
Submitted:
08 January 2026
Posted:
09 January 2026
You are already at the latest version
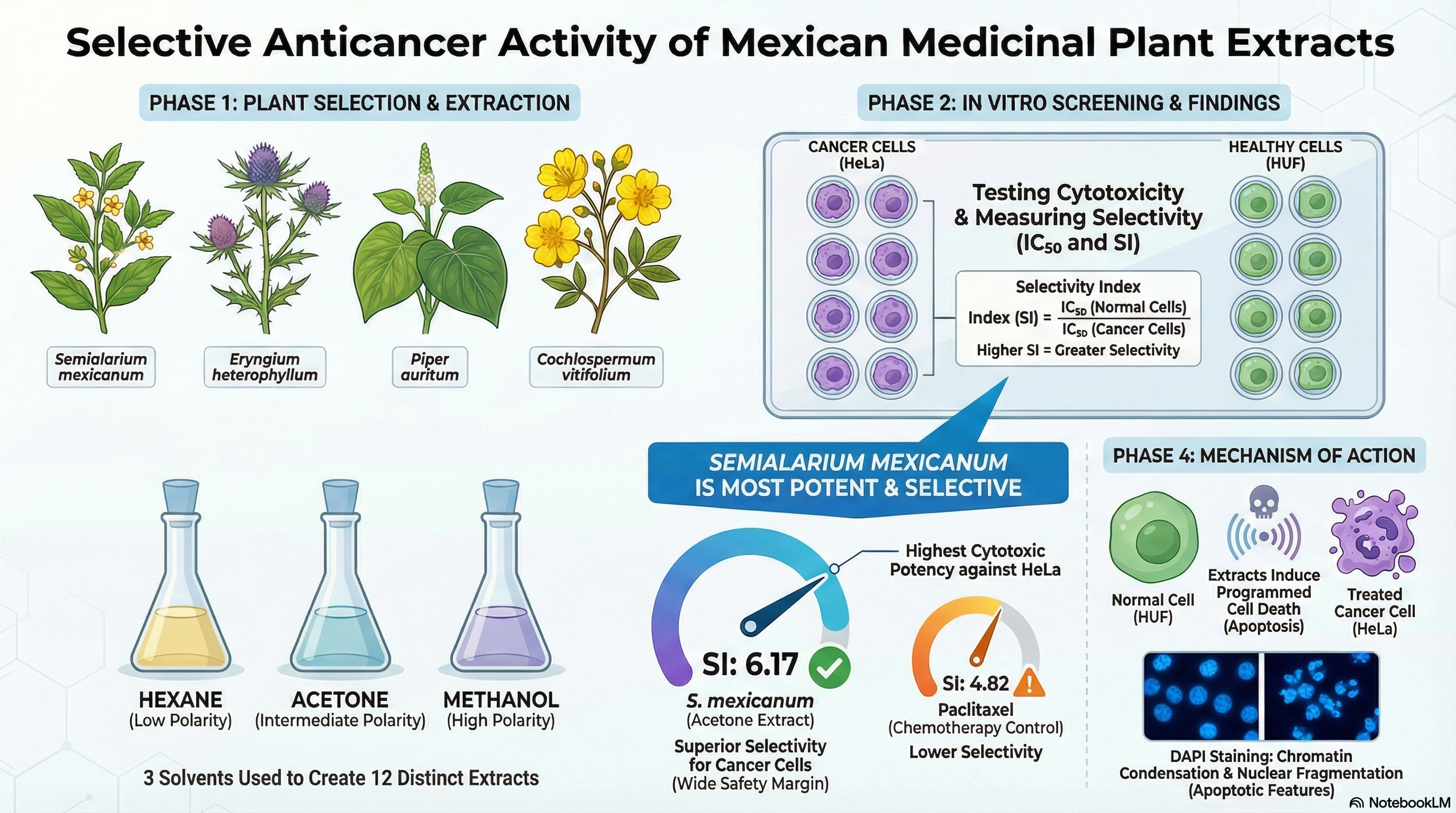
Abstract
Background: Medicinal plants used in traditional Mexican medicine represent a valu-able source of bioactive compounds with potential anticancer activity. Beyond cyto-toxic potency, selectivity toward cancer cells over normal cells is a critical toxicologi-cal parameter for identifying safer therapeutic candidates. This study aimed to evalu-ate the selective cytotoxic and antiproliferative effects of extracts from four Mexican medicinal plants across human cancerous and non-cancerous cell lines. Methods: Hexane, acetone, and methanolic extracts from Semialarium mexicanum, Eryngium heterophyllum, Piper auritum, and Cochlospermum vitifolium were evaluated in a panel of human cancer cell lines and non-tumoral models, including primary human uterine fibroblasts (HUF). Cytotoxicity was assessed after 48 h of treatment using in-creasing extract concentrations, and selectivity indices were calculated. Cell cycle dis-tribution and nuclear morphology analyses were performed to explore antiprolifera-tive effects. Additionally, GC–MS-based chemical profiling was conducted on selected extracts to obtain a tentative characterization of major bioactive constituents. Results: The extracts exhibited differential cytotoxic profiles depending on plant spe-cies and solvent polarity. Semialarium mexicanum, particularly its hexane extract, showed the highest cytotoxic potency and selectivity toward HeLa cervical cancer cells, achieving selectivity indices higher than those observed for paclitaxel when compared with HUF cells. Hexane extracts of Semialarium mexicanum and Eryngium heterophyllum, were generally more active than polar extracts, whereas Piper auritum displayed limited cytotoxicity and Cochlospermum vitifolium showed moderate, sol-vent-dependent effects. Cell cycle perturbations and nuclear alterations supported an antiproliferative response. Chemical profiling suggested the presence of lipophilic triterpenoid-related compounds in non-polar extracts and phenolic constituents in polar fractions. Conclusions: These findings provide in vitro evidence of selective anticancer activity of Mexican medicinal plant extracts and establish a basis for future mechanistic stud-ies medicinal plant extracts and lay the groundwork for future mechanistic investigations.
Keywords:
Mexican medicinal plants
; selective cytotoxicity
; antiproliferative activity
; Semialarium mexicanum
; natural products
; cancer cell lines
1. Introduction
Traditional Mexican Medicine (TMM) represents an ancestral therapeutic system deeply rooted in the indigenous, mestizo, and rural cultures of Mexico. This medical tradition is primarily based on the empirical use of medicinal plants, often integrated with ritual and cultural practices, and has been formally recognized by the World Health Organization as a fundamental component of biocultural heritage and a valuable source of bioactive compounds with pharmacological potential [1,2,3,4,5].
Mexico is considered one of the world’s most important reservoirs of medicinal plant diversity. To date, more than 4,000 plant species with documented medicinal use have been reported, of which at least 300 are traditionally employed to treat conditions associated with inflammation, tumors, “lumps,” cancer-like disorders, and hematological or reproductive diseases.1−4 Many of these species belong to genera with wide geographical distribution and longstanding ethnomedical relevance, including Bursera, Eryngium, Hippocratea excelsa, (syn. Semialarium mexicanum), Cochlospermum, Justicia, Artemisia (e.g., A. ludoviciana), and Piper [1,2,3,4,5,6,7,8,9].
Despite their extensive traditional use, systematic scientific evidence supporting the cytotoxic and antiproliferative activities of many Mexican medicinal plants remains limited. This gap has driven a growing interest in their experimental evaluation using in vitro cancer models, aimed at identifying extracts or metabolites with potential antitumoral properties [6]. The renewed global interest in natural products over recent decades is largely attributable to their remarkable chemical diversity, particularly their richness in secondary metabolites such as terpenoids, flavonoids, alkaloids, lignans, and quinones. These compounds are known to modulate key biological processes involved in tumor progression, including cell proliferation, apoptosis, oxidative stress, and oncogenic signaling pathways [10].
Notably, it has been estimated that approximately 60% of currently approved anticancer drugs are derived directly or indirectly from natural products, underscoring the critical importance of rigorously investigating medicinal plants with documented traditional use as potential sources of novel anticancer agents [10]. In this context, the systematic evaluation of Mexican medicinal plant extracts represents a scientifically justified and culturally relevant strategy for the discovery of selective cytotoxic and antiproliferative compounds.
2. Results
2.1. Plant Material
The four species are widely distributed in Mexico. According to the Global Biodiversity Information Facility (GBIF; Figure 1), C. vitifolium has 414 occurrence records in the country (GBIFa), E. heterophyllum 428 (GBIFb), P. auritum 1,824 (GBIFc), and S. mexicanum 542 (GBIFd) [11,12,13,14]. The specimens were collected in rural and semi-urban areas of the states of Morelos, Guerrero, and the State of Mexico (Figure 1). C. vitifolium was collected in Temixco, Morelos (18.8606 N, 99.2319 W); E. heterophyllum in Ixtapan de la Sal, State of Mexico (18.8903 N, 99.6733 W); P. auritum in Yautepec, Morelos (18.8917 N, 99.1208 W); and S. mexicanum in Iguala, Guerrero (18.4031 N, 99.5331 W; Figure 1).
The taxonomic identification was carried out by the specialist, Master of Science Gabriel Flores Franco, and a representative sample of each species was deposited in the Herbarium of the Autonomous University of the State of Morelos (HUMO-UAEM), where the corresponding registration codes were assigned, as shown in Table 1.
2.1.1. Plant Material Extraction
Twelve extracts were obtained from the plants C. vitifolium, E. heterophyllum, P. auritum and S. mexicanum, organic solvents with ascending polarity were used (hexane, acetone, and methanol). The extracts were prepared from 150 g of ground plant material. The yields obtained from the extracts are shown in Table 2.
The methanolic extract of P. auritum had the highest yield at 16.2%, followed by the methanolic extract of E. heterophyllum (9.81%). On the other hand, the hexane extract of C. vitifolium showed the lowest yield, followed by S. mexicanum (0.49% and 1.14%, respectively).
2.1.2. Total Polyphenol Content
The total polyphenol content of the extracts was determined using the Folin–Ciocalteu method and expressed as milligrams of gallic acid equivalents per gram of extract (mg GAE/g). The results revealed significant differences among the analyzed species and the solvents used Table 3.
The acetonic extract of C. vitifolium (CA) exhibited the highest concentration of phenolic compounds (231.19 ± 2.00 mg GAE/g), followed by the acetonic and methanolic extracts of S. mexicanum (SA and SM), with 205.98 ± 2.87 and 180.06 ± 1.55, respectively. In contrast, the methanolic and acetonic extracts of E. heterophyllum (EM, EA) and P. auritum (PM), as well as the methanolic extract of C. vitifolium (CM), showed substantially lower concentrations, ranging from 33.78 to 56.51 mg GAE/g.
In general, the extracts obtained with acetone showed higher phenolic concentrations compared to those obtained with methanol, suggesting that compounds of intermediate polarity predominate in the extract composition of these species. This result is consistent with previous reports indicating that solvents of intermediate polarity favor the recovery of these metabolites [15]. The difference was particularly notable in C. vitifolium, where the acetonic extract (CA) exhibited a significantly higher yield than its methanolic counterpart (CM).
An exception was P. auritum, whose methanolic extract (PM) showed a better yield than the acetonic one (PA). This finding suggests that the dominant phenolic fraction in P. auritum consists of more polar compounds, which are better solubilized in methanol. Overall, these results confirm that both plant species and solvent polarity are determining factors in the efficiency of polyphenol extraction.
The high total polyphenol content observed in S. mexicanum and C. vitifolium has important biological implications. The elevated phenolic content in these extracts could be directly associated with their ethnomedicinal uses and previously reported anti-inflammatory properties [16,17,18], as the activity of these compounds is largely linked to their antioxidant capacity.
2.1.3. Effect of Mexican Medicinal Plant Extracts on Cell Viability in Human Cancer and Normal Cell Lines
To evaluate the cytotoxic effects of the hexane, acetone, and methanol extracts of C. vitifolium, E. heterophyllum, P. auritum, and S. mexicanum, selected medicinal plants from Mexican Traditional Medicine, their impact on cell viability was assessed using a panel of human cancer cell lines, including cervical cancer (HeLa and SiHa), prostate cancer (PC-3), breast cancer (MCF-7), and lung cancer (H1299).
In parallel, cytotoxicity was also evaluated in non-cancerous human cell models, comprising normal human dermal fibroblasts (HFF-1), primary uterine fibroblast cells (HUF), as well as immortalized but non-tumorigenic human cell lines, including human keratinocytes (HaCaT) and human embryonic kidney cells (HEK-293), in order to estimate the selectivity of the extracts toward malignant cells.
Given the large number of treatments derived from the three extraction solvents per plant species and the multiple cell lines evaluated, representative concentration–response curves were selected based on cytotoxic potency, selectivity toward cancer cells, and relevance across different tumor tissue origins. As shown in Figure 2, the hexane, acetone, and methanol extracts of S. mexicanum and E. heterophyllum, as well as the methanol extract of C. vitifolium, induced a concentration-dependent reduction in cell viability in cancer cell lines derived from distinct tissues, including cervix, breast, lung, and prostate, with this effect being particularly pronounced in cervical cancer cells (HeLa).
Subsequently, based on the initial evaluation of extract effects on cell viability, concentration–response curves were generated and expressed as the percentage of proliferation inhibition as a function of extract concentration (Figure 3). These curves were used to quantitatively determine the half-maximal inhibitory concentration (IC50), defined as the concentration of a compound or extract required to inhibit 50% of cellular proliferation or viability relative to untreated control cells under identical experimental conditions. IC50 values provide a robust and comparative measure of biological potency, enabling the assessment of cytotoxic and antiproliferative effects of different extracts across multiple cell lines.
All extract treatments were systematically analyzed, and the resulting cytotoxicity data are summarized as IC50 values in Table 4, reported as mean ± SD from three independent experiments performed in triplicate.
Overall, the extracts displayed cytotoxic effects that were dependent on both the extraction solvent and the cell line, revealing differential responses among tumor types. The lowest IC50 values were mainly observed in cervical cancer cell lines (HeLa and SiHa), particularly for S. mexicanum extracts, indicating higher sensitivity of these cells. In contrast, lung (H1299), breast (MCF-7), liver (HepG2), and prostate (PC-3) generally exhibited slightly higher and more varied IC50 values compared to HeLa and SiHa cells, and the extracts of Piper auritum were the least cytotoxic. Suggesting a more moderate, tissue-dependent response. Paclitaxel, used as a positive control, showed consistently low IC50 values, confirming the validity of the experimental model.
To assess the selectivity of the evaluated extracts, their effects were further analyzed in a panel of non-cancerous or low malignant potential cell models, enabling a biologically relevant comparison across normal and immortalized cells from different tissue origins. This panel included normal human dermal fibroblasts (HFF-1), widely used as a reference model of normal stromal cells17; spontaneously immortalized human keratinocytes (HaCaT), which represent a non-tumorigenic epithelial model [19]; and human embryonic kidney epithelial cells (HEK293), an immortalized but non-tumorigenic cell line originally generated by adenoviral transformation [20,21].
In addition, to provide a more tissue-specific comparison relevant to cervical cancer, the extracts were also evaluated in HUF cells, corresponding to primary normal cervical fibroblasts, which serve as a physiologically relevant non-cancerous model of cervical origin [22].
Notably, all non-cancerous cell lines consistently exhibited higher IC50 values than those observed in cancer cell lines, indicating reduced sensitivity to the evaluated extracts. Among the non-cancerous models, HUF and HFF-1 cells were the least sensitive, followed by HaCaT and HEK293 cells (Table 5).
Figure 4 presents a comparative analysis of the effects of the hexanic, acetonic, and methanolic extracts of Semialarium mexicanum and Eryngium heterophyllum, which were previously identified as the most active species based on their cytotoxic responses in the cervical cancer cell lines HeLa and SiHa.
Overall, non-cancerous cells exhibited lower sensitivity to the extracts compared to cervical cancer cells. Among them, primary uterine fibroblast cells (HUF) and fibroblasts (HFF-1) were the least affected.
Notably, HeLa cells remained the most sensitive to all treatments, confirming a preferential cytotoxic effect toward cervical cancer cells. This response pattern indicates a selective cytotoxic profile of the evaluated extracts, characterized by enhanced activity against cancer cells while maintaining a favorable safety margin in normal and immortalized cell lines.
This selectivity is further supported by the selectivity index (SI) values summarized in Table 6. To facilitate a comparative visualization of the selectivity profile, a heatmap Figure 5 representation of the SI was generated using the values summarized in Table 6. The SI was calculated as the ratio between the IC50 obtained in non-cancerous cell lines (HFF-1, HaCaT, and HUF) and the IC50 determined in cervical cancer cell lines (HeLa and SiHa), where higher SI values indicate greater selectivity toward cancer cells.
The hexanic, acetonic, and methanolic extracts of S. mexicanum show SI >5 against at least one HeLa cell line, indicating high selectivity for tumor cells over HFF-1 and HUF. In comparison, E. heterophyllum exhibits moderate SI values (3–4.5).
Paclitaxel shows SI of 4.18 and 4.82 in HeLa-HaCaT and HeLa-HUF, respectively, with no data available for HFF-1, indicating good but not outstanding selectivity.
2.1.4. Morphological Assessment of Extract-Induced Cytotoxicity in Cancer and Normal Cells
To complement the quantitative data obtained from cell viability, IC50 determination, and proliferation inhibition assays, morphological changes induced by the most potent extracts were examined by microscopy. A concentration of 20 µg/mL, close to the IC50 in HeLa and SiHa cells, was applied to all treatments, including normal HUF fibroblasts, enabling direct comparison. At this concentration, HeLa (Figure 6) and SiHa cells (Figure 7) exhibited clear morphological alterations indicative of cytotoxicity, whereas HUF cells (Figure 8) remained largely unchanged relative to controls. These observations underscore the selective cytotoxic effects of the extracts on cancer cells while preserving the structural integrity of normal cells.
2.1.1. Effects of Hexanic, Acetonic, and Methanolic Extracts (Eryngium, Semialarium y Cochlospermum), on Cell Cycle Progression and Nuclear Morphology in HeLa Cells
While the primary aim of this study was to evaluate extract selectivity toward normal cells and identify those with a safe profile, we further explored their impact on cancer cell proliferation. Cell cycle analysis in HeLa cells (Figure 9) revealed that all extracts induced an increase in the sub-G1 population, indicative of DNA fragmentation and suggestive of cell death, with the hexanic extract of S. mexicanum producing the most pronounced effect, thereby corroborating the cytotoxicity and selectivity observed in previous assays.
To complement the evaluation of extract selectivity toward normal cells, we examined their effects on HeLa cell proliferation and nuclear morphology. Cell cycle analysis (Figure 9) revealed that all extracts induced an increase in the sub-G1 population, indicative of DNA fragmentation and suggestive of cell death, with the hexanic extract of S. mexicanum producing the most pronounced effect, corroborating previous cytotoxicity results. Consistently, DAPI staining (Figure 10) showed clear nuclear alterations: control cells (0.1% DMSO) displayed intact, uniformly stained nuclei, whereas positive controls (paclitaxel 20 and 100 nM) exhibited classic apoptotic features, including nuclear fragmentation and condensed apoptotic bodies. Treatment with 20 µg/mL of S. mexicanum, E. heterophyllum, and C. vitifolium induced varying degrees of chromatin condensation, nuclear fragmentation, and pyknotic nuclei, supporting the conclusion that the antiproliferative effects of these extracts could involve programmed cell death.
2.1.3. Chemical characterization
The hexane extract of S. mexicanum, analyzed by GC-MS and presented in Table 7 and Figure 11, generated greater toxicity in various cancer cell lines, with a special sensitivity observed in the cervical cancer cell lines (HeLa and SiHa). The chemical structures of these compounds were determined by analyzing their fragmentation patterns and comparing them with the NIST data library. The substances identified were benzene, 1,2,4,5-tetramethyl- (5) (23.90%) as the major constituent, followed by squalene (11) (11.43%), benzene, 1,2,3,5-tetramethyl- (4) (7.25%), friedelan-3-one (15) (5.73%), benzene,4-ethenyl-1,2-dimethyl- (7) (4.01%), 12-oleanen-3-yl acetate, (3.alpha.)- (12) (3.77%), benzene, 1-ethyl-3,5-dimethyl- (2) (3.72%), benzene, 1-ethyl-2,4-dimethyl- (1) (3.05%), cyanic acid, ethyl ester (9) (2.29%), stigmasta-3,5-diene (13) (2.23%), 4-ethylbenzoic acid, tridec-2-ynyl ester (8) (1.80%), β.-amyrone (14) (1.70%), indan, 1-methyl- (6) (1.55%), benzene, 2-ethyl-1,4-dimethyl- (3) (1.51%) and hexadecanoic acid, ethyl ester (10) (0.50%) (Fig.#). It is important to mention that some constituents present in the extract were not identified by this analysis.
3. Discussion
In this study, we demonstrate that hexane, acetone, and methanolic extracts of S. mexicanum exhibit pronounced selective cytotoxicity toward human cervical cancer cells, with SI greater than 5 in at least one HeLa cell line, indicating a marked preference for tumor cells over non-cancerous fibroblasts (HFF-1 and HUF). In contrast, extracts of E. heterophyllum and showed moderate SI values (3–4.5), reflecting cytotoxicity with a less discriminatory targeting of malignant cells. These findings highlight the importance of evaluating not only cytotoxic potency but also the therapeutic window: the ability to affect cancer cells without affecting healthy tissue, a key factor in the development of anticancer drugs and the determination of their safety.
Conventional chemotherapeutic agents, such as paclitaxel, maintain high potency but often lack sufficient selectivity, leading to dose-limiting toxicities in healthy tissues. In our model, paclitaxel exhibited SI values of 4.18 and 4.82 in the HeLa-HaCaT and HeLa-HUF comparisons, respectively, suggesting good, though not exceptional, selectivity. In contrast, S. mexicanum extracts achieved higher IS values even when effective concentrations exceeded those required for paclitaxel cytotoxicity, indicating a potentially wider safety margin—an essential characteristic for compounds derived from traditional medicinal plants that can reduce unwanted effects. Studies analyzing natural products have also highlighted the value of extracts that inhibit tumor cell viability with minimal effects on normal cells, reinforcing the therapeutic promise of plant-derived agents with selective profiles [23,24,25,26,27].
In all non-cancerous cell models analyzed (HFF-1, HUF, HaCaT, HEK293), consistently higher IC50 values were observed compared to cancer cells, suggesting lower sensitivity and supporting the safety profile of the extracts. Among these, HUF and HFF-1 fibroblasts were the least susceptible, followed by HaCaT keratinocytes and HEK293 cells. The inclusion of multiple non-cancerous cell lines, particularly HUF fibroblasts as a direct comparator with HeLa cervical cancer cells, reinforces the evidence of a preference for malignant over normal phenotypes [28,29,30,31,32].
The selective cytotoxicity of plant extracts has been documented in various studies with medicinal plants. For example, ethyl acetate fractions from Citrullus colocynthis demonstrated potent inhibition of the viability of pancreatic and cutaneous cancer cells, with minimal effects on normal BJ-1 cells, suggesting significant selectivity [33,34].
The selective cytotoxicity observed for S. mexicanum, particularly its hexane extract, may be partially explained by its chemical composition. GC–MS analysis of the hexanic stem extract revealed a complex mixture dominated by alkylated aromatic hydrocarbons and triterpenoid-related compounds, including benzene derivatives, squalene, β-amyrone, stigmasterol-related structures, and friedelan-3-one. Several of these compounds or related triterpenoids and sterols have been previously associated with antiproliferative and pro-apoptotic activities in cancer cell models, supporting their potential contribution to the observed biological effects [35,36,37,38,39].
Notably, squalene and pentacyclic triterpenoids such as amyrins and friedelane derivatives have been reported to modulate oxidative stress, membrane integrity, and apoptosis-related signaling pathways in tumor cells, while exhibiting relatively low toxicity toward non-malignant cells. The enrichment of such non-polar bioactive constituents in the hexane extract is consistent with its higher cytotoxic potency and selectivity compared to more polar extracts [40,41,42,43].
In contrast, acetonic and methanolic extracts of S. mexicanum showed a high content of total phenolic compounds, suggesting that phenolics may also contribute to the antiproliferative effects observed for these fractions. Phenolic compounds are widely recognized for their ability to interfere with cell cycle progression, induce oxidative imbalance, and activate programmed cell death pathways in cancer cells, while often displaying lower cytotoxicity toward normal cells [44,45,46,47].
Meanwhile, the cell cycle data revealed that all extracts significantly increased the sub-G1 population in HeLa cells, an indicator of DNA fragmentation associated with apoptotic processes, with the hexane extract from S. mexicanum producing the most pronounced effect [48].
This observation is consistent with reports indicating that plant extracts induce cell cycle disruption and apoptosis in cancer cells, a common mechanism attributed to phytochemicals such as flavonoids, alkaloids, and phenolic compounds that modulate cell death pathways. Furthermore, DAPI staining provided visual evidence of nuclear condensation and fragmentation in treated HeLa cells, further supporting the antiproliferative and cytotoxic effects of the extracts [49]. While the exact molecular mechanisms remain unclear, these combined findings suggest that the action of the extracts may involve programmed cell death pathways, a desirable attribute in anticancer agents [50,51].
Overall, the combined presence of lipophilic triterpenes and phenolic constituents in S. mexicanum extracts provides a plausible biochemical basis for the preferential cytotoxicity toward cancer cells observed in this study.
Although GC–MS provides tentative identification based on spectral matching, and does not allow definitive structural elucidation, the present chemical profiling offers a rational basis to explain the differential biological activity among extracts and supports future bioassay-guided fractionation and mechanistic studies to identify the compounds primarily responsible for the selective cytotoxic effects.
4. Materials and Methods
4.1. Collection of Plant Materials
The plant material was collected in various locations: Piper auritum (commonly known as “Hoja Santa”) in Álvaro Leonel, La Joya, Morelos (18° 53’ 30.0” N, 99° 07’ 15.8” W); Eryngium heterophyllum (“Hierba del Sapo”) in the municipality of Ixtapan de la Sal, State of Mexico (18° 53’ 25” N, 99° 40’ 24” W); Semialarium mexicanum (“Cancerina”) in Los Naranjos, Guerrero (18° 24’ 11.9” N, 99° 31’ 59.5” W); and Cochlospermum vitifolium (“Panicua”) in the Lomas del Carril neighborhood of Temixco, Morelos (18° 51’ 38.646” N, 99° 13’ 55.480” W). Once the specimen was deposited in the HUMO-UAEM Herbarium, the plant material was dried in an oven with air circulation at 50 °C and subsequently ground using an electric mill.
4.2. Plant Material Extraction
The plant material of each species, once powdered, was subjected to maceration using a series of organic solvents of increasing polarity (hexane, acetone, and methanol). For each species-solvent combination, 150 g of plant powder was placed in a 1 L amber flask with 1 L of the corresponding solvent. The mixtures were macerated for 72 hours, and the procedure was performed in triplicate. Following maceration, the extracts were filtered through Whatman No. 16 filter paper and concentrated to dryness under reduced pressure using a Büchi R-114 rotary evaporator.
Preliminary chromatographic analysis was carried out by thin-layer chromatography (TLC) and column chromatography (CC). For TLC, the plates were developed with two different reagents: 2-aminoethyl diphenylborinate (NP-PEG) as a specific reagent for flavonoid detection, and ammonium cerium (IV) sulfate as a universal developer. Visualization of the separated compounds was performed under ultraviolet light at wavelengths of 254 and 365 nm using a portable UV lamp (UVP UVGL-25).
4.3. Total Polyphenols
The total polyphenol content (TPC) was quantified using the Folin–Ciocalteu method [52]. Briefly, an aliquot of 28 μL of extract, dissolved in 8:2 H2O–MeOH (0.9 mg/mL), was mixed with 42 μL of Folin–Ciocalteu reagent (1 N). The mixture was kept on an orbital shaker at 80 ± 1 rpm and incubated at room temperature in the dark for 5 min. Then, 42 μL of Na2CO3 solution (20% w/v) and 168 μL of distilled water were added, followed by incubation under the same conditions for 30 min. Samples were immediately read against a blank on a Multiscan Go spectrophotometer at 760 nm. The TPC was calculated by comparison with a gallic acid standard curve (5–100 μg/mL; y = 0.0094x + 0.0803; R2 = 0.9999). Results are expressed as mg of gallic acid equivalents per g of dry extract (mg GAE/g extract). All determinations were performed in triplicate (n = 3) and are reported as mean ± standard deviation.
4.4. GC-MS Analysis
The hexane extract of H. excelsa was analyzed at the Laboratorio de Espectrometría de Masas del Instituto de Química, Universidad Nacional Autónoma de México. The analysis was performed using gas chromatography-mass spectrometry (GC-MS) with an Agilent Technologies 7890A gas chromatograph coupled to a Jeol GC Mate II mass spectrometer. Separation was carried out on a 30 m long, 0.32 mm internal diameter, 0.25 µm film thickness NP-5 capillary column (5% diphenyl/95% dimethylpolysilox-ane). Helium was used as the carrier gas at a constant flow rate of 1 mL/min. The in-jector temperature was maintained at 300 °C, and 1 µL of sample was injected in split mode (50:1). The samples were dissolved in hexane at a concentration of 1 mg/mL prior to injection. The oven temperature program started at 40 °C (maintained for 1 min) and increased to 300 °C at a rate of 8 °C/min. For detection, the mass spectrometer operated in electron impact ionization mode at 70 eV, recording a mass range of m/z 10 to 600 with a scan time of 1.3 s. The identification of the substances was made by comparison with data library of the National Institute of Standards and Technology (NIST).
4.5. Cell Culture and Cytotoxicity Assay
The cytotoxic activity of the plant extracts was evaluated using the crystal violet assay in a panel of human cancer and non-malignant cell lines, including human cancer cell lines HeLa (cervical adenocarcinoma), SiHa (cervical squamous cell carcinoma), HepG2 (hepatocellular carcinoma), PC3 (prostate adenocarcinoma), H1299 (non-small cell lung carcinoma), and MCF-7 (breast adenocarcinoma) were used in this study. Non-tumoral models included human uterine fibroblasts (HUF, primary cells), human foreskin fibroblasts (HFF-1), human embryonic kidney cells (HEK293), and human keratinocytes (HaCaT).
All cell lines were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA), except for HaCaT cells, which were kindly provided by Dr. Alexandre Toshirrico Cardoso Taketa, Full-Time Research Professor (Titular B) at the Centro de Investigación en Biotecnología, Universidad Autónoma del Estado de Morelos (UAEM), and primary human uterine fibroblasts (HUF), which were kindly donated by Dr. Heriberto Abraham Valencia González, researcher at the Instituto Nacional de Cancerología (INCan).
Cells were cultured at 37 °C in a humidified atmosphere with 5% CO2. Culture media and fetal bovine serum (FBS) were purchased from Invitrogen (Thermo Fisher Scientific, Waltham, MA, USA) and supplemented with 10% FBS and 1% penicillin–streptomycin. Specifically, HeLa and HaCaT cells were maintained in DMEM, MCF-7, SiHa and HEK293 cells in Eagle’s Minimum Essential Medium (EMEM), PC3 in RPMI, and HUF and HepG2 cells in DMEM/F-12. The remaining cell lines were cultured according to ATCC-recommended conditions.
For cytotoxicity evaluation, cells were seeded at a density of 5,000 cells per well in 96-well plates and allowed to adhere for 24 h. Cells were then treated with increasing concentrations of the plant extracts (200, 100, 50, 25, and 12.5 µg/mL) for 24 h. Paclitaxel was used as a positive control, while dimethyl sulfoxide (DMSO) served as the vehicle control, with a final concentration not exceeding 0.1% (v/v).
After treatment, cells were gently washed with phosphate-buffered saline (PBS), fixed with 4% paraformaldehyde and stained with 0.5% (w/v) crystal violet solution for 1 h. Excess dye was removed by thorough washing with distilled water, and plates were air-dried. The bound dye was solubilized using SDS 10%, and absorbance was measured at 570 nm using an automated ELISA reader (Promega, Madison, WI, USA). Cell viability was expressed as a percentage relative to untreated control cells.
Data Analysis
Concentration– response curves were generated, and IC50 values were calculated by nonlinear regression analysis using GraphPad PRISM software (version 8.0; GraphPad Software, San Diego, CA, USA). All assays were performed in triplicate and repeated in three independent experiments. Data are expressed as mean ± standard deviation (SD).
4.6. Cell Cycle Analysis
HeLa cells (1.5 × 105 cells/well) were seeded in 6-well plates. Exponentially growing cells were exposed for 24 h to the hexane, acetone, and methanol extracts of S. mexicanum, E. heterophyllum, and C. vitifolium at a concentration of 20 µg/mL, selected based on values close to their respective IC50. Paclitaxel (20 and 100 nM) was included as a positive control for G2/M phase arrest.
Following treatment, cells were trypsinized, collected as single-cell suspensions, centrifuged, and fixed overnight in cold 70% ethanol at −20 °C. Fixed cells were then treated with RNase A (0.01 M; Sigma-Aldrich) and stained with propidium iodide (PI; 7.5 µg/mL; Invitrogen) for 30 min in the dark. RNase treatment was included to eliminate RNA interference, allowing PI to bind specifically to cellular DNA.
Cell cycle distribution (G0/G1, S, and G2/M phases) was analyzed using a flow cytometer (FACS Lyric™, Becton Dickinson), acquiring 10,000 events per sample. All experiments were performed in triplicate and repeated in three independent experiments. Flow cytometry data were analyzed using FlowJo software (Tree Star Inc., Ashland, OR, USA) to generate DNA content histograms and quantify the percentage of cells in each cell cycle phase.
4.7. Nuclear Morphology Analysis by DAPI Staining
HeLa cells were seeded at a density of 1.25 × 105 cells per well in 6-well plates and allowed to attach overnight. Exponentially growing cells were treated for 24 h with the hexane, acetone, and methanol extracts of S. mexicanum, E. heterophyllum, and C. vitifolium at a concentration of 20 µg/mL, selected based on values close to their respective IC50. Paclitaxel (20 and 100 nM) was used as a positive control for apoptosis induction, while untreated cells served as the negative control.
After treatment, cells were washed twice with PBS and fixed with 4% paraformaldehyde for 15 min at room temperature. Fixed cells were subsequently permeabilized with 0.1% Triton X-100 in PBS for 5 min, washed with PBS, and stained with 4′,6-diamidino-2-phenylindole (DAPI; 1 µg/mL) for 10 min in the dark.
Nuclear morphology was examined using a fluorescence microscope equipped with a DAPI filter. Apoptotic features such as chromatin condensation, nuclear fragmentation, and apoptotic body formation were qualitatively assessed. Images were captured from randomly selected fields under identical acquisition settings. Experiments were performed in triplicate and repeated in three independent assays.
5. Conclusions
This study provides a comprehensive in vitro evaluation of the selective cytotoxic and antiproliferative effects of extracts from four Mexican medicinal plants (Semialarium mexicanum, Eryngium heterophyllum, Piper auritum, and Cochlospermum vitifolium) across a diverse panel of human cancer and non-cancerous cell lines, emphasizing selectivity as a decisive parameter in anticancer screening. Among the species analyzed, S. mexicanum emerged as the most promising candidate, exhibiting pronounced cytotoxic potency and preferential activity toward HeLa cervical cancer cells, particularly in its hexane extract, with selectivity indices exceeding those observed for paclitaxel when compared with primary human uterine fibroblasts (HUF).
Hexane extracts of S. mexicanum and E. heterophyllum were generally more active than their corresponding polar extracts, whereas P. auritum exhibited limited cytotoxicity and C. vitifolium displayed moderate, solvent-dependent effects. The use of physiologically relevant non-tumoral models, especially HUF cells as direct comparators for cervical cancer cell lines, strengthens the assessment of tumor selectivity.
Furthermore, the cell cycle alterations and nuclear morphological changes observed in HeLa cells support an antiproliferative response consistent with the activation of cell death pathways. When integrated with the chemical profile data, these biological effects suggest that the enrichment of lipophilic triterpenoids and phenolic components in S. mexicanum extracts could contribute to their selective cytotoxic profile.
Although confined to in vitro systems, the present study establishes a toxicologically informed screening strategy for medicinal plant extracts and supports the value of selectivity-guided evaluation as a critical step preceding chemical isolation, mechanistic elucidation, and more advanced safety assessments.
Author Contributions
Conceptualization, J.D and J.N.S.C; methodology, Y.A., G.A., K.L., A.G. and E.C..; software, J.F., C.M and A.N.; validation, S.M., J.F. and A.G.; formal analysis, Y.A., G.A., and K.L.; investigation, J.D, J.M..; resources, C.M and A.N.; data curation, J.U.; writing—original draft preparation, J.N.S.C.; writing—review and editing, J.D., E.S, E.A. A.G. and J.N.S.C.; visualization, E.C and E.A.; supervision, J.D.; project administration, J.N.S.C, S.M. and J.D.; funding acquisition, E.S and J.N.S.C. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
During the preparation of this manuscript, the authors used generative artificial intelligence tools (ChatGPT, Gemini, and NotebookLM) exclusively for the generation and formatting of graphical elements and illustrative images. The authors reviewed and edited all generated materials and take full responsibility for the content, interpretation, and scientific accuracy of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| TLA | Three letter acronym |
| LD | Linear dichroism |
References
- Soto, K. M.; Pérez Bueno, J. J.; Mendoza López, M. L.; Apátiga-Castro, M.; López-Romero, J. M.; Mendoza, S.; Manzano-Ramírez, A. Anti-oxidants in Traditional Mexican Medicine and Their Applications as Antitumor Treatments. Pharmaceuticals 2023, 16, 482. [Google Scholar] [CrossRef]
- Cuevas-Cianca, S. I.; González-Campos, R. E.; Mejía Méndez, J. L.; Sánchez Arreola, E.; Juárez, Z. N.; Hernández, L. R. Anticancer Properties of Mexican Medicinal Plants: An Updated Review. Nat. Prod. Commun. 2023, 18, 1–9. [Google Scholar] [CrossRef]
- Jacobo-Herrera, NJ; Jacobo-Herrera, FE; Zentella-Dehesa, A; Andrade-Cetto, A; Heinrich, M; Pérez-Plasencia, C. Medicinal plants used in Mexican traditional medicine for the treatment of colorectal cancer. J Ethnopharmacol. 2016, 179, 391–402. [Google Scholar] [CrossRef]
- Maldonado-Cubas, J.; San Martin-Martínez, E.; Quiroz-Reyes, C. N.; Casañas-Pimentel, R. G. Cytotoxic Effect of Semialarium mexicanum (Miers) Mennega Root Bark Extracts and Fractions against Breast Cancer Cells. Physiol. Mol. Biol. Plants 2018, 24, 1185–1201. [Google Scholar] [CrossRef]
- Cáceres-Castillo, D.; Mena-Rejón, G. J.; Cedillo-Rivera, R.; Quijano, L. 21β-Hydroxy-Oleanane-Type Triterpenes from Hippocratea excelsa. Phytochemistry 2008, 69, 1057–1064. [Google Scholar] [CrossRef] [PubMed]
- Araujo Leon, J. A.; Ruiz Ciau, D. V.; Coral Martinez, T. I.; Cantillo Ciau, Z. O. Comparative Fingerprint Analyses of Extracts from the Root Bark of Wild Hippocratea excelsa and “Cancerina” by High-Performance Liquid Chromatography. J. Sep. Sci. 2015, 38, 3870–3875. [Google Scholar] [CrossRef]
- Salleh, W. M. N. H. W. A Systematic Review of Botany, Phytochemicals and Pharmacological Properties of “Hoja Santa” (Piper auritum Kunth). Z. Naturforsch. C 2020, 76, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Guadarrama, A. B.; Rios, M. Y. Flavonoids, Sterols and Lignans from Cochlospermum vitifolium and Their Relationship with Its Liver Activity. Molecules 2018, 23, 1952. [Google Scholar] [CrossRef]
- Pérez-Muñoz, E. P.; Antunes-Ricardo, M.; Martínez-Ávila, M.; Guajardo-Flores, D. Eryngium Species as a Potential Ally for Treating Metabolic Syndrome and Diabetes. Front. Nutr. 2022, 9, 878306. [Google Scholar] [CrossRef] [PubMed]
- Newman, D. J.; Cragg, G. M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- GBIF Occurrence Download. Available online: https://doi.org/10.15468/dl.cym2fw (accessed on 11 November 2025).
- GBIF Occurrence Download. Available online: https://doi.org/10.15468/dl.vh2rm7 (accessed on 11 November 2025).
- GBIF Occurrence Download. Available online: https://doi.org/10.15468/dl.7stc5p (accessed on 11 November 2025).
- GBIF Occurrence Download. Available online: https://doi.org/10.15468/dl.4rg5pk (accessed on 19 November 2025).
- El Mannoubi, I. Impact of Different Solvents on Extraction Yield, Phenolic Composition, In Vitro Antioxidant and Antibacterial Activities of Deseeded Opuntia stricta Fruit. J. Umm Al-Qura Univ. Appl. Sci. 2023, 9, 176–184. [Google Scholar] [CrossRef]
- Sarmento-Filha, M. J.; Torres-Rêgo, M.; Daniele-Silva, A.; de Queiroz-Neto, M. F.; Rocha, H. A. O.; Camara, C. A.; Araújo, R. M.; da Silva-Júnior, A. A.; Silva, T. M. S.; Fernandes-Pedrosa, M. de F. Phytochemical Analysis by UPLC-QTOF-MS/MS and Evaluation of Antioxidant and Anti-Inflammatory Activities of the Extract and Fractions from Flowers of Cochlospermum vitifolium. S. Afr. J. Bot. 2022, 148, 293–306. [Google Scholar] [CrossRef]
- Perez, R. M.; Perez, S.; Zavala, M. A.; Salazar, M. Anti-Inflammatory Activity of the Bark of Hippocratea excelsa. J. Ethnopharmacol. 1995, 47, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Davì, F.; Taviano, M. F.; Acquaviva, R.; et al. Chemical Profile, Antioxidant and Cytotoxic Activity of a Phenolic-Rich Fraction from the Leaves of Brassica fruticulosa subsp. fruticulosa (Brassicaceae) Growing Wild in Sicily (Italy). Molecules 2023, 28, 2281. [Google Scholar] [CrossRef] [PubMed]
- Seo, M. D.; Kang, T. J.; Lee, C. H.; Lee, A. Y.; Noh, M. HaCaT Keratinocytes and Primary Epidermal Keratinocytes Have Different Transcriptional Profiles of Cornified Envelope-Associated Genes to T Helper Cell Cytokines. Biomol. Ther. (Seoul) 2012, 20, 171–176. [Google Scholar] [CrossRef]
- Graham, F. L.; Smiley, J.; Russell, W. C.; Nairn, R. Characteristics of a Human Cell Line Transformed by DNA from Human Adenovirus Type 5. J. Gen. Virol. 1977, 36, 59–74. [Google Scholar] [CrossRef]
- Topics, ScienceDirect. HEK293 Cell Line; Elsevier; Available online: https://www.sciencedirect.com/topics/immunology-and-microbiology/hek293-cell-line (accessed on 8 March 2025).
- Keyif, B.; Hacioglu, C. Boric Acid Suppresses Cell Survival by Triggering Endoplasmic Reticulum Stress-Induced Autophagy in Cervical Cancers. J. Cell. Mol. Med. 2025, 29, e70740. [Google Scholar] [CrossRef]
- Beutler, J. A. Natural Products as a Foundation for Drug Discovery. Curr. Protoc. Pharmacol. 2019, 86, e67. [Google Scholar] [CrossRef]
- Kellogg, J. J.; Paine, M. F.; McCune, J. S.; Oberlies, N. H.; Cech, N. B. Selection and Characterization of Botanical Natural Products for Research Studies: A NaPDI Center Recommended Approach. Nat. Prod. Rep. 2019, 36, 1196–1221. [Google Scholar] [CrossRef]
- Beutler, J. A. Natural Products as a Foundation for Drug Discovery. Curr. Protoc. Pharmacol. 2009, 46, 9.11.1–9.11.21. [Google Scholar] [CrossRef]
- Greenwell, M.; Rahman, P. K. Medicinal Plants: Their Use in Anticancer Treatment. Int. J. Pharm. Sci. Res. 2015, 6, 4103–4112. [Google Scholar] [CrossRef]
- Talib, W. H.; Baban, M. M.; Bulbul, M. F.; Al-Zaidaneen, E.; Allan, A.; Al-Rousan, E. W.; Ahmad, R. H. Y.; Alshaeri, H. K.; Alasmari, M. M.; Law, D. Natural Products and Altered Metabolism in Cancer: Therapeutic Targets and Mechanisms of Action. Int. J. Mol. Sci. 2024, 25, 9593. [Google Scholar] [CrossRef]
- Calderón-Montaño, J. M.; Martínez-Sánchez, S. M.; Jiménez-González, V.; Burgos-Morón, E.; Guillén-Mancina, E.; Jiménez-Alonso, J. J.; Díaz-Ortega, P.; García, F.; Aparicio, A.; López-Lázaro, M. Screening for Selective Anticancer Activity of 65 Extracts of Plants Collected in Western Andalusia, Spain. Plants 2021, 10, 2193. [Google Scholar] [CrossRef] [PubMed]
- López-Lázaro, M. A Simple and Reliable Approach for Assessing Anticancer Activity In Vitro. Curr. Med. Chem. 2015, 22, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- López-Lázaro, M. Two Preclinical Tests to Evaluate Anticancer Activity and to Help Validate Drug Candidates for Clinical Trials. Oncoscience 2015, 2, 91–98. [Google Scholar] [CrossRef]
- López-Lázaro, M. How Many Times Should We Screen a Chemical Library to Discover an Anticancer Drug? Drug Discov. Today 2015, 20, 167–169. [Google Scholar] [CrossRef]
- Adedokun, K. A.; Imodoye, S. O.; Bello, I. O.; Lanihun, A.-A.; Bello, I. O. Therapeutic Potentials of Medicinal Plants and Significance of Computational Tools in Anti-Cancer Drug Discovery. In Phytochemistry, Computational Tools and Databases in Drug Discovery; Egbuna, C., Rudrapal, M., Tijjani, H., Eds.; Elsevier: Amsterdam, The Netherlands, 2023; pp. 393–455. [Google Scholar] [CrossRef]
- Mandour, Y. M.; Refaat, E.; Hassanein, H. D. Anticancer Activity, Phytochemical Investigation and Molecular Docking Insights of Citrullus colocynthis (L.) Fruits. Sci. Rep. 2023, 13, 20038. [Google Scholar] [CrossRef]
- Mata, R.; Figueroa, M.; Navarrete, A.; Rivero-Cruz, I. Chemistry and Biology of Selected Mexican Medicinal Plants. Prog. Chem. Org. Nat. Prod. 2019, 108, 1–142. [Google Scholar] [CrossRef]
- Zengin, K.B.; Özmen, Ö.; Öztürk Civelek, D.; Şenol, H.; Sönmez, F. Exploring Anti-Cancer Properties of New Triazole-Linked Benzenesulfonamide Derivatives against Colorectal Carcinoma: Synthesis, Cytotoxicity, and In Silico Insights. Bioorg. Med. Chem. 2025, 119, 118060. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.T.; Tan, Y.J.; Oon, C.E. Benzimidazole and Its Derivatives as Cancer Therapeutics: The Potential Role from Traditional to Precision Medicine. Acta Pharm. Sin. B 2023, 13, 478–497. [Google Scholar] [CrossRef]
- Lou, S.; Dai, C.; Wu, Y.; et al. Betulonic Acid: A Review on Its Sources, Biological Activities, and Molecular Mechanisms. Eur. J. Pharmacol. 2025, 998, 177518. [Google Scholar] [CrossRef]
- Similie, D.; Minda, D.; Bora, L.; et al. An Update on Pentacyclic Triterpenoids Ursolic and Oleanolic Acids and Related Derivatives as Anticancer Candidates. Antioxidants 2024, 13, 952. [Google Scholar] [CrossRef]
- Smith, T. Squalene: Potential Chemopreventive Agent. Expert Opin. Investig. Drugs 2000, 9, 1841–1848. [Google Scholar] [CrossRef]
- Silva, L.W.P.E.; Almeida, T.C.; Teixeira, M.S.D.S.; et al. Antiproliferative Effects of the Triterpene Ursolic Acid Natural Product in Bladder and Ovarian Tumor Cell Lines. Drug Dev. Res. 2025, 86, e70172. [Google Scholar] [CrossRef]
- Nemli, E.; Saricaoglu, B.; Kirkin, C.; et al. Chemopreventive and Chemotherapeutic Potential of Betulin and Betulinic Acid: Mechanistic Insights from In Vitro, In Vivo and Clinical Studies. Food Sci. Nutr. 2024, 12, 10059–10069. [Google Scholar] [CrossRef]
- Similie, D.; Minda, D.; Bora, L.; Kroškins, V.; Lugiņina, J.; Turks, M.; Dehelean, C.A.; Danciu, C. An Update on Pentacyclic Triterpenoids Ursolic and Oleanolic Acids and Related Derivatives as Anticancer Candidates. Antioxidants 2024, 13, 952. [Google Scholar] [CrossRef] [PubMed]
- Bidooki, S.H.; Quero, J.; Sánchez-Marco, J.; et al. Squalene in Nanoparticles Improves Antiproliferative Effect on Human Colon Carcinoma Cells through Apoptosis by Disturbances in Redox Balance. Int. J. Mol. Sci. 2024, 25, 13048. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.P.; Li, S.; Chen, Y.M.; Li, H.B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef]
- Bouyahya, A.; Omari, N.E.; Bakrim, S.; Hachlafi, N.E.; Balahbib, A.; Wilairatana, P.; Mubarak, M.S. Advances in Dietary Phenolic Compounds to Improve Chemosensitivity of Anticancer Drugs. Cancers 2022, 14, 4573. [Google Scholar] [CrossRef]
- Singaravelan, N.; Tollefsbol, T.O. Polyphenol-Based Prevention and Treatment of Cancer through Epigenetic and Combinatorial Mechanisms. Nutrients 2025, 17, 616. [Google Scholar] [CrossRef] [PubMed]
- Yammine, A.; Namsi, A.; Vervandier-Fasseur, D.; Mackrill, J.J.; Lizard, G.; Latruffe, N. Polyphenols of the Mediterranean Diet and Their Metabolites in the Prevention of Colorectal Cancer. Molecules 2021, 26, 3483. [Google Scholar] [CrossRef]
- Riccardi, C.; Nicoletti, I. Analysis of Apoptosis by Propidium Iodide Staining and Flow Cytometry. Nat. Protoc. 2006, 1, 1458–1461. [Google Scholar] [CrossRef] [PubMed]
- Atale, N.; Gupta, S.; Yadav, U.; Rani, V. Cell-Death Assessment by Fluorescent and Nonfluorescent Cytosolic and Nuclear Staining Techniques. J. Microsc. 2014, 255, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Blowman, K.; Magalhães, M.; Lemos, M.; Cabral, C.; Pires, I. Anticancer Properties of Essential Oils and Other Natural Products. Evid. Based Complement. Altern. Med. 2018, 3149362. [Google Scholar] [CrossRef]
- Bernardini, S.; Tiezzi, A.; Laghezza Masci, V.; Ovidi, E. Natural Products for Human Health: An Historical Overview of the Drug Discovery Approaches. Nat. Prod. Res. 2018, 32, 1926–1950. [Google Scholar] [CrossRef]
- Teixeira, B. J.; Ataurima, I. M.; Inca, L. S.; Llica, E. R.; Cotos, M. R.; Manrique, P. E.; Alfaro, K. M. Evaluación del contenido de polifenoles totales y la capacidad antioxidante de los extractos etanólicos de los frutos de aguaymanto (Physalis peruviana L.) de diferentes lugares del Perú. Rev. Soc. Quím. Perú 2016, 82, e58. [Google Scholar] [CrossRef]
Figure 1.
Figure 1. Occurrence records of the four species in Mexico. Letter A indicates the collection site of C. vitifolium, B of E. heterophyllum, C of S. mexicanum, and D of P. auritum. For Mexico, the background color intensity represents elevation (m a.s.l.): light tones indicate low elevation, and dark tones indicate high elevation.
Figure 1.
Figure 1. Occurrence records of the four species in Mexico. Letter A indicates the collection site of C. vitifolium, B of E. heterophyllum, C of S. mexicanum, and D of P. auritum. For Mexico, the background color intensity represents elevation (m a.s.l.): light tones indicate low elevation, and dark tones indicate high elevation.
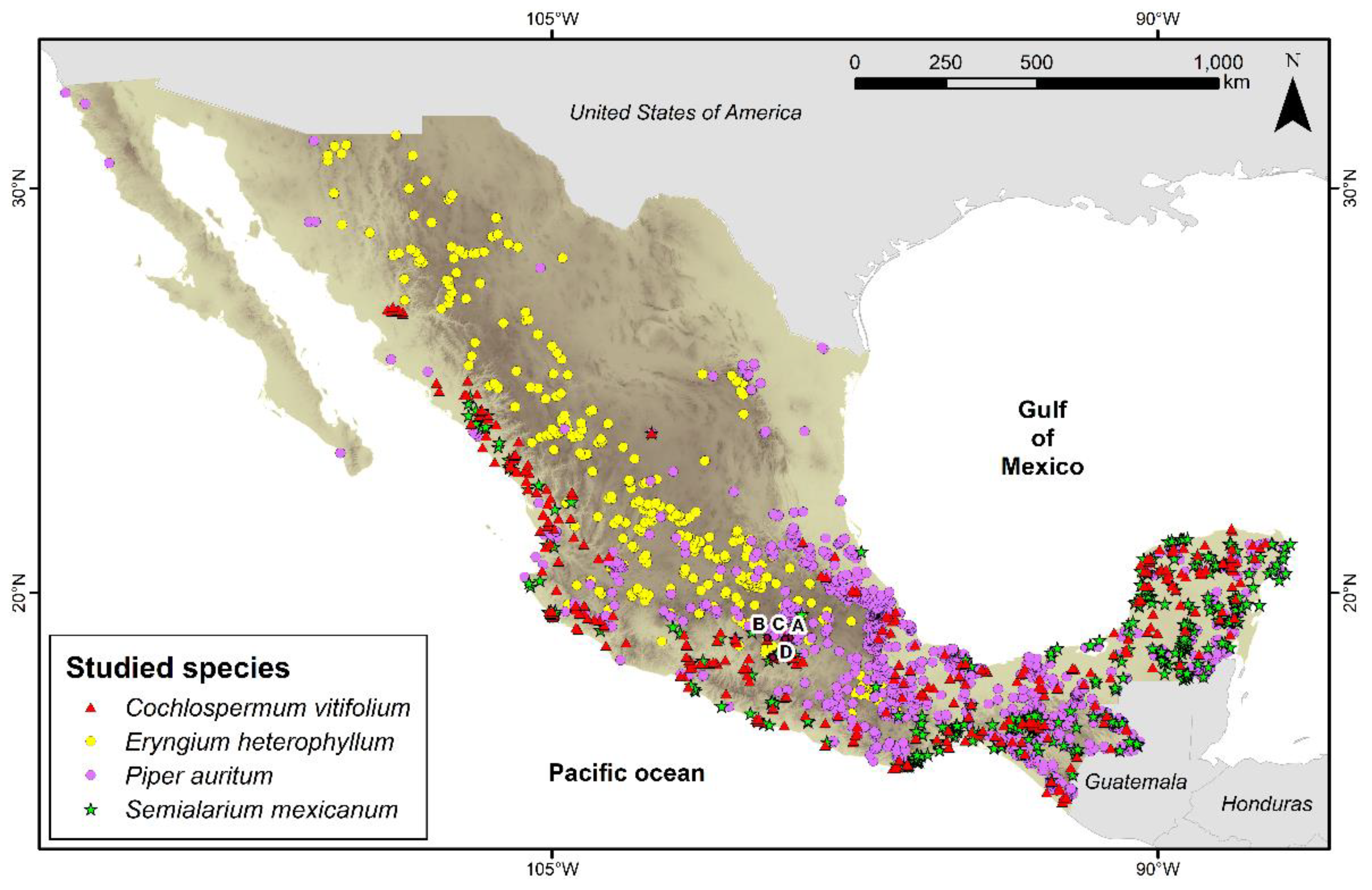
Figure 2.
Effect on HeLa cell viability of plant extracts. HeLa cells were treated with hexane, acetone, and methanol extracts from Semialarium mexicanum (a–c), Eryngium heterophyllum (d–f), and Cochlospermum vitifolium (g–i) at concentrations ranging from 0 to 200 µg/mL. Cell viability is expressed as percentage of the negative control (0.1% DMSO). All experiments were performed in triplicate, and data show a concentration-dependent decrease in cell viability, with extract- and species-dependent differences in potency.
Figure 2.
Effect on HeLa cell viability of plant extracts. HeLa cells were treated with hexane, acetone, and methanol extracts from Semialarium mexicanum (a–c), Eryngium heterophyllum (d–f), and Cochlospermum vitifolium (g–i) at concentrations ranging from 0 to 200 µg/mL. Cell viability is expressed as percentage of the negative control (0.1% DMSO). All experiments were performed in triplicate, and data show a concentration-dependent decrease in cell viability, with extract- and species-dependent differences in potency.
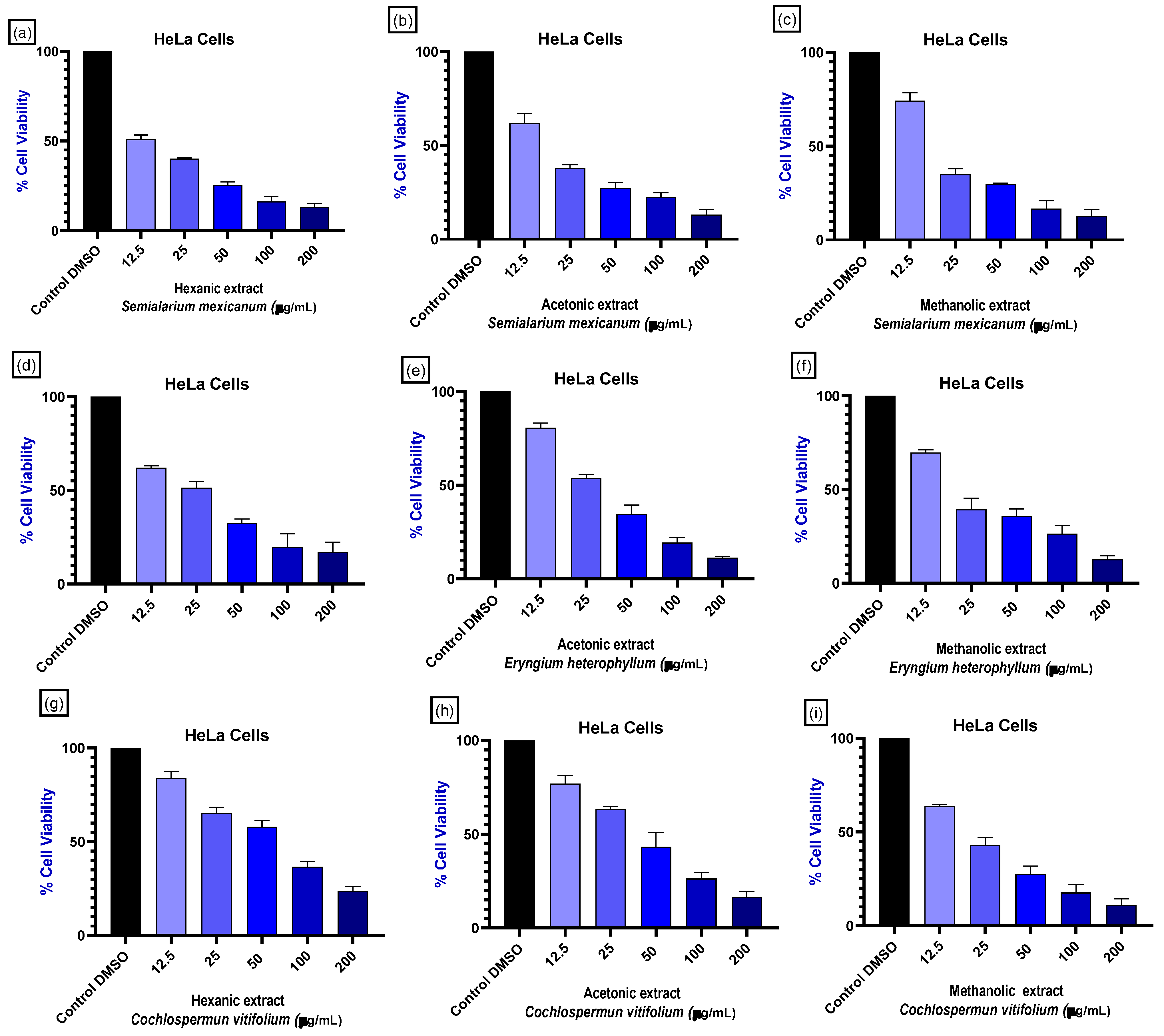
Figure 3.
Concentration–response curves showing proliferation inhibition in HeLa cells treated with plant extracts. Percentage of proliferation inhibition in HeLa cells treated with hexane, acetone, and methanolic extracts of Semialarium mexicanum (a–c), Eryngium heterophyllum (d–f), and Cochlospermum vitifolium (g–i) at concentrations ranging from 0 to 200 µg/mL. Curves were used to calculate IC50 values. Cells treated with 0.1% DMSO served as the negative control. Data are expressed as mean ± SD of three independent experiments performed in triplicate.
Figure 3.
Concentration–response curves showing proliferation inhibition in HeLa cells treated with plant extracts. Percentage of proliferation inhibition in HeLa cells treated with hexane, acetone, and methanolic extracts of Semialarium mexicanum (a–c), Eryngium heterophyllum (d–f), and Cochlospermum vitifolium (g–i) at concentrations ranging from 0 to 200 µg/mL. Curves were used to calculate IC50 values. Cells treated with 0.1% DMSO served as the negative control. Data are expressed as mean ± SD of three independent experiments performed in triplicate.
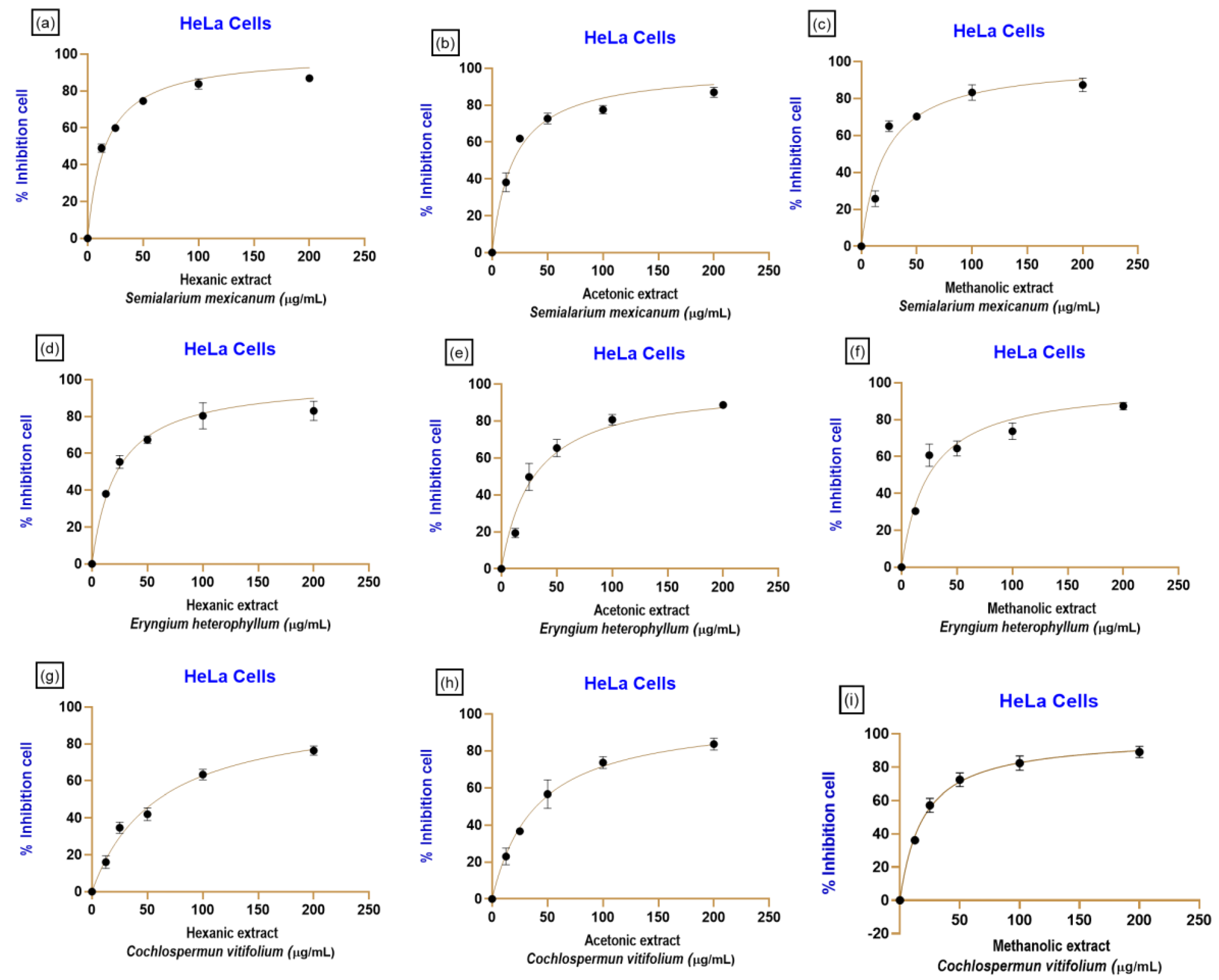
Figure 4.
Comparative cytotoxic effects of selected Mexican medicinal plant extracts across cancerous and non-cancerous cell lines. Bar graphs depict the percentage of cell viability inhibition induced by hexane, acetone, and methanolic extracts of Semialarium mexicanum and Eryngium heterophyllum in human cervical cancer cell lines HeLa and non-cancerous cell models, including HFF-1, HUF, HaCaT, and HEK293 cells. Values represent mean ± SD from three independent experiments performed in triplicate.
Figure 4.
Comparative cytotoxic effects of selected Mexican medicinal plant extracts across cancerous and non-cancerous cell lines. Bar graphs depict the percentage of cell viability inhibition induced by hexane, acetone, and methanolic extracts of Semialarium mexicanum and Eryngium heterophyllum in human cervical cancer cell lines HeLa and non-cancerous cell models, including HFF-1, HUF, HaCaT, and HEK293 cells. Values represent mean ± SD from three independent experiments performed in triplicate.
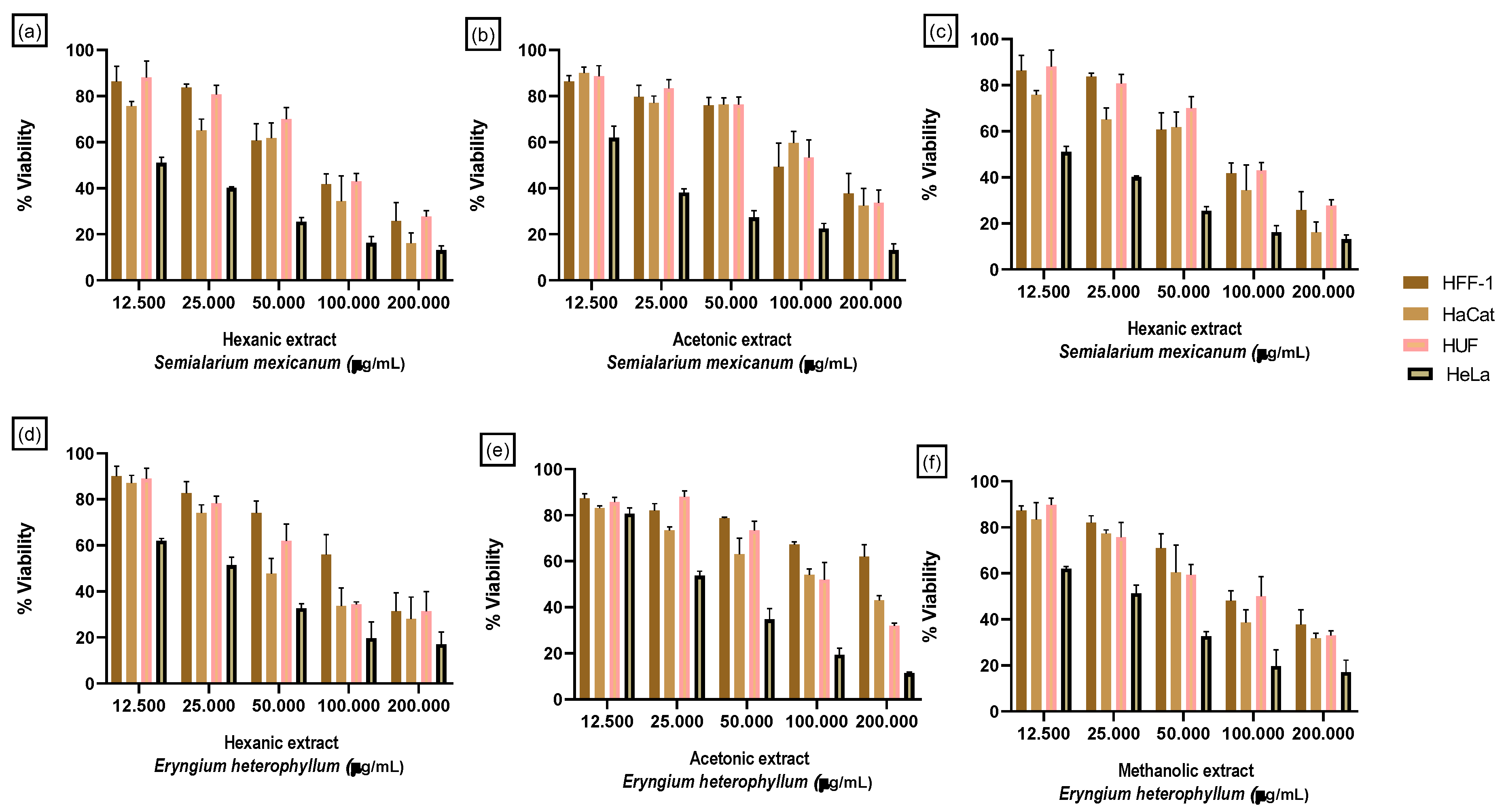
Figure 5.
Heatmap of SI values selected plant extracts. Heatmap representation of SI values calculated for hexane, acetone, and methanolic extracts of Eryngium heterophyllum and Semialarium mexicanum in cervical cancer cell lines (HeLa and SiHa) relative to non-cancerous cell models (HFF-1, HaCaT, and HUF). SI values were calculated as the ratio between IC50 values in non-cancerous cells and the corresponding IC50 values in cancer cells. Higher SI values indicate greater selectivity toward cancer cells. Paclitaxel was included as a reference compound. ND indicates not determined.
Figure 5.
Heatmap of SI values selected plant extracts. Heatmap representation of SI values calculated for hexane, acetone, and methanolic extracts of Eryngium heterophyllum and Semialarium mexicanum in cervical cancer cell lines (HeLa and SiHa) relative to non-cancerous cell models (HFF-1, HaCaT, and HUF). SI values were calculated as the ratio between IC50 values in non-cancerous cells and the corresponding IC50 values in cancer cells. Higher SI values indicate greater selectivity toward cancer cells. Paclitaxel was included as a reference compound. ND indicates not determined.
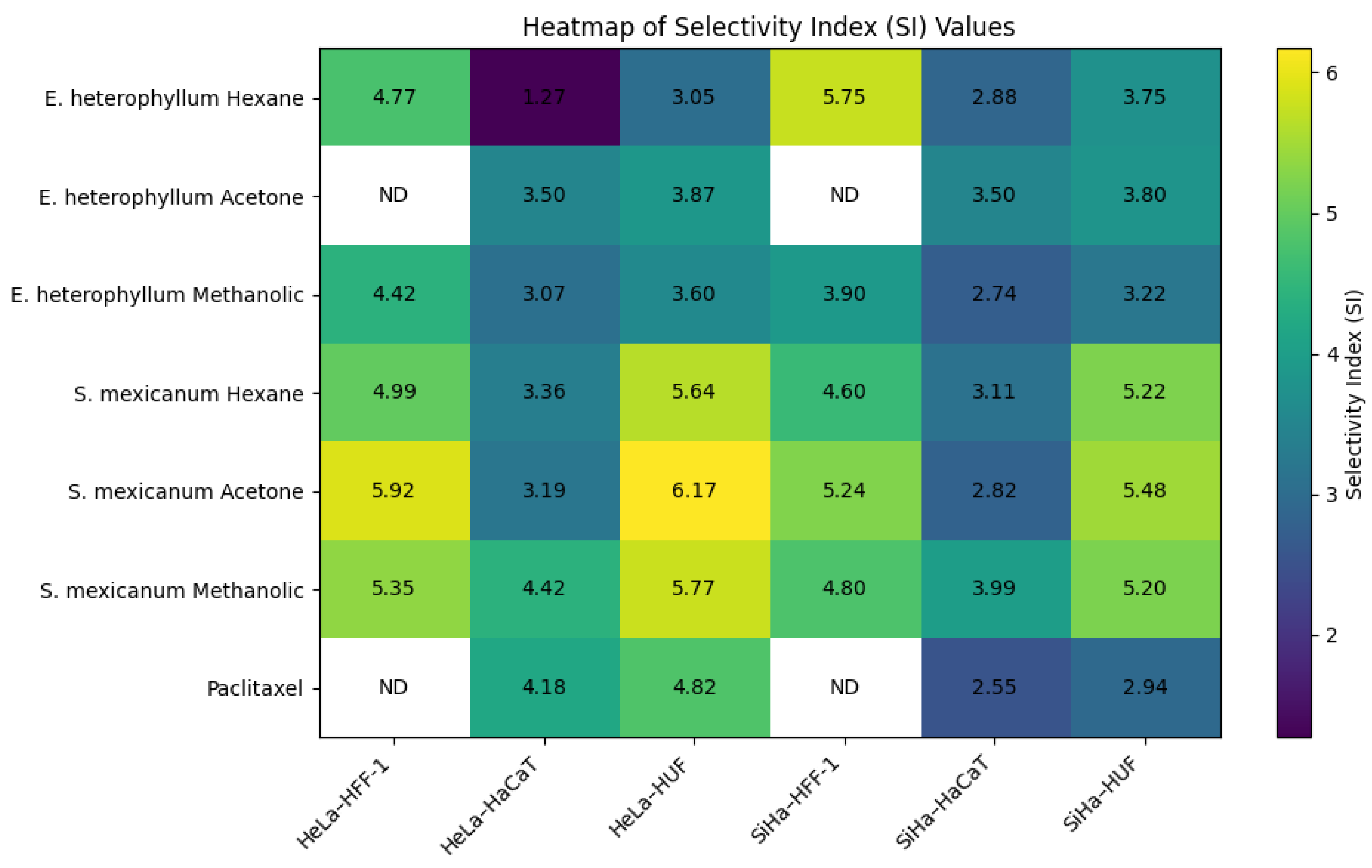
Figure 6.
Representative micrographs of HeLa cells treated for 48 h with hexanic, acetonic, and methanolic extracts of Semialarium mexicanum, Eryngium heterophyllum, and Cochlospermum vitifolium (20 µg/mL), compared to vehicle control (0.1% DMSO) and positive control paclitaxel (20 and 100 nM). The images reveal a marked reduction in cell density, morphological changes indicative of cell death, and the formation of aggregates following extract exposure. Images were acquired at 10X magnification.
Figure 6.
Representative micrographs of HeLa cells treated for 48 h with hexanic, acetonic, and methanolic extracts of Semialarium mexicanum, Eryngium heterophyllum, and Cochlospermum vitifolium (20 µg/mL), compared to vehicle control (0.1% DMSO) and positive control paclitaxel (20 and 100 nM). The images reveal a marked reduction in cell density, morphological changes indicative of cell death, and the formation of aggregates following extract exposure. Images were acquired at 10X magnification.
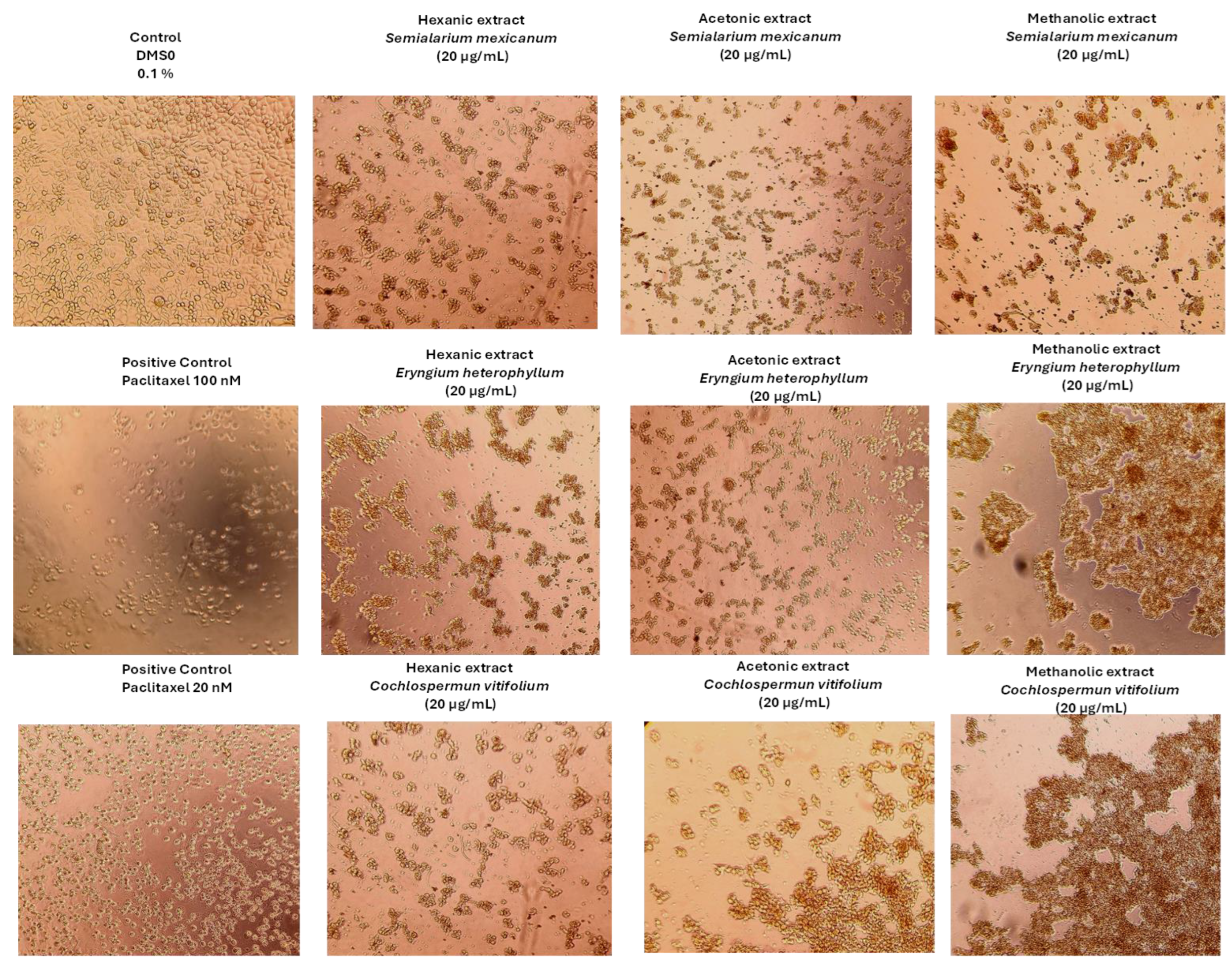
Figure 7.
Representative phase-contrast micrographs of SiHa cells after 48 h of treatment with hexanic, acetonic, and methanolic extracts of Semialarium mexicanum, Eryngium heterophyllum, and Cochlospermum vitifolium (20 µg/mL), compared to vehicle control (0.1% DMSO) and positive control paclitaxel (20 and 100 nM). The images show reduced confluency, cell detachment, and morphological changes indicative of extract-induced cell death. Images were acquired at 10X magnification.
Figure 7.
Representative phase-contrast micrographs of SiHa cells after 48 h of treatment with hexanic, acetonic, and methanolic extracts of Semialarium mexicanum, Eryngium heterophyllum, and Cochlospermum vitifolium (20 µg/mL), compared to vehicle control (0.1% DMSO) and positive control paclitaxel (20 and 100 nM). The images show reduced confluency, cell detachment, and morphological changes indicative of extract-induced cell death. Images were acquired at 10X magnification.

Figure 8.
Representative phase-contrast micrographs of HUF cells after 48 h of treatment with hexanic, acetonic, and methanolic extracts of Semialarium mexicanum, Eryngium heterophyllum, and Cochlospermum vitifolium (20 µg/mL), compared to vehicle control (0.1% DMSO) and positive control paclitaxel (20 and 100 nM). At this concentration, no significant morphological changes were observed in HUF cells, supporting the selective cytotoxicity of the extracts toward cancer cells. Images were acquired at 10X magnification.
Figure 8.
Representative phase-contrast micrographs of HUF cells after 48 h of treatment with hexanic, acetonic, and methanolic extracts of Semialarium mexicanum, Eryngium heterophyllum, and Cochlospermum vitifolium (20 µg/mL), compared to vehicle control (0.1% DMSO) and positive control paclitaxel (20 and 100 nM). At this concentration, no significant morphological changes were observed in HUF cells, supporting the selective cytotoxicity of the extracts toward cancer cells. Images were acquired at 10X magnification.

Figure 9.
Cell cycle analysis by flow cytometry of cells treated with extracts from Semialarium mexicanum, Eryngium heterophyllum, and Cochlospermum vitifolium in HeLa cells. Representative histograms show the DNA content distribution (propidium iodide staining) following treatment with hexanic, acetonic, and methanolic extracts 20 µg/mL. Negative control (0.1% DMSO) displaying a standard distribution of G1, S, and G2/M phases. Positive controls with Paclitaxel (100 nM and 20 nM) showing apoptosis induction (increased SubG1 sub-population) and G2/M phase arrest.
Figure 9.
Cell cycle analysis by flow cytometry of cells treated with extracts from Semialarium mexicanum, Eryngium heterophyllum, and Cochlospermum vitifolium in HeLa cells. Representative histograms show the DNA content distribution (propidium iodide staining) following treatment with hexanic, acetonic, and methanolic extracts 20 µg/mL. Negative control (0.1% DMSO) displaying a standard distribution of G1, S, and G2/M phases. Positive controls with Paclitaxel (100 nM and 20 nM) showing apoptosis induction (increased SubG1 sub-population) and G2/M phase arrest.
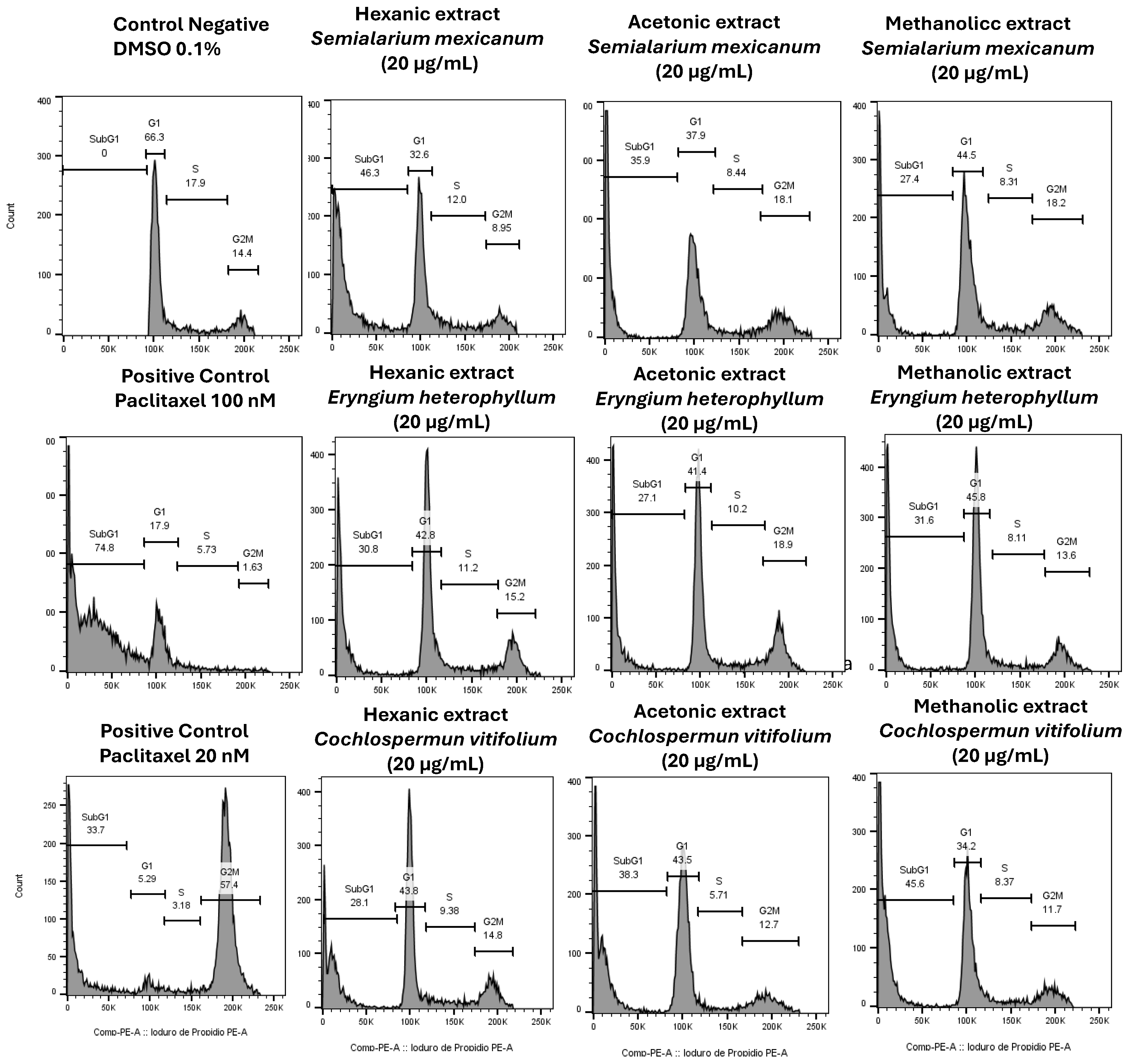
Figure 10.
Nuclear morphology of HeLa cells assessed by DAPI staining. Representative fluorescence micrographs (10× magnification) of cells treated for 24 h with hexanic, acetonic, and methanolic extracts (20 µg/mL) of Semialarium mexicanum, Eryngium heterophyllum, and Cochlospermum vitifolium in HeLa cells. Negative controls (0.1% DMSO) display intact nuclei with uniform chromatin, whereas positive controls (paclitaxel 20 and 100 nM) and extract-treated cells exhibit apoptotic features, including chromatin condensation, nuclear fragmentation, and brightly fluorescent apoptotic bodies.
Figure 10.
Nuclear morphology of HeLa cells assessed by DAPI staining. Representative fluorescence micrographs (10× magnification) of cells treated for 24 h with hexanic, acetonic, and methanolic extracts (20 µg/mL) of Semialarium mexicanum, Eryngium heterophyllum, and Cochlospermum vitifolium in HeLa cells. Negative controls (0.1% DMSO) display intact nuclei with uniform chromatin, whereas positive controls (paclitaxel 20 and 100 nM) and extract-treated cells exhibit apoptotic features, including chromatin condensation, nuclear fragmentation, and brightly fluorescent apoptotic bodies.
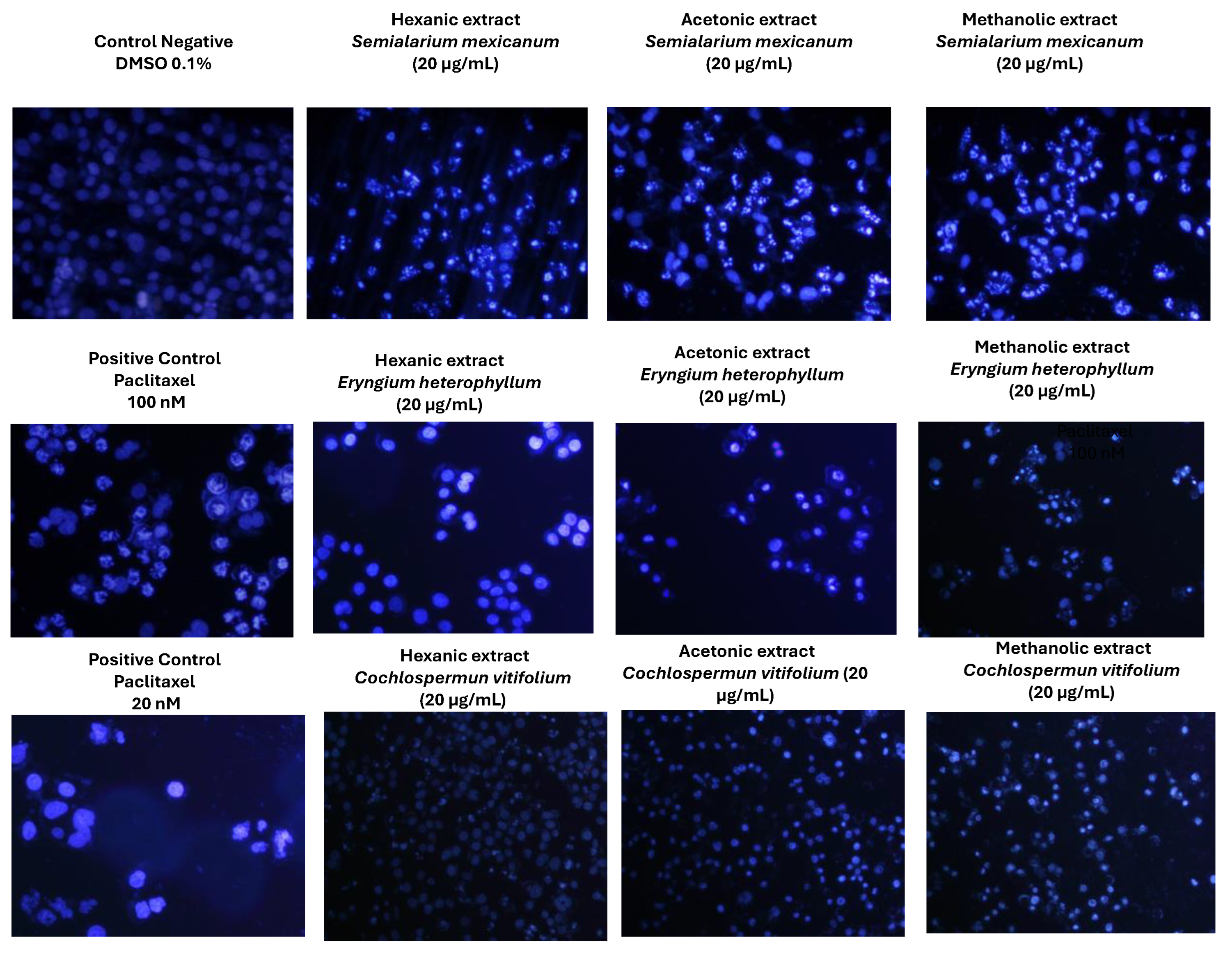
Figure 11.
Chemical structures of compounds 1–15 of hexane extract of S. mexicanum.
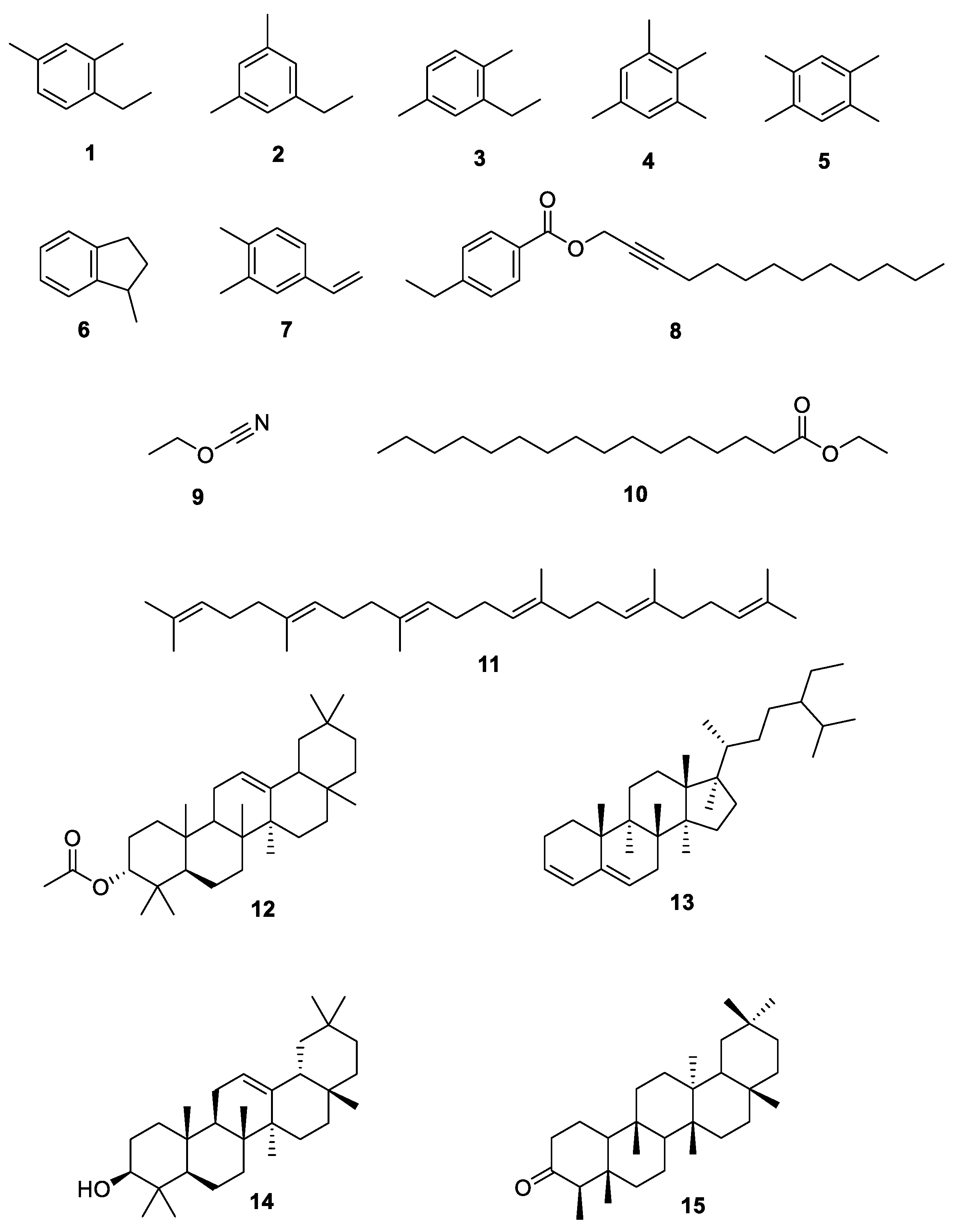

Table 1.
Taxonomic information, geographic origin, and collection parameters of the plant species under study.
Table 1.
Taxonomic information, geographic origin, and collection parameters of the plant species under study.
| Name | Family | Voucher | Plant organ used | Collection date | Collection site |
|
Cochlospermum vitifolium |
Bixaceae | 28904 | trunk and stem | 4/02/2024 | Lomas del Carril neighborhood of Temixco, Morelos |
|
Eryngium heterophyllum |
Apiaceae | 29010 | whole plant without roots | 6/02/2024 | State Of Mexico |
| Piper auritum | Piperaceae | 28935 | aerial plant parts | 7/02/2024 | Álvaro Leonel, La Joya, Morelos |
|
Semialarium mexicanum |
Celastraceae | 32942 | trunk and stem | 15/02/2024 | Los Naranjos, Guerrero |
Table 2.
Yields of hexanic, acetone and methanolic extracts of C. vitifolium, E. heterophyllum, P. auritum and S. mexicanum.
Table 2.
Yields of hexanic, acetone and methanolic extracts of C. vitifolium, E. heterophyllum, P. auritum and S. mexicanum.
| Plant species | Hexane | Acetone | Methanol | |||
| Extract (g) | Extract yields (%) | Extract (g) | Extract yields (%) | Extract (g) | Extract yields (%) | |
| Cochlospermum vitifolium | 0.73 | 0.49 | 1.92 | 1.28 | 12.08 | 9.39 |
| Eryngium heterophyllum | 1.92 | 1.28 | 3.36 | 2.24 | 14.71 | 9.81 |
| Piper auritum | 3.21 | 2.14 | 24.03 | 4.17 | 24.30 | 16.2 |
| Semialarium mexicanum | 1.71 | 1.14 | 6.90 | 4.60 | 14.29 | 9.53 |
Table 3.
Total Phenolic Content (mg GAE/g extract) of Methanolic and Acetonic Extracts from Semialarium mexicanum, Cochlospermum vitifolium, Eryngium heterophyllum, Piper auritum itum.
Table 3.
Total Phenolic Content (mg GAE/g extract) of Methanolic and Acetonic Extracts from Semialarium mexicanum, Cochlospermum vitifolium, Eryngium heterophyllum, Piper auritum itum.
| Extract |
Total phenolic content (mg GAE/g extract) |
| EM | 33.78 ± 1.26 |
| CA | 56.51 ± 0.46 |
| SM | 180.06 ± 1.55 |
| PM | 54.82 ± 1.27 |
| CA | 231.19 ± 2.00 |
| EA | 40.79 ± 0.82 |
| SA | 205.98 ± 2.87 |
| PA | 45.68 ± 1.02 |
Semialarium mexicanum methanolic extract = SM; Semialarium mexicanum acetonic extract = SA; Cochlospermum vitifolium methanolic extract = CM; Cochlospermum vitifolium acetonic extract = CA; Eryngium heterophyllum methanolic extract = EM; Eryngium heterophyllum acetonic extract = EA; Piper auritum methanolic extract = PM; Piper auritum acetonic extract = PA.
Table 4.
IC50 values (mean ± SD) of plant extracts and paclitaxel in human cancer cell lines. IC50 values for plant extracts are expressed in µg/mL, whereas paclitaxel values are expressed in nM. Data represent the mean ± SD of three independent experiments performed in triplicate.
Table 4.
IC50 values (mean ± SD) of plant extracts and paclitaxel in human cancer cell lines. IC50 values for plant extracts are expressed in µg/mL, whereas paclitaxel values are expressed in nM. Data represent the mean ± SD of three independent experiments performed in triplicate.
| Plant species | Extract | Cancerous cell lines | |||||
| HeLa | SiHa | HepG2 | PC-3 | H1299 | MCF-7 | ||
| Eryngium heterophyllum | Hexane | 24.58 ± 4.0 | 20.4 ± 2.4 | 34.0 ± 4.5 | 28.3±4.1 | 29.5 ± 7.5 | 32.7 ± 3.7 |
| Acetone | 29.35 ± 3.9 | 29.4 ± 3.5 | 32.6 ± 3.9 | 40.2± 6.9 | 45.7 ± 2.5 | 49.3 ± 3.5 | |
| Methanolic | 24.5 ± 3.6 | 27.45± 3.7 | 40.4 ± 3.7 | 29.8±5.7 | 65.7 ± 7.2 | 59.3 ± 3.7 | |
| Cochlospermun vitifolium | Hexane | 59.5 ± 2.59 | 55.78± 4.0 | 82.6 ± 2.4 | 68.4±2.6 | 78.5 ± 3.2 | 58.3 ± 6.1 |
| Acetone | 39.6 ± 4.1 | 42.3± 5 | 45.5 ± 5.1 | 32.1±3.7 | 62.1 ± 4.5 | 42.3 ± 2.4 | |
| Methanolic | 19.19± 3.3 | 20.45± 2.5 | 20.4 ± 2.3 | 30.1±3.2 | 35.1 ± 5.7 | 30.1 ± 6.2 | |
|
Semialarium mexicanum |
Hexane | 15.9 ± 1.8 | 17.16±2.8 | 24.7.± 2.7 | 23.8±4.5 | 38.02 ± 10.1 | 26.48 ± 6.4 |
| Acetone | 19.3 ± 2.8 | 21.78±3.2 | 37.0 ± 2.7 | 29.5±5.5 | 45 ± 4.3 | 29.4 ± 3.3 | |
| Methanolic | 21.2 ± 3.1 | 23.5 ± 2.7 | 24.4 ± 3.7 | 51.2±4.8 | 41 ± 7.2 | 66.8 ± 6.3 | |
| Piper auritum | Hexane | 220.2 ± 4.6 | 240.7 ± 7.1 | 199.5 ± 2.9 | 201.4±2.4 | 245 ± 20.7 | 289.9 ± 14.5 |
| Acetone | 230.6 ± 2.0 | 230.6 ± 2.0 | 212.7 ± 3.3 | 251.7±2.2 | 277.1 ± 27.4 | 256.2 ± 16.9 | |
| Methanolic | 50.7 ± 5.1 | 75.5 ± 5.1 | 160.4 ± 2.2 | 78.5±2.7 | 121.4 ± 14.5 | 135.78 ± 12.8 | |
| Paclitaxel nM | 15.7 ± 1.6 | 25.7 ± 2.2 | 68.7 ± 17.9 | 70.56 ±4.9 | 70.79 ± 2.7 | 108.52 ± 7.2 | |
Table 5.
IC50 values (mean ± SD) of the most cytotoxic extracts from Semialarium mexicanum and Eryngium heterophyllum in non-cancerous cell lines.
Table 5.
IC50 values (mean ± SD) of the most cytotoxic extracts from Semialarium mexicanum and Eryngium heterophyllum in non-cancerous cell lines.
| Plant species | Extract |
Human foreskin fibroblasts |
Primary uterine fibroblast cells | Immortalized human keratinocytes |
Embryonic kidney immortalized cell line |
|
| HFF-1 | HUF | HaCat | HeK-293 | |||
|
Semialarium mexicanum |
Hexane | 79.46±5.5 | 89.71± 4.3 | 53.53.±6.2 | 40.4 ±5.5 | |
| Acetone | 114.2±6.1 | 119.2±4.7 | 61.59 ±6.9 | 49.17±5.3 | ||
| Methanolic | 113.5±5.4 | 122.4 ±4.2 | 93.79 | 89.2±2.7 | ||
|
Eryngium heterophyllum |
Hexane | 117.3±5.9 | 75.8±5.1 | 58.85±6.0 | 41.67±6.7 | |
| Acetone | *ND | 113.9±3.7 | 104.0±3.2 | 75.41±6.1 | ||
| Methanolic | 108.5± 4.3 | 88.6±5.0 | 75.25±6.2 | 70.2±4.4 | ||
| Paclitaxel (nM) | *ND | 75.7±5.0 | 65.79±5.0 | 37.8±2.5 | ||
*ND:not determined.
Table 6.
SI values of Eryngium heterophyllum and Semialarium mexicanum extracts in HeLa and SiHa cells calculated using non-cancerous cell lines (HFF-1, HUF and HaCat as reference.
Table 6.
SI values of Eryngium heterophyllum and Semialarium mexicanum extracts in HeLa and SiHa cells calculated using non-cancerous cell lines (HFF-1, HUF and HaCat as reference.
| Plant species | Extract | HeLa | SiHa cells | ||||
| HFF-1 | HaCat | HUF | HFF-1 | HaCat | HUF | ||
|
Eryngium heterophyllum |
Hexane |
4.77 | 1.27 | 3.05 | 5.75 | 2.88 | 3.75 |
| Acetone | *ND | 3.5 | 3.87 | *ND | 3.5 | 3.8 | |
| Methanolic | 4.42 | 3.07 | 3.6 | 3.9 | 2.74 | 3.22 | |
|
Semialarium Mexicanum |
Hexane | 4.99 | 3.36 | 5.64 | 4.6 | 3.11 | 5.22 |
| Acetone | 5.92 | 3.19 | 6.17 | 5.24 | 2.82 | 5.48 | |
| Methanolic | 5.35 | 4.42 | 5.77 | 4.8 | 3.99 | 5.2 | |
| Paclitaxel | *ND | 4.18 | 4.82 | *ND | 2.55 | 2.94 | |
Table 7.
Chemical compounds identified in hexanic extract of S. mexicanum stem by GC-MS analysis.
| No | Name | *RT | Peak area % | Match % |
| 1 | Benzene, 1-ethyl-2,4-dimethyl- | 5.95 | 3.05 | 88.3 |
| 2 | Benzene, 1-ethyl-3,5-dimethyl- | 6.06 | 3.72 | 94.5 |
| 3 | Benzene, 2-ethyl-1,4-dimethyl- | 6.41 | 1.51 | 92.4 |
| 4 | Benzene, 1,2,3,5-tetramethyl- | 6.58 | 7.25 | 95.5 |
| 5 | Benzene, 1,2,4,5-tetramethyl- | 6.65 | 23.90 | 97.4 |
| 6 | Indan, 1-methyl- | 6.96 | 1.55 | 77.1 |
| 7 | Benzene,4-ethenyl-1,2-dimethyl- | 7.13 | 4.01 | 90.5 |
| 8 | 4-Ethylbenzoic acid, tridec-2-ynyl ester | 8.03 | 1.80 | 85.7 |
| 9 | Cyanic acid, ethyl ester | 10.94 | 2.29 | 84.9 |
| 10 | Hexadecanoic acid, ethyl ester | 19.47 | 0.50 | 78.8 |
| 11 | Squalene | 27.72 | 11.43 | 95.6 |
| 12 | 12-Oleanen-3-yl acetate, (3.alpha.)- | 29.07 | 3.77 | 79.6 |
| 13 | Stigmasta-3,5-diene | 29.69 | 2.23 | 76.7 |
| 14 | β.-Amyrone | 31.36 | 1.70 | 75.7 |
| 15 | Friedelan-3-one | 32.79 | 5.73 | 89.6 |
| RT = Retention Time* | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



