
Submitted:
16 January 2026
Posted:
20 January 2026
You are already at the latest version
Abstract
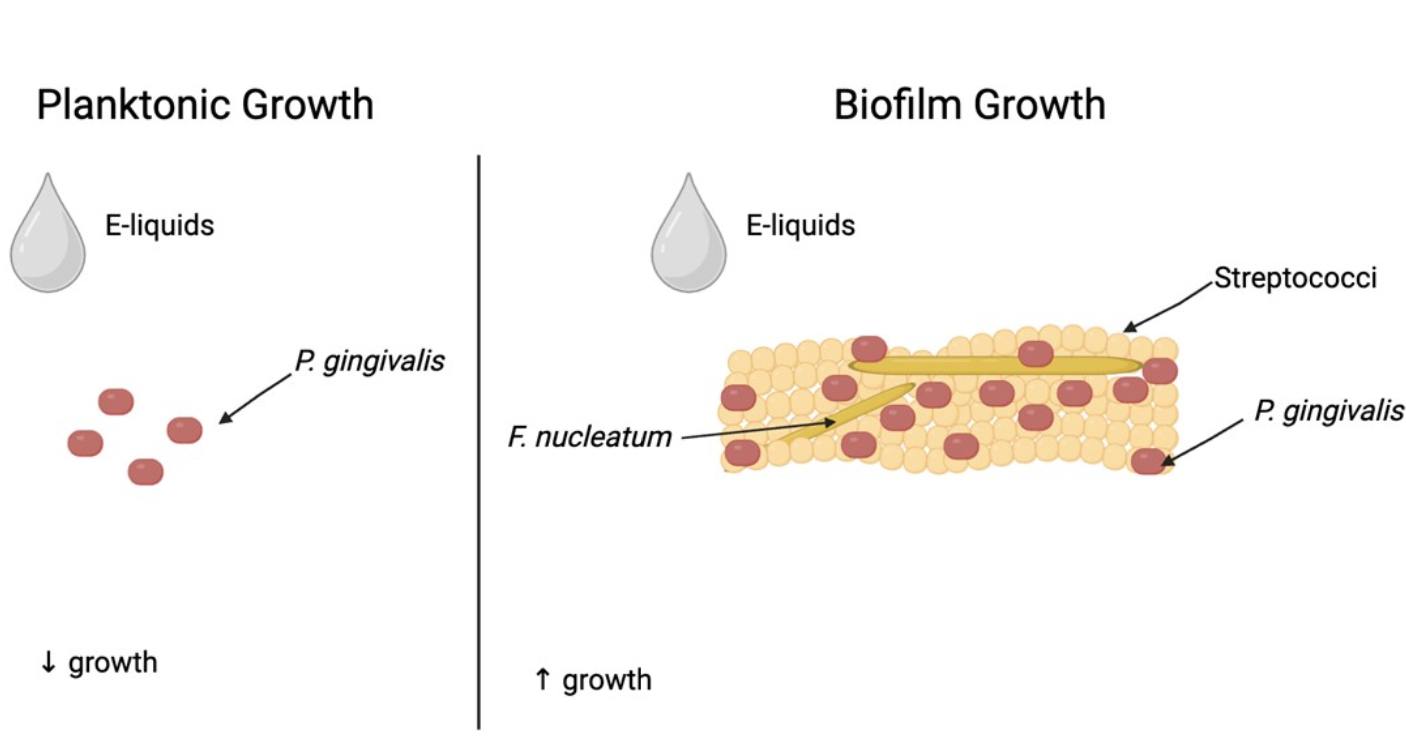
Background: The increasing use of electronic cigarettes (ECIGs), especially among youth, has raised concerns about the impact of vaping on oral health. While ECIGs are often marketed as a safer alternative, existing literature suggests their use may have detrimental effects on the pulmonary and cardiovascular systems. The oral cavity is the first point of contact for ECIG aerosol, and new reports link vaping to the onset of periodontal disease. It is critical to understand the potential effects of vaping on the oral microbiome, which affects systemic health. This study investigates how flavored E-liquids and commensal bacteria influence the growth of Porphyromonas gingivalis, a periodontal pathogen, under planktonic and biofilm conditions. Methods: P. gingivalis was grown planktonically in the presence of the supernatants of four streptococcal species (Streptococcus gordonii, Streptococcus intermedius, Streptococcus mitis, and Streptococcus oralis) and flavored E-liquids (tobacco, menthol, cinnamon, strawberry, and blueberry) under anaerobic conditions. Multispecies biofilms, including all species above and Fusobacterium nucleatum, were also grown anaerobically and quantified by crystal violet assays, qPCR, and CFU counts. Re-sults: Although E-liquids inhibit P. gingivalis growth under planktonic conditions, the presence of commensal supernatants partially mitigates this effect. However, P. gingivalis growth in multispecies biofilms is increased by E-liquid treatments. Conclusion: This study highlights the enhanced growth of P. gingivalis as part of an oral microbial community in the presence of E-liquids. This dysbiosis can lead to oral diseases, such as periodontitis, and ultimately systemic pathologies.
Keywords:
e-liquids
; bacterial growth
; bactericidal
; biofilms
; commensal bacteria
; electronic cigarettes
; oral cavity
; streptococci
; Porphyromonas gingivalis
1. Introduction
The use of electronic cigarettes (ECIGs) has become increasingly popular, especially among adolescents [1,2]. These devices were first introduced in China in 2003 as a safer alternative to smoking [3,4], and have since exploded into a worldwide public health issue [5,6]. In their simplest form, ECIGs consist of a battery, a heating coil, a removable cartridge that holds the E-liquid, and a mouthpiece [3]. E-liquids consist of vegetable glycerin, propylene glycol, various flavoring agents, and different concentrations of nicotine [7]. When the coil heats the E-liquid, it produces an aerosol that is inhaled, similar to traditional cigarette smoke, a process known as vaping [7].
Young adults are the primary target of ECIG marketing [3], and the myriad of palatable E-liquid flavors are used as the enticement [8]. These young adults are able to acquire vaping products even though many states restrict the sale to minors. Vaping is also marketed as a healthier alternative to traditional cigarette smoking or as a means to abstain from the latter [9,10] due to its minimal ingredient list and non-combustible property. Whether or not this is true, existing literature shows that vaping in of itself can be harmful [7,11,12]. For example, vaping has been linked with damage to the pulmonary and cardiovascular systems [13]. Thus far, the flavoring components of E-liquids have been shown to cause the most detrimental effects on various models [14,15,16,17,18,19,20,21]. However, high concentrations of nicotine in the E-liquid, the formation of volatile organic compounds from the heating of the E-liquid, and the presence of trace amounts of metals leached from the ECIG device itself [22] may contribute to harmful vaping-induced effects in the airway and oral cavity. While there is substantial research on the effects of E-liquids on the respiratory system, little work has examined their effects on the oral cavity.
The oral cavity contains one of the highest concentrations of bacteria in the human body, harboring over 700 microbial species [23]. These microorganisms typically exist as biofilms, structured communities of bacteria adhered to oral surfaces, which include live organisms, dead cells, microbial metabolites, an exopolysaccharide layer that covers the community (slime layer), extracellular DNA and nutrients that flow in and out of the biofilm [24]. Oral biofilms live symbiotically with the host and play a central role in homeostasis and oral health [25,26]. Oral pathogens reside in these complex microbial communities, and the balance between health and disease depends on the multispecies interactions with the host [27]. These interactions include symbiotic and antagonistic mechanisms within oral biofilms. For example, the gram-negative anaerobe and periodontal pathogen Porphyromonas gingivalis can coexist with commensals in a mutualistic balance as commensals remove oxygen, enabling P. gingivalis to thrive [28]. Oral commensal Streptococcus gordonii, an early colonizer of oral surfaces, produces ornithine, which is further metabolized by another gram-negative anaerobe, Fusobacterium nucleatum, generating polyamines and putrescine, which accelerate maturation and dispersal of P. gingivalis [29]. In symbiosis with the host, S. gordonii produces arginine deiminase, which metabolizes arginine to generate ammonia, thereby neutralizing acids in dental plaque and protecting the tooth enamel from demineralization [30]. In contrast, many commensal streptococci help maintain microbial balance on oral surfaces by producing hydrogen peroxide, minimizing the overgrowth of pathogens, and thereby helping maintain homeostasis [31]. A prior study conducted by our team showed that the oral commensals S. gordonii and Streptococcus intermedius defend against invasion of P. gingivalis into oral epithelial cells [32], exemplifying that oral homeostasis is dependent upon interactions, both symbiotic and antagonistic, between microbial species, leading to a balance for both the microbes and the host.
Studies by our group have shown that traditional cigarette smoke is more harmful to oral commensal streptococci compared to flavorless E-liquid or its aerosol [33,34], however, the effects of flavored E-liquids on oral commensals are dose-dependent. At lower E-liquid concentrations (1%), commensal bacteria are not adversely affected; in fact, some strains exhibit improved growth under these conditions [35]. At higher E-liquid concentrations (up to 5%), a clear dose-dependent reduction in biofilm formation is evident [35]. The oral mucosa, which interacts with oral commensal bacteria, is also affected by 1% E-liquids, as evidenced by pathophysiological responses in oral keratinocytes [16]. Disruptions include compromised wound healing, increased oxidative stress, altered mucin expression, and elevated production of pro-inflammatory cytokines. Together, these findings indicate that E-liquids can directly destabilize oral homeostasis.
Emerging clinical evidence reveals concerning patterns of oral health deterioration among ECIG users. Studies comparing periodontal parameters among cigarette smokers, vapers, and non-smokers have shown that conventional smokers exhibit the poorest periodontal status with elevated inflammatory mediators, while vapers demonstrate intermediate and unique outcomes between smokers and non-smokers [36,37]. In addition to periodontal disease, vapers were more likely to have untreated dental caries compared to non-smokers [38,39,40]. Microbiome analyses also display an increase in the abundance of pathogenic bacteria, including P. gingivalis, among vapers [41,42,43,44]. These clinical findings suggest that vaping presents different oral health risks compared to smoking, affecting various tissues of the oral cavity. Thus far, such observational studies indicate that vaping may also be associated with an increased risk for periodontal disease, and that P. gingivalis plays a key role. Furthermore, it is essential to note that oral and systemic health are intimately related. For example, periodontal disease is associated with a range of systemic conditions, including diabetes and cardiovascular disease [45,46]. Therefore, vaping may have short and long-term effects on both oral and systemic health.
This study aims to assess the effects of E-liquids on (i) the planktonic growth of P. gingivalis alone and in the presence of oral commensal streptococci supernatants, individually or pooled together; and (ii) the growth of P. gingivalis in multispecies biofilms. We hypothesize that E-liquids ± flavors inhibit P. gingivalis growth, but streptococci or their by-products present in the supernatant may mitigate this toxic effect either planktonically or in multispecies biofilms. Understanding these effects may help clarify how vaping influences P. gingivalis within oral microbial communities, which could lead to periodontal disease.
2. Materials and Methods
2.1. Reagents and Supplies
Reagents and supplies for this study were purchased from Fisher Scientific (Waltham, MA, USA) unless otherwise explicitly noted.
2.2. Stock E-Liquids
The base flavorless E-liquid solution was prepared by mixing the humectants, propylene glycol and glycerol, in a 1:1 v/v ratio (Liquid Nicotine Wholesalers, Phoenix, AZ, USA), then spiked with 20 mg/mL of (S)-(-)-nicotine, 99% (Alpha Aesar, Tewksbury, MA, USA). Stock flavors, including tobacco, cinnamon, strawberry and blueberry (Liquid Nicotine Wholesalers, Phoenix, AZ, USA), and menthol (Vapor Vapes, Sand City, CA, USA), were added to the base flavorless E-liquid at a final concentration of 5% (v/v) as outlined in our previous protocols [35,47,48] E-liquids were stored at 4 °C after preparation.
2.3. Saliva Preparation
Saliva samples were collected from a minimum of five healthy donors under IRB approval (Cuadra_S19_18). In accordance with previously published protocols [35,47] the following inclusion criteria were met: (i) non-smokers and non-vapers, (ii) in good health at the time of donation, (iii) no antibiotic use within the three months preceding donation, and (iv) no consumption of food or beverages other than water within two hours prior to donation. Raw saliva samples were stored at -20 °C until processing, then thawed and pooled on ice. Dithiothreitol was added to a final concentration of 2.5 mM, and samples were gently stirred on ice for 10-15 min. The saliva was subsequently centrifuged at 4500× g for 90 min, after which the supernatant was collected and diluted 1:4 (v/v) with distilled water. The diluted saliva was filter-sterilized using a 0.45 µm vacuum filtration system. Sterile saliva was stored at -20 °C for up to one year or at 4 °C for up to two weeks prior to use.
2.4. Bacterial Strains and Growth Conditions
All bacterial stocks were stored at -80 °C, and the purity of each strain was routinely verified by 16S rRNA gene sequencing (Genewiz, South Plainfield, NJ, USA) to confirm species identity.
Facultative oral streptococci used in this study include S. gordonii DL1, S. intermedius 0809, Streptococcus mitis UF2, and Streptococcus oralis SK139. These strains were kindly provided by Dr. Robert Burne from the University of Florida, College of Dentistry in Gainesville, Florida, USA. In addition, anaerobic strains Porphyromonas gingivalis W83 and Fusobacterium nucleatum ATCC 49,526 were provided by Dr. Progulske-Fox and Dr. Kesavalu, respectively, at the University of Florida College of Dentistry in Gainesville, Florida, USA.
Oral streptococci were cultured in brain heart infusion (BHI) agar or BHI broth supplemented with 5 μg/mL porcine hemin at 37 °C with 5% CO2, as described previously [30,33,43,44]. Anaerobes F. nucleatum and P. gingivalis were grown in Tryptic Soy Agar (TSA) supplemented with 5% sheep’s blood, 1 μg/mL menadione, and in Tryptic Soy Broth (TSB) with 1 mg/mL yeast extract (TSBY) with 1 μg/mL menadione and 5 μg/mL porcine hemin [32,49]. P. gingivalis strain W83 is resistant to 30 μg/mL gentamicin and was routinely grown in TSBY and blood agar with this antibiotic. Anaerobes were cultured in a BACTRON Anaerobic Chamber (Sheldon Manufacturing, Inc., Cornelius, OR, USA) at 37 °C, 90% nitrogen, 5% hydrogen, and 5% CO2.
2.5. Preparation of Commensal Supernatants
S. gordonii, S. intermedius, S. mitis, and S. oralis cultures were grown in BHI at 37 °C, 5% CO2 to the late exponential phase depending on microbial species. Supernatants were adjusted to physiological pH (7.0-7.8) using incremental additions of 1 M NaOH. Cultures were then centrifuged at 15,000 x g for 10 min, and clarified supernatants were filter-sterilized using 0.45 μm filters. Filtered supernatants were aliquoted, labeled, and stored at -80 °C until use in subsequent assays.
2.6. Growth Curves of P. gingivalis with E-Liquids and Oral Commensal Supernatants
2.6.1. P. gingivalis Planktonic Growth
Treatments were prepared by diluting E-liquids or their components (see 2.2) to a final concentration of 2% (v/v) in TSBY. Commensal supernatants (see 2.5) were diluted to 20% (v/v) in TSBY. Combined treatments included both 2% (v/v) E-liquid and 20% (v/v) commensal supernatants in TSBY. All treatment and control media (TSBY alone) were added to 96-well plates at 100 µL per well. P. gingivalis W83 was grown anaerobically on blood agar for two days. P. gingivalis colonies were resuspended in TSBY and adjusted to an absorbance of 0.5 at 595 nm. Then, 100 µL of P. gingivalis suspension was added to the above treatment (and control) media in 96-well plates, yielding a final volume of 200 µL per well. With this, the bacteria’s absorbance is diluted to roughly 0.25, and the treatments are diluted to 1% E-liquids and 10% commensal supernatants. Six wells were used to measure 200 µL of blank media to subtract media absorbance. Cultures were incubated anaerobically at 37 °C, and growth was monitored by measuring absorbance at 595 nm at defined time points up to 30 h using a µQuant monochromatic microplate reader (MTX Lab Systems, Bradenton, FL, USA) equipped with the Gen5 version 1.1 software (BioTek, Winooski, VT, USA). To ensure continuous anaerobic growth of P. gingivalis, each time point was read in its own 96-well plate without returning it to the anaerobic incubator.
2.6.2. P. gingivalis CFU Quantification at 24 Hours of Planktonic Growth
At 24 hours of growth (see 2.6.1), bacterial viability was additionally assessed by colony-forming unit (CFU) counting. Cultures were serially diluted 1:10 up to 1:107 in phosphate-buffered saline (PBS), 10 µL of the 1:105, 1:106, and 1:107 dilutions were spot-plated in triplicate onto blood agar and incubated anaerobically for two days. Colonies were counted using a dissecting microscope.
2.7. Quantification of Multispecies Biofilms Exposed to E-Liquids
2.7.1. Crystal Violet
Salivary pellicles were formed by coating acetone-treated sterile 96-well plates with 100 µL of processed human saliva per well and incubating overnight at 4 °C. Multispecies streptococcal biofilms were formed as previously described [33,43] with a few adjustments. Briefly, S. gordonii, S. intermedius, S. mitis, and S. oralis were grown overnight (see 2.4) and adjusted to the same absorbance of 0.8. The four strains were then mixed at a 1:1:1:1 ratio, yielding a final volume of 12 mL (3 mL each species). Saliva-coated wells were inoculated with 100 µL of the bacteria mixture and incubated for 1 h at 37 °C, 5% CO2 to allow adherence to the surface. Non-adherent bacteria were removed by washing wells three times with 100 µL sterile PBS. Then, 100 µL of 50% TSBY was added to all wells, and the plate was incubated anaerobically for 24 h. F. nucleatum and P. gingivalis were grown anaerobically in TSBY, adjusted to the same absorbance (0.8), and combined at a 1:1 ratio. The pre-established streptococcal biofilms were washed twice with sterile PBS as above, and 100 µL of the anaerobe mixture was added to each biofilm. Co-cultures were incubated for four hours at 37 °C anaerobically to allow for bacterial integration into the streptococcal biofilms. Following integration, wells were washed twice with PBS, and biofilms were exposed to 1% (v/v) E-liquids ± flavors or 5% hydrogen peroxide (Px control) in 50% TSBY and incubated anaerobically at 37 °C for an additional 24 h.
To quantify total biofilm biomass, biofilms were washed three times with PBS, stained with 100 µL of 5% crystal violet for 10 min, and rinsed up to seven times with deionized water. Next, 100 µL of 3% acetic acid was added to all wells, and the plates were shaken at 400 rpm for 1 minute to ensure complete extraction of crystal violet from the bacteria. Acetic acid solution with any crystal violet was passed to a new and clear 96-well plate. Absorbance was measured at 595 nm using the plate reader and Gen5 software (see 2.6), and readings were used as an index of total biofilm biomass.
2.7.2. qPCR
Salivary pellicles were formed by coating acetone-treated sterile 12-well plates with 2 mL of processed human saliva per well and incubating overnight at 4 °C. S. gordonii, S. intermedius, S. mitis, and S. oralis multispecies biofilms were established as above (see 2.7.1), with a final volume of 2 mL TSBY per well, anaerobically for 24 h. P. gingivalis and F. nucleatum were grown, mixed, and integrated to streptococcal biofilms in a final volume of 2 mL as described above (see 2.7.1). Following integration, biofilms were grown with 1% (v/v) E-liquids ± flavors or 5% Px or control for 24 as indicated above (see 2.7.1).
To quantify total bacterial DNA and P. gingivalis W83 DNA, biofilm DNA was collected using the DNeasy Blood & Tissue Kit from QIAGEN (REF: 69506) following the manufacturer’s instructions. The isolated DNA was stored in microcentrifuge tubes at -20℃. TaqMan assays at 20X for 16S with the VIC fluorescent reporter and PG0717 with the FAM fluorescent reporter, along with the fast 2X master mix, were used for quantitative Polymerase Chain Reaction (qPCR). The StepOnePlus Real-Time PCR machine (Applied Biosystems, Foster City, CA, USA) was used to run 50 cycles, with each cycle spending 5 seconds at 95℃, and 20 seconds at 60℃. Ct values were analyzed using the 2-ΔCt method.
2.7.3. Biofilm Sonication for Bacterial Dispersal and Subsequent CFU Counting
Multispecies biofilms were grown and treated with E-liquids as described above (see 2.7.2). Following treatments, media were removed, and the wells were washed twice with sterile PBS. Then, 3 mL PBS was added to the biofilms in the wells. Bacteria were dissociated from biofilms via probe sonication (Hielscher Ultrasonics USA, Inc. West Milford, NJ, USA) while keeping the plates on ice. Prior to each sonication session, the probe was cleaned with ethanol. The probe was submerged in PBS in each well without contacting the plate surface, and the samples were sonicated at 80% amplitude for three 10-second pulses separated by 15-second rest intervals. The resulting single cell suspensions were serially diluted 1:10 up to 1:104 in PBS, and 10 µL from each dilution were spot-plated in triplicate. For commensal streptococci, suspensions were plated onto BHI agar, then incubated aerobically at 37 °C in 5% CO2 for 24 h. For P. gingivalis, suspensions were plated onto blood agar and incubated anaerobically at 37 °C for 48 h prior to colony counting.
2.8. Statistical Analysis
All data are reported as means ± standard error of the means (SEM). All comparisons within line graphs (planktonic growth curves) and bar graphs (CFU, crystal violet and qPCR assays) were made using two-way ANOVA and one-way ANOVA, respectively, followed by Bonferroni multiple comparisons test. Statistical significance is indicated when p < 0.05. Version 5 of Prism (GraphPad Software, San Diego, CA, USA) was used to generate all graphs and perform all statistical tests.
3. Results
3.1. Effect of E-Liquid Components on P. gingivalis Planktonic Growth
P. gingivalis planktonic growth, beginning at 8 hours, was significantly inhibited when exposed to 1% E-liquids ± flavors (Figure 1). The E-liquid humectant propylene glycol and the mixture of glycerol and propylene glycol also demonstrate a similar significant inhibitory effect, but to a lesser extent. Glycerol alone shows a significant inhibition only at 30 hours (Figure 1). These results suggest that E-liquid humectants and flavors inhibit the planktonic growth of P. gingivalis, with flavoring agents having the most drastic effects.
3.2. Effects of E-Liquids and Individual Commensal Supernatants on P. gingivalis Planktonic Growth
To test the putative beneficial effects of oral commensal streptococcal supernatants, P. gingivalis planktonic growth was measured in the presence of both the E-liquids (menthol and cinnamon) and commensal supernatants (Figure 2). The growth of P. gingivalis is differentially influenced by the supernatants of oral commensal streptococci, especially with S. mitis supernatant. In addition, S. gordonii and S. oralis supernatants also decrease P. gingivalis growth, but to a lower degree (Figure 2A). In contrast, no appreciable effect is observed with S. intermedius supernatant. As expected from Figure 1, both 1% cinnamon and menthol E-liquids severely inhibit P. gingivalis planktonic growth (Figure 2B and 2D). Notably, when P. gingivalis is grown in the presence of both commensal supernatants and E-liquids, growth is slightly higher compared to E-liquid treatments alone (Figure 2C and 2E). Supernatants from S. gordonii, S. intermedius, and S. mitis, but not S. oralis, significantly increase P. gingivalis growth in the presence of menthol-flavored E-liquid between 24 and 30 hours. In addition, all four commensal supernatants increase P. gingivalis growth in the presence of cinnamon-flavored E-liquid between 20 and 30 hours. Overall, commensal supernatants appear to limit the suppressive effects of the E-liquids, with a slightly greater effect associated with the menthol E-liquid (Figure 2). These results suggest that unknown metabolites in the supernatants from oral commensals can partially mitigate the toxicity of flavored E-liquids per se, thereby modulating P. gingivalis proliferation.
3.3. Effects of E-Liquids and Mixed Commensal Supernatants on P. gingivalis Planktonic Growth
The growth of P. gingivalis is monitored for 30 hours under conditions of exposure to E-liquids with and without mixed commensal supernatants. As expected, all E-liquids tested delay planktonic growth kinetics significantly compared to control, with the most significant inhibitions observed with cinnamon and menthol flavors (Figure 3). In the presence of mixed commensal supernatants, P. gingivalis exhibits growth rates similar to control. However, when P. gingivalis growth is delayed by E-liquids, mixed commensal supernatants do not significantly improve growth kinetics (Figure 3). The results of this experiment indicate that all E-liquids delay the growth of P. gingivalis, as seen in Figure 1, and that mixing the four supernatants together dampens the effects each individual supernatant has on E-liquid inhibition (Figure 2).
3.4. CFU Counts of P. gingivalis at 24 Hour of Planktonic Growth with E-Liquids and Mixed Supernatants
Figure 4 shows P. gingivalis CFUs/mL at the 24-hour time-point of planktonic growth shown in Figure 3. The P. gingivalis control yields 2.7 × 1011 CFUs/mL, but when exposed to 10% mixed commensal supernatants, P. gingivalis CFU/mL nearly doubles to 5.6 × 1011. However, when exposed to E-liquids, CFU counts significantly decrease approximately an order of magnitude, except for when exposed to strawberry E-liquid (Figure 4). P. gingivalis CFU counts in the presence of mixed supernatants from all four commensals and E-liquids also show a significant decrease compared to control and control with mixed supernatants. CFU counts are not significantly different between treatments with E-liquids alone as compared to their respective E-liquid plus supernatants. These results indicate that all E-liquids significantly decrease the planktonic growth of P. gingivalis and that the mixed commensal supernatants do not alter these effects.
3.5. Quantification of E-Liquid Effects on Multispecies Biofilm Biomass via Crystal Violet Assay
The crystal violet assay was employed to test the effects of E-liquids on total biofilm biomass of oral multispecies biofilms, including the four gram-positive commensals and the two gram-negative anaerobes. Figure 5 displays that biofilm biomass of oral bacteria is significantly affected by 1% E-liquids with cinnamon and blueberry compared to the control, but not with the other flavor treatments. The hydrogen peroxide treatment is used as a control and causes an expected significant reduction in biofilm biomass. These results indicate that at least some E-liquids tested reduce the total biofilm biomass.
3.6. Quantification of E-Liquid Effects on Multispecies Biofilm Biomass via qPCR
To test the effects of E-liquids on total bacterial DNA and P. gingivalis DNA within the multispecies biofilms, a qPCR assay was performed. Total bacterial DNA (Figure 6A) and P. gingivalis DNA (Figure 6B) quantifications yield no significant differences between treatment groups and their respective controls. These data indicate that levels of both bacterial and P. gingivalis DNA are not significantly altered following E-liquid treatments on biofilms. This could be due to DNA quantification accounting for both live and dead bacteria as well as DNA found in the slime layer surrounding biofilms.
3.7. Quantification of E-Liquid Effects on Multispecies Biofilm Biomass via CFU Counting
To test the effects of E-liquids on viable commensals and P. gingivalis within the multispecies biofilms following 1% E-liquid treatments, a CFU viability test was performed. Across all E-liquid conditions, both commensals (Figure 7A) and P. gingivalis (Figure 7B) show increased CFU counts compared with controls, but only the treatment with menthol E-liquid significantly increases viable counts. The results indicate that P. gingivalis and commensal bacteria CFUs are slightly higher in biofilms when exposed to E-liquids.
3.8. Comparison of E-Liquid Effects on P. gingivalis Growth Planktonically and in Multispecies Biofilms
To compare the effects of E-liquids on P. gingivalis growth, both planktonically and in multispecies biofilms, the CFUs from Figure 4 and Figure 7B are compared side by side and expressed as percent of control in Table 1. Under planktonic conditions, P. gingivalis growth is severely decreased with percent values ranging between 4% and 27% after exposure to 1% E-liquids. On the other hand, in multispecies biofilms exposed to 1% E-liquids, the relative percentages of P. gingivalis CFUs are similar and in some cases higher than the control. For example, menthol E-liquid treatment induces an 87% increase over control while flavorless and blueberry E-liquids yield a 42% increase. Taken altogether, these results indicate that E-liquids favor P. gingivalis growth in oral multispecies biofilms.
4. Discussion
Homeostasis in the oral cavity is maintained by interactions between oral microbes and the host [50,51]. P. gingivalis often enters as an opportunistic invader [52,53,54]. Studying P. gingivalis in multispecies biofilms, such as the species in this study, offers a more realistic model of oral microbiology. Our results show that E-liquids are detrimental to P. gingivalis when isolated and grown planktonically. However, when P. gingivalis is exposed to E-liquids as part of a multispecies biofilm its growth enhanced.
When P. gingivalis is grown planktonically, exposure to E-liquids ± flavors induces a dramatic decrease in growth and CFU production (Figure 1 and Figure 4), but in the presence of commensal supernatants, the toxicity of the flavoring agents is mitigated, thereby promoting P. gingivalis growth (Figure 2). These results indicate that commensal bacteria may provide protective mediators or metabolites that partially counteract the toxic effects of E-liquids. It is for this reason that the effects of E-liquids on P. gingivalis were tested in the context of multispecies biofilms. Within these microbial communities, commensal bacteria may alter the microenvironment by providing metabolites that neutralize the E-liquid toxicity. Furthermore, because of the anaerobic nature of P. gingivalis, this species typically tends to live in the deeper layers of oral biofilms, away from toxic agents, such as E-liquids and their components, which dilute to sub-lethal concentrations as they approach the P. gingivalis microenvironment.
Figure 1 suggests that E-liquid components inhibit P. gingivalis growth, particularly flavored E-liquids such as cinnamon, blueberry, and menthol. Meanwhile, E-liquid humectants, propylene glycol and glycerol, affect P. gingivalis growth to a lesser extent, suggesting that the flavoring agents are the components of E-liquids that are most disruptive to P. gingivalis growth. This is consistent with previous literature suggesting that exposure to flavored E-liquids has a dose-dependent inhibitory effect on the planktonic growth of oral bacteria [48]. The present investigation aligns with other studies suggesting that flavored E-liquids inhibit the growth of oral bacteria similar to aerosols [55].
Figure 2A demonstrates that commensal supernatants, especially those of S. mitis, decrease the growth of P. gingivalis, which agrees with others. For example, S. mitis produces an abundant amount of hydrogen peroxide in comparison to the other commensals, inhibiting the biofilm formation of cariogenic species Streptococcus mutans [56]. In addition to hydrogen peroxide, commensal bacteria play a protective role in maintaining microbial balance by producing competitive scavenging metabolites, reactive nitrogen intermediates, and bacteriocins, all of which help protect against pathogenic species [57]. However, supernatants from commensal species seem to partially enhance P. gingivalis planktonic growth when exposed to menthol and cinnamon E-liquids (Figure 2C and 2E). Previous studies from our group have shown that cinnamon and menthol flavors have the greatest inhibitory impact on the growth of oral bacteria [35,47,48]. This study displays the same trend with P. gingivalis, as seen in Figure 1, and hence these two flavors were used to test the effects of individual commensal supernatants in Figure 2. Molecular approaches indicate that the ecological balance in the oral cavity is maintained through antagonistic and mutualistic interactions among species [28,58,59,60]. Such interactions are mediated by metabolites [28,51,59,60,61,62] that could also participate in the tolerance to external toxic materials [63], such as E-liquids. The materials responsible for such beneficial effects remain unknown and should be investigated.
The protective effects noticed by the four individual commensal supernatants (Figure 2) are lost when mixed at a 1:1:1:1 ratio (Figure 3 and Figure 4). Likely, such protective materials in the supernatants either fall below the threshold of activity when diluted 1:4, or unknown biochemical interactions change their functions. Although this study tests a small number of flavors compared to the plethora of flavors available [64], the flavors tested are well-represented in the scientific literature and help us understand the tolerance of P. gingivalis by itself to these external agents.
Six species were chosen to grow in vitro oral biofilms, and test the viability of P. gingivalis in this context. In addition to all four commensals, F. nucleatum was included because it is required as a bridging species between P. gingivalis and the four commensal streptococci [29]. Most flavors, except cinnamon and blueberry, do not significantly alter biofilm biomass (Figure 5). Despite the reductions in the cinnamon and blueberry treatments, the overall magnitude of biomass reduction across treatments is minimal. These findings extend prior research showing that vaping induces variable effects on oral bacteria, including commensal streptococci, depending on the dose, physical state (liquid vs. aerosol) and flavors [35,47,48,55]. The reduced biomass may be attributable to the E-liquid components or their flavorings. For example, trans-cinnamaldehyde, a component of the cinnamon flavor, can inhibit biofilm formation of S. mutans [65]. Crystal violet staining quantifies total biomass, making it difficult to determine the exact biofilm composition.
Biofilms exposed to E-liquids show minimal differences relative to the control in terms of both 16S (total bacteria) and PG0717 (P. gingivalis only) DNA (Figure 6). These results indicate that the biofilm microenvironment offers substantial protection to P. gingivalis from E-liquids. While individual commensal supernatants partially protect P. gingivalis against E-liquids (Figure 2), biofilm communities offer a greater degree of protection, not only to P. gingivalis but to the commensals as well. Despite the established bactericidal properties of E-liquids [47], to our knowledge this is the first report of vaping-induced increase of P. gingivalis in the context of polymicrobial oral biofilms in vitro, and is consistent with studies that indicate that vaping contributes to dysbiosis in oral biofilms [66].
E-liquid treatments yield a modest increase in CFU counts from multispecies biofilms for both commensal streptococci (Figure 7A) and P. gingivalis (Figure 7B) relative to the untreated control. In contrast to planktonic results (Figure 1 to 4), as well as previous studies reporting antimicrobial/antibiofilm effects of flavored E-liquids on oral bacteria and streptococcal communities [35,48], these data suggest that biofilms mitigate E-liquid toxicity compared to planktonic growth, consequently yielding higher CFU counts. Even though total biofilm biomass (Figure 5) and DNA levels (Figure 6) show minimal changes after E-liquid exposure, it is likely that multispecies biofilms may specifically increase the number of CFUs (Figure 7) rather than increasing overall biofilm mass, slime layer, dead cells, or total DNA. In other words, E-liquids may increase CFUs within the biofilms (i.e., live material) and not the slime layer or other non-living materials, without producing a substantial change in crystal violet staining or DNA readouts. Notably, menthol stands out as the only treatment causing a significant CFU increase for both commensals and P. gingivalis (Figure 7). This provides a basis for why polymicrobial interactions in a multispecies biofilm could alter responses to the same stressor. This supports the broader idea that polymicrobial interactions and metabolite exchange within oral biofilms can change how stressors affect individual species, like P. gingivalis [57,61,67].
Our findings demonstrate that while E-liquid flavoring agents are directly toxic to P. gingivalis in planktonic culture, individual commensal supernatants modestly improved growth of this anaerobe. Table 1 illustrates the striking contrast between planktonic and biofilm growth: while E-liquids reduce P. gingivalis viability during planktonic growth, multispecies biofilms conditions reverse this effect, with P. gingivalis CFUs exceeding the control levels across all tested flavors. Therefore, when multispecies oral biofilms are exposed to E-liquids, P. gingivalis thrives. This helps explain clinical observations reporting that vapers house elevated P. gingivalis levels and face an increased risk of periodontal disease compared to non-vapers [42,66]. Vapers exhibit a unique periodontal microbiome characterized by the increase of pathogenic species, alongside increased inflammatory markers, and other clinical signs of periodontitis[43]. The enhanced P. gingivalis colonization observed in vapers has serious systemic implications as the pathogen has been linked to cardiovascular disease, arthritis risk, and diabetes [45,46].
5. Conclusions
In conclusion, this study demonstrates that when P. gingivalis is exposed to E-liquids in a biofilm, greater growth is observed than when exposed to E-liquids alone. This study confirms that E-liquids significantly alter microbial composition and provide a controlled in vitro rendition of the effects of vaping, supporting in vivo reports of the same kind. E-liquids disturb oral biofilms and seem to increase dysbiosis and, by extension, susceptibility to oral and systemic disease. The broader health implications of these findings warrant consideration, as disruptions in oral microbial communities can impact systemic health.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, G.C; methodology, G.C.; software, GC and DP; validation, GC and DP; formal analysis, DP; investigation, SS, JA, DB, DB; resources, GC; data curation, GC; writing—original draft preparation, SS, JA, DB, DB; writing—review and editing, GC and DP; visualization, GC and DP; supervision, GC; project administration, GC; funding acquisition, GC. All authors have read and agreed to the published version of the manuscript.
Funding
Funding was procured from intramural grants at Muhlenberg College.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board of Muhlenberg College (IRB approval code Cuadra_S19_18, 13 May 2019).
Informed Consent Statement
Verbal informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The contributions presented in the study are included in the publication. Any further questions or inquiries can be directed to the corresponding author.
Acknowledgments
Authors would like to thank Jonathan Jarrah, Sara Gill, and Angela Mamari for their technical assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Selekman, J. Vaping: It’s All a Smokescreen—ProQuest Available online: https://www.proquest.com/docview/2184907265?parentSessionId=4ogZxMBzh2Lk87ECvsLC4jJLiQ6qMS%2Bbx%2F0J9sfUzyU%3D&sourcetype=Scholarly%20Journals (accessed on 23 July 2024).
- Glantz, S.A.; Bareham, D.W. E-Cigarettes: Use, Effects on Smoking, Risks, and Policy Implications. Annu Rev Public Health 2018, 39, 215–235. [CrossRef]
- Palazzolo, D.L. Electronic Cigarettes and Vaping: A New Challenge in Clinical Medicine and Public Health. A Literature Review. Front. Public Health 2013, 1. [CrossRef]
- Famiglietti, A.; Memoli, J.W.; Khaitan, P.G. Are Electronic Cigarettes and Vaping Effective Tools for Smoking Cessation? Limited Evidence on Surgical Outcomes: A Narrative Review. J Thorac Dis 2021, 13, 384–395. [CrossRef]
- Besaratinia, A.; Tommasi, S. Vaping: A Growing Global Health Concern. EClinicalMedicine 2019, 17, 100208. [CrossRef]
- Lyzwinski, L.N.; Naslund, J.A.; Miller, C.J.; Eisenberg, M.J. Global Youth Vaping and Respiratory Health: Epidemiology, Interventions, and Policies. npj Prim. Care Respir. Med. 2022, 32, 14. [CrossRef]
- Eaton, D.; Kwan, L.; Stratton, K. Public Health Consequences of E-Cigarettes; The National Academies Press: Washington DC, 2018; ISBN 978-0-309-46834-3.
- Hajek, P.; Etter, J.-F.; Benowitz, N.; Eissenberg, T.; McRobbie, H. Electronic Cigarettes: Review of Use, Content, Safety, Effects on Smokers and Potential for Harm and Benefit. Addiction 2014, 109, 1801–1810. [CrossRef]
- Ghosh, S.; Drummond, M.B. Electronic Cigarettes as Smoking Cessation Tool: Are We There? Curr Opin Pulm Med 2017, 23, 111–116. [CrossRef]
- While Less Harmful than Cigarettes, e-Cigarettes Still Pose Risks Available online: https://truthinitiative.org/research-resources/emerging-tobacco-products/while-less-harmful-cigarettes-e-cigarettes-pose (accessed on 8 January 2026).
- Cuadra, G.; Hanel, A.; Christian, N.; Pecorelli, S.; Ha, V.; Tomov, S.; Palazzolo, D.; Cuadra, G.; Hanel, A.; Christian, N.; et al. Perspective Chapter: The Impact of Electronic Cigarettes on the Oral Microenvironment; IntechOpen, 2025; ISBN 978-1-83635-438-3.
- Izquierdo-Condoy, J.S.; Naranjo-Lara, P.; Morales-Lapo, E.; Hidalgo, M.R.; Tello-De-la-Torre, A.; Vásconez-Gonzáles, E.; Salazar-Santoliva, C.; Loaiza-Guevara, V.; Rincón Hernández, W.; Becerra, D.A.; et al. Direct Health Implications of E-Cigarette Use: A Systematic Scoping Review with Evidence Assessment. Front. Public Health 2024, 12. [CrossRef]
- Holliday, R.; Kist, R.; Bauld, L. E-Cigarette Vapour Is Not Inert and Exposure Can Lead to Cell Damage. Evid Based Dent 2016, 17, 2–3. [CrossRef]
- Muthumalage, T.; Prinz, M.; Ansah, K.O.; Gerloff, J.; Sundar, I.K.; Rahman, I. Inflammatory and Oxidative Responses Induced by Exposure to Commonly Used E-Cigarette Flavoring Chemicals and Flavored e-Liquids without Nicotine. Front Physiol 2018, 8, 1130. [CrossRef]
- Clapp, P.W.; Lavrich, K.S.; van Heusden, C.A.; Lazarowski, E.R.; Carson, J.L.; Jaspers, I. Cinnamaldehyde in Flavored E-Cigarette Liquids Temporarily Suppresses Bronchial Epithelial Cell Ciliary Motility by Dysregulation of Mitochondrial Function. Am J Physiol Lung Cell Mol Physiol 2019, 316, L470–L486. [CrossRef]
- Shamim, A.; Herzog, H.; Shah, R.; Pecorelli, S.; Nisbet, V.; George, A.; Cuadra, G.A.; Palazzolo, D.L. Pathophysiological Responses of Oral Keratinocytes After Exposure to Flavored E-Cigarette Liquids. Dentistry Journal 2025, 13, 60. [CrossRef]
- Shah, R.; Luo, E.D.; Shaffer, C.A.; Tabakha, M.; Tomov, S.; Minton, S.H.; Brown, M.K.; Palazzolo, D.L.; Cuadra, G.A. Polarization of THP-1-Derived Human M0 to M1 Macrophages Exposed to Flavored E-Liquids. Toxics 2025, 13, 451. [CrossRef]
- Ramenzoni, L.L.; Schneider, A.; Fox, S.C.; Meyer, M.; Meboldt, M.; Attin, T.; Schmidlin, P.R. Cytotoxic and Inflammatory Effects of Electronic and Traditional Cigarettes on Oral Gingival Cells Using a Novel Automated Smoking Instrument: An In Vitro Study. Toxics 2022, 10, 179. [CrossRef]
- Shaikh, Z.N.; Alqahtani, A.; Almela, T.; Franklin, K.; Tayebi, L.; Moharamzadeh, K. Effects of Electronic Cigarette Liquid on Monolayer and 3D Tissue-Engineered Models of Human Gingival Mucosa. J Adv Periodontol Implant Dent 2019, 11, 54–62. [CrossRef]
- Palazzolo, D.L.; Nelson, J.M.; Ely, E.A.; Crow, A.P.; Distin, J.; Kunigelis, S.C. The Effects of Electronic Cigarette (ECIG)-Generated Aerosol and Conventional Cigarette Smoke on the Mucociliary Transport Velocity (MTV) Using the Bullfrog (R. Catesbiana) Palate Paradigm. Front Physiol 2017, 8, 1023. [CrossRef]
- Allbright, K.; Villandre, J.; Crotty Alexander, L.E.; Zhang, M.; Benam, K.H.; Evankovich, J.; Königshoff, M.; Chandra, D. The Paradox of the Safer Cigarette: Understanding the Pulmonary Effects of Electronic Cigarettes. Eur Respir J 2024, 63, 2301494. [CrossRef]
- Palazzolo, D.L.; Crow, A.P.; Nelson, J.M.; Johnson, R.A. Trace Metals Derived from Electronic Cigarette (ECIG) Generated Aerosol: Potential Problem of ECIG Devices That Contain Nickel. Front Physiol 2016, 7, 663. [CrossRef]
- HOMD: Human Oral Microbiome Database Available online: https://www.homd.org/# (accessed on 28 May 2025).
- Damyanova, T.; Paunova-Krasteva, T. What We Still Don’t Know About Biofilms—Current Overview and Key Research Information. Microbiology Research 2025, 16, 46. [CrossRef]
- Deo, P.N.; Deshmukh, R. Oral Microbiome: Unveiling the Fundamentals. J Oral Maxillofac Pathol 2019, 23, 122–128. [CrossRef]
- Kolenbrander, P.E.; Andersen, R.N.; Blehert, D.S.; Egland, P.G.; Foster, J.S.; Palmer, R.J. Communication among Oral Bacteria. Microbiol. Mol. Biol. Rev. 2002, 66, 486–505, table of contents.
- Jenkinson, H.F.; Lamont, R.J. Oral Microbial Communities in Sickness and in Health. Trends Microbiol 2005, 13, 589–595. [CrossRef]
- Periasamy, S.; Kolenbrander, P.E. Mutualistic Biofilm Communities Develop with Porphyromonas Gingivalis and Initial, Early, and Late Colonizers of Enamel. J. Bacteriol. 2009, 191, 6804–6811. [CrossRef]
- Sakanaka, A.; Kuboniwa, M.; Shimma, S.; Alghamdi, S.A.; Mayumi, S.; Lamont, R.J.; Fukusaki, E.; Amano, A. Fusobacterium Nucleatum Metabolically Integrates Commensals and Pathogens in Oral Biofilms. mSystems 2022, 7, e0017022. [CrossRef]
- Dong, Y.; Chen, Y.-Y.M.; Snyder, J.A.; Burne, R.A. Isolation and Molecular Analysis of the Gene Cluster for the Arginine Deiminase System from Streptococcus Gordonii DL1. Appl Environ Microbiol 2002, 68, 5549–5553. [CrossRef]
- Redanz, S.; Cheng, X.; Giacaman, R.A.; Pfeifer, C.S.; Merritt, J.; Kreth, J. Live and Let Die: Hydrogen Peroxide Production by the Commensal Flora and Its Role in Maintaining a Symbiotic Microbiome. Mol Oral Microbiol 2018, 33, 337–352. [CrossRef]
- Hanel, A.N.; Herzog, H.M.; James, M.G.; Cuadra, G.A. Effects of Oral Commensal Streptococci on Porphyromonas Gingivalis Invasion into Oral Epithelial Cells. Dentistry Journal 2020, 8, 39. [CrossRef]
- Cuadra, G.A.; Smith, M.T.; Nelson, J.M.; Loh, E.K.; Palazzolo, D.L. A Comparison of Flavorless Electronic Cigarette-Generated Aerosol and Conventional Cigarette Smoke on the Survival and Growth of Common Oral Commensal Streptococci. Int J Environ Res Public Health 2019, 16, e1669. [CrossRef]
- Nelson, J.M.; Cuadra, G.A.; Palazzolo, D.L. A Comparison of Flavorless Electronic Cigarette-Generated Aerosol and Conventional Cigarette Smoke on the Planktonic Growth of Common Oral Commensal Streptococci. Int J Environ Res Public Health 2019, 16. [CrossRef]
- Xu, C.P.; Palazzolo, D.L.; Cuadra, G.A. Mechanistic Effects of E-Liquids on Biofilm Formation and Growth of Oral Commensal Streptococcal Communities: Effect of Flavoring Agents. Dent J (Basel) 2022, 10, 85. [CrossRef]
- Thomas, S.C.; Xu, F.; Pushalkar, S.; Lin, Z.; Thakor, N.; Vardhan, M.; Flaminio, Z.; Khodadadi-Jamayran, A.; Vasconcelos, R.; Akapo, A.; et al. Electronic Cigarette Use Promotes a Unique Periodontal Microbiome. mBio 13, e00075-22. [CrossRef]
- Cichońska, D.; Kusiak, A.; Goniewicz, M.L. The Impact of E-Cigarettes on Oral Health—A Narrative Review. Dentistry Journal 2024, 12, 404. [CrossRef]
- Aldakheel, F.M.; Alduraywish, S.A.; Jhugroo, P.; Jhugroo, C.; Divakar, D.D. Quantification of Pathogenic Bacteria in the Subgingival Oral Biofilm Samples Collected from Cigarette-Smokers, Individuals Using Electronic Nicotine Delivery Systems and Non-Smokers with and without Periodontitis. Arch Oral Biol 2020, 117, 104793. [CrossRef]
- Tabnjh, A.K.; Alizadehgharib, S.; Campus, G.; Lingström, P. The Effects of Electronic Smoking on Dental Caries and Proinflammatory Markers: A Systematic Review and Meta-Analysis. Front Oral Health 2025, 6, 1569806. [CrossRef]
- Gaur, S.; Agnihotri, R. The Role of Electronic Cigarettes in Dental Caries: A Scoping Review. Scientifica (Cairo) 2023, 2023, 9980011. [CrossRef]
- Alkattan, R.; Tashkandi, N.; Mirdad, A.; Ali, H.T.; Alshibani, N.; Allam, E. Effects of Electronic Cigarettes on Periodontal Health: A Systematic Review and Meta-Analysis. International Dental Journal 2025, 75, 2014–2024. [CrossRef]
- Figueredo, C.A.; Abdelhay, N.; Figueredo, C.M.; Catunda, R.; Gibson, M.P. The Impact of Vaping on Periodontitis: A Systematic Review. Clin Exp Dent Res 2021, 7, 376–384. [CrossRef]
- Pushalkar, S.; Paul, B.; Li, Q.; Yang, J.; Vasconcelos, R.; Makwana, S.; González, J.M.; Shah, S.; Xie, C.; Janal, M.N.; et al. Electronic Cigarette Aerosol Modulates the Oral Microbiome and Increases Risk of Infection. iScience 2020, 23, 100884. [CrossRef]
- Yang, I.; He, X.; Jeon, J.; Claussen, H.; Arthur, R.; Cushenan, P.; Weaver, S.R.; Luo, R.; Black, M.; Shannahan, J.; et al. The Impact of Vaping Behavior on Functional Changes within the Subgingival Microbiome. Sci Rep 2025, 15, 34374. [CrossRef]
- Hajishengallis, G. Periodontitis: From Microbial Immune Subversion to Systemic Inflammation. Nat Rev Immunol 2015, 15, 30–44. [CrossRef]
- Mealey, B.L. Influence of Periodontal Infections on Systemic Health. Periodontol. 2000 1999, 21, 197–209.
- Christian, N.; Burden, D.; Emam, A.; Brenk, A.; Sperber, S.; Kalu, M.; Cuadra, G.; Palazzolo, D. Effects of E-Liquids and Their Aerosols on Biofilm Formation and Growth of Oral Commensal Streptococcal Communities: Effect of Cinnamon and Menthol Flavors. Dent J (Basel) 2024, 12, 232. [CrossRef]
- Fischman, J.S.; Sista, S.; Lee, D.; Cuadra, G.A.; Palazzolo, D.L. Flavorless vs. Flavored Electronic Cigarette-Generated Aerosol and E-Liquid on the Growth of Common Oral Commensal Streptococci. Front Physiol 2020, 11, 585416. [CrossRef]
- Werheim, E.R.; Senior, K.G.; Shaffer, C.A.; Cuadra, G.A. Oral Pathogen Porphyromonas Gingivalis Can Escape Phagocytosis of Mammalian Macrophages. Microorganisms 2020, 8, 1432. [CrossRef]
- Liu, Y.; Qv, W.; Ma, Y.; Zhang, Y.; Ding, C.; Chu, M.; Chen, F. The Interplay between Oral Microbes and Immune Responses. Front. Microbiol. 2022, 13. [CrossRef]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The Oral Microbiota: Dynamic Communities and Host Interactions. Nat Rev Microbiol 2018, 16, 745–759. [CrossRef]
- Mysak, J.; Podzimek, S.; Sommerova, P.; Lyuya-Mi, Y.; Bartova, J.; Janatova, T.; Prochazkova, J.; Duskova, J. Porphyromonas Gingivalis: Major Periodontopathic Pathogen Overview. Journal of Immunology Research 2014, 2014, 476068. [CrossRef]
- Tribble, G.D.; Lamont, R.J. Bacterial Invasion of Epithelial Cells and Spreading in Periodontal Tissue. Periodontology 2000 2010, 52, 68–83. [CrossRef]
- How, K.Y.; Song, K.P.; Chan, K.G. Porphyromonas Gingivalis: An Overview of Periodontopathic Pathogen below the Gum Line. Front. Microbiol. 2016, 7. [CrossRef]
- Catala-Valentin, A.; Bernard, J.N.; Caldwell, M.; Maxson, J.; Moore, S.D.; Andl, C.D. E-Cigarette Aerosol Exposure Favors the Growth and Colonization of Oral Streptococcus Mutans Compared to Commensal Streptococci. Microbiol Spectr 2022, 10, e02421-21. [CrossRef]
- Williams, I.; Tuckerman, J.S.; Peters, D.I.; Bangs, M.; Williams, E.; Shin, I.J.; Kaspar, J.R. A Strain of Streptococcus Mitis Inhibits Biofilm Formation of Caries Pathogens via Abundant Hydrogen Peroxide Production. Applied and Environmental Microbiology 2025, 91, e02192-24. [CrossRef]
- Baty, J.J.; Stoner, S.N.; Scoffield, J.A. Oral Commensal Streptococci: Gatekeepers of the Oral Cavity. J Bacteriol 2022, 204, e0025722. [CrossRef]
- Avila, M.; Ojcius, D.M.; Yilmaz, O. The Oral Microbiota: Living with a Permanent Guest. DNA Cell Biol. 2009, 28, 405–411. [CrossRef]
- Zhou, W.; Huang, S.; Huang, X. Editorial: Mutualistic and Antagonistic Interactions in the Human Oral Microbiome. Front Microbiol 16, 1731807. [CrossRef]
- Kuramitsu, H.K.; He, X.; Lux, R.; Anderson, M.H.; Shi, W. Interspecies Interactions within Oral Microbial Communities. Microbiology and Molecular Biology Reviews 2007, 71, 653–670. [CrossRef]
- Miller, D.P.; Fitzsimonds, Z.R.; Lamont, R.J. Metabolic Signaling and Spatial Interactions in the Oral Polymicrobial Community. J Dent Res 2019, 98, 1308–1314. [CrossRef]
- Kuboniwa, M.; Houser, J.R.; Hendrickson, E.L.; Wang, Q.; Alghamdi, S.A.; Sakanaka, A.; Miller, D.P.; Hutcherson, J.A.; Wang, T.; Beck, D.A.C.; et al. Metabolic Crosstalk Regulates Porphyromonas Gingivalis Colonization and Virulence during Oral Polymicrobial Infection. Nature Microbiology 2017, 2, 1493–1499. [CrossRef]
- Bostanghadiri, N.; Kouhzad, M.; Taki, E.; Elahi, Z.; Khoshbayan, A.; Navidifar, T.; Darban-Sarokhalil, D. Oral Microbiota and Metabolites: Key Players in Oral Health and Disorder, and Microbiota-Based Therapies. Front Microbiol 2024, 15, 1431785. [CrossRef]
- Havermans, A.; Krüsemann, E.J.Z.; Pennings, J.; de Graaf, K.; Boesveldt, S.; Talhout, R. Nearly 20 000 E-Liquids and 250 Unique Flavour Descriptions: An Overview of the Dutch Market Based on Information from Manufacturers. Tob Control 2021, 30, 57–62. [CrossRef]
- Balasubramanian, A.R.; Vasudevan, S.; Shanmugam, K.; Lévesque, C.M.; Solomon, A.P.; Neelakantan, P. Combinatorial Effects of Trans-Cinnamaldehyde with Fluoride and Chlorhexidine on Streptococcus Mutans. J Appl Microbiol 2021, 130, 382–393. [CrossRef]
- Panariello, B.; Dias Panariello, F.; Misir, A.; Barboza, E.P. An Umbrella Review of E-Cigarettes’ Impact on Oral Microbiota and Biofilm Buildup. Pathogens 2025, 14, 578. [CrossRef]
- Smith, T.P.; Clegg, T.; Ransome, E.; Martin-Lilley, T.; Rosindell, J.; Woodward, G.; Pawar, S.; Bell, T. High-Throughput Characterization of Bacterial Responses to Complex Mixtures of Chemical Pollutants. Nat Microbiol 2024, 9, 938–948. [CrossRef]
Figure 1.
Thirty-hour growth curves illustrating the effects of E-liquids or their humectants on P. gingivalis planktonic growth in TSBY. Each point represents the mean ± SEM; n = 7. * = p < 0.05, ** = p < 0.01 and *** = p < 0.001 as compared to P. gingivalis (control). The bubbles and arrows indicate the presence of significance.
Figure 1.
Thirty-hour growth curves illustrating the effects of E-liquids or their humectants on P. gingivalis planktonic growth in TSBY. Each point represents the mean ± SEM; n = 7. * = p < 0.05, ** = p < 0.01 and *** = p < 0.001 as compared to P. gingivalis (control). The bubbles and arrows indicate the presence of significance.
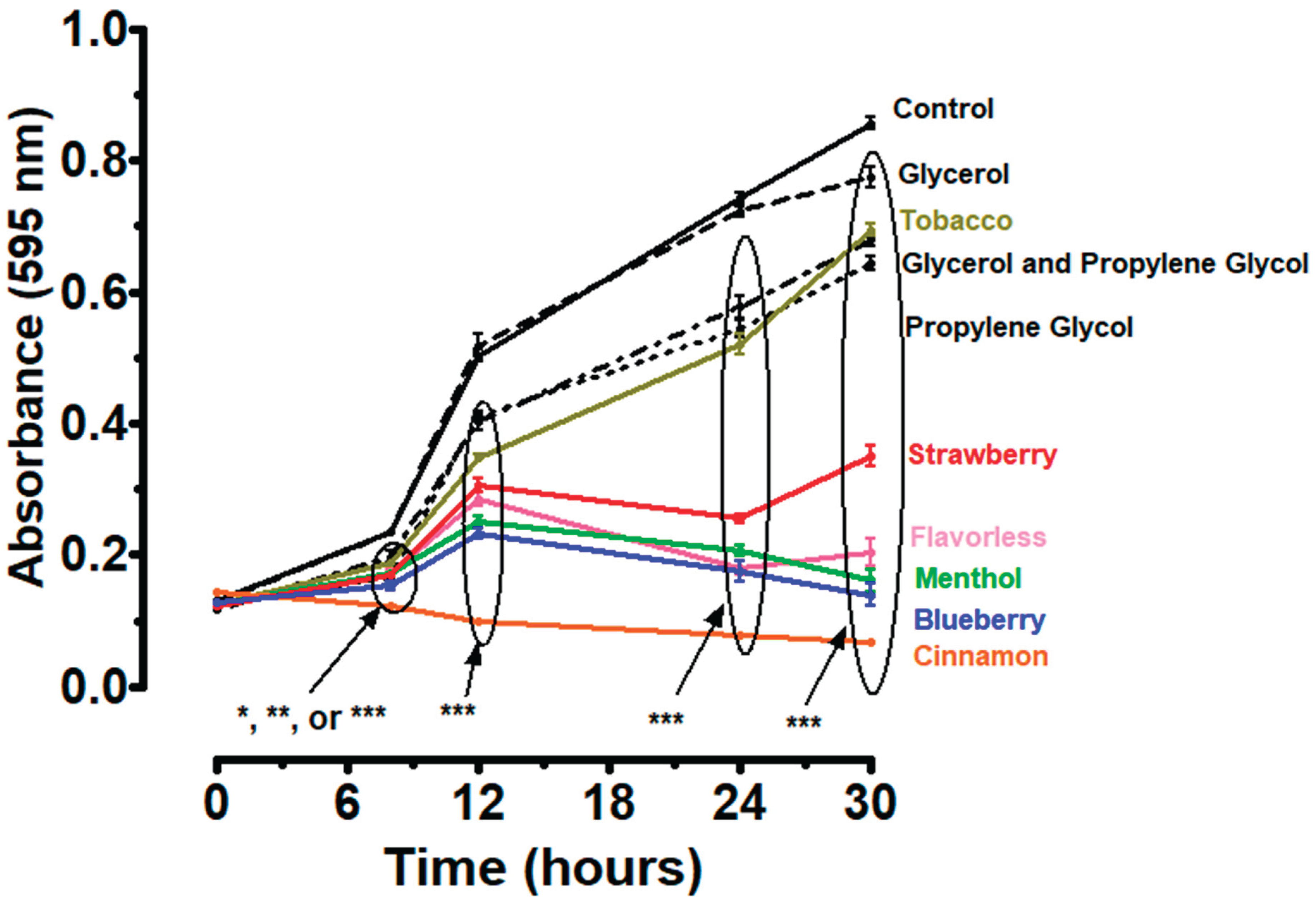
Figure 2.
Thirty-hour growth curves illustrating the effects of individual commensal supernatants and E-liquid treatments on P. gingivalis (Pg) growth. Each point represents the mean ± SEM; n = 4-8. * = p < 0.05, ** = p < 0.01, and *** = p < 0.001 as compared to P. gingivalis control (A, B, D) or as compared to P. gingivalis + menthol (C), or P. gingivalis + cinnamon (E) E-liquids. Supernatants of S. gordonii, S. intermedius, S. mitis, and S. oralis are indicated by Sg, Si, Sm, and So, respectively. Figures C and E are enhanced views from figures B and D, respectively, and indicate comparisons between E-liquids alone and E-liquids with commensal supernatants.
Figure 2.
Thirty-hour growth curves illustrating the effects of individual commensal supernatants and E-liquid treatments on P. gingivalis (Pg) growth. Each point represents the mean ± SEM; n = 4-8. * = p < 0.05, ** = p < 0.01, and *** = p < 0.001 as compared to P. gingivalis control (A, B, D) or as compared to P. gingivalis + menthol (C), or P. gingivalis + cinnamon (E) E-liquids. Supernatants of S. gordonii, S. intermedius, S. mitis, and S. oralis are indicated by Sg, Si, Sm, and So, respectively. Figures C and E are enhanced views from figures B and D, respectively, and indicate comparisons between E-liquids alone and E-liquids with commensal supernatants.

Figure 3.
Thirty-hour growth curves illustrating the effects of mixed commensal supernatants and/or E-liquid (flavorless, tobacco, menthol, cinnamon, strawberry, and blueberry) treatments on P. gingivalis (Pg) growth. Supernatants are a 1:1:1:1 mixture of all four commensal supernatants. Each point represents the mean ± SEM; n = 4-6. * = p < 0.05, ** = p < 0.01 and *** = p < 0.001 as compared to P. gingivalis control.
Figure 3.
Thirty-hour growth curves illustrating the effects of mixed commensal supernatants and/or E-liquid (flavorless, tobacco, menthol, cinnamon, strawberry, and blueberry) treatments on P. gingivalis (Pg) growth. Supernatants are a 1:1:1:1 mixture of all four commensal supernatants. Each point represents the mean ± SEM; n = 4-6. * = p < 0.05, ** = p < 0.01 and *** = p < 0.001 as compared to P. gingivalis control.
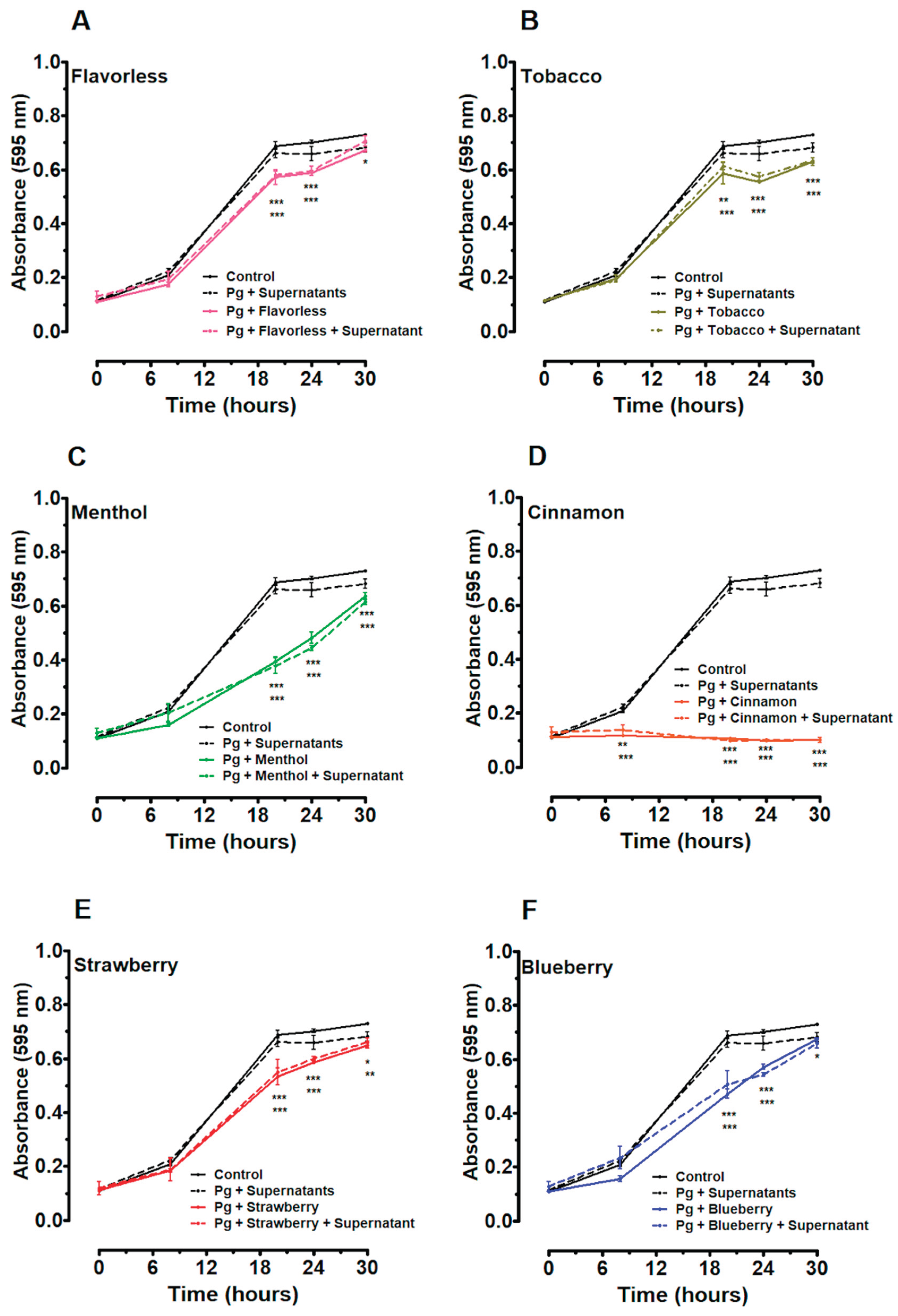
Figure 4.
P. gingivalis CFUs/mL at 24h of planktonic growth. Each value represents the mean ± SEM of CFUs, n = 8 to 12. * = p < 0.05, ** = p < 0.01 and *** = p < 0.001 as compared to P. gingivalis (Pg) control. *** = p < 0.001 as compared to Pg + supernatant. The black and red horizontal lines indicate levels of Pg control and Pg + supernatant, respectively.
Figure 4.
P. gingivalis CFUs/mL at 24h of planktonic growth. Each value represents the mean ± SEM of CFUs, n = 8 to 12. * = p < 0.05, ** = p < 0.01 and *** = p < 0.001 as compared to P. gingivalis (Pg) control. *** = p < 0.001 as compared to Pg + supernatant. The black and red horizontal lines indicate levels of Pg control and Pg + supernatant, respectively.

Figure 5.
Crystal violet assay of multispecies biofilms. Multispecies biofilm biomass after treatments with 1% E-liquids ± flavors in 50% TSBY. Each bar represents the mean ± SEM of absorbance (n = 10 to 12). ** = p < 0.01 and *** = p < 0.001 as compared to control. The black horizontal line indicates the level of control biofilms.
Figure 5.
Crystal violet assay of multispecies biofilms. Multispecies biofilm biomass after treatments with 1% E-liquids ± flavors in 50% TSBY. Each bar represents the mean ± SEM of absorbance (n = 10 to 12). ** = p < 0.01 and *** = p < 0.001 as compared to control. The black horizontal line indicates the level of control biofilms.
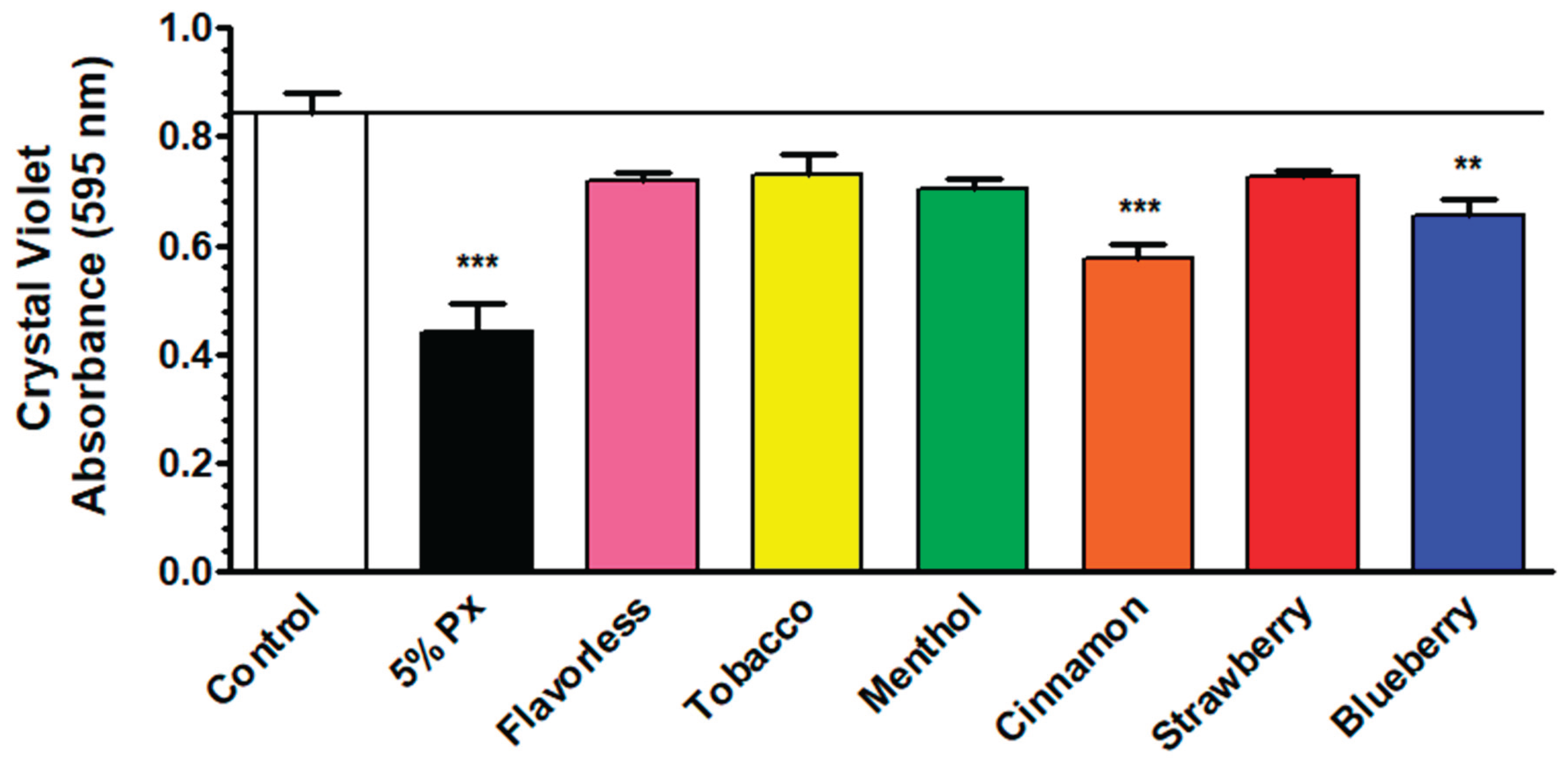
Figure 6.
Quantitative PCR results comparing total bacterial DNA (A) and P. gingivalis DNA (B) from multispecies biofilms after treatment with 1% E-liquids ± flavorings in 50% TSBY. Each bar represents the mean ± SEM of 2-ΔCt (n = 8 to 16). The black horizontal lines indicate the level of control biofilms.
Figure 6.
Quantitative PCR results comparing total bacterial DNA (A) and P. gingivalis DNA (B) from multispecies biofilms after treatment with 1% E-liquids ± flavorings in 50% TSBY. Each bar represents the mean ± SEM of 2-ΔCt (n = 8 to 16). The black horizontal lines indicate the level of control biofilms.

Figure 7.
CFU counts comparing commensal bacteria (A) and P. gingivalis (B) from multispecies biofilms after treatment with 1% E-liquids ± flavorings in 50% TSBY. Each bar represents the mean ± SEM of CFUs/well (n = 13 to 18). * = p < 0.05, ** = p < 0.01, and *** = p < 0.001 as compared to commensal bacteria or P. gingivalis controls. The black horizontal lines indicate the level of control biofilms.
Figure 7.
CFU counts comparing commensal bacteria (A) and P. gingivalis (B) from multispecies biofilms after treatment with 1% E-liquids ± flavorings in 50% TSBY. Each bar represents the mean ± SEM of CFUs/well (n = 13 to 18). * = p < 0.05, ** = p < 0.01, and *** = p < 0.001 as compared to commensal bacteria or P. gingivalis controls. The black horizontal lines indicate the level of control biofilms.
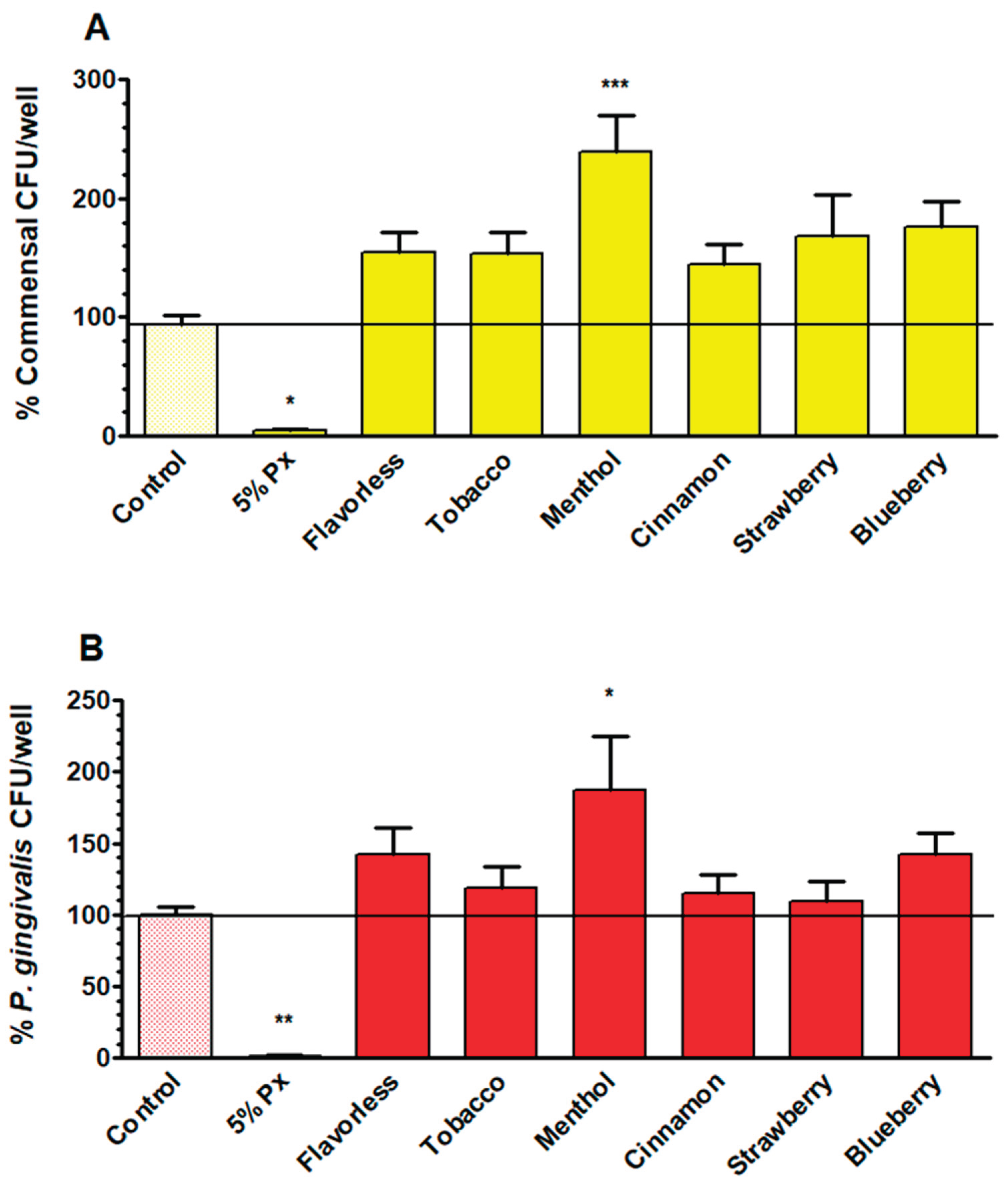

Table 1.
Comparison of P. gingivalis CFUs after Planktonic Growth and Multispecies Biofilm growth while exposed to 1% E-liquids and flavors.
Table 1.
Comparison of P. gingivalis CFUs after Planktonic Growth and Multispecies Biofilm growth while exposed to 1% E-liquids and flavors.
| Control | Flavorless | Tobacco | Menthol | Cinnamon | Strawberry | |
| Planktonic* | 100 | 6 | 14 | 11 | 4 | 27 |
| Biofilm** | 100 | 142 | 119 | 187 | 115 | 109 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



