
Submitted:
19 January 2026
Posted:
20 January 2026
You are already at the latest version
Abstract
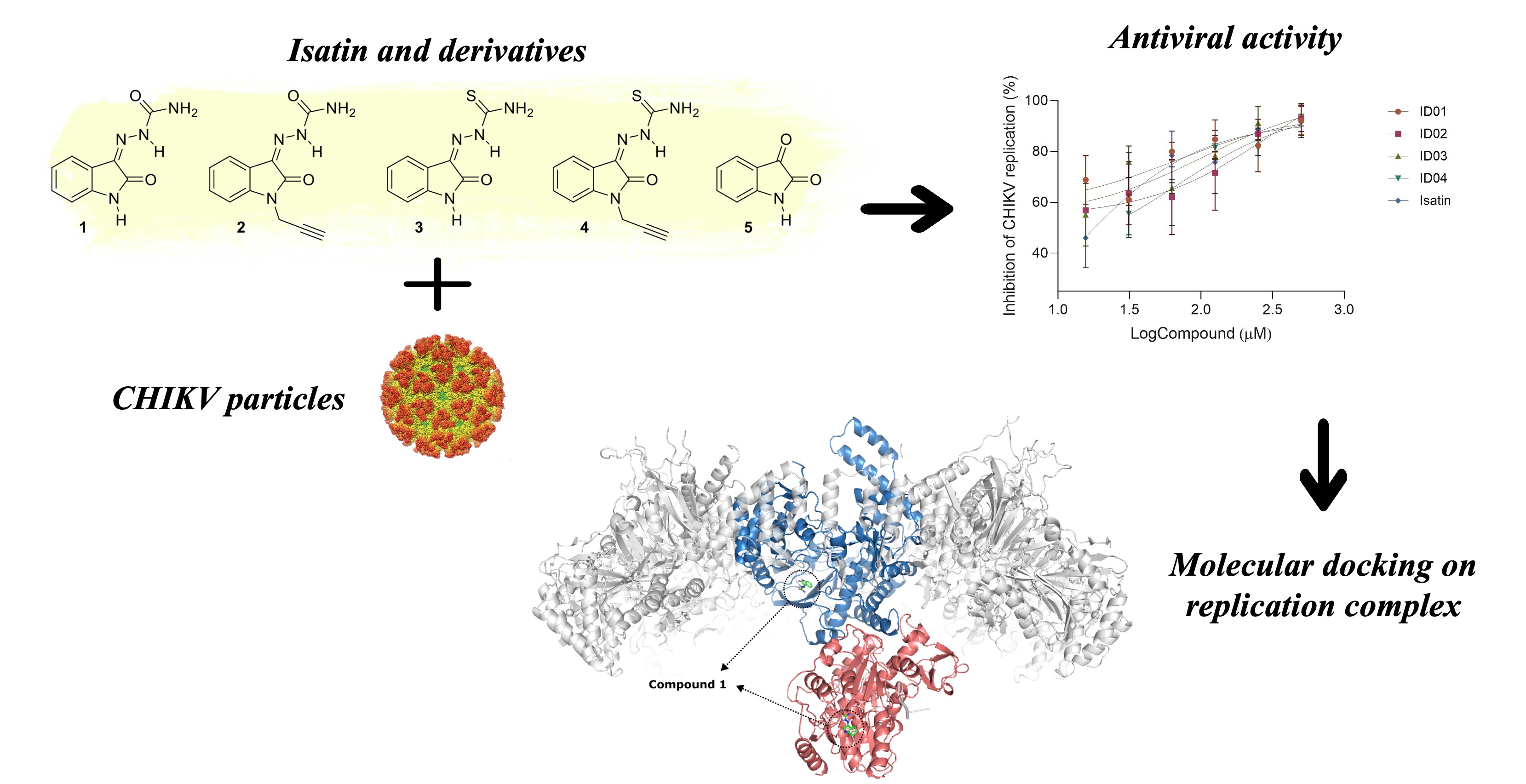
Chikungunya virus (CHIKV), a re-emerging arbovirus of the Alphavirus genus, causes acute febrile illness often followed by persistent polyarthritis or arthralgia. Due to its rapid transmission and the absence of approved antivirals or licensed vaccines, there is an urgent need for effective therapeutic strategies. Among potential antiviral targets, the nonstructural proteins nsP2 (a helicase/protease) and nsP4 (an RNA-dependent RNA polymerase) are essential for viral replication. In this study, we evaluated five compounds based on the isatin scaffold: one parent isatin molecule and four synthetic derivatives (ID11–ID14) functionalized with semicarbazone or thiosemicarbazone groups. Cytotoxicity assays confirmed low toxicity for all compounds, with CC₅₀ values above 500 µM. Using a CHIKV replicon model in BHK-CHIKV-NCT cells, compounds ID13 and ID14 significantly reduced EGFP fluorescence, indicating strong inhibition of viral replication. Infectious particle assays supported these results, with all compounds displaying dose-dependent antiviral effects and EC₅₀ values between 20 and 33.6 µM. No significant reduction in viral entry was observed in entry inhibition assays, suggesting that the compounds act post-entry by targeting replication. Molecular docking simulations revealed a predicted propensity of all compounds to interact with important residues of nsP2 and nsP4. Notably, compound ID14 showed the strongest predicted affinity for both enzymes, while compound ID13 demonstrated the highest antiviral efficacy in vitro. These findings highlight isatin derivatives as promising candidates for CHIKV antiviral development and support further exploration of dual-target strategies focusing on viral replication proteins.
Keywords:
chikungunya virus
; alphavirus
; small-molecule screening
; antiviral compounds
; molecular docking
; viral replication inhibitors
1. Introduction
Chikungunya virus (CHIKV), a member of the alphavirus genus of the Togaviridae family, is the etiologic agent of Chikungunya fever (CHIKF), manifested by a self-limited high fever associated with rash, myalgia, and debilitating polyarthritis or arthralgia, which may persist for months or even years after virus clearance [1,2]. Since its re-emergence, it has been considered a significant global health concern, particularly in tropical and subtropical regions [3].
Although the CHIKV vaccine has recently been approved and available for community use, immunocompromised individuals and other vulnerable groups may be not eligible for vaccination, exhibit worse side effects or reduced vaccine efficacy[4]. Additionally, limited population access to vaccines in endemic regions reinforces the need for antiviral therapies [5]. Thus, CHIKV antivirals remain essential for CHIKF management. Moreover, patients with prolonged acute CHIKF symptoms often develop persistent joint pain, which makes an early antiviral intervention of utmost importance [6]. Currently, clinical management relies solely on symptomatic relief, without attacking virus life cycle [7]. Therefore, development of antivirals that directly target the viral replication cycle is critical.
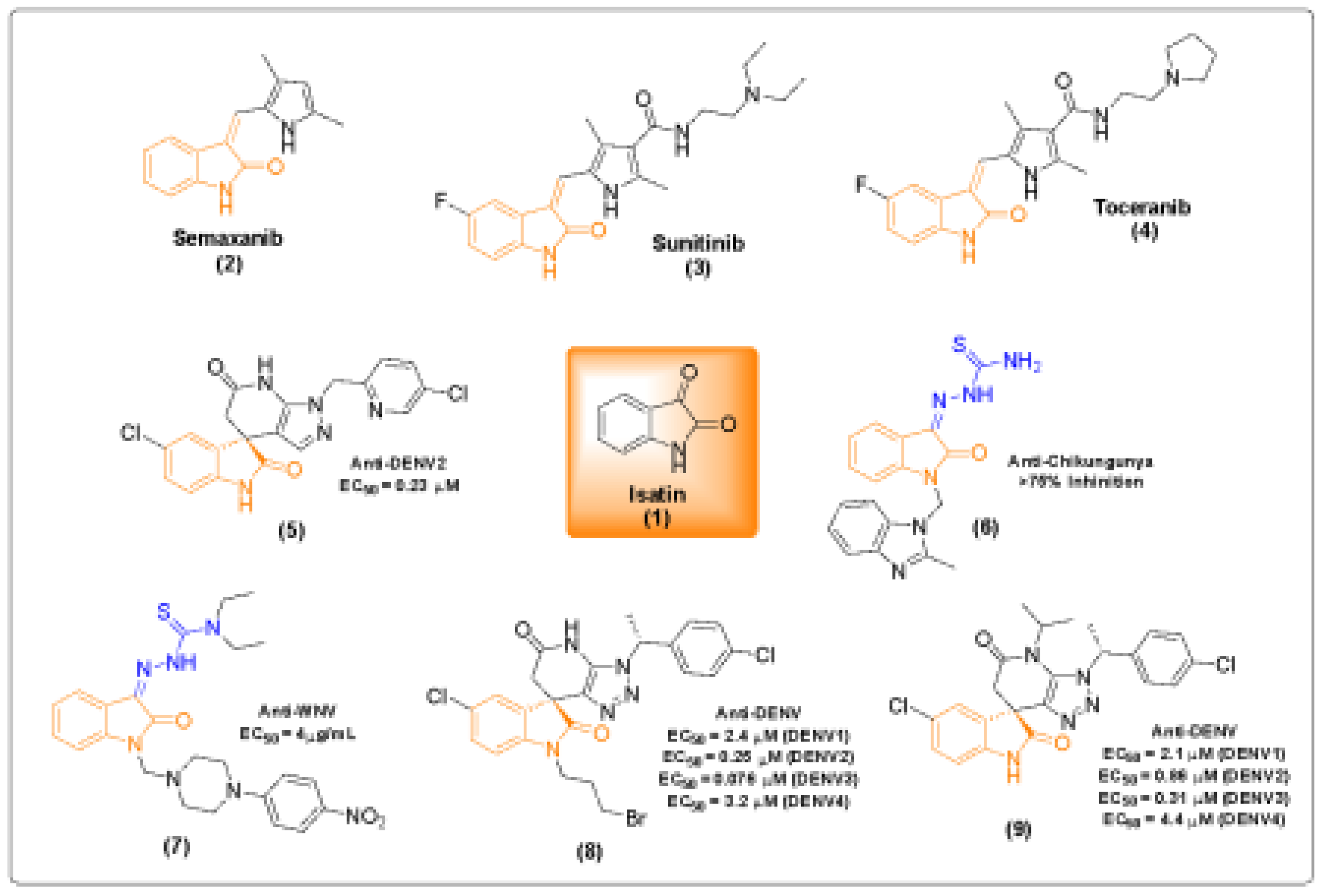
Among CHIKV proteins, the nonstructural proteins 2 and 4 (nsP2 and nsP4) are of particular interest for antiviral therapies, since it is the unique protease and helicase, and the RNA-dependent RNA polymerase, respectively. Blocking each of these proteins results in an impairment of virus replication, therefore, hampering virus propagation in the host [8,9]. Not surprisingly, most FDA-approved antivirals focused on protease and replicase viral enzymes [10,11]. In recent years, a wide variety of compounds, including natural extracts, synthetic organic molecules, and structurally diverse small molecules, have been investigated as potential antiviral agents against CHIKV. These studies highlight the relevance of multiple chemical classes, such as phytocompounds, alkaloids, terpenoids, and nucleoside analogs, which act through distinct mechanisms to inhibit viral replication [12,13]. Among these, heterocyclic compounds, particularly those based on the isatin (1H-indole-2,3-dione) (1) scaffold have gained attention due to their structural versatility and broad pharmacological activity. Semaxanib (2), sunitinib (3), and toceranib (4) are examples of clinically successful isatin-based drugs (Figure 1). The isatin structure demonstrate a range of biological and pharmacological properties, including anticancer, antimicrobial, anti-inflammatory and antiviral activities, including some arboviruses, such as CHIKV, JEV (Japanese encephalitis virus), WNV (West Nile virus) and DENV (dengue virus) [14,15]. Another significant class of biologically active compounds in medicinal and pharmaceutical chemistry is the hydrazone derivatives, as well, have been reported in medicinal chemistry as a strategic structure, since they have an azomethine pharmacophore, widely associated to several biological activities [16,17].
Taking the urgent need for CHIKV antiviral and the relevance of isatin as a promising pharmacophore, in the present study, we evaluated the antiviral potential of isatin and four isatin-derivatives (ID) compounds against CHIKV infection, focusing on their ability to inhibit viral replication through interactions with nsP2 and nsP4. We assessed both cellular cytotoxicity and antiviral activity of those compounds through classic and innovative methods. We also performed molecular docking studies to explore the likelihood of isatin derivatives to interact with nsP2 and nsP4. Our data indicates that our compounds are able to effectively block CHIKV replication through interactions with both nsP2 and nsP4.
2. Material and Methods
2.1. Chemistry
All reagents and solvents used were of analytical grade. The melting points (m.p. values) were determined with a Fisher-Johns apparatus and are uncorrected. The infrared (IR) spectra were recorded using a Perkin-Elmer FT-IR spectrophotometer (models 1600 Series and EspectroOne), equipped with a double beam, and measured on anhydrous potassium bromide tablets. Absorption values are expressed in wavenumbers, with cm⁻¹ as the unit of measurement. NMR 1H spectra were obtained in deuterated DMSO-d6 were acquired from Sigma-Aldrich (São Paulo, Brazil) using a Varian Unity Plus 500.00 MHz spectrometer. Chemical shifts (δ) are expressed in ppm and the coupling constant (J) in Hertz. Reactions were routinely monitored by thin layer chromatography (TLC) on silica gel pre-coated F254 Merck plates.
2.2. Synthesis
2.2.1. Procedure for Preparing 2-(2-oxoindolin-3-ylidene)hydrazine-1-Carboxamide (11)
In a round-bottom flask, 27.1 mmol of semicarbazide hydrochloride (10a), 27.1 mmol of sodium acetate, 42 mL of water, and 18 mL of ethanol were added. The mixture was stirred for 30 minutes. Subsequently, 13.6 mmol of isatin (1) was added. The reaction mixture was stirred at room temperature for 48 h and monitored by thin-layer chromatography (TLC), using a hexane/ethyl acetate (3:7) mixture as the eluent. Afterwards, the reaction mixture was poured onto ice, the resulting solid was filtered, and washed with cold water until completely purification. The compound 11 was obtained as a yellow solid with 61% yield.
Yield: 61%. m.p.: 266-268 ºC. IR (cm-1; KBr): 1347; 1601; 1694; 3237. 1H NMR (500.00 MHz, DMSO- d6, TMS, δ in ppm): 6.91 (d, 1H, J = 7.8 Hz, H-7); 7.06 (t, 1H, J = 7.8 Hz, H-5); 7.32 (t, 1H, J = 7.8 Hz, H-6); 7.61 (d, 1H, J = 7.8 Hz, H-4); 10.02 (s, 1H); 10.55 (s, 1H); 10.95 (s, 1H); 11.72 (s, 1H).
2.2.2. Procedure for Preparing 2-(2-Oxoindolin-3-ylidene)Hydrazine-1-Carbothioamide (13)
In a round-bottom flask, 1.36 mmol of isatin (1), 2.72 mmol of thiosemicarbazide (10b), 14 mL of water, and 6 mL of ethanol were added. The mixture was stirred at room temperature for 48 h. The reaction was monitored by thin-layer chromatography (TLC), using a hexane/ethyl acetate (3:7) mixture as the eluent. Subsequently, the reaction mixture was poured onto ice, filtered, and the resulting solid was washed with cold water until completely purification. The compound 13 was obtained as a yellow solid with 94% yield.
Yield: 94%. m.p.: 243-245 ºC. IR (cm-1; KBr): 856 and 1256; 1593; 1672; 3266. 1H NMR (500.00 MHz, DMSO- d6, TMS, δ in ppm): 6.93 (d, 1H, J = 7.8 Hz, H-7); 7.08 (t, 1H, J = 7.8 Hz, H-5); 7.34 (t, 1H, J = 7.8 Hz, H-6); 7.66 (d, 1H, J = 7.8 Hz, H-4); 8.51 (s, 1H, H-4’); 8.85 (s, 1H, H-4”); 12.47 (s, 1H, H-2’).
2.2.3. Procedure for Preparing 1-(prop-2-yn-1-yl)-2,3-Dihydro-1H-Indole-2,3-Dione (16)
In a Schlenk tube, 0.680 mmol of isatin (1), 0.68 mmol of K2CO3, and 2 mL of DMF were added. The reaction was maintained under stirring at 50 °C for 15 minutes. After this period, 1.0 mmol of propargyl bromide (15) was added dropwise with the aid of a syringe. The reaction continued for 2 h and was monitored by TLC, using a hexane/ethyl acetate 3:7 elution system. The reaction mixture was poured onto ice, was filtered and washed with cold water. The product was purified by recrystallization using ethanol as solvent. Recrystallization was performed using ethanol. The intermediate 16 was obtained as an orange solid with 79% yield.
Yield: 79%. m.p.: 158-160 ºC. IRν (cm-1; KBr): 1612; 1737; 2923; 3262. 1H NMR (500.00 MHz, DMSO-d6, TMS, δ in ppm): 3.24 (t, 1H, J = 2.4 Hz, H-3’); 4.55 (d, 2H, J = 2.4 Hz, H-1’); 7.19 (t, 1H, J = 7.8 Hz, H-5); 7.24 (d, 1H, J = 7.8 Hz, H-7); 7.59 (d, 1H, J = 7.8 Hz, H-4); 7.72 (t, 1H, J = 7.8 Hz, H-6).
2.2.4. Procedure for Preparing 2-(2-oxo-1-(prop-2-yn-1-yl)Indolin-3-Ylidene)Hydrazinecarboxamide (12)
In a round-bottom flask, 21.6 mmol of semicarbazide hydrochloride (10a), 21.6 mmol of sodium acetate, 42 mL of water, and 18 mL of ethanol were added. The mixture was stirred for 1 hour, after which 11.8 mmol of propargylated isatin (16) was added. The reaction mixture was stirred at room temperature for 72 h. The progress of the reaction was monitored by thin-layer chromatography (TLC), using a hexane/ethyl acetate (3:7) mixture as the eluent. Upon completion, the reaction mixture was poured onto ice, and the resulting solid was filtered and washed several times with cold water until completely purification. The compound 12 was obtained as yellow solid with 85% yield.
Yield: 85%. m.p.: 178-180 °C. IR (cm-1; KBr): 1603; 1692; 3427, 3294. 1H NMR (500.00 MHz, DMSO-d6, TMS, δ in ppm): 3.14-3.16 (m, 1H, H-3”); 4.58 (d, 2H, J = 2.4 Hz, H-1”), 6.80 (s, 2H, NH2); 7.14-7.18 (m, 2H, H-5 and H-7); 7.46 (t, 1H, J = 7.5 and 7.8 Hz, H-6); 8.14 (d, 1H, J = 7.5 Hz, H-4); 10.20 (s, 1H, H-2’).
2.2.5. Procedure for Preparing 2-(2-oxo-1-(prop-2-yn-1-yl)Indolin-3-ylidene)Hydrazinecarbothioamide (14)
In a round-bottom flask, 6.48 mmol of propargylated isatin (16), 12.9 mol of recrystallized thiosemicarbazide (10b), 14 mL of water, and 6 mL of ethanol were added. The mixture was stirred at room temperature for 48 h. The reaction was monitored by thin-layer chromatography (TLC), using a hexane/ethyl acetate (3:7) mixture as the eluent. Finally, the reaction mixture was poured onto ice, filtered, and the solid was washed several times with cold water until completely purification. The compound 14 was obtained as yellow solid with 98% yield.
Yield: 98%. m.p.: 185-187 °C. IR (cm-1; KBr): 1241; 1593; 1702; 3392, 3236.1H NMR (500.00 MHz, DMSO-d6, TMS, δ in ppm): 3.23 (t, 1H, J = 2.4 Hz, H-3”); 4.62 (d, 2H, J = 2.4 Hz, H-1”); 7.19 (td, 1H, J = 7.3 and 8.3 Hz, H-5); 7.23 (d, 1H, J = 7.8 Hz, H-7); 7.47 (td, 1H, J = 7.8 and 8.3 Hz, H-6); 7.75 (d, 1H, J = 7.3 Hz, H-4); 8.59 (s, 1H, NH2); 8.94 (s, 1H, NH2); 12.28 (s, 1H, H-2’).
2.3. Cytotoxicity Assay in BHK-21 Cells
BHK-21 cells were seeded in 96-well plates at a density of 1 × 10⁴ cells per well and cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin. After 24 h, the cells were treated with the compounds at concentrations ranging from 15.62 to 500 µM and incubated for an additional 24 h at 37 °C with 5% CO₂. For the MTT assay, 10 µL of MTT solution (5 mg/mL in PBS) was added to each well, and the plates were incubated for 4 h. The formazan crystals formed were dissolved in 100 µL of DMSO, and the absorbance was measured at 570 nm using a microplate reader. Cell viability was calculated as a percentage relative to untreated controls. For the trypan blue exclusion assay, cells were trypsinized, mixed with 0.4% trypan blue solution, stained with trypan blue and counted according to the protocol described by Lebeau et al., 2019 [24]. The assays were performed in triplicate, and the results were used to determine the non-toxic concentration range for subsequent antiviral assays.
2.4. Cell-Based Replicon Assay for CHIKV Inhibition
BHK-CHIKV-NCT cells were seeded in opaque-white 96-well plates with a clear bottom at a density of 3 × 10⁴ cells per well and cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 1% penicillin–streptomycin, and 5 μg/mL puromycin. After 24 h, cells were treated with the selected compounds at concentrations ranging from 15.62 to 500 µM for an additional 24 h. This cell line, kindly provided by Prof. Andres Merits (University of Tartu, Estonia), constitutively expresses the CHIKV replicase proteins along with two reporter genes: EGFP and Renilla luciferase
The inhibition of CHIKV replication was assessed by monitoring the suppression of EGFP fluorescence using a microplate reader (excitation/emission: 478/508 nm, bandwidth: 5 nm). The half-maximal inhibitory concentration (IC₅₀) values for both cell viability and EGFP fluorescence inhibition were calculated using nonlinear regression analysis. For Renilla luciferase activity, assays were carried out with the Renilla Luciferase Assay Kit (Promega, USA), following the manufacturer’s instructions. All experiments were performed in triplicate.
2.5. Viral Replication Inhibition Assay
BHK-21 cells were seeded into a 6-well plate with 1x10⁶ cells per well, and the plates were incubated at 37 °C with 5% CO₂ for 24 h. The cells were then infected with CHIKV, strain RJ5, genotype ECSA at a multiplicity of infection (MOI) of 0.2 and incubated for 2 h with agitation every 20 minutes[25]. Following infection, the virus-containing medium was replaced with fresh DMEM containing the compounds at concentrations ranging from 15.62 to 500 µM. Controls included virus-infected cells treated with 0.5% DMSO (vehicle control) and uninfected cells treated with the compounds. The plates were incubated for an additional 24 h at 37 °C with 5% CO₂ prior to viral quantification by RT-PCR.
2.6. Viral Entry Inhibition Assay
BHK-21 cells were seeded into a 6-well plate with 1x10⁶ cells per well, and the plates were incubated at 37 °C with 5% CO₂ for 24 h. The cells were then infected with CHIKV, as described above, together with different compounds concentration. The compounds were screened to a final concentration of 20 or 32 µM, selected based on their IC₅₀ values determined in the antiviral assay described in Section 2.5. The viral particles and compounds remained in contact with the cells during a 2-hour incubation, with agitation every 20 minutes to ensure homogeneity. After the 2-hour incubation, the medium containing the virus and compounds was removed and replaced with MEM supplemented with serum. The plates were then incubated for an additional 24 h to allow the experiment to proceed. Total RNA was extracted from the infected cells using TRIzol reagent, and viral RNA levels were quantified by qRT-PCR as described in Section 2.7. The reduction in viral RNA levels relative to the vehicle control was used as a measure of the antiviral efficacy of the compounds. All experiments were performed in triplicate, and the results were expressed as mean ± standard deviation.
2.7. Virus Quantification by RT-qPCR
Total RNA from CHIKV-infected BHK-21 cells treated with vehicle or the compounds in Section 2.5 and Section 2.6 was extracted using TRIzol reagent (Thermo Fisher Scientific) according to the manufacturer’s instructions. The RNA was converted into cDNA using the High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific). The quantification of the CHIKV genome was performed with 100 ng of cDNA and 300 ng of the following primers:
| CHIKV_Fw | AAAGGGCAAACTCAGCTTCAC |
| CHIKV_Rv | GCCTGGGCTCATCGTTATTC |
The qRT-PCR was performed on the QuantStudio™ 3 Real-Time PCR System using PowerUp™ SYBR™ Green Master Mix (Thermo Fisher Scientific) according to the manufacturer’s instructions. A standard curve established from a serial dilution of the viral stock was used to determine the virus load[26]. All experiments were performed in triplicate, and the results were expressed as mean ± standard deviation.
2.8. Molecular Modeling and Docking of Isatins with nsP4 and nsP2 Proteins of CHIKV
The three-dimensional structure of nsP2 was retrieved from the Protein Data Bank (PDB ID: 6JIM), while the structure of nsP4 was modeled using AlphaFold2, based on the reference replication complex structure available in the PDB (ID: 7Y38) [27]. Molecular docking was performed using the HADDOCK 2.4 server, which allows flexible docking and refinement of protein–ligand interactions. For nsP4, particular attention was given to the catalytic residues Asp466 and Asp467, while for nsP2, the key residues analyzed were Lys192, Gln285, and Arg312 [28]. A docking grid was centered around these critical catalytic residues, focusing on pockets previously described for nsP2 and nsP4. The docking scores for each compound are summarized in Table 2, while the main protein–ligand interactions are illustrated in Figure 6 and Figure 7. The structural representations were generated using PyMOL (version 2.5.7, Schrödinger, LLC), and the 2D interaction diagrams were prepared with LigPlot+ (version 2.2, European Bioinformatics Institute). As a negative control, Benzaldehyde Semicarbazone was included as a decoy ligand.
2.9. Data Analysis
The data were analyzed using GraphPad Prism statistical software (version 10.4.1, GraphPad Software, Inc.). For the viability and viral replication assays, IC50 values were calculated based on dose-response curves using nonlinear regression analysis. The results were expressed as mean ± standard error of the mean (SEM) from at least three independent experiments. Statistical significance was determined using a one-way ANOVA test, with a p-value of <0.05 considered statistically significant.
3. Results and Discussion
3.1. Chemistry
Synthetic strategy of hydrazone-isatin derivatives (11-14), tested in this study, has been outlined in Scheme 1. Initially, the nucleophilic addition reaction between isatin (1) and the respective semicarbazide hydrochloride (10a) or thiosemicarbazide (10b) were dissolved in water/etanol, at room temperature for 48 h, afforded the synthesis of isatin-semicarbazone (11) and isatin-thiosemicarbazone (13) in 61% and 94% yield, respectively[29,30,31]. To obtain N-propargylated isatin (16) in 79% yield, isatin (1) was stirred continuously for 2 h with propargyl bromide (15) in presence of K2CO3 in minimum amount of DMF at 50 °C for 15 minutes [32,33,34]; Finally, the propargylated isatin (16) was dissolved in ethanol/water and stirred with semicarbazide hydrochloride (10a) or thiosemicarbazide (10b) in water and ethanol, for 72 h at room temperature to obtain isatin-semicarbazone tethered terminal alkynes 12 and 14, in 85% and 98% yield, respectively[30,35]. All the synthesized isatin derivatives (11-14) were satisfactorily characterized by spectral techniques and the data are according to literature. To facilitate sample identification throughout the Results section, the compounds will be referred to as Isatin (compound 1) and Isatin Derivatives (ID11–ID14), corresponding to compounds 11 to 14, respectively.
3.3. Antiviral Activity Using a CHIKV Replicon Model
The BHK-CHIKV-NCT cell line harbors a non-cytotoxic CHIKV replicon encoding the viral nonstructural proteins (nsPs) along with two reporter genes: Renilla luciferase (Rluc) fused to nsP3 and EGFP, expressed as a fusion protein with puromycin acetyltransferase (Pac) under the control of the subgenomic promoter.. EGFP is cleaved from Pac through the Foot-and-Mouth Disease Virus (FMDV) 2A autoprotease sequence, allowing its release into the cytoplasm. When cultured in 96-well plates, BHK-CHIKV-NCT cells display robust luminescent and fluorescent signals, which can be easily quantified using a microplate reader. This replicon system faithfully recapitulates the RNA replication phase of CHIKV and enables monitoring of viral replication through reporter activity. Thus, any interference with the replication complex directly impacts the expression of EGFP and Rluc [36].
Treatment with Isatin, ID13, and ID14 resulted in a significant, dose-dependent reduction in EGFP fluorescence, indicating inhibition of viral replication. The most pronounced effects were observed for Isatin (85%), ID13 (80%), and ID14 (60%) at the highest concentration tested (500 µM) (Figure 2). Fluorescence microscopy images of cells treated with the compounds at 500 µM are shown in Supplementary Figure S2, obtained using the EVOS M5000 system (Thermo Fisher Scientific). To further validate these findings, Rluc activity assays were performed with ID13 and ID14. The compounds exhibited IC₅₀ values of 313 µM (ID13) and 352 µM (ID14), confirming their ability to interfere with replicon activity. Collectively, these results indicate that the tested compounds disrupt the activity of the CHIKV replication complex, which is expected to impact the production of new viral particles, as investigated in the subsequent assay.
3.2. Cytotoxicity Analysis
To assess compound safety, cell cytotoxicity was evaluated using MTT and trypan blue exclusion assays after 24 h of treatment with each compound. Both assays confirmed minimal cytotoxicity and preserved cell viability at the tested concentrations, indicating compatibility with the cellular model. Isatin exhibited moderate toxicity (70–75% viability) at concentrations between 125 and 500 µM in the trypan blue assay. In contrast, isatin derivatives ID11–ID14 showed no signs of cytotoxicity in either assay. The complete cytotoxicity profile is presented as Supplementary Figure S1.
3.4. Antiviral Activity Against Infectious Viral Particles
To directly evaluate the effect of the compounds on CHIKV replication, assays were performed using infectious viral particles. Compound concentrations were optimized based on preliminary tests. All compounds reduced viral titers in a concentration-dependent manner, with EC₅₀ values ranging from 20 to 33 µM (Table 1, Figure 3). Among them, ID13 showed the highest efficacy (EC₅₀ = 20.0 µM), followed by ID11 (EC₅₀ = 20.0 µM) and Isatin (EC₅₀ = 24.6 µM). Interestingly, while ID11 and ID13 did not exhibit strong antiviral activity in the replicon cell model, they were highly effective in the infectious virus assay. This discrepancy suggests that the replicon system may not fully capture the antiviral activity of these compounds, highlighting the importance of complementary experimental models. Overall, these findings reinforce the antiviral potential of Isatin derivatives against CHIKV.
*EC50, 50% effective concentration, CC50, 50% cytotoxic concentration.
Several studies have identified small molecules with significant antiviral activity against CHIKV by targeting the viral replication. These compounds have demonstrated in vitro efficacy with EC₅₀ values ranging from 0.12 to 70 μM, which are comparable to the ranges observed for the compounds evaluated in the present study. Among the direct-acting inhibitors, MADTP-314 and CHVB-032 have shown notable results, with EC₅₀ values of 26 μM and 2.7 μM, respectively, in Vero cells infected with the CHIKV IO 899 strain [37]. Repositioned compounds have also proven to be effective in CHIKV replication inhibition, as the case of mycophenolic acid, which exhibited an EC₅₀ of 0.21 μM for CHIKV inhibition in Vero cells. However, this compound presented significant cytotoxic effects in mammalian cells, with CC₅₀ values around 30 μM, which may limit its therapeutic application [38]. Sofosbuvir, a clinically approved antiviral for hepatitis C, also demonstrated anti-CHIKV activity, with an EC₅₀ of 2.7 μM in Huh-7 cells [39]. Favipiravir, another approved antiviral, showed an EC₅₀ of 25 μM in Vero-A cells for CHIKV [40]. Additionally, MBZM-N-IBT, an isatin-derivative compound, showed inhibitory activity against the ECSA strain of CHIKV with an EC₅₀ of 38.7 μM [23]. Similarly, silymarin, a flavonoid-rich compound, exhibited antiviral effects in Vero cells with an EC₅₀ of 35 μM against the ECSA lineage [41].
The similarity in the EC₅₀ ranges between previously reported compounds and those evaluated in this study suggests that the mechanisms of action may be comparable. Additionally, the isatin-derivative compounds tested here showed no evidence of cytotoxicity in mammalian cells, with all presenting CC₅₀ values above 500 μM. This indicates a favorable safety profile and a high selectivity index, reinforcing their potential as lead compounds for further antiviral development.
3.5. Mechanism of Action: Entry vs. Replication Inhibition
To investigate Isatin and ID 11-14 mechanisms of action, a viral entry inhibition assay was performed by treating cells with the compounds together with CHIKV infection. No significant differences were observed between treated and untreated conditions, indicating that the compounds do not interfere with viral adsorption, therefore, with viral entry (Figure 4). These results corroborate with our hypothesis that the compounds impair CHIKV infection by inhibition of viral replication rather than entry process.
3.6. Molecular Docking Analysis and Correlation with Biological Assays
The nonstructural proteins (nsPs) of CHIKV are essential for viral transcription and translation. Notably, both nsP2 and nsP4 are among the primary targets for the development of antivirals against alphaviruses, as they play essential roles in the viral replication process [42,43]. nsP2 contains two functional domains: a protease domain and an ATP-dependent helicase domain, while nsP4 functions as the RNA-dependent RNA polymerase (RdRp). Given the chemical nature of the isatin-derived molecules tested in this study, whose structures are analogous to nucleotides, the helicase domain of nsP2 and the polymerase domain of nsP4 were selected as potential binding sites for molecular docking simulations.
Docking analysis revealed that all compounds showed favorable binding affinities with both nsP2 and nsP4 (Table 2), supporting their potential role as inhibitors of the CHIKV replication machinery. Importantly, the decoy ligand (benzaldehyde semicarbazone) displayed a lower binding score, thereby validating the specificity of the docking approach.
3.6.1. Interactions with nsP2 (Helicase Domain)
The helicase domain of nsP2 is a promising target for antiviral development due to its essential role in viral replication. This domain exhibits ATPase and RNA helicase activity, which are critical for RNA unwinding and viral replication progression, making it a key target for antiviral strategies. Inhibiting this activity could disrupt viral replication, making it an effective antiviral strategy [28]. While inhibitors for human helicases exist, no specific inhibitors for viral helicases have been reported so far [44].
For nsP2, Lys192, Gln285, and Arg312 were identified as critical residues for ligand stabilization[28]. All tested compounds (Isatin and ID11–14) showed similar docking scores, suggesting a conserved binding mode within the nsP2 helicase pocket. Among them, ID14 exhibited the most favorable binding score, followed by ID12, ID11, ID13, and Isatin. The interactions were mainly stabilized through hydrogen bonds and electrostatic interactions, with ID14 forming multiple hydrogen bonds with Gly191, Lys192, Ser193, and Glu253. In contrast, Isatin engaged fewer residues, interacting primarily with Gly191 and Lys192 (Figure 5).
3.6.2. Interactions with nsP4 (Polymerase Domain)
Similarly, nsP4 was selected for in silico analyses due to its role as the viral RNA-dependent RNA polymerase, a key enzyme in CHIKV genome replication. Given its central function in viral propagation, targeting nsP4 could effectively disrupt CHIKV replication, making it a promising candidate for antiviral development.
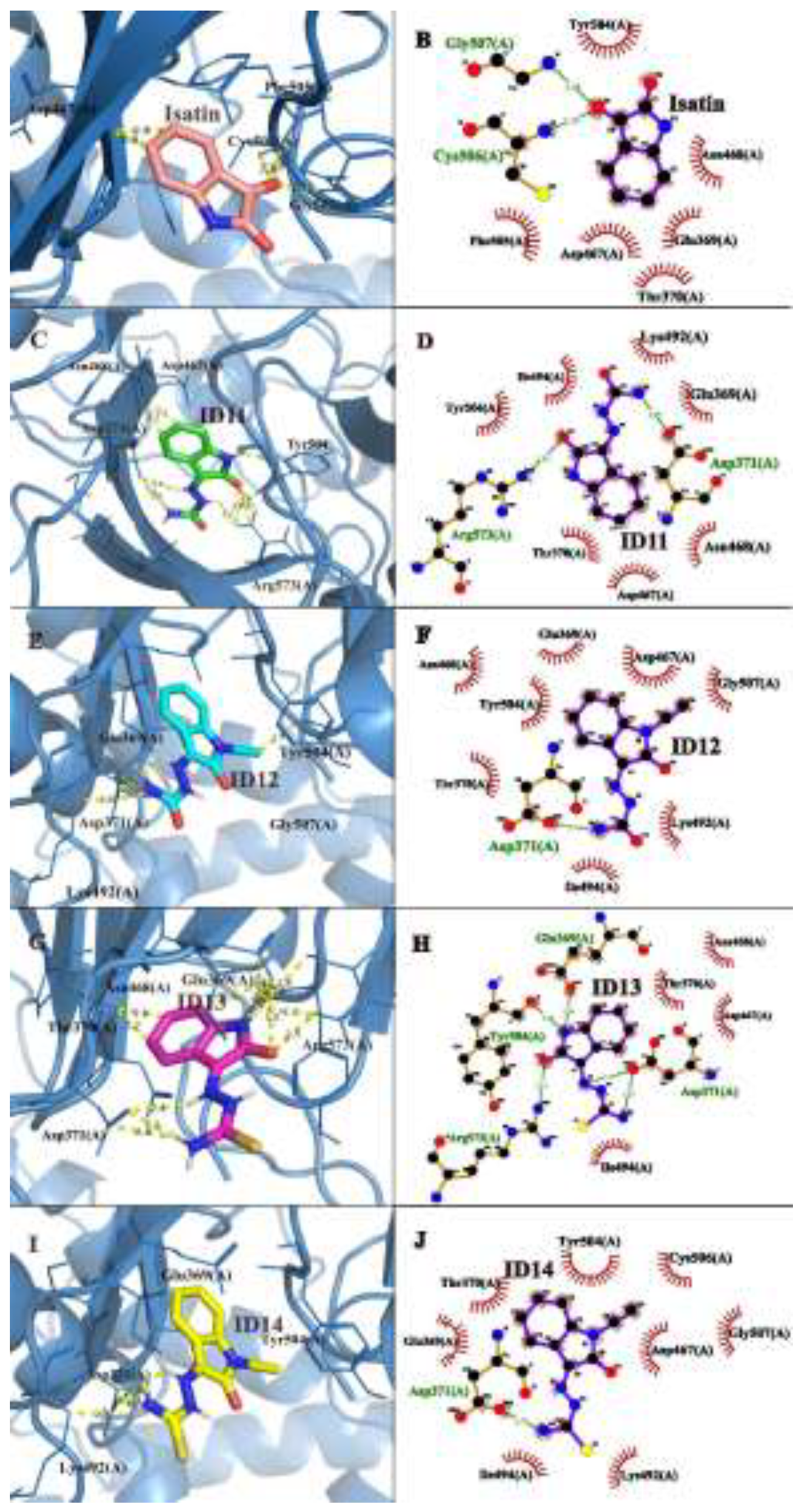
For nsP4, Asp371 and Asp467 were identified as critical residues for ligand stabilization. Compounds ID11 to ID14 exhibited favorable interactions, primarily involving hydrogen bonding and hydrophobic contacts with key residues. Among them, compound ID14 again displayed the highest docking score (-32.4 ± 0.3), forming multiple hydrogen bonds with Asp371 and additional interactions with Tyr504, Glu369, and Thr370. Compounds ID12 and ID11 followed, with docking scores ranging from -31.9 to -29.8, while compound ID13 showed slightly lower affinity in this target. Isatin presented the weakest binding (-27.4 ± 0.2), interacting mainly with Gly507 and Cys506 (Figure 6).
Figure 6.
Molecular docking representation of Isatin and ID11-14 with CHIKV nsP4. The figure presents both three-dimensional (A–E) and two-dimensional (F–J) representations of the docking interactions between nsP4 and the tested compounds. Panels A to E correspond to the 3D docking interactions of Isatin and ID11-14, respectively, while panels F to J display the 2D interaction maps for the same compounds. Key binding residues and interaction types, such as hydrogen bonds and hydrophobic interactions, are highlighted. Images generated with PyMOL© and LigPlot ©.
Figure 6.
Molecular docking representation of Isatin and ID11-14 with CHIKV nsP4. The figure presents both three-dimensional (A–E) and two-dimensional (F–J) representations of the docking interactions between nsP4 and the tested compounds. Panels A to E correspond to the 3D docking interactions of Isatin and ID11-14, respectively, while panels F to J display the 2D interaction maps for the same compounds. Key binding residues and interaction types, such as hydrogen bonds and hydrophobic interactions, are highlighted. Images generated with PyMOL© and LigPlot ©.
3.7. Structural Insights into the Replication Complex
To better illustrate the impact of these interactions, Figure 7 shows an overlay of nsP4 (in blue) and the helicase domain of nsP2 (in pink) within the CHIKV replication complex, highlighting the binding sites of the ID compounds. Compound ID11 was used as a reference due to its representative binding profile among the tested derivatives, allowing for a comparative structural analysis. As illustrated, all compounds demonstrated a clear tendency to interact with both nsP2 and nsP4, suggesting that the molecules may have the potential to engage more than one target within the replication complex, rather than binding exclusively to a single protein domain.
Figure 7.
CHIKV replication complex highlighting the interaction sites of the tested compounds. nsP4 is shown in blue, while the helicase domain of nsP2 is in pink. The binding positions of ID11, used as a reference, are indicated by dotted circles, representing the interaction sites of the tested molecules. Images generated with PyMOL©.
Figure 7.
CHIKV replication complex highlighting the interaction sites of the tested compounds. nsP4 is shown in blue, while the helicase domain of nsP2 is in pink. The binding positions of ID11, used as a reference, are indicated by dotted circles, representing the interaction sites of the tested molecules. Images generated with PyMOL©.
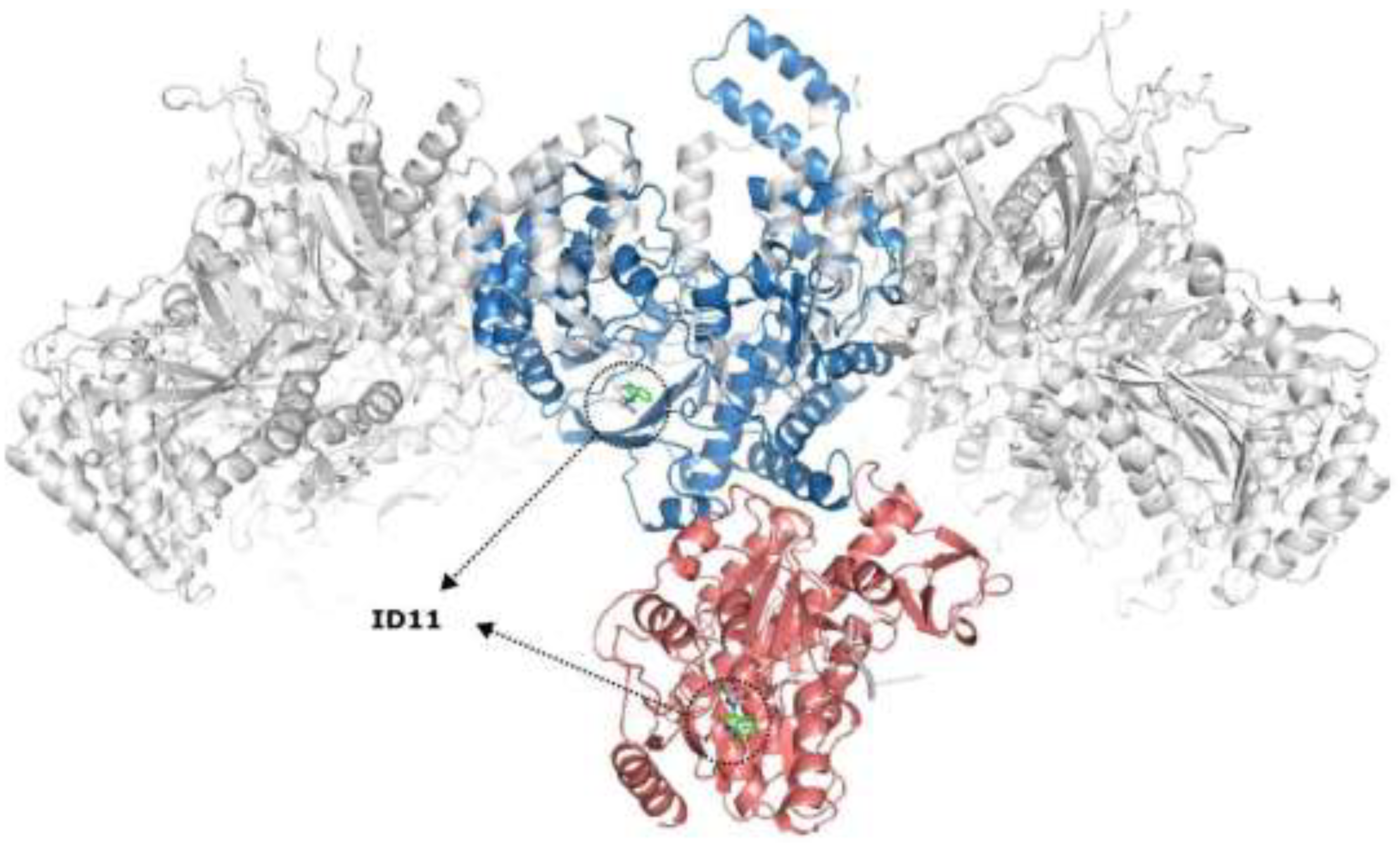
The binding of ID11–ID14 to both nsP2 and nsP4 suggests that these compounds may interfere with the functional coordination between the helicase and polymerase domains, potentially impairing viral genome replication. Simultaneous engagement of these proteins could generate a synergistic effect, significantly reducing the efficiency of the replication process. These results indicate that the ID compounds inhibit CHIKV replication, possibly through interactions with nsP2 and nsP4. The integration of biological and computational data underscores the potential of these molecules as antiviral candidates, with ID14 and ID13 emerging as the most promising leads for further optimization.
4. Overview and Conclusion
This study combined in vitro and in silico approaches to investigate the antiviral potential of five isatin-derived compounds against CHIKV. These molecules, based on nitrogen-containing heterocyclic scaffolds such as pyrimidine, pyrazine, and purine, are structurally related to nucleotide analogs and have previously been recognized for their broad-spectrum antiviral activity [45]. Three experimental assays were performed to determine the antiviral mechanism of these compounds. The replicon system revealed that Isatin, ID13 and ID14 significantly reduced GFP fluorescence, suggesting inhibition of viral replication. In the assay using infectious CHIKV particles, all compounds reduced viral titers in a dose-dependent manner (EC₅₀: 20–33.6 µM). However, no effect was observed in the viral entry inhibition assay, confirming that the compounds act post-entry, likely targeting replication machinery.
Molecular-docking results revealed that all ID compounds exhibit favorable affinities to the two key CHIKV replication enzymes, nsP2 (helicase) and nsP4 (RNA-dependent RNA polymerase). Among them, ID14 showed the lowest docking scores for both targets (−34.9 for nsP2 and −32.4 for nsP4) and ranked among the most potent antivirals in cell replicon-based assays. Conversely, ID13 produced the strongest in vitro inhibition despite only intermediate docking scores, indicating that additional factors, such as metabolic stability or intracellular accumulation, may modulate overall potency. Taken together, the parallel reduction in viral replication and the predicted interactions with both nsP2 and nsP4 support a dual-target antiviral mechanism for this series of ID compounds.
Our results are in accordance with previous studies that also highlight the potential of synthetic small molecules as CHIKV inhibitors. Nucleoside analogs such as sofosbuvir, favipiravir, and ribavirin have demonstrated efficacy in inhibiting CHIKV RdRp, reinforcing the therapeutic relevance of targeting viral replication [39,46,47]. Similarly, Mishra et al. (2020) reported that a hybrid isatin-based compound could inhibit multiple stages of the CHIKV life cycle, further validating the potential of isatin derivatives as multitarget antivirals.
Additionally, recent reviews emphasize the importance of focusing on CHIKV nonstructural proteins, especially nsP2 and nsP4, as promising targets for antiviral development [43,48,49]. Our findings reinforce this strategy by demonstrating that isatin-based compounds can engage both targets, potentially enhancing antiviral efficacy through dual inhibition.
This study consists of an innovative contribution to CHIKV research by integrating experimental assays with molecular docking to elucidate the mechanism of action of five Isatin Derivatives compounds. Unlike many previous studies that focus exclusively on screening or computational modeling, our approach not only confirms the antiviral activity of these synthetic molecules in vitro but also identifies nsP2 and nsP4 as dual molecular targets. This dual-target strategy is particularly promising in the context of CHIKV, where treatment options remain limited. By advancing the understanding of how isatin-based scaffolds interact with key viral enzymes, this work lays a solid foundation for the rational design of novel antivirals with improved efficacy and specificity.
In summary, our study highlights the potential of isatin-derived compounds as CHIKV inhibitors, with dual inhibition of nsP2 and nsP4 as a proposed mechanism. ID14 demonstrated the strongest tendency to interact with both targets, while ID13 exhibited the most potent antiviral effect in vitro, suggesting that additional pharmacokinetic properties influence efficacy.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Mayara Silva - Conceptualization; Data curation; Formal analysis; Investigation; Funding acquisition; Methodology; Visualization; Roles/Writing - original draft; Writing - review & editing; Tamires Carmencitta - Investigation; Methodology; Clara Lima - Investigation; Methodology; Carolaine da Silva - Investigation; Methodology; Flaviana Rodrigues Fintelman Dias - Investigation; Methodology; Luiz C. S. Pinheiro - Investigation; Methodology; Diego Allonso - Conceptualization; Data curation; Formal analysis; Funding acquisition; Investigation; Methodology; Project administration; Supervision; Validation; Visualization; Roles/Writing - original draft; Writing - review & editing; Ronaldo Mohana - Funding acquisition; Investigation; Methodology; Project administration; Supervision; Validation; Visualization; Writing - review & editing; Vinicius Rangel - Conceptualization; Data curation; Formal analysis; Funding acquisition; Investigation; Methodology; Project administration; Supervision; Validation; Visualization; Roles/Writing - original draft; Writing - review & editing.
Funding
This study was funded by Fundação Carlos Chagas Filho de Amparo à Pesquisa no Estado do Rio de Janeiro (FAPERJ) (grants E-26/210.403/2022, E-26/201.303/2022, E-26/206.004/2022) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (grant 444229/2023-0).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no financial conflict of interest with the contents of this article.
References
- Hua, C.; Combe, B. Chikungunya Virus-Associated Disease. Curr. Rheumatol. Rep. 2017, 19, 69. [CrossRef]
- Mason, P.J.; Haddow, A.J. An Epidemic of Virus Disease in Southern Province, Tanganyika Territory, in 1952–1953. Trans. R. Soc. Trop. Med. Hyg. 1957, 51, 238–240. [CrossRef]
- Zeller, H.; Van Bortel, W.; Sudre, B. Chikungunya: Its History in Africa and Asia and Its Spread to New Regions in 2013-2014. J. Infect. Dis. 2016, 214, S436–S440. [CrossRef]
- Weber, W.C.; Streblow, D.N.; Coffey, L.L. Chikungunya Virus Vaccines: A Review of IXCHIQ and PXVX0317 from Pre-Clinical Evaluation to Licensure. BioDrugs 2024, 38, 727–742. [CrossRef]
- Guignard, A.; Praet, N.; Jusot, V.; Bakker, M.; Baril, L. Introducing New Vaccines in Low- and Middle-Income Countries: Challenges and Approaches. Expert Rev. Vaccines 2019, 18, 119–131. [CrossRef]
- Ojeda Rodriguez, J.A.; Haftel, A.; Walker, J.R., III Chikungunya Fever. In StatPearls; StatPearls Publishing: Treasure Island (FL), 2025.
- Webb, E.; Michelen, M.; Rigby, I.; Dagens, A.; Dahmash, D.; Cheng, V.; Joseph, R.; Lipworth, S.; Harriss, E.; Cai, E.; et al. An Evaluation of Global Chikungunya Clinical Management Guidelines: A Systematic Review. EClinicalMedicine 2022, 54, 101672. [CrossRef]
- Law, Y.-S.; Wang, S.; Tan, Y.B.; Shih, O.; Utt, A.; Goh, W.Y.; Lian, B.-J.; Chen, M.W.; Jeng, U.-S.; Merits, A.; et al. Interdomain Flexibility of Chikungunya Virus nsP2 Helicase-Protease Differentially Influences Viral RNA Replication and Infectivity. J. Virol. 2021, 95. [CrossRef]
- Tan, Y.B.; Chmielewski, D.; Law, M.C.Y.; Zhang, K.; He, Y.; Chen, M.; Jin, J.; Luo, D. Molecular Architecture of the Chikungunya Virus Replication Complex. Sci. Adv. 2022, 8, eadd2536. [CrossRef]
- Borges, P.H.O.; Ferreira, S.B.; Silva, F.P., Jr Recent Advances on Targeting Proteases for Antiviral Development. Viruses 2024, 16, 366. [CrossRef]
- Behrouz, S.; Kühl, N.; Klein, C.D. N-Sulfonyl Peptide-Hybrids as a New Class of Dengue Virus Protease Inhibitors. Eur. J. Med. Chem. 2023, 251, 115227. [CrossRef]
- Dansana, J.; Purohit, P.; Panda, M.; Meher, B.R. Recent Advances in Phytocompounds as Potential Chikungunya Virus Non-Structural Protein 2 Protease Antagonists: A Systematic Review. Phytomedicine 2025, 136, 156359. [CrossRef]
- Rocha, D.C.P.; Sisnande, T.; Gavino-Leopoldino, D.; Guimarães-Andrade, I.P.; Cruz, F.F.; Assunção-Miranda, I.; Mendonça, S.C.; Leitão, G.G.; Simas, R.C.; Mohana-Borges, R.; et al. Antiviral, Cytoprotective, and Anti-Inflammatory Effect of Ampelozizyphus Amazonicus Ducke Ethanolic Wood Extract on Chikungunya Virus Infection. Viruses 2023, 15. [CrossRef]
- Elsaman, T.; Mohamed, M.S.; Eltayib, E.M.; Abdel-Aziz, H.A.; Abdalla, A.E.; Munir, M.U.; Mohamed, M.A. Isatin Derivatives as Broad-Spectrum Antiviral Agents: The Current Landscape. Med. Chem. Res. 2022, 31, 244–273. [CrossRef]
- De Moraes Gomes, P.A.T.; Pena, L.J.; Leite, A.C.L. Isatin Derivatives and Their Antiviral Properties against Arboviruses: A Review. Mini Rev. Med. Chem. 2019, 19, 56–62. [CrossRef]
- Kostova, I.; Saso, L. Advances in Research of Schiff-Base Metal Complexes as Potent Antioxidants. Curr. Med. Chem. 2013, 20, 4609–4632. [CrossRef]
- Tumosienė, I.; Jonuškienė, I.; Kantminienė, K.; Mickevičius, V.; Petrikaitė, V. Novel N-Substituted Amino Acid Hydrazone-Isatin Derivatives: Synthesis, Antioxidant Activity, and Anticancer Activity in 2D and 3D Models in Vitro. Int. J. Mol. Sci. 2021, 22, 7799. [CrossRef]
- Xu, Z.; Zhao, S.-J.; Lv, Z.-S.; Gao, F.; Wang, Y.; Zhang, F.; Bai, L.; Deng, J.-L. Fluoroquinolone-Isatin Hybrids and Their Biological Activities. Eur. J. Med. Chem. 2019, 162, 396–406. [CrossRef]
- Varun; Sonam; Kakkar, R. Isatin and Its Derivatives: A Survey of Recent Syntheses, Reactions, and Applications. Medchemcomm 2019, 10, 351–368. [CrossRef]
- Xu, J.; Xie, X.; Ye, N.; Zou, J.; Chen, H.; White, M.A.; Shi, P.Y.; Zhou, J. Design, Synthesis, and Biological Evaluation of Substituted 4,6-Dihydrospiro[[1,2,3]triazolo[4,5- B]pyridine-7,3′-Indoline]-2′,5(3 H)-Dione Analogues as Potent NS4B Inhibitors for the Treatment of Dengue Virus Infection. Journal of medicinal chemistry 2019, 62, 7941–7960. [CrossRef]
- Bal, T.R.; Anand, B.; Yogeeswari, P.; Sriram, D. Synthesis and Evaluation of Anti-HIV Activity of Isatin Beta-Thiosemicarbazone Derivatives. Bioorg. Med. Chem. Lett. 2005, 15, 4451–4455. [CrossRef]
- Zou, B.; Chan, W.L.; Ding, M.; Leong, S.Y.; Nilar, S.; Seah, P.G.; Liu, W.; Karuna, R.; Blasco, F.; Yip, A.; et al. Lead Optimization of Spiropyrazolopyridones: A New and Potent Class of Dengue Virus Inhibitors. ACS Med. Chem. Lett. 2015, 6, 344–348. [CrossRef]
- Mishra, P.; Kumar, A.; Mamidi, P.; Kumar, S.; Basantray, I.; Saswat, T.; Das, I.; Nayak, T.K.; Chattopadhyay, S.; Subudhi, B.B.; et al. Inhibition of Chikungunya Virus Replication by 1-[(2-Methylbenzimidazol-1-Yl) Methyl]-2-Oxo-Indolin-3-Ylidene] Amino] thiourea(MBZM-N-IBT). Sci. Rep. 2016, 6, 20122. [CrossRef]
- Lebeau, P.F.; Chen, J.; Byun, J.H.; Platko, K.; Austin, R.C. The Trypan Blue Cellular Debris Assay: A Novel Low-Cost Method for the Rapid Quantification of Cell Death. MethodsX 2019, 6, 1174–1180. [CrossRef]
- Cunha, M.S.; Cruz, N.V.G.; Schnellrath, L.C.; Medaglia, M.L.G.; Casotto, M.E.; Albano, R.M.; Costa, L.J.; Damaso, C.R. Autochthonous Transmission of East/Central/South African Genotype Chikungunya Virus, Brazil. Emerg. Infect. Dis. 2017, 23, 1737–1739. [CrossRef]
- Lanciotti, R.S.; Calisher, C.H.; Gubler, D.J.; Chang, G.J.; Vorndam, A.V. Rapid Detection and Typing of Dengue Viruses from Clinical Samples by Using Reverse Transcriptase-Polymerase Chain Reaction. J. Clin. Microbiol. 1992, 30, 545–551. [CrossRef]
- Freire, M.C.L.C.; Basso, L.G.M.; Mendes, L.F.S.; Mesquita, N.C.M.R.; Mottin, M.; Fernandes, R.S.; Policastro, L.R.; Godoy, A.S.; Santos, I.A.; Ruiz, U.E.A.; et al. Characterization of the RNA-Dependent RNA Polymerase from Chikungunya Virus and Discovery of a Novel Ligand as a Potential Drug Candidate. Sci. Rep. 2022, 12, 10601. [CrossRef]
- Law, Y.-S.; Utt, A.; Tan, Y.B.; Zheng, J.; Wang, S.; Chen, M.W.; Griffin, P.R.; Merits, A.; Luo, D. Structural Insights into RNA Recognition by the Chikungunya Virus nsP2 Helicase. Proc. Natl. Acad. Sci. U. S. A. 2019, 116, 9558–9567. [CrossRef]
- Olomola, T.O.; Bada, D.A.; Obafemi, C.A. Synthesis and Antibacterial Activity of Two Spiro [indole] Thiadiazole Derivatives. Toxicol. Environ. Chem. 2009, 91, 941–946. [CrossRef]
- Saranya, S.; Haribabu, J.; Vadakkedathu Palakkeezhillam, V.N.; Jerome, P.; Gomathi, K.; Rao, K.K.; Hara Surendra Babu, V.H.; Karvembu, R.; Gayathri, D. Molecular Structures, Hirshfeld Analysis and Biological Investigations of Isatin Based Thiosemicarbazones. J. Mol. Struct. 2019, 1198, 126904. [CrossRef]
- Haj Mohammad Ebrahim Tehrani, K.; Hashemi, M.; Hassan, M.; Kobarfard, F.; Mohebbi, S. Synthesis and Antibacterial Activity of Schiff Bases of 5-Substituted Isatins. Chin. Chem. Lett. 2016, 27, 221–225. [CrossRef]
- Yu, B.; Qi, P.-P.; Shi, X.-J.; Huang, R.; Guo, H.; Zheng, Y.-C.; Yu, D.-Q.; Liu, H.-M. Efficient Synthesis of New Antiproliferative Steroidal Hybrids Using the Molecular Hybridization Approach. Eur. J. Med. Chem. 2016, 117, 241–255. [CrossRef]
- Huong, T.-T.-L.; Dung, D.-T.-M.; Huan, N.-V.; Cuong, L.-V.; Hai, P.-T.; Huong, L.-T.-T.; Kim, J.; Kim, Y.-G.; Han, S.-B.; Nam, N.-H. Novel N-Hydroxybenzamides Incorporating 2-Oxoindoline with Unexpected Potent Histone Deacetylase Inhibitory Effects and Antitumor Cytotoxicity. Bioorg. Chem. 2017, 71, 160–169. [CrossRef]
- Silva, M.; Gonçalves, J.C.O.; Oliveira-Campos, A.M.F.; Rodrigues, L.M.; Esteves, A.P. Synthesis of Novel Glycoconjugates Derived from Alkynyl Heterocycles through a Click Approach. Synth. Commun. 2013, 43, 1432–1438. [CrossRef]
- Kumar, A.; Lal, K.; Kumar, V.; Murtaza, M.; Jaglan, S.; Paul, A.K.; Yadav, S.; Kumari, K. Synthesis, Antimicrobial, Antibiofilm and Computational Studies of Isatin-Semicarbazone Tethered 1,2,3-Triazoles. Bioorg. Chem. 2023, 133, 106388. [CrossRef]
- Pohjala, L.; Utt, A.; Varjak, M.; Lulla, A.; Merits, A.; Ahola, T.; Tammela, P. Inhibitors of Alphavirus Entry and Replication Identified with a Stable Chikungunya Replicon Cell Line and Virus-Based Assays. PLoS One 2011, 6, 1–13. [CrossRef]
- Gigante, A.; Gómez-SanJuan, A.; Delang, L.; Li, C.; Bueno, O.; Gamo, A.-M.; Priego, E.-M.; Camarasa, M.-J.; Jochmans, D.; Leyssen, P.; et al. Antiviral Activity of [1,2,3]triazolo[4,5- D ]pyrimidin-7(6 H )-Ones against Chikungunya Virus Targeting the Viral Capping nsP1. Antiviral Res. 2017, 144, 216–222. [CrossRef]
- Khan, M.; Dhanwani, R.; Patro, I.K.; Rao, P.V.L.; Parida, M.M. Cellular IMPDH Enzyme Activity Is a Potential Target for the Inhibition of Chikungunya Virus Replication and Virus Induced Apoptosis in Cultured Mammalian Cells. Antiviral Res. 2011, 89, 1–8. [CrossRef]
- Ferreira, A.C.; Reis, P.A.; de Freitas, C.S.; Sacramento, C.Q.; Villas Bôas Hoelz, L.; Bastos, M.M.; Mattos, M.; Rocha, N.; Gomes de Azevedo Quintanilha, I.; da Silva Gouveia Pedrosa, C.; et al. Beyond Members of the Flaviviridae Family, Sofosbuvir Also Inhibits Chikungunya Virus Replication. Antimicrob. Agents Chemother. 2019, 63. [CrossRef]
- Abdelnabi, R.; Jochmans, D.; Verbeken, E.; Neyts, J.; Delang, L. Antiviral Treatment Efficiently Inhibits Chikungunya Virus Infection in the Joints of Mice during the Acute but Not during the Chronic Phase of the Infection. Antiviral Res. 2018, 149, 113–117. [CrossRef]
- Lani, R.; Hassandarvish, P.; Chiam, C.W.; Moghaddam, E.; Chu, J.J.H.; Rausalu, K.; Merits, A.; Higgs, S.; Vanlandingham, D.; Abu Bakar, S.; et al. Antiviral Activity of Silymarin against Chikungunya Virus. Sci. Rep. 2015, 5, 11421. [CrossRef]
- Abu Bakar, F.; Ng, L.F.P. Nonstructural Proteins of Alphavirus-Potential Targets for Drug Development. Viruses 2018, 10. [CrossRef]
- Battisti, V.; Urban, E.; Langer, T. Antivirals against the Chikungunya Virus. Viruses 2021, 13, 1307.
- Navarro, H.; Scott, J.E.; Smith, G.R.; Ghiabi, P.; Gibson, E.; Loppnau, P.; Harding, R.J.; Hossain, M.A.; Bose, M.R.; Pearce, K.H.; et al. Identification of Inhibitors of Chikungunya Virus nsP2 ATPase. bioRxivorg 2025.
- Santana, A.C.; Silva Filho, R.C.; Menezes, J.C.J.M.D.S.; Allonso, D.; Campos, V.R. Nitrogen-Based Heterocyclic Compounds: A Promising Class of Antiviral Agents against Chikungunya Virus. Life (Basel) 2020, 11, 16. [CrossRef]
- Metibemu, D.S.; Adeyinka, O.S.; Falode, J.; Crown, O.; Ogungbe, I.V. Inhibitors of the Structural and Nonstructural Proteins of Alphaviruses. ACS Infect. Dis. 2024, 10, 2507–2524. [CrossRef]
- Furuta, Y.; Gowen, B.B.; Takahashi, K.; Shiraki, K.; Smee, D.F.; Barnard, D.L. Favipiravir (T-705), a Novel Viral RNA Polymerase Inhibitor. Antiviral Res. 2013, 100, 446–454. [CrossRef]
- Rabelo, V.W.-H.; Paixão, I.C.N. de P.; Abreu, P.A. Targeting Chikungunya Virus by Computational Approaches: From Viral Biology to the Development of Therapeutic Strategies. Expert Opin. Ther. Targets 2020, 24, 63–78. [CrossRef]
- Wang, M.; Wang, L.; Leng, P.; Guo, J.; Zhou, H. Drugs Targeting Structural and Nonstructural Proteins of the Chikungunya Virus: A Review. Int. J. Biol. Macromol. 2024, 262, 129949. [CrossRef]
Figure 1.
Clinically successful isatin-based drugs such as semaxanib (2), sunitinib (3), and toceranib (4), which have been applied in the treatment of various diseases, along with potential antiviral isatin and hydrazone derivatives (5-9) [18,19,20,21,22,23].
Scheme 1.
Synthetic route used to prepare compounds 11-14.
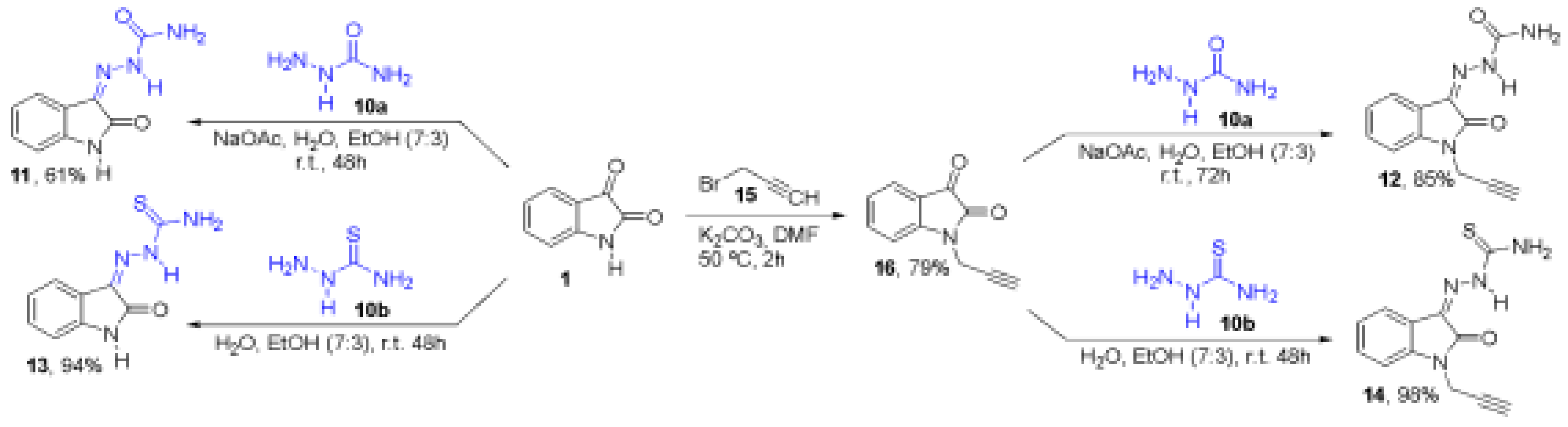
Figure 2.
GFP fluorescence in BHK-CHIKV-NCT cells treated with Isatin and Isatin derivatives 11-14 for 24 h. Error bars represent the standard deviation from three independent experiments. The colors indicate the different tested compounds, while the dashed lines at 75% and 50% mark the fluorescence reduction in response to treatment with the compounds. Statistical analysis was performed using one-way ANOVA. (n = 3) *<0,1; **<0,01; ***< 0,001; ****< 0,0001.
Figure 2.
GFP fluorescence in BHK-CHIKV-NCT cells treated with Isatin and Isatin derivatives 11-14 for 24 h. Error bars represent the standard deviation from three independent experiments. The colors indicate the different tested compounds, while the dashed lines at 75% and 50% mark the fluorescence reduction in response to treatment with the compounds. Statistical analysis was performed using one-way ANOVA. (n = 3) *<0,1; **<0,01; ***< 0,001; ****< 0,0001.
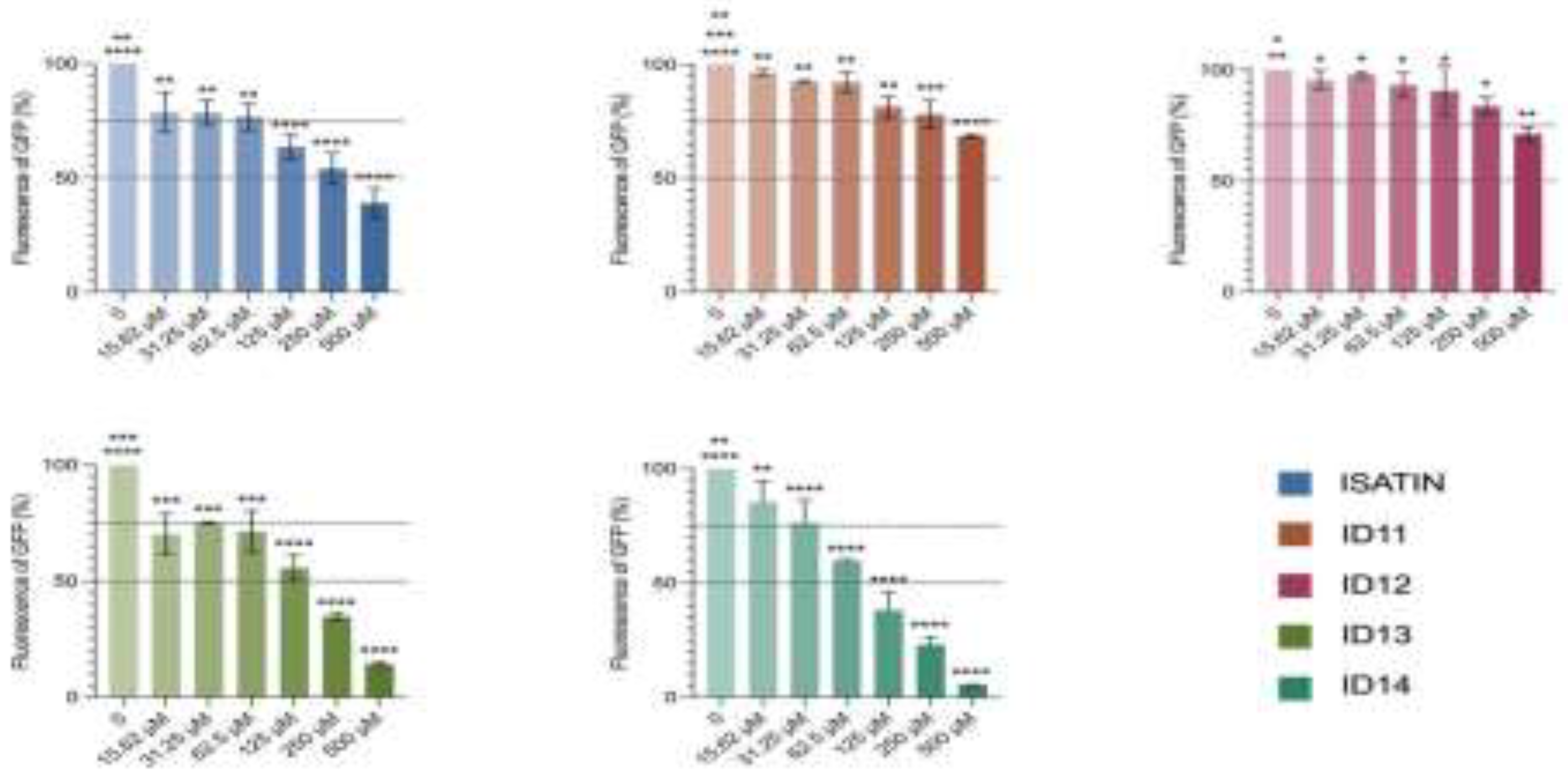
Figure 3.
The dose-dependent curves of compounds with the anti-CHIKV activity. BHK-21 cells infected with CHIKV MOI 0.2 were treated with Isatin and ID11-14, at concentrations curve (15 to 500 µM) for 24h at 37 ºC, 5% CO2. (n=3).
Figure 3.
The dose-dependent curves of compounds with the anti-CHIKV activity. BHK-21 cells infected with CHIKV MOI 0.2 were treated with Isatin and ID11-14, at concentrations curve (15 to 500 µM) for 24h at 37 ºC, 5% CO2. (n=3).
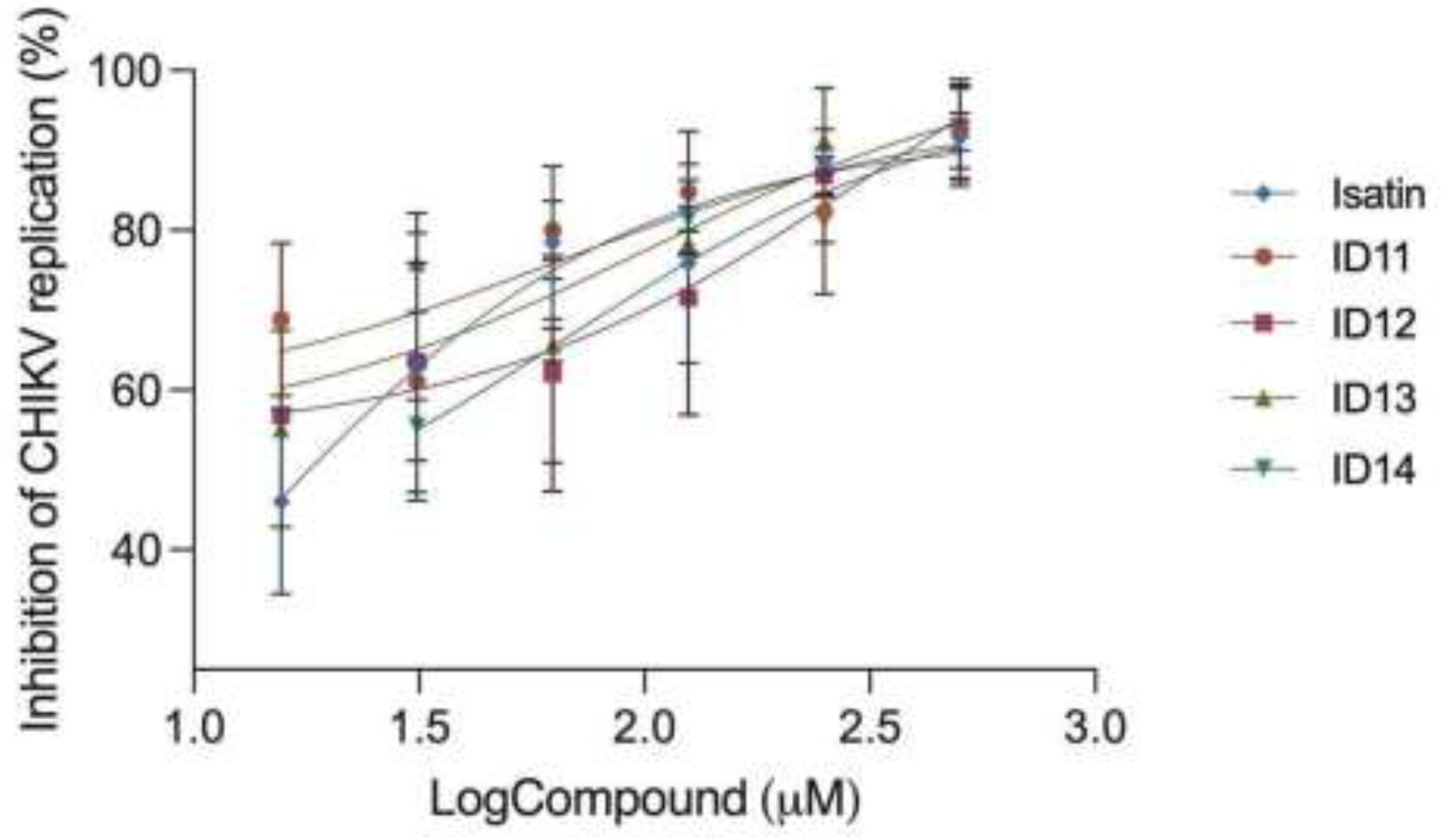
Figure 4.
Virus Entry Inhibition Assay. Cells were treated with the compounds simultaneously with the viral particle infection at MOI = 0.2. Treatment was performed using ID12 and ID14 at 32 μM and Isatin, ID11, and ID13 at 20 μM, based on their IC50 values obtained from antiviral activity assays with infectious CHIKV particles. No significant differences were observed between treated and untreated (NT) conditions. Statistical analysis was performed using one-way ANOVA. (n = 3). .
Figure 4.
Virus Entry Inhibition Assay. Cells were treated with the compounds simultaneously with the viral particle infection at MOI = 0.2. Treatment was performed using ID12 and ID14 at 32 μM and Isatin, ID11, and ID13 at 20 μM, based on their IC50 values obtained from antiviral activity assays with infectious CHIKV particles. No significant differences were observed between treated and untreated (NT) conditions. Statistical analysis was performed using one-way ANOVA. (n = 3). .
Figure 5.
Molecular docking representation of Isatin and ID11-14 with CHIKV nsP2. The figure presents both three-dimensional (A–E) and two-dimensional (F–J) representations of the docking interactions between nsP2 and the tested compounds. Panels A to E correspond to the 3D docking interactions of Isatin and ID11-14, respectively, while panels F to J display the 2D interaction maps for the same compounds. Key binding residues and interaction types, such as hydrogen bonds and hydrophobic interactions, are highlighted. Images generated with PyMOL© and LigPlot ©.
Figure 5.
Molecular docking representation of Isatin and ID11-14 with CHIKV nsP2. The figure presents both three-dimensional (A–E) and two-dimensional (F–J) representations of the docking interactions between nsP2 and the tested compounds. Panels A to E correspond to the 3D docking interactions of Isatin and ID11-14, respectively, while panels F to J display the 2D interaction maps for the same compounds. Key binding residues and interaction types, such as hydrogen bonds and hydrophobic interactions, are highlighted. Images generated with PyMOL© and LigPlot ©.
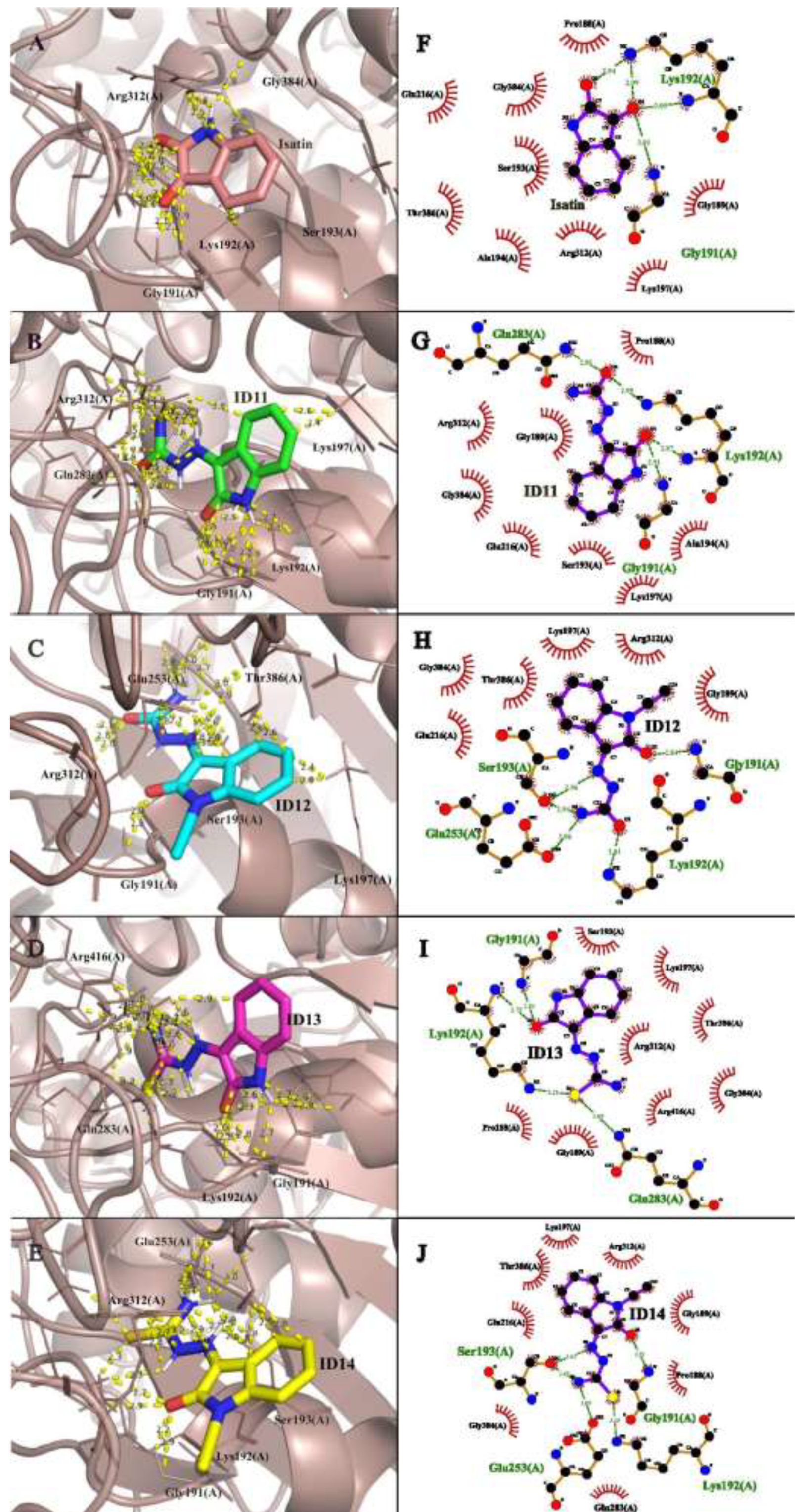

Table 1.
Anti-CHIKV activity of compounds Isatin and ID11-14 in BHK-CHIKV-NCT cells and against infectious CHIKV particles*.
Table 1.
Anti-CHIKV activity of compounds Isatin and ID11-14 in BHK-CHIKV-NCT cells and against infectious CHIKV particles*.
| Compound | EGFPEC50 (μM) | EC50 (μM) | CC50 (μM) | SI (CC50 / EC50) |
| Isatin | 340.6 | 20.2 | >500 | 24.7 |
| ID11 | >500 | 24.6 | >500 | 34.2 |
| ID12 | >500 | 32.6 | >500 | 15.3 |
| ID13 | 189.7 | 20.0 | >500 | 25 |
| ID14 | 152.92 | 33.6 | >500 | 14.9 |
Table 2.
Molecular docking results for isatins derivatives compounds and NSP2 and NSP4 CHIKV Proteins.
Table 2.
Molecular docking results for isatins derivatives compounds and NSP2 and NSP4 CHIKV Proteins.
| Compound | NSP2 | NSP4 |
| Score | Score | |
| simone..tammaro | -31.3 ± 0.1 | -27.4 ± 0.2 |
| simone..tammaro | -33.8 ± 0.1 | -29.8 ± 0.4 |
| simone..tammaro | -32.9 ± 0.5 | -31.9 ± 0.2 |
| simone..tammaro | -32.5 ± 0.3 | -29.8 ± 0.2 |
| simone..tammaro | -34.9 ± 0.5 | -32.4 ± 0.3 |
| simone..tammaro | -21.1±0.2 | -22.6±0.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



