
Submitted:
06 January 2026
Posted:
07 January 2026
You are already at the latest version
Abstract
Addiction is a neuropsychiatric disorder characterised by compulsive substance use despite harmful consequences. Ketamine a dissociative anaesthetic increasingly misused among young people has become a global public health concern, necessitating the search for effective neuroprotective interventions. N-acetylcysteine (NAC) a glutathione precursor with antioxidant and anti inflammatory properties, has shown promise in mitigating substance-induced neurotoxicity. This study investigated the neuroprotective effects of NAC on ketamine induced cerebellar alterations in Wistar rats. Sixty adult Wistar rats (120–150 g) were randomly assigned to six groups. Group A received distilled water (control); Groups B and C received NAC (500 or 1000 mg/kg, orally) Group D received ketamine (15 mg/kg, intraperitoneally) while Groups E and F received ketamine followed by NAC (500 or 1000 mg/kg, respectively). Ketamine was administered for 10 days followed by NAC treatment from days 11 to 24. Behavioural assessments including open-field Y-maze, and catalepsy tests, were conducted on day 25. Animals were then euthanised for biochemical analyses of total antioxidant capacity (TAC) malondialdehyde (MDA), tumour necrosis factor-alpha (TNF-alpha), and interleukins IL-1 beta, IL-6, and IL-10. Cerebellar tissues were processed for histological evaluation. Ketamine exposure induced hyperlocomotion, increased rearing, working memory deficits, oxidative stress, and elevated pro-inflammatory cytokines, with a concomitant reduction in anti-inflammatory markers. NAC treatment at both doses significantly attenuated these behavioural and biochemical disturbances. Histological examination revealed marked cerebellar neurodegeneration, including Purkinje and granule cell loss, in ketamine-treated rats, whereas NAC particularly at 1000 mg/kg largely preserved cerebellar cytoarchitecture. In conclusion, NAC exerted significant neuroprotective effects against ketamine-induced behavioural, biochemical, and structural cerebellar damage in rats, supporting its potential therapeutic relevance in mitigating ketamine-related neurotoxicity.
Keywords:
antioxidant
; neuroinflammation
; neuroprotection
; N-methyl-D-aspartate antagonist
1. Introduction
Addiction is a chronic neuropsychiatric disorder characterised by a compulsive drive to use substances despite harmful consequences [1,2,3]. Among commonly abused drugs worldwide, ketamine has attracted increasing attention due to its anaesthetic, dissociative, and reinforcing properties [4,5,6]. Although originally developed for clinical use, persistent and excessive ketamine consumption has been linked to multisystem toxicity, including significant neurobiological alterations [7,8]. Preclinical studies have consistently demonstrated that even subanaesthetic doses of ketamine can induce marked neurobehavioural and neuromorphological changes [9,10,11]. While ketamine use disorder is less prevalent than opioid or psychostimulant dependence, chronic misuse is associated with enduring cognitive and psychological impairments, particularly deficits in memory and executive function [12]. These disturbances may resemble features of psychiatric conditions such as schizophrenia, although the underlying mechanisms differ [7,9,10,11]. Emerging evidence further suggests that prolonged ketamine misuse may exacerbate pre-existing mental health disorders, including psychosis and schizophrenia spectrum conditions [1,13].
Recreational ketamine use typically occurs via intranasal, oral, smoked, or injectable routes, often to induce euphoria, hallucinations, and dissociative states [7]. These dissociative effects manifest as depersonalisation, derealisation, and distortions in time, space, and bodily perception [14,15]. Experimental studies show that repeated exposure to subanaesthetic doses results in persistent neurocognitive impairments affecting attention, memory, and executive processes—domains also disrupted in animal models of schizophrenia [9,10,11]. Although ketamine is generally safe when used under controlled clinical conditions, non-medical use carries substantial neurological and societal risks. Its neurotoxicity has been linked to dysregulation of N-methyl-D-aspartate receptors, leading to calcium overload, mitochondrial dysfunction, oxidative stress, and apoptotic cell death [16]. These mechanisms underscore the need to identify interventions capable of counteracting ketamine-induced neural injury.
N-acetylcysteine (NAC) is a widely available therapeutic agent, clinically used in acetaminophen toxicity and as a mucolytic [17]. Beyond these applications, NAC exhibits potent antioxidant, anti-inflammatory, neuroprotective, and glutamatergic modulatory properties [9,18]. Importantly, NAC crosses the blood–brain barrier, increases brain glutathione levels, and helps restore glutamate homeostasis [19,20]. Through these mechanisms, NAC has emerged as a promising candidate for the treatment of neuropsychiatric and substance-use disorders [19,20,21]. Taken together, these observations highlight the need to investigate N-acetylcysteine as a potential therapeutic strategy for mitigating ketamine-induced neurotoxicity and its associated neurobehavioural disturbances.
2. Materials and Methods
2.1. Chemicals and Drugs
N-Acetylcysteine (1000mg, Swanson Health Products, Fargo, ND 58104 USA), ketamine injection (50mg/10ml, Swiss Parenterals Ltd), normal saline, distilled water, Assay kits for Malondialdehyde, Total antioxidant capacity, Interleukin-1β, Interleukin-6 (Biovision Inc., Milpitas, CA, USA).
2.2. Animals
Wistar rats weighing 120–150 g were procured from Empire Breeders, Osogbo, Osun State, Nigeria. The animals were housed in groups of six within wooden cages placed in temperature-controlled quarters (22–25 °C) under a 12-hour light/dark cycle. All rats were maintained on commercially available standard rodent chow (caloric composition: 29% protein, 13% fat, 58% carbohydrate) from weaning and had unrestricted access to food and water, except during behavioural assessments. All experimental procedures were carried out in accordance with approved institutional guidelines and conformed to the provisions of the European Council Directive (EU 2010/63) on the protection of animals used for scientific purposes.
2.3. Experimental Methodology
Sixty young adult, male Wistar rats (120–150 g) were randomly assigned into six groups (n = 10) and fed standard rodent chow, with daily feed intake recorded. Group A received distilled water (2 ml/kg, i.p, and 10ml/kg orally). Rat in groups B and C were administered distilled water i.p and NAC orally at 500 and 100 mg/kg respectively. Group D (Ketamine control) was administered ketamine (15 mg/kg, i.p.) [9,10,11] and distilled water orally, while rats in groups E and F received ketamine i.p. and NAC at 500 and 1000 mg/kg, orally, Intraperitoneal injection of distilled water or ketamine was from days 1-10 while oral administration of distilled water or NAC was from days 11-24. Body weight was measured weekly using a sensitive scale balance. On day 25, animals were exposed to the behavioural paradigm including; open field and Y-maze. On day 26, animals were euthanized and blood collected via cardiac puncture was used for the assessment of oxidative stress and inflammatory markers (TAC, MDA, TNF-α, IL-6, IL-10, IL-1β). The cerebellum as sectioned and processed for general histology using H&E and Cresyl Fast Violet staining.
2.4. Open Field Novelty Induced Behaviours
The open-field test assesses exploratory activity, anxiety-related behaviours as well as arousal, inhibition and investigatory activity. It is also useful for evaluating stereotypic behaviours such as grooming. In this study, grooming, rearing, and horizontal locomotion were recorded in the open-field arena over a 10-minute period. Spontaneous locomotor activity was measured in a 60 × 60 × 40 cm activity cage with a floor divided into 16 equal squares marked in blue and painted white. Recorded parameters included horizontal locomotion (number of floor units traversed with all paws), rearing frequency (number of times the rat stood on its hind limbs, with or without forelimb support on the cage wall), and grooming frequency (body-cleaning actions such as paw and face washing, body licking, and pubic grooming indicative of stereotypy). Behavioural scoring was conducted over 10 minutes [22,23]. The apparatus was cleaned with 70% ethanol between animals to eliminate olfactory cues.
2.5. Y maze Spatial Working Memory
The Y-maze spatial working memory test exploits the innate tendency of rodents to explore novel environments. The apparatus consists of a wooden maze with three identical arms arranged at 120° angles to form a “Y” shape. Each arm measures approximately 15 inches in length and 3.5 inches in width, with walls about 3 inches high. Each rat was placed in one arm of the maze and allowed to explore freely, with arm entries recorded when the animal’s tail had completely entered the next arm. The sequence of arm entries was documented as previously described [23].
2.6. Biochemical Tests
2.6.1. Estimation of MDA Content (Lipid Peroxidation)
The 2-Thiobarbituric Acid Reactive Substances (TBARS) assay was used to quantify lipid peroxidation products, including lipid hydroperoxides and aldehydes, which increase under oxidative stress. Results were expressed as malondialdehyde (MDA) equivalents, a major decomposition product of polyunsaturated fatty acid peroxides. In this assay, MDA reacts with 2-thiobarbituric acid at 25 °C to form a pink chromophore with a maximal absorbance at 532 nm, required reagents included 2-thiobarbituric acid (indicator), a 10% acid solution in dimethyl sulfoxide (acid reagent), and a 10 mM malondialdehyde tetrabutylammonium salt (MDA standard), prepared in a 96-well microplate format. Blood samples were collected and processed immediately. A saturated ammonium sulfate solution (100 µL) was added to 0.5 mL of plasma in a microcentrifuge tube, followed by the addition of 35 mg of trichloroacetic acid (TCA). The mixture was vortexed until a cloudy precipitate formed, then centrifuged, and the supernatant transferred to a clean tube. For the assay, 200 µL of either the standard or sample was mixed with 200 µL of the indicator solution. The blank consisted of 200 µL of sample and 200 µL of the acid reagent. All tubes were incubated at room temperature for 45 minutes. After incubation, 150 µL from each tube was transferred into a microplate, and absorbance was read at 532 nm [11].
2.6.2. Antioxidant Status
Total antioxidant capacity was measured using the Trolox Equivalent Antioxidant Capacity Assay that is based on the ability of antioxidants within a sample to react with oxidised products as previously described by [24].
2.6.3. Inflammatory Markers
Interleukin1β, IL-6, IL-10 and TNF level was assayed using enzyme-linked immunosorbent assay (ELISA) techniques with commercially available kits (Enzo Life Sciences Inc. NY, USA).
2.7. Tissue Histology
Rat brains were dissected, sectioned and fixed in neutral buffered formol-calcium. The cerebellum was processed for paraffin-embedding, cut at 5 µm and stained with haematoxylin and eosin and cresyl fast violet stains as previously described [25]
2.8. Photomicrography
Histological slides of the cerebellum were examined under an Olympus binocular light microscope. Images were captured using a Canon PowerShot 2500 Digital camera.
2.9. Statistical Analysis
Data was analysed using Chris Rorden’s ANOVA for Windows (version 0.98). Analysis of data was by One-way analysis of variance (ANOVA) and a post-hoc test (Tukey HSD) was used. Results were expressed as mean ± S.E.M and p < 0.05 was taken as the accepted level of significant difference.
3. Results
3.1. Effect of N-Acetyl Cysteine on Body Weight and Feed Intake
Figure 1 and Figure 2 illustrate the effects of N-acetylcysteine (NAC) on relative changes in body weight and feed intake, respectively, in ketamine-treated rats. There was a significant increase in body weight with KET (group D) and KET/NAC 1000MG (group F), compared to control (group A). Compared to KET (group D), body weight increased in KET/NAC1000 (group F).
In Figure 2, feed intake increased significantly (p < 0.05) with NAC500 (group B), NAC1000 (group C), KET (group D), KET/NAC500 (group E), and KET/NAC1000 (group F) compared to control (group A). Compared to KET (group D), feed intake increased significantly (p < 0.05) with KET/NAC500 (group E) and KET/NAC 1000 (group F). Over the treatment period, percentage change in feed intake increased with KET (184.8g) compared to control (115.6g). Compared to KET (184.8g), feed intake increased with KET/NAC500 (208.4g), and KET/ NAC1000 (211.4g), respectively
3.2. Effect of N-Acetyl Cysteine on Exploratory Behaviours in the Open-Field Box
Figure 3 and Figure 4 show the effects of NAC on horizontal locomotion and rearing activities respectively, in ketamine-treated rats. Figure 3 presents the effect of quercetin on horizontal locomotor activity, measured as the number of line crossings over a 10-minute period. There was a significant increase with KET compared to control (p< 0.05). Compared to KET, horizontal locomotion increased significantly (p< 0.05) with KET/NAC500, and decreased with KET/NAC1000.
Figure 4 shows the effect of quercetin on rearing activity, measured as the number of rearings within a 10-minute period. Rearing activity increase significantly (p<0.05) with KET compared to control. Compared to KET, rearing activity increased significantly (p< 0.05) with KET/NAC500, and decreased with KET/NAC1000 respectively.
3.3. Effect of N-Acetyl Cysteine on Self-Grooming Behaviour
Figure 5 shows the effect of quercetin on self-grooming behaviours in ketamine-treated rats. There was significant decrease with NAC/500, NAC/1000 and increase with KET, and KET/NAC500 compared to control. Compared to KET, self-grooming increased with KET/NAC500 and decrease with KET/NAC1000 respectively.
3.4. Effect of N-Acetyl Cysteine on Spatial Working Memory
Figure 6 shows the effect of NAC on spatial working memory in the Y- maze. Spatial working memory scores measured as % alternation/5 min increased significantly (p < 0.001) in groups administered NAC/500, NAC/1000 and KET/NAC 1000 and decreased with KET, and KET/NAC500 compared to control. Compared to KET, working memory scores increased significantly with KET/NAC1000
3.5. Effect of N-Acetyl Cysteine on Oxidative Stress Parameters and Inflammatory Cytokines
Table 1 shows the effect of NAC on oxidative stress parameters and levels of inflammatory cytokines in bromocriptine treated rats. Lipid peroxidation measured as Malondialdehyde (MDA) concentration increased significantly with KET compared to control. Compared to KET, the MDA levels decreased significantly (p<0.05) with KET/NAC500 and KET/NAC1000. Total antioxidant capacity (TAC) exhibited a visual increase with KET compared to control. Compared to KET, level of TAC increased significantly (p<0.05) with KET/NAC500, and decreased visually with KET/NAC1000.
Table 2 shows effect of NAC on levels of pro- and anti-inflammatory cytokines in ketamine treated rats. Levels of pro-inflammatory cytokines (IL-6, IL-1β, TNF-ᾱ) increased significantly with KET compared to control. While there was a significant decrease with KET/NAC500 and KET/NAC1000 compared to KET. Level of anti-inflammatory cytokine (IL-10), decreased significantly with KET compared to control and increased significantly with KET/NAC500 and KET/NAC1000 compared to KET.
3.6. Effect of N Acetyl Cysteine on the Cerebellar Cortex Histomorphology
Figure 7 (A-F) and 8 (A-F) show representative photomicrographs of haematoxylin and eosin (H&E) and cresyl fast violet (CFV) stained sections of the rat cerebellar cortex respectively. Examination of H&E-stained sections from groups A, B and C showed well-defined layers of the cerebellar cortex. Within the neuropil, multipolar Purkinje cells with large vesicular nuclei, granule neurons with open-faced nuclei and scant cytoplasm, and small molecular layer cells were clearly visible (Figure 7A, B & C). The neuropil, which appears pink in the H&E sections, was well preserved in the CFV-stained slides (Figure 8A, B & C).
In the groups treated with KET/NAC500 (Figure 7E, 8E) and KET/NAC1000 (Figure 7F, 8F), Purkinje cells with normal morphology were observed among granule and molecular layer cells in both H&E and CFV stained sections. In contrast, the group administered ketamine at 15 mg/kg (Figure 7D, 8D) exhibited numerous molecular layer cells, along with degenerating Purkinje and granule cells interspersed with some normal Purkinje and granule cells. Signs of neuronal injury included pale-staining granule cells with shrunken nuclei, indicating ongoing neuronal degeneration.
Discussion
The present study examined the modulatory effects of N-acetylcysteine (NAC) on ketamine-induced behavioural alterations, oxidative stress, neuroinflammation, and cerebellar histomorphology in adult rats. Overall, the findings demonstrate that NAC—particularly at 1000 mg/kg—significantly attenuates ketamine-associated neurobehavioural and biochemical disturbances, with clear dose-dependent effects across several outcome measures.
Ketamine administration significantly increased body weight and feed intake relative to controls. Reports on ketamine’s effects on body weight in rodents are inconsistent and appear to depend on dose, duration, age, and physiological state [26]. While some studies report weight loss following ketamine exposure in healthy animals [27,28], others have observed weight gain under specific experimental conditions [29]. The increase observed in this study may partly reflect the age of the animals used. Ketamine also increased feed intake, likely through modulation of hypothalamic feeding circuits and metabolic regulation [26]. Interestingly, NAC alone similarly increased feed intake, suggesting a possible influence on appetite regulation via glutamatergic or antioxidant mechanisms [30]. Co-administration of NAC with ketamine further enhanced both body weight and feed intake, particularly at 1000 mg/kg. These findings suggest that NAC does not suppress ketamine-associated hyperphagia but may improve overall physiological resilience or metabolic efficiency rather than exacerbate dysregulation.
Ketamine markedly increased locomotor activity and rearing behaviour, consistent with its stimulant and dissociative effects mediated by NMDA receptor antagonism and downstream dopaminergic and glutamatergic dysregulation [6,9,10]. NAC exerted dose-dependent behavioural modulation. At 500 mg/kg, NAC further enhanced ketamine-induced activity, suggesting partial synergism with ketamine’s excitatory profile. In contrast, 1000 mg/kg NAC significantly reduced locomotion and rearing, likely reflecting stabilisation of glutamatergic transmission, reduced oxidative stress, and restoration of neurochemical balance [33,34]. Self-grooming behaviour, a sensitive index of emotionality and stereotypy [35], was increased following ketamine exposure, consistent with stress-related behavioural activation [9,10,36]. NAC alone reduced grooming behaviour, and this effect was maintained in the KET/NAC1000 group, indicating anxiolytic or anti-stereotypic properties at higher doses. Conversely, increased grooming in the KET/NAC500 group further supports a biphasic, dose-dependent behavioural effect of NAC.
Ketamine significantly impaired working memory performance, in line with its established disruption of prefrontal and hippocampal function [37,38,39]. NAC improved memory performance when administered alone, consistent with its pro-cognitive effects mediated by antioxidant activity and glutamate regulation [40]. Notably, high-dose NAC (1000 mg/kg) reversed ketamine-induced memory deficits, whereas the lower dose exacerbated impairment, underscoring the importance of dose optimisation in therapeutic applications. Biochemically, ketamine increased oxidative stress and neuroinflammation, evidenced by elevated MDA, IL-6, IL-1β, and TNF-α levels alongside reduced IL-10 [27]. NAC significantly reversed these alterations, reflecting its glutathione-restoring, antioxidant, and anti-inflammatory properties. While total antioxidant capacity increased with NAC 500 mg/kg, the less pronounced response at 1000 mg/kg may reflect compensatory redox dynamics at higher doses [40]. Histologically, ketamine induced marked cerebellar neurodegeneration, including Purkinje cell shrinkage, granule cell loss, and cortical disorganisation, consistent with excitotoxic and inflammatory injury [41,42]. NAC co-treatment preserved cerebellar architecture, with near-normal Purkinje and granule cell morphology and restoration of Nissl substance, supporting structural neuroprotection.
Conclusion
This study demonstrates that N-acetylcysteine exerts significant neuroprotective and behavioural modulatory effects against ketamine-induced alterations in rats. Ketamine exposure produced hyperlocomotion, stereotypy, cognitive impairment, oxidative stress, neuroinflammation, and cerebellar neurodegeneration. NAC mitigated these effects in a dose-dependent manner, with the 1000 mg/kg dose showing the most consistent restorative impact across behavioural, biochemical, and histological parameters. These findings highlight NAC’s potential as a therapeutic adjunct in conditions characterised by glutamatergic dysregulation, oxidative stress, and neuroinflammation. Further studies are required to clarify underlying molecular mechanisms and to explore translational relevance in neuropsychiatric disorders.
Funding
None.
Ethical Approval
Ethical approval for this study was granted by the Ethical Committee of the Faculty of Basic Medical Sciences (ERC/FBMS/099/2025, date 15 January 2025).
Availability of data and materials
Data generated during and analysed during the course of this study are available from the corresponding author on request.
Competing interests
All authors of this paper declare that there is no conflict of interest related to the content of this manuscript.:
References
- Zou, Z.; Wang, H.; Uquillas, F.D.; Wang, X.; Ding, J.; Chen, H. Definition of Substance and Non-substance Addiction. Adv Exp Med Biol. 2017, 1010, 21–41. [Google Scholar] [CrossRef]
- Onaolapo, O.J.; Onaolapo, A.Y. Melatonin in drug addiction and addiction management: Exploring an evolving multidimensional relationship. World J. Psychiatry 2018, 8, 64–74. [Google Scholar] [CrossRef]
- Onaolapo, A.; Onaolapo, O. Food additives, food and the concept of ‘food addiction’: Is stimulation of the brain reward circuit by food sufficient to trigger addiction? Pathophysiology 2018, 25, 263–276. [Google Scholar] [CrossRef]
- Vujović, K.S.; Jotić, A.; Medić, B.; Srebro, D.; Vujović, A.; Žujović, J.; Opanković, A.; Vučković, S. Ketamine, an Old–New Drug: Uses and Abuses. Pharmaceuticals 2023, 17, 16. [Google Scholar] [CrossRef] [PubMed]
- Marongiu, S.; van Eijk, M.; Gresnigt, F.M.; Croes, E.A.; Franssen, E.J. Rising incidence of recreational ketamine use: Clinical cases and management in emergency settings. Toxicol. Rep. 2025, 14, 101940. [Google Scholar] [CrossRef] [PubMed]
- Onaolapo, A.; Ayeni, O.; Ogundeji, M.; Ajao, A.; Onaolapo, O.; Owolabi, A. Subchronic ketamine alters behaviour, metabolic indices and brain morphology in adolescent rats: Involvement of oxidative stress, glutamate toxicity and caspase-3-mediated apoptosis. J. Chem. Neuroanat. 2019, 96, 22–33. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Zhao, J.; Li, L.; Wang, Y.; Zhang, Y.; Li, Y.; Chen, Y.; Liu, W.; Gao, L. Administration of Ketamine Causes Autophagy and Apoptosis in the Rat Fetal Hippocampus and in PC12 Cells. Front. Cell. Neurosci. 2018, 12, 21. [Google Scholar] [CrossRef]
- Chang, C.-T.; Hseu, Y.-C.; Thiyagarajan, V.; Lin, K.-Y.; Way, T.-D.; Korivi, M.; Liao, J.-W.; Yang, H.-L. Chalcone flavokawain B induces autophagic-cell death via reactive oxygen species-mediated signaling pathways in human gastric carcinoma and suppresses tumor growth in nude mice. Arch. Toxicol. 2017, 91, 3341–3364. [Google Scholar] [CrossRef]
- Onaolapo, O.J.; Paul, T.B.; Onaolapo, A.Y. Comparative effects of sertraline, haloperidol or olanzapine treatments on ketamine-induced changes in mouse behaviours. Metab. Brain Dis. 2017, 32, 1475–1489. [Google Scholar] [CrossRef]
- Onaolapo, O.J.; Ademakinwa, O.Q.; Olalekan, T.O.; Onaolapo, A.Y. Ketamine-induced behavioural and brain oxidative changes in mice: an assessment of possible beneficial effects of zinc as mono- or adjunct therapy. Psychopharmacology 2017, 234, 2707–2725. [Google Scholar] [CrossRef] [PubMed]
- Onaolapo, A.Y.; Aina, O.A.; Onaolapo, O.J. Melatonin attenuates behavioural deficits and reduces brain oxidative stress in a rodent model of schizophrenia. Biomed. Pharmacother. 2017, 92, 373–383. [Google Scholar] [CrossRef]
- Treatment for Stimulant Use Disorders: Updated 2021 [Internet]. Rockville (MD): Substance Abuse and Mental Health Services Administration (US); 1999. (Treatment Improvement Protocol (TIP) Series, No. 33.) Chapter 3—Medical Aspects of Stimulant Use Disorders. Available online: https://www.ncbi.nlm.nih.gov/books/NBK576550/.
- Chung, A.-N.; Huang, M.-C.; Liu, T.-H.; Chang, H.-M.; Chen, P.-Y.; Liu, Y.-L.; Bavato, F. Ketamine-dependent patients with persistent psychosis have higher neurofilament light chain levels than patients with schizophrenia. Asian J. Psychiatry 2024, 100, 104167. [Google Scholar] [CrossRef]
- Strous, J.F.M.; Weeland, C.J.; van der Draai, F.A.; Daams, J.G.; Denys, D.; Lok, A.; Schoevers, R.A.; Figee, M. Brain Changes Associated With Long-Term Ketamine Abuse, A Systematic Review. Front. Neuroanat. 2022, 16, 795231. [Google Scholar] [CrossRef]
- Ding, R.; Li, Y.; Du, A.; Yu, H.; He, B.; Shen, R.; Zhou, J.; Li, L.; Cui, W.; Zhang, G.; et al. Changes in hippocampal AMPA receptors and cognitive impairments in chronic ketamine addiction models: another understanding of ketamine CNS toxicity. Sci. Rep. 2016, 6, 38771. [Google Scholar] [CrossRef]
- Liu, F.; Paule, M.G.; Ali, S.; Wang, C. Ketamine-Induced Neurotoxicity and Changes in Gene Expression in the Developing Rat Brain. Curr. Neuropharmacol. 2011, 9, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Tenório, M.C.d.S.; Graciliano, N.G.; Moura, F.; de Oliveira, A.C.M.; Goulart, M.O.F. N-Acetylcysteine (NAC): Impacts on Human Health. Antioxidants 2021, 10, 967. [Google Scholar] [CrossRef] [PubMed]
- Sahasrabudhe, S.A.; Terluk, M.R.; Kartha, R.V. N-acetylcysteine Pharmacology and Applications in Rare Diseases—Repurposing an Old Antioxidant. Antioxidants 2023, 12, 1316. [Google Scholar] [CrossRef]
- Hara, Y.; Mckeehan, N.; Dacks, P.A.; Fillit, H.M. EVALUATION OF THE NEUROPROTECTIVE POTENTIAL OF N-ACETYLCYSTEINE FOR PREVENTION AND TREATMENT OF COGNITIVE AGING AND DEMENTIA. J. Prev. Alzheimer's Dis. 2017, 4, 201–206. [Google Scholar] [CrossRef]
- Holmay, M.J.B.; Terpstra, M.; Coles, L.D.; Mishra, U.; Ahlskog, M.B.; Öz, G.; Cloyd, J.C.; Tuite, P.J. N-acetylcysteine Boosts Brain and Blood Glutathione in Gaucher and Parkinson Diseases. Clin. Neuropharmacol. 2013, 36, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Onaolapo, O.J.; Onaolapo, A.Y. Nutrition, nutritional deficiencies, and schizophrenia: An association worthy of constant reassessment. World J. Clin. Cases 2021, 9, 8295–8311. [Google Scholar] [CrossRef]
- Ajao, J.; Akinsehinwa, A.; Onaolapo, O.; Onaolapo, A. Ethanol Extract of Muira Puama (Ptychopetalum Olacoides) ameliorates Aluminium Chloride-induced changes in Behaviour and Cerebral cortex Histomorphology in Wistar Rats. 2025, 1, 22–29. [Google Scholar] [CrossRef]
- Hassan, L.; Folorunso, K.; Kehinde, E.; Afolabi, O.; Olawale, A.; Lawal, M. Dietary supplementation with Powdered Ginger root ameliorates Lead Carbonate-induced Ovarian Injuries in Adult Wistar Rats. 2025, 1, 134–139. [Google Scholar] [CrossRef]
- Onaolapo, A.Y.; Olofinnade, A.T.; Onaolapo, O.J. Anxiogenic, memory-impairing, pro-oxidant and pro-inflammatory effects of sodium benzoate in the mouse brain. The J. Psychiatry Neurol. Sci. 2021, 34, 14–22. [Google Scholar] [CrossRef]
- Onaolapo, A.; Onaolapo, O.; Nwoha, P. Methyl aspartylphenylalanine, the pons and cerebellum in mice: An evaluation of motor, morphological, biochemical, immunohistochemical and apoptotic effects. J. Chem. Neuroanat. 2017, 86, 67–77. [Google Scholar] [CrossRef]
- Słupski, J.; Mechlińska, A.; Włodarczyk, A.; Kwaśny, A.; Szarmach, J.; Słupska, A.; Cubała, W.J. Appetite measures as correlates of clinical response in mood disorders treated with ketamine: systematic review. Front. Nutr. 2025, 12, 1616859. [Google Scholar] [CrossRef] [PubMed]
- L, E.; Onaolapo, O.; Onaolapo, A. Omega-3 fatty acid reverses Ketamine-induced hyperlocomotion, memory deficit and cerebral cortex neuronal injury in rats. 2025, 1, 46–57. [Google Scholar] [CrossRef]
- Parise, E.M.; Alcantara, L.F.; Warren, B.L.; Wright, K.N.; Hadad, R.; Sial, O.K.; Kroeck, K.G.; Iñiguez, S.D.; Bolaños-Guzmán, C.A. Repeated Ketamine Exposure Induces an Enduring Resilient Phenotype in Adolescent and Adult Rats. Biol. Psychiatry 2013, 74, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wang, Y.; Sun, X.; Lian, B.; Sun, H.; Wang, G.; Du, Z.; Li, Q.; Sun, L. Short- and long-term antidepressant effects of ketamine in a rat chronic unpredictable stress model. Brain Behav. 2017, 7, e00749–e00749. [Google Scholar] [CrossRef]
- Melanson, B.; Leri, F. Effect of ketamine on the physiological responses to combined hypoglycemic and psychophysical stress. IBRO Neurosci. Rep. 2021, 11, 81–87. [Google Scholar] [CrossRef]
- Hurley, M.M.; Resch, J.M.; Maunze, B.; Frenkel, M.M.; A Baker, D.; Choi, S. N-acetylcysteine decreases binge eating in a rodent model. Int. J. Obes. 2016, 40, 1183–1186. [Google Scholar] [CrossRef]
- Kokkinou, M.; Ashok, A.H.; Howes, O.D. The effects of ketamine on dopaminergic function: meta-analysis and review of the implications for neuropsychiatric disorders. Mol. Psychiatry 2017, 23, 59–69. [Google Scholar] [CrossRef]
- Cherneva, D.I.; Kehayova, G.; Dimitrova, S.; Dragomanova, S. The Central Nervous System Modulatory Activities of N-Acetylcysteine: A Synthesis of Two Decades of Evidence. Curr. Issues Mol. Biol. 2025, 47, 710. [Google Scholar] [CrossRef]
- Phensy, A.; Duzdabanian, H.E.; Brewer, S.; Panjabi, A.; Driskill, C.; Berz, A.; Peng, G.; Kroener, S. Antioxidant Treatment with N-acetyl Cysteine Prevents the Development of Cognitive and Social Behavioral Deficits that Result from Perinatal Ketamine Treatment. Front. Behav. Neurosci. 2017, 11, 106–106. [Google Scholar] [CrossRef]
- Onaolapo, OJ; Onaolapo, YAY; Akanmu, MA; Olayiwola, G. Caffeine and sleep-deprivation mediated changes in open-field behaviours, stress response and antioxidant status in mice. Sleep Science 2016, 9(3), 236–43. [Google Scholar] [CrossRef] [PubMed]
- Popik, P.; Cyrano, E.; Piotrowska, D.; Holuj, M.; Golebiowska, J.; Malikowska-Racia, N.; Potasiewicz, A.; Nikiforuk, A. Effects of ketamine on rat social behavior as analyzed by DeepLabCut and SimBA deep learning algorithms. Front. Pharmacol. 2024, 14, 1329424. [Google Scholar] [CrossRef] [PubMed]
- Roussy, M.; Luna, R.; Duong, L.; Corrigan, B.; Gulli, R.A.; Nogueira, R.; Moreno-Bote, R.; Sachs, A.J.; Palaniyappan, L.; Martinez-Trujillo, J.C. Ketamine disrupts naturalistic coding of working memory in primate lateral prefrontal cortex networks. Mol. Psychiatry 2021, 26, 6688–6703. [Google Scholar] [CrossRef]
- Tang, W.K.; Lau, C.G.; Ungvari, G.S.; Lin, S.-K.; Lane, H.-Y. Recovery of cognitive functioning following abstinence from ketamine. Addict. Behav. 2019, 99, 106081. [Google Scholar] [CrossRef]
- A Morgan, C.J.; Mofeez, A.; Brandner, B.; Bromley, L.; Curran, H.V. Acute Effects of Ketamine on Memory Systems and Psychotic Symptoms in Healthy Volunteers. Neuropsychopharmacology 2003, 29, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Cherneva, D.I.; Kehayova, G.; Dimitrova, S.; Dragomanova, S. The Central Nervous System Modulatory Activities of N-Acetylcysteine: A Synthesis of Two Decades of Evidence. Curr. Issues Mol. Biol. 2025, 47, 710. [Google Scholar] [CrossRef]
- Onaolapo, A.Y.; Onaolapo, O.J. Peripheral and Central Glutamate Dyshomeostasis in Neurodegenerative Disorders. Curr. Neuropharmacol. 2021, 19, 1069–1089. [Google Scholar] [CrossRef]
- Choudhury, D.; Autry, A.E.; Tolias, K.F.; Krishnan, V. Ketamine: Neuroprotective or Neurotoxic? Front. Neurosci. 2021, 15, 672526. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of N-Acetylcysteine (NAC) on relative change in body weight in KET treated rats. A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg). Each bar represents Mean ± S.E.M, *p < 0.05 vs. control, #p<0.05 significant difference from KET, number of rats per treatment group =10. KET: Ketamine, NAC: N-Acetylcysteine.
Figure 1.
Effect of N-Acetylcysteine (NAC) on relative change in body weight in KET treated rats. A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg). Each bar represents Mean ± S.E.M, *p < 0.05 vs. control, #p<0.05 significant difference from KET, number of rats per treatment group =10. KET: Ketamine, NAC: N-Acetylcysteine.
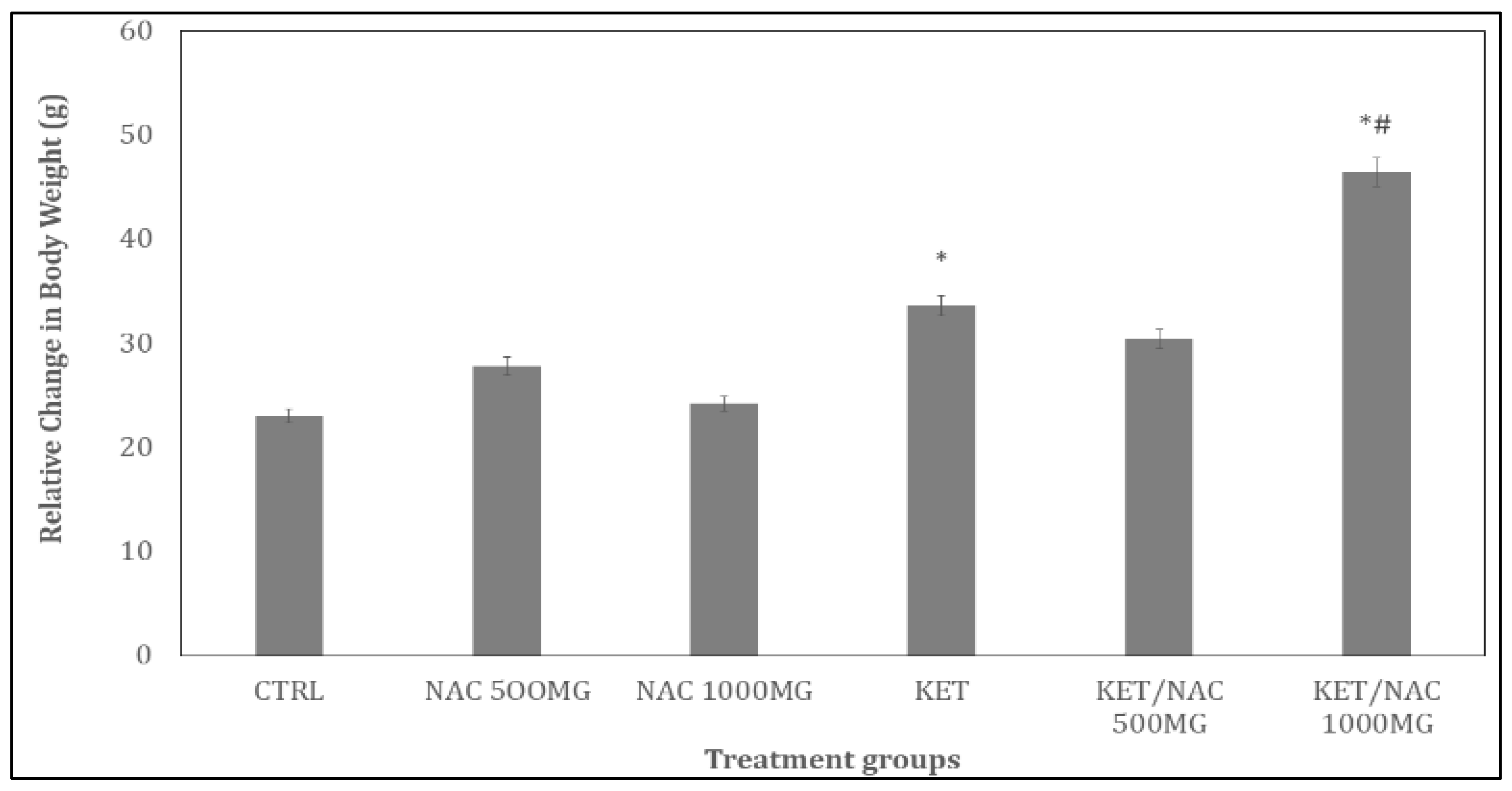
Figure 2.
Effect of N-Acetylcysteine (NAC) on relative change in feed intake in KET treated rats. A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg). Each bar represents Mean ± S.E.M, *p < 0.05 vs. control, #p<0.05 significant difference from KET, number of rats per treatment group =10. KET: Ketamine, NAC: N-Acetylcysteine.
Figure 2.
Effect of N-Acetylcysteine (NAC) on relative change in feed intake in KET treated rats. A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg). Each bar represents Mean ± S.E.M, *p < 0.05 vs. control, #p<0.05 significant difference from KET, number of rats per treatment group =10. KET: Ketamine, NAC: N-Acetylcysteine.
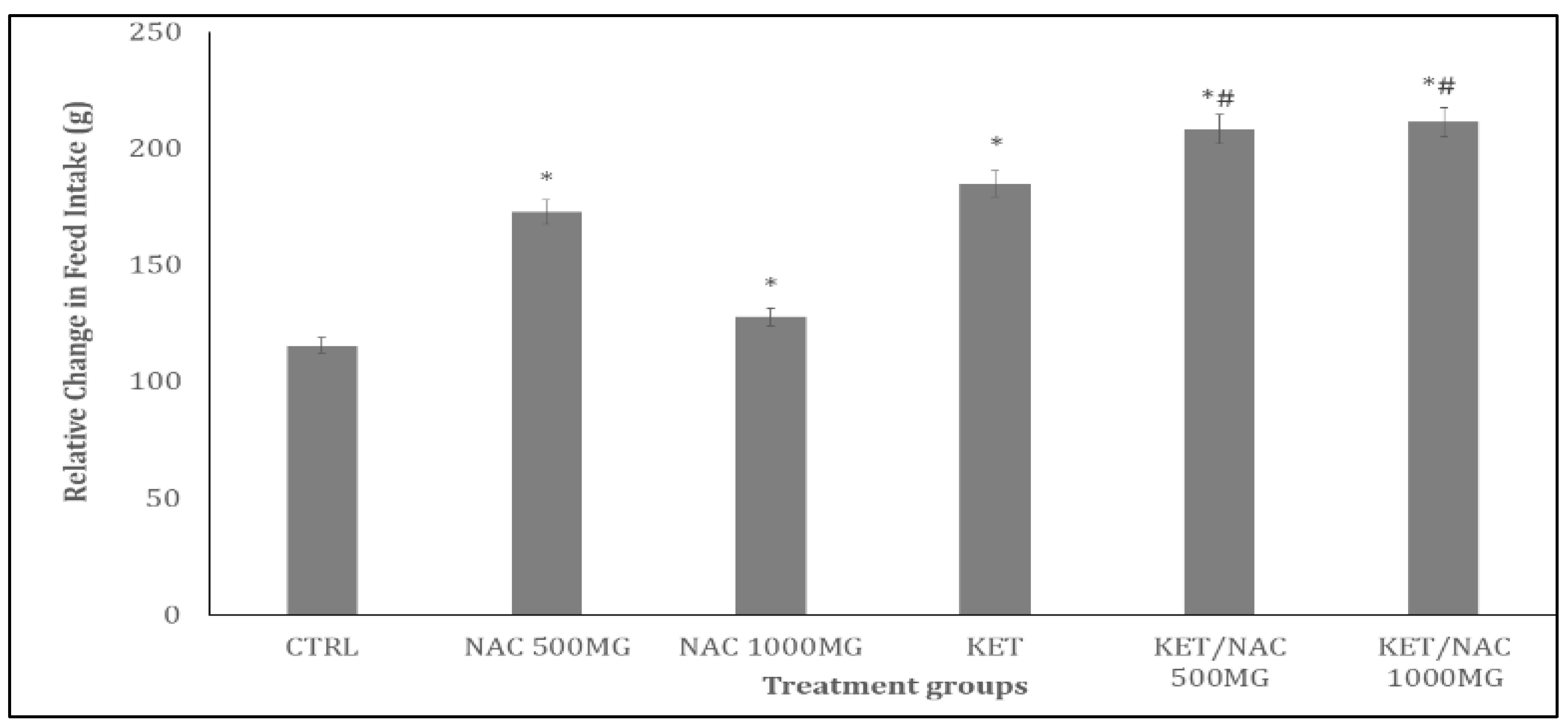
Figure 3.
Effect of N-Acetylcysteine (NAC) on horizontal locomotion in KET treated rats. A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg). Each bar represents Mean ± S.E.M, *p < 0.05 vs. control, #p<0.05 significant difference from KET, number of rats per treatment group =10. KET: Ketamine, NAC: N-Acetylcysteine.
Figure 3.
Effect of N-Acetylcysteine (NAC) on horizontal locomotion in KET treated rats. A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg). Each bar represents Mean ± S.E.M, *p < 0.05 vs. control, #p<0.05 significant difference from KET, number of rats per treatment group =10. KET: Ketamine, NAC: N-Acetylcysteine.
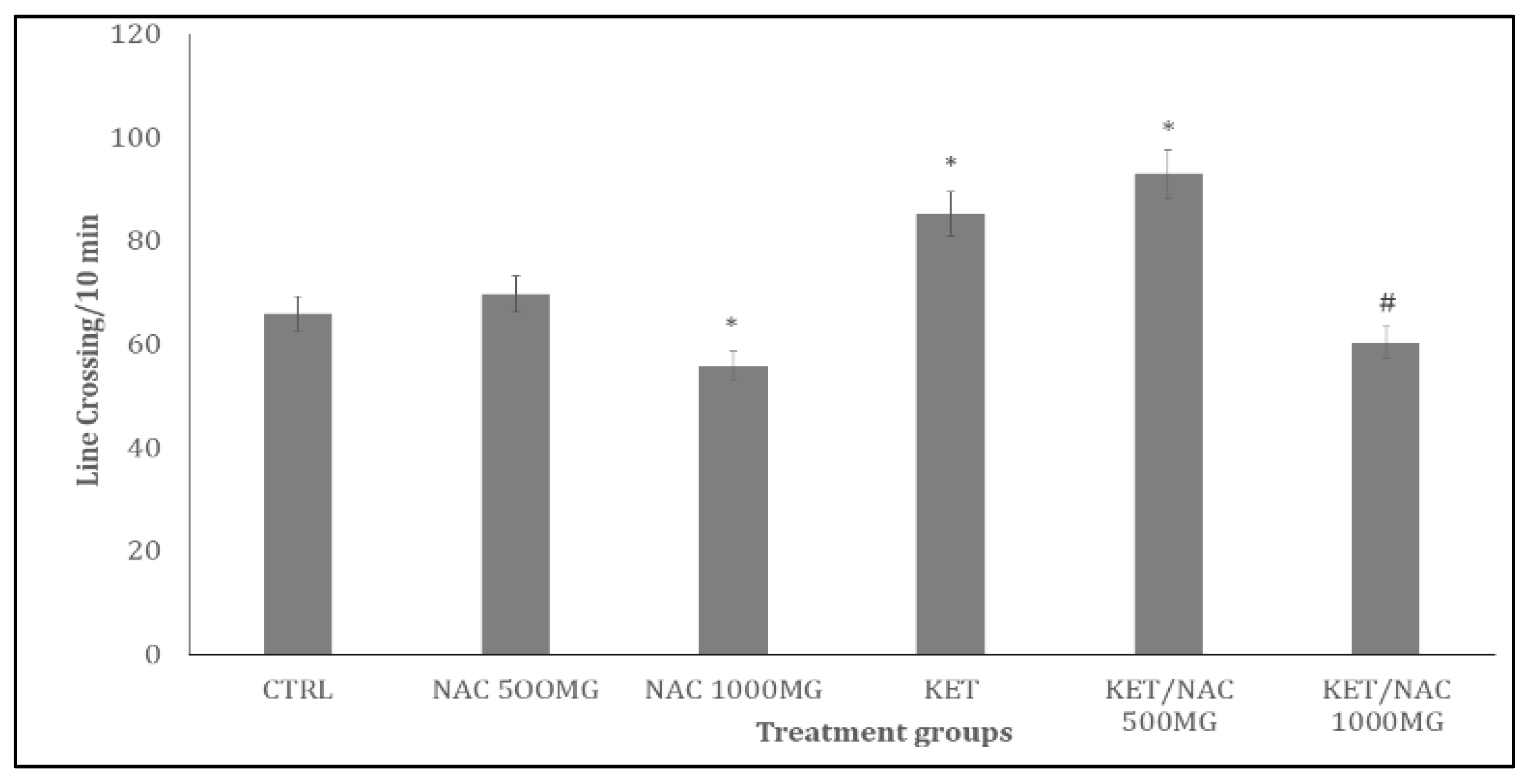
Figure 4.
Effect of N-Acetylcysteine (NAC) on rearing activity in KET treated rats. A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg). Each bar represents Mean ± S.E.M, *p < 0.05 vs. control, #p<0.05 significant difference from KET, number of rats per treatment group =10. KET: Ketamine, NAC: N-Acetylcysteine.
Figure 4.
Effect of N-Acetylcysteine (NAC) on rearing activity in KET treated rats. A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg). Each bar represents Mean ± S.E.M, *p < 0.05 vs. control, #p<0.05 significant difference from KET, number of rats per treatment group =10. KET: Ketamine, NAC: N-Acetylcysteine.
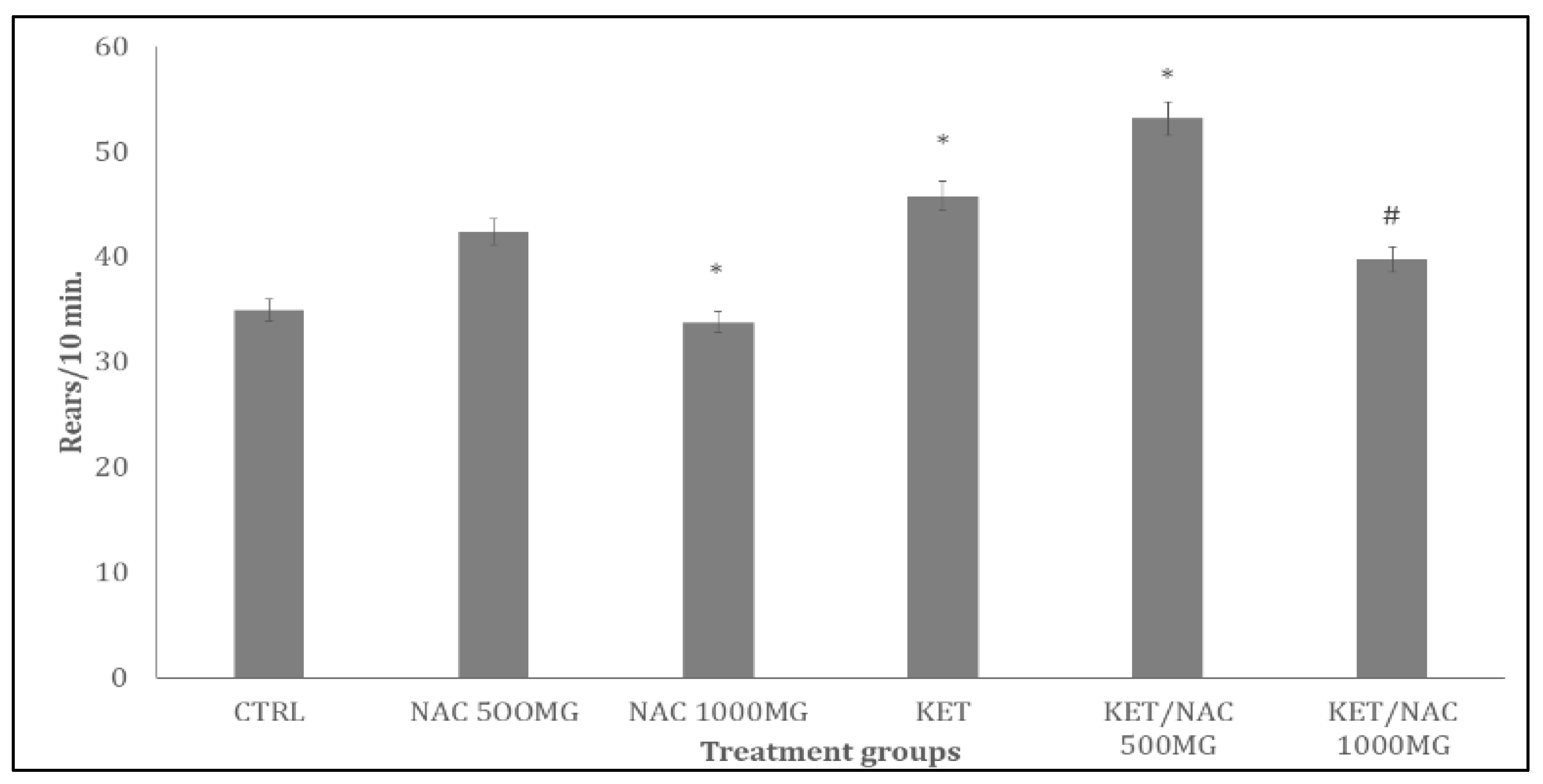
Figure 5.
Effect of N-Acetylcysteine (NAC) on self-grooming behaviour in KET treated rats. A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg). Each bar represents Mean ± S.E.M, *p < 0.05 vs. control, #p<0.05 significant difference from KET, number of rats per treatment group =10. KET: Ketamine, NAC: N-Acetylcysteine.
Figure 5.
Effect of N-Acetylcysteine (NAC) on self-grooming behaviour in KET treated rats. A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg). Each bar represents Mean ± S.E.M, *p < 0.05 vs. control, #p<0.05 significant difference from KET, number of rats per treatment group =10. KET: Ketamine, NAC: N-Acetylcysteine.
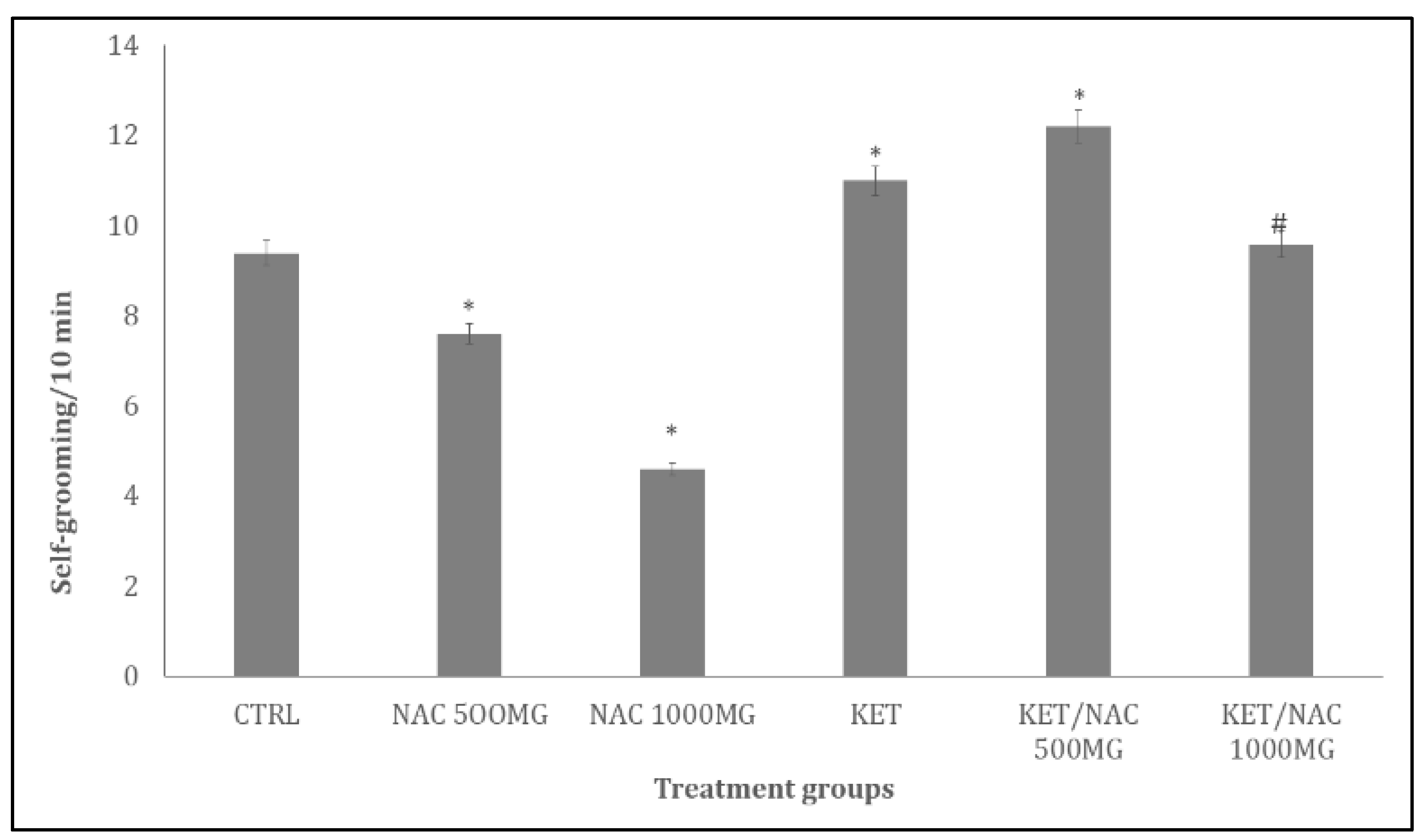
Figure 6.
Effect of N-Acetylcysteine (NAC) on Y maze spatial working memory in KET treated rats. A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg).Each bar represents Mean ± S.E.M, *p < 0.05 vs. control, #p<0.05 significant difference from KET, number of rats per treatment group =10. KET: Ketamine, NAC: N-Acetylcysteine.
Figure 6.
Effect of N-Acetylcysteine (NAC) on Y maze spatial working memory in KET treated rats. A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg).Each bar represents Mean ± S.E.M, *p < 0.05 vs. control, #p<0.05 significant difference from KET, number of rats per treatment group =10. KET: Ketamine, NAC: N-Acetylcysteine.
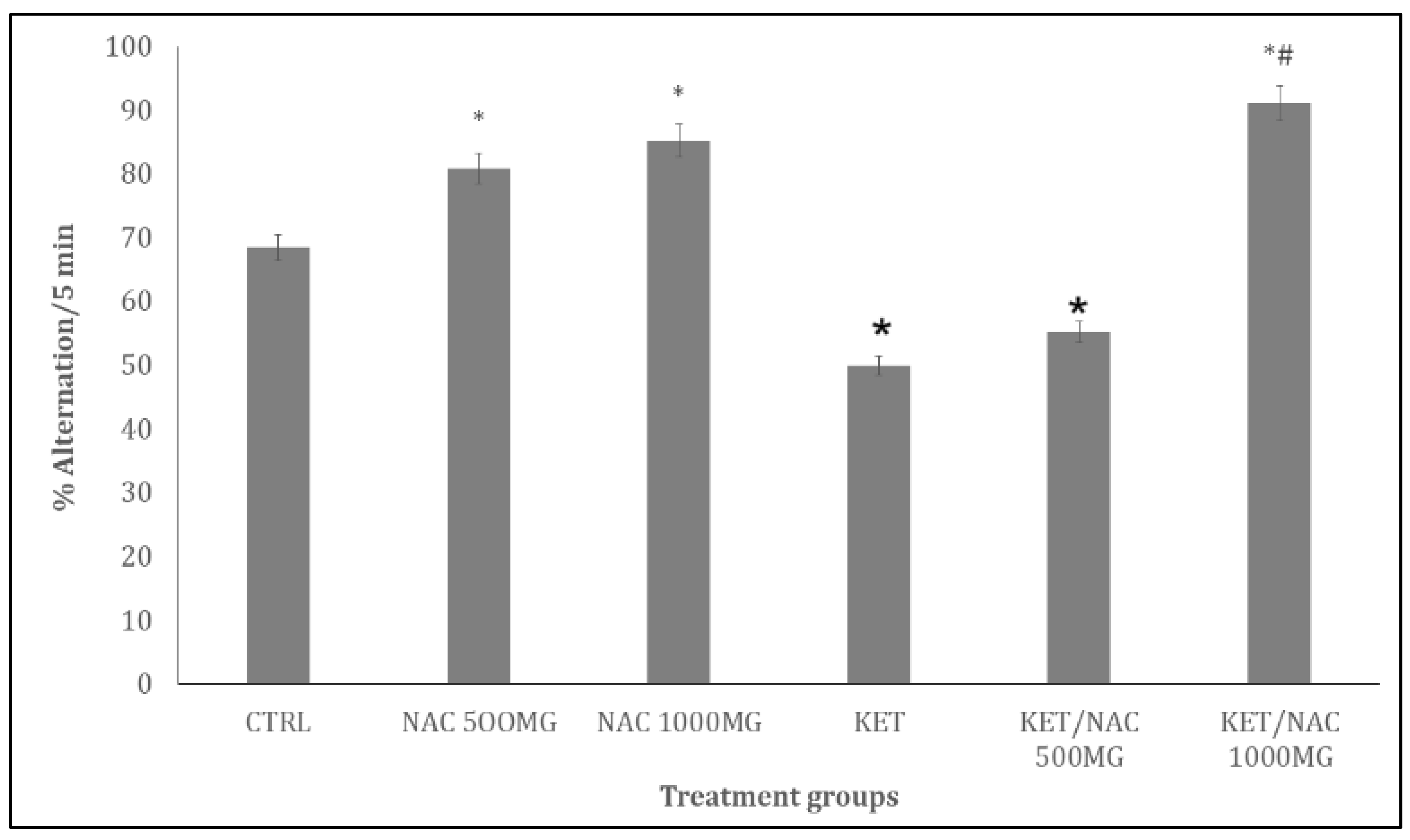
Figure 7.
Photomicrograph of haematoxylin and eosin-stained cerebellar cortex in A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg) TNF: Tumour necrosis factor -α, IL: Interleukin. Showing granule cells (GC) in the granule cell layers (GL), Purkinje cells (PC) in the Purkinje layers (PL) and molecular layers (ML) (Mag. X400).
Figure 7.
Photomicrograph of haematoxylin and eosin-stained cerebellar cortex in A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg) TNF: Tumour necrosis factor -α, IL: Interleukin. Showing granule cells (GC) in the granule cell layers (GL), Purkinje cells (PC) in the Purkinje layers (PL) and molecular layers (ML) (Mag. X400).
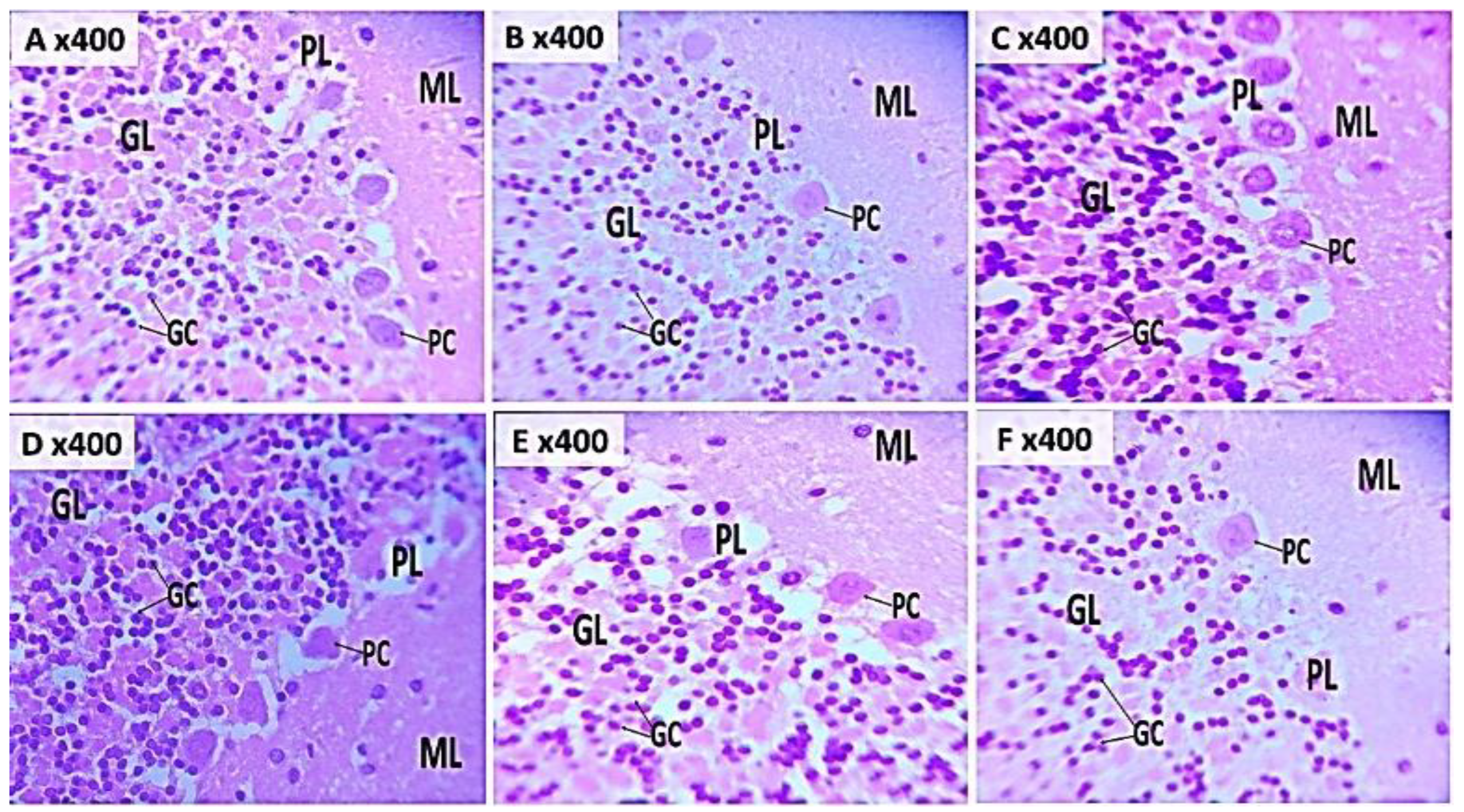
Figure 8.
Photomicrograph of cresyl fast violet-stained sections of the rat cerebellar cortex in groups A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg) TNF: Tumour necrosis factor -α, IL: Interleukin. Showing granule cells (GC) in the granular layers (GL), Purkinje cells (PC) in the Purkinje layer (PL). (Mag. X400).
Figure 8.
Photomicrograph of cresyl fast violet-stained sections of the rat cerebellar cortex in groups A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg) TNF: Tumour necrosis factor -α, IL: Interleukin. Showing granule cells (GC) in the granular layers (GL), Purkinje cells (PC) in the Purkinje layer (PL). (Mag. X400).
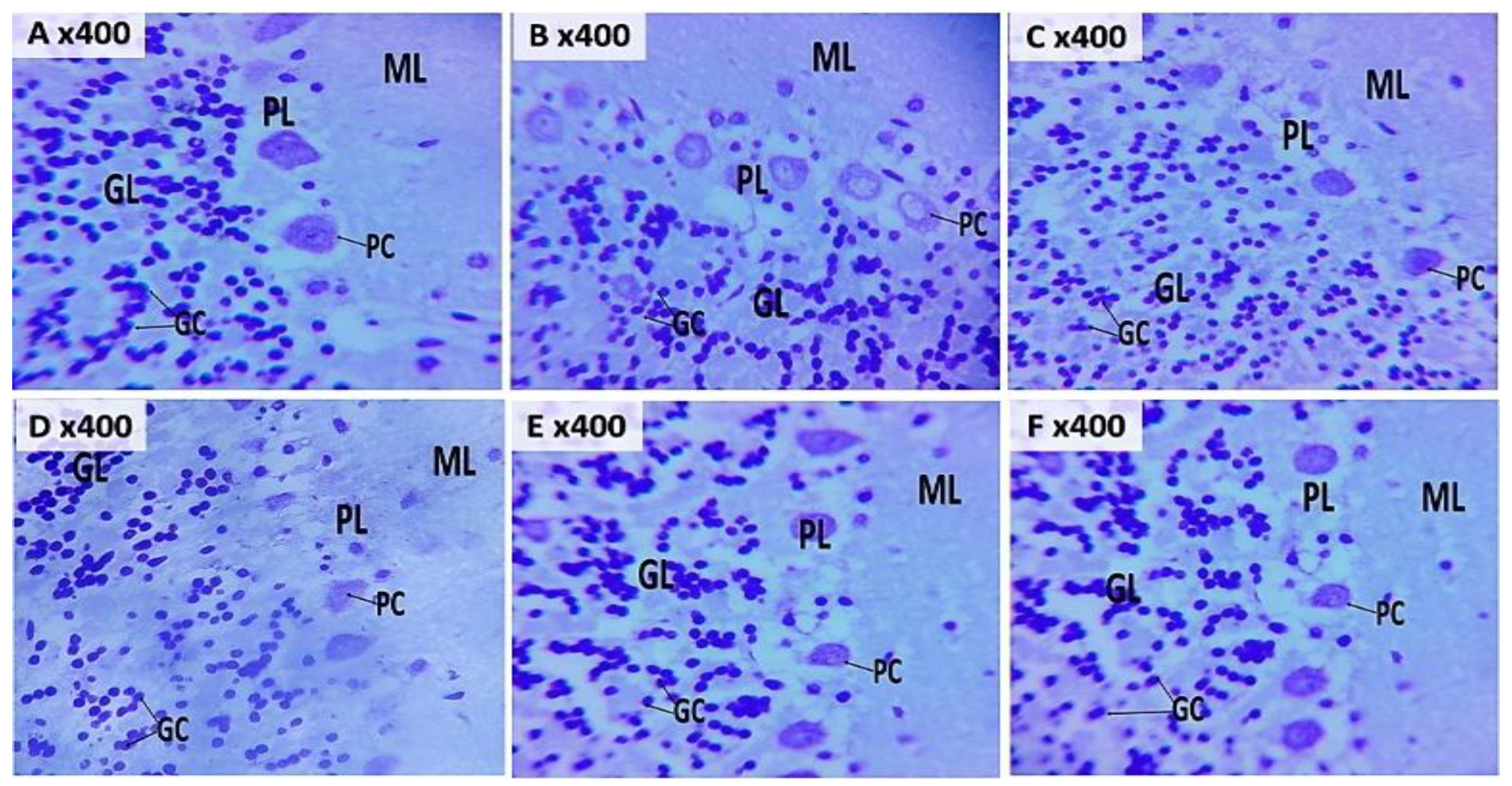

Table 1.
Effect of N-acetyl cysteine on oxidative stress markers.
| Groups | MDA (µm) | TAC (mM TE) |
| A: Control | 0.73±0.01 | 10.48±0.15 |
| B: NAC500 | 0.8±0.03 | 11.36±3.16 |
| C: NAC1000 | 0.68±0.01* | 12.67±0.03* |
| D: Ketamine | 15.81±0.91* | 5.88±3.63* |
| E: KET/NAC500 | 5.31±0.24*# | 26.53±2.71*# |
| F: KET/NAC1000 | 4.7±0.44*# | 17.8±2.03*# |
Data presented as Mean ± S.E.M, *p < 0.05 significant difference from A, #p < 0.05 significant difference from D, number of rats per treatment group =10. A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg), MDA: Malondialdehyde, TAC: Total antioxidant capacity.
Table 2.
Effect of N-acetyl cysteine on inflammatory cytokines.
| Groups | IL-6 pg/ml | TNF-α pg/ml | IL-1β ng/ml | IL-10 pg/ml |
| A | 8.33±0.22 | 11.73±0.23 | 4.57±0.5 | 6.28±0.04 |
| B | 2.03±0.09* | 9.59±0.11* | 2.82±0.08* | 10.55±0.23* |
| C | 2.33±0.01* | 11.45±0.36 | 4.9±0.19 | 6.03±0.01* |
| D | 11.45±4.17* | 35.98±0.18* | 12.2±0.89* | 0.61±0.17* |
| E | 2.11±0.03*# | 14.57±0.24*# | 4.08±0.33# | 7.66±0.2*# |
| F | 2.99±0.17* | 19.24±0.36*# | 3.31±0.16*# | 7.7±0.25*# |
Data presented as Mean ± S.E.M, *p < 0.05 significant difference from A, #p < 0.05 significant difference from D, number of rats per treatment group =10. A: Control B: NAC at 500 mg/kg, C: NAC at 1000 mg/kg, D: Ketamine at 15 mg/kg, E: Ketamine + NAC (500 mg/kg) F: Ketamine + NAC (1000 mg/kg) TNF: Tumour necrosis factor -α, IL: Interleukin.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



