all 12 base-to-base conversions (both transitions and transversions). For example, multiple pegRNAs were designed for prime editing of three tomato genes, SlGAI, SLALS2 and SLPDS1, and the desired mutations were obtained in the last two, even if with very low efficiencies [75]. To enhance its efficiency in dicotyledonous plants, prime editing system has been optimised by changing promoters, terminators, nCas and pegRNA variants, including RNA chaperones and using a viral replicon system, obtaining editing efficiencies up to 38% on SlALS gene [76]. Another high-efficiency Csy4-based prime editing system has been developed for dicots, and in particular tomato, in order to knock-in a heat-shock element into the promoter of SlLIN5 gene and obtain a better yield under heat stress conditions [77]. Moreover, a dual prime editing system was demonstrated to facilitate precise deletion, replacement and inversion of large DNA fragments in N. benthamiana and tomato with high editing efficiency [78]. The prime editing technology also proved to allow successful generation of nucleotide transitions and transversions but also short defined indels in potato [79].
3. CRISPR/Cas-Mediated Transcription Activation and Inhibition
The inactivation of both the nuclease domains converts the Cas9 into a generic RNA-guided DNA-binding protein that is catalytically inactive (dCas9). It is then possible to directly fuse effectors to the dCas9, transforming the dCas9-effector fusion into an easily programmable artificial transcription factor when paired with a target-specific gRNA [80]. One type of effector that can be fused to dCas9 is a transcriptional activator that will be guided on the desired genomic locus by the complementarity of gRNA and will bind the regulatory region (promoter or enhancer) thus upregulating the expression of the target gene (CRISPRa) (Figure 3A) [81]. Even the modification of epigenetic marks can change the level of transcription of a target gene (Figure 3C).
While most functional genomic studies have relied on the CRISPR/Cas mediated induction of loss-of-function mutations, gain-of-function approaches offer unique insights [82]. In fact, some examples of applications of CRISPRa have been reported in particular in tomato to obtain upregulation of target genes.
The catalytic domain of the tomato SlATX1 gene (a histone methyltransferase) was fused to the dCas9 and targeted to the promoter region of the SlPR-1 gene. Epigenetically edited tomato plants showed enhanced SlPR-1 transcription that increased biotic stress tolerance and resistance to the pathogenic bacteria Clavibacter michiganensis subsp. michiganensis [83]. A similar approach has been adopted to target the SlPAL2 promoter with the same SlATX1 domain fused to a dCas12; the resulting epigenetic mark caused an upregulation of SlPAL2 expression in tomato, thus enhancing lignin accumulation and conferring increased resistance to C. michiganensis subsp. michiganensis without significant pleiotropic effects [84]. Likewise, epigenetic activation through CRISPRa of SlWRKY29 gene was found to be linked to changes in chromatic state and somatic embryo induction in tomato, useful to develop a protocol for indirect somatic embryogenesis from cotyledonary explants [85].
A stress-inducible gene regulation has been developed in tomato by fusing the transmembrane domain of a membrane-bound NAC transcription factor with dCas9, so that the complex is sequestered to the plasma membrane under normal conditions and is released in response to heat induction. This way tomato immunity against bacterial speck disease was enhanced under elevated temperatures by activating SlCBP60g and SlSARD1 genes and also heat-stress tolerance has been engineered through activation of heat-responsive transcription factors, SlNAC2 and SlHSFA6b [86].
Alternative activation strategies have been developed, such as CRISPR–Act3.0 that proved to be able to recruit additional activators based on the SunTag system and increase activation efficiency of endogenous genes by 240-fold in tomato [87]. MoonTag, another new activator able to recruit the VP64 activation domain, was tested in tomato hairy roots and proved to be more efficient than the older SunTag system in stably activating the expression of the luciferase reporter gene by binding to its synthetic promoter [88].
Another possible application of CRISPR/Cas systems consists in fusing a synthetic transcriptional repressor domain to a dCas9 to induce target gene silencing (Figure 3B). This strategy known as CRISPR interference system (CRISPRi) offers several advantages over previous forms of gene repression, in particular RNA interference (RNAi) because it does not interfere with the miRNA pathway and has simpler design rules that simplify targeting multiple genes with high levels of knockdown an low off-target effects [80]. Different repressor domains have been tested in plants [89] and an example in potato demonstrated that fusing dCas9 to a KRAB (Krupple associated box) domain, suppression of the StSGT1 gene expression through CRISPRi approach produced plants with altered content of steroidal glycoalkoids, and in particular low α-solanine and unchanged α-chaconine levels [90].
4. Alternative Cas Proteins
One of the major limitations of CRISPR/Cas9 systems is the compulsory necessity of a PAM sequence close to the edit site that restricts the target regions to sequences with high GC content (“NGG” for SpyCas9) [91]. Even if some efforts have been done to engineer near-PAMless Cas9 variants [92], only very few applications have been reported in plants. For example, a SpCas9-NG variant that recognizes a NGN PAM has been engineered and proved to be efficient first in model plant Physcomitrella patens and then also in tomato and potato for both classical CRISPR-generated gene knock-out and cytosine base editing [93].
Exploring the extreme diversity of CRISPR loci [94], many other Cas proteins have been identified with different nuclease activity and different PAM specificity. Among other CRISPR systems, Cas12a (originally identified as Cpf1) may offer a good alternative to Cas9 [95] since it requires “T” rich PAMs (e.g. TTTV), thus expanding the possibilities for the target region choice. It also generates staggered ended cuts that may promote site-directed integration events through HDR (Figure 4B) [96]. Cas12a variants with altered PAM specificities have also been engineered [97,98]. The efficiency of Cas12a has been tested in tomato protoplasts, and resulted similar to Cas9 but with a higher probability of more and larger deletions induced [99]. Vu et al. [63] also proved the efficiency of CRISPR/Cas12a mediated genome editing in tomato inserting ANT1 gene sequence through HDR in order to obtain visually recognisable anthocyanin rich tomatoes; the same technique was then applied to SlHKT1;2 gene and a salt tolerant line was obtained [63]. Further optimisation of the gene targeting system have been performed by using small chemical molecules to inhibit the NHEJ pathway, treating with chemical such as silver nitrate and selecting high performance and thermotolerant Cas12a, like the one isolated from Lachnospiraceae bacterium [100].
Another Cas protein that can be engineered for knockdown is Cas13. It has RNase activity and when associated with crRNA it forms an RNA-guided RNA targeting complex that recognises and cleaves ssRNA (Figure 4C). This system can be used to target both viral RNA and endogenous mRNA [101]. CRISPR/Cas13 mechanism is very similar to RNAi, but its high specificity results in low probability of off-target events and high knockdown efficiency. Transgenic potato lines expressing Cas13a/gRNA constructs efficiently suppressed Potato virus Y (PVY) accumulation in the plant and reduced disease symptoms [102,103,104]. Moreover, a CRISPR/Cas13 system targeting with multiple crRNAs the genome of potato spindle tuber viroid (PSTVd) has been applied in tomato plants via transient expression and into N. benthamiana through transgenic methods. This resulted in reduced PSTVd accumulation and alleviated disease symptoms [105]. Recently, the knockdown of PDS endogenous transcript was demonstrated in N. benthamiana, A. thaliana and tomato using Cas13a delivered through Agrobacterium tumefaciens infiltration. This study also proved that gene silencing can be induced even in the absence of Cas13 by exploiting only the crRNA with the Argonaute proteins and the plant RNAi machinery [106].
5. RNPs Mediated Genome Editing in Protoplasts
Most commonly CRISPR/Cas reagents are delivered into plants as plasmid DNA constructs through Agrobacterium-mediated transformation or particle bombardment. In both methods, the plasmid DNA is likely to integrate randomly into one or more genomic loci, resulting in stable and prolonged expression of the CRISPR/Cas components in the host genome [107]. To meet the requirements of the current regulation on genetically modified organisms (GMOs) emerging at European level, developing gene-edited lines without the integration of foreign DNA into the host genome is gaining increasing importance. Delivery of pre-assembled ribonucleoprotein complexes (RNPs) into protoplasts through either electroporation, polyethylene glycol (PEG) or nanoparticles is a promising alternative way to obtain DNA-free edited plants (Figure 5). The presence of the cell wall in plant cells is the major challenge in delivering the big sized Cas9 protein along with the negatively charged gRNA, and also in vitro regeneration from edited cells is often difficult and highly species-specific. Nevertheless, some achievements have been made in species within the Solanaceae [108]. Efficient protocols for protoplasts isolation and PEG mediated transfection of RNPs have been established in both the wild tetraploid Solanum peruvianum, targeting genes involved in the small interfering RNAs biogenesis (SpRDR6 and SpSGS3), in the pathogen-related peptide precursors (SpPR-1 and SpProSys) and in fungal tolerance (SpMlo1) [109], and in the cultivated Solanum lycopersicum. In the latter, two genes involved in strigolactone biosynthesis (i.e. SlCCD7 and SlCCD8) were edited, but only calli were regenerated [110]; furthermore, SlSP and SlSP5G genes responsible for the control of growth habit and flowering were targeted in tomato protoplasts from four cultivars (namely Red Setter, Alisa Craig, M82 and Moneymaker) and edited plants were regenerated [111]. Another example of application of this technique has been recorded in tomato protoplasts cv. Heinz 1706, targeting SlPelo gene for tomato yellow leaf curl virus resistance [112]. Similarly, PEG-mediated transfection of potato protoplasts has been optimised and exploited to obtain DNA free edited plants with lower tuber browning due to StPPO2 knockout [40,113], modified starch composition by editing of StGBSS gene [39,114] or genes encoding starch branching enzymes [115], enhanced tolerance to Phytophthora infestans by targeting the susceptibility gene Signal Responsive 4 (SR4) [116]. The state of the art is different for eggplant and sweet pepper, that are highly recalcitrant to in vitro regeneration from protoplasts, thus making it more difficult to apply transformation protocols. However, some results have been achieved in pepper with RNPs mediated transfection of protoplasts: gene editing has been highlighted on two different genes, CaPAD1 or CaMLO2, even if with very low editing efficiency, but no plant regeneration was obtained [117,118,119]. No CRISPR/Cas mediated editing has ever been demonstrated in eggplant protoplasts instead, although a protocol for protoplasts isolation and PEG mediated transfection with vectors encoding fluorescent proteins has been published [120].
Electroporation is another widely employed technique for protoplast transfection with exogenous DNA. A transient electric field applied to the protoplasts induces temporary pores in the cell membrane, allowing the entry of foreign DNA. This method has been mainly exploited for protoplasts fusion [121] and transient gene expression [122], but no applications for gene editing have been reported so far in Solanaceae.
Nanoparticles can also be used as an alternative delivery method for RNPs: for example, metallic nanoparticles coated with CRISPR/Cas reagents have been used for particle bombardment of rice protoplasts [123], but this strategy is used preferably to penetrate plant tissues or embryos more than cell wall free protoplasts and is used more frequently in monocotyledonous species. It has been demonstrated that addition of anionic polymer polyglutamic acid to standard PEG transfection protocols significantly improved editing efficiencies in Nicotiana benthamiana protoplasts relative to RNPs alone.
Lipofection seems to be a new promising alternative that allows to deliver RNP complexes into naked plant cells thanks to cationic lipids that facilitate the interaction between the membranes and RNPs thus allowing the uptake of the exogenous material [124]. An example of lipofection-mediated transfection approach for delivery of Cas9/gRNA RNPs into tobacco (Nicotiana tabacum) protoplasts has been described by Liu et al. [125] with transfection efficiency reaching up to 66% and targeted mutagenesis frequency up to 6%.
6. Virus Induced Gene Editing
Since in vitro regeneration is highly species-specific and sometimes even cultivar-specific [126] and it represents one of the major bottlenecks in plant transformation, alternative strategies have been developed to avoid this step.
Virus-induced gene editing (VIGE) is a promising technique in this perspective. It uses viruses as transient delivery vehicles for CRISPR/Cas components and takes advantage of their ability to replicate inside the plant cells to increase accumulation of Cas protein and gRNA within the host cell, ultimately resulting in a higher editing efficiency. To this end, viral genomes are engineered to produce viral vectors encoding CRISPR components able to spread in the plant. VIGE vectors can be divided into two categories according to their cargo capacity and the reagents that may be delivered (Cas9 is a large protein, so it is difficult to fit its long coding sequence in the small viral capsid). The first category includes viral vectors expressing Cas9 and gRNAs that are used to infect wild-type plants. Somatic tissues are edited as the viral infection spreads systemically through the plant. Following tissue culture, infected leaves can be used to regenerate transgene-free edited progeny (Figure 6A). Otherwise, if the infection reaches the meristem, the resulting seeds may also be edited, achieving transgenerational heritability (Figure 6B). In the second category fall viral vectors expressing gRNAs fused to RNA mobility signals, such as Flowering Locus T (FT) mRNA or transfer RNAs (tRNAs). These are infiltrated into Cas9-overexpressing plants where mobile gRNAs spread systemically possibly entering also the shoot apical meristem in order to induce editing in the germline. Collected seeds provide edited progeny bypassing tissue culture procedures, but the Cas9 transgene needs to be segregated through backcrossing with wild-type plants or self-pollination cycles (Figure 6C) [127,128]. Viral constructs are usually delivered to the plant through A. tumefaciens previously transformed with viral plasmids; agroinoculation is usually done through leaf syringe infiltration, but other procedures can also be used, such as vacuum infiltration, agrospray or agrodrench [127]. The delivery of CRISPR reagents via VIGE was first attempted in N. benthamiana using a viral vector based on Tobacco Rattle Virus (TRV) [129,130], but many improvements have been implemented to optimise the virus host range, tissue specificity, cargo capacity and efficiency of the system.
Examples of VIGE application in Solanaceous crops have been reported, mainly using Tobacco Rattle Virus (TRV), Potato Virus X (PVX) or Tomato Spotted Wilt Virus (TSWV). Lee et al. [131] were able to knockout SlPDS gene infecting tomato with TRV or PVX and regenerate edited plants by subsequent in vitro culture, and they also demonstrated the applicability of the system in potato and eggplant. By fusing the gRNA targeting PDS to the sequence of tomato FT, efficient and rapid generation of heritable genome editing was achieved in tomato [132]. Kang et al. [133] showed that low temperatures significantly enhanced VIGE efficiency of TRV and PVX vectors in both cotyledons and systemic upper leaves of tomato and Oh et al. [134] highlighted the importance of the promoter driving the expression of Cas9 in order to increase the efficiency of the TRV system and its trans-generational heritability in tomato. PVX has also been efficiently engineered to apply VIGE in tomato knocking out SlPDS and SlSGR1 genes [135]. Other functional genes involved in the anthocyanins biosynthesis pathway (namely SlCHS1, SlCHI and SlDFR) have been successfully edited in tomato leaves through TRV-mediated VIGE and the same technique was also applied on unripen fruits to edit SlEIL3, a positive regulator of the ethylene signalling pathway during tomato ripening, thus delaying the ripening in the infiltrated sectors [136]. In addition, TSWV, that has a larger cargo capacity compared to TRV and PVX, has been engineered to deliver CRISPR/Cas9, CRISPR/Cas12a and base editing systems for knockout of different functional genes in N. benthamiana; the same system has also been efficiently applied to tomato, chili pepper, sweet pepper and habanero pepper [137]. To improve gRNA mobility in the infected plant, a cleaving hammerhead ribozyme (HH) sequence was added to the TRV2-gRNA construct along with a FT mobile RNA or other mobile tRNAs. The editing on CaPDS and CaFA was successful in Cas9-transgenic pepper lines [138]. TSWV was also used in sweet pepper to edit PDS gene via VIGE and symptomatic leaf tissues from infected plants were used as explants for tissue culture in order to regenerate pepper plants with targeted mutations, that also demonstrated to be germline transmissible [139].
7. De Novo Induction of Meristems
Another recently developed technique allows to simplify or completely avoid tissue culture by reprogramming genome-edited somatic cells into meristems through co-expressing developmental regulators (DRs), such as WUS, IPT and STM, and genome editing components, thus enabling direct regeneration of genome-edited plants from somatic cells [140] without requiring dedifferentiation into callus-like structures and then differentiation into shoots and roots. This method enables the creation of either transgenic or gene-edited shoots from de novo induced meristems that proved to be able to produce flowers and seeds, ultimately transmitting transgenes or gene edits to the next generation. Since different combinations of developmental regulators could induce the formation of meristems in different dicot species, the most suitable combination of DRs was first optimised in N. benthamiana and then in tomato by delivering constructs expressing different DRs into young seedlings through A. tumefaciens coculture. The possibility of generating gene-edited meristems was also demonstrated transforming seedlings constitutively expressing Cas9 with a vector containing DRs and a gRNA targeting PDS gene [141]. The same constructs were cloned into a T-DNA backbone producing geminiviral replicons derived from bean yellow dwarf virus (BeYDV). These replicons are able to replicate after delivery to plant cells, increasing copy number and leading to high levels of gene expression, even without integration in the genome, potentially generating transgene-free edited lines. Maher et al. [141] also demonstrated that genetically modified meristems could be induced on soil-grown Cas9-overexpressing plants by pruning to remove axillary meristems and perfusing the cutting sites with a suspension of A. tumefaciens cultures expressing DRs and gRNA cassettes. Gene-edited plants were obtained directly from the resulting shoots, and the induced mutations were inheritable (Figure 7). This protocol has been applied also in tomato and potato providing a generalizable in planta delivering method that halves the time required for generating gene-edited plants [142]. Similarly, RNA viral vectors expressing gRNAs were delivered to leaves or sites near axillary meristems to induce new edited shoots to form from somatic cells (Figure 7). To enhance the induction of shoots, RNA viral vectors were delivered along with the cytokinin biosynthesis gene IPT: this system provided abundant, phenotypically normal, gene-edited shoots from soil-grown tomato plants [143]. This system overcomes the bottleneck in applying VIGE to dicotyledonous crops and reduces the dependency on tissue culture, which can be inefficient in recalcitrant species or cultivars.
Another similar approach to induce transgenic and gene-edited de novo meristems exploits a synthetic cascade made up by a wound-induced transcriptional regulator (WIND1) that triggers the expression of DR genes along with gene-editing reagents. This system was applied in planta to non-meristematic internodes of N. benthamiana to regenerate de novo shoots with mutations in PDS gene and IPT proved to be the most effective DR among those tested for regeneration. The same synthetic toolkit was successfully applied to both tomato and soybean [144]. Additional DRs have been tested at first in snapdragons (Antirrhinum majus) and then validated in tomato, highlighting that PLT5 provided the highest in planta transformation efficiency [145]. The same DR significantly improved the formation of transgenic calli and somatic embryos in sweet pepper through in vitro tissue culture.
Moreover, an Agrobacterium-mediated transient expression technology has been applied in tomato to deliver nCas fused to a cytidine deaminase and a viral vector harbouring a gRNA targeting WS gene, involved in the regulation of miRNA expression and plant growth. After agroinfiltration of these reagents, homozygous base edited plants were obtained and a more than 70% were Cas-free, demonstrating the efficacy of a transient system for base editing that combines different approaches [146].
8. Research and Field Trials for CRISPRed Plants in Europe
CRISPR technology has revolutionized how and how fast improved crops can be produced, allowing precise modifications in the plants’ genome to control desirable traits. These new traits may help address global agricultural challenges, such as pressures of climate change, food security issues and also producing plant material that meets industrial requirements. Scientific research and laboratory experiments are the starting point to produce new improved varieties, but the plants’ behaviour in controlled environment must be validated in field conditions to ensure that the desired phenotype is maintained also in a context with multiple stimuli and stresses.
The cultivation of GMOs in the EU was first regulated by the 2001/18/EC Directive [147] that requires a strict risk assessment procedure before authorisation and establishes traceability and labelling criteria for GM products. By the way, New Genomic Techniques (NGTs), mainly based on genome editing strategies such as CRISPR/Cas and cisgenesis, have been developed after 2001 thus making the current legislation outdated. Considering that in principle these techniques produce genomic modifications comparable with those achieved through conventional breeding or classical mutagenesis with no foreign DNA introduction in the genomes, NGT products could be suitable for an exemption from the GMO Directive [148]. In this view, a new regulatory framework on plants obtained through NGTs has been proposed in July 2023 by the European Commission [149]. This proposal distinguishes between NGT category 1 plants, considered equivalent to those produced by conventional breeding or occurred naturally and therefore out of the GMO regulation, and all other GM plants, included in NGT category 2 and falling under the 2001 Directive. In February 2024, the European Parliament voted in favour of this proposal and in March 2025 also the European Council supported it with slight changes. After the trilogue phase, the 4th of December 2025 the Council has reached a provisional agreement with the European Parliament on a set of rules that establish a legal framework for new genomic techniques (NGTs) [150,151].
Nevertheless, the experimental release of GM plant is harmonized across the EU through part B of the Directive 2001/18/EC. Following article 6.1 of the Directive [147], anyone planning to release GMOs in the environment for experimental purposes must notify the competent authority of the member state in which the release is planned. The notification must contain a technical dossier and an environmental risk assessment and is processed by the authority before giving permission or not for the field trial [152]. This procedure delegates to single EU states the evaluation and authorisation of field trial requests.
A very different approach has been adopted in the UK after Brexit, with the Precision Breeding Act in 2023 [153] and following Precision Breeding Regulations [154] that entered into full force in November 2025. These regulations introduce the Precision Bred Organisms (PBOs) category that, according to the technical guidance published by the Advisory Committee on Environmental Release (ACRE) [155], includes plants with epigenetic changes, NGT products, and some cisgenic events as long as functional transgenes, such as selectable markers, have been removed (for example, by segregation) and genetic changes introduced could have resulted from traditional processes and selection. Interestingly, in this regulation the decisive point is the nature of the change in the genome, rather than the technique that was used [152], and PBOs can follow simplified procedures for field release after being included in a public national register.
Despite a great number of genome-edited crop plants have already been produced, as reported in peer-reviewed scientific publications and as summarised in the publicly accessible online database managed by the European Sustainable Agriculture Through Genome Editing (EU-SAGE) (https://www.eu-sage.eu/genome-search), in this regulatory landscape only a few examples of NGT plants have been brought to the field for experimentations.
Focusing on Solanaceae, 3 and 12 field trials respectively have been authorised respectively for S. lycopersicum and S. tuberosum in different European countries (Table 1). These events mainly focus on the improvement of crops for their tolerance to biotic and abiotic stresses, for their nutritional value and for their content of industrially useful molecules. In particular, tomato dmr6-1 mutants [10] with enhanced tolerance to drought stress and Phytophthora infestans infection, and d27 and ccd7 mutants with higher resistance to broomrapes infestation [156] have been accepted for field trials in Italy, while tomatoes engineered for the accumulation of provitamin D3 by genome editing of Sl7-DR2 gene [35] are cultivated for field trials in UK and even a clinical trial to test the impact of vitamin D biofortified tomatoes on serum levels of 25(OH)D is ongoing in the UK (https://www.clinicaltrials.gov/study/NCT07142759). Many more NGT events on potato have been accepted for field trials in northern Europe, particularly dealing with tolerance to late blight, general resistance to pathogens, altered starch composition for industrial purposes and content of antinutritional compounds like glycoalkaloids.
9. Conclusions and Perspectives
The advent of NGTs, and in particular CRISPR/Cas technology, has radically changed crop improvement both by making precise modifications possible in desired genetic loci and by speeding up the time required for generating novel improved varieties.
CRISPR/Cas9 technique (Figure 1) has been the first to be developed and is the most used one to produce targeted knockouts in genes linked to desired phenotypes; further optimisations allowed to produce also knock-ins and allele replacements, even if with a significantly lower efficiency. The arsenal of CRISPR techniques has been expanded with the advent of base editing and prime editing (Figure 2), both exploiting mutagenized Cas9 nickases fused with adenine/cytidine deaminases or a reverse transcriptase, respectively. These methods allow base-to-base substitutions and substantially every kind of mutation, although still with lower efficiency. By fusing a dCas9 with activator or repressor domains, is also possible to activate or inhibit the expression of target genes in a precise manner (Figure 3). Other Cas proteins, like Cas12 and Cas13a (Figure 4), have also been isolated and used for gene editing and RNA editing.
All the above-mentioned techniques usually involve a delivery system that leaves transgenes integrated in the recipient genome, which is undesirable in a transgene-free perspective. Therefore, delivery of RNPs into protoplasts can be a valid alternative guaranteeing the editing without integration of any foreign DNA in the genome (Figure 5). The downside of this technique is that regenerating in vitro from a single naked cell is usually difficult and highly genotype dependant, thus VIGE can provide a valid alternative by boosting the editing signal through viral vectors agroinfiltrated into somatic tissues (Figure 6). Ultimately, de novo induction of meristems allows to spread the editing in soil-grown plants through co-expression of CRISPR reagents with developmental regulator genes (Figure 7).
The ultimate objective of crop improvement programs is to reach the market with new crop cultivars that can better respond to the challenges that agriculture has to face. However, the regulatory scenario on GM crops varies widely across countries, ranging from relatively permissive frameworks, such as those adopted in the United States, Australia, and Japan—where certain gene-edited plants are exempt from GMO regulation—to the stringent regulatory regimes enforced in New Zealand and the EU. This is the reason why even if a considerable number of gene-edited crops have been produced worldwide, only a few have been approved and even less have made it to the market so far.
The first CRISPR-edited food ever reaching the consumers in 2021 is the ‘Sicilian Rouge’ tomatoes, which contain high amounts of γ-aminobutyric acid (GABA). They have been developed by Sanatech Seed Co., Ltd, a startup from the University of Tsukuba (Japan), by targeting the autoinhibitory domain of SlGAD3 gene, whose removal increases the activity of the glutamic acid decarboxylase enzyme that catalyses the decarboxylation of glutamate to GABA [157]. These tomatoes are claimed to have a five- to sixfold increase in GABA content (0.95 mg per gram of fresh weight), thus conferring a health-promoting effect by lowering blood pressure and promoting relaxation through oral intake of GABA, even if no clear evidence is available on this yet [158].
Besides, another CRISPRed product has already been marketed in the USA since 2023 by the biotech company Pairwise, that produced less pungent mustard greens by intervening in the metabolism of the glucosinolates, responsible for the bitter and pungent taste [159]. Furthermore, other genome-edited crops have been approved in different countries but are still waiting for commercialisation: for example, the waxy corn from Corteva Agriscience with high starch content approved in Canada, the non-browning romaine lettuce by Interexon approved in the US, wheat varieties resistant to powdery mildew developed by the Chinese Academy of Sciences approved in China and non-browning bananas by Tropic Biosciences approved in the Philippines.
Of course the hope is that more and more CRISPRed foods will get the approval, reach the fields and ultimately the market to help fighting the challenges that global agriculture has to face. Alongside, the regulatory frameworks need to evolve, particularly in the countries with a more stringent regulation, in order to adapt to the NGTs and their products.
Author Contributions
Writing—original draft preparation, M.F.; writing—review and editing, M.F., A.A. and A.M.; supervision, A.M. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| CRISPR | Clustered Regularly Interspaced Short Palindromic Repeats |
| DNA | Deoxyribonucleic Acid |
| RNA | Ribonucleic acid |
| Cas | CRISPR associated |
| PAM | Protospacer adjacent motif |
| GMO | Genetically modified organism |
References
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. ‘CRISPR/Cas Genome Editing and Precision Plant Breeding in Agriculture’. Annu. Rev. Plant Biol. 2019, vol. 70(no. 1), 667–697. [Google Scholar] [CrossRef]
- Razzaq. ‘Modern Trends in Plant Genome Editing: An Inclusive Review of the CRISPR/Cas9 Toolbox’. IJMS 2019, vol. 20(no. 16), 4045. [Google Scholar] [CrossRef]
- Doudna, J. A.; Charpentier, E. ‘The new frontier of genome engineering with CRISPR-Cas9’. Science 2014, vol. 346(no. 6213), 1258096. [Google Scholar] [CrossRef]
- Bortesi, L.; Fischer, R. ‘The CRISPR/Cas9 system for plant genome editing and beyond’. Biotechnology Advances 2015, vol. 33(no. 1), 41–52. [Google Scholar] [CrossRef]
- Li, J.-F. ‘Multiplex and homologous recombination–mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9’. Nat Biotechnol 2013, vol. 31(no. 8), 688–691. [Google Scholar] [CrossRef]
- Jaganathan, D.; Ramasamy, K.; Sellamuthu, G.; Jayabalan, S.; Venkataraman, G. ‘CRISPR for Crop Improvement: An Update Review’. Front. Plant Sci. 2018, vol. 9, 985. [Google Scholar] [CrossRef]
- Brooks, C.; Nekrasov, V.; Lippman, Z. B.; Van Eck, J. ‘Efficient Gene Editing in Tomato in the First Generation Using the Clustered Regularly Interspaced Short Palindromic Repeats/CRISPR-Associated9 System’. PLANT PHYSIOLOGY 2014, vol. 166(no. 3), 1292–1297. [Google Scholar] [CrossRef] [PubMed]
- Butler, N. M.; Atkins, P. A.; Voytas, D. F.; Douches, D. S. ‘Generation and Inheritance of Targeted Mutations in Potato (Solanum tuberosum L.) Using the CRISPR/Cas System’. PLoS ONE 2015, vol. 10(no. 12), e0144591. [Google Scholar] [CrossRef]
- Li, R. ‘CRISPR/Cas9-Based Knock-Out of the PMR4 Gene Reduces Susceptibility to Late Blight in Two Tomato Cultivars’. IJMS 2022, vol. 23(no. 23), 14542. [Google Scholar] [CrossRef] [PubMed]
- Maioli. ‘Knock-out of SlDMR6-1 in tomato promotes a drought-avoidance strategy and increases tolerance to Late Blight’. Plant Stress 2024, 100541. [Google Scholar] [CrossRef]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. ‘Rapid generation of a transgene-free powdery mildew resistant tomato by genome deletion’. Sci Rep 2017, vol. 7(no. 1), 482. [Google Scholar] [CrossRef]
- Santillán Martínez, M. I. ‘CRISPR/Cas9-targeted mutagenesis of the tomato susceptibility gene PMR4 for resistance against powdery mildew’. BMC Plant Biol 2020, vol. 20(no. 1), 284. [Google Scholar] [CrossRef]
- Thomazella, D. P. de T. ‘Loss of function of a DMR6 ortholog in tomato confers broad-spectrum disease resistance’. Proceedings of the National Academy of Sciences 2021, vol. 118(no. 27), e2026152118. [Google Scholar] [CrossRef] [PubMed]
- Li, R. ‘Less is more: CRISPR/Cas9-based mutations in DND1 gene enhance tomato resistance to powdery mildew with low fitness costs’. BMC Plant Biol 2024, vol. 24(no. 1), 763. [Google Scholar] [CrossRef]
- Bi, W. CRISPR /Cas9-guided editing of a novel susceptibility gene in potato improves Phytophthora resistance without growth penalty’. Plant Biotechnology Journal 2024, vol. 22(no. 1), 4–6. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, M. ‘CRISPR/Cas9 genome editing of potato StDMR6-1 results in plants less affected by different stress conditions’. Horticulture Research 2024, p. uhae130. [Google Scholar] [CrossRef] [PubMed]
- Kieu, N. P.; Lenman, M.; Wang, E. S.; Petersen, B. L.; Andreasson, E. ‘Mutations introduced in susceptibility genes through CRISPR/Cas9 genome editing confer increased late blight resistance in potatoes’. Sci Rep 2021, vol. 11(no. 1), 4487. [Google Scholar] [CrossRef]
- Sun, K. ‘Silencing of six susceptibility genes results in potato late blight resistance’. Transgenic Res 2016, vol. 25(no. 5), 731–742. [Google Scholar] [CrossRef]
- Sun, K. ‘Silencing susceptibility genes in potato hinders primary infection with Phytophthora infestans at different stages’. Horticulture Research 2022, vol. 9, uhab058. [Google Scholar] [CrossRef]
- Bi, W. ‘Potato DMP2 positively regulates plant immunity by modulating endoplasmic reticulum homeostasis’. JIPB 2025, vol. 67(no. 6), 1568–1581. [Google Scholar] [CrossRef]
- Razzaq, H. A.; Ijaz, S.; Haq, I. U.; Khan, I. A. ‘Functional inhibition of the StERF3 gene by dual targeting through CRISPR/Cas9 enhances resistance to the late blight disease in Solanum tuberosum L.’. Mol Biol Rep 2022, vol. 49(no. 12), 11675–11684. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, M. ‘Genome editing of NPR3 confers potato resistance to Candidatus Liberibacter spp.’. Plant Biotechnology Journal 2024, vol. 22(no. 9), 2635–2637. [Google Scholar] [CrossRef]
- Kuroiwa, K.; Thenault, C.; Nogué, F.; Perrot, L.; Mazier, M.; Gallois, J.-L. ‘CRISPR-based knock-out of eIF4E2 in a cherry tomato background successfully recapitulates resistance to pepper veinal mottle virus’. Plant Science 2022, vol. 316, 111160. [Google Scholar] [CrossRef]
- Pramanik, D. ‘CRISPR/Cas9-Mediated Generation of Pathogen-Resistant Tomato against Tomato Yellow Leaf Curl Virus and Powdery Mildew’. IJMS 2021, vol. 22(no. 4), 1878. [Google Scholar] [CrossRef]
- Tashkandi, M.; Ali, Z.; Aljedaani, F.; Shami, A.; Mahfouz, M. M. Engineering resistance against Tomato yellow leaf curl virus via the CRISPR/Cas9 system in tomato’. Plant Signaling & Behavior 2018, vol. 13(no. 10), e1525996. [Google Scholar] [CrossRef]
- Lucioli; Tavazza, R.; Baima, S.; Fatyol, K.; Burgyan, J.; Tavazza, M. ‘CRISPR-Cas9 Targeting of the eIF4E1 Gene Extends the Potato Virus Y Resistance Spectrum of the Solanum tuberosum L. cv. Desirée’. Front. Microbiol. 2022, vol. 13, 873930. [Google Scholar] [CrossRef]
- Noureen; Khan, M. Z.; Amin, I.; Zainab, T.; Mansoor, S. ‘CRISPR/Cas9-Mediated Targeting of Susceptibility Factor eIF4E-Enhanced Resistance Against Potato Virus Y’. Front. Genet. 2022, vol. 13, 922019. [Google Scholar] [CrossRef]
- Hu, Z. ‘The protein kinase CPK28 phosphorylates ascorbate peroxidase and enhances thermotolerance in tomato’. Plant Physiology 2021, vol. 186(no. 2), 1302–1317. [Google Scholar] [CrossRef]
- Kakeshpour, T. ‘CGFS -type glutaredoxin mutations reduce tolerance to multiple abiotic stresses in tomato’. Physiologia Plantarum 2021, vol. 173(no. 3), 1263–1279. [Google Scholar] [CrossRef] [PubMed]
- Li, R. ‘CRISPR/Cas9-Mediated SlNPR1 mutagenesis reduces tomato plant drought tolerance’. BMC Plant Biol 2019, vol. 19(no. 1), 38. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yuan, Y.; Feng, B.; Deng, W. ‘CRISPR/Cas9-mediated gene-editing technology in fruit quality improvement’. Food Quality and Safety 2020, vol. 4(no. 4), 159–166. [Google Scholar] [CrossRef]
- Liu, X. ‘Biofortification of tomatoes with beta-carotene through targeted gene editing’. International Journal of Biological Macromolecules 2025, vol. 327, 147396. [Google Scholar] [CrossRef]
- Yang, Q. ‘CRISPR/cas9 Allows for the Quick Improvement of Tomato Firmness Breeding’. CIMB 2024, vol. 47(no. 1), 9. [Google Scholar] [CrossRef]
- Do, J. H. ‘Development of a Genome-Edited Tomato With High Ascorbate Content During Later Stage of Fruit Ripening Through Mutation of SlAPX4’. Front. Plant Sci. 2022, vol. 13, 836916. [Google Scholar] [CrossRef] [PubMed]
- Li, J. ‘Biofortified tomatoes provide a new route to vitamin D sufficiency’. Nat. Plants 2022, vol. 8(no. 6), 611–616. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, M. ‘Enhancing tolerance to Phytophthora spp. in eggplant through DMR6–1 CRISPR/Cas9 knockout’. Plant Stress 2024, vol. 14, 100691. [Google Scholar] [CrossRef]
- Maioli. Simultaneous CRISPR/Cas9 Editing of Three PPO Genes Reduces Fruit Flesh Browning in Solanum melongena L.’, Front. Plant Sci. 2020, vol. 11. [Google Scholar] [CrossRef]
- Kodackattumannil, P. ‘Hidden pleiotropy of agronomic traits uncovered by CRISPR-Cas9 mutagenesis of the tyrosinase CuA-binding domain of the polyphenol oxidase 2 of eggplant’. Plant Cell Rep 2023, vol. 42(no. 4), 825–828. [Google Scholar] [CrossRef]
- Andersson, M.; Turesson, H.; Nicolia, A.; Fält, A.-S.; Samuelsson, M.; Hofvander, P. ‘Efficient targeted multiallelic mutagenesis in tetraploid potato (Solanum tuberosum) by transient CRISPR-Cas9 expression in protoplasts’. Plant Cell Rep 2017, vol. 36(no. 1), 117–128. [Google Scholar] [CrossRef] [PubMed]
- González, M. N. ‘Reduced Enzymatic Browning in Potato Tubers by Specific Editing of a Polyphenol Oxidase Gene via Ribonucleoprotein Complexes Delivery of the CRISPR/Cas9 System’. Front. Plant Sci. 2020, vol. 10, 1649. [Google Scholar] [CrossRef]
- Zheng, Z.; Ye, G.; Zhou, Y.; Pu, X.; Su, W.; Wang, J. Editing sterol side chain reductase 2 gene ( StSSR2 ) via CRISPR/Cas9 reduces the total steroidal glycoalkaloids in potato’. All Life 2021, vol. 14(no. 1), 401–413. [Google Scholar] [CrossRef]
- Ducreux, L. J. M. ‘Metabolic engineering of high carotenoid potato tubers containing enhanced levels of -carotene and lutein’. Journal of Experimental Botany 2004. [Google Scholar] [CrossRef]
- Nakayasu, M. ‘Generation of α-solanine-free hairy roots of potato by CRISPR/Cas9 mediated genome editing of the St16DOX gene’. Plant Physiology and Biochemistry 2018, vol. 131, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Ye, G.; Zhou, Y.; Pu, X.; Su, W.; Wang, J. Editing sterol side chain reductase 2 gene ( StSSR2 ) via CRISPR/Cas9 reduces the total steroidal glycoalkaloids in potato’. All Life 2021, vol. 14(no. 1), 401–413. [Google Scholar] [CrossRef]
- Kumar, M.; Ayzenshtat, D.; Rather, G. A.; Aisemberg, E.; Goldshmidt, A.; Bocobza, S. ‘Breaking the glass ceiling of stable genetic transformation and gene editing in the popular pepper cv Cayenne’. Journal of Experimental Botany 2025, vol. 76(no. 10), 2688–2699. [Google Scholar] [CrossRef] [PubMed]
- Bulle, M.; Venkatapuram, A. K.; Abbagani, S.; Kirti, P. B. ‘CRISPR/Cas9 based genome editing of Phytoene desaturase (PDS) gene in chilli pepper (Capsicum annuum L.)’. Journal of Genetic Engineering and Biotechnology 2024, vol. 22(no. 2), 100380. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, H.; Li, J. ‘De novo domestication: retrace the history of agriculture to design future crops’. Current Opinion in Biotechnology 2023, vol. 81, 102946. [Google Scholar] [CrossRef] [PubMed]
- Li, T. ‘Domestication of wild tomato is accelerated by genome editing’. Nat Biotechnol 2018, vol. 36(no. 12), 1160–1163. [Google Scholar] [CrossRef]
- Rehman, F.; Gong, H.; Bao, Y.; Zeng, S.; Huang, H.; Wang, Y. ‘CRISPR gene editing of major domestication traits accelerating breeding for Solanaceae crops improvement’. Plant Mol Biol 2022, vol. 108(no. 3), 157–173. [Google Scholar] [CrossRef]
- Zsögön. ‘De novo domestication of wild tomato using genome editing’. Nat Biotechnol 2018, vol. 36(no. 12), 1211–1216. [Google Scholar] [CrossRef]
- Lemmon, Z. H. ‘Rapid improvement of domestication traits in an orphan crop by genome editing’. Nature Plants 2018, vol. 4(no. 10), 766–770. [Google Scholar] [CrossRef]
- Swinnen, G.; Goossens, A.; Pauwels, L. ‘Lessons from Domestication: Targeting Cis-Regulatory Elements for Crop Improvement’. Trends in Plant Science 2016, vol. 21(no. 6), 506–515. [Google Scholar] [CrossRef]
- Rodríguez-Leal, D.; Lemmon, Z. H.; Man, J.; Bartlett, M. E.; Lippman, Z. B. ‘Engineering Quantitative Trait Variation for Crop Improvement by Genome Editing’. Cell 2017, vol. 171(no. 2), 470–480.e8. [Google Scholar] [CrossRef]
- Li, Q.; Feng, Q.; Snouffer, A.; Zhang, B.; Rodríguez, G. R.; Van Der Knaap, E. ‘Increasing Fruit Weight by Editing a Cis-Regulatory Element in Tomato KLUH Promoter Using CRISPR/Cas9’. Front. Plant Sci. 2022, vol. 13, 879642. [Google Scholar] [CrossRef]
- Steinert, J.; Schiml, S.; Puchta, H. ‘Homology-based double-strand break-induced genome engineering in plants’. Plant Cell Rep 2016, vol. 35(no. 7), 1429–1438. [Google Scholar] [CrossRef] [PubMed]
- Danilo; Perrot, L.; Mara, K.; Botton, E.; Nogué, F.; Mazier, M. ‘Efficient and transgene-free gene targeting using Agrobacterium-mediated delivery of the CRISPR/Cas9 system in tomato’. Plant Cell Rep 2019, vol. 38(no. 4), 459–462. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q. ‘CRISPR/Cas9-induced Targeted Mutagenesis and Gene Replacement to Generate Long-shelf Life Tomato Lines’. Sci Rep 2017, vol. 7(no. 1), 11874. [Google Scholar] [CrossRef]
- Hayut, S. Filler; Bessudo, C. Melamed; Levy, A. A. ‘Targeted recombination between homologous chromosomes for precise breeding in tomato’. Nat Commun 2017, vol. 8(no. 1), 15605. [Google Scholar] [CrossRef]
- Dahan-Meir, T. ‘Efficient in planta gene targeting in tomato using geminiviral replicons and the CRISPR /Cas9 system’. The Plant Journal 2018, vol. 95(no. 1), 5–16. [Google Scholar] [CrossRef]
- Čermák, T.; Baltes, N. J.; Čegan, R.; Zhang, Y.; Voytas, D. F. ‘High-frequency, precise modification of the tomato genome’. Genome Biol 2015, vol. 16(no. 1), 232. [Google Scholar] [CrossRef] [PubMed]
- Hegde, N.; Joshi, S.; Soni, N.; Kushalappa, A. C. ‘The caffeoyl-CoA O-methyltransferase gene SNP replacement in Russet Burbank potato variety enhances late blight resistance through cell wall reinforcement’. Plant Cell Rep 2021, vol. 40(no. 1), 237–254. [Google Scholar] [CrossRef] [PubMed]
- J. Van Eck; Cornell University; USA; and The Boyce Thompson Institute. ‘Genome editing of tomatoes and other Solanaceae’. In Burleigh Dodds Series in Agricultural Science; Cornell University, USA, Willmann, M. R., Eds.; Burleigh Dodds Science Publishing, 2021; pp. 421–440. [Google Scholar] [CrossRef]
- Vu, T. V. ‘Highly efficient homology-directed repair using CRISPR/Cpf1-geminiviral replicon in tomato’. Plant Biotechnology Journal 2020, vol. 18(no. 10), 2133–2143. [Google Scholar] [CrossRef] [PubMed]
- Butler, N. M.; Baltes, N. J.; Voytas, D. F.; Douches, D. S. ‘Geminivirus-Mediated Genome Editing in Potato (Solanum tuberosum L.) Using Sequence-Specific Nucleases’. Front. Plant Sci. 2016, vol. 7. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Doudna, J. A. ‘CRISPR–Cas9 Structures and Mechanisms’. Annu. Rev. Biophys. 2017, vol. 46(no. 1), 505–529. [Google Scholar] [CrossRef]
- Anzalone, V.; Koblan, L. W.; Liu, D. R. ‘Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors’. Nat Biotechnol 2020, vol. 38(no. 7), 824–844. [Google Scholar] [CrossRef]
- Shimatani, Z. ‘Targeted base editing in rice and tomato using a CRISPR-Cas9 cytidine deaminase fusion’. Nat Biotechnol 2017, vol. 35(no. 5), 441–443. [Google Scholar] [CrossRef]
- Veillet, F. ‘Expanding the CRISPR Toolbox in P. patens Using SpCas9-NG Variant and Application for Gene and Base Editing in Solanaceae Crops’. IJMS 2020, vol. 21(no. 3), 1024. [Google Scholar] [CrossRef]
- Sretenovic, S. ‘Exploring C-To-G Base Editing in Rice, Tomato, and Poplar’, Front. Genome Ed. 2021, vol. 3, 756766. [Google Scholar] [CrossRef]
- Hunziker, J.; Nishida, K.; Kondo, A.; Kishimoto, S.; Ariizumi, T.; Ezura, H. ‘Multiple gene substitution by Target-AID base-editing technology in tomato’. Sci Rep 2020, vol. 10(no. 1), 20471. [Google Scholar] [CrossRef]
- Yuan, S. ‘Efficient base editing in tomato using a highly expressed transient system’. Plant Cell Rep 2021, vol. 40(no. 4), 667–676. [Google Scholar] [CrossRef]
- Zong, Y. ‘Efficient C-to-T base editing in plants using a fusion of nCas9 and human APOBEC3A’. Nat Biotechnol 2018, vol. 36(no. 10), 950–953. [Google Scholar] [CrossRef]
- Veillet, F. ‘The Solanum tuberosum GBSSI gene: a target for assessing gene and base editing in tetraploid potato’. Plant Cell Rep 2019, vol. 38(no. 9), 1065–1080. [Google Scholar] [CrossRef]
- Li, J. ‘Plant base editing and prime editing: The current status and future perspectives’. JIPB 2023, vol. 65(no. 2), 444–467. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y. ‘Precise genome modification in tomato using an improved prime editing system’. Plant Biotechnology Journal 2021, vol. 19(no. 3), 415–417. [Google Scholar] [CrossRef] [PubMed]
- Vu, T. V.; Nguyen, N. T.; Kim, J.; Song, Y. J.; Nguyen, T. H.; Kim, J.-Y. ‘Optimized dicot prime editing enables heritable desired edits in tomato and Arabidopsis’. Nat. Plants 2024, vol. 10(no. 10), 1502–1513. [Google Scholar] [CrossRef]
- Lou, H. ‘Engineering source-sink relations by prime editing confers heat-stress resilience in tomato and rice’. Cell 2025, vol. 188(no. 2), 530–549.e20. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y. ‘Precise deletion, replacement and inversion of large DNA fragments in plants using dual prime editing’. Nat. Plants 2025, vol. 11(no. 2), 191–205. [Google Scholar] [CrossRef]
- Perroud, P.-F. ‘Prime Editing in the model plant Physcomitrium patens and its potential in the tetraploid potato’. Plant Science 2022, vol. 316, 111162. [Google Scholar] [CrossRef]
- La Russa, M. F.; Qi, L. S. ‘The New State of the Art: Cas9 for Gene Activation and Repression’. Molecular and Cellular Biology 2015, vol. 35(no. 22), 3800–3809. [Google Scholar] [CrossRef]
- Chen, M.; Qi, L. S. Repurposing CRISPR System for Transcriptional Activation’, in RNA Activation, vol. 983. In Advances in Experimental Medicine and Biology; Li, L.-C., Ed.; Springer Singapore: Singapore, 2017; vol. 983. pp. 147–157. [Google Scholar] [CrossRef]
- McLaughlin, J. E.; Kue Foka, I. C.; Lawton, M. A.; Di, R. ‘CRISPR activation: identifying and using novel genes for plant disease resistance breeding’. Front. Genome Ed. 2025, vol. 7, 1596600. [Google Scholar] [CrossRef]
- García-Murillo, L. ‘CRISPRa-mediated transcriptional activation of the SlPR-1 gene in edited tomato plants’. Plant Science 2023, vol. 329, 111617. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Toro, M.; De Folter, S.; Alvarez-Venegas, R. ‘CRISPR/dCas12a-mediated activation of SlPAL2 enhances tomato resistance against bacterial canker disease’. PLoS ONE 2025, vol. 20(no. 3), e0320436. [Google Scholar] [CrossRef]
- Valencia-Lozano. ‘Editing of SlWRKY29 by CRISPR-activation promotes somatic embryogenesis in Solanum lycopersicum cv. Micro-Tom’. PLoS ONE 2024, vol. 19(no. 4), e0301169. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee; Jodder, J.; Chowdhury, S.; Das, H.; Kundu, P. ‘A novel stress-inducible dCas9 system for solanaceous plants’. International Journal of Biological Macromolecules 2025, vol. 308, 142462. [Google Scholar] [CrossRef]
- Pan. ‘CRISPR–Act3.0 for highly efficient multiplexed gene activation in plants’. Nat. Plants 2021, vol. 7(no. 7), 942–953. [Google Scholar] [CrossRef]
- Casas-Mollano, J. A.; Zinselmeier, M. H.; Sychla, A.; Smanski, M. J. ‘Efficient gene activation in plants by the MoonTag programmable transcriptional activator’. Nucleic Acids Research 2023, vol. 51(no. 13), 7083–7093. [Google Scholar] [CrossRef]
- Xu, L. ‘The evaluation of active transcriptional repressor domain for CRISPRi in plants’. Gene 2023, vol. 851, 146967. [Google Scholar] [CrossRef]
- Bhatt, R.; Tiwari, B. S. ‘CRISPRi/dCas9-KRAB mediated suppression of Solanidine galactosyltransferase (sgt1) in Solanum tuberosum leads to the reduction in α-solanine level in potato tubers without any compensatory effect in α-chaconine.’. Biocatalysis and Agricultural Biotechnology 2024, vol. 58, 103133. [Google Scholar] [CrossRef]
- Bernabé-Orts, J. M. ‘Assessment of Cas12a-mediated gene editing efficiency in plants’. Plant Biotechnology Journal 2019, vol. 17(no. 10), 1971–1984. [Google Scholar] [CrossRef]
- Walton, R. T.; Christie, K. A.; Whittaker, M. N.; Kleinstiver, B. P. ‘Unconstrained genome targeting with near-PAMless engineered CRISPR-Cas9 variants’. Science 2020, vol. 368(no. 6488), 290–296. [Google Scholar] [CrossRef]
- Veillet. ‘Expanding the CRISPR Toolbox in P. patens Using SpCas9-NG Variant and Application for Gene and Base Editing in Solanaceae Crops’. IJMS 2020, vol. 21(no. 3), 1024. [Google Scholar] [CrossRef]
- Makarova, K. S. ‘An updated evolutionary classification of CRISPR–Cas systems including rare variants’. Nat Microbiol 2025. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay; Kancharla, N.; Javalkote, V. S.; Dasgupta, S.; Brutnell, T. P. ‘CRISPR-Cas12a (Cpf1): A Versatile Tool in the Plant Genome Editing Tool Box for Agricultural Advancement’. Front. Plant Sci. 2020, vol. 11, 584151. [Google Scholar] [CrossRef] [PubMed]
- Zetsche. ‘Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System’. Cell 2015, vol. 163(no. 3), 759–771. [Google Scholar] [CrossRef]
- Gao, L. ‘Engineered Cpf1 variants with altered PAM specificities’. Nat Biotechnol 2017, vol. 35(no. 8), 789–792. [Google Scholar] [CrossRef]
- Tóth. ‘Improved LbCas12a variants with altered PAM specificities further broaden the genome targeting range of Cas12a nucleases’. Nucleic Acids Research 2020, vol. 48(no. 7), 3722–3733. [Google Scholar] [CrossRef]
- Slaman, E.; Kottenhagen, L.; De Martines, W.; Angenent, G. C.; De Maagd, R. A. ‘Comparison of Cas12a and Cas9-mediated mutagenesis in tomato cells’. Sci Rep 2024, vol. 14(no. 1), 4508. [Google Scholar] [CrossRef]
- Vu, T. V.; Doan, D. T. H.; Tran, M. T.; Sung, Y. W.; Song, Y. J.; Kim, J.-Y. ‘Improvement of the LbCas12a-crRNA System for Efficient Gene Targeting in Tomato’. Front. Plant Sci. 2021, vol. 12, 722552. [Google Scholar] [CrossRef]
- Kavuri, N. R.; Ramasamy, M.; Qi, Y.; Mandadi, K. ‘Applications of CRISPR/Cas13-Based RNA Editing in Plants’. Cells 2022, vol. 11(no. 17), 2665. [Google Scholar] [CrossRef]
- Zhan, X. ‘Generation of virus-resistant potato plants by RNA genome targeting’. Plant Biotechnology Journal 2019, vol. 17(no. 9), 1814–1822. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Liu, W.; Nie, B.; Zhang, F.; Zhang, J. ‘Cas13d-mediated multiplex RNA targeting confers a broad-spectrum resistance against RNA viruses in potato’. Commun Biol 2023, vol. 6(no. 1), 855. [Google Scholar] [CrossRef]
- Noureen. Broad-spectrum resistance against multiple PVY-strains by CRSIPR/Cas13 system in Solanum tuberosum crop’. GM Crops & Food 2022, vol. 13(no. 1), 97–111. [Google Scholar] [CrossRef]
- Khoo, Y. W. ‘Resistance of the CRISPR-Cas13a Gene-Editing System to Potato Spindle Tuber Viroid Infection in Tomato and Nicotiana benthamiana’. Viruses 2024, vol. 16(no. 9), 1401. [Google Scholar] [CrossRef]
- Sharma, V. K. ‘CRISPR guides induce gene silencing in plants in the absence of Cas’. Genome Biol 2022, vol. 23(no. 1), 6. [Google Scholar] [CrossRef]
- Laforest, L. C.; Nadakuduti, S. S. Advances in Delivery Mechanisms of CRISPR Gene-Editing Reagents in Plants’, Front. Genome Ed. 2022, vol. 4, 830178. [Google Scholar] [CrossRef]
- Yang, S. H.; Kim, S. W.; Lee, S.; Koo, Y. ‘Optimized protocols for protoplast isolation, transfection, and regeneration in the Solanum genus for the CRISPR/Cas-mediated transgene-free genome editing’. Appl Biol Chem 2024, vol. 67(no. 1), 21. [Google Scholar] [CrossRef]
- Lin, C.-S. ‘DNA-free CRISPR-Cas9 gene editing of wild tetraploid tomato Solanum peruvianum using protoplast regeneration’. Plant Physiology 2022, vol. 188(no. 4), 1917–1930. [Google Scholar] [CrossRef]
- Nicolia; Andersson, M.; Hofvander, P.; Festa, G.; Cardi, T. ‘Tomato protoplasts as cell target for ribonucleoprotein (RNP)-mediated multiplexed genome editing’. Plant Cell Tiss Organ Cult 2021, vol. 144(no. 2), 463–467. [Google Scholar] [CrossRef]
- Liu, Y.; Andersson, M.; Granell, A.; Cardi, T.; Hofvander, P.; Nicolia, A. ‘Establishment of a DNA-free genome editing and protoplast regeneration method in cultivated tomato (Solanum lycopersicum)’. Plant Cell Rep 2022, vol. 41(no. 9), 1843–1852. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Kang, B.-C.; Han, J.-S.; Lee, J. M. ‘DNA-free genome editing in tomato protoplasts using CRISPR/Cas9 ribonucleoprotein delivery’. Hortic. Environ. Biotechnol. 2024, vol. 65(no. 1), 131–142. [Google Scholar] [CrossRef]
- González, M. N. ‘Comparative potato genome editing: Agrobacterium tumefaciens-mediated transformation and protoplasts transfection delivery of CRISPR/Cas9 components directed to StPPO2 gene’. Plant Cell Tiss Organ Cult 2021, vol. 145(no. 2), 291–305. [Google Scholar] [CrossRef]
- Andersson, M. ‘Genome editing in potato via CRISPR-Cas9 ribonucleoprotein delivery’. Physiologia Plantarum 2018, vol. 164(no. 4), 378–384. [Google Scholar] [CrossRef]
- Zhao, X. ‘Amylose starch with no detectable branching developed through DNA-free CRISPR-Cas9 mediated mutagenesis of two starch branching enzymes in potato’. Sci Rep 2021, vol. 11(no. 1), 4311. [Google Scholar] [CrossRef]
- Moon, K.-B. ‘Editing of StSR4 by Cas9-RNPs confers resistance to Phytophthora infestans in potato’. Front. Plant Sci. 2022, vol. 13, 997888. [Google Scholar] [CrossRef] [PubMed]
- Choi; Shin, H.; Kim, C. Y.; Park, J.; Kim, H. Highly efficient CRISPR/Cas9-RNP mediated CaPAD1 editing in protoplasts of three pepper ( Capsicum annuum L.) cultivars’. Plant Signaling & Behavior 2024, vol. 19(no. 1), 2383822. [Google Scholar] [CrossRef]
- Kim; Choi, J.; Won, K.-H. ‘A stable DNA-free screening system for CRISPR/RNPs-mediated gene editing in hot and sweet cultivars of Capsicum annuum’. BMC Plant Biology 2020, vol. 20(no. 1), 449. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-H.; Kim, H. ‘Harnessing CRISPR/Cas9 for Enhanced Disease Resistance in Hot Peppers: A Comparative Study on CaMLO2-Gene-Editing Efficiency across Six Cultivars’. IJMS 2023, vol. 24(no. 23), 16775. [Google Scholar] [CrossRef]
- Wang, Y. ‘A highly efficient mesophyll protoplast isolation and PEG-mediated transient expression system in eggplant’. Scientia Horticulturae 2022, vol. 304, 111303. [Google Scholar] [CrossRef]
- Fish, N.; Karp, A.; Jones, M. G. K. ‘Production of somatic hybrids by electrofusion in Solanum’. Theoret. Appl. Genetics 1988, vol. 76(no. 2), 260–266. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.; Ooms, G.; Jones, M. G. K. ‘Transient gene expression in electroporated Solanum protoplasts’. Plant Mol Biol 1989, vol. 13(no. 5), 503–511. [Google Scholar] [CrossRef]
- Banakar, R.; Schubert, M.; Collingwood, M.; Vakulskas, C.; Eggenberger, A. L.; Wang, K. ‘Comparison of CRISPR-Cas9/Cas12a Ribonucleoprotein Complexes for Genome Editing Efficiency in the Rice Phytoene Desaturase (OsPDS) Gene’. Rice 2020, vol. 13(no. 1), 4. [Google Scholar] [CrossRef]
- Ghogare, R.; Ludwig, Y.; Bueno, G. M.; Slamet-Loedin, I. H.; Dhingra, A. ‘Genome editing reagent delivery in plants’. Transgenic Res 2021, vol. 30(no. 4), 321–335. [Google Scholar] [CrossRef]
- Liu, W. ‘Lipofection-mediated genome editing using DNA-free delivery of the Cas9/gRNA ribonucleoprotein into plant cells’. Plant Cell Rep 2020, vol. 39(no. 2), 245–257. [Google Scholar] [CrossRef] [PubMed]
- Li, R. ‘CRISPR/Cas9-Based Knock-Out of the PMR4 Gene Reduces Susceptibility to Late Blight in Two Tomato Cultivars’. IJMS 2022, vol. 23(no. 23), 14542. [Google Scholar] [CrossRef] [PubMed]
- Uranga, M. ‘Virus-Induced Genome Editing’. In CRISPR and Plant Functional Genomics, 1st edn; CRC Press: Boca Raton, 2024; pp. 81–106. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, S.; Li, X.; Zhang, R.; Li, J. ‘Virus-Induced Gene Editing and Its Applications in Plants’. IJMS 2022, vol. 23(no. 18), 10202. [Google Scholar] [CrossRef]
- Ali, Z.; Abul-faraj, A.; Piatek, M.; Mahfouz, M. M. ‘Activity and specificity of TRV-mediated gene editing in plants’. Plant Signaling & Behavior 2015, vol. 10(no. 10), e1044191. [Google Scholar] [CrossRef] [PubMed]
- Ellison, E. E.; Nagalakshmi, U.; Gamo, M. E.; Huang, P.; Dinesh-Kumar, S.; Voytas, D. F. ‘Multiplexed heritable gene editing using RNA viruses and mobile single guide RNAs’. Nat. Plants 2020, vol. 6(no. 6), 620–624. [Google Scholar] [CrossRef]
- Lee, S.-Y. ‘Development of virus-induced genome editing methods in Solanaceous crops’. Horticulture Research 2024, vol. 11(no. 1), uhad233. [Google Scholar] [CrossRef]
- Lee, H.; Baik, J. E.; Kim, K.-N. Development of an efficient and heritable virus-induced genome editing system in Solanum lycopersicum’. Horticulture Research 2025, vol. 12(no. 4), uhae364. [Google Scholar] [CrossRef]
- Kang, G. H.; Ko, Y.; Lee, J. M. ‘Enhancing virus-mediated genome editing for cultivated tomato through low temperature’. Plant Cell Rep 2025, vol. 44(no. 1), 22. [Google Scholar] [CrossRef]
- Oh, Y. ‘Heritable virus-induced germline editing in tomato’. The Plant Journal 2025, vol. 122(no. 1), e70115. [Google Scholar] [CrossRef] [PubMed]
- Uranga, M. ‘RNA virus-mediated gene editing for tomato trait breeding’. Horticulture Research 2024, vol. 11(no. 1), uhad279. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, D.; Dai, Y.; Liu, C. ‘Efficient tobacco rattle virus-induced gene editing in tomato mediated by the CRISPR/Cas9 system’. Biotechnology Journal 2024, vol. 19(no. 5), 2400204. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhao, C.; Sun, K.; Deng, Y.; Li, Z. ‘Engineered biocontainable RNA virus vectors for non-transgenic genome editing across crop species and genotypes’. Molecular Plant 2023, vol. 16(no. 3), 616–631. [Google Scholar] [CrossRef]
- Kang, B. ‘Virus-induced systemic and heritable gene editing in pepper ( Capsicum annuum L.)’. The Plant Journal 2025, vol. 122(no. 5), e70257. [Google Scholar] [CrossRef]
- Zhao, C.; Lou, H.; Liu, Q.; Pei, S.; Liao, Q.; Li, Z. ‘Efficient and transformation-free genome editing in pepper enabled by RNA virus-mediated delivery of CRISPR/Cas9’. JIPB 2024, vol. 66(no. 10), 2079–2082. [Google Scholar] [CrossRef]
- Ji, X.; Yang, B.; Wang, D. ‘Achieving Plant Genome Editing While Bypassing Tissue Culture’. Trends in Plant Science 2020, vol. 25(no. 5), 427–429. [Google Scholar] [CrossRef]
- Maher, M. F.; Nasti, R. A.; Vollbrecht, M.; Starker, C. G.; Clark, M. D.; Voytas, D. F. ‘Plant gene editing through de novo induction of meristems’. Nat Biotechnol 2020, vol. 38(no. 1), 84–89. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Li, C.; Gao, C. ‘Applications of CRISPR–Cas in agriculture and plant biotechnology’. Nat Rev Mol Cell Biol 2020, vol. 21(no. 11), 661–677. [Google Scholar] [CrossRef]
- Liu, D. ‘Heritable gene editing in tomato through viral delivery of isopentenyl transferase and single-guide RNAs to latent axillary meristematic cells’. Proc. Natl. Acad. Sci. U.S.A. 2024, vol. 121(no. 39), e2406486121. [Google Scholar] [CrossRef]
- Kshetry, O. ‘A synthetic transcription cascade enables direct in planta shoot regeneration for transgenesis and gene editing in multiple plants’. Molecular Plant 2025, vol. 18(no. 12), 2066–2081. [Google Scholar] [CrossRef] [PubMed]
- Lian, Z. ‘Application of developmental regulators to improve in planta or in vitro transformation in plants’. Plant Biotechnology Journal 2022, vol. 20(no. 8), 1622–1635. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S. ‘Efficient base editing in tomato using a highly expressed transient system’. Plant Cell Rep 2021, vol. 40(no. 4), 667–676. [Google Scholar] [CrossRef]
- European Parliament. ‘Directive 2001/18/EC of the European Parliament and of the Council of 12 March 2001 on the deliberate release into the environment of genetically modified organisms and repealing Council Directive 90/220/EEC.’ Apr. 17, 2001. 08 May 2025. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=CELEX:32001L0018&rid=1.
- Habets, M. G. J. L.; Macnaghten, P. ‘From genes to governance: Engaging citizens in the new genomic techniques policy debate’. Plants People Planet 2024, p. ppp3.10626. [Google Scholar] [CrossRef]
- European Commission. ‘Proposal for a REGULATION OF THE EUROPEAN PARLIAMENT AND OF THE COUNCIL on plants obtained by certain new genomic techniques and their food and feed, and amending Regulation (EU) 2017/625’. 05 Jul 2023. Available online: https://food.ec.europa.eu/plants/genetically-modified-organisms/new-techniques-biotechnology_en.
- Council of the EU. ‘New genomic techniques: Council and Parliament strike deal to boost the competitiveness and sustainability of our food systems’. 04 Dec 2025. Available online: https://www.consilium.europa.eu/en/press/press-releases/2025/12/04/new-genomic-techniques-council-and-parliament-strike-deal-to-boost-the-competitiveness-and-sustainability-of-our-food-systems/.
- European Commission. Directorate-General for Health and Food Safety, EU’s rules new genomic techniques. Publications Office of the European Union: LU, 2025; Available online: https://data.europa.eu/doi/10.2875/1144622.
- Zimny, T. ‘Regulation of GMO field trials in the EU and new genomic techniques: will the planned reform facilitate experimenting with gene-edited plants?’. BioTechnologia 2023, vol. 104(no. 1), 75–83. [Google Scholar] [CrossRef]
- UK Parliament, Genetic Technology (Precision Breeding) Act 2023. 2023. Available online: https://www.legislation.gov.uk/ukpga/2023/6/contents.
- UK Parliament, The Genetic Technology (Precision Breeding) Regulations 2025. 2025. Available online: https://www.legislation.gov.uk/uksi/2025/581/contents/made.
- ACRE, ACRE guidance on producing precision bred plants. 2025. Available online: https://www.gov.uk/government/publications/acre-guidance-on-producing-precision-bred-plants.
- Nicolia. Editing strigolactone biosynthesis genes in tomato reveals novel phenotypic effects and highlights D27 as a key target for parasitic weed resistance’. In Genetics; 06 Feb 2025. [Google Scholar] [CrossRef]
- Sakthivel; Balasubramanian, R.; Sampathrajan, V.; Veerasamy, R.; Appachi, S. V.; K. K., K. ‘Transforming tomatoes into GABA-rich functional foods through genome editing: A modern biotechnological approach’. Funct Integr Genomics 2025, vol. 25(no. 1), 27. [Google Scholar] [CrossRef]
- Waltz, E. ‘GABA-enriched tomato is first CRISPR-edited food to enter market’. Nat Biotechnol 2022, vol. 40(no. 1), 9–11. [Google Scholar] [CrossRef]
- Karlson, D. ‘Targeted Mutagenesis of the Multicopy Myrosinase Gene Family in Allotetraploid Brassica juncea Reduces Pungency in Fresh Leaves across Environments’. Plants 2022, vol. 11(no. 19), 2494. [Google Scholar] [CrossRef]
Figure 1.
CRISPR/Cas9 system and subsequent DNA repair mechanisms: NHEJ (left) and HDR (right). (Created in BioRender.com).
Figure 1.
CRISPR/Cas9 system and subsequent DNA repair mechanisms: NHEJ (left) and HDR (right). (Created in BioRender.com).
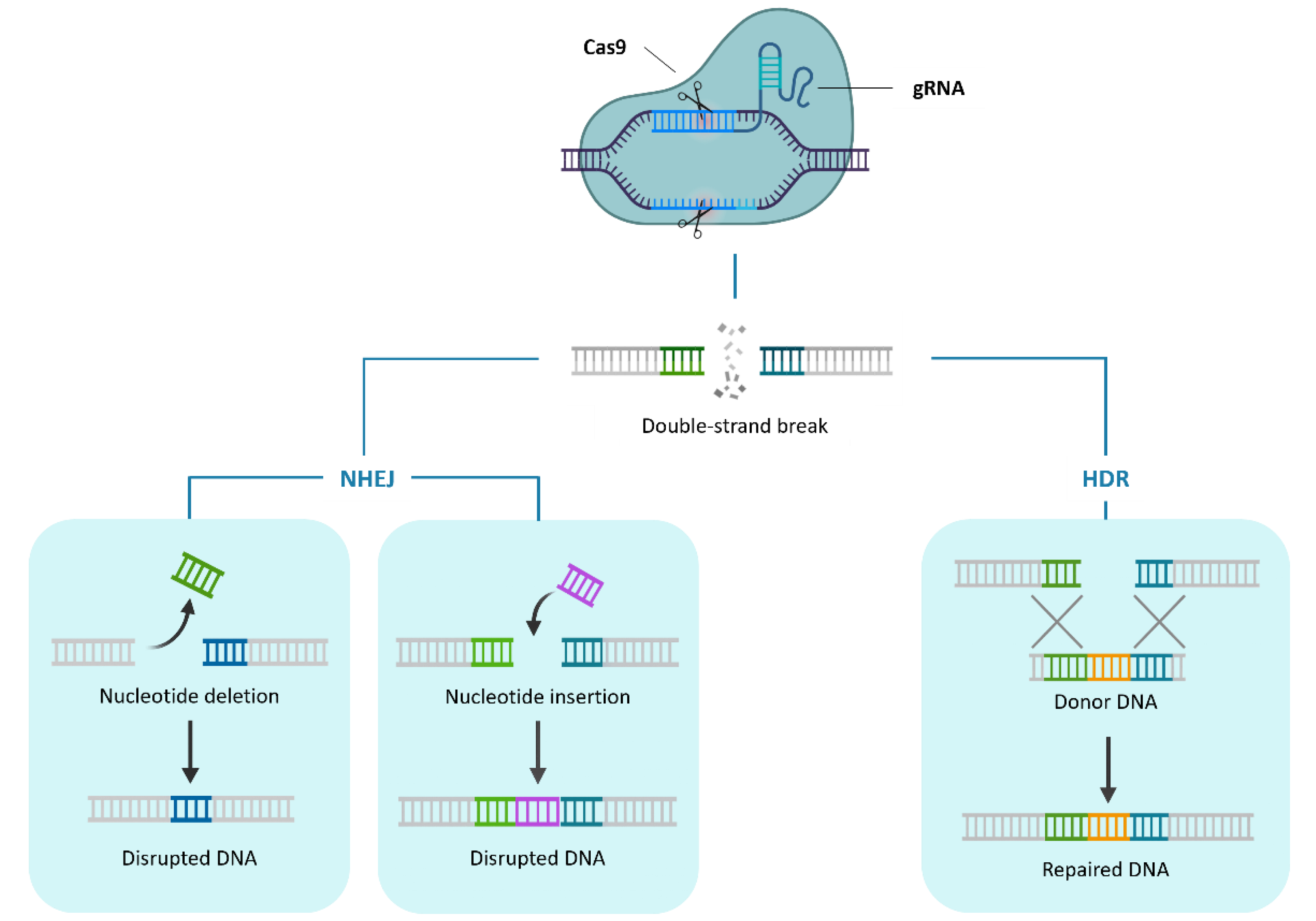
Figure 2.
CRISPR/Cas9 mediated base editing (A) and prime editing (B) systems. (Created in BioRender.com).
Figure 2.
CRISPR/Cas9 mediated base editing (A) and prime editing (B) systems. (Created in BioRender.com).
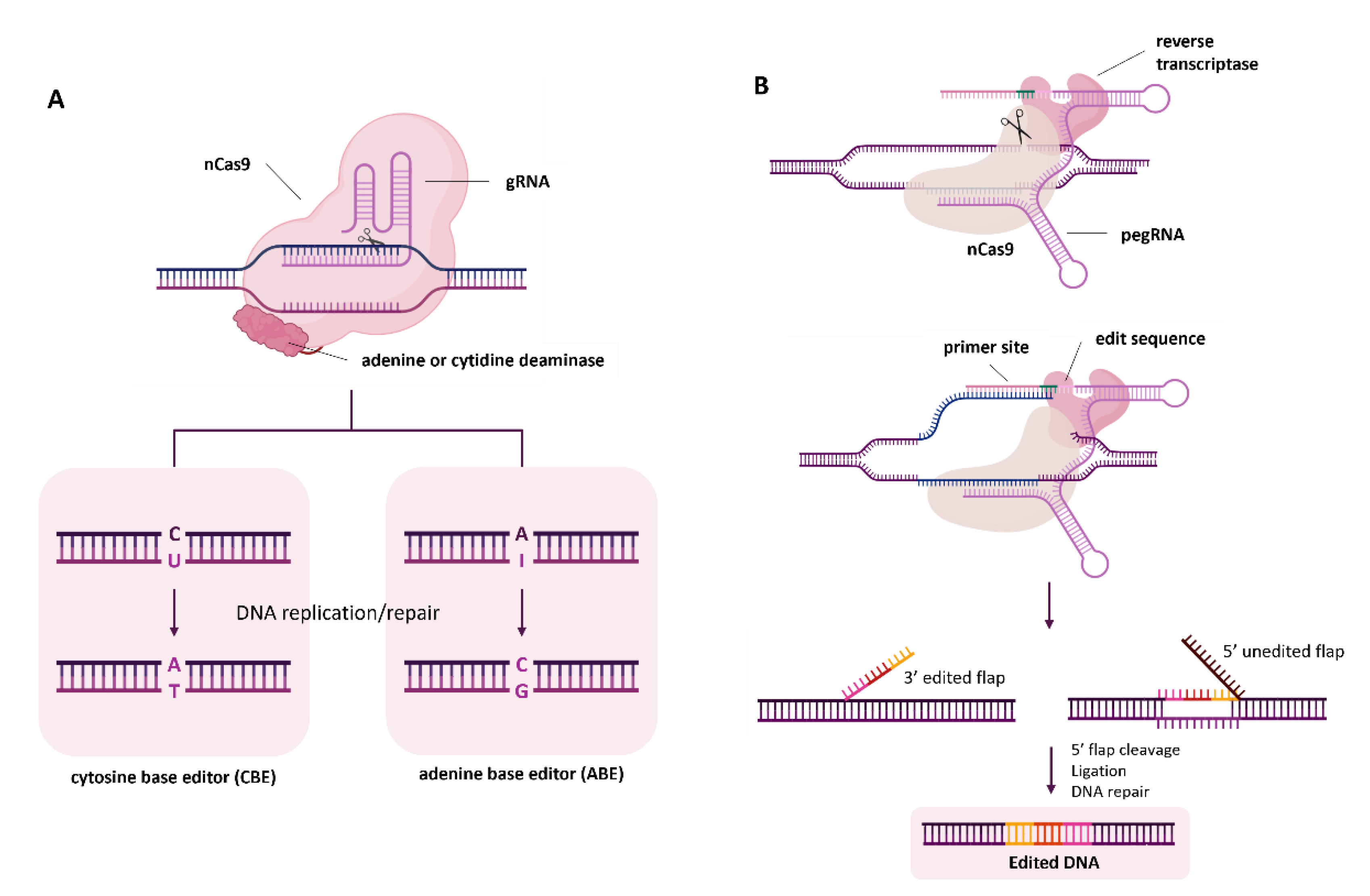
Figure 3.
CRISPR/Cas9-mediated transcription activation and inhibition systems: CRISPRa (A), CRISPRi (B) and epigenetic activation (C). (Created in BioRender.com).
Figure 3.
CRISPR/Cas9-mediated transcription activation and inhibition systems: CRISPRa (A), CRISPRi (B) and epigenetic activation (C). (Created in BioRender.com).
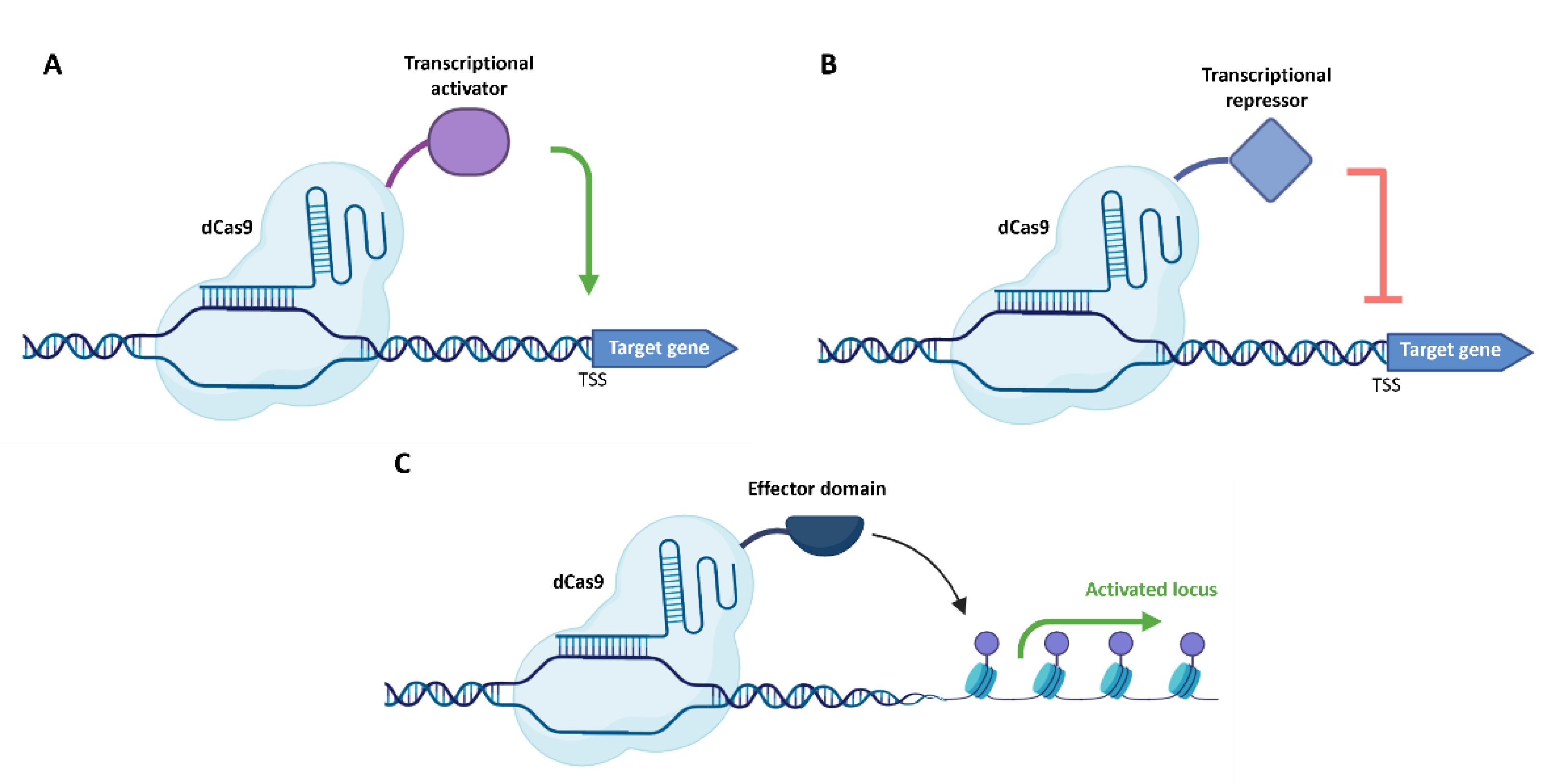
Figure 4.
Different Cas proteins and their CRISPR machinery: CRISPR/Cas9 (A), CRISPR/Cas12 (B), CRISPR/Cas13a (C). (Created in BioRender.com).
Figure 4.
Different Cas proteins and their CRISPR machinery: CRISPR/Cas9 (A), CRISPR/Cas12 (B), CRISPR/Cas13a (C). (Created in BioRender.com).
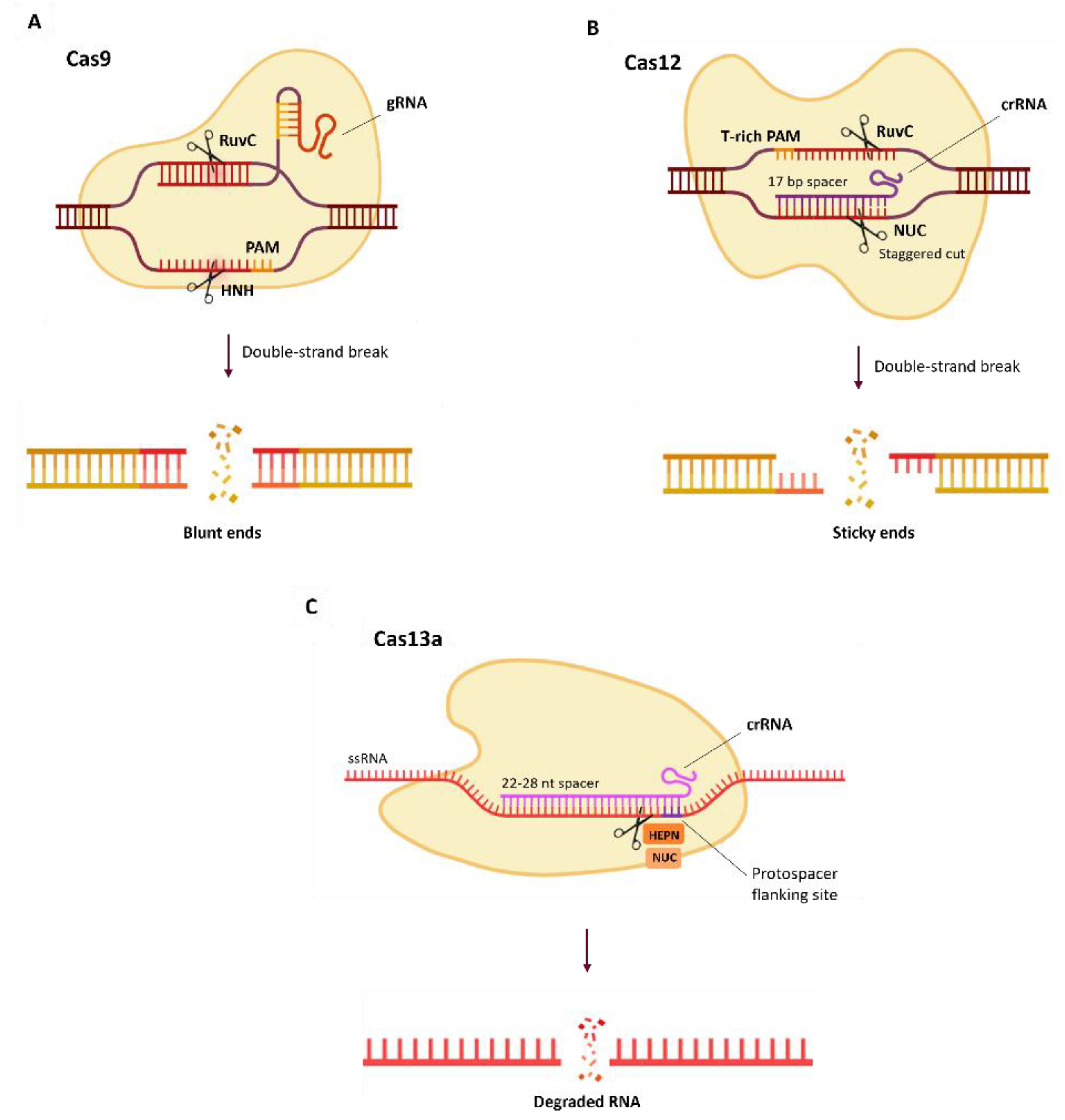
Figure 5.
CRISPR/Cas9 RNPs mediated gene editing in protoplasts. CRISPR reagents can be delivered into plant cells as RNPs. Particle bombardment is mainly used to deliver RNPs into explants, while PEG-mediated transfection and lipofection are used for delivery into protoplasts. Nanoparticles are emerging methods for RNP delivery. Transformed cells are then used for plant regeneration and edit detection. (Created in BioRender.com).
Figure 5.
CRISPR/Cas9 RNPs mediated gene editing in protoplasts. CRISPR reagents can be delivered into plant cells as RNPs. Particle bombardment is mainly used to deliver RNPs into explants, while PEG-mediated transfection and lipofection are used for delivery into protoplasts. Nanoparticles are emerging methods for RNP delivery. Transformed cells are then used for plant regeneration and edit detection. (Created in BioRender.com).
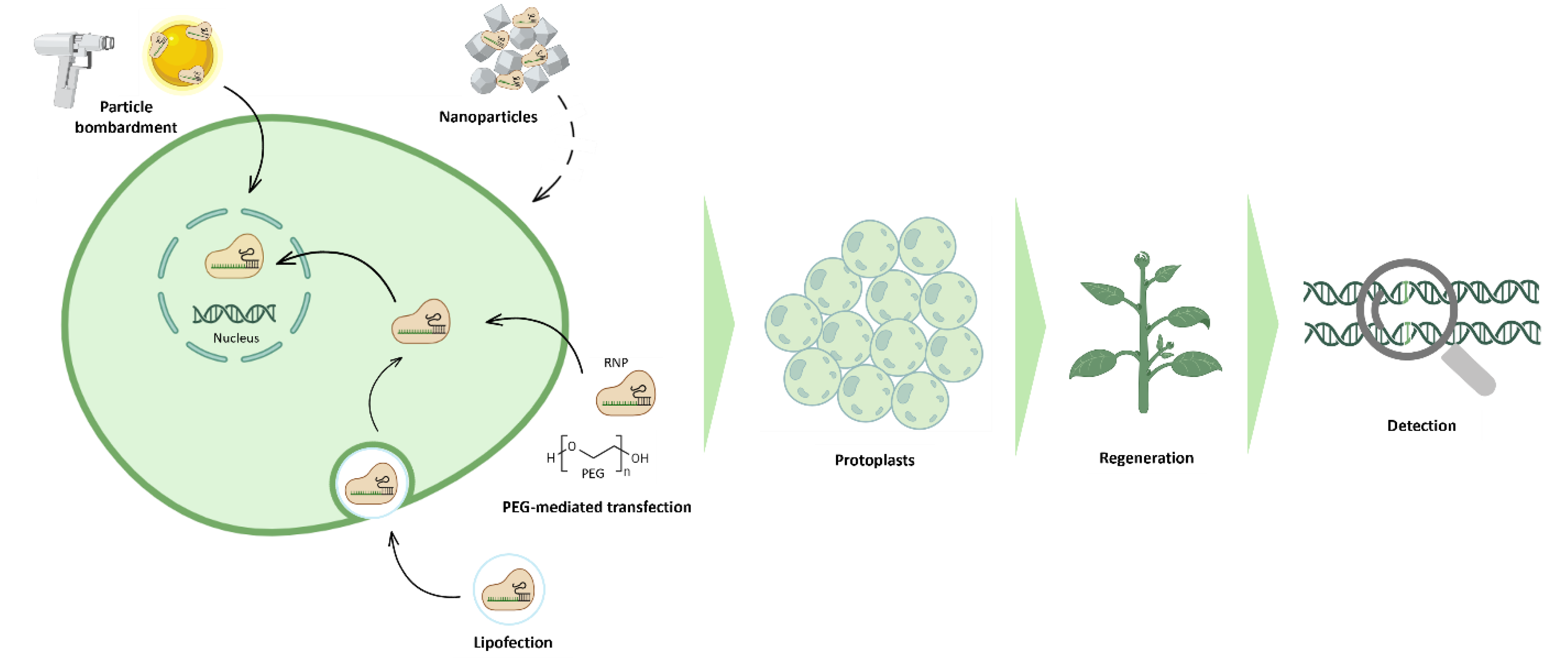
Figure 6.
VIGE strategies to obtain heritable gene editing in plants. (A) VIGE vectors that deliver both Cas9 and the gRNA spread systemically in the plant. Subsequent tissue culture from infected leaves is needed to regenerate gene-edited plants that can then produce gene-edited Cas-free seeds. (B) If the viral infection reaches the meristem, edited seeds and progeny may be obtained. (C) VIGE vectors expressing the gRNA fused to mobile elements are used to infect plants that stably express Cas9, thus producing gene-edited Cas9 transgenic seeds. The edited generation needs to be crossed with wild-type plants or self-pollinated to segregate the Cas9 gene. (Created in BioRender.com).
Figure 6.
VIGE strategies to obtain heritable gene editing in plants. (A) VIGE vectors that deliver both Cas9 and the gRNA spread systemically in the plant. Subsequent tissue culture from infected leaves is needed to regenerate gene-edited plants that can then produce gene-edited Cas-free seeds. (B) If the viral infection reaches the meristem, edited seeds and progeny may be obtained. (C) VIGE vectors expressing the gRNA fused to mobile elements are used to infect plants that stably express Cas9, thus producing gene-edited Cas9 transgenic seeds. The edited generation needs to be crossed with wild-type plants or self-pollinated to segregate the Cas9 gene. (Created in BioRender.com).
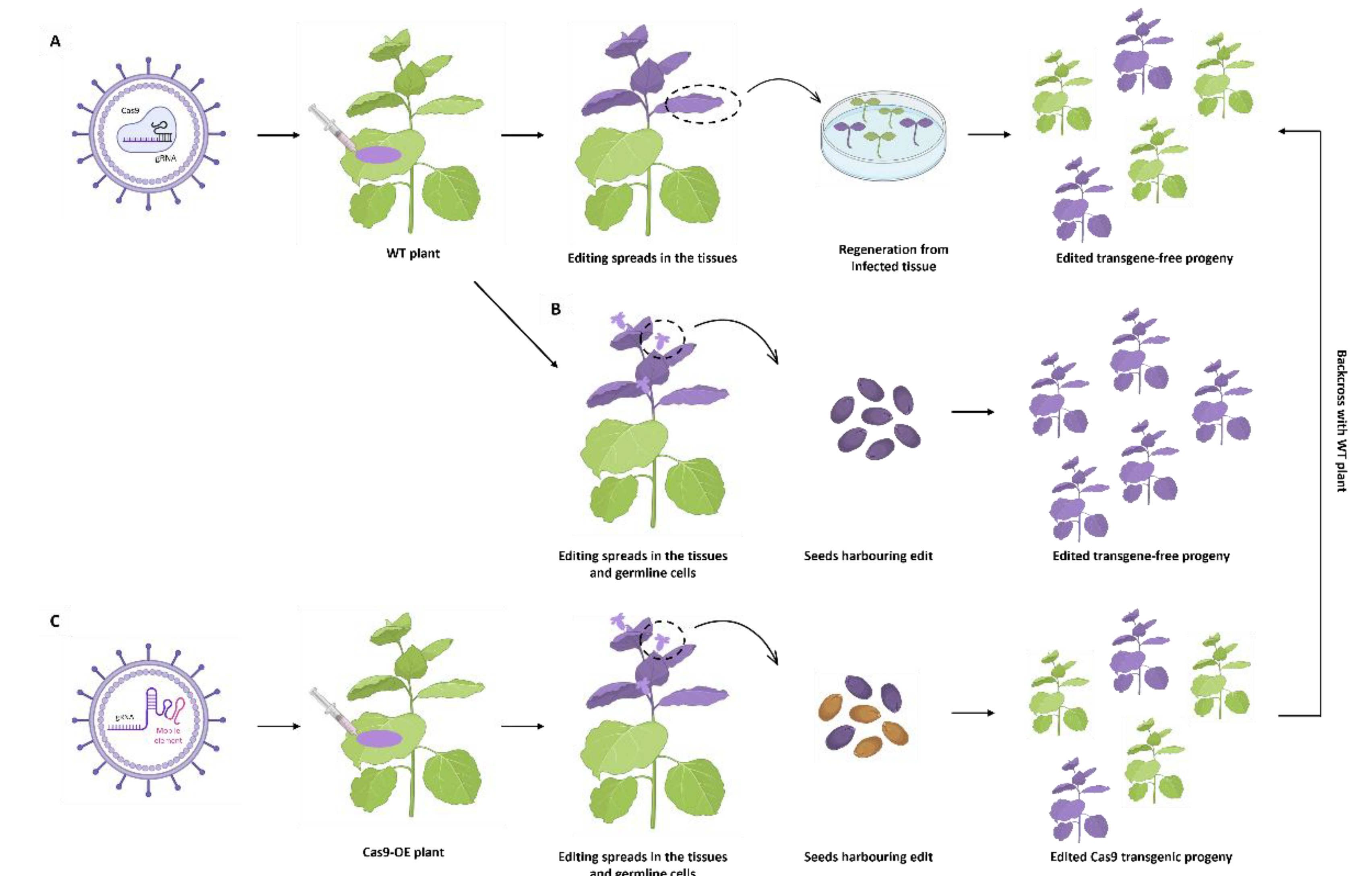
Figure 7.
Induction of genetically modified meristems on soil-grown plants. Schematic representation of the procedure to induce genetically modified meristem on soil-grown plants: A. tumefaciens culture is perfused in the pruned sites, where DRs expression stimulates new gene-edited meristems (orange) to develop into a whole plant. (Created in BioRender.com).
Figure 7.
Induction of genetically modified meristems on soil-grown plants. Schematic representation of the procedure to induce genetically modified meristem on soil-grown plants: A. tumefaciens culture is perfused in the pruned sites, where DRs expression stimulates new gene-edited meristems (orange) to develop into a whole plant. (Created in BioRender.com).
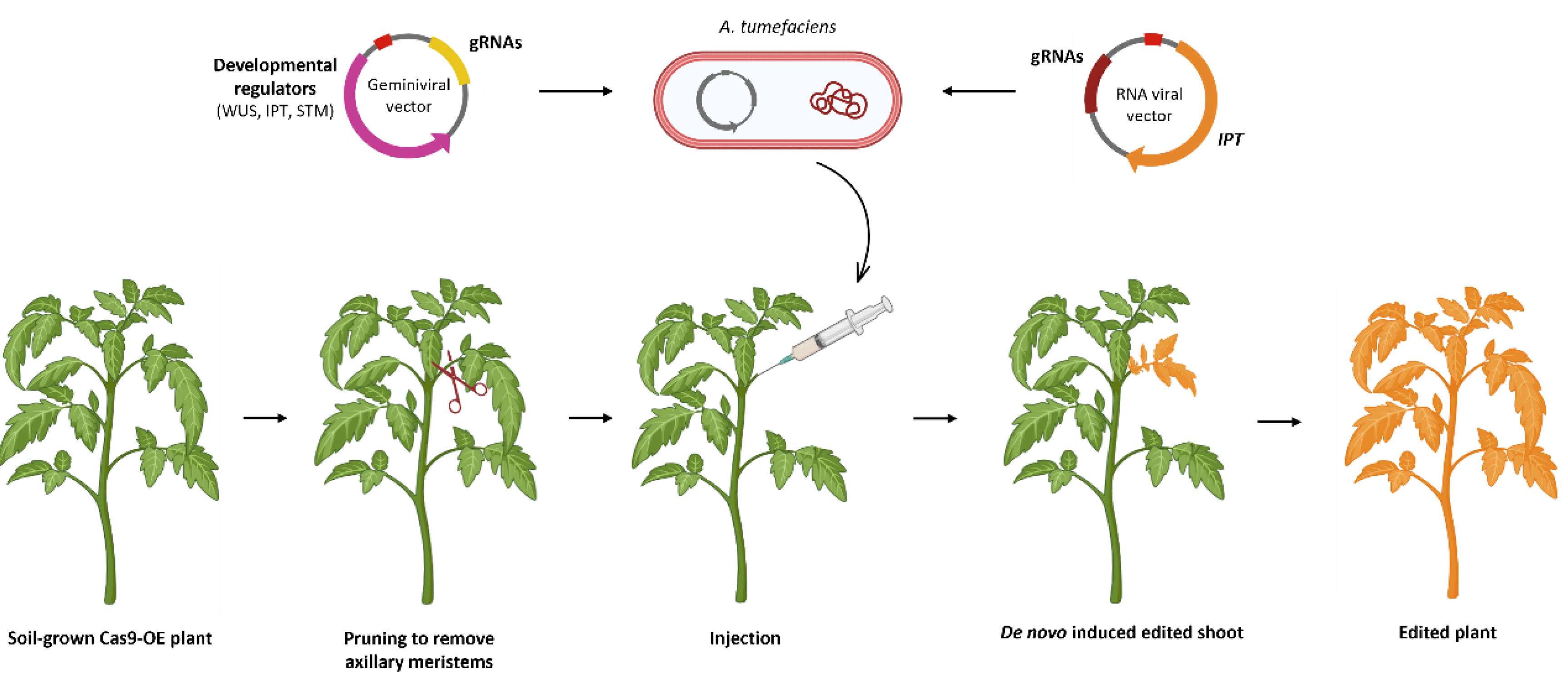

Table 1.
Authorised field trials in Europe for NGT varieties in Solanaceae species.
| Species | Veriety | Gene | Nation | Notification number | Trait |
|---|---|---|---|---|---|
| S. lycopersicum | San Marzano | DMR6-1 | Italy | B/IT/25/03 | Tolerance to abiotic and biotic stress |
| S. lycopersicum | Alisa Craig | D27, CCD7 | Italy | B/IT/24/02 | Reduced strigolactones - resistance razmakto Orobanche spp. |
| S. lycopersicum | MoneyMaker | 7-DR2 | UK | 23/Q06 | Conversion of provitamin D3 to vitamin razmakD3 in sunlight |
| S. tuberosum | Ydun | DMR6-1 | Denmark | B/DK/24/23110 | Tolerance to late blight |
| S. tuberosum | Ydun | DMR6-1 | Denmark | B/DK/54/StDMR6-1 LoF Ydun | Altered sensitvity razmakto blight |
| S. tuberosum | Wotan | GBSS1 | Denmark | B/DK/55/Waxy Wotan | Amylopectin only starch |
| S. tuberosum | Various | Rpi-chc1, Rpi-cap1, ELR1, Peru / DND1, PMR4, DMR6-1, SR4 | Netherlands | B/NL/25/007 | Increased resistance / lower susceptibility to Phytophthora infestans |
| S. tuberosum | Kuras | DMR6-1 | Denmark | B/DK/25/Kuras | Tolerance to late blight |
| S. tuberosum | Ydun | CBP-1 | Denmark | B/DK/25/Ydun_v1 | Tolerance to late blight |
| S. tuberosum | NA | GBSS, SSS3, SSS2 | Sweden | B/SE/19/5614 | Lack of amylose starch |
| S. tuberosum | NA | DMR6, CHL1, AsS1, PiS1 | Sweden | B/SE/21/3359 | Altered resistance razmakto pathogens |
| S. tuberosum | NA | CHL1 | Sweden | B/SE/20/1726 | Altered resistance razmakto pathogens |
| S. tuberosum | NA | DMR6-1 | Sweden | B/SE/23/3093 | Altered resistance razmakto pathogens |
| S. tuberosum | Various | GBSS, SSS, SBE | Sweden | B/SE/22/23780 | Tailored starch quality |
| S. tuberosum | Various | GBSS, SSS3, SSS2, SSS1, SSS4, SSS5, SSS6, SBE1, SBE2 / RMA1H1, SMO1, DWF1, GAME9, DWF7, GAME6, GAME11, GAME4, GAME12 / DMR6, Parakletosis, eIF4E | Sweden | B/SE/24/20422 | Altered starch quality / decreased content of antinutritional compounds (TGA) / reduced susceptibility to late blight and PVY |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



