Submitted:
12 January 2026
Posted:
13 January 2026
You are already at the latest version
Abstract
The double-slit experiment, a cornerstone of quantum mechanics, is traditionally viewed as a paradoxical demonstration of wave-particle duality. This article posits that its core dynamic—superposition, interference, and environment-driven localization—is not a unique quantum phenomenon but a fundamental computational principle implemented by the brain. We introduce the Ze framework, arguing that the brain operates as a biological interferometer. Cognitive systems maintain multiple generative hypotheses in a state of active interference (superposition), analogous to the quantum wavefunction passing through both slits. "Which-path" information, supplied by sensory data, action, and social context, forces cognitive decoherence, localizing perception and decision into a single narrative. Sleep is recast as an intrinsic quantum eraser, periodically degrading which-path information to restore cognitive flexibility and prevent pathological hyper-localization. The framework structurally links quantum decoherence, Bayesian active inference, and the neurobiology of sleep and wake cycles. It provides a transdiagnostic model for psychopathology, where disorders like psychosis and PTSD are seen as dysregulations of this interference-localization cycle. We conclude that the brain does not observe quantum reality; it actively instantiates its core logic, making the double-slit experiment a continuous, lived process of resolving ambiguity to survive and understand the world.
Keywords:
active inference
; cognitive decoherence
; double-slit analogy
; free energy principle
; predictive processing
; sleep
; transdiagnostic psychiatry
1. The Central Thesis: From Quantum Paradox to Cognitive Principle
The quantum double-slit experiment is often presented as a mysterious phenomenon exclusive to the microscopic world. This paper argues that the core mechanism of this experiment is not a quantum curiosity but a fundamental principle of adaptive systems engaged in active inference (Friston, 2010). We posit that the adult human brain operates continuously in this double-slit mode. It perpetually constructs multiple, often incompatible, interpretations of sensory data (Hohwy, 2016), minimizes a quantity formally analogous to free energy (Friston & Kiebel, 2009), and is forced by environmental demands to either maintain a superposition of hypotheses or localize onto a single, actionable narrative.
This framework, termed the Ze framework, proposes that cognition is a dynamic interference pattern between competing generative models. The “which-path” information that destroys quantum interference finds its cognitive analogue in sensory binding, linguistic framing, goal-directed action, and social feedback—all of which force a path localization (Clark, 2013). Consequently, phenomena like sleep, insight, and psychopathology can be reinterpreted through the physics of interference and decoherence, suggesting a profound structural isomorphism between the resolution of uncertainty in quantum systems and in the brain (Bruza, Wang, & Busemeyer, 2015).
2. Interference as a Cognitive State
In the Ze framework, cognitive interference corresponds to the simultaneous, active maintenance of multiple incompatible hypotheses about the world (Friston & Kiebel, 2009). This is not merely parallel processing; it is the maintenance of a probabilistic superposition within the brain’s generative model (Knill & Pouget, 2004). Conversely, cognitive localization is the process by which this superposition collapses into a single, stabilized interpretation.
- States Permitting Interference: Sleep, imagination, and insight are characterized by a heightened tolerance for interference. During sleep, sensory gating reduces “which-path” information, allowing for the hyper-associative recombination of memory traces (Lewis, Knoblich, & Poe, 2018; Tononi & Cirelli, 2014). Imagination and mind-wandering, supported by the default mode network, involve holding present reality and counterfactual possibilities in mind simultaneously (Buckner & Carroll, 2007; Raichle, 2015). Insight involves a relaxation of top-down constraints, allowing interference between remote concepts before a sudden, new solution localizes (Jung-Beeman et al., 2004).
- States Suppressing Interference: Effective waking action demands the suppression of interference. Goal-directed wakefulness requires localization onto a single model of the world for sensorimotor control (Milner & Goodale, 2008; Cisek & Kalaska, 2010). Focused attention acts as a cognitive “which-path” detector, selectively amplifying sensory evidence for one hypothesis and forcing rapid localization (Feldman & Friston, 2010; Meng & Tong, 2004). Neuromodulators like norepinephrine sharpen this competition (Aston-Jones & Cohen, 2005).

Table 1.
Cognitive and Quantum States of Interference and Localization.
| Cognitive Phenomenon (Ze Framework) | Quantum Analogue (Double-Slit) | Primary Neural/Physiological Correlate |
|---|---|---|
| Cognitive Interference (Ambiguity, contemplation, dreaming) | Wave-like behavior (Superposition through both slits, interference pattern) | Co-activation of competing neural assemblies; Default Mode Network activity; High entropy states. |
| “Which-Path” Information (Sensory fixation, action commitment, social feedback) | Path measurement (Detector at slit, recording which path is taken) | Increased precision-weighting of prediction errors; Norepinephrine-mediated gain; Gamma-band synchronization for binding. |
| Cognitive Localization (Perceptual decision, categorical choice, narrative stabilization) | Particle-like behavior (Collapse to a single localized point on detector) | Winner-take-all inhibitory competition; Synchronization of winning neural coalition; Suppression of rival representations. |
| Sleep as Cognitive Eraser (Synaptic downscaling, memory recombination) | Quantum eraser experiment (Retroactive erasure of path information restores interference) | Thalamic sensory gating; Reduced noradrenergic tone; Slow-wave oscillations (SWS); Theta-gamma coupling in REM. |
3. “Which-Path” Information and Cognitive Decoherence
In quantum mechanics, acquiring “which-path” information destroys interference. An isomorphic process of cognitive decoherence is fundamental to brain function. Here, the brain’s internal models constitute the “system,” and the sensorimotor stream provides the “environment” (Friston & Kiebel, 2009). As evidence accrues for a specific interpretation, the probability distribution over causes sharpens, suppressing alternatives—a cognitive analogue of decoherence (Bruza et al., 2015). This “measurement” is inherent to the brain’s active engagement with the world to minimize free energy (Hohwy, 2016).
Which-path information is conveyed through several channels:
- Sensory Fixation & Binding: Focused perception provides high-precision data, anchoring interpretation. Gamma-band synchronization may mediate this binding (Engel & Singer, 2001).
- Linguistic Labeling: Attaching a verbal label commits the system to a categorical schema, suppressing alternatives (Herz & von Clef, 2001).
- Social Feedback: Confirmation or disagreement from others provides direct Bayesian evidence, often overriding personal interpretations (Zaki, Schirmer, & Mitchell, 2011).
- Goal-Directed Action: A motor commitment is the ultimate which-path measurement. The proprioceptive feedback uniquely validates the associated generative model (Cisek & Kalaska, 2010; Friston et al., 2016).
The accumulation of convergent which-path information exponentially accelerates localization by creating an unsustainable conflict (ΔF) between models, mediated by neuromodulatory systems like the locus coeruleus-norepinephrine system (Aston-Jones & Cohen, 2005; Feldman & Friston, 2010).
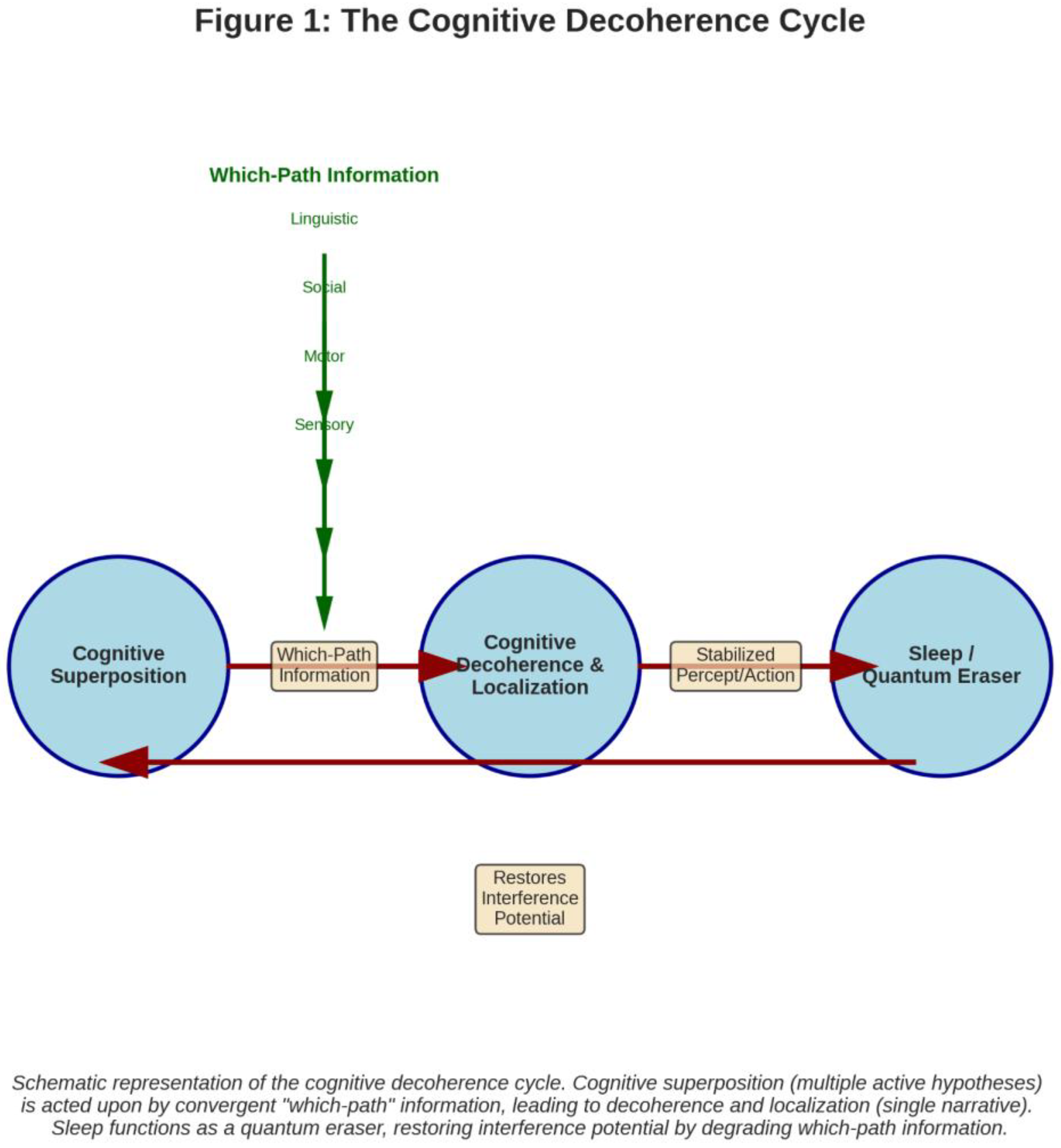
Figure 1.
The Cognitive Decoherence Cycle (A schematic diagram showing a “Cognitive Superposition” state (multiple active hypotheses) being acted upon by convergent “Which-Path Information” (sensory, motor, social, linguistic arrows). This leads to “Cognitive Decoherence & Localization” (a single selected narrative). A “Sleep/Quantum Eraser” arrow loops back from localization to superposition, showing the reset function.).
Figure 1.
The Cognitive Decoherence Cycle (A schematic diagram showing a “Cognitive Superposition” state (multiple active hypotheses) being acted upon by convergent “Which-Path Information” (sensory, motor, social, linguistic arrows). This leads to “Cognitive Decoherence & Localization” (a single selected narrative). A “Sleep/Quantum Eraser” arrow loops back from localization to superposition, showing the reset function.).
4. Sleep as a Cognitive Quantum Eraser
The delayed-choice quantum eraser shows that erasing which-path information can restore interference (Walborn et al., 2002). Sleep performs a precisely analogous function (Hobson & Friston, 2012). It actively degrades the precision-weighted “which-path record” etched into synaptic weights during wakefulness (Yang et al., 2014), acting as a cognitive quantum eraser.
The mechanism involves:
- Sensory Disconnection: Thalamic gating attenuates external evidence (McCormick & Bal, 1997).
- Neuromodulatory Reversal: Norepinephrine and serotonin drop during SWS, lowering precision-weighting. Cholinergic dominance in REM promotes hyper-association (Pace-Schott & Hobson, 2002; Hobson & Friston, 2012).
This flattens the free energy landscape, reducing the ΔF between models via synaptic downscaling (Tononi & Cirelli, 2006), and restores conditions for interference.
- Slow-Wave Oscillations (SWS): Orchestrate the reactivation and recombination of memory traces, not simple replay (Diekelmann & Born, 2010; Lewis & Durrant, 2011).
- REM Sleep (Theta-Gamma Coupling): Creates an ideal environment for associative linking of memories, emotions, and concepts (Walker & van der Helm, 2009).
Thus, sleep is a necessity for de-localization, preventing cognitive rigidity and enabling memory flexibility and creativity (Lewis et al., 2018).
5. Two Generative Models: The Ze Duality
The Ze framework proposes the brain is organized around two core, competing generative models whose interaction produces cognitive interference.
- Model A (Forward, Sensorimotor): Pragmatic, causal, and prospective. It predicts sensory consequences of actions to minimize surprise through movement (Friston et al., 2016). Associated with dorsal visual streams, frontoparietal networks, and the cerebellum (Milner & Goodale, 2008; Wolpert, Miall, & Kawato, 1998). It demands localization for action and dominates during focused tasks (Aston-Jones & Cohen, 2005).
- Model B (Inverse, Reconstructive): Reflective, diagnostic, and often retrospective. It infers the causes of data to build coherent narratives about the past, others’ minds, and counterfactuals (Hassabis & Maguire, 2009). Associated with the Default Mode Network (DMN), ventral visual stream, and hippocampus (Buckner & Carroll, 2007; Raichle, 2015). It tolerates ambiguity and parallel interpretations.
Interference occurs at their interface when data is ambiguous enough for both models (e.g., an ambiguous social cue). Localization is forced either by Model A’s imperative for action or Model B’s triumph of a more coherent narrative (e.g., an “Aha!” moment) (Jung-Beeman et al., 2004). The anterior cingulate cortex monitors this conflict, and the dorsolateral prefrontal cortex implements control (Botvinick, Cohen, & Carter, 2004).
Figure 2.
The Ze Duality and the Interference-Localization Cycle (A diagram showing Model A and Model B receiving Ambiguous Sensory Input. Their outputs create a “Zone of Cognitive Interference” (superposition). Arrows from “Which-Path Information” and “Imperative for Action” apply pressure, leading to a “Point of Localization” where one model’s output is selected, resulting in a “Stabilized Percept/Action.” A feedback loop labeled “Sleep/Erasure” resets the system back to the interference zone.).
Figure 2.
The Ze Duality and the Interference-Localization Cycle (A diagram showing Model A and Model B receiving Ambiguous Sensory Input. Their outputs create a “Zone of Cognitive Interference” (superposition). Arrows from “Which-Path Information” and “Imperative for Action” apply pressure, leading to a “Point of Localization” where one model’s output is selected, resulting in a “Stabilized Percept/Action.” A feedback loop labeled “Sleep/Erasure” resets the system back to the interference zone.).
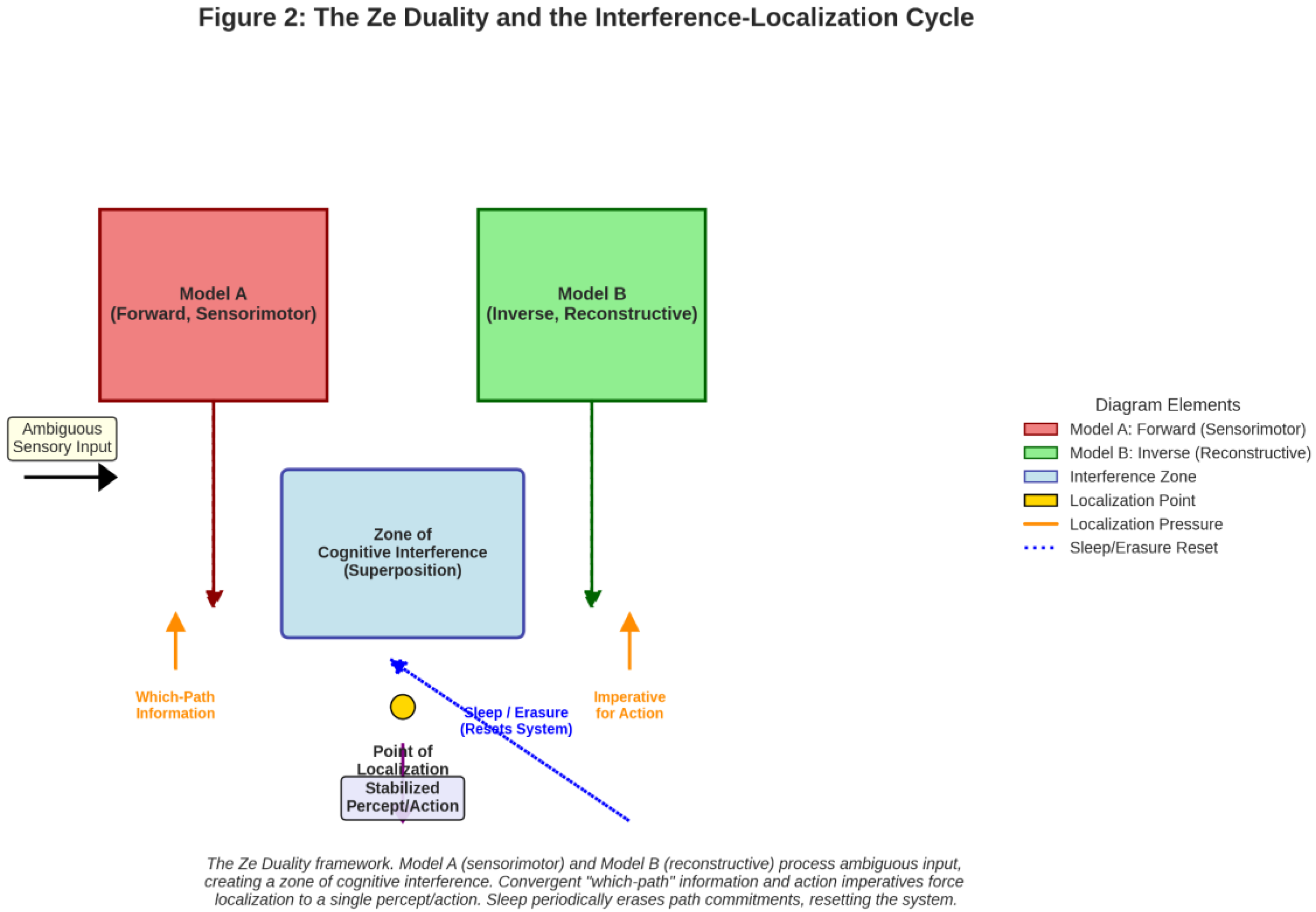
Table 2.
The Ze Duality: Two Generative Models of the Brain.
| Characteristic | Model A: Forward (Sensorimotor) | Model B: Inverse (Reconstructive) |
|---|---|---|
| Core Function | Predict consequences of action; minimize prediction error through movement. | Infer causes of sensory data; construct explanatory narratives. |
| Temporal Focus | Prospective (“What will happen if I do that?”) | Retrospective/Counterfactual (“What caused this? What if?”) |
| Primary Demand | Localization. Requires a single, unambiguous model for effective action. | Tolerates Interference. Can hold multiple interpretations in parallel. |
| Neuroanatomical Correlates | Dorsal visual stream, frontoparietal action networks, cerebellum, basal ganglia. | Default Mode Network (mPFC, PCC, angular gyrus), ventral visual stream, hippocampus. |
| Dominant States | Focused task engagement, threat response, skilled performance. | Mind-wandering, reminiscence, social reasoning, creative brainstorming. |
| Dysfunctional Extremes | Perseveration/Compulsion: Pathological, rigid action loops (e.g., OCD rituals). | Psychosis/Dissociation: Uncontrolled narrative generation detached from sensory evidence. |
6. Localization as a Forced Process
Cognitive localization is not a conscious choice but a forced, subpersonal computational outcome (Friston, 2010), analogous to quantum decoherence (Zurek, 2003). Perceptual resolution often occurs pre-consciously (Dehaene et al., 2006), and the “Aha!” moment feels involuntary (Jung-Beeman et al., 2004).
Localization is forced when:
- The free energy difference (ΔF) between models exceeds a stability threshold, creating an unsustainable gradient (Friston & Kiebel, 2009).
- The environment provides unambiguous sensory support, selectively lowering the free energy of one model (Feldman & Friston, 2010).
- The imperative for action generates proprioceptive predictions that can only be fulfilled by one model, forcing a collapse to avoid catastrophic prediction error (Cisek & Kalaska, 2010; Friston et al., 2016).
The feeling of conscious will is a post-hoc narrative of this forced transition (Haggard, 2008).
7. Structural Isomorphism: Molecules and the Brain
The similarity is not metaphorical but a structural isomorphism. Large molecules in interference experiments possess internal degrees of freedom (iDOFs) that can entangle with their path, causing self-decoherence (Arndt et al., 1999; Zurek, 2003). Similarly, the brain’s recursive, hierarchical architecture means every part is both processor and environment for others, creating a rich internal medium for which-path information (Friston, 2010; Hohwy, 2016).
Both systems exhibit self-decoherence: molecules via thermal vibration (Hackermüller et al., 2004); the brain via active neuromodulatory gain control to accelerate localization (Aston-Jones & Cohen, 2005). The brain achieves operational closure through the perception-action cycle, becoming its own primary environment (Clark, 2013; Friston et al., 2016).
The unifying principle: any complex, self-interacting system inferring hidden states will exhibit a trade-off between maintaining superposition (interference) and committing to one state (localization) to minimize free energy (Friston & Kiebel, 2009).
8. Psychopathology: Dysregulation of the Which-Path/Eraser Cycle
Mental disorders can be reformulated as dysregulations of the interference-localization cycle (Friston, Stephan, Montague, & Dolan, 2014).
-
Excessive Interference (Failed Which-Path Generation):
- ○
- Psychosis: Abnormally weak precision on sensory evidence (failing which-path information) allows Model B’s narratives to operate in uncontrolled interference, leading to hallucinations and delusions (Sterzer et al., 2018; Corlett et al., 2019; Fletcher & Frith, 2009).
- ○
- Dissociation: A failure to integrate which-path information into a coherent self-model, leading to fragmented consciousness (Lanius, Vermetten, & Pain, 2010).
-
Excessive Localization (Failed Erasure):
- ○
- PTSD: A traumatic memory forms an ultra-strong which-path record. Failed sleep-dependent erasure/integration leaves it hyper-localized and intrusive (Brewin, 2015; Tononi & Cirelli, 2014).
- ○
- Depression: Hyper-localization onto a negative narrative (Model B), compounded by poor sleep (reduced erasure), creates cognitive rigidity (Roiser, Elliott, & Sahakian, 2012; Riemann, Krone, Wulff, & Nissen, 2020).
- ○
- OCD: An intrusive thought (interference) is met with a compulsive action—a maladaptive, self-generated which-path measurement to force temporary, fragile localization (Robbins, Vaghi, & Banca, 2019).
Blurred sleep-wake boundaries in these disorders are a direct symptom of this core dysregulation (Muto et al., 2016).
9. Altered States of Consciousness as Ze Regimes
- Coma: A global suppression of both models, halting active inference. The apparatus is powered down (Laureys, 2005; Alkire, Hudetz, & Tononi, 2008).
- General Anesthesia: Induces a widespread, artificial localization without interpretation. It disrupts network integration, creating a uniform, low-complexity state that precludes coherent interference (Brown, Lydic, & Schiff, 2010; Pal et al., 2020).
- Psychedelics (e.g., psilocybin, LSD): Attenuate which-path information (reducing precision of high-level priors), thereby amplifying interference. This is evidenced by DMN disintegration, increased entropy, and global connectivity (Carhart-Harris et al., 2012, 2014; Carhart-Harris & Friston, 2019).
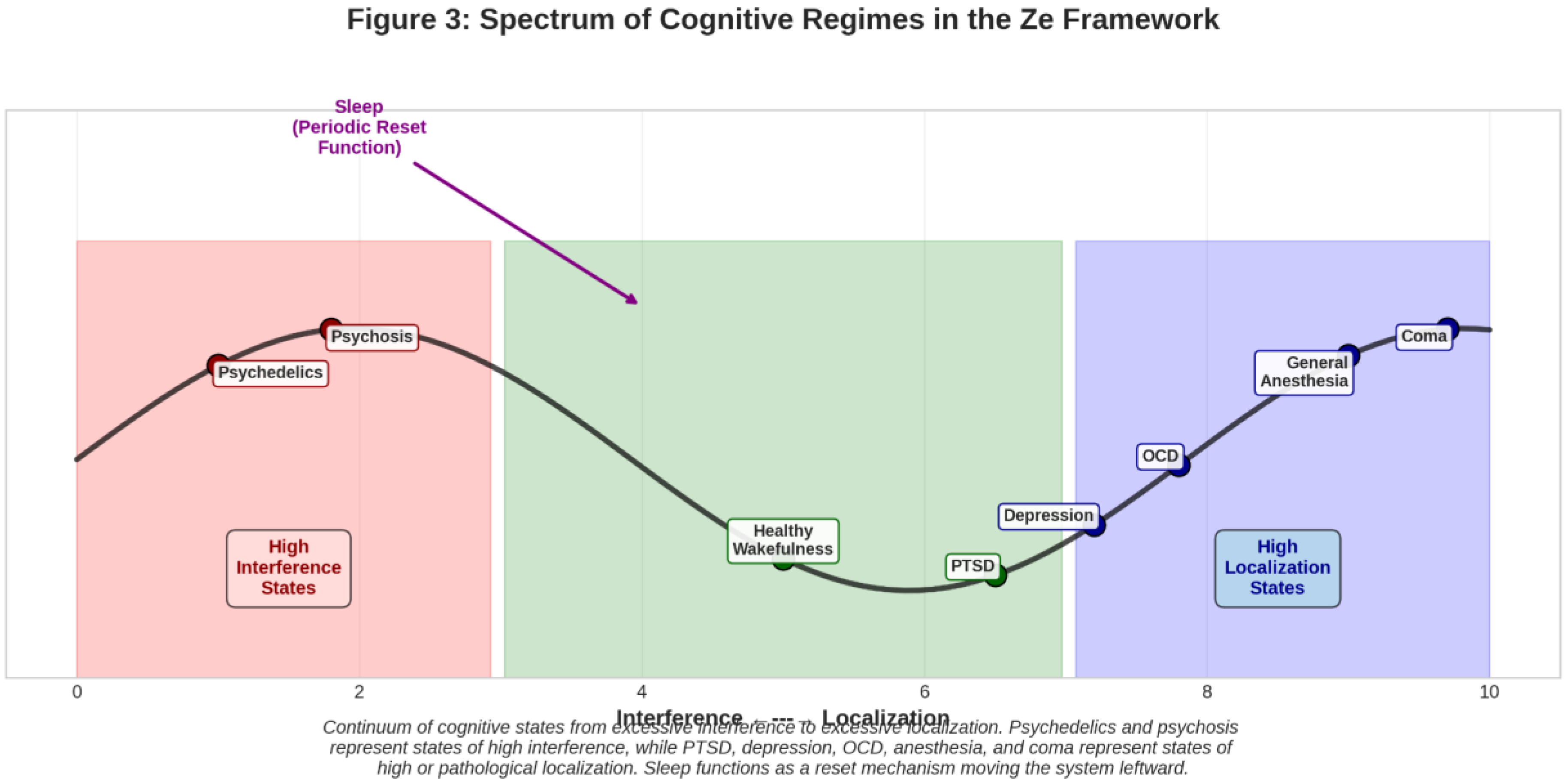
Figure 3.
Spectrum of Cognitive Regimes in the Ze Framework (A one-dimensional spectrum with “Excessive Interference” on the left, “Healthy Balance” in the center, and “Excessive Localization” on the right. Key states are plotted along it: Psychedelics and Psychosis towards the left; Coma and General Anesthesia towards the far right (as null/artificial states); PTSD, Depression, and OCD to the right of center; Sleep as a periodic resetting function moving the system leftwards from localized states.).
Figure 3.
Spectrum of Cognitive Regimes in the Ze Framework (A one-dimensional spectrum with “Excessive Interference” on the left, “Healthy Balance” in the center, and “Excessive Localization” on the right. Key states are plotted along it: Psychedelics and Psychosis towards the left; Coma and General Anesthesia towards the far right (as null/artificial states); PTSD, Depression, and OCD to the right of center; Sleep as a periodic resetting function moving the system leftwards from localized states.).
10. Against Copenhagen: Locality Without an Observer
The Ze framework, grounded in decoherence theory, forcefully rejects the Copenhagen notion that an observer causes collapse (Heisenberg, 1958). Localization is a physical, observer-independent process (Zurek, 2003). In the brain, “which-path” information from the environment and the brain’s own structure forces resolution via winner-take-all dynamics (Engel & Singer, 2001; Feldman & Friston, 2010). Consciousness is a consequence of localization (e.g., global availability of the winning model), not its cause (Dehaene & Naccache, 2001; Hohwy, 2016). Neural precursors to decisions occur before conscious awareness (Libet et al., 1983; Soon, Brass, Heinze, & Haynes, 2008).
The unifying mechanistic principle is the minimization of variational free energy. The brain implements the same physics of uncertainty resolution as the double-slit system, at a higher level of complexity (Friston, 2010, 2013).
11. Conclusions: The Brain as an Interferometric Inference Engine
The double-slit experiment reveals a fundamental operational mode for any complex system performing active inference. The brain is a biological interferometer where:
- Interference is the norm: The default is a superposition of hypotheses.
- Localization is a forced event: Driven by free energy gradients, environmental evidence, and action imperative.
- Sleep is the essential eraser: Periodically resetting path commitments to restore flexibility.
This tripartite scheme provides a unified framework linking quantum physics, cognitive neuroscience, and psychiatry. The brain does not observe quantum reality; it is an evolved embodiment of its core computational logic. The famous experiment is a continuous, lived process within us—the ceaseless, self-decohiring inference that is the fundamental game of staying alive.
References
- Alkire, M. T.; Hudetz, A. G.; Tononi, G. Consciousness and anesthesia. Science 2008, 322(5903), 876–880. [Google Scholar] [CrossRef]
- Arndt, M.; Nairz, O.; Vos-Andreae, J.; Keller, C.; van der Zouw, G.; Zeilinger, A. Wave–particle duality of C60 molecules. Nature 1999, 401(6754), 680–682. [Google Scholar] [CrossRef]
- Aston-Jones, G.; Cohen, J. D. An integrative theory of locus coeruleus-norepinephrine function: adaptive gain and optimal performance. Annual Review of Neuroscience 2005, 28, 403–450. [Google Scholar] [CrossRef] [PubMed]
- Botvinick, M. M.; Cohen, J. D.; Carter, C. S. Conflict monitoring and anterior cingulate cortex: an update. Trends in Cognitive Sciences 2004, 8(12), 539–546. [Google Scholar] [CrossRef]
- Brewin, C. R. Re-experiencing traumatic events in PTSD: new avenues in research on intrusive memories and flashbacks. European Journal of Psychotraumatology 2015, 6(1), 27180. [Google Scholar] [CrossRef] [PubMed]
- Brown, E. N.; Lydic, R.; Schiff, N. D. General anesthesia, sleep, and coma. New England Journal of Medicine 2010, 363(27), 2638–2650. [Google Scholar] [CrossRef] [PubMed]
- Bruza, P. D.; Wang, Z.; Busemeyer, J. R. Quantum cognition: a new theoretical approach to psychology. Trends in Cognitive Sciences 2015, 19(7), 383–393. [Google Scholar] [CrossRef]
- Buckner, R. L.; Carroll, D. C. Self-projection and the brain. Trends in Cognitive Sciences 2007, 11(2), 49–57. [Google Scholar] [CrossRef]
- Carhart-Harris, R. L.; Friston, K. J. REBUS and the anarchic brain: toward a unified model of the brain action of psychedelics. Pharmacological Reviews 2019, 71(3), 316–344. [Google Scholar] [CrossRef]
- Carhart-Harris, R. L.; Erritzoe, D.; Williams, T.; Stone, J. M.; Reed, L. J.; Colasanti, A.; Nutt, D. J. Neural correlates of the psychedelic state as determined by fMRI studies with psilocybin. Proceedings of the National Academy of Sciences 2012, 109(6), 2138–2143. [Google Scholar] [CrossRef]
- Carhart-Harris, R. L.; Leech, R.; Hellyer, P. J.; Shanahan, M.; Feilding, A.; Tagliazucchi, E.; Nutt, D. The entropic brain: a theory of conscious states informed by neuroimaging research with psychedelic drugs. Frontiers in Human Neuroscience 2014, 8, 20. [Google Scholar] [CrossRef]
- Cisek, P.; Kalaska, J. F. Neural mechanisms for interacting with a world full of action choices. Annual Review of Neuroscience 2010, 33, 269–298. [Google Scholar] [CrossRef] [PubMed]
- Clark, A. Whatever next? Predictive brains, situated agents, and the future of cognitive science. Behavioral and Brain Sciences 2013, 36(3), 181–204. [Google Scholar] [CrossRef]
- Corlett, P. R.; Horga, G.; Fletcher, P. C.; Alderson-Day, B.; Schmack, K.; Powers, A. R. Hallucinations and strong priors. Trends in Cognitive Sciences 2019, 23(2), 114–127. [Google Scholar] [CrossRef] [PubMed]
- Dehaene, S.; Naccache, L. Towards a cognitive neuroscience of consciousness: basic evidence and a workspace framework. Cognition 2001, 79(1–2), 1–37. [Google Scholar] [CrossRef]
- Dehaene, S.; Changeux, J. P.; Naccache, L.; Sackur, J.; Sergent, C. Conscious, preconscious, and subliminal processing: a testable taxonomy. Trends in Cognitive Sciences 2006, 10(5), 204–211. [Google Scholar] [CrossRef] [PubMed]
- Diekelmann, S.; Born, J. The memory function of sleep. Nature Reviews Neuroscience 2010, 11(2), 114–126. [Google Scholar] [CrossRef]
- Engel, A. K.; Singer, W. Temporal binding and the neural correlates of sensory awareness. Trends in Cognitive Sciences 2001, 5(1), 16–25. [Google Scholar] [CrossRef]
- Feldman, H.; Friston, K. J. Attention, uncertainty, and free-energy. Frontiers in Human Neuroscience 2010, 4, 215. [Google Scholar] [CrossRef]
- Fletcher, P. C.; Frith, C. D. Perceiving is believing: a Bayesian approach to explaining the positive symptoms of schizophrenia. Nature Reviews Neuroscience 2009, 10(1), 48–58. [Google Scholar] [CrossRef]
- Friston, K. The free-energy principle: a rough guide to the brain? Trends in Cognitive Sciences 2009, 13(7), 293–301. [Google Scholar] [CrossRef]
- Friston, K. The free-energy principle: a unified brain theory? Nature Reviews Neuroscience 2010, 11(2), 127–138. [Google Scholar] [CrossRef]
- Friston, K. Life as we know it. Journal of The Royal Society Interface 2013, 10(86), 20130475. [Google Scholar] [CrossRef]
- Friston, K.; Kiebel, S. Predictive coding under the free-energy principle. Philosophical Transactions of the Royal Society B: Biological Sciences 2009, 364(1521), 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.; Daunizeau, J.; Kilner, J.; Kiebel, S. J. Action and behavior: a free-energy formulation. Biological Cybernetics 2010, 102(3), 227–260. [Google Scholar] [CrossRef]
- Friston, K.; FitzGerald, T.; Rigoli, F.; Schwartenbeck, P.; Pezzulo, G. Active inference: a process theory. Neural Computation 2016, 29(1), 1–49. [Google Scholar] [CrossRef]
- Friston, K. J.; Stephan, K. E.; Montague, R.; Dolan, R. J. Computational psychiatry: the brain as a phantastic organ. The Lancet Psychiatry 2014, 1(2), 148–158. [Google Scholar] [CrossRef]
- Hackermüller, L.; Hornberger, K.; Brezger, B.; Zeilinger, A.; Arndt, M. Decoherence of matter waves by thermal emission of radiation. Nature 2004, 427(6976), 711–714. [Google Scholar] [CrossRef] [PubMed]
- Haggard, P. Human volition: towards a neuroscience of will. Nature Reviews Neuroscience 2008, 9(12), 934–946. [Google Scholar] [CrossRef]
- Hassabis, D.; Maguire, E. A. The construction system of the brain. Philosophical Transactions of the Royal Society B: Biological Sciences 2009, 364(1521), 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- Heisenberg, W. Physics and philosophy: The revolution in modern science; Harper & Row, 1958. [Google Scholar]
- Herz, R. S.; von Clef, J. The influence of verbal labeling on the perception of odors: Evidence for olfactory illusions? Perception 2001, 30(3), 381–391. [Google Scholar] [CrossRef]
- Hobson, J. A.; Friston, K. J. Waking and dreaming consciousness: Neurobiological and functional considerations. Progress in Neurobiology 2012, 98(1), 82–98. [Google Scholar] [CrossRef]
- Hohwy, J. The self-evidencing brain. Noûs 2016, 50(2), 259–285. [Google Scholar] [CrossRef]
- Jaba, T. Dasatinib and quercetin: short-term simultaneous administration yields senolytic effect in humans. Issues and Developments in Medicine and Medical Research 2022, Vol. 2, 22–31. [Google Scholar]
- Jung-Beeman, M.; Bowden, E. M.; Haberman, J.; Frymiare, J. L.; Arambel-Liu, S.; Greenblatt, R.; Kounios, J. Neural activity when people solve verbal problems with insight. PLoS Biology 2004, 2(4), e97. [Google Scholar] [CrossRef]
- Knill, D. C.; Pouget, A. The Bayesian brain: the role of uncertainty in neural coding and computation. Trends in Neurosciences 2004, 27(12), 712–719. [Google Scholar] [CrossRef]
- Lanius, R. A.; Vermetten, E.; Pain, C. (Eds.) The impact of early life trauma on health and disease: The hidden epidemic; Cambridge University Press, 2010. [Google Scholar]
- Laureys, S. The neural correlate of (un)awareness: lessons from the vegetative state. Trends in Cognitive Sciences 2005, 9(12), 556–559. [Google Scholar] [CrossRef]
- Lewis, P. A.; Durrant, S. J. Overlapping memory replay during sleep builds cognitive schemata. Trends in Cognitive Sciences 2011, 15(8), 343–351. [Google Scholar] [CrossRef]
- Lewis, P. A.; Knoblich, G.; Poe, G. How memory replay in sleep boosts creative problem-solving. Trends in Cognitive Sciences 2018, 22(6), 491–503. [Google Scholar] [CrossRef] [PubMed]
- Libet, B.; Gleason, C. A.; Wright, E. W.; Pearl, D. K. Time of conscious intention to act in relation to onset of cerebral activity (readiness-potential). Brain 1983, 106(3), 623–642. [Google Scholar] [CrossRef] [PubMed]
- McCormick, D. A.; Bal, T. Sleep and arousal: thalamocortical mechanisms. Annual Review of Neuroscience 1997, 20, 185–215. [Google Scholar] [CrossRef]
- Meng, M.; Tong, F. Can attention selectively bias bistable perception? Differences between binocular rivalry and ambiguous figures. Journal of Vision 2004, 4(7), 539–551. [Google Scholar] [CrossRef]
- Milner, A. D.; Goodale, M. A. Two visual systems re-viewed. Neuropsychologia 2008, 46(3), 774–785. [Google Scholar] [CrossRef]
- Muto, V.; Jaspar, M.; Meyer, C.; Kussé, C.; Chellappa, S. L.; Degueldre, C.; Phillips, C. Local modulation of human brain responses by circadian rhythmicity and sleep debt. Science 2016, 353(6300), 687–690. [Google Scholar] [CrossRef] [PubMed]
- Pace-Schott, E. F.; Hobson, J. A. The neurobiology of sleep: genetics, cellular physiology and subcortical networks. Nature Reviews Neuroscience 2002, 3(8), 591–605. [Google Scholar] [CrossRef]
- Pal, D.; Li, D.; Dean, J. G.; Brito, M. A.; Liu, T.; Fryzel, A. M.; Mashour, G. A. Level of consciousness is dissociable from electroencephalographic measures of cortical connectivity, slow oscillations, and complexity. Journal of Neuroscience 2020, 40(3), 605–618. [Google Scholar] [CrossRef]
- Raichle, M. E. The brain’s default mode network. Annual Review of Neuroscience 2015, 38, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Riemann, D.; Krone, L. B.; Wulff, K.; Nissen, C. Sleep, insomnia, and depression. Neuropsychopharmacology 2020, 45(1), 74–89. [Google Scholar] [CrossRef] [PubMed]
- Robbins, T. W.; Vaghi, M. M.; Banca, P. Obsessive-Compulsive Disorder: Puzzles and Prospects. Neuron 2019, 102(1), 27–47. [Google Scholar] [CrossRef]
- Roiser, J. P.; Elliott, R.; Sahakian, B. J. Cognitive mechanisms of treatment in depression. Neuropsychopharmacology 2012, 37(1), 117–136. [Google Scholar] [CrossRef]
- Soon, C. S.; Brass, M.; Heinze, H. J.; Haynes, J. D. Unconscious determinants of free decisions in the human brain. Nature Neuroscience 2008, 11(5), 543–545. [Google Scholar] [CrossRef]
- Sterzer, P.; Adams, R. A.; Fletcher, P.; Frith, C.; Lawrie, S. M.; Muckli, L.; Corlett, P. R. The predictive coding account of psychosis. Biological Psychiatry 2018, 84(9), 634–643. [Google Scholar] [CrossRef] [PubMed]
- Tkemaladze, J. Reduction, proliferation, and differentiation defects of stem cells over time: a consequence of selective accumulation of old centrioles in the stem cells? Molecular Biology Reports 2023, 50(3), 2751–2761. [Google Scholar] [CrossRef] [PubMed]
- Tkemaladze, J. Editorial: Molecular mechanism of ageing and therapeutic advances through targeting glycative and oxidative stress. Front Pharmacol 2024, 14, 1324446. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tkemaladze, J. Old Centrioles Make Old Bodies. Annals of Rejuvenation Science 2026, 1(1). [Google Scholar] [CrossRef]
- Tkemaladze, J. Visions of the Future. Longevity Horizon 2026, 2(1). [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



