
Submitted:
09 January 2026
Posted:
12 January 2026
You are already at the latest version
Abstract
The endoplasmic reticulum (ER) stress is triggered by the accumulation of unfolded or misfolded proteins in the lumen of the ER, caused by factors like calcium depletion, oxidative imbalance, or high cellular protein demand. Current evidence suggests that ER stress is involved in the onset of diverse cardiovascular diseases, including hypertension. We previously showed that in normotensive and hypertensive animals, endothelins (ETs) regulate norepinephrine transporter (NET) activity and expression through impaired calcium management and oxidative stress. In the present study we sought to establish whether the ER stress was involved in ETs-induced NET impairment in salt-dependent hypertension. Hypertension was induced in rats by DOCA injections and NaCl in the drinking water. At week 4, sulfisoxazole (dual ET receptor antagonist) was orally administered for seven days. Blood pressure and heart rate were weekly monitored. At the end of the experiment, the adrenal medulla was dissected to perform electron microscopy studies and to assess NET expression and activity, ER stress markers, and ER-dependent apoptosis. Hypertensive rats showed elevated systolic blood pressure, reduced NET activity and expression, and significant ultrastructural changes in the adrenal medulla compatible with ER stress that were reversed by sulfisoxazole administration. Markers of ER stress (Bip, pPERK, p-eIF2α, CHOP, XBP1and IRE-1) were enhanced in DOCA-salt rats but decreased by sulfisoxazole. Furthermore, increased ER-dependent apoptosis was attenuated by the ETA/ETB antagonist. Sulfisoxazole treatment reduced blood pressure and ER stress improving NET function. These findings suggest that NET impairment in DOCA-salt hypertension results from ET-induced ER stress and ER-dependent apoptosis.
Keywords:
endothelins
; hypertension
; sulfisoxazole
; endothelin receptors
; endoplasmic reticulum stress
; neuronal norepinephrine transporter
1. Introduction
Strong clinical and experimental evidence sustains that the endoplasmic reticulum (ER) stress is one of the underlying mechanisms involved in the onset of several cardiovascular diseases [1,2]. Recent studies have shown a close relationship between ER stress and hypertension [1,2,3]. The ER is critical for the proper folding, maturation, and secretion of transmembrane and secreted proteins, but also functions as a calcium reservoir [4]. Diverse factors like calcium depletion, oxidative stress, or high cellular protein demand [1,2,3,4] disturb the ER, causing the accumulation of unfolded or misfolded proteins in the lumen, that leads to ER stress. To restore protein folding homeostasis, a well-conserved cellular mechanism called the unfolded protein response (UPR) is triggered. If the stress is prolonged or severe, the cells fail to recover, and apoptosis is activated [4,5].
Endothelins (ETs), the most potent vasopressor substances known, are a family of three isopeptides (ET-1, ET-2, and ET-3) widely distributed in different tissues and cell types. Their effects are mediated by two pharmacologically well-characterized receptors, ETA and ETB, which can activate diverse signaling pathways [6,7]. ETs (mainly ET-1 and ET-3) are implicated in the pathogenesis of hypertension and other cardiovascular diseases due to their vasoactive properties [8,9]. Numerous studies have shown that the ET system is activated in clinical and experimental hypertension [8,9]. A close relationship between the sympathetic nervous system and ETs was observed in patients with essential hypertension, where ET-1 contributes to increased sympathetic vascular tone through ETA receptors, both in normotensive and hypertensive subjects. Furthermore, hypertensive patients show increased susceptibility to the sympatho-excitatory effect of endogenous ET-1 [10,11]. Released catecholamines from the adrenal medulla (AM) significantly contribute to the development and maintenance of DOCA-salt hypertension [12,13]. In addition, plasma ETs are increased and enhance adrenomedullary catecholamine release in normotensive and hypertensive animals [13,14]. ETs also increase phosphoinositide turnover, oxidative stress, and calcium handling in the adrenal medulla and other tissues [15].
The norepinephrine neuronal transporter (NET) removes norepinephrine from the synaptic gap, thus ending noradrenergic signaling [16,17]. NET, a family member of the Na+/Cl- dependent membrane transporters, exhibits multiple cytoplasmic glycosylation and phosphorylation sites [16,17,18]. The glycosylated isoform of the protein represents the functionally active transporter. In hypertension and congestive heart failure, NET function is impaired at the sympathetic presynaptic terminals [19,20]. Moreover, NET density and norepinephrine uptake are diminished in noradrenergic nerve endings and PC12 cells following norepinephrine administration due to posttranscriptional changes induced by oxidative stress [21,22]. We previously reported that ET-1 and ET-3 differentially modulate NET activity, its internalization, and kinetic parameters in diverse brain areas in normotensive and hypertensive animals [23,24,25].
In the present study, we sought to determine the role of the ET system in regulating NET in the AM of DOCA-salt hypertensive rats, with a particular focus on the involvement of the ER stress.
2. Materials and Methods
2.1. Animals and Experimental Design
Male Sprague-Dawley rats (Outbred Crl:CE) weighing 125±5 g from the Facultad de Farmacia y Bioquímica of the Universidad de Buenos Aires facility were used in the experiments. Animals were housed in steel cages and maintained at 22-24°C and 45-55% humidity in a controlled room with a 12h light/dark cycle under the care of qualified personnel. All procedures followed the recommendations of the Guidelines for the Care and Use of Laboratory Animals (National Institutes of Health publication N85-23, 1985; revised 1996). The experimental protocols were approved by the Animal Care Committee of the Facultad de Farmacia y Bioquimica of the University of Buenos Aires (CICUAL-FFYB #0045151). Every effort was made to minimize the number of animals used and the stress caused by experimental procedures. Animals were randomly divided into control (normotensive rats) and DOCA-salt groups. Hypertension was induced by subcutaneous injections of 15mg/kg DOCA (MP Biomedicals, LLC OH, USA), twice a week, dissolved in sesame oil as vehicle and the administration of 1% NaCl in the drinking water for 5 weeks [26,27]. At week 4 a set of animals from the control and DOCA-salt groups received 1mg/kg sulfisoxazole (SFX) in the food [28]. Animals were daily weighed to adjust SFX dose. At the end of week 5, animals were euthanized and AM dissected. The experimental protocol is illustrated in Figure 1.
2.2. Methods
2.2.1. Norepinephrine Uptake Assay
Neuronal norepinephrine uptake was measured as previously detailed [23]. Briefly, AMs were preincubated at 37°C for 30 min in a gassed (95% O2 +5% CO2) standard Krebs solution (pH 7.4) containing 100µM hydrocortisone and 50µM pargyline to inhibit extraneuronal norepinephrine uptake and monoaminoxidase activity, respectively (MP Biomedicals, LLC OH, USA). Tissues were then incubated for 5 min with 100 nmol/L de 3H-norepinephrine. The reaction was stopped by adding cold Krebs buffer, followed by three 5-min washes with Krebs and homogenization in distilled water. Tritium activity was measured in the supernatants by usual scintillation counting methods (Wallac-WinSpectral 1414, Liquid Scintillation Counter; Turku, Finland), and proteins in the pellets according to Bradford et al., (1976). Data was expressed as percentage of the control group ± SD [23].
2.2.2. Western Blot Assay
For Western blot assay, samples (50µg protein) were separated on SDS-PAGE gel and electro-transferred to PVDF membranes (GE Healthcare, Amersham Biosciences, Little Chalfont, UK), then blocked with 5% nonfat dry milk, washed, and exposed to primary antibodies (Table 1) at 4°C overnight. Membranes were washed and incubated with horseradish peroxidase (HRP)-conjugated secondary antibody for 2 h at room temperature. Membranes were developed using a bioluminescent Western blotting detection system (Kalium Technologies, Buenos Aires, Argentina) and exposed to X ray films or determined by a digital system (GeneGnome XXQ, Syngene, Maryland, MD, USA). β-actin was used as loading control. Data was normalized to β-actin and expressed as percentage of the control group ± S.E.M.
2.2.3. Quantitative Reverse Transcription Real-Time PCR (RT-qPCR)
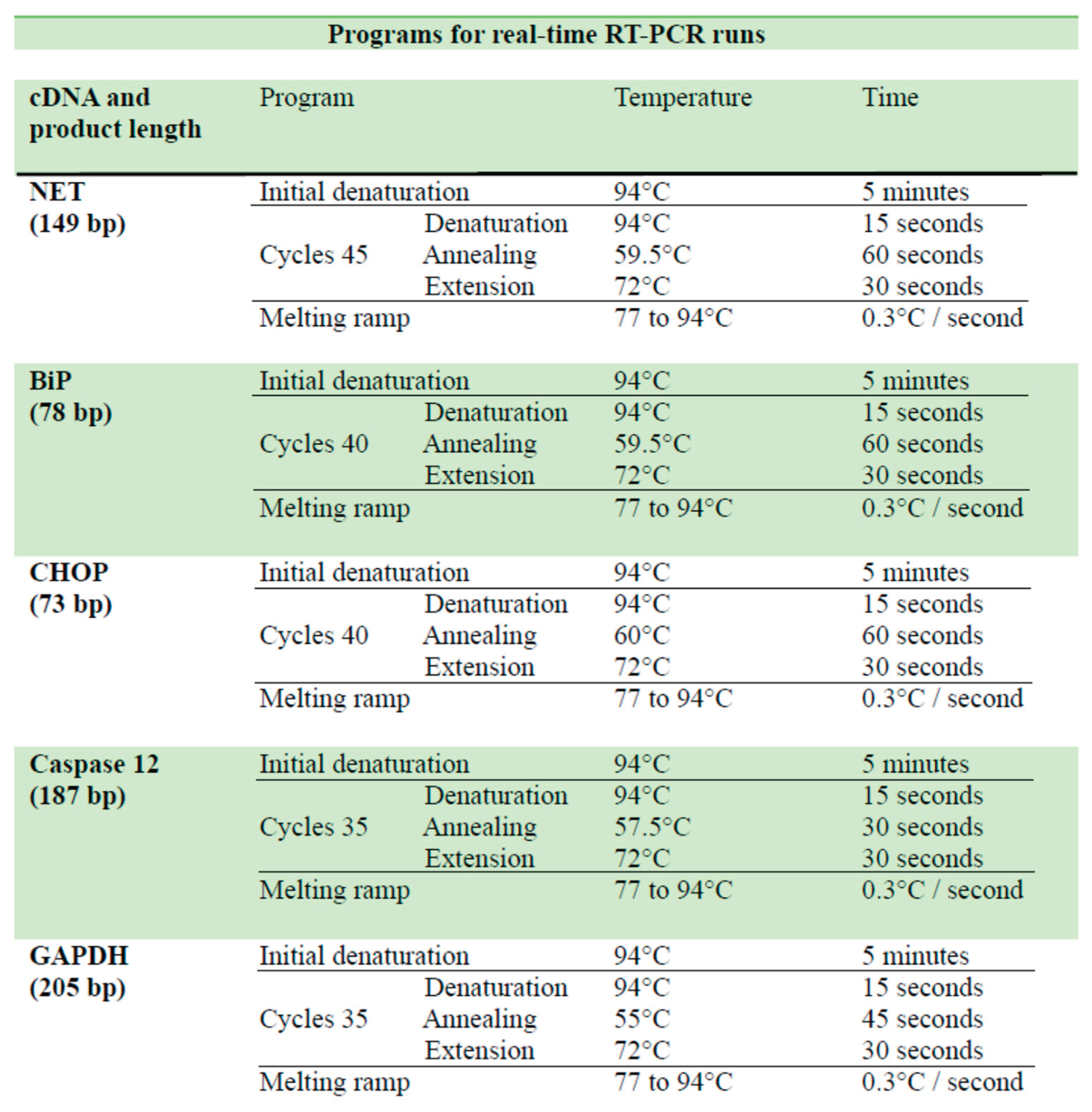
NET, binding immunoglobulin protein (Bip), C/EBP homologous protein (CHOP), caspase 12, and GADPH mRNA expression were examined by RT-qPCR. Total RNA was isolated from AM samples by ReagentTMfollowing the manufacturer´s instructions and incubated with RQ1 RNase-free DNase (Promega, Madison, WI, USA) at 37°C for 40 min to eliminate contaminating DNA. RNA quality and quantity were evaluated by 1% agarose gel electrophoresis and UV spectrometry, respectively. Reverse transcription was performed by adding MMLV reverse transcriptase (Promega, WI, USA), an RNase inhibitor, dNTPs, and oligo-dT primers (Genbiotech, Buenos Aires, Argentina). The products were confirmed by conventional PCR and verified using an agarose gel to ensure the presence of a single band corresponding to the primer pair used. The real time PCR was carried out in the same reaction mixture containing cDNA AM, GoTaq G2 DNA polymerase, GoTaq polymerase G2 buffer (Promega, MI, USA), dNTPs (Geniotech, Buenos Aires, Argentina), MgCl2, and EvaGreen as a fluorophore (Biotium, San Francisco, USA). Primers and PCR conditions are shown in Table 2 and Table 3. Results were normalized to GADPH expression (housekeeping gene). Quantification was performed using the 2−ΔΔCt method.
2.2.4. Transmission Electron Microscopy Studies
AMs were fixed by immersion in 3% w/v paraformaldehyde for 24 h at 4°C. Following washes in phosphate-buffered saline (PBS), tissues were kept in 0.32M sucrose in 0.1M PBS (pH 7.4) for 24 h at 4°C and then postfixed with 1.5% w/v osmium tetroxide in 0.1 mmol/L PBS for 2 h at 4 °C. Tissues were then washed and contrasted with 2% w/v uranyl acetate, dehydrated and embedded in Spurr medium kit (Ted Pella,Redding, CA, USA). Ultrathin sections were performed with an ultramicrotome Porter Bloom MT 1 and collected in 300 mesh copper grids, contrasted with uranyl acetate and stained with Reynolds solution, and photographed in a Gatam 1000 V coupled to a Zeiss EM109T Electron Microscope.
2.2.5. Terminal Deoxynucleotidyl Transferase-Mediated dUTP-Biotin DNA-Nick End Labelling (TUNEL) Assay
Apoptosis was quantified by the TUNEL assay (Promega, WI, USA). Adrenomedullary tissue was fixed in 5% buffered formaldehyde overnight, dehydrated in 70% ethanol, embedded in paraffin, and 5 µm-thick sections were obtained. Sections were then deparaffinized with xylene, hydrated with decreasing ethanol concentrations, and stained for breaks in DNA. Results were expressed as the number of apoptotic nuclei/10 fields (Leica ICC50W camera coupled to a Leica DM500 microscope). At least three sections per animal were counted and three animals per group.
2.3. Statistical Analysis
Results are expressed as the mean ± standard error of the mean (SD). The statistical analysis was performed by the analysis of variance (ANOVA) followed by the Student Newman Keuls test. A p value of 0.05 or less was considered statistically significant.
3. Results
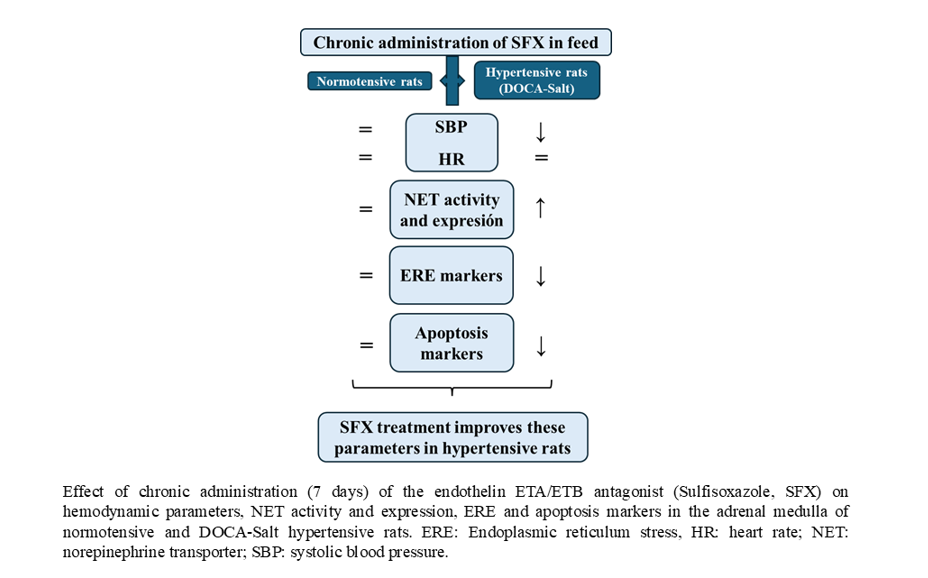
3.1. Hemodynamic Parameters
3.2. NET Activity and Expression
SFX inhibited the decrease in neuronal norepinephrine uptake observed in the AM of hypertensive rats but did not affect normotensive animals (Figure 3A). Protein expression of functional NET (glycosylated form) was increased in the AM of DOCA-salt rats following SFX administration without changes in normotensive animals (Figure 3B,C). Similar results were observed in mRNA NET (Figure 3D).
3.3. Ultrastructural Changes in the Adrenal Medulla Following SFX Administration
No changes in the chromaffin cells of normotensive animals were observed following SFX administration (Figure 4). However, in hypertensive animals, adrenomedullary cells exhibited edema, endoplasmic reticulum swelling, few granules, nuclear interdigitation, cytoplasmic vacuolization, and disruption of mitochondrial membrane. SFX treatment prevented the ultrastructural changes in chromaffin cells induced by hypertension. Cytoplasmic vacuolization and endoplasmic reticulum swelling were significantly reduced. Mitochondria exhibited conserved morphology, but cytoplasmic microvacuoles were observed (Figure 4).
3.4. ER Stress Makers Following SFX Administration
In DOCA-salt hypertensive rats, Bip protein and mRNA expression were increased in the AM (supporting UPR activation), but SFX administration prevented it (Figure 5A–C). Furthermore, in the AM the expression of enhanced protein kinase-like ER kinase (p-PERK) and downstream effector, the eukaryotic initiation factor 2 p-(p-eIF2α) was also increased in hypertension but partially reduced by SFX (Figure 6A–C). pPERK also activates CHOP, which stimulates apoptosis by regulating members of the Bcl2 family. CHOP protein and mRNA expression were augmented in the chromaffin cells of hypertensive rats but reduced by SFX administration (Figure 6A,D,E). In DOCA-salt rats, both inositol-requiring enzyme-1 (IRE-1) and XBP1 were enhanced, but the ETA/ETB receptor antagonist significantly diminished them (Figure 7A–C). It is worth noting that SFX did not modify the expression of UPR proteins in the AM of normotensive animals.
3.5. Effect of SFX Treatment on Apoptosis
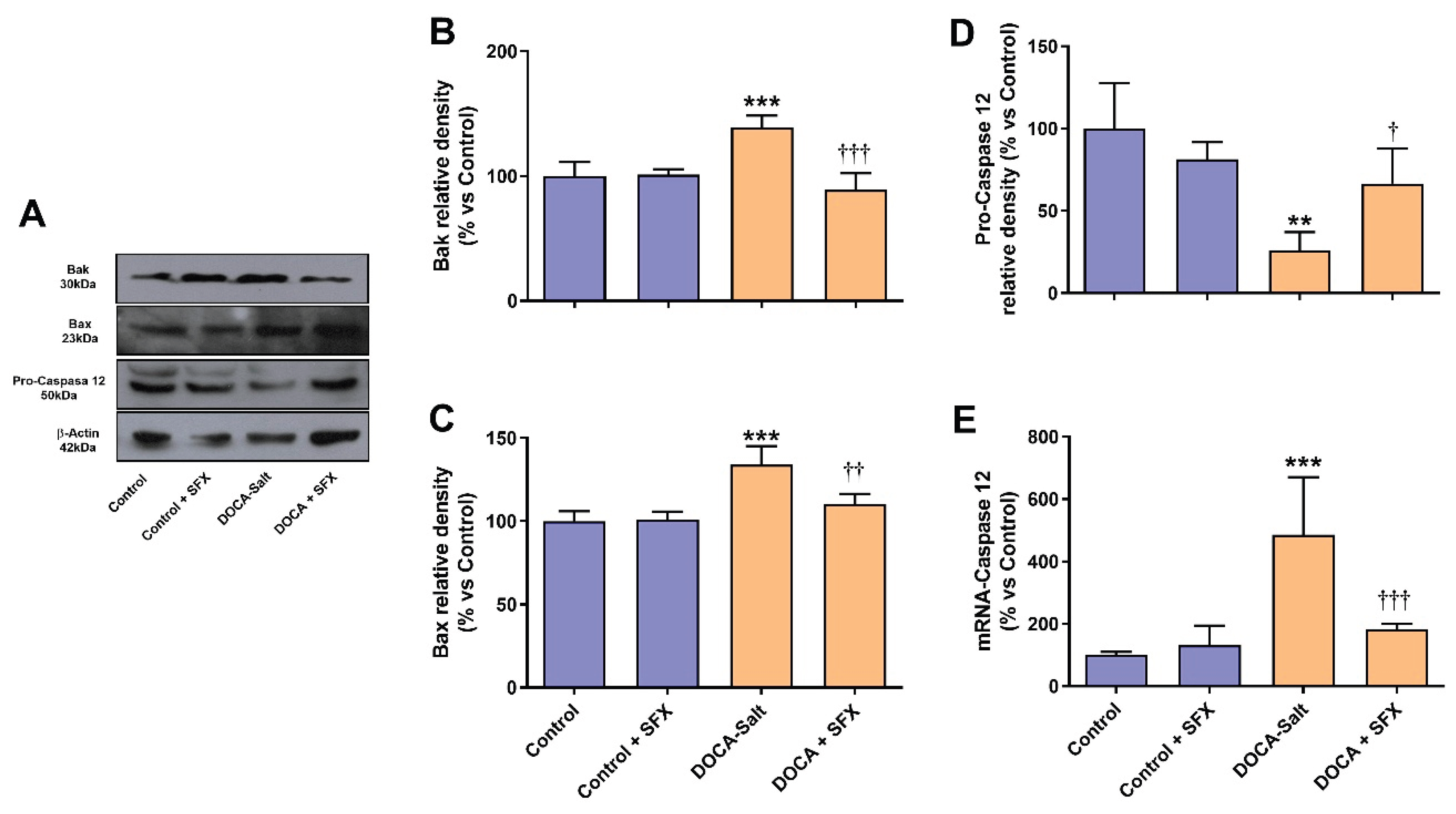
The increased number of apoptotic nuclei induced by hypertension in the AM was significantly reduced by SFX administration (Figure 7A,B). Surprisingly, SFX induced apoptosis in the AM of normotensive rats although to a significant lesser extent than in hypertensive animals (Figure 7A,B). The adrenomedullary expression of Bak and Bax was enhanced in hypertension, but the ETA/ETB antagonist prevented the expression of both proapoptotic proteins (Figure 8A–C). Caspase 12, the only member of the caspase family located in the ER, mediates ER-dependent apoptosis by activating caspases 3 and 9. Procaspase 12 expression diminished in the AM of hypertensive rats, but SFX administration prevented it. In accordance, augmented mRNA caspase 12 expression in the AMof DOCA-salt rats was restored to control values by SFX.
4. Discussion
Despite numerous studies the pathogenesis of hypertension, particularly salt-sensitive hypertension remains to be fully understood. The present study advances our knowledge about the underlying mechanisms involved in DOCA-salt hypertension in the AM, focusing on ETs contribution to catecholamine imbalance. The major findings were that one-week SFX administration to DOCA-salt hypertensive rats decreased systolic blood pressure, normalized NET activity, and reduced ER stress and ER-dependent apoptosis in the AM.
The brain and peripheral catecholamine system is enhanced in salt-dependent hypertension. In 1972, Champlain and van Ameringen showed that chemical sympathectomy with 6-OH-DA or bilateral adrenalectomy, performed before DOCA and salt treatment, reduced blood pressure in hypertensive rats more markedly than in normotensive animals, although it did not fully normalize it. However, after both chemical sympathectomy and bilateral adrenalectomy, DOCA-salt animals exhibited blood pressure levels comparable to normotensive controls, indicating that the combined sympathetic and adrenal medullary activity is essential in this model [29]. In addition, it was reported that catecholamine synthesis and release is increased in clinical and experimental hypertension [30,31,32]. The AM synthesizes ETs and expresses both ET receptors [15]. ET-1 evokes adrenomedullary catecholamine release in normotensive and DOCA-salt hypertensive animals [33].
Figure 9.
Bak, Bax and Caspase 12 determination following sulfisoxazole (SFX, 1g/Kg) administration to normotensive and DOCA-salt hypertensive rats. Protein expression was determined by Western blot whereas mRNA expression by RT-qPCR as detailed in Materials and Methods. A) Representative immunoblots. B) Bak protein expression. ***: p<0.001 vs. control; †††: p<0.001 vs. DOCA-salt. C) Bax protein expression. ***: p<0.001 vs. control; ††: p<0.01 vs. DOCA-salt. D) Pro-Caspase 12 protein expression. **: p<0.01 vs. control; †: p<0.05 vs. DOCA-salt. E) Levels of mRNA Caspase 12. ***: p<0.001 vs. control; †††: p<0.001 vs. DOCA-salt. Results were expressed as the mean ± SD. Number of experiments: 3-5.
Figure 9.
Bak, Bax and Caspase 12 determination following sulfisoxazole (SFX, 1g/Kg) administration to normotensive and DOCA-salt hypertensive rats. Protein expression was determined by Western blot whereas mRNA expression by RT-qPCR as detailed in Materials and Methods. A) Representative immunoblots. B) Bak protein expression. ***: p<0.001 vs. control; †††: p<0.001 vs. DOCA-salt. C) Bax protein expression. ***: p<0.001 vs. control; ††: p<0.01 vs. DOCA-salt. D) Pro-Caspase 12 protein expression. **: p<0.01 vs. control; †: p<0.05 vs. DOCA-salt. E) Levels of mRNA Caspase 12. ***: p<0.001 vs. control; †††: p<0.001 vs. DOCA-salt. Results were expressed as the mean ± SD. Number of experiments: 3-5.
Present findings further support the role of ETs in hypertension. The administration of the ETA/ETB antagonist SFX to DOCA-salt hypertensive rats reduced blood pressure at 48 h whereas no effect was observed when given to control animals. However, heart rate showed no changes in either group following SFX administration. Some authors reported no changes in heart rate, but others found a slight increase in hypertensive rats. These controversial findings may be explained by the observation that in DOCA-salt hypertensive rats, the baroreflex response is impaired even before systolic blood pressure elevation [34,35]. ETB receptors play a role in baroreflex impairment, that it is further aggravated in animals with increased sodium intake. This observation may contribute to salt dependent hypertension [36]. Furthermore, and in accordance with our results it was shown that ER stress inhibition reduces systolic blood pressure in DOCA-salt hypertensive rats [37].
Both the peripheral and brain endothelin system play a relevant role in salt sensitive hypertension [11,15,38]. Peripherally, ETs regulate vascular tone by inducing vasoconstriction through ETA/ETB receptor activation on vascular smooth muscle cells or by promoting vasodilation via ETB receptors on endothelial cells [8,9,15]. Peripheral vascular resistance is regulated by ET-1 through its potent vasoconstriction, stimulation of catecholamine release from the AM and its synergic interaction with angiotensin II [39]. In DOCA-salt hypertension, circulating ET-1 levels and ET receptor activity are elevated, supporting a significant contribution of ETs to increased vascular tone and blood pressure [11,15,38]. Accordingly, peripheral blockade with specific ETA antagonists or dual ETA/ETB receptor antagonists has been shown to reduce blood pressure in various animal models and in certain forms of hypertension including salt-dependent hypertension [8,9,15]. The antibacterial agent sulfanilamide, SFX, has been used as an ET receptor antagonist. It has been reported to reduce pulmonary artery pressure in an experimental model of pulmonary hypertension [28], enhance the analgesic effect of clonidine [40], and protect the retina from ischemic insults, such as that occurring in glaucoma [41]. In addition, SFX shows anticancer properties by suppressing the secretion of extracellular vesicles and altering vesicle cargo in cancer cells [42]. To our knowledge, the present study is the first to evaluate SFX on blood pressure. Notably, SFX has an IC50 value of 0.66 μM for ETA receptors and 22 μM for ETB receptors, suggesting a stronger selectivity towards ETA. However, SFX IC50 values are comparable to those of other dual ET antagonists such as bosentan and darusentan (IC50 ETA/ETB: 20 and 62, respectively). Specific ETA receptor antagonists such as atrasentan, sitaxentan and BQ123 show high ETA/ETB selectivity with ratios of 950, 7000 and 857 respectively.
In the present study we also demonstrate that SFX prevented the reduction in NET activity and expression in the AM of hypertensive rats. The diminished NET activity observed in DOCA-salt hypertension results from decreased expression of the glycosylated form of the transporter, which represents the mature functional form essential for transport stability, activity and normal surface trafficking. A previous study showed that ET-1 increases norepinephrine release in the AM of normotensive and hypertensive rats [33]. Additionally, ET-1 through ETA activation inhibits cardiac NET activity and that the ETA receptor antagonist darusentan reduces norepinephrine uptake and NET binding sites in rats with heart failure induced by transverse aortic constriction [43]. NET dysfunction has also been reported in hypertension and congestive heart failure likely due to increased interstitial norepinephrine [22]. NE administration reduces both NET density and norepinephrine uptake within sympathetic nerve terminals and PC12 cells [21,22]. In a previous study we reported that ET-1 and ET-3 differentially regulated NET activity, internalization, and kinetic properties in diverse brain regions in normotensive and hypertensive rats [23,24,25]. Our present and earlier findings show that increased adrenomedullary norepinephrine in DOCA-salt hypertension impairs NET synthesis and maturation. In accordance, studies in PC12 showed that exposure to norepinephrine negatively regulates NET by diverse mechanisms resulting in ER stress.
In the present work we show that SFX significantly reduced ER stress in the AM of hypertensive rats. Increasing evidence supports that ER stress is an early event in the pathogenesis of cardiovascular diseases, including hypertension [1,3,44]. Calcium depletion, oxidative stress, or high cellular protein demand induces the accumulation of misfolded and unfolded protein in the ER, which triggers the UPR [4,5]. This well-conserved cellular response is mediated by the activation of the stress sensor proteins PERK, IRE-1, and ATF6 to restore ER homeostasis [4,5]. Activation of the UPR clears misfolded proteins from the ER, reduces the flow of proteins to the ER, and increases the expression of proteins like chaperones and foldases tending to protect the cell. If the stress is severe or chronic the ER homeostasis is not restored so programmed cell death is activated [4,5]. Our results show that ER homeostasis is almost restored by SFX in the AM of hypertensive rats. Several evidence supports that ER stress is involved in the pathogenesis of experimental and human hypertension [3,44]. Administration of tauroursodeoxycholic acid (TUDCA), an inhibitor of ER stress, to DOCA-salt rats enhances myocardial contractility while reducing cardiac apoptosis and fibrosis [45]. In the same animal model, TUDCA also improves hepatic function [46]. Various studies show that ET-1 leads to ER stress in diverse cell types. In kidney cells ETs induce ER stress and NLRP3 inflammasome activation, that were prevented by an fosfoamidon (inhibitor of endothelin-converting enzyme) exposure [47]. In atherosclerotic plaques, ET-1 levels are elevated, and this peptide is a key contributor to the ER stress observed in atherosclerosis [48].
In the present study blockade of ET receptors attenuated ER stress and ER-dependent apoptosis in DOCA-salt hypertensive rats. SFX reduced Bip protein and gene expression supporting attenuation of ER stress. Under unstressed conditions Bip binds to the three stress sensor proteins PERK, IRE1 and activating transcription factor 6 (ATF 6). However, when the stress is triggered, Bip preferentially binds to unfolded proteins resulting in the release and activation of the stress sensor proteins that mediate the UPR. In addition, Bip expression reduction is indicative of less protein load to the ER [4,5].
In accordance, SFX also reduced the sensor protein PERK expression and downstream effectors, eIF2α and CHOP. Upon BiP dissociation, PERK is activated by autophosphorylation and oligomerization. Activated PERK (pPERK) phosphorylates eIF2α leading to a general reduction in translation initiation [50]. In the AM of hypertensive rats both proteins (p-PERK and p-eIF2α) were enhanced but SFX reduced their expression supporting decreased protein load to the ER. PERK also activates CHOP that stimulates programmed cell death by regulating members of the Blc2 family and increasing proapoptotic proteins like Bax, Puma, Bim and Bak among others. Blockade of ET receptors by SFX reduced the increase in CHOP protein and mRNA expression observed in the AM of DOCA-salt rats supporting diminished ER-dependent apoptosis. These findings were further supported by the reduced expression of the proapoptotic proteins Bax and Bak. ER stress induces conformational changes in Bax and Bak that favors ER calcium release to the cytoplasm that cleaves procaspase 12 to yield caspase 12 [51]. In hypertensive rats, procaspase 12 expression was decreased in the AM supporting an increase in caspase 12 that was prevented by SFX. It should be noted that caspase 12 is the only member of the caspase family in the ER that mediates ER-dependent apoptosis through caspase 3 and 9 activation.
Blockade of ET receptors also reduced the expression of another sensor stress protein, inositol-requiring enzyme-1 (IRE-1) and its downstream effector XBP1 that were increased in hypertension. Increasing evidence supports the idea that XBP1 is associated with the pathogenesis of cardiovascular diseases [52]. The IRE-1/XBP1 branch of the UPR upregulates the expression of several chaperones and foldases, causes ER expansion and attenuates protein synthesis [53]. The changes in ER stress markers induced by SFX in the AM of hypertensive rats were further supported by electron microscopy studies. To enhance protein-folding capacity, the UPR expands the ER size through increased biogenesis components mediated by various UPR signaling pathways. SFX treatment prevented the ultrastructural changes in chromaffin cells induced by hypertension like edema, ER swelling, reduced granules, nuclear interdigitation, cytoplasmic vacuolization, and disruption of mitochondrial membrane.
5. Conclusion
In summary, we provide evidence that in salt sensitive hypertension ETA/ETB blockade by SFX not only significantly reduces systolic blood pressure but also improves NET function by attenuating ER stress in the AM. Whether ET-1 effect is direct or mediated by ET-1-induced norepinephrine release is presently unknown. Both present and previous findings support that the ET system in the AM impairs NET function through ET-induced ER stress and ER-mediated apoptosis. The observation that SFX significantly improved systolic blood pressure and NET function in the AM suggests that SFX may represent a promising therapeutic option for the treatment of salt-dependent hypertension.
Author Contributions
Conceptualization, M.S.V.; investigation, M.I.S., F.A.I.C. and M.F.F.; validation, M.I.S. and F.A.I.C.; methodology, M.I.S. and F.A.I.C.; funding acquisition, M.S.V. and L.G.B.; supervision, M.S.V.; writing—original draft preparation, M.S.V.; review and editing, M.S.V. and L.G.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the University of Buenos Aires (UBACYT 0020220100159BA and CONICET (PIP 11220210100433CO) to M.S.V. and L.G.B., respectively.
Institutional Review Board Statement
Experimental protocols were approved by the Animal Care Committee of the Faculty of Pharmacy and Biochemistry, University of Buenos Aires (CICUAL-FFYB #0045151).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data is contained within the article.
Acknowledgments
The authors thank Dr. Mónica Navarro and Mrs. Gabriela Nocetti for their excellent technical assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Liu, M-Q.; Chen, Z.; Chen, L-X. Endoplasmic reticulum stress: a novel mechanism and therapeutic target for cardiovascular diseases. Acta Pharmacol. Sin. 2016, 37, 425–443. [Google Scholar] [CrossRef]
- Ochoa, C.D.; Wu, R.F.; Terada, L.S. ROS signaling and ER stress in cardiovascular disease. Mol. Asp. Med. 2018, 63, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Balhara, M.; Neikirk, K.; Marshall, A.; Hinton, A., Jr.; Kirabo, A. Endoplasmic reticulum stress in hypertension and salt sensitivity of blood pressure. Curr. Hypertens. Rep. 2024, 26, 273–290. [Google Scholar] [CrossRef] [PubMed]
- Almanza, A.; Carlesso, A.; Chintha, C.; Creedican, S.; Doultsinos, D.; Leuzzi, B.; Luis, A.; McCarthy, N.; Montibeller, L.; More, S.; Papaioannou, A.; Püschel, F.; Sassano, M.L.; Skoko, J.; Agostinis, P.; de Belleroche, J.; Eriksson, L.A.; Fulda, S.; Gorman, A.M.; Healy, S.; Kozlov, A.; Muñoz-Pinedo, C.; Rehm, M.; Chevet, E.; Samali, A. Endoplasmic reticulum stress signalling – from basic mechanisms to clinical applications. FEBS J. 2019, 286(2), 241–278. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, C.; Gu, R.; Han, R.; Li, Z.; Xu, X. Endoplasmic reticulum stress in diseases. MedComm 2024, 5, e701. [Google Scholar] [CrossRef]
- Davenport, A.P.; Hyndman, K.A.; Dhaun, N.; Southan, C.; Kohan, D.E.; Pollock, J.S.; Pollock, D.M.; Webb, D.J.; Maguire, J.J. Endothelin. Pharmacol. Rev. 2016, 68, 357–418. [Google Scholar] [CrossRef]
- Enevoldsen, F.C.; Sahana, J.; Wehland, M.; Grimm, D.; Infanger, M.; Krüger, M. Endothelin receptor antagonists: status quo and future perspectives for targeted therapy. J. Clin. Med. 2020, 9, 824. [Google Scholar] [CrossRef]
- Barton, M.; Yanagisawa, M. Endothelin: 30 Years from discovery to therapy. Hypertension 2019, 74, 1232–1265. [Google Scholar] [CrossRef] [PubMed]
- Dhaun, N.; Webb, D.J. Endothelins in cardiovascular biology and therapeutics. Nat. Rev. Cardiol. 2019, 16, 491–502. [Google Scholar] [CrossRef]
- Bruno, R.M.; Sudano, I.; Ghiadoni, L.; Masi, L.; Taddei, S. Interactions between sympathetic nervous system and endogenous endothelin in patients with essential hypertension. Hypertension 2011, 57, 79–84. [Google Scholar] [CrossRef]
- Schiffrin, E.L.; Pollock, D.M. Endothelin system in hypertension and chronic kidney disease. Hypertension 2024, 81, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Drolet, G.; Bouvier, M.; de Champlain, J. Enhanced sympathoadrenal reactivity to haemorrhagic stress in DOCA-salt hypertensive rats. J. Hypertens. 1989, 7, 237–242. [Google Scholar] [CrossRef]
- Lange, D.L.; Haywood, J.R.; Hinojosa-Laborde, C. Role of the adrenal medullae in male and female DOCA-salt hypertensive rats. Hypertension 1998, 31 1 Pt 2, 403–408. [Google Scholar] [CrossRef]
- Gómez-Guzmán, M.; Jiménez, R.; Sánchez, M.; Zarzuelo, M.J.; Galindo, P.; Quintela, A.M.; LópezSepúlveda, R; Romero, M.; Tamargo, J.; Vargas, F.; Pérez-Vizcaíno, F.; Duarte, J. Epicatechin lowers blood pressure, restores endothelial function, and decreases oxidative stress and endothelin-1 and NADPH oxidase activity in DOCA-salt hypertension. J. Free. Radic. Biol. Med. 2012, 52(1), 70–79. [Google Scholar] [CrossRef]
- Kohan, D.; Rossi, N.F.; Inscho, E.W.; Pollock, D.M. Regulation of blood pressure and salt homeostasis by endothelin. Physiol. Rev. 2011, 91, 1–77. [Google Scholar] [CrossRef]
- Kristensen, A.S.; Andersen, J.; Jørgensen, T.N.; Sørensen, L.; Eriksen, J.; Loland, C.J.; Strømgaard, K.; Gether, U. SLC6 Neurotransmitter transporters: Structure, function, and regulation. Pharmacol. Rev. 2011, 63, 585–640. [Google Scholar] [CrossRef]
- Bermingham, D.P.; Blakely, R.D. Kinase-dependent regulation of monoamine neurotransmitter transporters. Pharmacol. Rev. 2016, 68, 888–953. [Google Scholar] [CrossRef]
- Jayanthi, L.D.; Ramamoorthy, S. Role of phosphorylation of serotonin and norepinephrine transporters in animal behavior: Relevance to neuropsychiatric disorders. Int. J. Mol. Sci. 2025, 26, 7713. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, C.; Jordan, J. Norepinephrine transporter function and human cardiovascular disease. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H1273–H1282. [Google Scholar] [CrossRef]
- Wehrwein, E.A.; Novotny, M.; Swain, G.M.; Parker, L.M.; Esfahanian, M.; Spitsbergen, J.M.; Habecker, B.A.; Kreulen, D.L. Regional changes in cardiac and stellate ganglion norepinephrine transporter in DOCA-salt hypertension. Auton. Neurosci. 2013, 179(1-2), 99–107. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Iwai, C.; Qin, F.; Liang, C-S. Norepinephrine induces endoplasmic reticulum stress and downregulation of norepinephrine transporter density in PC12 cells via oxidative stress. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H2381–H2389. [Google Scholar] [CrossRef]
- Liang, C-S. Cardiac sympathetic nerve terminal function in congestive heart failure. Acta Pharmacol. Sin. 2007, 28(7), 921–927. [Google Scholar] [CrossRef]
- Hope, S.I.; Schmipp, J.; Rossi, A.H.; Bianciotti, L.G.; Vatta, M.S. Regulation of the neuronal norepinephrine transporter by endothelin-1 and -3 in the rat anterior and posterior hypothalamus. Neurochem. Int. 2008, 53, 207–213. [Google Scholar] [CrossRef]
- Abramoff, T.; Guil, M.J.; Morales, V.P.; Hope, S.I.; Soria, C.; Bianciotti, L.G.; Vatta, M.S. Enhanced assymetrical noradrenergic transmission in the olfactory bulb of deoxycorticosterone acetate-salt hypertensive rats. Neurochem. Res. 2013, 38, 2063–2071. [Google Scholar] [CrossRef]
- Abramoff, T.; Guil, M.J.; Morales, V.P.; Hope, S.I.; Höcht, C.; Bianciotti, L.G.; Vatta, M.S. Involvement of endothelins in deoxycorticosterone acetate–salt hypertension through the modulation of noradrenergic transmission in the rat posterior hypothalamus. Exp. Physiol. 2015, 100.6, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Guil, M.J.; Schöller, M.I.; Cassinotti, L.R.; Biancardi, V.C.; Pitra, S.; Bianciotti, L.G.; Stern, J.E.; Vatta, M.S. Role of endothelin receptor type A on catecholamine regulation in the olfactory bulb of DOCA-salt hypertensive rats: Hemodynamic implications. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 165527. [Google Scholar] [CrossRef]
- Cassinotti, L.; Guil, M.; Bianciotti, L.; Vatta, M. Role of brain endothelin receptor type B (ETB) in the regulation of tyrosine hydroxylase in the olfactory bulb of DOCA-salt hypertensive rats. Curr. Vasc. Pharmacol. 2023, 21(4), 246–256. [Google Scholar] [CrossRef] [PubMed]
- Uchino, T.; Sanyal, S.N.; Yamane, M.; Kaku, T.; Takebashi, S.; Shimaoka, T.; Shimada, T.; Noguchi, T.; Ono, K. Rescue of Pulmonary Hypertension with an Oral Sulfonamide Antibiotic Sulfisoxazole by Endothelin Receptor Antagonistic Actions. Hypertens. Res. 2008, 31, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- De Champlain, J.; Van Ameringen, MR. Regulation of blood pressure by sympathetic nerve fibers and adrenal medulla in normotensive and hypertensive rats. Circ. Res. 1972, 31(4), 617–628. [Google Scholar] [CrossRef]
- Reims, H.M.; Fossum, E.; Høieggen, A.; Moan, A.; Eide, I.; Kjeldsen, S.E. Adrenal medullary overactivity in lean, borderline hypertensive young men. Am. J. Hypertens. 2004, 17(7), 611–618. [Google Scholar] [CrossRef]
- Quang, V.T.; Hammes, S.R. Recent Insights on Circulating Catecholamines in Hypertension. Curr. Hypertens. Rep. 2014, 16, 498. [Google Scholar] [CrossRef]
- Currie, G.; Freel, E.M.; Perry, C.G.; Dominiczak, A.F. Disorders of blood pressure regulation-role of catecholamine biosynthesis, release, and metabolism. Curr. Hypertens. Rep. 2012, 14, 38–45. [Google Scholar] [CrossRef]
- Lange, D.L.; Haywood, J.R.; Hinojosa-Laborde, C. Endothelin enhances and inhibits adrenal catecholamine release in deoxycorticosterone acetate-salt hypertensive rats. Hypertension 2000, 35 1 Pt 2, 385–390. [Google Scholar] [CrossRef]
- Wang, D.S.; Xie, H.H.; Shen, F.M.; Cai, G.J.; Su, D.F. Blood pressure variability, cardiac baroreflex sensitivity and organ damage in experimentally hypertensive rats. Clin. Exp. Pharmacol. Physiol. 2005, 32(7), 545–52. [Google Scholar] [CrossRef]
- Basting, T.; Lazartigues, E. DOCA-salt hypertension: an update. Curr. Hypertens. Rep. 2017, 19(4), 32. [Google Scholar] [CrossRef]
- Becker, B.K.; Johnston, J.G.; Young, C.M.; Torres Rodriguez, A.A.; Jin, C.; Pollock, D.M. Endothelin B receptors impair baroreflex function and increase blood pressure variability during high salt diet. Auton. Neurosci. Basic Clin. 2021, 232, 102796. [Google Scholar] [CrossRef] [PubMed]
- Hana, S.; Bala, N.B.; Sadib, G.; Usanmaz, S.E.; Tuglu, M.M.; Uludag, M.O.; Demirel-Yilmaz, E. Inhibition of endoplasmic reticulum stress protected DOCA-salt hypertension-induced vascular dysfunction. Vasc. Pharmacol. 2019, 113, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Burnier, M. Update on Endothelin Receptor Antagonists in Hypertension. Curr. Hypertens. Rep. 2018, 20, 51. [Google Scholar] [CrossRef] [PubMed]
- Kostov, K. The Causal Relationship between Endothelin-1 and Hypertension: Focusing on Endothelial Dysfunction, Arterial Stiffness, Vascular Remodeling, and Blood Pressure Regulation. Life 2021, 11, 986. [Google Scholar] [CrossRef]
- Boxwalla, M.; Matwyshyn, G.; Puppala, B.L.; Andurkar, S.V.; Gulati, A. Involvement of imidazoline and opioid receptors in the enhancement of clonidine-induced analgesia by sulfisoxazole. Can. J. Physiol. Pharmacol. 2010, 88(5), 541–552. [Google Scholar] [CrossRef]
- Syed, H.; Safa, R.; Chidlow, G.; Osbornem, N.N. Sulfisoxazole, an endothelin receptor antagonist, protects retinal neurones from insults of ischemia/reperfusion or lipopolysaccharide. Neurochem. Int. 2006, 48, 708–717. [Google Scholar] [CrossRef]
- Im, E-J; Lee, C-H; Moon, P-G; et al. Sulfisoxazole inhibits the secretion of small extracellular vesicles by targeting the endothelin receptor A. Nature Commun 2019, 10, 1387. [Google Scholar] [CrossRef]
- Backs, J; Bresch, E; Lutz, M; Kristen, AV; Haass, M. Endothelin-1 inhibits the neuronal norepinephrine transporter in hearts of male rats. Cardiovasc Res 2005, 67(2), 283–290. [Google Scholar] [CrossRef]
- Young, CN. Endoplasmic reticulum stress in the pathogenesis of hypertension. Exp Physiol 2017, 102.8, 869–884. [Google Scholar] [CrossRef]
- Bala, NB; Hana, S; Kiremitcib, S; Sadic, G; Uludaga, O; Demirel-Yilmazd, E. Hypertension-induced cardiac impairment is reversed by the inhibition of endoplasmic reticulum stress. J Pharm Pharmacol 2019, 71, 1809–1821. [Google Scholar] [CrossRef] [PubMed]
- Bala, NB; Han, S; Kiremitci, S; Uludag, MO; Demirel-Yilmaz, E. Reversal of deleterious effect of hypertension on the liver by inhibition of endoplasmic reticulum stress. Mol Biol Rep 2020, 47, 2243–2252. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y-H; Zheng, C-M; Chou, Ch-L; et al. Therapeutic effect of endothelin-converting enzyme inhibitor on chronic kidney disease through the inhibition of endoplasmic reticulum stress and the NLRP3 inflammasome. Biomedicines 2021, 9, 398. [Google Scholar] [CrossRef] [PubMed]
- Jain, A. Endothelin-1–induced endoplasmic reticulum stress in disease. J Pharmacol Exp Ther 2013, 346, 163–172. [Google Scholar] [CrossRef]
- Harding, HP; Zhang, Y; Ron, D. Protein translation and folding are coupled by an endoplasmic-reticulum-resident kinase. Nature 1999, 397, 271–274. [Google Scholar] [CrossRef]
- Hong, J; Kim, K; Kim, J-H; Park, Y. The role of endoplasmic reticulum stress in cardiovascular disease and exercise. Int J Vasc Med 2017, 2049217. [Google Scholar] [CrossRef]
- Wang, S; Binder, P; Fang, Q; et al. Endoplasmic reticulum stress in the heart: insights into mechanisms and drug targets. Br J Pharmacol 2018, 175, 1293–1304. [Google Scholar] [CrossRef] [PubMed]
- Liu, S; Ding, H; Li, Y; Zhang, X. Molecular mechanism underlying the role of the XBP1s in cardiovascular diseases. J Cardiovasc Dev Dis 2022, 9, 459. [Google Scholar] [CrossRef] [PubMed]
- Dufey, E; Sepúlveda, D; Rojas-Rivera, D; Hetz, C. Cellular mechanisms of endoplasmic reticulum stress signaling in health and disease. 1. An overview. Am J Physiol Cell Physiol 2017, 307, C582–C594. [Google Scholar] [CrossRef]
Figure 1.
Experimental design. SFX: sulfisoxazole.
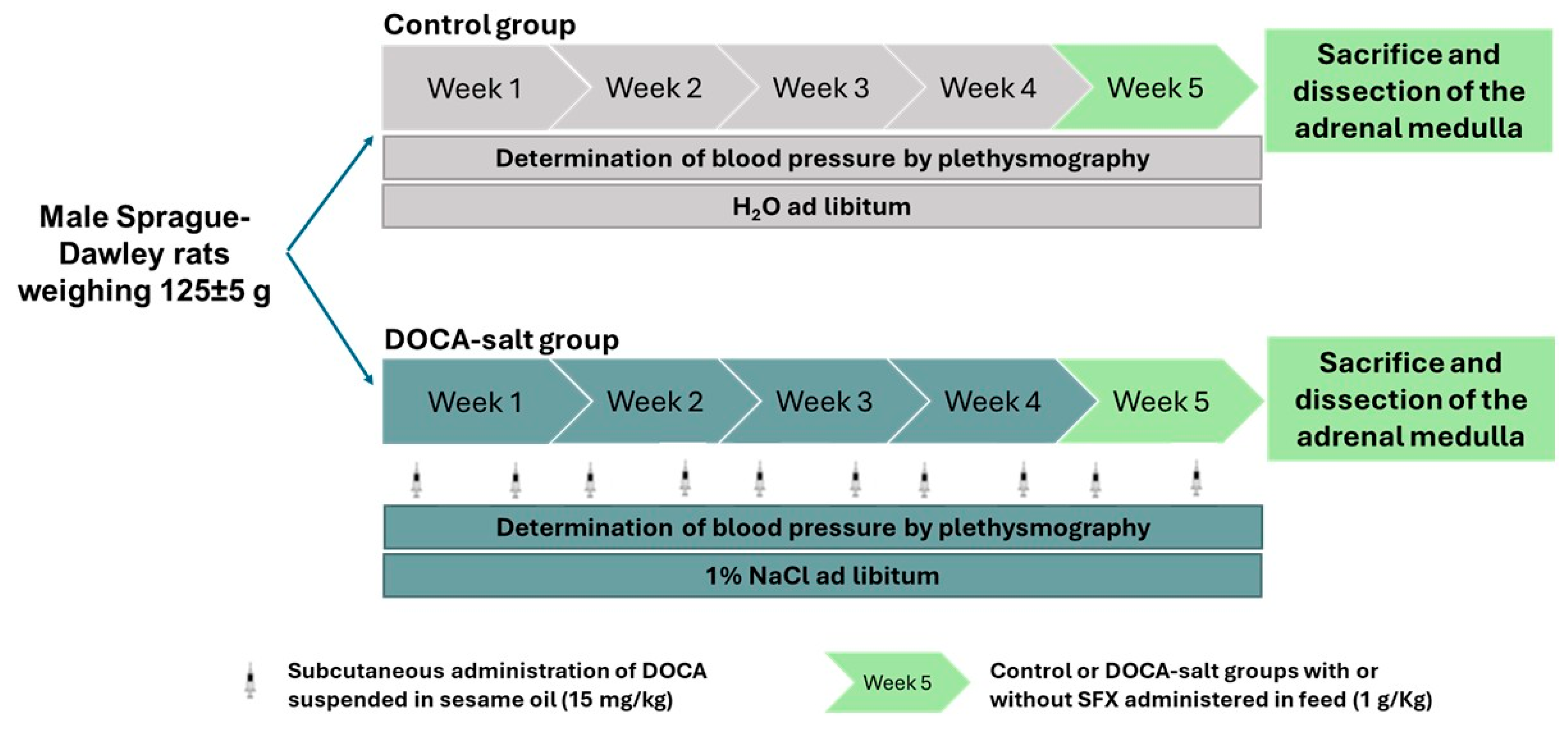
Figure 2.
Systolic blood pressure (SBP) and heart rate (HR) following sulfisoxazole (SFX, 1g/Kg) administration to normotensive and DOCA-salt hypertensive rats. SBP and HR were determined by tail-cuff plethysmography as described in Materials and Methods and expressed as mmHg ± SD and beats per minute (BPM) ± SD, respectively. A) Time course of SBP following SFX treatment: ***: p<0.001 vs. control; †††: p<0.001 vs. DOCA-salt; ‡: p<0.05, ‡‡: p<0.01 and ‡‡‡: p<0.001 vs. SFX. B) SBP average. Each bar represents the area under each curve (shown in A) between 0 and 168 h. ***: p<0.001 vs. control; †††: p<0.001 vs. DOCA-salt; ‡‡‡: p<0.001 vs. SFX. C) Time course of HR following SFX treatment. D) HR Average. Each bar represents the area under each curve (shown in C) between 0 and 168 h. Number of animals: 22 per experimental group.
Figure 2.
Systolic blood pressure (SBP) and heart rate (HR) following sulfisoxazole (SFX, 1g/Kg) administration to normotensive and DOCA-salt hypertensive rats. SBP and HR were determined by tail-cuff plethysmography as described in Materials and Methods and expressed as mmHg ± SD and beats per minute (BPM) ± SD, respectively. A) Time course of SBP following SFX treatment: ***: p<0.001 vs. control; †††: p<0.001 vs. DOCA-salt; ‡: p<0.05, ‡‡: p<0.01 and ‡‡‡: p<0.001 vs. SFX. B) SBP average. Each bar represents the area under each curve (shown in A) between 0 and 168 h. ***: p<0.001 vs. control; †††: p<0.001 vs. DOCA-salt; ‡‡‡: p<0.001 vs. SFX. C) Time course of HR following SFX treatment. D) HR Average. Each bar represents the area under each curve (shown in C) between 0 and 168 h. Number of animals: 22 per experimental group.
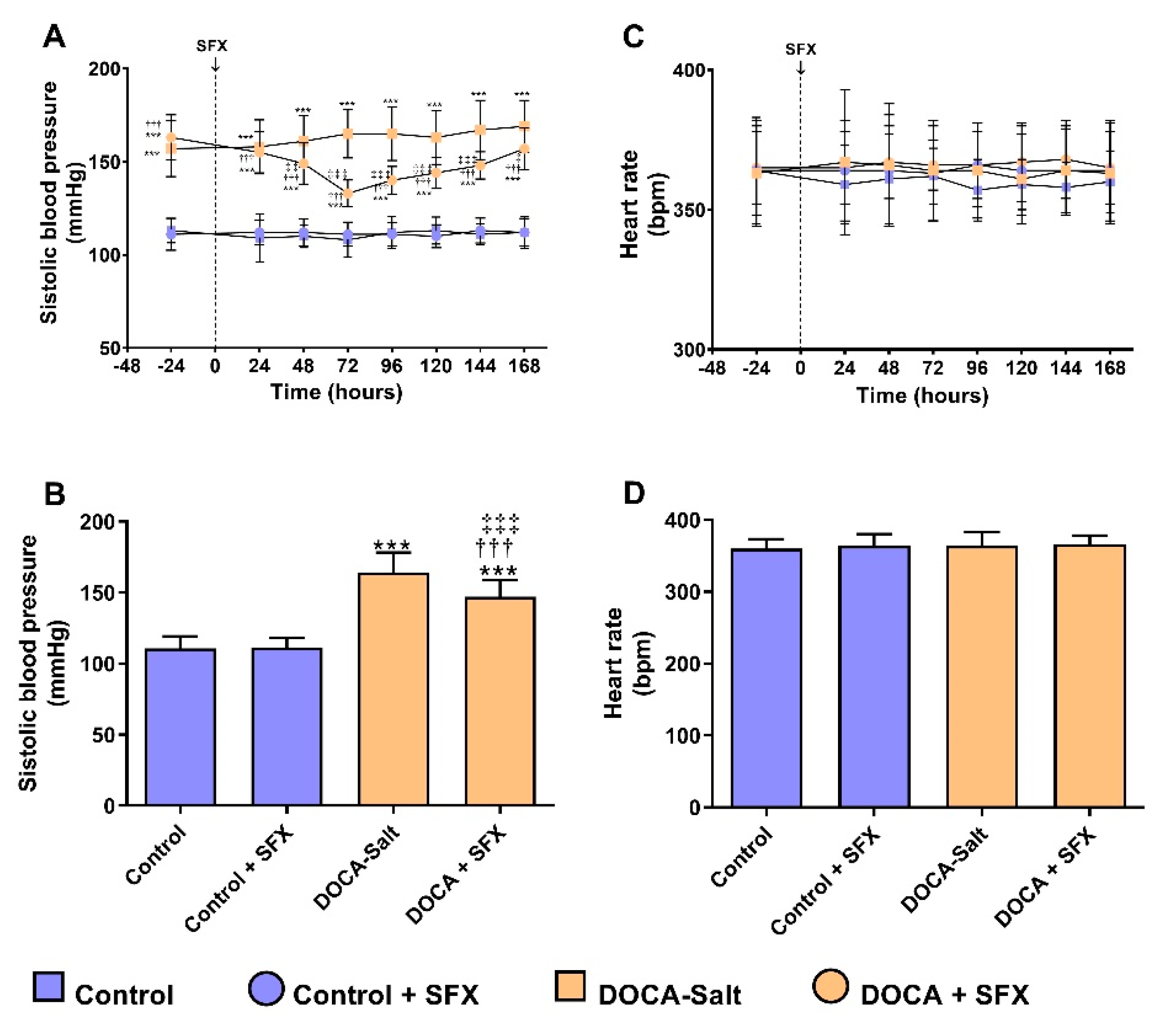
Figure 3.
Neuronal norepinephrine transporter (NET) activity and expression following sulfisoxazole (SFX, 1g/Kg) administration to normotensive and DOCA-salt hypertensive rats. A) NET activity. The transporter activity was evaluated by neuronal norepinephrine uptake as described in Materials and Methods. **: p<0.01 vs. control; †: p<0.05 vs. DOCA-salt. B) Representative NET immunoblots. C) Protein expression of functional NET (glycosylated form). NET expression was determined by Western blot as described in Materials and Methods. ***: p<0.001 vs. control; ††: p<0.01 vs. DOCA-salt. D) Levels of mRNA NET. The mRNA expression was measured by RT-qPCR as described in Materials and Methods. **: p<0.01 and ***: p<0.001 vs. control; †: p<0.05 vs. DOCA-salt. Results were expressed as the mean ± SD. Number of experiments: 3-5.
Figure 3.
Neuronal norepinephrine transporter (NET) activity and expression following sulfisoxazole (SFX, 1g/Kg) administration to normotensive and DOCA-salt hypertensive rats. A) NET activity. The transporter activity was evaluated by neuronal norepinephrine uptake as described in Materials and Methods. **: p<0.01 vs. control; †: p<0.05 vs. DOCA-salt. B) Representative NET immunoblots. C) Protein expression of functional NET (glycosylated form). NET expression was determined by Western blot as described in Materials and Methods. ***: p<0.001 vs. control; ††: p<0.01 vs. DOCA-salt. D) Levels of mRNA NET. The mRNA expression was measured by RT-qPCR as described in Materials and Methods. **: p<0.01 and ***: p<0.001 vs. control; †: p<0.05 vs. DOCA-salt. Results were expressed as the mean ± SD. Number of experiments: 3-5.
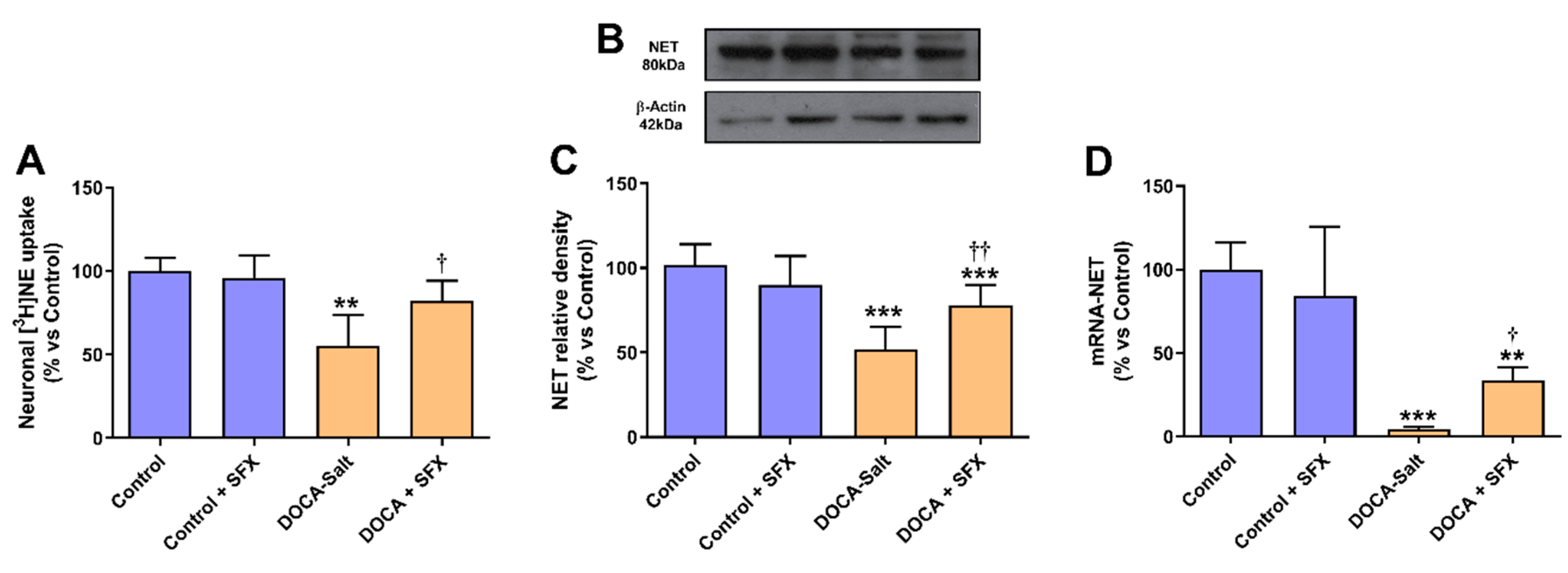
Figure 4.
Ultrastructural changes in the adrenal medulla following sulfisoxazole (SFX) administration. SFX prevents most of the ultrastructural changes in the chromaffin cells of hypertensive DOCA-Salt rats. Representative transmission electron micrographs of chromaffin cells derived from control animals (A), SFX (B), DOCA-salt (C), DOCA-salt + SFX (D). Number of experiments: 3.
Figure 4.
Ultrastructural changes in the adrenal medulla following sulfisoxazole (SFX) administration. SFX prevents most of the ultrastructural changes in the chromaffin cells of hypertensive DOCA-Salt rats. Representative transmission electron micrographs of chromaffin cells derived from control animals (A), SFX (B), DOCA-salt (C), DOCA-salt + SFX (D). Number of experiments: 3.
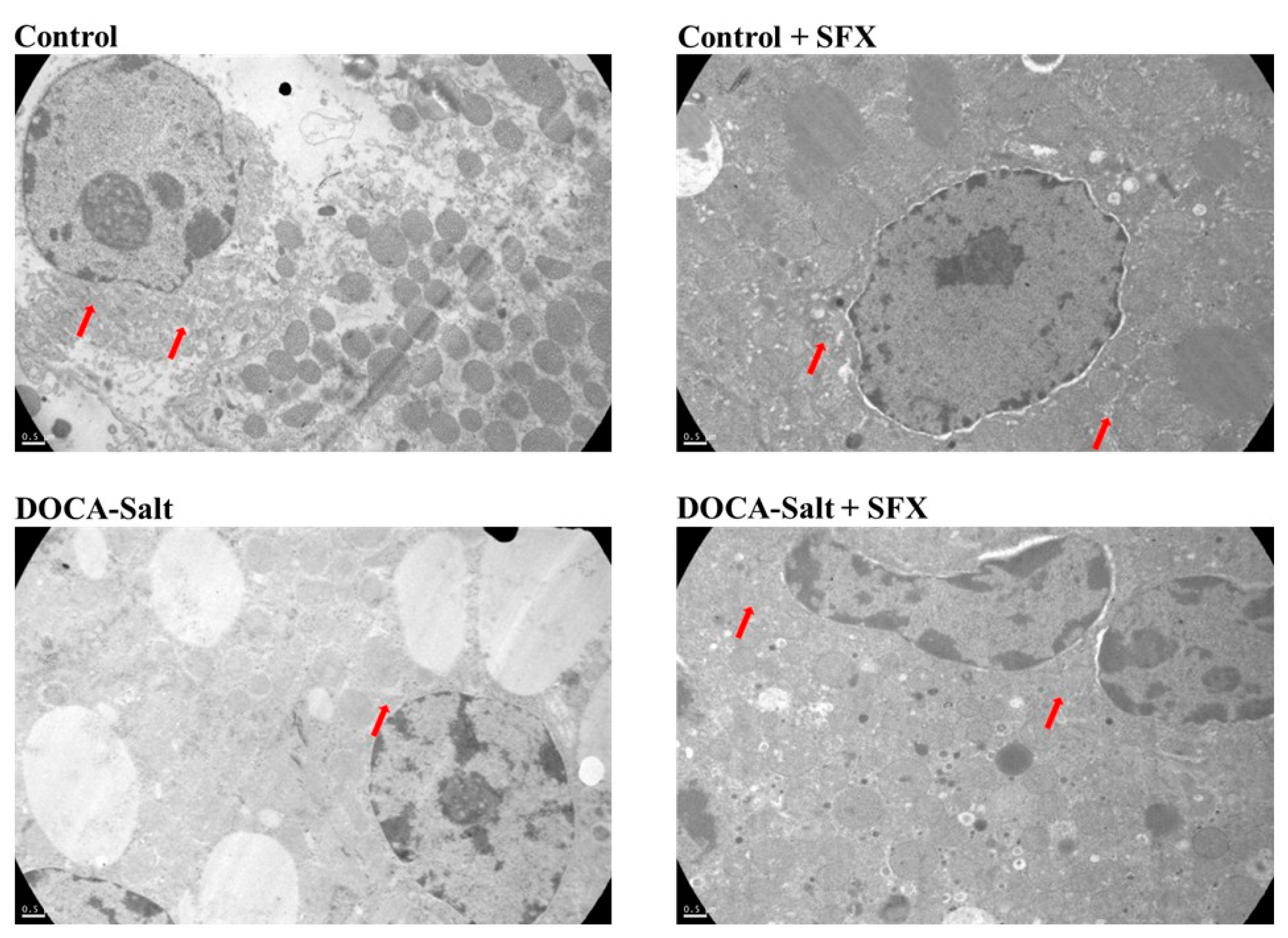
Figure 5.
Bip expression following sulfisoxazole (SFX, 1g/Kg) administration to normotensive and DOCA-salt hypertensive rats. Bip protein expression was determined by Western blot whereas mRNA Bip by RT-qPCR as detailed in Materials and Methods. A) Representative immunoblots. B) Bip protein expression. **: p<0.01 vs. control; ††: p<0.01 vs. DOCA-salt. C) Levels of mRNA Bip. ***: p<0.001 vs. control; †††: p<0.001 vs. DOCA-salt. Results were expressed as the mean ± SD. Number of experiments: 3.
Figure 5.
Bip expression following sulfisoxazole (SFX, 1g/Kg) administration to normotensive and DOCA-salt hypertensive rats. Bip protein expression was determined by Western blot whereas mRNA Bip by RT-qPCR as detailed in Materials and Methods. A) Representative immunoblots. B) Bip protein expression. **: p<0.01 vs. control; ††: p<0.01 vs. DOCA-salt. C) Levels of mRNA Bip. ***: p<0.001 vs. control; †††: p<0.001 vs. DOCA-salt. Results were expressed as the mean ± SD. Number of experiments: 3.
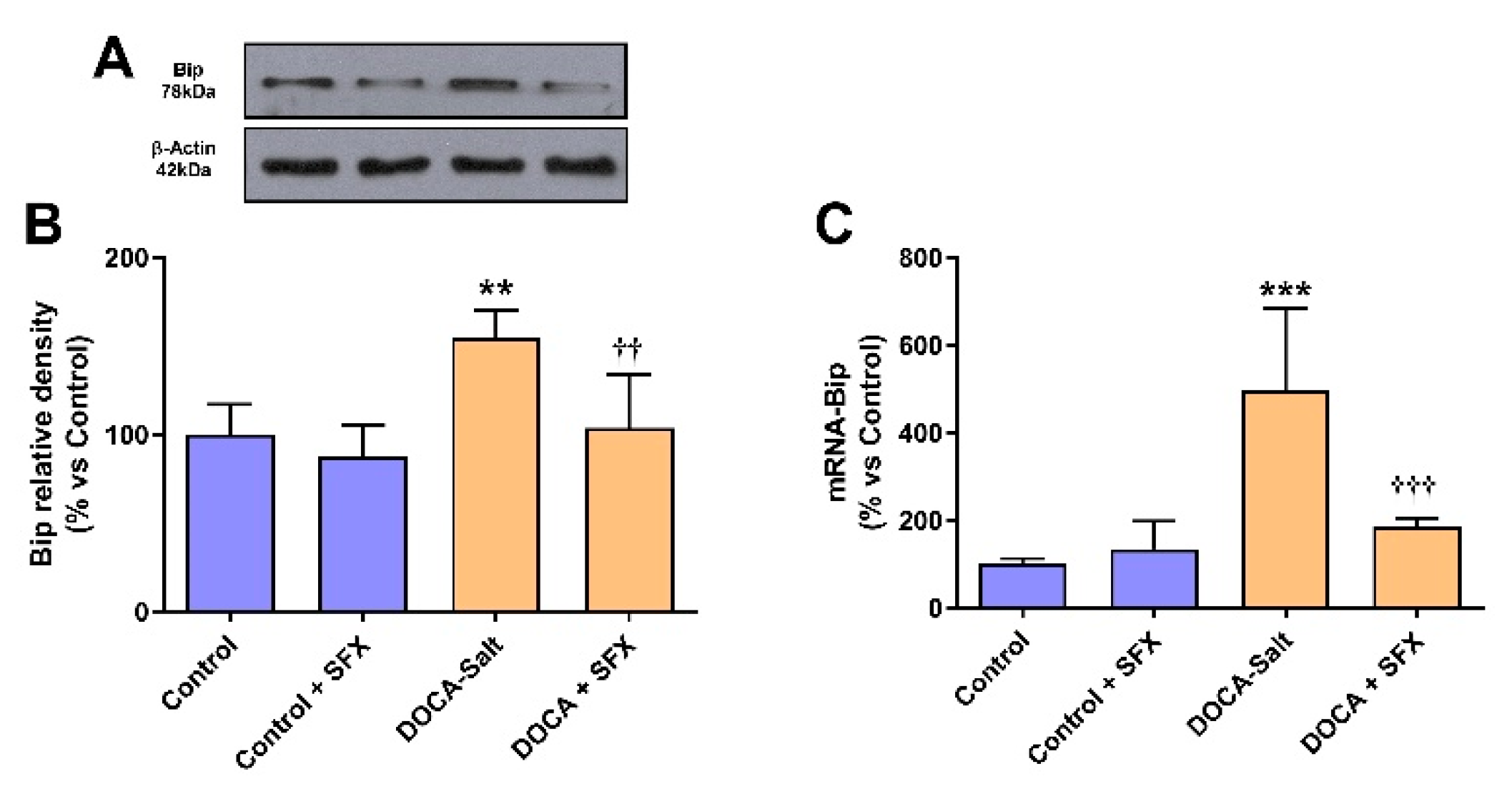
Figure 6.
PERK, eIF2α and CHOP expression following sulfisoxazole (SFX, 1g/Kg) administration to normotensive and DOCA-salt hypertensive rats. Protein expression was determined by Western blot whereas mRNA expression by RT-qPCR as detailed in Materials and Methods. A) Representative immunoblots. B) Phosphorylated PERK protein expression. **: p<0.01 and ***: p<0.001 and vs. control; ††: p<0.01 vs. DOCA-salt; ‡: p<0.05 vs. control + SFX. C) Determination of eIF2α phosphorylation. It was evaluated by the ratio between phosphorylated eIF2α and total protein (p-eIF2α/t-eIF2α) as described in Materials and Methods. ** p<0.01 vs. control; †† p<0.01 vs. DOCA-salt. D) CHOP protein expression. **: p<0.01 and ***: p<0.001 and vs. control; †††: p<0.001 vs. DOCA-salt. E) Levels of mRNA CHOP. *: p<0.05 and ***: p<0.001 vs. control; †††: p<0.001 vs. DOCA-salt; ‡: p<0.05 vs. control + SFX. Results were expressed as the mean ± SD. Number of experiments: 3-5.
Figure 6.
PERK, eIF2α and CHOP expression following sulfisoxazole (SFX, 1g/Kg) administration to normotensive and DOCA-salt hypertensive rats. Protein expression was determined by Western blot whereas mRNA expression by RT-qPCR as detailed in Materials and Methods. A) Representative immunoblots. B) Phosphorylated PERK protein expression. **: p<0.01 and ***: p<0.001 and vs. control; ††: p<0.01 vs. DOCA-salt; ‡: p<0.05 vs. control + SFX. C) Determination of eIF2α phosphorylation. It was evaluated by the ratio between phosphorylated eIF2α and total protein (p-eIF2α/t-eIF2α) as described in Materials and Methods. ** p<0.01 vs. control; †† p<0.01 vs. DOCA-salt. D) CHOP protein expression. **: p<0.01 and ***: p<0.001 and vs. control; †††: p<0.001 vs. DOCA-salt. E) Levels of mRNA CHOP. *: p<0.05 and ***: p<0.001 vs. control; †††: p<0.001 vs. DOCA-salt; ‡: p<0.05 vs. control + SFX. Results were expressed as the mean ± SD. Number of experiments: 3-5.
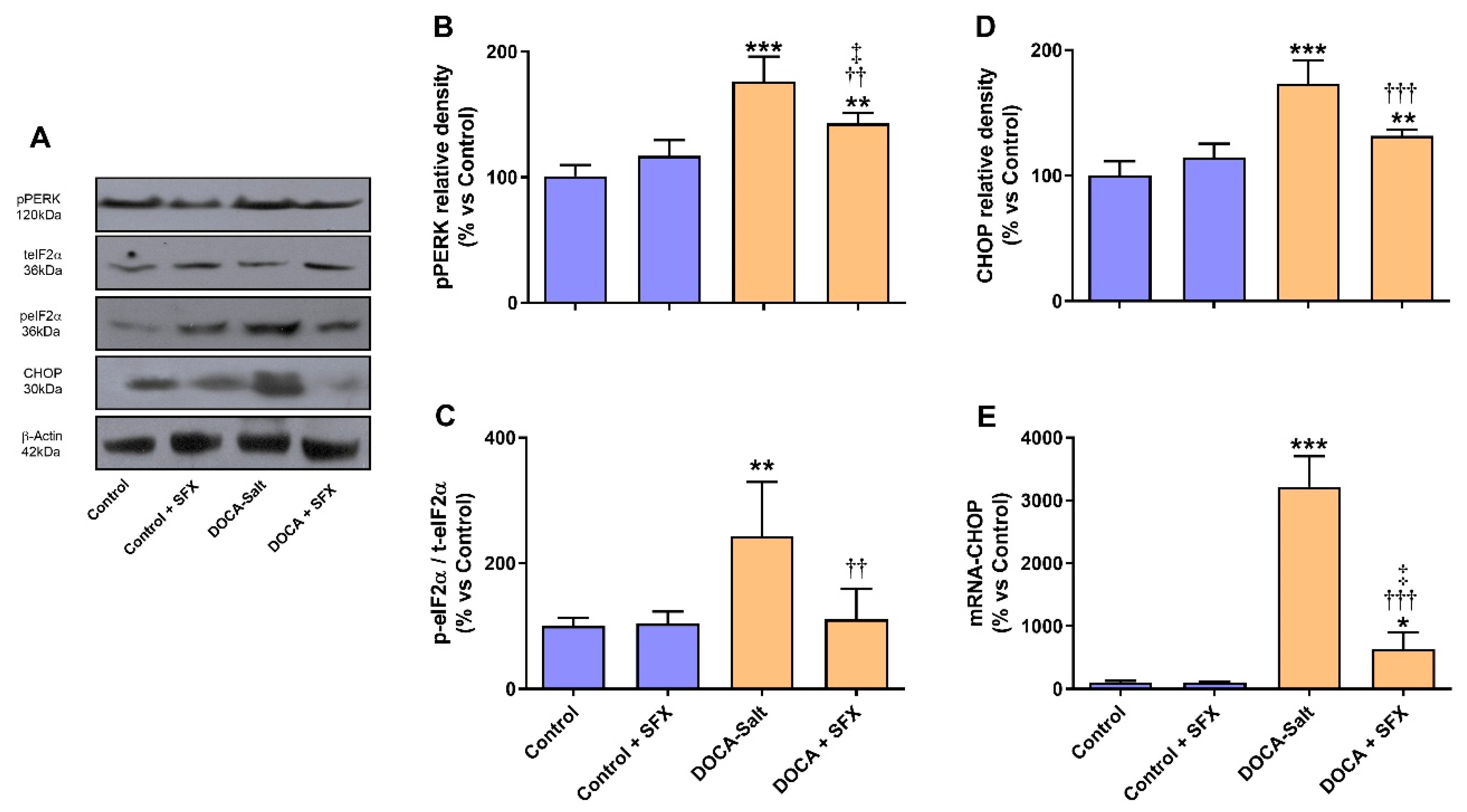
Figure 7.
IRE and Xbp1 protein expression following sulfisoxazole (SFX, 1g/Kg) administration to normotensive and DOCA-salt hypertensive rats. Protein expression was determined by Western blot as detailed in Materials and Methods. A) Representative immunoblots. B) IRE protein expression. **: p<0.01 vs. control; ††: p<0.01 vs. DOCA-salt. C) Xbp1protein expression. **: p<0.01 vs. control; ††: p<0.01 vs. DOCA-salt. Results were expressed as the mean ± SD. Number of experiments: 3.
Figure 7.
IRE and Xbp1 protein expression following sulfisoxazole (SFX, 1g/Kg) administration to normotensive and DOCA-salt hypertensive rats. Protein expression was determined by Western blot as detailed in Materials and Methods. A) Representative immunoblots. B) IRE protein expression. **: p<0.01 vs. control; ††: p<0.01 vs. DOCA-salt. C) Xbp1protein expression. **: p<0.01 vs. control; ††: p<0.01 vs. DOCA-salt. Results were expressed as the mean ± SD. Number of experiments: 3.
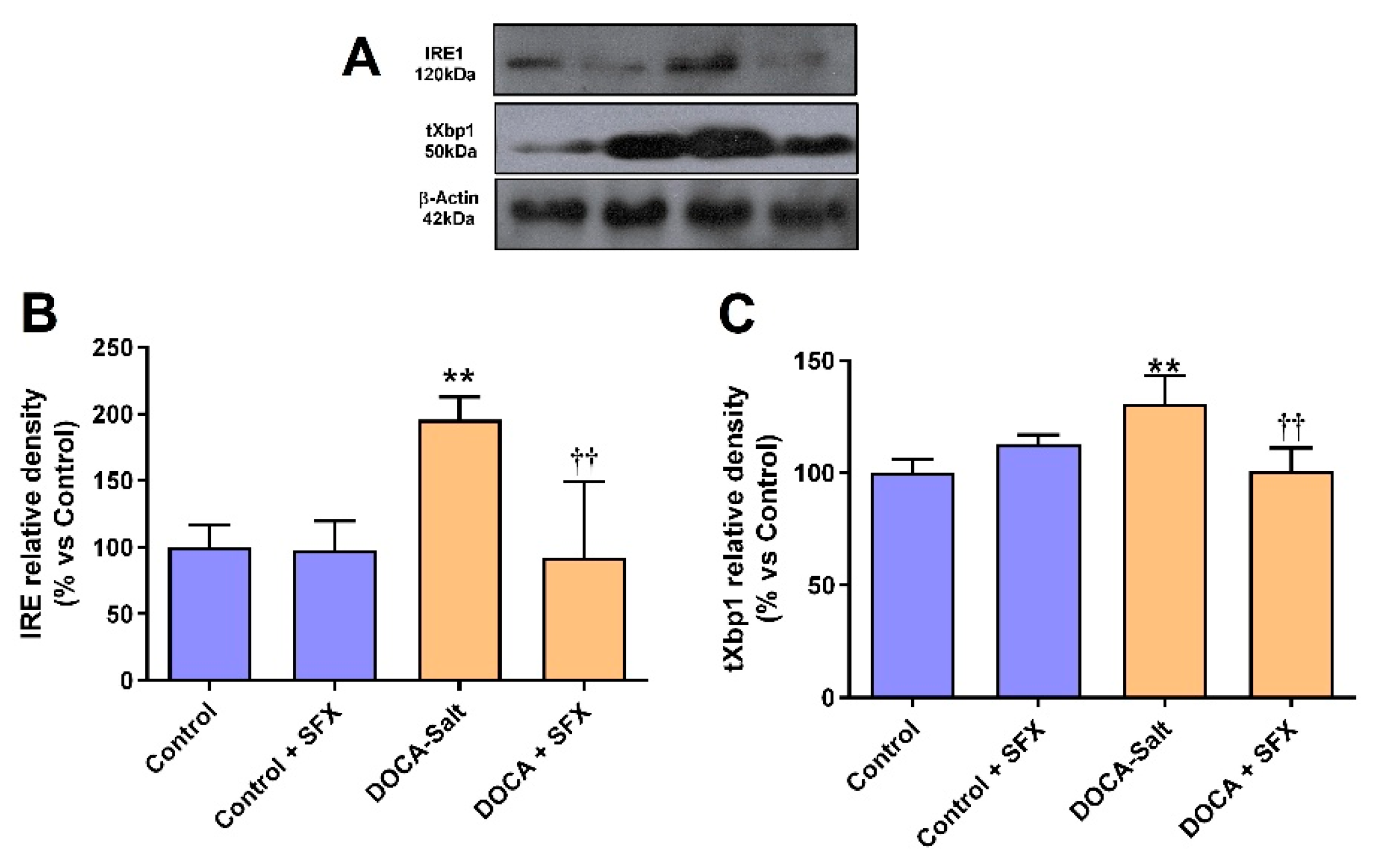
Figure 8.
Evaluation of apoptosis by tunnel assay following sulfisoxazole (SFX, 1g/Kg) administration to normotensive and DOCA-salt hypertensive rats. A) Representative microscopy images. B) Quantitative analysis of TUNEL positive cells. ***: p<0.001 vs. control; †††: p<0.01 vs. DOCA-salt. Number of experiments: 4.
Figure 8.
Evaluation of apoptosis by tunnel assay following sulfisoxazole (SFX, 1g/Kg) administration to normotensive and DOCA-salt hypertensive rats. A) Representative microscopy images. B) Quantitative analysis of TUNEL positive cells. ***: p<0.001 vs. control; †††: p<0.01 vs. DOCA-salt. Number of experiments: 4.
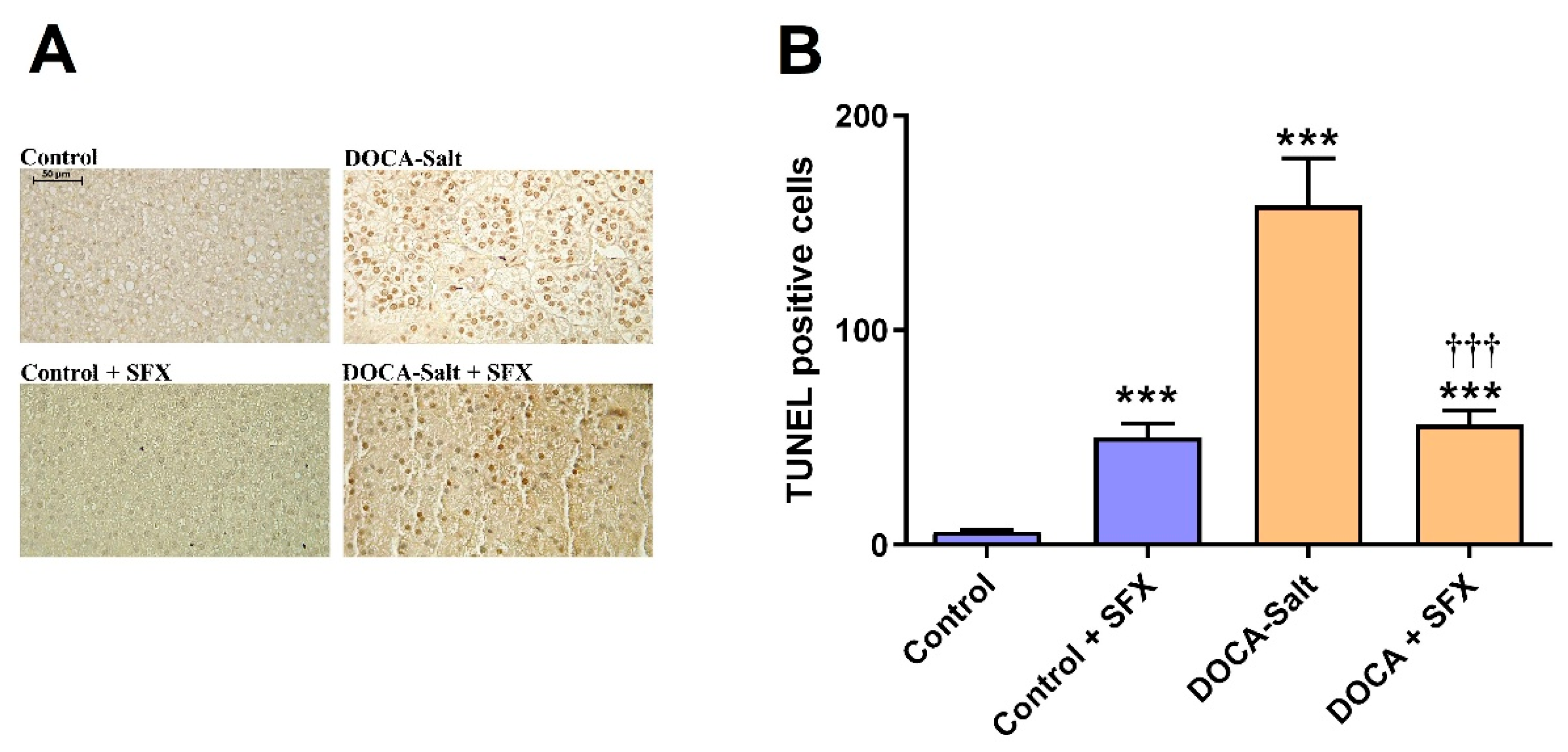

Table 1.
Primary and secondary antibodies. Details of the antibodies used for Western blot assays. All secondary antibodies are HRP-conjugated.
Table 1.
Primary and secondary antibodies. Details of the antibodies used for Western blot assays. All secondary antibodies are HRP-conjugated.
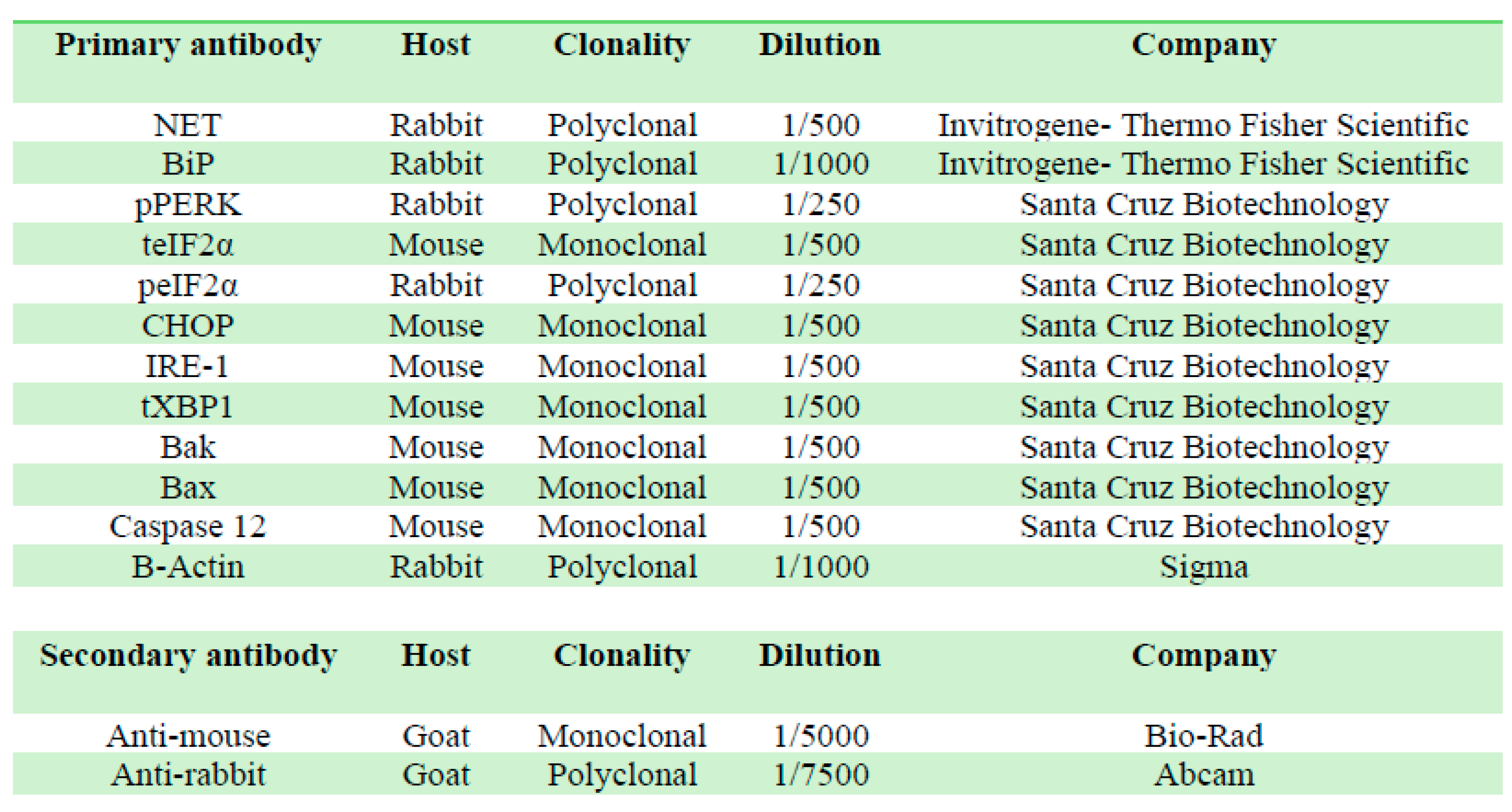 |
Table 2.
Primers sequence of NET, BiP, CHOP, caspase 12 and GAPDH in RT-qPCR.
 |
Table 3.
Real-time RT-PCR programs. Run settings used in real-time RT-PCR for the amplification of NET, BiP, CHOP, caspase 12 and GAPDH cDNA.
Table 3.
Real-time RT-PCR programs. Run settings used in real-time RT-PCR for the amplification of NET, BiP, CHOP, caspase 12 and GAPDH cDNA.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



