
Submitted:
08 January 2026
Posted:
09 January 2026
You are already at the latest version
Abstract
Dog assisted interventions (DAIs) are an established procedure to support military staff but their implementation during active warfare has not yet been systematically studied. Alongside, the welfare of therapy dogs participating in DAIs during war remains unexplored. Therapy dogs may develop clinically relevant emotional disorders, including trauma-related stress responses, analogous to human psychopathologies. The present study sought to monitor physiological arousal in therapy dogs performing DAI sessions with their handlers in two Ukrainian military hospitals (Vinnyzja and Kyiv). Thus, biomarkers of hypothalamic-pituitary-adrenal (HPA) axis activity including salivary, urinary and hair cortisol concentrations in Ukrainian (UA) therapy dogs were assessed to capture their acute and long-term stress responses. Moreover, cortisol levels from German (GE) therapy dogs, performing similar DAIs under peaceful conditions were gathered to compare cortisol levels between dogs. Results suggest that GE dogs exhibited significantly higher urinary and hair cortisol levels and significantly lower salivary cortisol concentrations, reflecting alterations in longer-term glucocorticoid secretion that is possibly caused by war-related stimulation in the UA cohort. In contrast, no significant differences in salivary cortisol emerged as a consequence of performing DAIs. The present findings suggest an environmental impact on therapy dogs’ cortisol secretion rather than involvement in DAIs.
Keywords:
therapy dog
; war
; PTSD
; mental health
; Ukraine
; dog-assisted intervention
; cortisol
; welfare
; stress
; dog-assisted services
1. Introduction
On February 24, 2022, the Russian Federation launched a full-scale invasion of Ukraine, initiating a prolonged armed conflict with profound humanitarian consequences. War and large-scale crises constitute complex, multidimensional challenges, producing enduring physical, psychological and emotional damages affecting not only human but also non-human animals [1,2,3] working and living in affected environments [3,4,5,6]. From a One Health perspective the tripartite system environment, human and non-human animal is deeply interlinked and essential when assessing mental health outcomes and welfare risks [7,8].
A substantial body of research examined the mental health outcome of war exposure in civilians, refugees and military personnel. The most prevalent disorders, described post-deployment or post-conflict, are post-traumatic stress disorder (PTSD), depression and anxiety [9,10,11,12]. Exposure to traumatic events involving threats to life or physical integrity markedly increase mental health disorders, with PTSD being the most prominent and debilitating [2,10,11]. Given its severity, chronicity and impact on functioning, PTSD warrants special attention in both human and non-human animal comparative health research. Empirical evidence addressing psychological stress during active warfare, particularly referring to animals, has so far remained scarce [13,14,15].
Studies manifest that dogs are capable of developing clinically relevant behavioral, emotional and physiological disorders analogous to human psychopathologies, including trauma-related stress responses [3,5,6]. Chronic stress exposure, cumulative trauma, social instability and insufficient recovery periods - hallmarks of war environments - are known contributors to anxiety disorders, impaired social functioning and avoidance behaviors in dogs [3,5,6]. Salden et al. (2023) [16] in their systematic review on the behavioral, emotional and psychological effects of exposure to high stress and/or traumatic environments described increased anxiety, hypervigilance and PTSD in affected dogs. War impacts humans and dogs alike, all experience and endure the invasion accompanying numerous stressors: noise, loss, stress, disruption to life, grief, pain, [5,6,17], creating deeply intertwined histories of trauma and adaptation strategies [2,17,18].
Animal suffering in war does not invariably derive from deliberate harmful human treatment; the worst outcomes frequently result from the structural violence inherent in the organized activity that represents war as such. Both endure trying circumstances with challenging travel, life and logistics [19,20]. The exigencies of war, however, do produce the most egregious examples of abuse, with thousands of dogs, horses or zoo animals involuntarily exposed to suffering and death, including mass execution [18,19,20]. Shorn of unexamined anthropocentric suppositions, the narrative of nonhuman animals in war unveils various facets of the nature of human relationships with other animals. However, we also realize that the human-dog partnership, derived from the needs and vulnerabilities shared, testifies to human dependence for this extra-species comfort [18]. The roles dogs had and have in war is wide-ranging: They have been mine detectors, trench guards, foodbearers, message or equipment carrier underground and across no-man’s land [20,21]. Patron, a terrier trained as an explosive detection dog is a symbol and mascot of Ukrainian resilience. The dog was awarded a medal for dedicated services by president Selenskyj in May 2022 [22]. However, dogs also serve as emotional support, confidant and comrade which illustrates both their instrumental value and exposure to harm [23]. Kateryna et al. (2023) demonstrated that companion animals significantly support civilians’ emotional regulation, reducing anxiety, feelings of helplessness, abandonment and fear during air raids and displacement [24]. Dogs provide constant companionship and physical closeness, routine and a sense of responsibility, helping individuals preserve psychological stability amid chaos. These findings align with broader evidence that dogs promote emotional regulation through non-judgmental presence, tactile comfort and reinforcement of safety and hope [24,25]. The bidirectional bond likely enhances resilience for both human and dog, although it might expose dogs to secondary or synchronized stress [26].
Within the military context the dog-human bond is particularly pronounced. Trust, reliance and interdependence between soldier and dog have historically enhanced operational effectiveness and psychological resilience [20,27,28]. Rethinking the roles of dogs in war brought about novel ideas and concepts: In the U.S., programs were developed in the early 2000, in which veterans with PTSD were being treated by pairing them with dog-human teams for goal-orientated dog-assisted interventions (DAIs) [27,28,29]. Widespread evidence supports the effectiveness of such programs [27,28], nonetheless, the implementation of DAIs during active armed conflict and its implication for canine stress, welfare and ethical responsibility has, to date, not been systematically examined.
From a One Health and welfare perspective it is critical to recognize that canine wellbeing is closely linked to the animal handler’s mental health [26,29,30]. Psychological distress in handlers has been shown to correlate with increased stress related behaviours in their dogs, underscoring the bidirectional nature of human–animal emotional transmission [26,30,31] and reflecting psychological and physiological spillover within human-dog teams. This interdependence raises ethical concerns where both human and dog participants are exposed to heightened and persistent stressors [4,26,30]. Therefore, evaluating stress responses in therapy dogs working in wartime environments is essential not only for safeguarding animal welfare but also for ensuring the sustainability and ethical integrity of DAIs. Addressing this gap will contribute to a more comprehensive understanding of a shared vulnerability, resilience and guidance across species in armed conflict.
Stress is an adaptive psycho-physiological response to actual or perceived threats, enabling an organism to efficiently cope with challenges. Initially, noradrenaline is secreted from the locus coeruleus in the brain stem to mobilize the body by facilitating actions that promote survival [17,32]. Moreover, the hypothalamic–pituitary–adrenal (HPA) axis is stimulated to release glucocorticoids to modulate metabolic rates. Behavioral consequences of these adaptations include heightened arousal and vigilance. Under acute conditions, these dynamics typically subside once the stressor is removed [32]. However, when stress persists over time in high intensity, or is poorly regulated, it can lead to chronic dysfunction of the HPA axis, causing prolonged elevations in cortisol which, in turn, precede adrenal exhaustion once the bodily resources are depleted [32]. Elevated stress levels have been identified as a key psychological consequence in humans during the ongoing Russian invasion of Ukraine [17,33,34].
Stress reactions to traumatic events can be acute or chronic. Acute stress involves intense emotional and physiological responses that occur within the first month following trauma exposure and gradually diminish [35]. For some individuals, however, distress persists or intensifies over time. Prolonged or repeated exposure to stressors such as violence, displacement, or bereavement substantially increase the threat of developing chronic psychological conditions, including PTSD [35,36]. Although these dynamics are well documented, little attention has been paid to dogs exposed to such stressors.
While most existing studies have examined the physiological and emotional states of therapy dogs in health care or education contexts under peaceful conditions [27,28], the present investigation analyzed data gathered during an enduring war situation. We sought to evaluate biomarkers of stress-related arousal in Ukrainian (UA) therapy dogs who teamed up with their handlers to visit PTSD-affected military personnel in Ukraine. Assessing complementary matrices of cortisol allows the assessment of both acute (saliva) and integrated/short-term (urine) HPA axis activation, whereas hair cortisol provides a longer-term index of chronic stress [37,38,39]. To provide a systematic monitoring of the HPA axis, we collected first-morning urine samples to calculate the urinary cortisol (UCC) and cortisol-to-creatinine ratio (UCCR). Furthermore we sampled saliva to measure salivary cortisol before and after DAIs. In addition, hair samples were analyzed from all participating dogs. German (GE) therapy dogs performing analogous DAI sessions under peaceful conditions were used as a control cohort.
The primary research question was: Do dogs working in DAIs with their handlers visiting PTSD-affected soldiers in a wartime setting show evidence of elevated physiological stress? How do acute (saliva), integrated (UCC) and long-term (hair) HPA-related stress measures compare with data gathered from a German cohort of therapy dogs? Secondary aims were to (a) describe inter-individual variability between dogs, (b) examine whether repeated saliva sampling reveals acute stress peaks that are not captured by morning UCC, and (c) discuss welfare implications for sustained deployment in conflict zones.
2. Materials and Methods
To demonstrate the effects of DAIs on glucocorticoid secretion in saliva, urine and hair in dogs from distinct living contexts, those in war-affected regions (Kyiv and Winnyzja, Ukraine) and dogs in a non-war environment (Germany), we employed a mixed design with both between-subject factors (Ukraine vs. Germany) and within-subject factors (pre- and post-intervention cortisol levels).
2.1. Therapy Dog Sample
Both, in person and online participant recruitment of human-dog teams were carried out in 2025 from January to June. Study respondents were invited by the authors via convenience sampling methodology based on personal invitation or online recruitment on social media. Inclusion criteria for UA and GE therapy dog handlers were adult age and either to have a valid DAI certificate awarded by an ISAAT- or ESAAT [40,41] recognized organization or currently attending an ISAAT- or ESAAT certified DAI education program.
2.2. Salivary Cortisol Analysis
Cortisol concentrations in canine saliva were determined on a working day with a scheduled DAI session. Thus, saliva samples were taken by the experimenter prior to interaction with recipients and immediately after the interaction using LABOklin saliva swabs (LABOklin, Germany). To minimize contamination, food was withheld from dogs for 20 minutes before saliva collection. To absorb saliva, the swab was gently inserted into the dog’s buccal cavity, targeting the lower gum line behind the last molar, where saliva tends to pool, and held in place for 30 seconds before being transferred to a collection tube. To estimate baseline variability, each dog was considered its own control. To determine salivary cortisol, an Enzyme-linked immunosorbent assay (ELISA) was used (LABOklin, Germany). Salivary cortisol is reported in µg·dL⁻¹.
2.3. Urine Cortisol Analysis
First naturally voided morning urine samples were collected by the handlers for cortisol analysis to minimize diurnal variation caused by DAI sessions. Samples were obtained by using a sterilized ladle and the urine was immediately transferred using a disposable pipette into a polypropylene tube. Minimum required volume was 0,5-2mL urine. Samples were handled with minimum delay and stored at -20 °C or lower in a freezer. To determine urinary cortisol an Enzyme-linked immunosorbent assay (ELISA) was used (LABOklin, Germany). Urinary cortisol is reported in nmol·L⁻¹.
2.4. Urine-Cortisol-Creatinine Ratio Analysis
To determine the Urine-Cortisol-Creatinine Ratio (UCCR), the collected morning urine was utilized and processed according to the upper protocol. To measure urinary creatinine, a standard enzymatic assay was used to account for urine concentration variability (LABOklin, Germany). The UCCR corrects for urine concentration, making it more reliable than raw cortisol measurements alone by reflecting integrated cortisol secretion over time. UCCR measures cortisol (and its metabolites) concentrations in urine normalized to creatinine, a muscle-derived metabolite. Dividing cortisol by creatinine, which is excreted at a stable rate, provides a concentration-independent standardized index. The ratio describes cortisol per unit of creatinine, making cortisol measurements samples comparable.
2.5. Hair Cortisol Analysis
A tuft of 2-centimeter-long hair was collected by the experimenter from each dog for cortisol analysis. Samples were retrieved from the base of the back of the neck of each dog by their handler. Collection of samples was done by cutting the hair as close to the skin as possible without damaging the skin [39,42]. The desired weight of the hair sample was 40 mg or more. Collected hair samples were wrapped in aluminum foil to protect them from sunlight and stored at room temperature in a dark environment. Subsequent extraction of cortisol was performed based on the protocol by Davenport, Tiefenbacher, Lutz, Novak, and Meyer (2006)[39]. HCC is reported in pg·mg⁻¹. To control for factors like coat colour and season, samples from all groups (Germany, Kyiv, Vinnyzja) were collected within similar time frames and seasons, ensuring comparable variation [42,43].
2.6. Statistics
All statistical analyses were performed using JASP software (version 0.18.3 for Windows; JASP Team, 2024). Graphical representations were also generated in JASP. Statistical significance was set at p < 0.05 for all analyses. Data were assessed for normality and homogeneity of variance using the Shapiro–Wilk and Levene tests, respectively. As the assumption of normality was violated, nonparametric tests were applied. Specifically, the Kruskal–Wallis ANOVA was used to compare groups, followed by Dunn’s post hoc test for pairwise comparisons. Protein and creatinine concentrations in urine, as well as the UCCR, were compared across all dog groups. Data are presented as medians and ranges. Group differences in categorical variables were analyzed using Fisher’s exact test, while differences in continuous variables were examined with the Kruskal–Wallis test. Cronbach’s alpha was calculated to assess the internal consistency reliability of the applied scales.
To analyze differences in stress markers (salivary cortisol [SCC], urinary cortisol [UCC], and hair cortisol [HCC]) and UCCR among dog groups at two levels—Level 1: Kyiv (KY) vs. Vinnyzja (WI) vs. Germany (GE), and Level 2: Ukraine (UA) vs. Germany (GE)—one-way ANOVAs were performed with group as the fixed factor and cortisol markers as dependent variables (α = 0.05). In cases of variance heterogeneity or violations of normality assumptions, Welch ANOVAs were applied. For post-hoc comparisons in the three-group analysis, Games-Howell tests were conducted.
Effect sizes were calculated as Cohen’s d for t-tests and partial eta-squared (ηₚ²) for ANOVAs. Following established conventions, Cohen’s d values of 0.20, 0.50, and 0.80 represent small, medium, and large effects, respectively [44]. Similarly, ηₚ² values of 0.01, 0.06, and 0.14 correspond to small, medium, and large effects, respectively [45,46].
3. Results
3.1. Demographics
As depicted in Table 1, the final sample consisted of n = 56 dogs. On average, dogs had a mean age of 5.23 years (SD = 4.02, range 1–17 years), with 27 female (48.2 %) and 28 male dogs (50 %), one dog’s sex had not been documented (1.8 %).
In Table 2, the UA therapy dogs cohort was divided into subgroups, resulting in overall k = 3 groups, of which n = 17 dogs were assessed in Kyjiw (KY dogs), n = 19 assessed in Vinnyzja (WI dogs) and n = 20 assessed in Germany (GE dogs).
In sum, the sample sizes did not differ significantly (F3groups (2,53) = 1.96, p = .15, η² = .07, F2groups (1,54) = 3.83, p = .06, η² = .07). Sex was equally distributed ((χ²3groups (2, N = 55) = 2.67, p = .26, V = .22, χ²2groups (1, N = 55) = .04, p = .84, V = .03), while the Shapiro-Wilk test for age was significant (Wyears = 0.84, p < .001; Wmonths = 0.85, p <.001).
Using Independent Samples T-Tests, we examined if the dog’s sex influenced cortisol markers. No significant differences could be found between male and female dogs in their cortisol levels, neither for their UCC, t(49) = 1.76, p = .20, d = 0.29, nor for their HCC, t(39) = 1.67, p = .10, d = 0.32, or the pre and post intervention values of the SCC, t(26) = 0.90, p = .38, d = 0.34, t(19) = 0.87, p = .40, d = 0.42.
3.2. Salivary Cortisol (SCC)
To examine potential group differences, time effects, and their interaction, mixed-design ANOVAs were conducted. All assumptions were tested and met. Sphericity was not a concern because the within-subjects factor included only two measurement points. Homogeneity of error variances was confirmed using Levene’s test (p > .05). The mixed-design ANOVA comparing the three groups (Kyiv [KY], Vinnyzja [WI], and Germany [GE]) revealed no significant main effect of time, F(1, 12) = 0.23, p = .64, ηₚ² = .02, indicating that salivary cortisol levels did not significantly change from before to after the intervention.
Figure 1.
Salivary cortisol concentrations (SCC, expressed in µg/dL) before (T1) and after (T2) a DAI session in UA therapy dogs (KY dogs and WI dogs) and GE therapy dogs. (GE (German), UA (Ukrainian), KY = Kyjiw, WI = Vinnyzja)).
Figure 1.
Salivary cortisol concentrations (SCC, expressed in µg/dL) before (T1) and after (T2) a DAI session in UA therapy dogs (KY dogs and WI dogs) and GE therapy dogs. (GE (German), UA (Ukrainian), KY = Kyjiw, WI = Vinnyzja)).
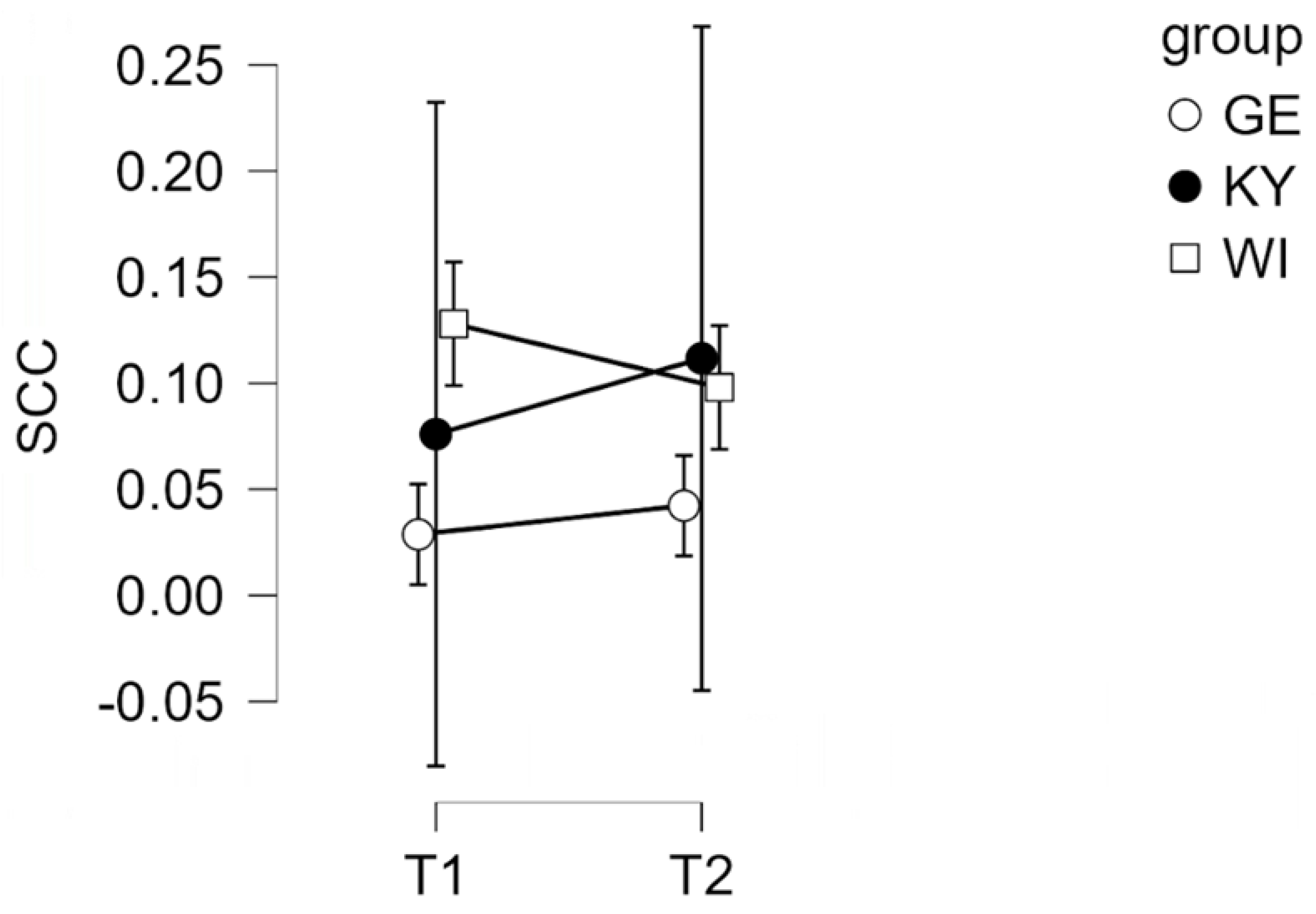
The main effect of group was not statistically significant, F(2, 12) = 3.11, p = .08, ηₚ² = .34, but showed a statistical trend toward group differences. No significant time × group interaction was found (p > .05), suggesting that changes in cortisol levels over time did not differ between groups.
3.3. Urinary Cortisol (UCC)
For the UCC, homogeneity of variances was asserted using Levene’s Test which showed that equal variances could be assumed for both levels of group comparisons (p3groups = .54, p2groups = .90). As shown in Figure 2, GE therapy dogs had the highest mean UCC levels (M = 187.9, SD = 78.57, n = 17), followed by WI dogs (M = 158.2, SD = 75.81, n = 18), and KY dogs showing the lowest levels (M = 123.4, SD = 63.15, n = 17). Post-hoc Tukey comparisons revealed that this effect was driven by a significant difference between the GE and KY groups (MDiff = 64.51, p = .03, d = .89), marked by a large effect size, with GE dogs exhibiting substantially higher UCC levels than those from Kyiv.
When the data were analyzed using a two-group comparison (Ukraine [KY + WI] UA vs. Germany [GE]), the one-way ANOVA also revealed a significant main effect with a medium effect size, F(1, 50) = 4.82, p = .03, ηₚ² = .09. As depicted in Figure 3, the data shows that dogs from Germany had significantly higher UCC levels (M = 187.9, SD = 78.57, n = 17) compared to dogs from Ukraine (M = 140.3, SD = 70.79, n = 35).
3.4. Urine-Cortisol-Creatinine Ratio analysis (UCCR)
UCCR values of the majority of dogs were within the physiologically acceptable reference range (<10-12), however, with substantial inter-individual variability. UCCR values ranged from 0.18 to 41.44 with a median of 8.98. Three individuals displayed a marked decrease and two dogs were extreme outliers with a marked increase (>33.46 and 41.44 ×10⁻⁶). Latter outliers reflect pathological variability indicating a substantial activation of the HPA axis and may be indicative of Hypercortisolism or severe and sustained stress exposure.
3.5. Hair Cortisol Concentrations (HCC)
HCC was assessed as a third and more long-term indicator of cortisol levels. Levene’s test revealed that the assumption of equal variances was violated for both levels of group comparisons (three-group comparison: p = .002; two-group comparison: p < .001). Consequently, Welch’s ANOVA and Games–Howell post-hoc tests were applied. The one-way Welch ANOVA comparing the three groups (Kyiv [KY], Winnyzja [WI], and Germany [GE]) did not show a significant main effect of location on HCC, F(2, 21.47) = 2.21, p = .13, ηₚ² = .11. In the absence of statistical significance, the pattern of group means followed the same direction as observed for UCC levels. GE dogs showed the highest mean HCC (M = 41.56, SD = 52.00, n = 16), followed by KY dogs (M = 18.17, SD = 5.04, n = 19), and WI dogs with the lowest levels (M = 12.58, SD = 11.18, n = 6), as illustrated in Figure 4.
Accounting for the absence of statistical significance, no further comparison between the UA and GE cohorts was calculated. Of note, a significant correlation was found between HCC and urinary cortisol-to-creatinine ratios (τ = 0.30, p < 0.001).
4. Discussion
The armed conflict in Ukraine has placed an unprecedented strain upon both civilians and military personnel, intensifying trauma related mental health factors. As one complementary approach, DAIs may be integrated alongside conventional care as a promising psychosocial tool drawing on the human-animal bond to support emotional regulation and reduce stress. Within the context of warfare, dogs may offer accessible, low-stigma support for individuals affected by trauma. DAIs represents an innovative approach within the One Health framework, however, effective interventions require careful consideration of the welfare of all participating individuals.
Previous studies have emphasized that dogs participating in therapeutic programs are exposed to both endogenous and exogenous stressors, which may deeply impact their behaviour, health and emotional wellbeing [47,48,49,73,78]. Dogs participating in DAI occupy a dual role: they provide psychosocial and physiological benefits to vulnerable humans, while themselves being exposed to the emotional and physical demands of the intervention setting [73]. Because animal welfare and handler–dog functioning are crucial for sustainable, ethical DAI deployment, objective physiological measures of stress in dogs are essential. Cortisol, as the primary glucocorticoid produced by the adrenal glands, has vital regulatory effects on physiology and metabolism. Secreted in high levels, it impacts glucose homeostasis, mediates stress adaptation mechanism and modulates immune functions, thereby modeling the dogs comprehensive metabolic state [32,50].
4.1. Salivary Cortisol (SCC)
To better understand the impact of DAI on canine welfare, salivary cortisol was measured in participating dogs as a physiological indicator of stress. Cortisol, a key hormone of the HPA axis, is central in regulating emotional arousal, energy mobilization and adaptive responses to environmental demands [32,50]. As a non-invasive measure it offers insight into neuroendocrine pathways, reflecting acute physiological and emotion states [71,72]. In our study, SCC levels did not change significantly pre- to post-intervention. This suggests that the activities involved did not elicit acute physiological stress responses in the participating dogs. Likewise, no significant interaction between time and group was observed, demonstrating comparable cortisol stability across the different locations. Although the main effect of the group was not statistically significant, the pattern of means indicated slightly higher SCC concentrations in GE dogs compared to UA dogs, suggesting potential contextual or environmental influences on baseline stress levels rather than intervention-related effects.
These results are consistent with previous studies reporting that animal-assisted interventions (AAI) and structured human–dog interactions do not typically elevate cortisol concentrations in well-adapted dogs [48,50,51]. Glenk et al. (2014) [52] similarly found that therapy dogs showed stable or even reduced salivary cortisol during animal-assisted sessions, suggesting that positive social engagement can buffer physiological stress responses. Other research also emphasizes the role of habituation and handler familiarity in maintaining low stress indicators during such interactions [29,41,53,54]. Overall, the absence of significant changes in salivary cortisol in the present study supports the interpretation that the intervention was not perceived as stressful by the dogs. Instead, it aligns with evidence that dogs accustomed to human interaction and structured tasks can remain physiologically stable during therapeutic or social settings.
4.2. Urinary Cortisol (UCC)
UCC represents another non-invasive biomarker of physiological stress in dogs that illustrates the activity of the HPA axis. When a dog experiences a physical or psychological stressor, the HPA axis is stimulated, resulting in the secretion of cortisol from the adrenal cortex [50,52]. Cortisol then circulates in the bloodstream and is metabolized and excreted in urine, permitting UCC to function as an integrated measure of systemic cortisol output over several hours prior to sampling [37,63,64].
Variations in UCC occur when an animal’s arousal or stress state shifts — for instance, during exposure to novel environments, social challenges, handling, or physical exertion. Elevated UCC levels usually specify amplified HPA axis activity, while stable or lower levels suggest habituation, calmness, or positive welfare states [59,63,64]. UCC provides a reliable and ethically adequate method for evaluating welfare in dogs [37,50,53] and as a welfare indicator, UCC is valued for capturing physiological responses that are not easily observable solely through behavior. UCC offers evidence of how dogs experience and cope with environmental or social conditions [57,58]. Studies frequently utilize UCC to evaluate whether participation leads to stress reduction, arousal, or stability [59,60].
In our study, GE dogs displayed the highest mean UCC, followed by WI and KY dogs. These results suggest cross-location variation in basal stress physiology rather than intervention-related effects given the absence of changes in SCC. Comparable findings have been reported in studies demonstrating that environmental context, training, daily routines, and handling practices can influence cortisol excretion in dogs [56,57,58] . Consistent with previous research, the absence of a significant time effect or interaction indicates that the intervention itself did not induce quantifiable stress responses [52,54,55]. Instead, the observed group alterations may reflect divergent management conditions, social environments, or levels of habituation to human interaction across study sites.
Although GE dogs showed significantly higher urinary cortisol levels than UA dogs, this difference may reflect contextual rather than interventional factors. Several explanations are plausible. GE dogs may experience more structured routines, enrichment and greater coaching, which can elevate baseline cortisol without indicating distress [49,52,55]. In contrast, dogs from Ukraine, particularly those living amid the ongoing war, may have adapted to chronic environmental challenges in ways that alter their stress physiology [55,56,57]. Prolonged or repeated exposure to unpredictable or threatening conditions can lead to hypoactivation of the HPA axis, resulting in lower basal cortisol concentrations — a phenomenon observed in both human and other animals under sustained adversity [59,60,61,62] .
Thus, the comparatively lower UCC levels in UA dogs may not automatically be interpreted as indicative of better welfare, but rather as a possible sign of physiological adaptation or blunted stress reactivity after prolonged exposure to stressors associated with wartime environments, such as noise, displacement, or reduced predictability. Conversely, higher UCC values in GE dogs may reflect greater stimulation or heightened arousal in more stable and controllable conditions. Future studies combining physiological, behavioral, and contextual data are essential to disentangle whether these differences reflect adaptive modulation or welfare disparities between populations living under markedly different socio-environmental circumstances.
UCCR is the accepted gold standard for quantifying proteinuria in veterinary medicine [63,64]. It is utilized to evaluate systemic conditions, making it a reliable parameter for longitudinal monitoring [64]. Furthermore, it is a non-invasive biomarker for assessing stress and diagnosing endocrine disorders in dogs. Creatinine, a small molecule formed as a byproduct of muscle metabolism, is mainly influenced by the amount of muscle mass and correlates positively with body weight. UCCR provides a complementary measure to salivary and urinary protocols and by normalizing cortisol excretion, the effects of urine dilution are minimized. According to Del Baldo et al. (2022), reference UCCR values for healthy dogs are approximately 3.38 × 10⁻⁶ (range: 1.11–17.32 × 10⁻⁶) [63]. UCCR values greater than 30 may indicate hyperadrenocorticism, stress, or non-adrenal illness [64]. Zeugwetter et al. (2010) reported median UCCR values of 9.6 × 10⁻⁶ (range: 3.9–88.2 × 10⁻⁶), 2.5 × 10⁻⁶ (1.0–8.2 × 10⁻⁶), and 14.7 × 10⁻⁶ (3.2–401.7 × 10⁻⁶) in healthy dogs [64].
Our UCCR data illustrate from a stress-physiology standpoint three distinct cortisol phenotypes. The majority of dogs display adaptive stress response levels within normal distribution reference limits. Dogs with markedly elevated UCCR levels display an excessive HPS axis activation and dogs with profoundly low UCCR levels show a suppressed stress response. These patterns underscores UCCR utility as screening biomarker for stress-related endocrine dysregulation, identifying hyper- as well as hypo-reactive cortisol phenotype deviations displaying maladaptive stress responses.
4.3. Hair Cortisol (HCC)
HCC is a relatively recent method used to assess physiological stress. It has to be taken into account that it varies with season, coat type, sex, age and growth of hair, as it takes a varied period of time before a new segment of hair is at the skin surface and can be measured [65]. Comparable to other freely circulating molecules, cortisol becomes incorporated into growing hair follicles and thereby generates a measurable plasma cortisol level. HCC levels intercorrelate with saliva and serum cortisol and make it a valuable tool for assessing long-term physiological stress. HCC evinces cumulative cortisol secretion over weeks to months, integrating repeated activation of the HPA axis into a stable, retrospective marker of chronic physiological stress [65,66]. Elevated HCC values can result from sustained extrinsic stimulation, social activity, or ongoing stress exposure, while lower concentrations may indicate either low stress load or HPA-axis downregulation following prolonged adversity [65,67,68].
The analysis of HCC revealed no statistically significant effect of location, although the mean values followed the same pattern observed for urinary cortisol. GE therapy dogs displayed the highest average HCC levels, followed by KY and WI dogs. The observed tendency toward higher HCC in GE dogs may reflect great exogenous engagement and activity levels or stressful living conditions [66,69,70]. Conversely, UA dogs’ lower HCC might indicate adaptive physiological modulation under chronic contextual strain such as exposure to unpredictable conditions related to the ongoing war, resulting in a blunted cortisol response over time [52,69,70,71].
At first glance, it might seem counterintuitive that dogs living in a war-affected country would show lower cortisol levels than those in peaceful environments. However, research in both human and animal physiology shows that chronic, prolonged stress can lead to hypoactivation of the HPA axis [52,67,70,72]. This adaptive process, known as HPA down-regulation or allostatic overload, reflects the body’s attempt to maintain stability under repeated or uncontrollable stress [72,73,74]. In the case of Ukrainian dogs, living for several years amid war may have led to such chronic adaptation. Continuous exposure to unpredictable noise, displacement, and reduced environmental stability could have altered HPA responsiveness, producing lower basal cortisol concentrations [75]. These findings are consistent with previous studies showing that HCC is sensitive to extrinsic and intrinsic factors but not necessarily a direct measure of negative stress [70,71,76]. Instead, HCC provides a long-term index of allostatic load, offering valuable insight into how dogs physiologically adjust to their living contexts.
Similar mechanisms have been described in dogs engaged in highly stressful DAI settings [74,75,77], where initial hypercortisolism is followed by a “burnout” phase with reduced cortisol output [73,77,78]. Lower HCC in chronically ill dogs for instance reflected HPA-axis adaptation or dysregulation rather than simply lower stress [76,78].
4.4. Handler Related Factors
Mitropoulos et al. (2025) demonstrated that HCC in dogs is not only impacted by dog-level variables like age or sex, but also by handler-related factors [75]. They found that older handlers were associated with higher HCC, suggesting handler demographics and handler-dog attachment influence long-term cortisol parameters in their dogs. Barcelos et al. (2024) in their review likewise emphasized the bidirectional nature of the dog-handler relationships, pointing out that handler stress and mental health challenges can exacerbate canine behavioral problems, thereby creating a negative feedback loop [76,77]. A One Health welfare approach is recommended addressing both, handler and dog wellbeing simultaneously and with an interdisciplinary collaboration [75,76,77].
With reference to handlers a vital aspect is the necessity of enhanced ethological knowledge and adequate stress perception. Erichsmeier et al. (2025) examined dog behaviour during DAIs in concurrence with handlers’ perceptions of their dogs’ stress [77]. They described that dogs displayed numerous observable stress related behaviors, indicating physiological or emotional arousal and decreased well-being, which may reflect increased allostatic load. Handler perception was incongruous, as they commonly described their dogs as not or only minimally stressed. This fundamental welfare risk of chronic and/or undetected elevated stress levels of dogs participating in DAIs likely results in a dysregulation of the HPA axis over time, provoking long-term welfare concerns [77,78]. Despite substantial behavioral indicators of discomfort, handlers frequently underestimate dogs' stress, which may result in delay or repress adequate rest or workload adjustment. This highlights that objective behavioral monitoring and targeted handler education is pivotal to canine well-being [77,78,79].
In this context, Cortesi et al. (2025) examined stress and burnout in dogs participating in DAIs, analyzing and contrasting it with the handler evaluation [78]. They observed a discrepancy in that handlers commonly recognized acute stress (avoidance behaviours, reduced cooperation) but were seldom able to identify chronic stress, emotional withdrawal, learned helplessness or burnout in their dogs [78]. These findings suggest that dogs partaking regularly in DAIs may face health and welfare risks.
One solution is enhanced and more effective monitoring of the handlers, assessment of high risk environments, optimized management strategies for handlers to protect their dogs, in particular regarding duration, frequency and recovery time of an intervention [50,79]. Given the recent research outcomes, reliance on handler opinion alone holds the distinct potential to underestimate cumulative stress and strongly emphasizes the need for standardized welfare guidelines and substantive training programs for all handlers [77,78,79].
4.5. Animal Welfare Strategies
Strategies to improve canine welfare even and especially during wartime and crisis may focus on providing environmental and social conditions that support both physical and psychological well-being. Environmental enrichment, including physical and cognitive stimulation, agency, and safe exploration, can promote engagement without causing over-arousal [79,80,81]. Predictable routines and safe spaces are essential and handling schedules can help reduce physiological stress. The quality of the human–dog relationship plays a key role, as caregiver training that encourages positive interactions and minimizes aversive handling can buffer stress responses and support healthy cortisol regulation [76,81,82]. Studies have shown that therapy dogs with handlers who manage stress effectively display lower cortisol reactivity in challenging settings [81,82], supporting the idea that improving handler well-being can directly enhance animal welfare in applied contexts. For instance, Roth et al. (2021) examined cortisol levels in dogs before and after owner-targeted stress-reduction programs, reporting modest decreases in canine HCC following improvements in owner well-being [83]. Longitudinal studies incorporating genetic markers, wearable stress monitors, and environmental assessments are expected to further clarify how shared environments and relationships influence this synchronization.
Recognizing this bidirectional relationship has significant implications for both animal welfare and human mental health [81,82,83,84]. Interventions aimed at reducing handler stress may indirectly benefit the dog. Mindfulness, non-aversive positive reinforcement-based training, structured exercise routines, and rituals have been associated with improved behavioral outcomes and reduced stress markers [81,82,84]. Additionally, programs that support owners—such as education on canine body language, consistent routines, and the promotion of secure attachment behaviors—may help to minimize stress transmission within the dyad.
5. Conclusions
Existing research has not yet yielded conclusive evidence regarding the explanatory pathways or the specific physiological mechanisms through which benefits of human-animal interactions are conferred. A more comprehensive synthesis of the available evidence is therefore essential to understand the potential impacts of human–dog interactions on both physical and psychological health, as well as to identify the underlying mechanisms that mediate these effects. Such insights are critical for developing evidence-based approaches to enhance a One Health Welfare approach.
Future research should combine behavioral, physiological, and environmental assessments to predict health outcomes and evaluate how interventions, such as enrichment and structured routines, modulate welfare over time. Longitudinal studies tracking dogs in high-stress contexts would help clarify whether observed cortisol patterns reflect adaptive physiological modulation or indicate hidden welfare risks.
5.1. Limitations and Future Directions
Limitations of this study include its cross-sectional design, which prevents causal inferences and potential biases arising from convenience sampling in both Ukrainian and German respondents. Given the exploratory nature of the study, the want of sample representativeness and limited generalizability should be acknowledged. Moreover, the small number of human-dog teams involved further constrains the applicability of the findings to broader populations. Nonetheless, the present results contribute valuable insights into glucocorticoid responses of therapy dogs in DAIs during crisis and warfare. To enhance generalizability, future studies should apply probability-sampling methods. Furthermore, longitudinal research designs are recommended to examine recovery trajectories, investigate attachment dynamics between handlers and dogs in conflict settings and evaluate whether structured interventions can mitigate the psychological and physiological burdens associated with wartime deployment over time.
Author Contributions
Conceptualization, S.F. and S.K.; methodology, S.F.; software, JABS; validation, S.F. and L.M.G; formal analysis A.D.H.; investigation, S.F.; resources, S.F.; data curation S.F.; writing—original draft preparation, S.F.; L.M.G writing—review and editing, S.F., L.M.G.; supervision, S.F.; project administration, S.F., S.K.; funding acquisition, S.F.. All authors have read and agreed to the published version of the manuscript.
Funding
This research received an external fund from Kynos-Stiftung, Germany https://kynos-stiftung.de/.
Institutional Review Board Statement
This study adhered to the ethical principles outlined in the Declaration of Helsinki and was approved by the appropriate institutional ethics review board of the Bioethics Commission of the National University of Life and Environmental Science of Ukraine protocol code 025/2024, date of approval 30.12.2024. All human participants provided informed consent prior to participation. Particular attention was given to the protection of vulnerable populations, including individuals affected by war and trauma.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Personal data is unavailable due to privacy or ethical restrictions.
Acknowledgments
“During the preparation of this manuscript/study, the author(s) used chatGTP, 2022 version for the purposes of citation, grammar and spelling. The authors have reviewed and edited the output and take full responsibility for the content of this publication.”.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Shvets, A.; Sereda, I.; Lopin, E. The medical and social importance of mental and behavior disorders among military personnel in peacetime and warfare. Rom. J. Mil. Med. 2021, 124, 481–486. [Google Scholar] [CrossRef]
- Krasnodemska, I.; Savitskaya, M.; Berezan, V.; Tovstukha, O.; Rodchenko, L. Psychological consequences of warfare for combatants: Ways of social reintegration and support in Ukraine. Amazon. Investig. 2023, 12, 78–87. [Google Scholar] [CrossRef]
- Alupo, C. Canine PTSD . Ph.D. Thesis, Swedish University of Agricultural Sciences (SLU), Skara, Sweden, 2017. [Google Scholar]
- Alger, J.M.; Alger, S.F. Canine soldiers, mascots, and stray dogs in U.S. wars: Ethical considerations. In Animals and War; Hediger, R., Ed.; Brill: Leiden, The Netherlands, 2013; pp. 77–104. [Google Scholar]
- Ksenofontova, A.A.; Voinova, O.A.; Ivanov, A.A.; Ksenofontov, D.A. Behavioral veterinary medicine: A new direction in the study of behavioral disorders in companion animals. Int. Trans. J. Eng. Manag. Appl. Sci. Technol. 2020, 11, 11A10O. [Google Scholar]
- Haq, A.U. Canine psychiatry: Addressing animal psychopathologies. Behav. Sci. 2017, 6, 7. [Google Scholar]
- Wells, D.L. Domestic dogs and human health: An overview. Br. J. Health Psychol. 2007, 12, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Liguori, G.; Costagliola, A.; Lombardi, R.; Paciello, O.; Giordano, A. Human–animal interaction in animal-assisted interventions (AAIs): Zoonosis risks, benefits, and future directions—A One Health approach. Animals 2023, 13, 1592. [Google Scholar] [CrossRef]
- Giacomuzzi, S.; Kocharian, A.; Barinova, N.; Barinov, S. Experiencing and coping with trauma in warfare and military conflicts. Psychol. Couns. Psychother. 2021, 16, 40–44. [Google Scholar] [CrossRef]
- Naeem, A.; Sikder, I.; Wang, S.; Barrett, E.S.; Fiedler, N.; Ahmad, M.; Haque, U. Parent–child mental health in Ukraine in relation to war trauma and drone attacks. Compr. Psychiatry 2025, 139, 152590. [Google Scholar] [CrossRef] [PubMed]
- Mottola, F.; Gnisci, A.; Kalaitzaki, A.; Vintilă, M.; Sergi, I. The impact of the Russian–Ukrainian war on the mental health of Italian people after two years of the pandemic: Risk and protective factors as moderators. Front. Psychol. 2023, 14, 1154502. [Google Scholar] [CrossRef]
- Murthy, R.S.; Lakshminarayana, R. Mental health consequences of war: A brief review of research findings. World Psychiatry 2006, 5, 25–30. [Google Scholar]
- Bienvenu, T.C.; Dejean, C.; Jercog, D.; Aouizerate, B.; Lemoine, M.; Herry, C. The advent of fear conditioning as an animal model of post-traumatic stress disorder: Learning from the past to shape the future of PTSD research. Neuron 2021, 109, 2380–2397. [Google Scholar] [CrossRef]
- Stucki, S. Animal warfare law and the need for an animal law of peace: A comparative reconstruction. Am. J. Comp. Law 2023, 71, 189–233. [Google Scholar] [CrossRef]
- Stucki, S. Beyond Animal Warfare Law: Humanizing the 'War on Animals' and the Need for Complementary Animal Rights; MPIL Research Paper 2021-10, Max Planck Institute for Comparative Public Law & International Law, 2021.
- Salden, S.; Wijnants, J.; Baeken, C.; Saunders, J.; De Keuster, T. Trauma and its behavioral aftermath: A systematic review of the impact of disaster deployment on working dogs. In Proceedings of the 5th European Veterinary Congress of Behavioural Medicine and Animal Welfare (EVCBMAW 2023); Vol. 9, No. 1; pp. 80–81.
- Sacoor, C.; Marugg, J.D.; Lima, N.R.; Empadinhas, N.; Montezinho, L. Gut–brain axis impact on canine anxiety disorders: New challenges for behavioral veterinary medicine. Vet. Med. Int. 2024, 2024, 2856759. [Google Scholar] [CrossRef]
- Hediger, R. Animals in war. In The Palgrave International Handbook of Animal Abuse Studies; Maher, J., Pierpoint, H., Beirne, P., Eds.; Palgrave Macmillan: London, UK, 2017; pp. 475–494. [Google Scholar] [CrossRef]
- Milburn, J.; Van Goozen, S. Animals and the ethics of war: a call for an inclusive just-war theory. Int. Relat. 2023, 37, 423–448. [Google Scholar] [CrossRef]
- Lemish, M.G. War Dogs: A History of Loyalty and Heroism; University of Nebraska Press: Lincoln, NE, USA, 2022. [Google Scholar]
- Tsouparopoulou, C.; Recht, L. Dogs and equids in war in third millennium BC Mesopotamia. In Fierce Lions, Angry Mice and Fat-tailed Sheep: Animal Encounters in the Ancient Near East; pp. 279–289.
- Ukraine, ua. Patron, the Ukrainian Bomb-Detecting Dog. Ukraine.ua, 2022. Available online: https://ukraine.ua/faq/patron-ukrainian-bomb-sniffing-dog/ (accessed on 08 December 2025).
- Walsh, E.A.; Meers, L.L.; Samuels, W.E.; Boonen, D.; Claus, A.; Duarte-Gan, C.; Normando, S. Human–dog communication: How body language and non-verbal cues are key to clarity in dog-directed play, petting and hugging behaviour by humans. Appl. Anim. Behav. Sci. 2024, 272, 106206. [Google Scholar] [CrossRef]
- Kateryna, M.; Trofimov, A.; Vsevolod, Z.; Tetiana, A.; Liudmyla, K. The role of pets in preserving the emotional and spiritual wellbeing of Ukrainian residents during Russian hostilities. J. Relig. Health 2023, 62, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Sandvik, K.B. The Ukrainian refugee crisis: Unpacking the politics of pet exceptionalism. Int. Migr. 2023, 61, 292–304. [Google Scholar] [CrossRef]
- Hunt, M.; Otto, C.M.; Serpell, J.A.; Alvarez, J. Interactions between handler well-being and canine health and behavior in search and rescue teams. Anthrozoös 2012, 25, 323–335. [Google Scholar] [CrossRef]
- Leighton, S.C.; Nieforth, L.O.; O’Haire, M.E. Assistance dogs for military veterans with PTSD: A systematic review, meta-analysis, and meta-synthesis. PLoS ONE 2022, 17, e0274960. [Google Scholar] [CrossRef]
- Rodriguez, K.E.; LaFollette, M.R.; Hediger, K.; Ogata, N.; O’Haire, M.E. Defining the PTSD service dog intervention: Perceived importance, usage, and symptom specificity of psychiatric service dogs for military veterans. Front. Psychol. 2020, 11, 519718. [Google Scholar] [CrossRef] [PubMed]
- Barcelos, A.M.; Kargas, N.; Assheton, P.; Maltby, J.; Hall, S.; Mills, D.S. Dog owner mental health is associated with dog behavioural problems, dog care and dog-facilitated social interaction: A prospective cohort study. Sci. Rep. 2023, 13, 21734. [Google Scholar] [CrossRef]
- Koskela, A.; Törnqvist, H.; Somppi, S.; Tiira, K.; Kykyri, V.L.; Hänninen, L.; Kujala, M.V. Behavioral and emotional co-modulation during dog–owner interaction measured by heart rate variability and activity. Sci. Rep. 2024, 14, 25201. [Google Scholar] [CrossRef] [PubMed]
- Fung, A.; Mok, P.; Fung, W.K. The rich history and evolution of animal-assisted therapy. J. Altern. Complement. Integr. Med. 2024, 10, 443. [Google Scholar] [CrossRef]
- Glenk, L.M.; Kothgassner, O.D. Life out of balance: Stress-related disorders in animals and humans. In Comparative Medicine; Jensen-Jarolim, E., Ed.; Springer: Cham, Switzerland, 2017; pp. 95–112. [Google Scholar] [CrossRef]
- Johnson, R.J.; Antonaccio, O.; Botchkovar, E.; Hobfoll, S.E. War trauma and PTSD in Ukraine’s civilian population: Comparing urban-dwelling to internally displaced persons. Soc. Psychiatry Psychiatr. Epidemiol. 2022, 57, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Minervini, G.; Franco, R.; Marrapodi, M.M.; Fiorillo, L.; Cervino, G.; Cicciu, M. Post-traumatic stress, prevalence of temporomandibular disorders in war veterans: Systematic review with meta-analysis. J. Oral Rehabil. 2023, 50, 1101–1109. [Google Scholar] [CrossRef]
- Schincariol, A.; Orrù, G.; Otgaar, H.; Sartori, G.; Scarpazza, C. Posttraumatic stress disorder (PTSD) prevalence: An umbrella review. Psychol. Med. 2024, 1, 1–14. [Google Scholar] [CrossRef]
- Eshel, Y.; Kimhi, S.; Marciano, H.; Adini, B. Predictors of PTSD and psychological distress symptoms of Ukraine civilians during war. Disaster Med. Public Health Prep. 2023, 17, e429. [Google Scholar] [CrossRef]
- Kaszycka, K.; Goleman, M.; Krupa, W. Testing the level of cortisol in dogs. Animals 2025, 15, 1197. [Google Scholar] [CrossRef]
- van der Laan, J.E.; Vinke, C.M.; Arndt, S.S. Evaluation of hair cortisol as an indicator of long-term stress responses in dogs in an animal shelter and after subsequent adoption. Sci. Rep. 2022, 12, 5117. [Google Scholar] [CrossRef]
- Ghassemi Nejad, J.; Ghaffari, M.H.; Ataallahi, M.; Jo, J.H.; Lee, H.G. Stress concepts and applications in various matrices with a focus on hair cortisol and analytical methods. Animals 2022, 12, 3096. [Google Scholar] [CrossRef]
- https://www.esaat.org/.
- https://isaat.org/.
- Davenport, M.D.; Tiefenbacher, S.; Lutz, C.K.; Novak, M.A.; Meyer, J.S. Analysis of endogenous cortisol concentrations in the hair of rhesus macaques. Gen. Comp. Endocrinol. 2006, 147, 255–261. [Google Scholar] [CrossRef]
- Bennett, A.; Hayssen, V. Measuring cortisol in hair and saliva from dogs: Coat color and pigment differences. Domest. Anim. Endocrinol. 2010, 39, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Packer, R.M.; Davies, A.M.; Volk, H.A.; Puckett, H.L.; Hobbs, S.L.; Fowkes, R.C. What can we learn from the hair of the dog? Complex effects of endogenous and exogenous stressors on canine hair cortisol. PLoS ONE 2019, 14, e0216000. [Google Scholar] [CrossRef]
- Cohen, S. Perceived stress in a probability sample of the United States; 1988. [Google Scholar]
- Martin, E.L.; Martinez, D.A. The effect size in scientific publication. Educ. XX1 2023, 26, 9–17. [Google Scholar]
- Leslie, J.R.; Stephens, M.A.; Fotopoulos, S. Asymptotic distribution of the Shapiro–Wilk W for testing normality. Ann. Stat. 1986, 14, 1497–1506. [Google Scholar] [CrossRef]
- Foltin, S.; Glenk, L.M. Current perspectives on the challenges of implementing assistance dogs in human mental health care. Vet. Sci. 2023, 10, 62. [Google Scholar] [CrossRef] [PubMed]
- Glenk, L.M.; Foltin, S. Therapy dog welfare revisited: A review of the literature. Vet. Sci. 2021, 8, 226. [Google Scholar] [CrossRef]
- Townsend, L.; Gee, N.R. Recognizing and mitigating canine stress during animal assisted interventions. Vet. Sci. 2021, 8, 254. [Google Scholar] [CrossRef]
- Glenk, L.M.; Kothgassner, O.D.; Stetina, B.U.; Palme, R.; Kepplinger, B.; Baran, H. Therapy dogs’ salivary cortisol levels vary during animal-assisted interventions. Anim. Welfare 2013, 22, 369–378. [Google Scholar] [CrossRef]
- Kateřina, K.; Kristýna, M.; Radka, P.; Aneta, M.; Štěpán, Z.; Ivona, S. Evaluation of cortisol levels and behavior in dogs during animal-assisted interventions in clinical practice. Appl. Anim. Behav. Sci. 2024, 277, 106321. [Google Scholar] [CrossRef]
- Glenk, L.M.; Kothgassner, O.D.; Stetina, B.U.; Palme, R.; Kepplinger, B.; Baran, H. Salivary cortisol and behavior in therapy dogs during animal-assisted interventions: A pilot study. J. Vet. Behav. 2014, 9, 98–106. [Google Scholar] [CrossRef]
- Beerda, B.; Schilder, M.B.H.; Van Hooff, J.A.R.A.M.; De Vries, H.W.; Mol, J.A. Behavioural and hormonal indicators of enduring environmental stress in dogs. Anim. Welfare 2000, 9, 49–62. [Google Scholar] [CrossRef]
- Haubenhofer, D.K.; Kirchengast, S. Physiological arousal for companion dogs working with their owners in animal-assisted activities and animal-assisted therapy. J. Appl. Anim. Welfare Sci. 2006, 9, 165–172. [Google Scholar] [CrossRef]
- Dreschel, N.A.; Granger, D.A. Methods of collection for salivary cortisol measurement in dogs. Horm. Behav. 2009, 55, 163–168. [Google Scholar] [CrossRef]
- Spinella, G.; Tidu, L.; Grassato, L.; Musella, V.; Matarazzo, M.; Valentini, S. Military working dogs operating in Afghanistan theater: Comparison between pre- and post-mission blood analyses to monitor physical fitness and training. Animals 2022, 12, 617. [Google Scholar] [CrossRef]
- Mormède, P.; Andanson, S.; Aupérin, B.; Beerda, B.; Guémené, D.; Malmkvist, J.; et al. Exploration of the hypothalamic–pituitary–adrenal function as a tool to evaluate animal welfare. Physiol. Behav. 2007, 92, 317–339. [Google Scholar] [CrossRef]
- Corte, S.M.; Olivier, B.; Koolhaas, J.M. A new animal welfare concept based on allostasis. Physiol. Behav. 2005, 92, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Mbiydzenyuy, N.E.; Qulu, L.A. Stress, hypothalamic–pituitary–adrenal axis, hypothalamic–pituitary–gonadal axis, and aggression. Metab. Brain Dis. 2024, 39, 1613–1636. [Google Scholar] [CrossRef] [PubMed]
- Lehrner, A.; Daskalakis, N.; Yehuda, R. Cortisol and the hypothalamic–pituitary–adrenal axis in PTSD. In Posttraumatic Stress Disorder; 2016; pp. 265–290. [Google Scholar]
- Guilliams, T.G.; Edwards, L. Chronic stress and the HPA axis. Standard 2010, 9, 1–12. [Google Scholar]
- Del Baldo, F.; Gerou Ferriani, M.; Bertazzolo, W.; Luciani, M.; Tardo, A. M.; Fracassi, F. Urinary cortisol-creatinine ratio in dogs with hypoadrenocorticism. J Vet Inter Med 2022, 36, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Zeugswetter, F.; Bydzovsky, N.; Kampner, D.; Schwendenwein, I. Tailored reference limits for urine corticoid: creatinine ratio in dogs to answer distinct clinical questions. Vet Record 2010, 167, 997–1001. [Google Scholar] [CrossRef]
- Mariti, C.; Russo, G.; Mazzoni, C.; Borrelli, C.; Gori, E.; Habermaass, V.; Marchetti, V. Factors affecting hair cortisol concentration in domestic dogs: A focus on factors related to dogs and their guardians. Animals 2025, 15, 1901. [Google Scholar] [CrossRef] [PubMed]
- Roth, L.S.V.; Faresjö, Å.; Theodorsson, E.; Jensen, P. Hair cortisol varies with season and lifestyle and relates to human interactions in German shepherd dogs. Sci. Rep. 2016, 6, 19631. [Google Scholar] [CrossRef]
- Heimbürge, S.; Kanitz, E.; Otten, W. The use of hair cortisol for the assessment of stress in animals. Gen. Comp. Endocrinol. 2019, 270, 10–17. [Google Scholar] [CrossRef]
- Pan, C.; Xu, S.; Zhang, W.; Zhao, Y.; Song, M.; Zhao, J. Investigating hair cortisol dynamics in German Shepherd Dogs throughout pregnancy, lactation, and weaning phases, and its potential impact on the hair cortisol of offspring. Domest. Anim. Endocrinol. 2025, 92, 106921. [Google Scholar] [CrossRef]
- Mârza, S.M.; Munteanu, C.; Papuc, I.; Radu, L.; Diana, P.; Purdoiu, R.C. Behavioral, physiological, and pathological approaches of cortisol in dogs. Animals 2024, 14, 3536. [Google Scholar] [CrossRef]
- Ghassemi Nejad, J.; Ghaffari, M.H.; Ataallahi, M.; Jo, J.H.; Lee, H.G. Stress concepts and applications in various matrices with a focus on hair cortisol and analytical methods. Animals 2022, 12, 3096. [Google Scholar] [CrossRef]
- Kusch, J.M.; Matzke, C.C.; Lane, J.E. Social buffering reduces hair cortisol content in black-tailed prairie dogs during reproduction. Behaviour 2023, 160, 735–751. [Google Scholar] [CrossRef]
- Kudielka, B.M.; Bellingrath, S.; Hellhammer, D.H. Cortisol in burnout and vital exhaustion: An overview. G. Ital. Med. Lav. Ergon. 2006, 28, 34–42. [Google Scholar]
- Corsetti, S.; Ferrara, M.; Natoli, E. Evaluating stress in dogs involved in animal-assisted interventions. Animals 2019, 9, 833. [Google Scholar] [CrossRef]
- Kaltenegger, H.C.; Marques, M.D.; Becker, L.; Rohleder, N.; Nowak, D.; Wright, B.J.; Weigl, M. Prospective associations of technostress at work, burnout symptoms, hair cortisol, and chronic low-grade inflammation. Brain Behav. Immun. 2024, 117, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Mitropoulos, T.; Andrukonis, A. Dog owners’ job stress crosses over to their pet dogs via work-related rumination. Sci. Rep. 2025, 15, 16887. [Google Scholar] [CrossRef] [PubMed]
- Barcelos, A.M.; Kargas, N.; Mills, D. The effects of dog behavioural problems on owner well-being: A review of the literature and future directions. Pets 2024, 1, 53–69. [Google Scholar] [CrossRef]
- Erichsmeier, R.; Arney, D.; Soonberg, M. Behavioral observations of dogs during animal-assisted interventions and their handlers’ perceptions of their experienced level of stress. J. Appl. Anim. Welfare Sci. 2025, 1, 1–12. [Google Scholar] [CrossRef]
- Cortesi, B.C.; Palestrini, C.; Buttram, D.; Mazzola, S.; Cannas, S. Stress and burnout in dogs involved in animal assisted interventions: A survey of Italian handlers’ opinion. J. Vet. Behav. 2025, 78, 63–69. [Google Scholar] [CrossRef]
- Horowitz, A. Considering the “dog” in dog–human interaction. Front. Vet. Sci. 2021, 8, 642821. [Google Scholar] [CrossRef]
- Horowitz, A. Inside of a Dog: What Dogs See, Smell, and Know; Simon and Schuster: New York, USA, 2025. [Google Scholar]
- Santaniello, A.; Garzillo, S.; Cristiano, S.; Fioretti, A.; Menna, L.F. The research of standardized protocols for dog involvement in animal-assisted therapy: A systematic review. Animals 2021, 11, 2576. [Google Scholar] [CrossRef]
- Fine, A.H.; Griffin, T.C. Protecting animal welfare in animal-assisted intervention: Our ethical obligation. Semin. Speech Lang. 2022, 43, 8–23. [Google Scholar] [CrossRef] [PubMed]
- Roth, L.S.; Faresjö, Å.; Theodorsson, E.; Jensen, P. Hair cortisol varies with season and lifestyle and relates to human interactions in German shepherd dogs. Sci. Rep. 2016, 6, 19631. [Google Scholar] [CrossRef] [PubMed]
- Sundman, A.S.; Van Poucke, E.; Svensson Holm, A.C.; Faresjö, Å.; Theodorsson, E.; Jensen, P.; Roth, L.S. Long-term stress levels are synchronized in dogs and their owners. Sci. Rep. 2019, 9, 7391. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
Urinary cortisol concentrations (UCC, expressed nmol·L⁻¹in ) in UA therapy dogs (KY dogs and WI dogs) and GE therapy dogs. (GE (German), UA (Ukrainian), KY = Kyjiw, WI = Vinnyzja)).
Figure 2.
Urinary cortisol concentrations (UCC, expressed nmol·L⁻¹in ) in UA therapy dogs (KY dogs and WI dogs) and GE therapy dogs. (GE (German), UA (Ukrainian), KY = Kyjiw, WI = Vinnyzja)).
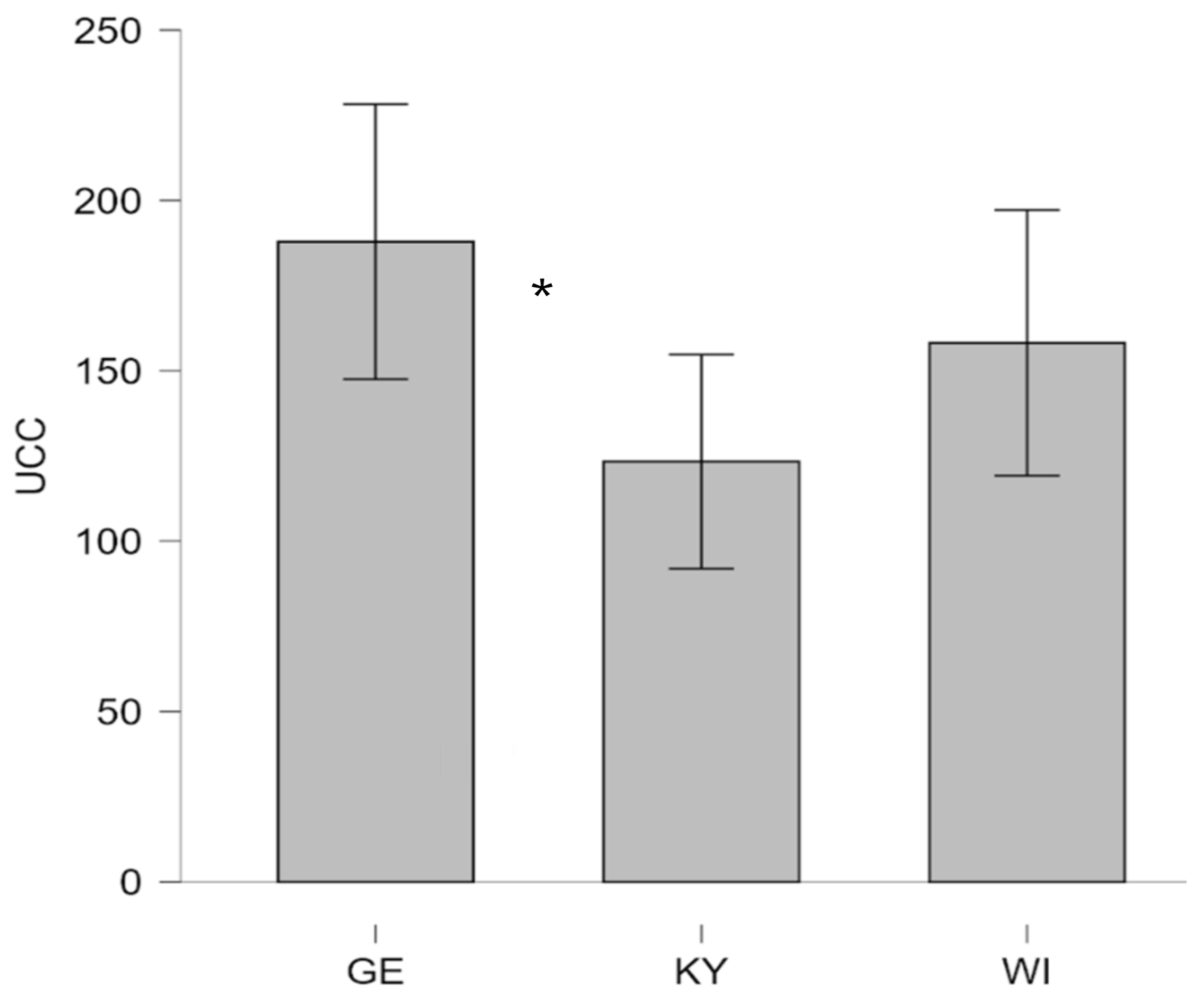
Figure 3.
Urinary cortisol concentrations (UCC, expressed nmol·L⁻¹in ) in UA therapy dogs and GE therapy dogs. (GE (German), UA (Ukrainian)).
Figure 3.
Urinary cortisol concentrations (UCC, expressed nmol·L⁻¹in ) in UA therapy dogs and GE therapy dogs. (GE (German), UA (Ukrainian)).
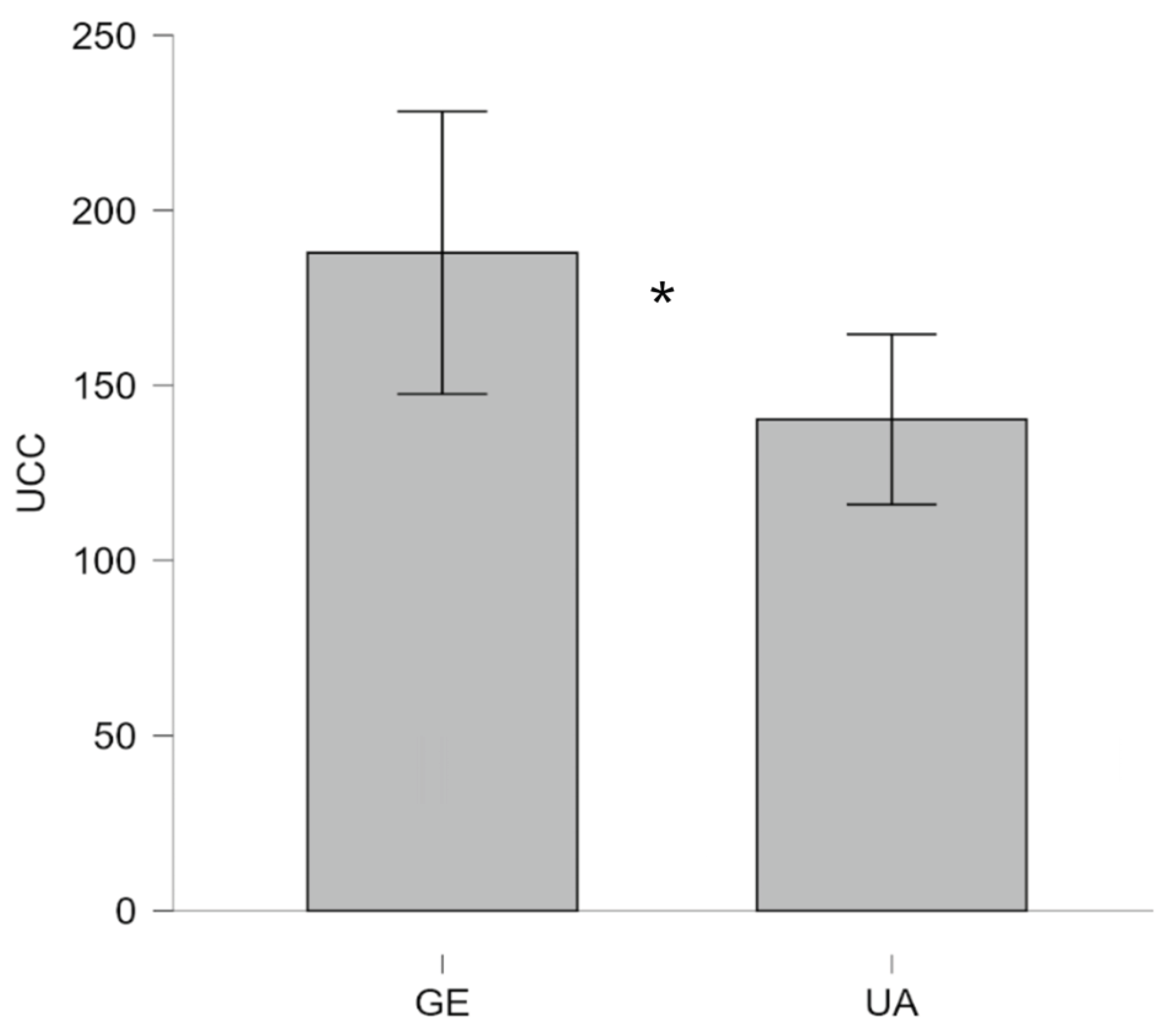
Figure 4.
Hair cortisol concentrations (HCC, expressed in pg·mg⁻¹) in UA therapy dogs (KY dogs and WI dogs) and GE therapy dogs. (GE (German), UA (Ukrainian), KY = Kyjiw, WI = Vinnyzja).
Figure 4.
Hair cortisol concentrations (HCC, expressed in pg·mg⁻¹) in UA therapy dogs (KY dogs and WI dogs) and GE therapy dogs. (GE (German), UA (Ukrainian), KY = Kyjiw, WI = Vinnyzja).
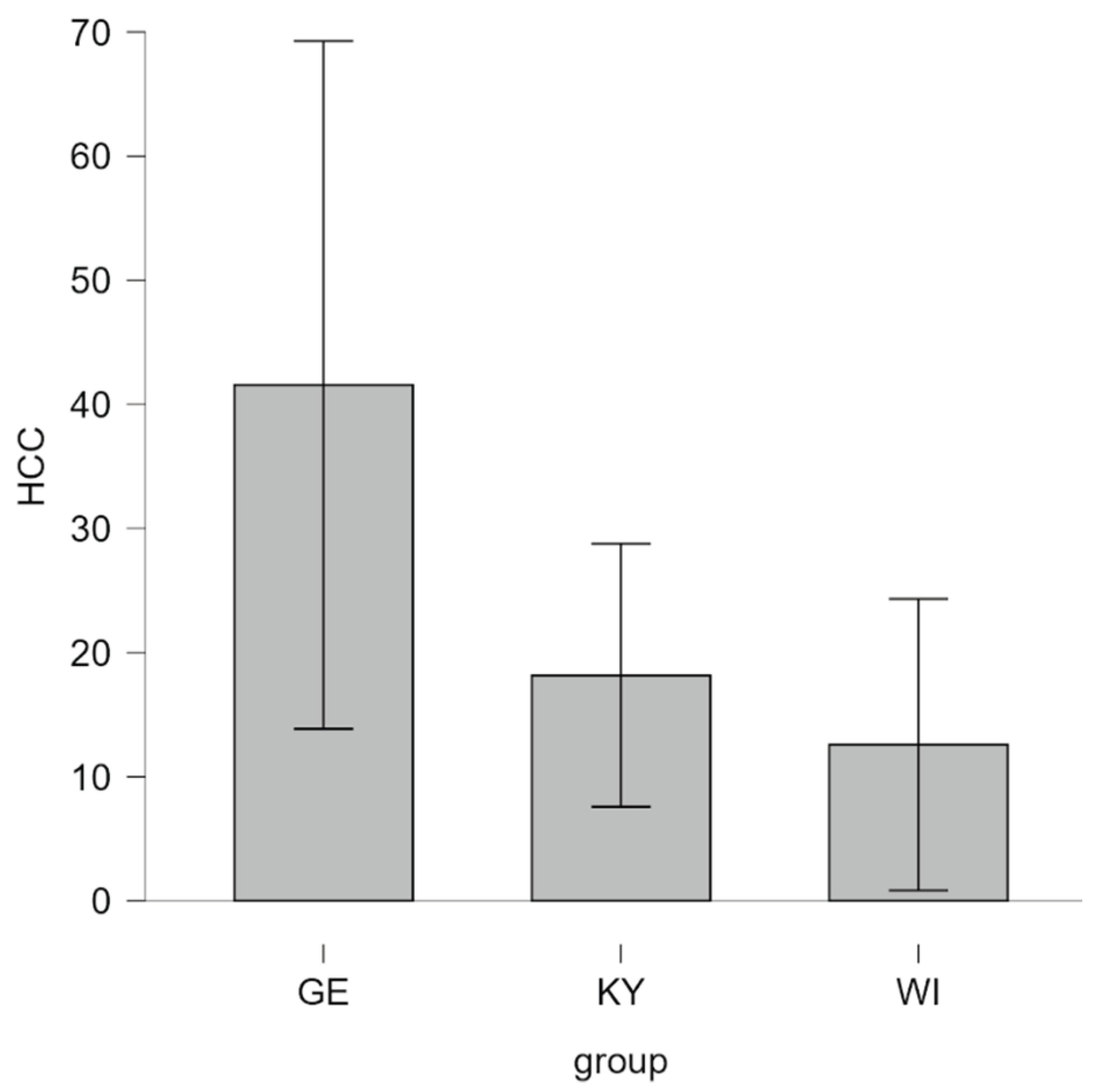

Table 1.
Age and sex of UA therapy dogs and GE therapy dogs. (GE (German), UA (Ukrainian)).
| GE | UA | GE | UA | ||||||||||||
| n | 17 | 39 | 17 | 39 | |||||||||||
| Age in years | Age in months | ||||||||||||||
| M | 3.765 | 5.872 | 48.06 | 74.46 | |||||||||||
| SD | 2.635 | 4.366 | 30.77 | 51.58 | |||||||||||
| W (Shapiro-Wilk) | 0.862 | 0.856 | 0.879 | 0.869 | |||||||||||
| p of Shapiro-Wilk | .017 | < .001 | .030 | < .001 | |||||||||||
| Minimum | 1.000 | 1.000 | 12.00 | 12.00 | |||||||||||
| Maximum | 11.00 | 19.00 | 134.0 | 233.0 | |||||||||||
| sex: female | n = 8 | n = 19 | |||||||||||||
| sex: male | n = 9 | n = 19 | |||||||||||||
| sex: missing | n = 1 | ||||||||||||||
Note. For Clarity, sex is given in the same table groupwise and independent of age.
Table 2.
Age and sex of UA therapy dogs (KY dogs and WI dogs) and GE therapy dogs. (GE (German), UA (Ukrainian; KY = Kyjiw, WI = Vinnyzja).
Table 2.
Age and sex of UA therapy dogs (KY dogs and WI dogs) and GE therapy dogs. (GE (German), UA (Ukrainian; KY = Kyjiw, WI = Vinnyzja).
| GE | KY | WI | GE | KY | WI | ||||||||
| n | 17 | 19 | 20 | 17 | 19 | 20 | |||||||
| Age in years | Age in months | ||||||||||||
| M | 3.765 | 5.579 | 6.150 | 48.06 | 71.47 | 77.30 | |||||||
| SD | 2.635 | 3.963 | 4.804 | 30.77 | 47.94 | 55.92 | |||||||
| W (Shapiro-Wilk) | 0.862 | 0.895 | 0.808 | 0.879 | 0.911 | 0.813 | |||||||
| p of Shapiro-Wilk | .017 | .040 | .001 | .030 | .079 | .001 | |||||||
| Minimum | 1.000 | 1.000 | 2.000 | 12.00 | 12.00 | 28.00 | |||||||
| Maximum | 11.00 | 17.00 | 19.00 | 134.0 | 206.0 | 233.0 | |||||||
| sex: female | n = 8 | n = 7 | n = 12 | ||||||||||
| sex: male | n = 9 | n = 12 | n = 7 | ||||||||||
| sex: missing | n = 1 | ||||||||||||
Note. For Clarity, sex is given in the same table groupwise and independent of age.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



