
Submitted:
14 January 2026
Posted:
14 January 2026
You are already at the latest version
Abstract
DNA-based cancer vaccines represent a safe and promising immunotherapeutic strategy, but their clinical efficacy is often limited by weak immunogenicity, primarily due to inefficient antigen cross-presentation. To overcome this challenge, the MHC class I trafficking domain (MITD) can be fused to tumor antigens to enhance their intracellular routing in dendritic cells (DCs), thereby promoting the efficiency of cross-presentation. In addition, incorporation of CD4⁺ T cell epitopes, such as PADRE or P2P16, can robustly activate CD4⁺ T cells, further amplifying antitumor immunity. Thus, combining MITD with CD4⁺ epitopes is expected to synergistically improve DNA vaccine potency. Mesothelin (MSLN), a tumor-associated antigen highly expressed in pancreatic cancer, was selected as the target in this study. We designed MSLN-targeted DNA vaccines incorporating MITD together with either PADRE or P2P16. In a Panc02 murine model, the MITD–PADRE construct, a novel design, elicited stronger immune responses and more effective antitumor activity compared to other formulations. To further counteract immunosuppression, we combined the vaccine with gemcitabine, which enhanced therapeutic efficacy. Together, these findings demonstrate that integrating PADRE with MITD in MSLN-targeted DNA vaccines offers a promising combinatorial strategy for advancing pancreatic cancer immunotherapy.
Keywords:
immunotherapy
; DNA vaccines
; MITD
; PADRE
; cross-presentation
1. Introduction
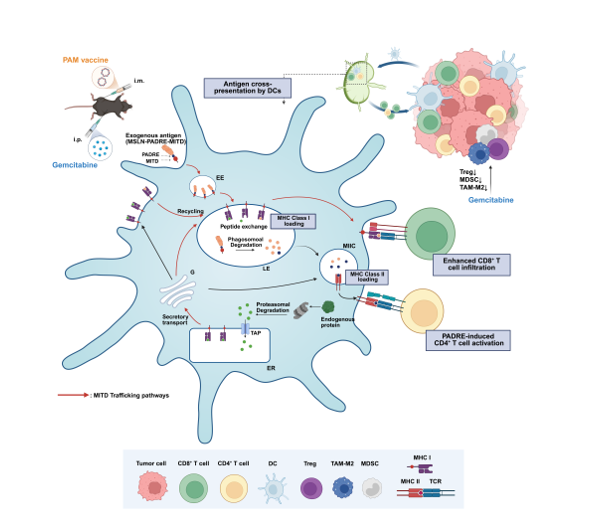
Pancreatic cancer ranks among the most lethal malignancies, accounting for 4.8% of cancer-related deaths, underscoring an urgent need for novel therapeutic strategies [1,2]. While cancer immunotherapy has made extraordinary strides in recent years, DNA tumor vaccines are particularly noteworthy due to their ability to activate CD8+ T cell immunity, high safety profile, and ease of production and storage [3]. However, obstacles such as poor immunogenicity have hindered the clinical translation of DNA vaccines. An effective DNA cancer vaccine must induce robust and sustained T cell responses, which critically depend on efficient antigen processing and presentation via MHC class I molecules. Notably, antigen cross-presentation by MHC class I molecules is an exceptionally inefficient process, with only ~150 peptides presented per second out of approximately 2 million generated [4,5,6,7].
The MHC class I trafficking domain (MITD), located within the cytoplasmic domain of the MHC class I molecule, contains a tyrosine-based sorting signal responsible for its recycling between endolysosomal compartments [8]. Cross-presentation of exogenous antigens by major histocompatibility complex (MHC) class I molecules is primarily mediated through two distinct pathways: the vacuolar pathway and the cytosolic pathway. In the vacuolar pathway, exogenous antigens are internalized via endocytosis and trafficked through endosomal compartments to lysosomes, where they are proteolytically degraded into antigenic peptides. Concurrently, MHC class I molecules are recycled from the plasma membrane or directly delivered from the Golgi apparatus to late endosomes, where the acidic microenvironment facilitates peptide exchange and loading. In the cytosolic pathway, internalized antigens are translocated from endosomal compartments into the cytosol and degraded by the proteasome. The shortened peptides are subsequently transported into the endoplasmic reticulum (ER) via TAP for loading onto MHC class I molecules, followed by trafficking to the cell surface. Alternatively, proteasome-generated peptides may be reimported into endosomal compartments, where they are loaded onto MHC class I molecules prior to surface presentation. The MITD mediates the targeting of MHC class I molecules to endosomal compartments through its tyrosine-based sorting motif (YXXA), thereby facilitating the loading of exogenous peptides and enhancing antigen cross-presentation [9]. Recent studies indicate that antigens conjugated to MITD exploit this dynamic trafficking pathway. They are rerouted to specific cellular compartments associated with antigen processing and presentation—including the endoplasmic reticulum (ER), Golgi apparatus, plasma membrane, lysosomes, and endosomes—depending on the maturation stage of dendritic cells (DCs)[7,10,11]. This precise intracellular compartmentalization consequently facilitates efficient antigen cross-presentation [4,8,12].
Incorporation of T-cell epitope peptides into cancer vaccines has been shown to promote immune cell activation, thereby enhancing vaccine immunogenicity.[4,13]. Consequently, incorporating a CD4+ T cell epitope into the MITD construct represents a potential strategy to further enhance CD4+ T cell activation. The universal CD4+ T cell epitope P2P16, derived from tetanus toxin, has been widely used in clinical applications. For example, the first clinical-stage tumor vaccine BNT116 employed a P2P16-MITD sequence as combined adjuvant elements [14,15,16]. Alternatively, the pan-DR epitope (PADRE), a synthetic peptide engineered to bind diverse HLA-DR alleles, has been shown to elicit more potent CD4+ T cell responses than P2P16[17,18].Therefore, attaching PADRE to the MITD sequence offers a novel strategy to enhance cellular immune responses induced by DNA vaccines targeting pancreatic cancer.
The selection of an appropriate antigen is critical for the efficacy of DNA tumor vaccines [19]. Mesothelin (MSLN), a GPI-anchored cell surface glycoprotein, is synthesized as a 69-kDa precursor protein and subsequently cleaved into two products: megakaryocyte potentiating factor (MPF) and mature membrane-bound MSLN [20,21,22,23,24]. Many MSLN-targeted therapeutic approaches for pancreatic cancer have been developed, including MSLN-directed antibodies, CAR-T cells, and vaccines. As a tumor-associated antigen, MSLN is highly expressed in pancreatic tumors but exhibits minimal expression in normal human tissues [25,26]. It is also reported to promote tumor cell invasion and migration by inducing epithelial - mesenchymal transition (EMT), significantly worsening pancreatic cancer prognosis [19,27,28]. While MSLN exhibits moderate intrinsic immunogenicity, its tumor - restricted expression and functional role in malignancy collectively make it as an ideal target for pancreatic cancer immunotherapy.
Dendritic cells (DCs) uptake, process, and present tumor antigens via MHC class I or II molecules, with cross-presentation being a critical pathway for MHC class I loading [29]. As type 1 conventional dendritic cells (cDC1s) are the primary mediators of MHC class I cross-presentation, our previous research built upon cDC1-targeting strategy via a vaccine platform based on XCL1 fusion [30]. In this study, we employed two distinct chimeric elements—P2P16-MITD and PADRE-MITD—to enhance both CD4+ helper T cell activation and cross-presentation to promoting immunogenicity of MSLN-targeted DNA vaccines. Furthermore, to reducing immunosuppression within the tumor microenvironment, we combined DNA vaccine with gemcitabine, aiming to boosting their antitumor efficacy [31,32]. Our findings demonstrate that this combination therapy significantly enhances antitumor responses in a murine pancreatic cancer model, establishing a promising translational approach for pancreatic cancer treatment.
2. Results
2.1. The Construction and Validation of DNA Vaccines
Mesothelin is a tumor-associated antigen which is highly expressed in pancreatic tumor cells. Therefore, we initially constructed a MSLN-targeted DNA vaccine based on the vaccine platform we early reported [30]. The DNA vaccine encodes a fusion protein which comprised of human XCL1(hXCL1) to target cDC1, MSLN and P2A peptide-linked IL-2 as adjuvant. Then we constructed two additional vaccines: CpVR-hXCL1/MSLN/PADRE-IL-2 (M-PA) and CpVR-hXCL1/MSLN/P2P16-IL-2 (M-P2) to compare the additive effects of the two CD4+ T cell epitopes PADRE/P2P16 on the DNA vaccines. Furthermore, to enhance the efficiency of antigen cross-presentation, we constructed two more DNA vaccines PAM/P2M by incorporating MITD to the end of the PADRE/P2P16 gene respectively (Figure 1A). The plasmid was verified by double digestion with Pst I/BamH I (Figure 1B), and the expression of the fusion proteins was analyzed by Westen blotting using anti-MSLN antibody (Figure 1C).
2.2. The Incorporation of MITD Significantly Enhances the Immunogenicity of DNA Vaccines
C57BL/6 mice were immunized with 100μg DNA vaccines by fast DNA immunization strategy [33] on day 0, 2, and 5 to compare the immunogenicity of the vaccines in vivo. An ELISpot assay was utilized to detect the IFN-γ secretion by antigen-specific T cells. Upon stimulation with MSLN protein, all five vaccines induced MSLN-specific T cell to produce IFN-γ, with a significant difference in contrast to the VR control group, demonstrating the potent immunogenicity of MSLN. In comparison to the M-P2 group, the M-PA group induced a greater number of IFN-γ spots, while the P2M group generated more IFN-γ spots than the PAM group upon the incorporation of MITD (Figure 2A, B). MSLN-specific antibody levels in mouse sera after immunization were measured by ELISA (Figure 2C). All five vaccine groups exhibited significantly elevated anti-MSLN antibody responses compared to the VR group, with PAM group showing the highest level, but the difference was not statistically significant. In more details, when P2P16 or PADRE was incorporated individually into the vaccine constructs, M-P2 induced a higher frequency of both CD8+ and CD4+ T cells (Figure 2D, E), while the M-PA group demonstrated greater proliferative capacity of CD8+ and CD4+ T cells. Neither M-PA nor M-P2 elicited a statistically significant enhancement in the induction of CD4+ and CD8+ T cell responses compared to the VR group. The addition of MITD to the vaccine markedly promoted CD4+ and CD8+ T cell proliferation in the PAM group, indicating its potential role in enhancing cellular immune response (Figure 2F, G). In the immunogenicity test, PAM and P2M each demonstrated distinct advantages in across different aspects; therefore, a further evaluation of their antitumor efficacy will be conducted in subsequent tumor challenge.
2.3. PADRE-MITD Exhibits Superior Antitumor Efficacy Compared with P2P16-MITD in a Murine Pancreatic Cancer Model
Next, we evaluated the therapeutic efficacy of MITD-incorporated DNA vaccines on MSLN stabled expressing pancreatic cancer (MSLN-EGFP+ Panc02) mouse model (Figure S1A,B). C57BL/6 mice (n=8-12) were inoculated with 2×106 MSLN-EGFP+ Panc02 cells (Figure S1C) on day 0 and received treatment on day 8. Based on fast DNA immunization strategy described above, two additional immunizations were administered on the day 9 (day 22) and day 11 (day 24) with 100μg vaccine dose each time (Figure 3A). Our previous research demonstrated that this regimen achieves better anti-tumor immune responses (data not show). Tumor growth in mice administered MSLN, PAM, and P2M was significant suppressed compared to VR (Figure 3B,C). Moreover, the average tumor size and weight in all three therapeutic groups were smaller than those in the VR group, particularly PAM.
In ELISpot assay, all three vaccine groups-MSLN, PAM and P2M revealed significant cellular immune response (Figure 3D,E). Notably, PAM induced the highest numbers of IFN-γ secreting MSLN-specific cells, with a significant increase compared to the other two groups. Then, we performed a flow cytometric analysis of immune cell populations in the spleen and tumor. In the spleen, the proportion of CD4+ T cells and CD8+ T cells in both PAM and P2M elicited significant increase compared to the VR group (Figure 3F). Similarly, the most significant enhancement in the levels and proliferation of CD4+ and CD8+ T cell were observed in the tumor among all the groups. (Figure 3G,H). The GzmB and Th1 cytokine (IFN-γ, IL-2, TNF-α) levels in tumor tissues were measured by qRT-PCR (Figure 3I,J), which showed significant elevation of GzmB and Th1 cytokine levels in all three therapeutic groups, with PAM exhibiting the highest levels of GzmB, indicating the strongest CD8+ T cell and NK cell cytotoxicity, as well as PAM showing the highest IFN-γ and TNF-α levels, reflecting its robust cellular immunity.
2.4. PAM Induces Relatively Weaker Immunosuppression in Tumor Microenvironment
To further investigate the factors hindering the antitumor effects of the vaccines, we conducted experiments to analyze immunosuppressive cells infiltrating into the tumor by flow cytometry and assessed Th2 cytokine expression in the tumor tissues by qRT-PCR. Both PAM and P2M group showed a significant increase in intratumoral MDSCs and Tregs (Figure 4A,B), with the P2M group exhibiting higher levels, though the difference was not statistically significant. We also analyzed intratumoral tumor-associated macrophages (TAMs), including the pro-inflammatory M1 and immunosuppressive M2 subtypes. The PAM group showed significantly higher levels of M1-TAMs compared to the other two therapeutical groups, suggesting an ascended pro-inflammatory potential. (Figure 4C) In contrast, the proportion of M2-TAMs was significantly reduced in all treatment groups compared to the VR group. Although both were effective, the PAM group showed a greater reduction than the P2M group, suggesting that PAM induces a lower level of immunosuppression. (Figure 4D). The M1/M2 ratio was used to assess the polarization status of TAMs. Compared to the VR group, all three therapeutic groups showed an inclination toward a pro-immunogenetic phenotype, with the PAM group indicating the highest degree of polarization, which was significantly different from the P2M group (Figure 4E).
Regarding of the Th2 cytokines, all three therapeutic groups showed increased expression of IL-4, IL-6, and IL-10 compared to the VR group, with the P2M group exhibiting the highest levels (Figure 4F). Elevated Th2 cytokines are associated with enhanced immunosuppression and reduced antitumor efficacy of the vaccines. These finding are consistent with the increased proportion of Tregs detected by flow cytometry which is described above, suggesting the PAM group exhibits comprehensively highest antitumor efficacy and relatively low immunosuppression, and indicating that combining PAM vaccine with strategies that alleviate immunosuppression might be a potential approach to further enhance its antitumor immunity.
2.5. Combination Therapy with Gem Elevated the Antitumor Efficacy of PAM
To further alleviate the immunosuppression of the DNA vaccines, we combined Gemcitabine with the PAM which proved the most efficient treatment regimen to test the therapeutic effect in pancreatic cancer mouse model [34,35]. C57BL/6 mice were inoculated 2×106 MSLN-EGFP+ Panc02 cells and started treatment on day 8 after tumor challenge. The mice received three injections of 100μg of DNA vaccines on day 8, day 10 and day 13, along with intraperitoneal administration of Gem on day 8, day 11, day 15 and day 18 with a dose of 15 mg/kg (Figure 5A). The tumors tissues were harvested at day 23, and the size and weight were shown in Figure 5B. Either single or combined-treatment demonstrated pronounced suppression to the tumor growth. In particular, mice treated with the combination of PAM and Gem revealed significantly lower volume and weight than in the VR group or in each monotherapy groups. The body weights of all mice remained within normal range throughout the treatment period (Figure 5C). No obvious differences in general health were observed in the Gem-treated group, suggesting no distinct toxicity at the given dose. Elispot assay showed that the combination therapy induced significantly higher IFN-γ secretion compared to either PAM or Gem monotherapy, reflecting the synergistic effect between the two agents and the efficacy of the combination therapy (Figure 5D).
Moreover, the proportion of CD4+ T cells in splenocytes observed a significantly increase in the combination group compared to the VR group (Figure 5E), however, CD8+ T cells in splenocytes did not observe significant difference between three therapeutic groups (data not show). Similarly, the intratumoral infiltration of CD45+ lymphocytes, CD4+T cells and CD8+ T cells in the combination group were increased (Figure 5F), exhibiting significantly higher levels than either the DNA vaccine or / and Gem monotherapy groups, with the proliferation of CD4+ T cells and CD8+ T cells remained the highest level in the combination group (Figure 5G). Additionally, intratumoral infiltration of other immune cells, including natural killer (NK) cells and dendritic cells (DCs), were also assessed (Fig. 5H). As the results showed, the combination group exhibited the highest level of NKs and DCs compared to either monotherapy group, further supporting the enhanced immunostimulatory effect of the combined treatment.
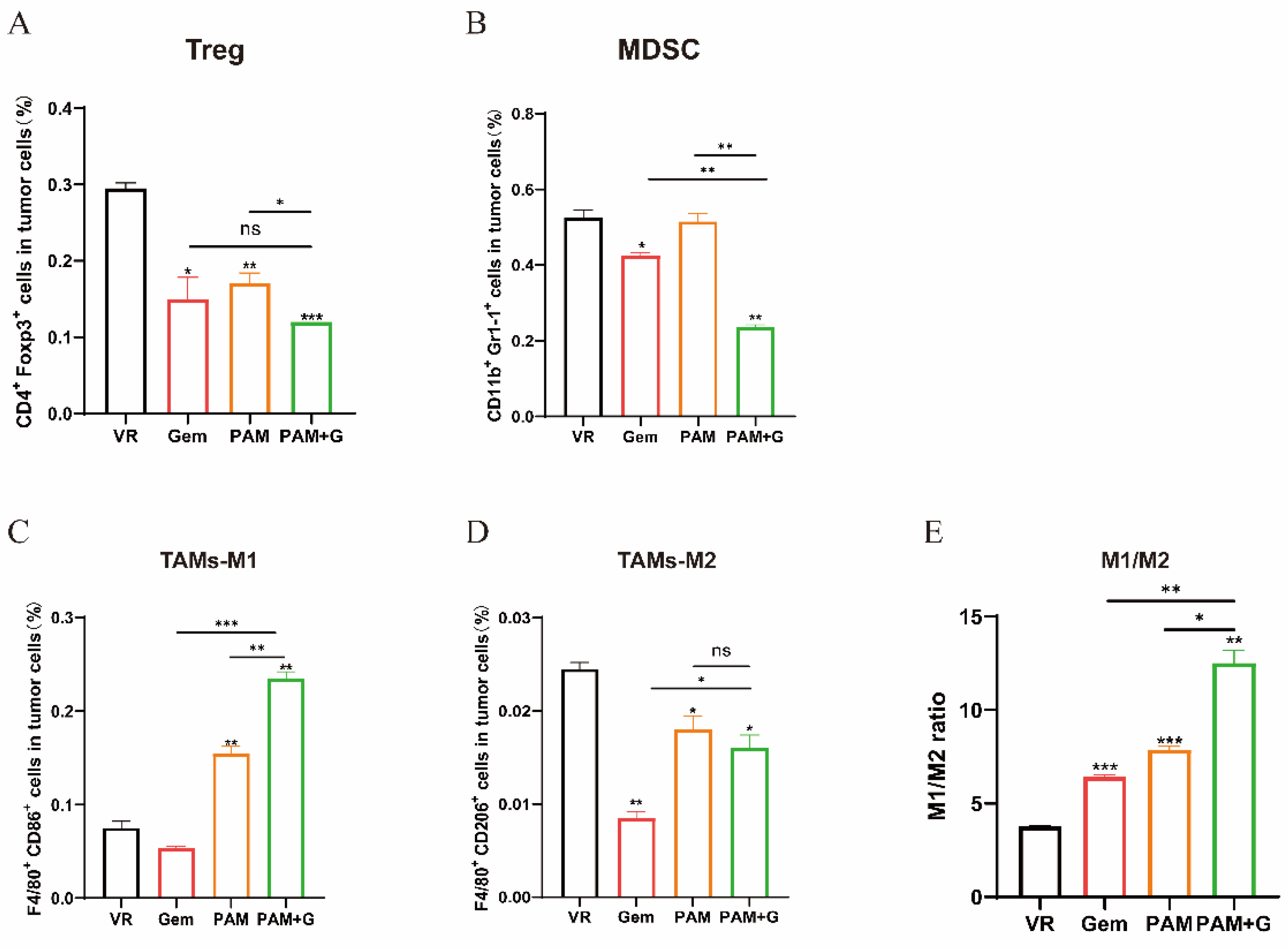
2.6. PAM Combined with Gemcitabine Further Reduces Immunosuppression in Tumor Microenvironment
To elucidate the effect of combination therapy on the tumor immunosuppressive microenvironment, we analyzed the intratumoral infiltrated immunosuppressive cells. Both Tregs and MDSCs in the combination group indicated a significant decrease in contrast to either the VR group or the PAM vaccine monotherapy groups (Figure 6A,B). The combination group induced significantly higher levels of M1-TAMs and lower levels of M2-TAMs compared to the other groups, the M1/M2 ratio indicated a pronounced shift toward a pro-immunogenic phenotype, which was the strongest among all the therapeutic groups and thus interprets a potential factor for the superior antitumor efficacy of the combination group as is presented in tumor growth curve (Figure 6C–E).
3. Discussion
The efficacy of a DNA cancer vaccine is primarily influenced by factors such as appropriate antigen selection, efficient antigen presentation and induction of robust CD4+ T helper cells and Cytotoxic CD8+ T cells activation [19]. MSLN overexpression is observed in 80-85% of pancreatic cancer while its expression in normal tissues is minimal [36,37]. Therefore, MSLN serves as an ideal target for pancreatic cancer immunotherapy, providing high tumor specificity and reducing the risk of off-target toxicity in normal tissues. However, due to the inherently limited immunogenicity of DNA vaccines, the vaccine containing MSLN antigen alone is insufficient to elicit a robust immune response capable of activating both CD4+ and CD8+ T cells for effective antitumor activity [38,39]. CD4+T cells play a pivotal role in tumor immunity, which promotes the reprogramming of CD8+ T cells and facilitate the induction of both effector and memory CD8+ T cell responses, within the tumor microenvironment, CD4+ T helper cells secrete IL-2 to recruit the CD8+ T cells, promotes their proliferation and upregulate granzyme B expression for the killing of target tumor cells [40,41]. Thus, we incorporated two universal CD4+ T cell epitope candidates, P2P16 and PADRE which is capable of binding to diverse HLA-II molecules and stimulate CD4+ T cell activation. We compared their individual effects on immune enhancement, although the results indicated that PADRE contributed to stronger IFN-γ secretion than P2P16, simply incorporating a CD4+ T cell epitope was insufficient to profoundly enhance the immunogenicity through immune cell activation. Consequently, we focused on additional factors that could potentially affect the immunogenicity of the vaccines and further attached another adjuvant element MITD to the CD4+ T cell epitopes. As hypothesized, MITD-attached vaccines indeed emerged an enhanced capability of inducing antigen-specific immune responses, which is likely due to the intracellular trafficking function mediated by MITD in immature and mature DCs, boosting the efficacy of antigen cross-presentation. Intriguingly [4], P2P16 conjugated to MITD elicited a stronger IFN-γ secretion in ELISpot assay compare to PADRE-MITD, which is not compatible with the results observed when the two epitopes were used individually. Reports suggesting that antigens attached to MITC would be rerouted into compartments where peptides derived from MHC I and loaded on MHC II molecules, which might be more beneficial for the processing of P2P16 within the endosomal or ER-associated pathways [4,42,43].
Due to the distinct immunogenic fluctuation in P2P16 and PADRE conjugated to MITD, both constructs along with MSLN group were selected for the subsequential antitumor efficacy study. The results indicates that the PAM group exhibited remarkably superior antitumor effect and higher level of in vivo immune response compared to the P2M group. We noticed that the declining intratumoral immunosuppressive cells in the PAM group may underlie the better antitumor efficacy compare to the P2M group. This highlights the potential of combination therapy which conceived to mitigate the intratumoral immunosuppression. Several studies revealed that low-dose Gemcitabine could promote to the depletion of regulatory T cells and reduce MDSCs production [44,45,46]. Therefore, we combined Gem with PAM vaccine, our results showed that the combination therapy significantly decreased intratumoral MDSCs and Tregs, and inhibited tumor growth .
In conclusion, our finding first indicates that the incorporation of adjuvant elements regimen PADRE-MITD profoundly promoted the immunogenicity and antitumor effect of MSLN-targeted vaccine through CD4+ helper T cell activation and cross-presentation enhancement. To mitigate the immunosuppression associated with the DNA vaccine, a gemcitabine combination therapy was employed and observed promising therapeutic efficacy. Our findings provide a novel strategy for pancreatic cancer treatment that overcome the obstacles of previous MSLN-targeted DNA vaccines, such as the insufficiency in antigen cross-presentation and the immunosuppression induced within the tumor microenvironment, underscoring a critical approach in clinical trials.
4. Materials and Methods
4.1. Animals and Cell Lines
Female C57BL/6 mice (6-8 weeks old) were purchased from Liaoning Changsheng Biotechnology Co., Ltd. (Liaoning, China) and raised in the animal experiment platform of the College of Life Sciences, Jilin University. All procedures were performed in accordance with China Law and were approved by the Ethics Committees of Jilin University. The murine Panc02 cell lines were provided by the National Center for Nanoscience and Technology.
The stable-expressing MSLN-EGFP+ Panc02 cell line was constructed in our lab. The pLVX-MSLN-EGFP lentiviral expression vector was constructed and co-transfected into 293T cells with 4μg of pLVX-MSLN-EGFP, 5μg of psPAX2 and 2.5μg VSVG. Viral supernatants were collected and concentrated for subsequent use. Panc02 cells were seeded in 6-well plates and cultured at 37℃ in a CO2 incubator. Concentrated lentiviral particles were then added to each well along with 10μl of polybrene to enhance infection. After 24 hours, the medium was replaced with DMEM containing 10% FBS. Following an additional 48 hours of incubation, puromycin was added at a final concentration of 6μg/ml for selection. After 7 days of selection, positively transduced cells were sorted by flow cytometry and expanded for further use.
4.2. Vaccine Construction
The plasmid pLVX-MSLN was purchased from Wuhan Miaoling Biotechnology Co., Ltd. (Wuhan, China). The plasmid CpVR-P2M and CpVR-PAM were synthesized by Nanjing Jinsirui Biotechnology Co., Ltd. (Nanjing, China). The plasmids MSLN, M-PA, M-P2, PAM, and P2M were constructed using the Seamless Assembly Cloning Kit according to the manufacturer’s instructions. The inserts were cloned into the Pst I/BamH I sites of the VR1012 vector, using the previously constructed plasmid CpVR-XCL1-MUC1 from our laboratory as the template. All plasmid were validated through DNA sequencing and performed Western blot to analysis the expression of the target proteins.
4.3. Protein Purification
To perform the ELISpot assay, human MSLN protein was extracted and purified. MSLN gene with His tag was constructed onto the prokaryotic expression vector pET30a and was transformed into BL21 expression receptor cells. Selected bacterial colonies were inoculated into LB broth containing kanamycin and incubated at 37℃ with shaking at 220rpm until the culture reached an OD600 of 0.6-0.8. Protein expression was then induced under IPTG (0.5mM) condition at 18℃ with shaking at 180 rpm for 16-18h. The bacterial was collected by centrifugation and lysed by sonication in an ice-water bath, the supernatant was then collected and filtered through a 0.22μm membrane filter. The protein was purified by a Ni column and eluted through a gradient concentration of imidazole.
4.4. In Vivo Immune Strategies
For immunogenicity analysis, the vaccine plasmid or vector was intramuscularly injected into the tibialis anterior muscles of both hind limbs (50μg in each limb) of C57BL/6 mice (n=5) through electroporation on days 0, 2 and 5. The mice were euthanized one week after the last immunization.
For antitumor effect analysis, 2×106 MSLN-EGFP+ Panc02 cells were subcutaneously injected into the right hind flank of the C57BL/6 mice(n=12). In the first therapeutic experiment, a homologous prime-boost immunization strategy (3D+2D) was applied to enhance immune responses. Tumor inoculation was performed on day 0, DNA vaccines were administered intramuscularly on days 8, 10, 13, and days 22, 24. Mice were euthanized 7 days after the final immunization for sample collection and analysis. In the second therapeutic experiment, mice were challenged with tumor cells on day 0. DNA vaccines were administered intramuscularly on days 8, 10, 13. Gemcitabine was administered via intraperitoneal injections on days 8, 11, 15 and 18. Mice were euthanized 5 days after the final Gemcitabine injection (i.e., 10 days after the last DNA immunization) for subsequent analysis. Mice body weights and tumor weights were measured by electric scale. The tumor inhibition rate (TIR) was calculated by the following formula: TIR (%)=[(average tumor weight of negative control group-average tumor weight of treatment group)/average tumor weight of negative control group] ×100%. The tumor size was measured every two days with a vernier caliper and the tumor volume was calculated by the formula: V= (length×width2)/2 (mm3). According to ethics guidelines, mice were euthanized if tumor volume exceed 1500 mm3 or if the health status of mice were severely affected.
4.5. Preparation of Single-Cell Suspension
Spleens from the vaccinated mice were harvested, mechanically dissociated, and filtered. Red Blood cells were lysed by RBC Lysis buffer (BioLegend, San Diego, CA, USA). The resulting cell suspensions were washed and resuspended in R10 (RPMI-1640 containing 10% FBS) medium to obtain a single cell suspension at a concentration of 1×107 cells/ml.
Tumor tissues were excised and placed in 6-well plates, the tissues were thoroughly minced with sterile scissors and digested with 2ml of digestion buffer (e.g., collagenase and DNase I) at 37℃ in a CO2 incubator for 2h. The resulted tumor tissue suspensions were then processed following the same protocol as for splenocytes including dissociation, filtration, RBCs lysis and resuspension in R10 medium for downstream assays.
4.6. IFN-γ ELISpot Assays
IFN-γ ELISpot assays were performed by the ELISpot kit (BD Biosciences, Franklin Lakes, NJ, USA) according to the manufacturer’s instructions.
4.7. Cell Staining and Flow Cytometry
For extracellular staining of splenocytes or tumor cells, after 2×106 cells were suspended in 100μl cell staining buffer in addition of 1-2μl fluorophore-conjugated antibodies, the cells were then incubated on ice in the dark for 15 min.
Following extracellular staining, cells were fix and permeabilized according to the manufacturer’s instructions using a cell fixation/permeabilization kit (eBioscience, San Diego, CA, USA) for intracellular staining. The supernatant was discarded and the remaining cells were suspended in 100μl staining buffer and blocked with 1μl anti-mouse CD16/32 Antibody. After incubation at room temperature in the dark for 15 min, 1-2μl of fluorophore-conjugated antibody was added, followed by an additional 30 min incubation at room temperature in the dark
After staining, cells were centrifuged at 350×g for 5 min, and the supernatant was discarded. The cells were then washed twice with R10 medium and finally resuspended in an appropriate volume of R10 for flow cytometric analysis.
4.8. Quantitative Real-Time PCR (qRT-PCR)
Total RNA was extracted from tumor tissues with TRIzol reagent (Invitrogen), and mRNA was then reverse-transcribed into cDNA using the PrimeScript 1st Strand cDNA Synthesis kit (Takara, Japan). The relative expression of target gene mRNA was normalized to GAPDH and calculated using the 2-ΔΔCt method.
4.9. Statistical Analysis
All experiments were performed with at least two independent replicates. Statistical analysis and graph generation were conducted using GraphPad Prism 8 software. Data are presented as mean ± standard error. Unpaired t-tests were used for comparisons between two groups, while one-way ANOVA was applied for comparisons among multiple groups. Tumor growth curves were analyzed using two-way ANOVA. A p-value of less than 0.05 was considered statistically significant.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: The establishment and validation of MSLN stable-expressed pancreatic cancer mouse model.
Author Contributions
Conceptualization: H.W., J.W., C.W., X.Y., and H.Z.; Methodology: S.C.; Validation: S.C., G.B.; Investigation: S.C., G.B., Q.W., J.L., X.Z, and Z.H.; Data curation: S.C., G.B.; Writing—original draft preparation: S.C., G.B.; Writing—review and editing: H.Z.; Supervision: H.Z.; Project administration: H.Z.; Funding acquisition: H.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Science & Technology Development Plan of Jilin Province (20240402044GH).
Institutional Review Board Statement
All animal experiments were performed in accordance with Chinese law and were approved by the Ethics Committee of Jilin University. Approval number: YNPZSY2024056.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and/or analysis during the current study are available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rojas, L.A.; et al. Personalized RNA neoantigen vaccines stimulate T cells in pancreatic cancer. Nature 2023, 618(7963), 144-+. [Google Scholar] [CrossRef]
- Sung, H.; et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Ca-a Cancer Journal for Clinicians 2021, 71(3), 209–249. [Google Scholar] [CrossRef]
- Arab, A.; Yazdian-Robati, R.; Behravan, J. HER2-Positive Breast Cancer Immunotherapy: A Focus on Vaccine Development. Archivum Immunologiae Et Therapiae Experimentalis 2020, 68(1). [Google Scholar] [CrossRef]
- Kreiter, S.; et al. Increased antigen presentation efficiency by coupling antigens to MHC class I trafficking signals. Journal of Immunology 2008, 180(1), 309–318. [Google Scholar] [CrossRef]
- Yewdell, J.W. Not such a dismal science: the economics of protein synthesis, folding, degradation and antigen processing. Trends in Cell Biology 2001, 11(7), 294–297. [Google Scholar] [CrossRef]
- Yewdell, J.W.; Reits, E.; Neefjes, J. Making sense of mass destruction: Quantitating MHC class I antigen presentation. Nature Reviews Immunology 2003, 3(12), 952–961. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; et al. MHCI trafficking signal-based mRNA vaccines strengthening immune protection against RNA viruses. Bioengineering & Translational Medicine 2025, 10(1). [Google Scholar] [CrossRef]
- Lizée, G.; et al. Control of dendritic cell cross-presentation by the major histocompatibility complex class I cytoplasmic domain. Nature Immunology 2003, 4(11), 1065–1073. [Google Scholar] [CrossRef]
- Lizée, G.; Basha, G.; Jefferies, W.A. Tails of wonder: endocytic-sorting motifs key for exogenous antigen presentation. Trends Immunol 2005, 26(3), 141–9. [Google Scholar] [CrossRef]
- Blander, J.M. Different routes of MHC-I delivery to phagosomes and their consequences to CD8 T cell immunity. Seminars in Immunology 2023, 66. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; et al. MHC class I trafficking signal improves induction of cytotoxic T lymphocyte using artificial antigen presenting cells. Biochemistry and Biophysics Reports 2025, 41. [Google Scholar] [CrossRef]
- Basha, G.; et al. MHC Class I Endosomal and Lysosomal Trafficking Coincides with Exogenous Antigen Loading in Dendritic Cells. Plos One 2008, 3(9). [Google Scholar] [CrossRef]
- Dengjel, J.; et al. Autophagy promotes MHC class II presentation of peptides from intracellular source proteins. Proceedings of the National Academy of Sciences of the United States of America 2005, 102(22), 7922–7927. [Google Scholar] [CrossRef]
- Diethelm-Okita, B.M.; et al. Epitope repertoire of human CD4+ T cells on tetanus toxin: identification of immunodominant sequence segments. The Journal of infectious diseases 1997, 175(2), 382–91. [Google Scholar] [CrossRef] [PubMed]
- Raju, R.; et al. Epitope repertoire of human CD4+ lines propagated with tetanus toxoid or with synthetic tetanus toxin sequences. Journal of autoimmunity 1996, 9(1), 79–88. [Google Scholar] [CrossRef]
- Sahin, U.; et al. An RNA vaccine drives immunity in checkpoint-inhibitor-treated melanoma. Nature 2020, 585(7823), 107+. [Google Scholar] [CrossRef]
- Alexander, J.; et al. The optimization of helper T lymphocyte (HTL) function in vaccine development. Immunologic Research 1998, 18(2), 79–92. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; et al. Development of high potency universal DR-restricted helper epitopes by modification of high affinity DR-blocking peptides. Immunity 1994, 1(9), 751–61. [Google Scholar] [CrossRef] [PubMed]
- Saxena, M.; et al. Therapeutic cancer vaccines. Nature Reviews Cancer 2021, 21(6), 360–378. [Google Scholar] [CrossRef]
- Hassan, R.; Bera, T.; Pastan, I. Mesothelin: A new target for immunotherapy. Clinical Cancer Research 2004, 10(12), 3937–3942. [Google Scholar] [CrossRef]
- Hassan, R.; Ho, M. Mesothelin targeted cancer immunotherapy. European Journal of Cancer 2008, 44(1), 46–53. [Google Scholar] [CrossRef]
- Chang, M.C.; et al. Mesothelin-specific cell-based vaccine generates antigen-specific immunity and potent antitumor effects by combining with IL-12 immunomodulator. Gene Therapy 2016, 23(1), 38–49. [Google Scholar] [CrossRef]
- Chen, J.N.; et al. Anti-mesothelin CAR-T immunotherapy in patients with ovarian cancer. Cancer Immunology Immunotherapy 2023, 72(2), 409–425. [Google Scholar] [CrossRef]
- Wittwer, N.L.; et al. An anti-mesothelin targeting antibody drug conjugate induces pyroptosis and ignites antitumor immunity in mouse models of cancer. Journal for Immunotherapy of Cancer 2023, 11(3). [Google Scholar] [CrossRef]
- Hollingsworth, R.E.; Jansen, K. Turning the corner on therapeutic cancer vaccines. Npj Vaccines 2019. [Google Scholar] [CrossRef]
- Zahm, C.D.; Colluru, V.T.; McNeel, D.G. DNA vaccines for prostate cancer. Pharmacology & Therapeutics 2017, 174, 27–42. [Google Scholar] [CrossRef]
- Hu, J.; et al. MSLN induced EMT, cancer stem cell traits and chemotherapy resistance of pancreatic cancer cells. Heliyon 2024, 10(8). [Google Scholar] [CrossRef]
- Zheng, C.; et al. Mesothelin regulates growth and apoptosis in pancreatic cancer cells through p53-dependent and -independent signal pathway. Journal of Experimental & Clinical Cancer Research 2012, 31. [Google Scholar] [CrossRef]
- Fu, C.; Jiang, A. Dendritic Cells and CD8 T Cell Immunity in Tumor Microenvironment. Frontiers in Immunology 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; et al. The XCL1-Mediated DNA Vaccine Targeting Type 1 Conventional Dendritic Cells Combined with Gemcitabine and Anti-PD1 Antibody Induces Potent Antitumor Immunity in a Mouse Lung Cancer Model. International Journal of Molecular Sciences 2024, 25(3). [Google Scholar] [CrossRef] [PubMed]
- Fabre, M.; et al. OMTX705, a Novel FAP-Targeting ADC Demonstrates Activity in Chemotherapy and Pembrolizumab-Resistant Solid Tumor Models. Clinical Cancer Research 2020, 26(13), 3420–3430. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.M.; et al. A phase I dose-escalation study of sibrotuzumab in patients with advanced or metastatic fibroblast activation protein-positive cancer. Clinical Cancer Research 2003, 9(5), 1639–1647. [Google Scholar]
- Liu, C.L.; et al. Fast DNA Vaccination Strategy Elicits a Stronger Immune Response Dependent on CD8+CD11c+ Cell Accumulation. Frontiers in Oncology 2021, 11. [Google Scholar] [CrossRef]
- Hardwick, N.R.; et al. p53-Reactive T Cells Are Associated with Clinical Benefit in Patients with Platinum-Resistant Epithelial Ovarian Cancer After Treatment with a p53 Vaccine and Gemcitabine Chemotherapy. Clinical Cancer Research 2018, 24(6), 1315–1325. [Google Scholar] [CrossRef]
- Natu, J.; Nagaraju, G.P. Gemcitabine effects on tumor microenvironment of pancreatic ductal adenocarcinoma: Special focus on resistance mechanisms and metronomic therapies. Cancer Letters 2023, 573. [Google Scholar] [CrossRef]
- Hagerty, B.L.; Takabe, K. Biology of Mesothelin and Clinical Implications: A Review of Existing Literature. World Journal of Oncology 2023, 14(5), 340–349. [Google Scholar] [CrossRef]
- Avula, L.R.; et al. Mesothelin Enhances Tumor Vascularity in Newly Forming Pancreatic Peritoneal Metastases. Molecular Cancer Research 2020, 18(2), 229–239. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, F.K.; et al. DNA fusion vaccines enter the clinic. Cancer Immunology Immunotherapy 2011, 60(8), 1147–1151. [Google Scholar] [CrossRef]
- Neeli, P.; et al. DNA vaccines against GPRC5D synergize with PD-1 blockade to treat multiple myeloma. Npj Vaccines 2024, 9(1). [Google Scholar] [CrossRef]
- Ahrends, T.; et al. CD4+ T Cell Help Confers a Cytotoxic T Cell Effector Program Including Coinhibitory Receptor Downregulation and Increased Tissue Invasiveness. Immunity 2017, 47(5), 848+. [Google Scholar] [CrossRef] [PubMed]
- Jiao, S.P.; et al. Differences in Tumor Microenvironment Dictate T Helper Lineage Polarization and Response to Immune Checkpoint Therapy. Cell 2019, 179(5), 1177+. [Google Scholar] [CrossRef]
- Wilson, N.S.; El-Sukkari, D.; Villadangos, J.A. Dendritic cells constitutively present self antigens in their immature state in vivo and regulate antigen presentation by controlling the rates of MHC class II synthesis and endocytosis. Blood 2004, 103(6), 2187–2195. [Google Scholar] [CrossRef]
- Roche, P.A.; Furuta, K. The ins and outs of MHC class II-mediated antigen processing and presentation. Nature Reviews Immunology 2015, 15(4), 203–216. [Google Scholar] [CrossRef] [PubMed]
- Le, H.K.; et al. Gemcitabine directly inhibits myeloid derived suppressor cells in BALB/c mice bearing 4T1 mammary carcinoma and augments expansion of T cells from tumor-bearing mice. International Immunopharmacology 2009, 9(7-8), 900–909. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, E.; et al. Gemcitabine reduces MDSCs, tregs and TGFβ-1 while restoring the teff/treg ratio in patients with pancreatic cancer. Journal of Translational Medicine 2016, 14. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, I.; et al. Low-dose gemcitabine depletes regulatory T cells and improves survival in the orthotopic Panc02 model of pancreatic cancer. International Journal of Cancer 2013, 133(1), 98–107. [Google Scholar] [CrossRef]
Figure 1.
Construction and Validation of DNA tumor vaccines. (A) Schematic diagrams of the DNA plasmids used in this study. (B) The DNA plasmids were digested with Pst I/BamH I, and was identified by Agarose gel electrophoresis. (C) 293T cells was transfected with the DNA plasmids respectively, and the expression of the fusion proteins was verified by Western Blotting, using anti-MSLN antibody as the primary antibody and HRP-conjugated Goat IgG as secondary antibody.
Figure 1.
Construction and Validation of DNA tumor vaccines. (A) Schematic diagrams of the DNA plasmids used in this study. (B) The DNA plasmids were digested with Pst I/BamH I, and was identified by Agarose gel electrophoresis. (C) 293T cells was transfected with the DNA plasmids respectively, and the expression of the fusion proteins was verified by Western Blotting, using anti-MSLN antibody as the primary antibody and HRP-conjugated Goat IgG as secondary antibody.
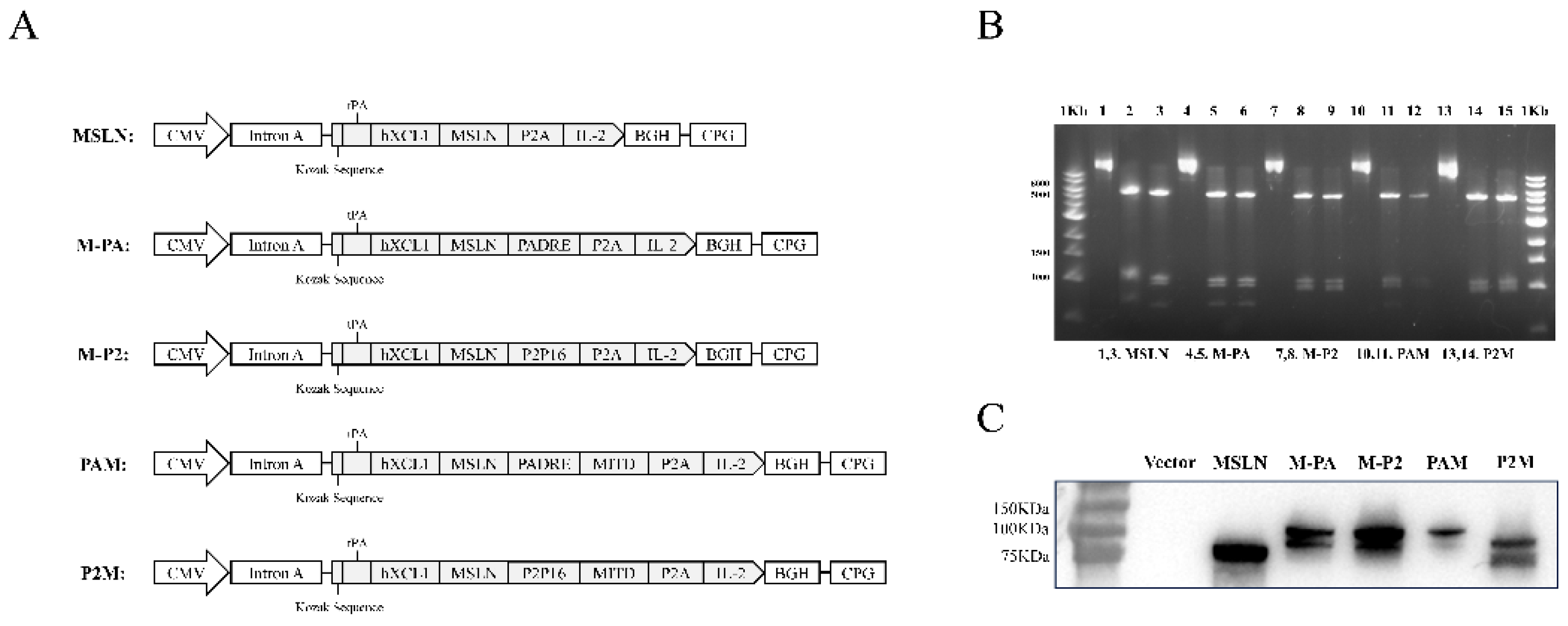
Figure 2.
Immunogenicity analysis of DNA vaccines. (A, B) Representative images of ELISpot and quantification of IFN-γ FSC in different immune groups after stimulation. MSLN Pro was utilized as specific stimulator. (C) Levels of MSLN-specific binding antibodies in the sera of mice were measured by ELISA. (D-G) The proportion of CD4+ T cells Activation (D), CD8+ T cells Activation (E), Proliferative CD4+ T cells (F), Proliferative CD8+ T cells (CD8+Ki67+) (G) in splenocytes were measured by flow cytometry. * P<0.05, ** P<0.01, *** P<0.001, **** P<0.0001, and ns: no significance.
Figure 2.
Immunogenicity analysis of DNA vaccines. (A, B) Representative images of ELISpot and quantification of IFN-γ FSC in different immune groups after stimulation. MSLN Pro was utilized as specific stimulator. (C) Levels of MSLN-specific binding antibodies in the sera of mice were measured by ELISA. (D-G) The proportion of CD4+ T cells Activation (D), CD8+ T cells Activation (E), Proliferative CD4+ T cells (F), Proliferative CD8+ T cells (CD8+Ki67+) (G) in splenocytes were measured by flow cytometry. * P<0.05, ** P<0.01, *** P<0.001, **** P<0.0001, and ns: no significance.
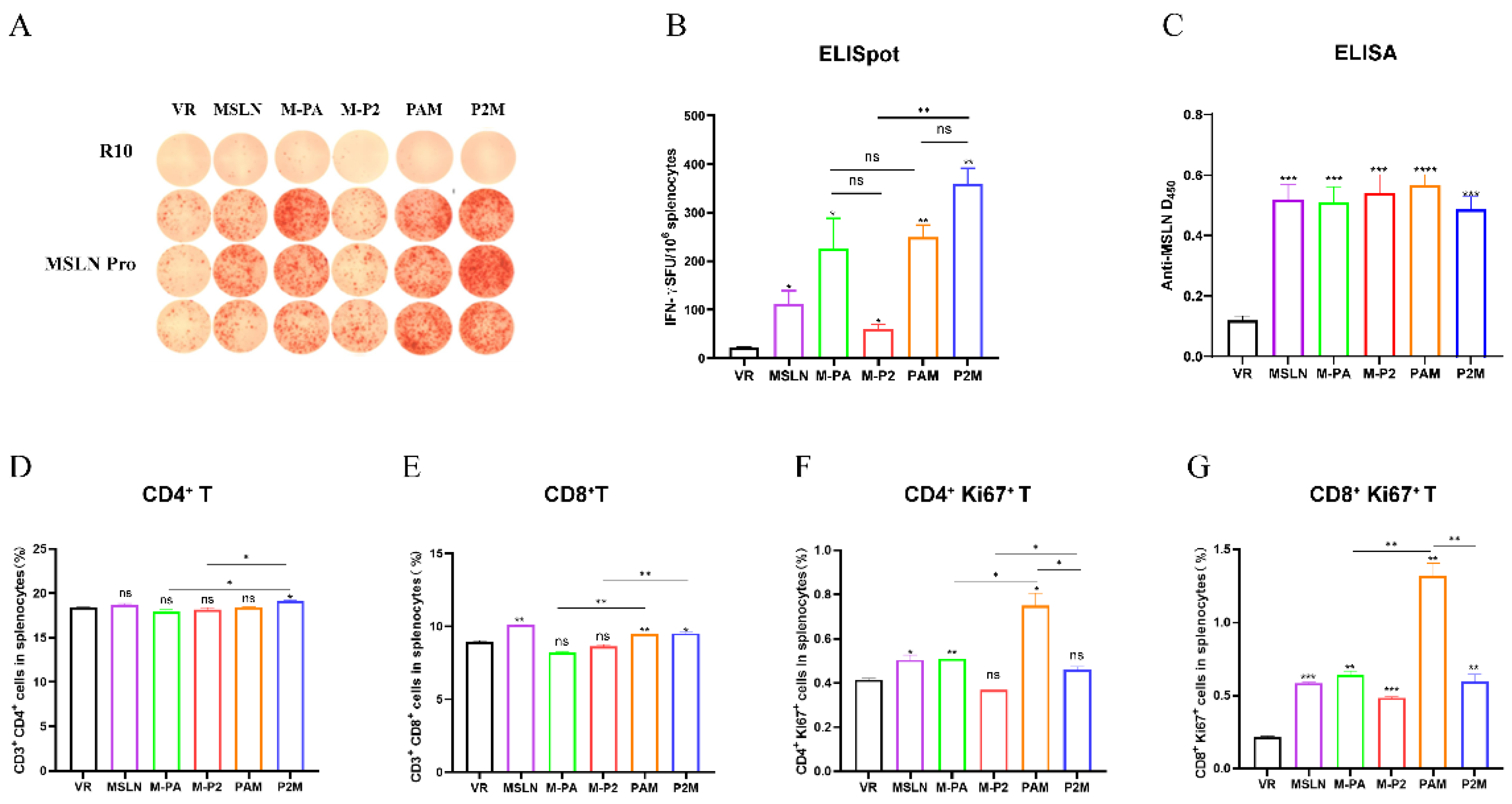
Figure 3.
Antitumor effect of the DNA vaccines and detection of related cellular immune responses. (A) Schematic diagram of the DNA vaccination strategy. C57BL/6 mice (n=8-12) were inoculated with 2×106 MSLN-EGFP+ Panc02 cells at day 0, and received treatment on day 8, 10, 13, 22 and day 24, the mice were euthanized on day 30 after tumor challenge. (B) Tumor growth curve was measured every two days. (C) Tumor weight analysis in different immunized groups. (D-E) Representative images of ELISpot and quantification of IFN-γ FSC in different immune groups after stimulation. MSLN Pro was utilized as specific stimulator. (F) The Proportion of CD3+CD4+ T cells and CD3+CD8+ T cells in splenocytes. (G-H) Proportion of tumor infiltrating lymphocytes, CD45+CD3+CD4+ T cells, CD45+CD3+CD8+ T cells (G) and Proliferative CD4+ T cells (CD4+Ki67+), Proliferative CD8+ T cells (CD8+Ki67+) (H) were measure by flow cytometry. (I-J) The expression levels of Gzmb (I), and Th1 cytokine (IFN-γ, IL-2, TNF-α) (J) was measure by qRT-PCR. * P<0.05, ** P<0.01, *** P<0.001, **** P<0.0001, and ns: no significance.
Figure 3.
Antitumor effect of the DNA vaccines and detection of related cellular immune responses. (A) Schematic diagram of the DNA vaccination strategy. C57BL/6 mice (n=8-12) were inoculated with 2×106 MSLN-EGFP+ Panc02 cells at day 0, and received treatment on day 8, 10, 13, 22 and day 24, the mice were euthanized on day 30 after tumor challenge. (B) Tumor growth curve was measured every two days. (C) Tumor weight analysis in different immunized groups. (D-E) Representative images of ELISpot and quantification of IFN-γ FSC in different immune groups after stimulation. MSLN Pro was utilized as specific stimulator. (F) The Proportion of CD3+CD4+ T cells and CD3+CD8+ T cells in splenocytes. (G-H) Proportion of tumor infiltrating lymphocytes, CD45+CD3+CD4+ T cells, CD45+CD3+CD8+ T cells (G) and Proliferative CD4+ T cells (CD4+Ki67+), Proliferative CD8+ T cells (CD8+Ki67+) (H) were measure by flow cytometry. (I-J) The expression levels of Gzmb (I), and Th1 cytokine (IFN-γ, IL-2, TNF-α) (J) was measure by qRT-PCR. * P<0.05, ** P<0.01, *** P<0.001, **** P<0.0001, and ns: no significance.
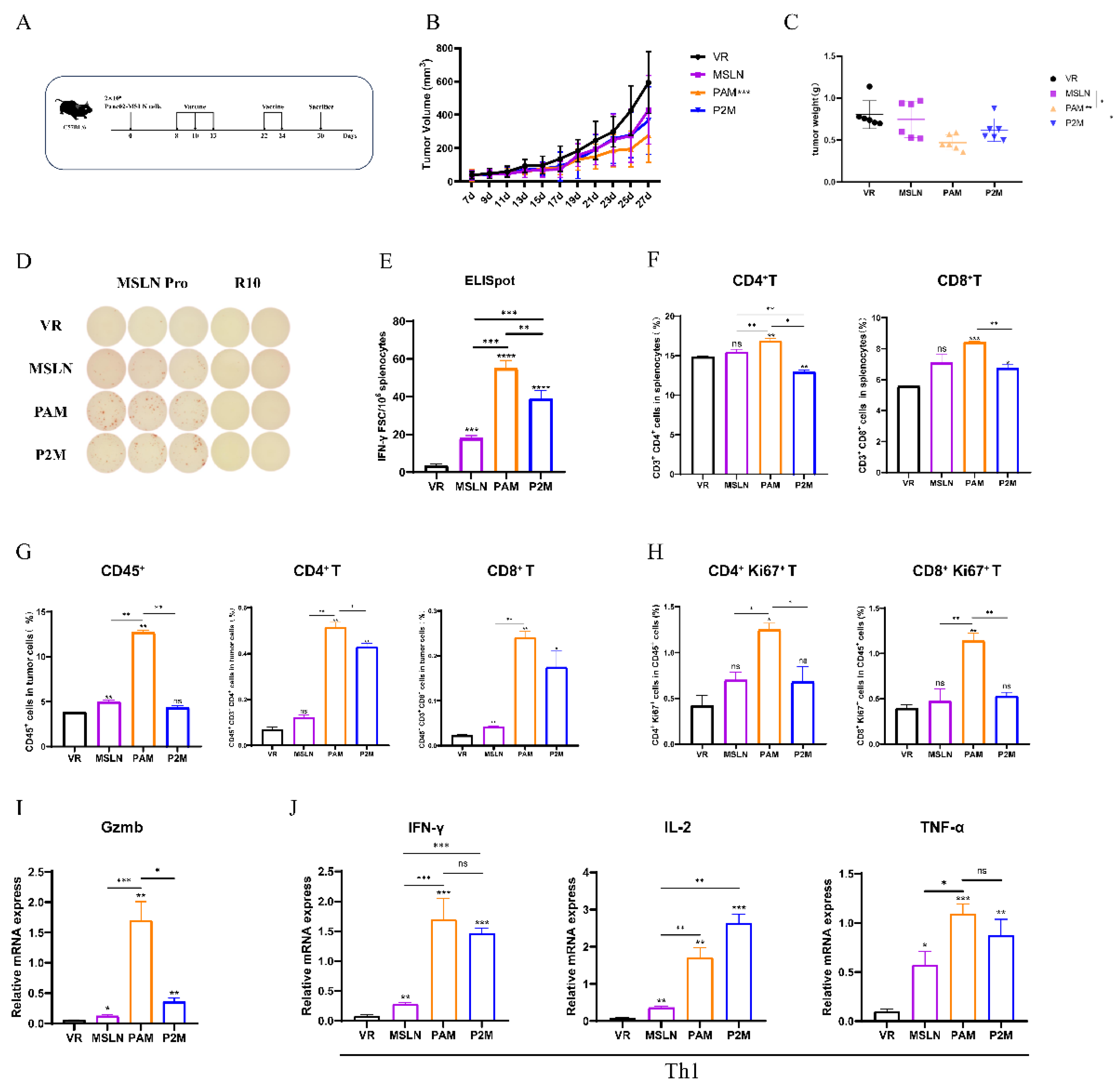
Figure 4.
Analysis of intratumoral immunosuppression in immunized tumor-bearing mice. (A-D) The proportion of tumor infiltrated MDSCs (A), Tregs (B), TAMs-M1 (C), TAMs-M2 (D) were measured by flow cytometry. (E) The ratio of M1/M2. (F) The expression levels of Th2 cytokines (IL-4, IL-6, IL-10) were measured by qRT-PCR. * P<0.05, ** P<0.01, *** P<0.001, and ns: no significance.
Figure 4.
Analysis of intratumoral immunosuppression in immunized tumor-bearing mice. (A-D) The proportion of tumor infiltrated MDSCs (A), Tregs (B), TAMs-M1 (C), TAMs-M2 (D) were measured by flow cytometry. (E) The ratio of M1/M2. (F) The expression levels of Th2 cytokines (IL-4, IL-6, IL-10) were measured by qRT-PCR. * P<0.05, ** P<0.01, *** P<0.001, and ns: no significance.
Figure 5.
PAM combined with Gem further enhances antitumor effect. (A) Schematic diagram of DNA vaccine combined with Gem immunization strategy. C57BL/6 mice (n=8-12) was inoculated with 2×106 MSLN-EGFP+Panc02 cells at day 0, and received DNA vaccine treatment on day 8, 10, 13, Gem treatment on day 8, 11, 15, 18. (B) Tumor growth and weight were measured in different immunized groups. (C) Body weight change curves were measured every two days. (D) Representative images of ELISpot and quantification of IFN-γ FSC in different immune groups after stimulation. MSLN Pro was utilized as specific stimulator. (E-G) The Proportion of CD3+CD4+ T cells in splenocytes (E), the proportion of tumor infiltrating lymphocytes, CD45+CD3+CD4+ T cells, CD45+CD3+CD8+ T cells (F) and Proliferative CD4+ T cells (CD4+Ki67+), Proliferative CD8+ T cells (CD8+Ki67+) (G) were measure by flow cytometry. (H) The proportion of tumor infiltrated NKs and DCs were measure by flow cytometry. * P<0.05, ** P<0.01, *** P<0.001, **** P<0.0001, and ns: no significance.
Figure 5.
PAM combined with Gem further enhances antitumor effect. (A) Schematic diagram of DNA vaccine combined with Gem immunization strategy. C57BL/6 mice (n=8-12) was inoculated with 2×106 MSLN-EGFP+Panc02 cells at day 0, and received DNA vaccine treatment on day 8, 10, 13, Gem treatment on day 8, 11, 15, 18. (B) Tumor growth and weight were measured in different immunized groups. (C) Body weight change curves were measured every two days. (D) Representative images of ELISpot and quantification of IFN-γ FSC in different immune groups after stimulation. MSLN Pro was utilized as specific stimulator. (E-G) The Proportion of CD3+CD4+ T cells in splenocytes (E), the proportion of tumor infiltrating lymphocytes, CD45+CD3+CD4+ T cells, CD45+CD3+CD8+ T cells (F) and Proliferative CD4+ T cells (CD4+Ki67+), Proliferative CD8+ T cells (CD8+Ki67+) (G) were measure by flow cytometry. (H) The proportion of tumor infiltrated NKs and DCs were measure by flow cytometry. * P<0.05, ** P<0.01, *** P<0.001, **** P<0.0001, and ns: no significance.
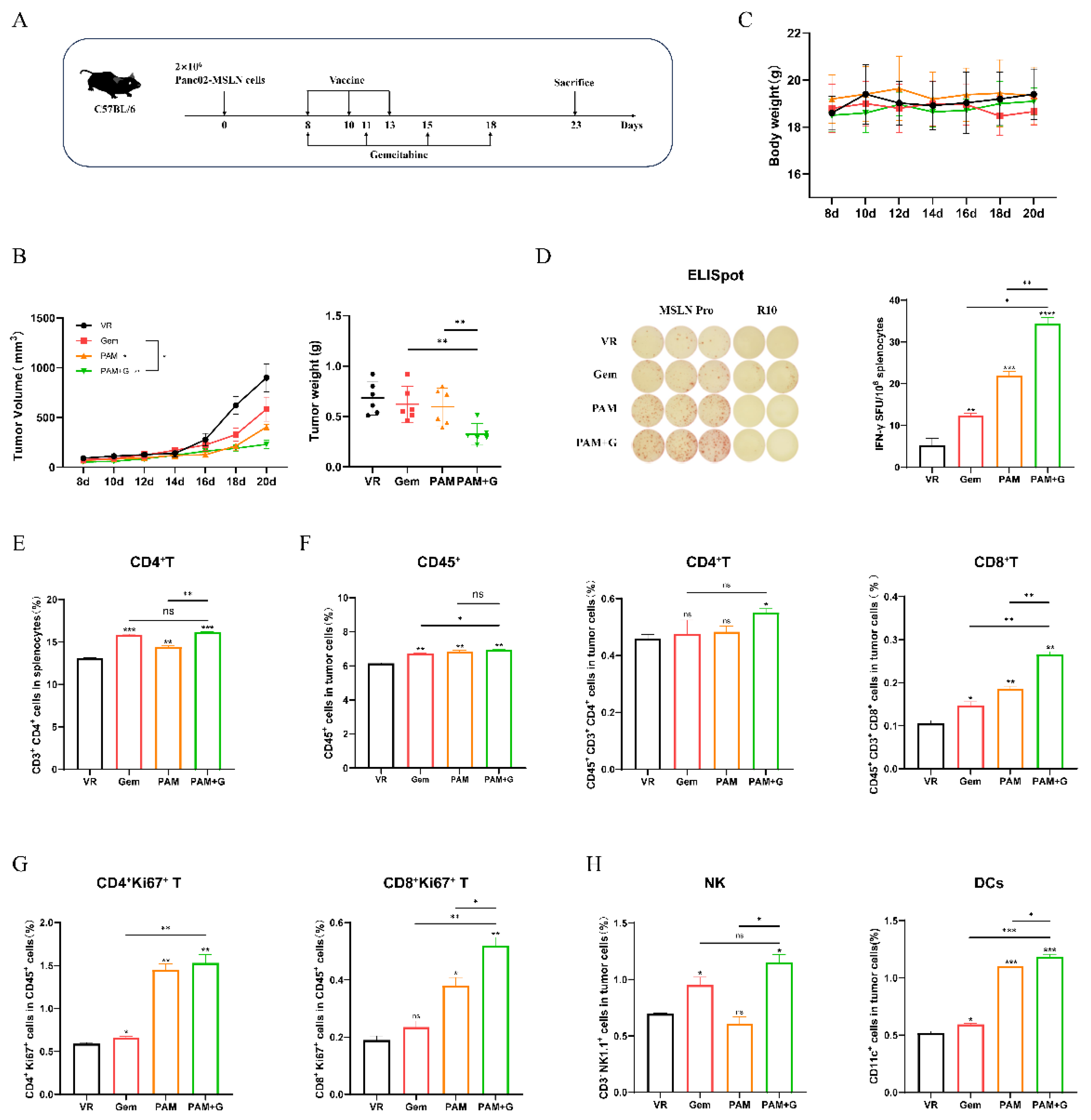
Figure 6.
Impact of Gem combination therapy on intratumoral immunosuppression. (A-D) The proportion of tumor infiltrated MDSCs (A), Tregs (B), TAMs-M1 (C), TAMs-M2 (D) were measured by flow cytometry. (E) The ratio of M1/M2. * P<0.05, ** P<0.01, *** P<0.001, and ns: no significance.
Figure 6.
Impact of Gem combination therapy on intratumoral immunosuppression. (A-D) The proportion of tumor infiltrated MDSCs (A), Tregs (B), TAMs-M1 (C), TAMs-M2 (D) were measured by flow cytometry. (E) The ratio of M1/M2. * P<0.05, ** P<0.01, *** P<0.001, and ns: no significance.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



