Submitted:
03 January 2026
Posted:
05 January 2026
Read the latest preprint version here
Abstract
The twin-arginine translocation (Tat) system is the only general pathway for the transport of folded proteins across energized biological membranes. It is found in the bacterial or archaeal cytoplasmic membrane, the plant thylakoid membrane or the inner membrane of plant mitochondria. The biological importance of this translocation system can be exemplified by the fact that no bacterial or plant photosynthesis and photosynthetic oxygen evolvement would exist on earth without this system. Despite many biochemical and biophysical studies, the Tat mechanism has been puzzling since the system was discovered in the 1990ies. Important characteristics of the Tat system could not be explained, and also recent high-resolution structures of the Tat system’s core with bound substrate did not lead scientists to a general transport mechanism. In this integrative review, we attempt to answer the key open questions with relevance to the Tat mechanism and thereby develop the first comprehensive explanation of how folded proteins are translocated across membranes by the Tat system.
Keywords:
twin-arginine translocation
; protein transport
; membrane proteins
; protein interactions
; protein targeting
; protein biochemistry
; biological membranes
; transport energetization
Introduction
Tat-dependently translocated proteins have N-terminal signal peptides with a highly conserved twin-arginine motif that is recognized by a TatBC complex in the membrane before a TatA-dependent translocation takes place [1]. While TatC is a polytopic membrane protein with six transmembrane helices, TatA and TatB have only a short N-terminal transmembrane helix (Figure 1A). They belong to the same protein family and require their membrane anchor, a subsequent amphipathic helix and only few more residues for functionality, whereas their not conserved C-terminal domain of unknown structure is dispensable [2,3,4]. There exist two-component TatAC systems in which a “bifunctional” TatA fulfills TatA and TatB functions [5,6,7]. Tat systems are found in all domains of life, particularly in organisms that need a general transport pathway for folded proteins [2]. Despite of decades of intensive research, the translocation mechanism was still puzzling and structural insights were needed. In their recent high-resolution Cryo-EM structures of TatBC complexes with bound substrate, the research groups of Sazanov as well as of Berks and Lea revealed many aspects of subunit interactions and substrate recognition on a molecular level [8,9]. The structures show that the TatBC complex is composed of three TatC and three TatB subunits (one of which may be substituted by TatA), which together form a structure resembling an inverted cup with its opening towards the cytoplasm covered by a triangle of amphipathic TatB helices (Figure 1B). A signal peptide is bound by interactions with the N-terminus and first cytoplasmic loop of TatC and sandwiched between the amphipathic helix and the N-terminal helix of two TatB protomers, which fully agrees with earlier biochemical and molecular biological data [10,11,12,13,14]. However, contradictory translocation modes are proposed (Figure 1C): One study favors a channel model with transport through a transient and symmetric opening at the top of the complex, which could further open and extent to a larger size by the aid of TatA in case of larger Tat substrates [8]. The other study excludes such an opening and instead favors a lateral passage of Tat substrates through an associated, membrane-thinning TatA oligomer that is recruited by “semi-specific interactions with the folded passenger domain”. The translocation mechanism is hypothesized to be accomplished not by a TatB3C3 but by a structurally asymmetric TatA1TatB2TatC3 complex [9]. However, mechanistic details beyond substrate-binding are left unclear for both proposed translocation modes.
Figure 1.
The components of the Tat pathway and the contradictory initial interpretations of the TatBC complex structure. A) The topology of TatB and TatC in the TatBC complex, and the topology of a single TatA in the membrane. The positions of the transmembrane helices of TatC (TM1-6) as well as the positions of the transmembrane helices and amphipathic helices of TatB and TatA are indicated. Note that a polar residue (blue) limits the stretch of hydrophobic residues in the TM of TatA to only 12 residues, which is important for membrane weakening. TatB is structurally related to TatA, but its TM is in an unusual orientation and positioned in the center of the resting TatBC complex. The structure of TatBC is taken from Deme et al. (PDB 9DZZ, Ref [9], and the structure of TatA has been generated by AF3 [61]. B) Views on the TatB3C3 complex from indicated perspectives, in one case with fully occupied primary signal peptide-binding sites (based on PDB 9DZZ and 9E01). C) and D): The different substrate passages proposed by the Zhao and Sazanov (C, Ref [8]) and Deme et al. (D, Ref [9]), who independently solved the TatBC complex structure with bound substrate. TatBC complex, grey; TatA, pink and orange; Tat substrate, red.
Figure 1.
The components of the Tat pathway and the contradictory initial interpretations of the TatBC complex structure. A) The topology of TatB and TatC in the TatBC complex, and the topology of a single TatA in the membrane. The positions of the transmembrane helices of TatC (TM1-6) as well as the positions of the transmembrane helices and amphipathic helices of TatB and TatA are indicated. Note that a polar residue (blue) limits the stretch of hydrophobic residues in the TM of TatA to only 12 residues, which is important for membrane weakening. TatB is structurally related to TatA, but its TM is in an unusual orientation and positioned in the center of the resting TatBC complex. The structure of TatBC is taken from Deme et al. (PDB 9DZZ, Ref [9], and the structure of TatA has been generated by AF3 [61]. B) Views on the TatB3C3 complex from indicated perspectives, in one case with fully occupied primary signal peptide-binding sites (based on PDB 9DZZ and 9E01). C) and D): The different substrate passages proposed by the Zhao and Sazanov (C, Ref [8]) and Deme et al. (D, Ref [9]), who independently solved the TatBC complex structure with bound substrate. TatBC complex, grey; TatA, pink and orange; Tat substrate, red.
We provide here a conclusive multistep model of how the Tat system transports a folded protein with an appropriate signal peptide is translocated across a biological membrane. This model can explain the hitherto puzzling facts of the Tat system at a structural level, which is why we believe that the principles of the Tat mechanism are now solved. In a first phase, the signal peptide of a Tat substrate is membrane inserted by TatB3C3. In a second phase, an associated TatA ring destabilizes and disrupts the membrane at the translocation site, thereby permitting translocation. In a third phase, the system resets and releases the translocated substrate. The system requires membrane energetization for the first two phases, and it likely requires a transient local depolarization for resetting.
Tat Transport Cannot Occur Through a Central Pore Generated by a TatB3C3 Complex
Zhao and Sazanow hypothesize that the TatB3C3 complex is designed for opening in its center, as only interactions of short parallel beta strands and few other interactions stabilize the proposed closed state of the complex (Figure 1C). The complex has highly tilted transmembrane helices in a thinned membrane environment and is therefore very unusual [8]. Although the extremely unusual shape of the complex must be related to the mechanism, Tat substrates cannot pass through the center of a TatBC pore with TatA remaining on the outside, even if it is assumed that a central pore can be laterally extended for larger substrates by the aid of associated TatA. There are multiple reasons for this:
(i) If larger TatA assemblies would be required for an optional pore extension only, the signal peptide alone would not cause the same TatA association as observed with full-length Tat substrates, but it does [15]. (ii) If TatBC-containing complexes would generally open with a central pore, there would not exist TatBC complexes with interacting large periplasmic loops that prevent opening, such as found in Myxococcus xanthus [9]. (iii) If transport would proceed through a central TatBC pore, the signal peptide would need to be reinserted somehow into the membrane to be cleaved by a signal peptidase (such as LepB), and there is no such pathway known [16]. (iv) An efficient but unknown membrane-reinsertion pathway would also be required for the Tat substrates that have membrane-integral N- or C-termini, and that can become part of larger membrane-protein complexes, such as the cytochrome bc1 complex [17,18]. (v) Some Tat substrates are small globular C-terminal domains of membrane proteins that are already membrane-inserted at their N-terminus when Tat transport of the C-terminal domain takes place. Such a transport would be impossible through a central pore formed by TatB and TatC components [19].
In contrast to Zhao and Sazanow, Deme et al. favor a passage through a laterally associated TatA oligomer that weakens the membrane [9] (Figure 1D). Such a translocation step was first suggested by us in 2003 [20] and later supported by various experimental data [21,22,23] and molecular dynamics simulations [24,25]. Based on their analyses, Deme et al. reasoned that one TatA is always in one of the three TatB binding sites under physiological conditions, resulting in a TatA1TatB2TatC3 complex [9]. They believe that this structural asymmetry in the resting state is functionally needed for activity, and that only one signal peptide-binding site is functional in Tat complexes, as only one site is not influenced by the bound TatA. Furthermore, they inferred that the translocation process is initiated after recruitment of a laterally associating TatA assembly by the aid of the globular domain of bound Tat substrates. Mechanistic explanations for a directional translocation through the lateral TatA assembly are not described. However, as outlined below in detail, an intrinsic structural asymmetry cannot be needed for the mechanism, and all binding sites can be equally functional, albeit they likely function sequentially.
All Substrate-Binding Sites Can Be Equally Functional
Based on double-tag purifications and structure analyses, Deme et al. concluded that all core complexes have a TatA, and they suggest that this intrinsic asymmetry is essential [9]. Indeed, the widely accepted 1:1 TatBC ratio relied mainly on slightly variable but convincing quantifications of radioactivity in SDS-PAGE bands of purified labeled Tat complexes [26]. TatA is associated with TatBC even in the absence of bound Tat substrates [27], and TatA can still be cross-linked to TatC when the system is in the resting state [28], all of which on the first view might support the TatA1TatB2TatC3 complex proposed by Deme et al. [9]. However, various biochemical studies strongly support a TatB3TatC3 composition of the core complex: (i) The increase of TatA-TatC cross-links at TM5 in response to active transport indicates that the increased TatA interaction is transient and thus likely results from a translocation-dependent approximation of TatC and TatA, rather than from a permanent exchange [29]. (ii) Pull-down experiments with E. coli TatABC preparations showed that TatA-antibodies pulled down only a small fraction of TatBC in comparison to the pull-down with TatB antibodies, with a 1:1 TatB/TatC ratio being retained, indicating that TatA is more loosely bound at other positions, and when analyzed by BN-PAGE, co-purified TatA readily dissociated, and it has yet been impossible to identify TatA as integral constituent of the TatBC-containing core complex [26,30]. (iii) The research group of Colin Robinson showed that an E. coli TatA F39A variant strongly altered the TatA interaction with TatBC, as antibody-independently detected by radiolabeled proteins, resulting in a drastically increased TatB/C ratio and absence of interacting TatA in fractions of purified complexes. This showed that the constitutive TatC interaction differs between TatA and TatB. TatA interacts weaker and this interaction depends on positions in the amphipathic helix, and alterations in the TatA association can generate additional binding sites for TatB [31]. (iv) Biochemical analyses by the research group of Ken Cline clearly demonstrated that, in the plant Tat system, each individual TatC can bind a substrate, and that all binding sites are independent [32], and finally (v) there are active TatAC systems in Bacillus and other species that simply cannot have this structural asymmetry, as their “bifunctional” TatA takes the roles of TatA and TatB, and it is highly unlikely that these systems are mechanistically significantly different [6]. The argument of Deme et al. that overproduced TatB suppresses Tat transport [9] does not show a requirement of a constitutive TatA subunit, as the effect may simply result from the a suppression of efficient TatA cluster association or the formation of non-functional mixed TatAB assemblies. In conclusion, in the resting TatABC-containing system of the E. coli type, TatA is more loosely bound at a different position than TatB, and thus TatB3TatC3 likely forms a stable core complex in which all three signal peptide binding sites are functional, as they are in TatAC complexes of the Bacillus-type.
Phase I of Tat Transport: Signal Peptide Insertion by TatBC Complexes
Previously Unanswered Key Questions That Helped to Uncover the TatBC Complex Mechanism
- -
- How can TatC catalyze the insertion of signal peptides without transport of the mature domain, and why does TatB prevent this insertase activity, as observed by Fröbel et al. [33]?
- -
- If this insertase activity is important for the mechanism, how can the signal peptide insertion be catalyzed without liberating the twin-arginine motif from its initial binding site, as observed by Gerard et al. [34]?
- -
- How can signal peptide-binding to TatBC complexes alone already cause the association of TatA clusters, as observed by Dabney-Smith et al. [15]?
TatB N-Terminal Helices Block Access to the Signal Peptide h-Region-Binding Site in TatC
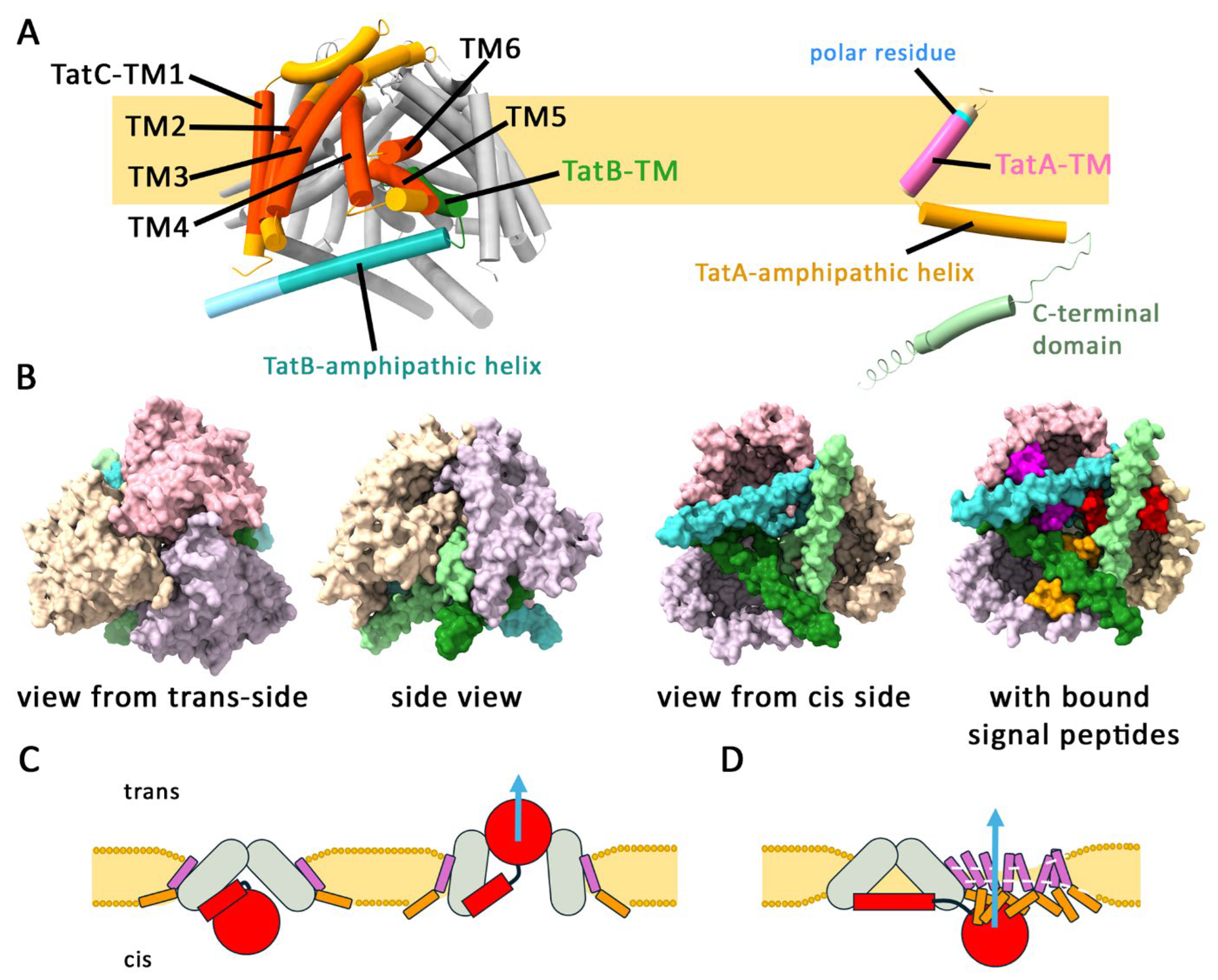
The twin-arginine motif of Tat signal peptides is bound by a cavity formed by one TatC and two TatB [9] (Figure 1B). The site is covered by a TatB amphipathic helix that must be lifted if Tat substrates enter the binding sites laterally from the membrane, as shown for selected E. coli and Streptomyces proteins [19,35]. In case of the Rieske protein of Streptomyces, this is a polytopic membrane proteins with a C-terminal domain and an internal signal peptide that mediates Tat transport, which proves that there must exist a lateral entry to the twin-arginine binding site in addition to possible binding from the cytoplasmic/stromal site [19]. TatC alone has a signal peptide insertase activity and thus must have a large signal peptide-binding site that is crossing the membrane [33]. Such a signal peptide binding site has been postulated also by the research group of Bil Clemons based on the crystal structure of a TatC monomer [36]. As the twin-arginine motif remains bound at its recognition site throughout transport [34], the hydrophobic (h)-region of the signal peptide must have access to a binding site on the TatC surface. This must reach from the twin-arginine binding site at the cytoplasmic side of TatC TM1/TM2 to the extracytoplasmic side of these helices. There is ample in vitro and in vivo cross-linking data showing that TatC interacts with the whole signal peptide, including the h-region [10,37]. However, as seen in the substrate-bound structures [8,9], the highly tilted N-terminal helix of TatB blocks the access of the signal peptide h-region to its binding site at TatC in the substrate-bound structures (Figure 2A). This explains the observation of Fröbel et al. that TatB blocks the TatC insertase activity [33]. A twin-arginine-dependent catalytic signal peptide insertion activity of TatC makes only sense if it is related to Tat transport. To function in membrane insertion of the signal peptide in the TatBC complex, the N-terminal TatB helix must therefore be moved away from this buried position in a step following signal peptide binding. The N-terminal helix of TatB interacts tightly with TM5 of the TM5/TM6 helix bundle of a neighboring TatC (Figure 2B). This whole helix bundle is highly tilted and connected to TM1-4 via a flexible linker sequence. The extremely tilted TatC TM5/TM6 protrude, together with the associated TatB N-terminal helix, into a cavity of the TatBC complex (Figure 2CD). This cavity is formed by TM1-4 of the three TatC protomers, which stably connect at the top of the complex and build a firm cage structure (Figure 2C). In summary, the TatBC complex forms a cage by helices 1-4 of the three TatC subunits. These helices are connected by a flexible linker to strongly tilted helices of TatC TM5/6 and the TatB N-terminus, which together protrude into the central cavity of the cage.
Figure 2.
The TatC TM5/TM6 domain positions TatB in a way that regulates signal peptide insertase activity of TatC. A) The TatB-TM prevents access of the signal peptide to its transmembrane-oriented binding site at TatC. B) The TatB-TM is tightly bound by TM5 of the TatC TM5/TM6 domain in the complex. C) The TatC TM5/TM6 domain with its associated TatB is positioned deep in a molecular cage generated by the TatC TM1-4 of the three subunits. D) View from the cytoplasmic side into the cup-shaped cavity formed by the TatBC complex, highlighting the three TatC TM5/TM6 domains that are tilted into the cavity. Images of the molecular structures in Figure 2, Figure 3 and Figure 4 were generated by use of ChimeraX 1.10.1 [83].
Figure 2.
The TatC TM5/TM6 domain positions TatB in a way that regulates signal peptide insertase activity of TatC. A) The TatB-TM prevents access of the signal peptide to its transmembrane-oriented binding site at TatC. B) The TatB-TM is tightly bound by TM5 of the TatC TM5/TM6 domain in the complex. C) The TatC TM5/TM6 domain with its associated TatB is positioned deep in a molecular cage generated by the TatC TM1-4 of the three subunits. D) View from the cytoplasmic side into the cup-shaped cavity formed by the TatBC complex, highlighting the three TatC TM5/TM6 domains that are tilted into the cavity. Images of the molecular structures in Figure 2, Figure 3 and Figure 4 were generated by use of ChimeraX 1.10.1 [83].
Biochemical and Structural Evidence for a Signal Peptide-Induced Domain Movement in TatBC
As the N-terminal helix of TatB has to move away to permit access of the signal peptide h-region to its TatC binding site, the TatC TM5/6 helix bundle must move together with the bound TatB. The necessary tilting at the hinge between TM4 and TM5 of TatC exposes the TatB N-terminal helix at the surface and forms a lateral opening and exit site for the inserted signal peptide (Figure 3 and Suppl. Video S1). When the TatBC complex laterally opens by the domain movement, vacant hydrophobic surfaces inside need to be occupied by membrane lipids. Such lipids have been identified in the structure [9], and they must be there in order to prevent a destabilization of the translocon. Accordingly, phosphatidylethanolamine has been identified as being essential for Tat transport [38]. The movement-induced exposure of the transmembrane-helix of TatB to the surface would generate a potential docking site for TatA in close proximity to the TatC TM5/TM6 domain. This is supported by the observation that TM5 contacts TatA under conditions of translocon saturation [29]. Other evidence comes from in vitro studies of Fröbel et al., who observed TatBC, RR- and pmf-dependent cross-links of Tat signal peptides to a single TatA [39]. These crosslinks can be explained by a placement of the inserted signal peptide in the lateral gate at the moved TatC TM5/TM6 domain in close proximity to the interacting TatA that is responsible for the TatA ring attachment. The absence of additional TatB or TatA crosslinks support the view that the E. coli TatB exchanges with TatA during transport, and thus such a transient exchange can be required in three-components Tat systems for efficient triggering of the TatA ring extraction. A permanent exchange is unlikely: Roland Freudl and coworkers found that single mutations in the N-terminal 6 residues could transform an E. coli TatA into a TatB [40]. The corresponding region in TatB is exactly the region that contacts the “periplasmic” TatC loop between TM5 and TM6 that is positioned deep in the complex in the resting state. This makes sense, as the N-terminal 6 residues of TatB are designed to stabilize the interaction that is required for the movement of TatB with the TM5/TM6 domain – and thus TatB must be bound to the TatC TM5/TM6 domain in the resting state, not TatA.
The conformational switch of the tilted helices would thus explain i) the signal peptide insertion, ii) the TatA mutations that render TatA bifunctional, iii) the formation of a TatA recruitment-site at the complex surface, iv) the transport-enhanced cross-links of TatC TM5 with TatA, and v) a lateral opening at the translocation site that would be required for the exit of the inserted signal peptide. But there is also experimental evidence for this key conformational switch coming from the structural analyses: Deme et al. did not mention but saw that the N-terminal helices of the TatB subunits became more flexibly positioned inside the complex in response to signal peptide-binding (Figure S8g in Ref [9]). This indicates a higher flexibility of the TatB and the tightly interacting TatC TM5/TM6 domain, which are therefore able to move in response to signal peptide-binding. The movement explains that signal peptide-binding triggers the recruitment of TatA oligomers already without globular mature domains of folded substrates, answering the key question of how TatA is recruited to the translocation site [15].
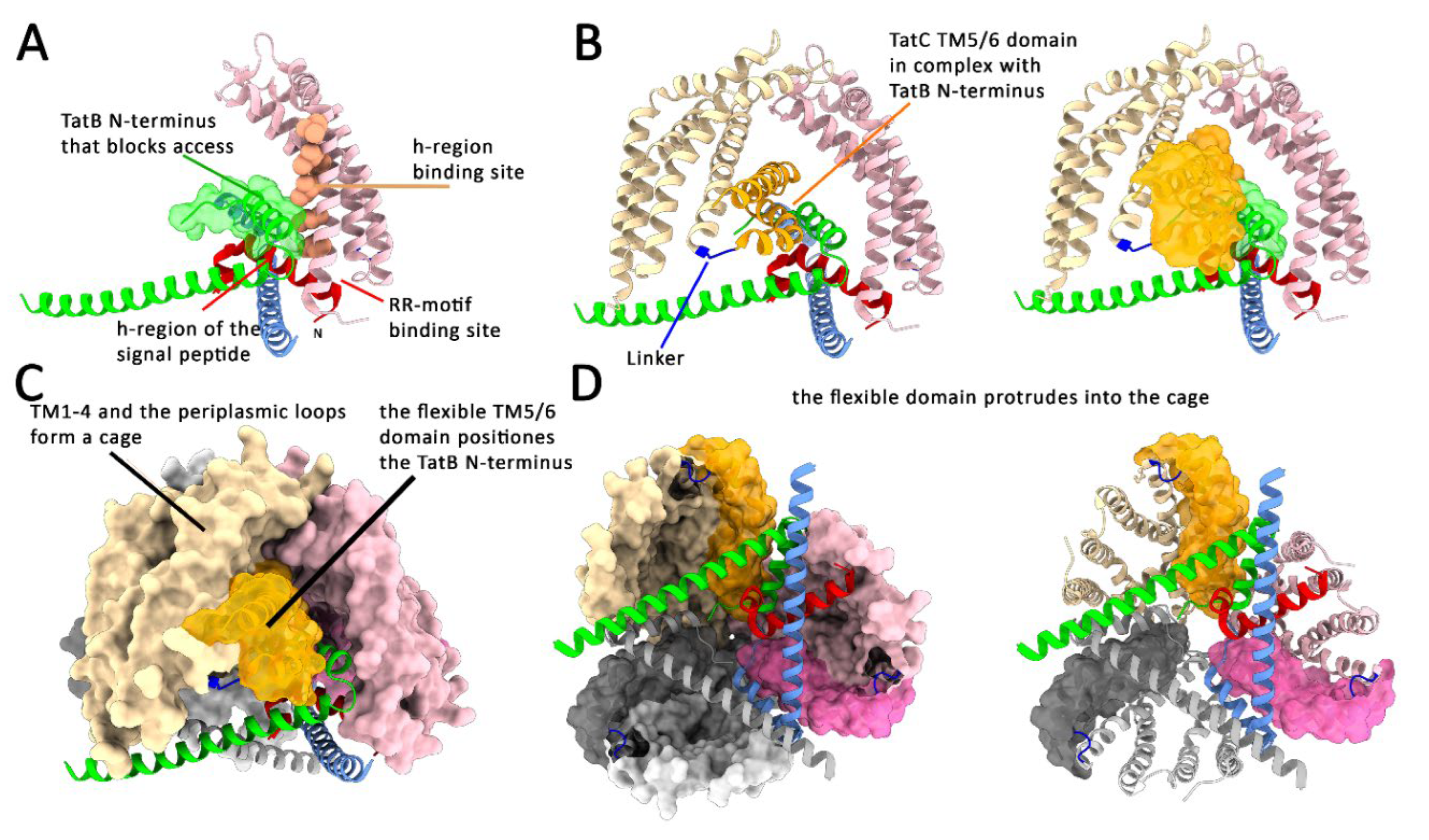
Figure 3.
Signal peptide insertion and its energetization. A) A likely movement of the TatC TM5/TM6 domain can position TatB on the surface and permit access of the signal peptide h-region to TatC, resulting in signal peptide insertion (see supplemental Video S1 for clarity). B) The movement can be energized by the electric field at the energized membrane. Especially a conserved aspartate of TatC (in E. coli D211) and a patch of arginines trigger the movement. The reason for its essential role was unclear so far. The conserved glutamic acid chain that is limiting the hydrophobic region of the transmembrane domain of TatB (in E. coli E8) also contributes to the movement.
Figure 3.
Signal peptide insertion and its energetization. A) A likely movement of the TatC TM5/TM6 domain can position TatB on the surface and permit access of the signal peptide h-region to TatC, resulting in signal peptide insertion (see supplemental Video S1 for clarity). B) The movement can be energized by the electric field at the energized membrane. Especially a conserved aspartate of TatC (in E. coli D211) and a patch of arginines trigger the movement. The reason for its essential role was unclear so far. The conserved glutamic acid chain that is limiting the hydrophobic region of the transmembrane domain of TatB (in E. coli E8) also contributes to the movement.
The Domain Movement Requires Membrane Energetization
But what would energize the domain movement and why is there no CryoEM structure of a signal peptide-bound TatBC complex with a switched conformation? The answer to these questions can be found in a biochemical in vitro study by the Matthias Müller research group, conducted in 2003, and confirmed by us in vivo in 2014 [10,41]: While the twin-arginine motif interacts with TatBC complexes already in the absence of the proton motive force (pmf), the TatC interactions to the h-region are absolutely dependent on a pmf and absent in the presence of the uncoupler CCCP. In agreement with the model and the structure is also the fact that TatB interactions of the h-region do not depend on a pmf: As TatB hinders access of the h-region to TatC in the resting state prior to the domain movement, the h-region must contact TatB in the presence of CCCP that blocks the domain movement. The membrane energetization has long been shown to be essential for Tat-dependent protein transport [42,43,44,45]. The research group of Siegfried Musser found that there is a ΔΨ-dependent step during Tat transport, and he suggested that “… instead of being consumed to energetically drive transport, it is possible that the Δψ is instead coupled to protein conformational changes (“gating reactions”) that are required for transport.” and “However, a reasonable scenario is that movement of certain charged regions within Tat proteins could be induced by a Δψ.” [46]. We think he was right: The highly tilted TatC TM5/TM6 helix has a conserved negatively charged spot on the “periplasmic” end of the TM5/TM6 domain, which is located deep in the membrane, which permits the use of the electric field for the movement of the domain into a conformation that places the negative charge closer to the periplasmic surface of the membrane (Figure 3). In E. coli, this residue (D211) has been demonstrated to be absolutely essential for Tat transport [47]. The movement is likely enhanced by TatB E8, which is even deeper positioned in the membrane and similarly moves toward the positively charged side of the energized membrane. This is likely the reason for the predominant occurrence of glutamic acid in TatB and bifunctional TatA at this position. In addition, often a positively charged spot of exposed arginines on the cytoplasmic face of the TM5/TM6 domain is embedded into the membrane and likely supports the domain movement (Figure 3). As this energetization is abolished by any membrane protein solubilization, sample preparation for CryoEM automatically locks the complex in the conformation prior to the switch, irrespective of substrate-binding, which is why CryoEM could not show the domain movement.
Upon Signal Peptide Insertion, the Globular Domain Relocates to the Translocation Site
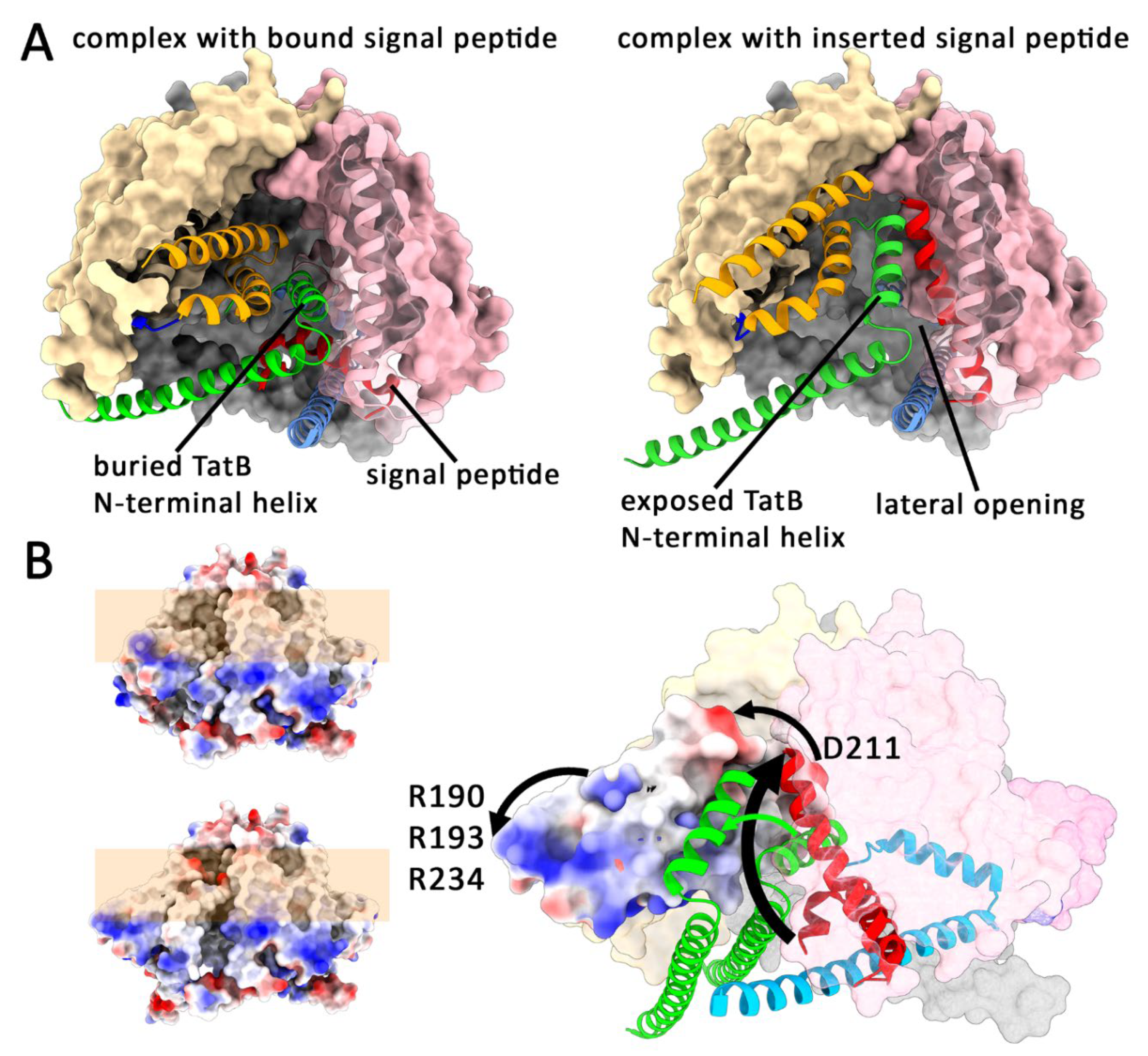
To pass through the membrane, the globular domain of the Tat substrate must be positioned at the lateral opening generated by the movement of the TM5/6 helix bundle. In the CryoEM structures that show the position prior to the movement, the globular domain is positioned at a more central position of the cis-side (in prokaryotes the cytoplasmic, in plant plastids and mitochondria the stromal side), making contacts to more than one TatB amphipathic helix [8,9]. This agrees with the crosslinking data of the research group of Matthias Müller [48]. At this time point, the signal peptide reaches the twin-arginine motif-binding site via a cavity underneath a TatB amphipathic helix. As three amphipathic helices form a closed triangle, this triangle requires opening to permit the passage of the mature domain to a lateral translocation site close to the signal peptide insertase site at TatC TM1/TM2. As already mentioned in the context of substrate-binding above, the amphipathic helix cannot be firmly bound, and thus the TatC TM5/TM6 domain movement with the bound TatB adjacent to the signal peptide insertion site will open the triangle formed by the TatB amphipathic helices. In other words, the TatC TM5/6 domain movement not only activates signal peptide insertase and generates a lateral opening that permits later TatA assembly and signal peptide release, it also enables the relocation of the globular domain of the substrate to the lateral opening of the TatBC complex. At the end of this phase I, the signal peptide is inserted, the N-terminal region of the mature protein is unfolded and pulled through the membrane by this insertase activity, and the globular domain is positioned on the cis site of the membrane at the lateral opening of the TatBC complex. This intermediate was first identified by the research group of Ralf Bernd Klösgen, who termed it “translocation intermediate 1” or “Ti-1” [49], and it is the pmf-dependent state where the substrate is deeply bound in the TatBC complex, as identified by Gerard and Cline [50].
Phase II of Tat Transport: Docking of TatA, Membrane-Disruption and Directional Translocation
Previously Unanswered Key Questions That Helped to Understand TatA-Dependent Translocation:
- -
- -
- What are the structural characteristics of the ordered cluster of at least 16 TatA protomers that is generated at TatBC upon RR-dependent binding of the signal peptide to TatBC, as observed by Carole Dabney Smith et al. [15]?
- -
- -
- Why does substrate-interaction with TatA quantitatively protect a specific region of the TatA in a TatBC-dependent way, as shown by us [21]?
- -
- Why do large TatA assemblies diffuse either slowly, as expected for membrane-integral proteins, or rapidly, as expected for membrane-associated proteins without transmembrane helix, as shown by Yves Bollen et al. [54]?
- -
- What is the energy-demanding step in phase II of Tat transport [46]?
A Simple Pulling of the Folded Protein Through a Disordered TatA Assembly Cannot Explain Tat Transport
When the TatC TM5/6 domain movement exposes the bound TatB at the surface, a docking of TatA to the translocation site starts, likely by an interaction with TatB and/or with its binding site at TM5. At least 16 interacting TatA protomers are recruited, and their interaction changes in a way that their C-terminal regions can be readily cross-linked [15]. It has been reported that TatA assemblies can weaken the membrane by local thinning and lipid disorder, which is currently believed to permit the passage of folded proteins next to TatBC complexes [23,25,55], proposed to be energized by a TatC-dependent pulling [20].
However, although the above described signal peptide insertion by TatBC exerts a force that pulls the N-terminal region of the folded domain into the membrane, this one pulling step cannot be responsible for the translocation of the folded protein: The plant Tat system has been shown to translocate small folded proteins that are connected to the signal peptide via an unfolded linker of at least 75 residues (→ 15 x GGGGS repeats) [52]. Similarly, we discovered that the E. coli Tat system is able to translocate unfolded linkers of 110 residues together with a C-terminally fused tightly folded ca. 10 kDa globular domain [51]. In addition, it has been shown that two globular proteins fused by a linker can be translocated by the Tat system [56]. These studies indicated that Tat substrates do not pass through the membrane via a single pulling stroke, and as the twin-arginine motif can remain bound to the same binding site until translocation is completed [34], there must be a second phase in Tat transport that completes the process after signal peptide insertion. This is further supported by bioenergetic analyses that point to a second, longer period of membrane energetization that is required to complete transport after phase I, which we know now is the above described Δψ-dependent signal insertase activity) [46]. But what is the nature of this “phase II” of transport, in which TatA somehow completes the translocation process?
TatA Must form a More Defined Structure to Permit Tat Transport
The association of TatA can either be unordered or ordered. We originally proposed unordered TatA arrangements when we invented the idea of membrane-weakening in 2003 [20]. Interacting flexible TatA transmembrane-helices and amphipathic helices could weaken the membrane, as experimentally shown [21,55], and TatA transmembrane-helices and amphipathic helices would need to be somehow pushed away by the membrane-crossing globular Tat substrate. An imposed directionality could be achieved by the fact that the signal peptide C-region is already translocated across the bilayer at this time point, and there is some energy provided by the refolding of the N-terminus of the globular Tat substrate that had been unfolded in the course of signal peptide insertion. However, this folding energy cannot be strictly required and it also cannot be a simple pulling mechanism, since fully folded proteins can be translocated that are bound to signal peptides via long unfolded linkers [51,52]. Without more energy, it is difficult to explain how interacting TatA helices and lipids can be reliably pushed away during transport. TatA obviously requires TatBC for translocation, and therefore at this phase II of transport there must be some contribution of TatBC other than pulling through a TatA assembly. In addition, if unordered TatA assemblies would catalyze a membrane passage, topology changes would not be controlled and TatA/substrate interactions during transport could generate irreversible holes that would harm the cells just like antimicrobial peptides. Further, if any kind of pulling or pressing would be responsible for the directionality of transport through TatA assemblies, the TatA assembly would need to be somehow in a fixed position relative to TatBC during translocation, as a highly flexible TatA assembly could move with the membrane without permitting a passage through the membrane. Moreover, it is hard to imagine that anything specific could be triggered by a pmf in an unordered TatA assembly, but phase II of Tat transport requires at least a low pmf [46]. Based on all these arguments, and given a much better alternative (see below), we exclude that an unordered assembly of TatA generates the pathway through the membrane.
TatA Forms Rings that Could Allow a Controlled TatBC-Dependent Membrane Perforation
An alternative to an unordered TatA cluster is an ordered, likely ring-shaped TatA complex that encloses a defined weakened membrane area. The membrane-weakening would require that the short transmembrane-helices are flexibly connected to the rigid structure, pointing to the center of the enclosed membrane area. It has been shown by crosslinking analyses that substrate-binding to TatBC induces an ordered interaction of TatA multiple protomers, which without substrate-binding interact differently [15,57]. A well-defined ring with identical TatA protomer interactions would best explain such a regular crosslinking and combines this with advantageous properties. In the already thinned membrane environment of the TatBC complex [9], docking of such a ring to TatBC could extend the thinned membrane area that surrounds TatBC into the inner space of the ring. Beyond thinning, the membrane would be further destabilized by the hydrophilic N-terminus of the substrate’s mature domain that crosses the membrane as a result form signal peptide insertion.
TatA can in principle form rings: When TatA was solubilized, purified and analyzed by electron microscopy, it showed rings or variable diameter [58]. Interestingly, the central pore of these rings was covered only on one side by electron density. The authors believed at that time in a gated pore mechanism and interpreted this electron density as lid on the cytoplasmic side. The lid-idea was discarded thereafter, and the rings were never understood. Note that smaller rings may have been generated during solubilization due to detergent sensitivity of larger rings. In another study, we found that with mild overproduction of TatABC, TatA formed hollow tubes with an inner diameter of ca. 6.7 nm and an outer diameter of about 11.5 nm, which were visible by electron microscopy in thin sections of cells after ultrastructure-preserving cryofixation and cryosubstitution [59]. These tubes had the dimensions of the rings reported before, and they appeared like stacks of multiple rings. Importantly, their formation depended on TatBC and therefore they originated at membranes. In the absence of TatB, the tubes were not hollow and contained a central, electron-dense region, indicating that TatB affected the TatA rings in a way that resulted in a hollow central space [59]. This hollow space was large enough to accommodate the largest known Tat substrates, such as the hetero dimeric FdnGH of formate dehydrogenase (142 kDa, ca. 6.4 nm, PDB1KQF, Ref [60]). In summary, TatA has the ability to assemble to ring structures of sufficient size to serve for translocation of folded proteins, even with more than one subunit.
AlphaFold 3 Predicts TatA Rings that Explain so Far Not Understood Findings
To function, the signal peptide release site of the TatBC complex must be open towards the TatA ring, which is why TatBC must be integral part of the rings that are present at active translocons. According to cross-linking data, at least 16 subunits reorganize at the substrate-bound translocon [15]. A closed ring of 22-26 TatA would be in the range of the dimensions of tube cross-sections or solubilized TatA rings [58,59]. AlphaFold 3 (AF3, Ref. [61]) models such TatA rings with amphipathic helices that interact in a stable closed circle that is the basis for the ring (Figure 4). The short transmembrane helices are seen in the center of the ring, which could easily insert into the membrane, if only amphipathic helices would be flexible in their orientation, which likely is the case (Figure 4B). After membrane insertion, the complex would have exactly the shape of the previously reported TatA rings, but what has been originally interpreted as a lid on the cytoplasmic side was likely the bundle of transmembrane-helices, and what has been assigned as transmembrane region were the polar and amphipathic TatA regions on the membrane surface [58].
Figure 4.
AlphaFold 3 predicts a ring structure for TatA that makes physiological sense and that explains the importance of conserved TatA residues. A) AF3 predicts ring structures for TatA in which the short transmembrane domains are positioned in the inner side of the ring. Shown is a 24mer. Note the hydrophobic cavity that is formed by the amphipathic helices and the transmembrane helices of the ring. B) Ribbon structure of the ring, highlighting the essential regions for TatA functionality, which are the transmembrane helix and the amphipathic helix. Note that the amphipathic helices are tightly interacting and form a stable ring, whereas the transmembrane helices are bound to the ring via the flexible hinge1 region. Note also the presence of a second hinge at the other end of the amphipathic helix. The two hinges can allow orientational changes to place the transmembrane helices in the membrane. C) The amphipathic helices interact in an ordered way that explains the important roles of the essential FK motif. D) Scheme of the two states of TatA rings that likely play a role for Tat transport, based on the AF3 structure, which explains TatA functionality.
Figure 4.
AlphaFold 3 predicts a ring structure for TatA that makes physiological sense and that explains the importance of conserved TatA residues. A) AF3 predicts ring structures for TatA in which the short transmembrane domains are positioned in the inner side of the ring. Shown is a 24mer. Note the hydrophobic cavity that is formed by the amphipathic helices and the transmembrane helices of the ring. B) Ribbon structure of the ring, highlighting the essential regions for TatA functionality, which are the transmembrane helix and the amphipathic helix. Note that the amphipathic helices are tightly interacting and form a stable ring, whereas the transmembrane helices are bound to the ring via the flexible hinge1 region. Note also the presence of a second hinge at the other end of the amphipathic helix. The two hinges can allow orientational changes to place the transmembrane helices in the membrane. C) The amphipathic helices interact in an ordered way that explains the important roles of the essential FK motif. D) Scheme of the two states of TatA rings that likely play a role for Tat transport, based on the AF3 structure, which explains TatA functionality.
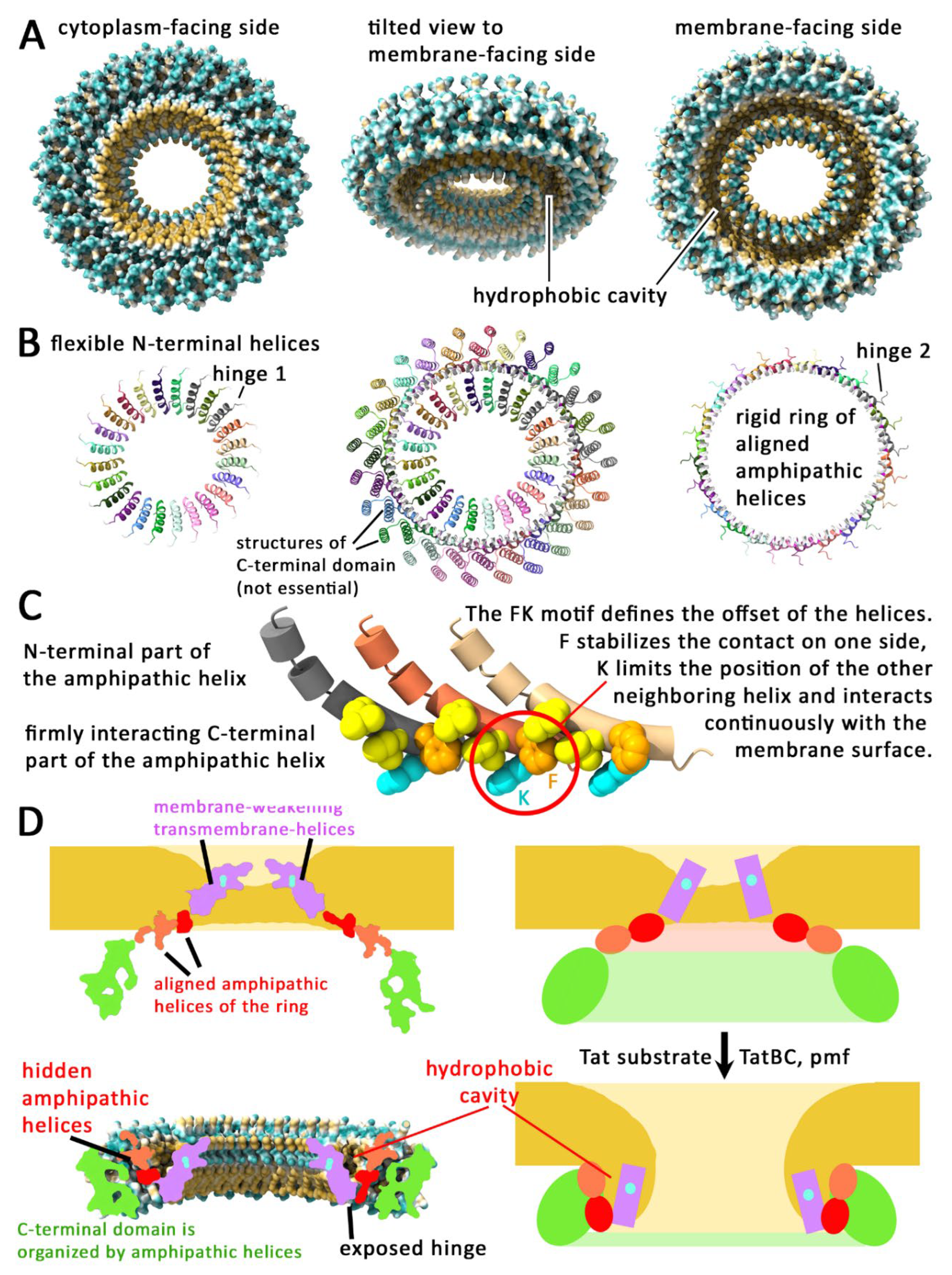
Based on multiple evidence, we believe that this AF3-predicted ring must strongly resemble the translocon-associated structure in the substrate-associated conformation: (i) The AF3-predicted ring attributes an essential role for ring formation to the amphipathic helix, and the amphipathic helix is known to be essential for Tat transport [3]. (ii) The AF3-predicted predicted ring explains the high conservation of the FK-motif (in plants and cyanobacteria FQ motif) in the amphipathic helix (in E. coli F39/K40, Ref 62), as this motif determines the staggered arrangement of the interacting helices that results in the defined ring: K interacts with the membrane surface and positions the end of the interacting amphipathic helix at its side (Q in plants and cyanobacteria that have glycolipids), and the preceding F has the tightest interaction with the neighboring helix (Figure 4C). This is experimentally confirmed, as TatA-F39A mutation results in irregular inactive TatA assemblies, indicating that firmly ordered amphipathic helices are essential functionality [31]. (iii) The AF3-predicted ring also explains the dominant negative phenotype of the F39A mutation, as all subunits of the ring require F39 to correctly interact with neighboring subunits [63]. The alignment and orientation of the amphipathic helices in the ring would also fully agree with biophysical analyses that indicated such an ordered and tilt orientation [64]. As seen below, the amphipathic helix interactions are of key importance for our proposed mechanism.
The Predicted TatA Rings Suggest a Membrane Disruption Mechanism That Is Experimentally Supported
Although details of the regular interaction of the amphipathic helices in the predicted TatA ring are in astonishing agreement with experimental data, which allows the assignment of functions of important conserved residues, the orientation of the amphipathic helices and the position of the transmembrane helices is surprising. In the predicted structure, only the C-terminal end of the amphipathic helices could make contacts to the membrane surface, with the conserved lysine of the FK motif being the key residue, while the hydrophobic side of the amphipathic helices hardly contact the membrane surface and instead mediate helix-helix interactions and form a hydrophobic cavity together with the N-terminal transmembrane-helices (Figure 4). This is possible, because the tilt orientation of the helices positions the N-terminal transmembrane helices inside the ring. To comfortably insert the transmembrane-helices into the membrane, the amphipathic helix would need a reorientation and/or a kink. Such a kink could be formed at the conserved internal flexible positions of the amphipathic helix (in E. coli G34), and also a reorientation could be easily achieved, as the amphipathic helices have flexible linkers at both ends (Figure 4). Therefore, the AF3-predicted ring could in principle insert into membranes. In the membrane-inserted state, the presence of multiple aligned amphipathic helices and likely disordered transmembrane helices would weaken the drag of the membrane, like proposed earlier [20].
As reorientation of the amphipathic helices thus has to be postulated to explain the membrane integration, we considered that this conformational transition might also function in the opposite direction: The short TatA transmembrane helices could be in principle pulled out of the destabilized membrane into the ring where they align at the inner border of the ring as seen in the AF3 structure. As the tilted amphipathic helices generate a large circular hydrophobic cavity with the pulled-out transmembrane helices, the interacting lipids of the destabilized bilayer in the center of the ring would be pulled into this cavity, resulting in a transient membrane disruption and formation of an aqueous channel (Figure 4). After this switch, the TatA ring would be transformed from an integral membrane protein to a membrane surface-attached ring. Note that the transmembrane helices have been found to have an intrinsic ability to flip out of the membrane core, which has been suggested by the research group of Anne Ulrich to play a key role in TatA functionality [65]. This finding is in full agreement with our proposed mechanism of a TatA-mediated membrane disruption.
As at this time point the Tat substrate is already anchored to the trans-side of the membrane by its inserted TatBC-bound signal peptide, the folded protein can rapidly diffuse through this open pore. Only this open pore explains how long linkers between signal peptides and globular domains can be tolerated [51,52]. The efficiency of directional transport will gradually decrease with linker-length: As the signalpeptide insertase activity of TatBC unfolded the N-terminal region of the mature domain, this region will fold again at the moment when the lipids are removed, thereby pulling the mature domain to the trans-side. The contribution of this pulling to directional transport will be abolished by linkers between the signal peptide and the globular domain. Importantly, this pulling is not essential for transport by our proposed mechanism, but only globular proteins will be transported efficiently. In agreement with this finding, it has been clearly demonstrated that unfolded artificial Tat substrates are successively less efficiently translocated the longer the unfolded region is [66]. The cytoplasmic turgor in bacteria and archaea will result in a solvent flow through the pore and thus may accelerate the process of translocation of folded proteins, but this solvent flow cannot be essential because thylakoids or mitochondria have no cell wall and thus no significant turgor.
The membrane is locally depolarized when the pore is formed, and therefore even Tat substrates with high isoelectric points are accepted. A transient local depolarization definitively takes place, as transport of each Tat substrate has been shown in the plant Tat system to be accompanied by the leakage of an estimated ca. 79,000 protons, which supports the membrane-disruption mechanism [67].
In summary, the details of the AF3 structure gave us the idea of a mechanism that involves a controlled transient membrane disruption, which can explain the so far unexplained translocation of linker-containing Tat substrates. The diverse structural and functional details that perfectly fit to this idea cannot be coincidental, and as the ring also explains many other open questions, as outlined below, we think to have found the correct translocation mechanism.
The Membrane Disruption Mechanism Explains the TatBC Dependence and the 2nd Energy Requirement
To disrupt the weakened membrane bilayer in the TatA ring, the amphipathic helices need to change their conformation in a coordinated and TatBC-dependent way, as the signal peptide has to be inserted before the membrane disruption takes place. The main reasons for the involvement of TatBC are likely (i) a required specific signal peptide insertion for the proteins that need to be translocated, and (ii) a required energetization and organization for the concerted conformational change of all TatA amphipathic helices in the ring. The signal peptide insertion is needed to anchor the transported protein to the trans-side, and the twin-arginine motif therein serves as binding site for the signal peptide insertase and as determinant for the selection of Tat substrates. The energetization is needed because TatA rings exist in two states – membrane anchored and membrane extracted – and there is an energy barrier to overcome the transition between these two states. TatA is a protein at the membrane surface that is only membrane-anchored with one short transmembrane helix that evolved to be extractable, and thus TatA alone has no way to couple membrane energetization to a conformational change, which is why TatBC must energize this step.
Two of our studies showed an influence of TatBC on TatA topology that we could not explain so far: From a study that compares the substrate-induced effect on the TatA amphipathic helix orientation in the absence and presence of TatBC, we learned that substrate-binds to TatA already in the absence of TatBC, causing TatA conformational changes, but only in the presence of TatBC the region G33-S35 becomes inaccessible by substrate-binding, showing that TatBC specifically influences the TatA transition upon substrate-binding [21]. This could be explained by the AF3-structure, as access to this TatA region is hidden by the helices that are formed by the C-terminus of TatA. This is another evidence that the AF3 model must resemble the transport-induced structure. In the absence of TatBC, the G33-S35 region is always exposed – irrespective of substrate-binding. This is also supported by a study that showed a transport-dependence of TatA-TatA interactions in the C-terminal domain of TatA [57], and by another study that demonstrated a reduced accessibility of central and C-terminal parts of the TatA amphipathic helix during active transport [53]. In conclusion, without active transport, the C-terminal domain does not fold like seen in the AF3 structure and the amphipathic helices changes its environment, which indicates that transport is achieved by a structural change of TatA rings at substrate-bound TatBC complexes.
If a binding site must be transiently generated for the TatA conformational transition that results in membrane disruption, then this should be the exposed TatB binding site or TatB itself that in phase I has moved to the lateral opening, which depended on the electric potential. In our model, the domain movement placed the binding site with the bound TatB half-way out of the inner surface of the membrane (Figure 3B). It is possible that TatB has to be transiently released from TatC TM5 to permit an optimum TatA interaction. In this case, TatB might remain in the near, possibly anchored by interactions of the amphipathic helices. Alternatively, TatB can mediate the TatA interaction without being released from TM5. The interaction of the TatA ring with the TatBC complex must have two effects: (i) the TatA ring must open at the lateral gate, permitting the passage of the mature domain N-terminus that was membrane inserted during signal peptide insertion, and (ii) the interaction must lift the TatA ring, triggering the concerted extraction of the ring, which would disrupt the membrane and generate an aqueous hole.
At the TatBC complex, there is no evidence for energetization-dependent movements other than the described movement of the TatC TM5/TM6 domain and the reorientation of the internal TatB to the surface. The reason for the pmf-requirement for phase II may thus be an effect of the pmf on the TatA ring and/or the need to maintain the moved globular domain with the TatB stably exposed at the surface until the correct docking to the TatA ring is accomplished. Minor movements back into the interior of the TatBC complex must be prevented, which would make the crucial TatA interaction impossible, and the formation or opening and lifting of the ring onto the membrane surface could not take place.
Experimental Evidence for a Transient Lifting of TatA Rings onto the Membrane Surface
The two above-described states of the TatA ring – membrane-inserted versus membrane-attached – explain the finding of the research group of Yves Bollen and Holger Lill that large TatA complexes switch between fast and slow diffusion [54]. In that study, it has even been stated that the fast diffusion is too fast for a trans-membrane protein complex consisting of multiple TatA monomers. Based on these findings, they proposed “…that TatA complexes switch between a slowly diffusing transmembrane conformation and a rapidly diffusing membrane-disrupting state that enables folded proteins to cross the membrane…” [54]. The Tat mechanism that we propose fully supports their finding. The two states of TatA complexes also agree with the fact that about half of the TatA protomers can be stripped off from membranes by carbonate washes, whereas the other half cannot [21].
Priming of TatA Rings by Bound Substrate, and TatBC-Dependent Membrane Disruption by TatA
The effect of membrane-weakening is enhanced by substrate-association with TatA, indicating that the globular domain of the protein substrate itself influences the TatA-mediated destabilization [21]. The enhancement of membrane weakening likely involves above-mentioned orientational changes of the substrate-associated TatA amphipathic helices [21,53]. The substrate interactions with TatA are important, as they increase translocation efficiency [37]. For transport, it is crucial that the associated TatA ring is attached to TatBC, as the substrate must be translocated while the signal peptide is still bound to TatC. Otherwise, the diffusion through the pore would be undirected. Interestingly, we have observed that Tat substrate interactions with TatA result in much of the conformational changes already in the absence of TatBC, suggesting that substrate-interaction prime the TatBC interaction [21]. Especially the important movement of the hinge region to the surface of the TatA complex is already accomplished without TatBC, which reflects the transition of the amphipathic helix to a straight conformation that is seen in the AF3 structure. This suggests that TatA forms rings already in the absence of TatBC, and that substrate-binding thus already puts the transmembrane helices under tension, with the amphipathic helices on the membrane surface being reoriented and ready to interact with TatBC, and only a minor TatBC-dependent event is required to initiate the extraction of the transmembrane helices out of the membrane. This is further supported by the fact that substrate-binding already results in increased TatA-dependent proton leakage in the absence of TatBC [21].
It is likely that substrate-binding induces the TatA ring formation, as the in vitro translated and membrane-inserted TatA was quantitatively transformed into the hinge-exposing TatA upon saturating the system with substrates, and the surface-exposed hinge is a characteristic of the ring structure [21]. Substrate- and transport-dependent crosslinking of at least 16 TatA subunits was achieved with positions that interact in the C-terminal TatA region [15], which is predicted to form a structured outer ring upon lifting of the structure out of the membrane. Disulfide-crosslinking of the flexible transmembrane helix was less efficient but also enhanced under transport conditions, likely reflecting the more ordered packing of these helices when they are positioned in the inner surface of the ring upon their membrane extraction [15]. Disulfide cross-links with single cysteines were much less efficient but could be enhanced by transport conditions, likely reflecting substrate effects on TatA assemblies that have not yet formed a ring, which agrees with the view that substrates induce ring formation [57]. In MD simulations, TatA oligomers of more than 10 protomers that did not contact substrates have been seen to cluster with stagged transmembrane helices and parallel aligned amphipathic helices, which would explain single cysteine disulfide cross-links in the amphipathic helix [25].
After transport, the signal peptide is in a membrane-spanning N-inside/C-outside orientation, as known for Tat transport [68], and thus anchors the folded Tat substrate to the trans-side of the membrane, an intermediate state that has been identified as Ti-2 in in vitro studies of the plant Tat system [69]. The membrane pore rapidly closes by influx of lipids from the surrounding. The lipid-contacts of the amphipathic and transmembrane helices prevent that membrane disruption can continue beyond the inner area of the ring. Tat transport is thus a spatially controlled perforation of the membrane.
As membrane disruption is transient, there will be only a short time window for the passage of the folded substrate through the transient pore that is formed by a TatA-dependent membrane-weakening and -disruption. In case of a delay of transport due to larger unfolded regions and/or exposed lipid-interacting hydrophobic regions of the substrate, membrane sealing prevents translocation in phase II without blocking the Tat system, as phase I is completed and the signal peptide released from TatBC into the membrane. We showed already that targeting of unfolded, not translocated Tat substrates to the translocon does not block translocation [70]. The completion of phase I and a failure of phase II explains also that what has been termed “large scale translocation reversal” in a study of the plant Tat system in which signal peptides were Tat-dependently cleaved without successful translocation of an artificial cargo protein [71].
Phase III: Closing of the Catalytic Cycle and Resetting of the System
Release of the Signal Peptide Into the Membrane, and Reset of the TatBC Complex
The transported Tat substrate is initially captured in the TatBC complex. According to studies of the Musser-group, the release of the Tat substrate from the Tat system “requires a significant fraction of the total transport time” [72]. As long as the signal peptide is not released from the TatBC complex, the TatC TM5/TM6 domain with its associated TatB cannot switch back into the resting position. As the initial movement of the domain depended on the membrane energetization, the movement back to the resting position is likely facilitated by the local depolarization of the membrane. Once the complex is in the resting state, membrane energetization can build up again to energize the next translocation.
Reset of the TatA Rings to Membrane-Inserted Clusters That Can Interact with Tat Substrates
After transport, the TatA ring is released from the translocation site, because the TatC TM5/TM6 domain with its bound TatB re-inserts into the interior of the TatB complex upon de-energetization of the membrane. This fully agrees with fluorescence-studies that showed TatA release upon addition of uncoupler that cause dissipation of the pmf [73]. If TatB has to exchange with TatA for a successful TatA-ring-opening and -lifting at the lateral opening, the TatA that is bridging the TatBC complex dissociates when the TatC TM5/TM6 domain moves back, and TatB reassociates with the TatC TM5/TM6 domain. After sealing of the membrane, the ring thus floats on the membrane surface, as supported by fluorescence-tracking experiments [54]. Dissociated rings likely spontaneously close, and the substrate can become captured in closed rings. Since substrate-binding had supported ring formation and a conformational transition of the TatA amphipathic helices that exposed the hinge to the cytoplasm, the lack of substrate contacts on the cis side after transport reverses the conformational transition and thus destabilizes the ring, which permits the spontaneous reinsertion of the TatA transmembrane helix into the membrane, reaching again a low-energy state. Thereby, smaller assemblies of membrane-inserted TatA are generated that are ready to assemble to rings again after substrate-binding. TatBC independent substrate interactions may thus be important for targeting. Signal peptides can interact with lipid bilayers, and, depending on the substrate, signal peptides can be efficiently recruited from a lipid-inserted state [35,74]. Therefore, it is likely that this substrate-binding is facilitated by signal peptide insertion into the membrane. This would explain why TatA-substrate interactions were twin-arginine independent but depend on the signal peptide, although the mature domain contacts are also important [37]. With in vitro systems that used highly overproduced TatABC components and radiolabeled Tat substrates, a TatA interaction was found to be RR- and even pmf-dependent, indicating that this in vitro system did not monitor processes that occurred prior to TatBC binding and signal peptide insertion [39]. Note that although the TatBC-independent TatA interactions likely can target substrates to TatBC in a TatA ring that is already primed for translocation, it is not necessarily the same ring that is used for transport, as TatA-dependent targeting may not be the only path to TatBC. In principle, TatA rings need to be recruited to the translocation site in the course of targeting or they can assemble de novo at the translocon. In the latter case, the Tat substrate that is already positioned on the membrane at the lateral gate may support this process, but signal peptides alone already result in the formation of the required ordered oligomer at the TatBC complex [15]. The dimensions of the ring imply a contact to more than only TatC TM5. Accordingly, a pmf-dependent predicted surface-exposed TatA contact site was discovered at TM4 of TatC in plant systems (pea plastids, L231), which is nearby [28]. The pmf thus somehow contributes to TatA recruitment, and also membrane-thinning at TatBC likely contributes to the recruitment of TatA oligomers, as a hydrophobic mismatch is generated by membrane-integral TatA [25,55]. Based on the observation that TatC can organize the alignment of multiple TatA tubes [59], we think that multiple rings can be associated with each TatBC complex, and possibly several TatBC complexes can be clustered together with multiple TatA rings. Such lateral binding sites are likely responsible for the residual TatA detected in purified TatBC complexes by Deme et al. [9]. There is clear evidence from studies that did not involve fluorescent protein tags that TatBC complexes interact with large TatA assemblies [30], and that both TatA clusters as well as TatBC complexes are predominantly found near cell poles, supporting the view that the larger TatA assemblies cluster in close vicinity of TatBC [25,59].
The release of the Tat substrate from TatBC sterically permits the movement of the TatC TM5/6 domain together with its bound TatB back into the resting position. As the initial movement of the domain depended on the membrane energetization, the movement back to the resting position is likely facilitated by the local depolarization of the membrane. As a consequence, it can be concluded that the free flow of protons and other ions through the membrane likely supports the recovery of the complex and thus is a requirement for Tat transport.
In this context, the highly controversial aspect of soluble TatA should be mentioned. The loosely membrane-attached TatA rings that are generated by the translocation mechanism could in principle also dissociate from the membrane and be able to attach again for reinsertion. This would explain the detection of soluble TatA in several organisms, such as Bacillus subtilis [75,76], Streptomyces lividans [77], archaea [78], or plants [79,80]. In case of Bacillus subtilis, the soluble PhoD-specific TatA has been reported to play a role in substrate-targeting to the membrane [75,76]. Such a targeting function was proposed for Streptomyces as well [77]. In Streptomyces, TatA and TatB have more overlapping functions and thus the systems more resemble TatAC systems of Bacillus [77]. TatA and TatB are both also in the cytoplasm, and part of the TatA and TatB can be washed from membranes by carbonate washes, which is why Streptomyces “TatB” could be rather a bifunctional TatA [81]. These aspects were widely neglected in the Tat literature and should be reassessed.
The Tat System Requires a Functional Asymmetry
The above-described translocation mechanism involves changes of the membrane energetization at the translocation site that must be coordinated. While the twin-arginine recognition and hence substrate-binding functions at all three TatC subunits even in the absence of membrane energetization, the signal peptide insertase activity and the TatA ring extraction transition require a membrane energetization whereas membrane disruption depolarizes the translocation site. It is thus crucial for the Tat system to assure a sequential translocation, coordinated by TatBC complexes. There are in principle two alternative ways to achieve a functional asymmetry: (i) When translocation is initiated at one site, a regulatory mechanism can block the initiation of translocation at the two other sites, or (ii) a structural asymmetry can exist that permits translocation at only one site. TatAC systems of the Bacillus type must use the regulatory mechanism, and also E. coli likely uses such a regulatory mechanism in TatB3C3 complexes. However, in phylogenetically distant other systems structural asymmetry may have evolved, as it is possible that TatC-containing complexes differ in the number of involved subunits. A good example may be the Tat system of the halophilic archaeon Halobacterium salinarium, which possesses in addition to a “normal” TatC1 a large TatC2, which consists of two fused TatC that are connected by two additional transmembrane helices and a large cytoplasmic loop [82]. TatC1 and TatC2 are both encoded in direct vicinity, and we believe that they together form an asymmetric complex with likely only one functional substrate-binding site.
Conclusions
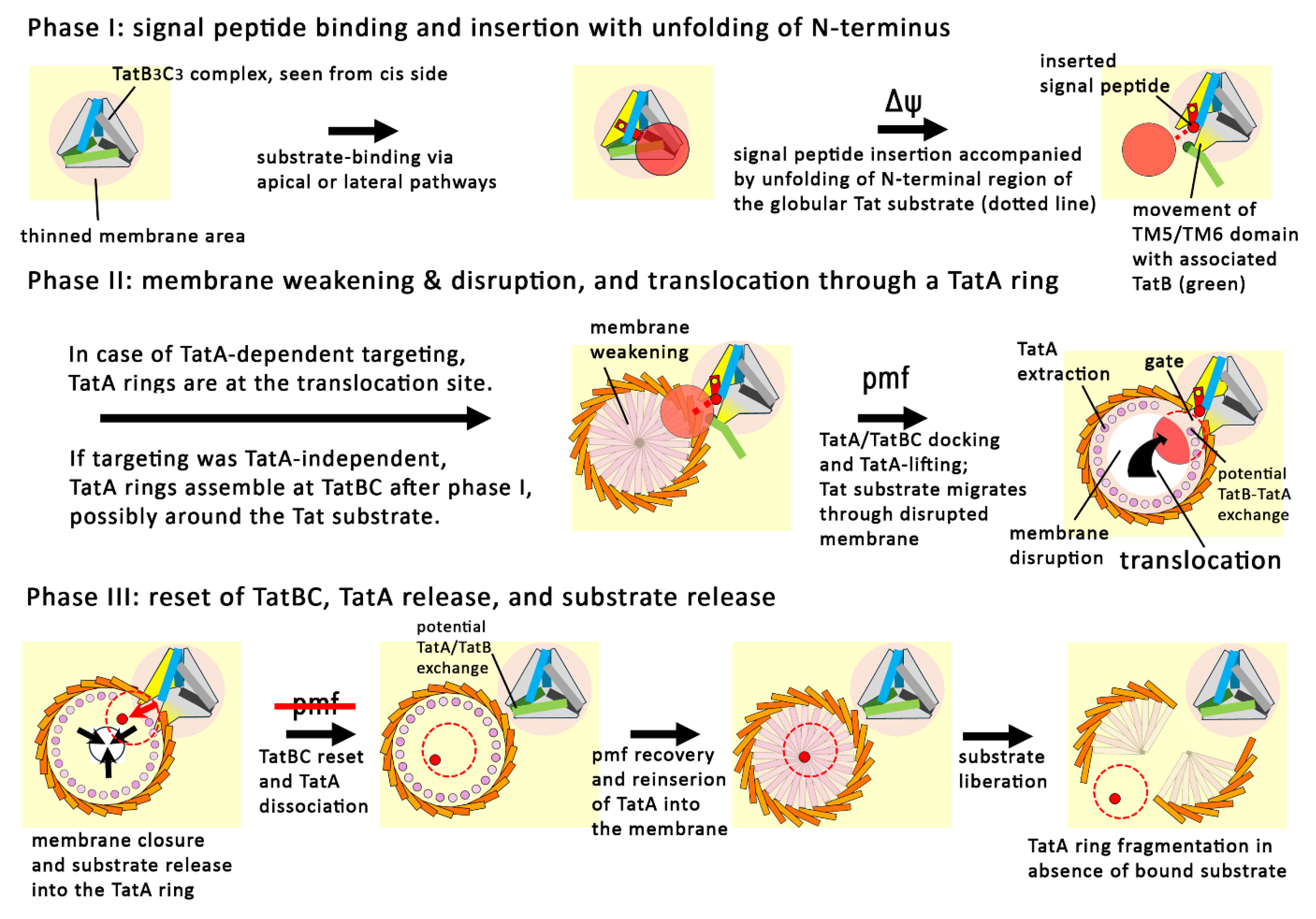
After more than 30 years of Tat research, the mechanism was still unclear and diverse puzzling results have been obtained that were not well decoded. To propose a convincing mechanism for Tat transport, it was important to respect the technically sound unexplained findings as facts, resulting in the key questions highlighted in this review. We were convinced that one day a mechanism could be found that explained all these facts, and this day has come. The solution of the TatBC complex structure with bound Tat substrates and the AF3 prediction of a meaningful TatA ring were the last puzzle pieces, and now everything could be integrated into a mechanistic model. Therefore, the key predictions of our model are already supported by experimental evidence. As schematically summarized in Figure 5, the transport occurs in two membrane energetization-dependent phases, with the signal peptide insertion and the membrane-disruption as main events, followed by a third phase for substrate release and translocon reset. Our model now permits to specifically address many details of the pathway, such as: What determines the targeting pathway to the translocon, and are distinct pathways preferred by distinct substrates or organisms? Is there a structural and/or functional heterogeneity of Tat systems from phylogenetically distant groups of organisms? Does the exposed TatB (or bifunctional TatA) need to be exchanged at TatC-TM5 by a TatA of a TatA ring? Does the exposed TatB or the TatA/B binding site at TatC TM5 open existing TatA rings or assemble TatA rings newly at the translocation site? Do the observed diverse TatA ring sizes represent detergent-induced fragments of a single physiologically relevant ring, or can TatA rings somehow adapt to the size of the translocated substrate? Many things are now understood, but Tat research remains exciting.
Figure 5.
Schematic summary of the series of events that have to take place for Tat transport of folded proteins across membranes. The process can be divided in three phases that comprise several steps, as indicated.
Figure 5.
Schematic summary of the series of events that have to take place for Tat transport of folded proteins across membranes. The process can be divided in three phases that comprise several steps, as indicated.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgments
We thank all former and current members of our group who in the last 30 years significantly contributed to the understanding of the Tat mechanism. This work was supported by the German Research Foundation (DFG) grant BR 2285/8-1 to TB.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Natale, P., Brüser, T., and Driessen, A. J. M. (2008) Sec- and Tat-mediated protein secretion across the bacterial cytoplasmic membrane - distinct translocases and mechanisms. Biochim Biophys Acta 1778, 1735–1756. [CrossRef] [PubMed]
- Yen, M.-R., Tseng, Y.-H., Nguyen, E. H., Wu, L.-F., and Saier, M. H. (2002) Sequence and phylogenetic analyses of the twin-arginine targeting (Tat) protein export system. Arch Microbiol 177, 441–450. [CrossRef] [PubMed]
- Lee, P. A., Buchanan, G., Stanley, N. R., Berks, B. C., and Palmer, T. (2002) Truncation analysis of TatA and TatB defines the minimal functional units required for protein translocation. J Bacteriol 184, 5871–5879. [CrossRef]
- Warren, G., Oates, J., Robinson, C., and Dixon, A. M. (2009) Contributions of the transmembrane domain and a key acidic motif to assembly and function of the TatA complex. J Mol Biol 388, 122–132.
- Barnett, J. P., Eijlander, R. T., Kuipers, O. P., and Robinson, C. (2008) A minimal Tat system from a gram-positive organism: a bifunctional TatA subunit participates in discrete TatAC and TatA complexes. J Biol Chem 283, 2534–2542. [CrossRef]
- Jongbloed, J. D., van der Ploeg, R., and van Dijl, J. M. (2006) Bifunctional TatA subunits in minimal Tat protein translocases. Trends Microbiol 14, 2–4. [CrossRef]
- Jongbloed, J. D. H., Grieger, U., Antelmann, H., Hecker, M., Nijland, R., Bron, S., and van Dijl, J. M. (2004) Two minimal Tat translocases in Bacillus. Mol Microbiol 54, 1319–1325. [CrossRef] [PubMed]
- Zhao, Z., and Sazanov, L. A. (2025) Structure of E. coli Twin-arginine translocase (Tat) complex with bound cargo. BioRxiv. [CrossRef]
- Deme, J. C., Bryant, O. j., Batista, M., Stansfeld, P. J., Berks, B. C., and Lea, S. M. (2025) Structure and substrate recognition by the Twin-arginine translocation (Tat) pathway core complex. BioRxiv. [CrossRef]
- Alami, M., Lüke, I., Deitermann, S., Eisner, G., Koch, H. G., Brunner, J., and Müller, M. (2003) Differential interactions between a twin-arginine signal peptide and its translocase in Escherichia coli. Mol Cell 12, 937–946. [CrossRef]
- Kreutzenbeck, P., Kroger, C., Lausberg, F., Blaudeck, N., Sprenger, G. A., and Freudl, R. (2007) Escherichia coli twin arginine (Tat) mutant translocases possessing relaxed signal peptide recognition specificities. J Biol Chem 282, 7903–7911. [CrossRef] [PubMed]
- Lausberg, F., Fleckenstein, S., Kreutzenbeck, P., Frobel, J., Rose, P., Muller, M., and Freudl, R. (2012) Genetic evidence for a tight cooperation of TatB and TatC during productive recognition of twin-arginine (Tat) signal peptides in Escherichia coli. PLoS ONE 7, e39867. [CrossRef] [PubMed]
- Ma, X., and Cline, K. (2013) Mapping the signal peptide binding and oligomer contact sites of the core subunit of the pea twin arginine protein translocase. Plant Cell 25, 999–1015. [CrossRef] [PubMed]
- Strauch, E. M., and Georgiou, G. (2007) Escherichia coli tatC mutations that suppress defective twin-arginine transporter signal peptides. J Mol Biol 374, 283–291. [CrossRef] [PubMed]
- Dabney-Smith, C., and Cline, K. (2009) Clustering of C-terminal stromal domains of Tha4 homo-oligomers during translocation by the Tat protein transport system. Mol Biol Cell. [CrossRef]
- Lüke, I., Handford, J. I., Palmer, T., and Sargent, F. (2009) Proteolytic processing of Escherichia coli twin-arginine signal peptides by LepB. Arch Microbiol 191, 919–925. [CrossRef]
- Hatzixanthis, K., Palmer, T., and Sargent, F. (2003) A subset of bacterial inner membrane proteins integrated by the twin-arginine translocase. Mol Microbiol 49, 1377–1390.
- Goosens, V. J., Monteferrante, C. G., and van Dijl, J. M. (2014) Co-factor insertion and disulfide bond requirements for twin-arginine translocase-dependent export of the Bacillus subtilis Rieske protein QcrA. J Biol Chem 289, 13124–13131. [CrossRef] [PubMed]
- Hopkins, A., Buchanan, G., and Palmer, T. (2014) Role of the twin arginine protein transport pathway in the assembly of the Streptomyces coelicolor cytochrome bc1 complex. J Bacteriol 196, 50–59. [CrossRef] [PubMed]
- Brüser, T., and Sanders, C. (2003) An alternative model of the twin arginine translocation system. Microbiol Res 158, 7–17. [CrossRef]
- Hou, B., Heidrich, E. S., Mehner-Breitfeld, D., and Brüser, T. (2018) The TatA component of the twin-arginine translocation system locally weakens the cytoplasmic membrane of Escherichia coli upon protein substrate binding. J Biol Chem 293, 7592–7605. [CrossRef] [PubMed]
- Asher, A. H., and Theg, S. M. (2021) Electrochromic shift supports the membrane destabilization model of Tat-mediated transport and shows ion leakage during Sec transport. Proc Natl Acad Sci U S A 118. [CrossRef] [PubMed]
- McNeilage, R., Ganesan, I., Keilman, J., and Theg, S. M. (2023) Cell-penetrating peptides stimulate protein transport on the Twin-arginine translocation pathway: evidence for a membrane thinning and toroidal pore mechanism. BioRxiv 2023.07.08.548235. [CrossRef]
- Rodriguez, F., Rouse, S. L., Tait, C. E., Harmer, J., Riso, A. de, Timmel, C. R., Sansom, M. S., Berks, B. C., and Schnell, J. R. (2013) Structural model for the protein-translocating element of the twin-arginine transport system. Proc Natl Acad Sci U S A 110, E1092-101. [CrossRef] [PubMed]
- Mehner-Breitfeld, D., Ringel, M. T., Tichy, D. A., Endter, L. J., Stroh, K. S., Lünsdorf, H., Risselada, H. J., and Brüser, T. (2022) TatA and TatB generate a hydrophobic mismatch important for the function and assembly of the Tat translocon in Escherichia coli. J Biol Chem 298, 102236. [CrossRef]
- Bolhuis, A., Mathers, J. E., Thomas, J. D., Barrett, C. M., and Robinson, C. (2001) TatB and TatC form a functional and structural unit of the twin-arginine translocase from Escherichia coli. J Biol Chem 276, 20213–20219. [CrossRef]
- Behrendt, J., and Brüser, T. (2014) The TatBC complex of the Tat protein translocase in Escherichia coli and its transition to the substrate-bound TatABC complex. Biochemistry 53, 2344–2354. [CrossRef] [PubMed]
- Aldridge, C., Ma, X., Gerard, F., and Cline, K. (2014) Substrate-gated docking of pore subunit Tha4 in the TatC cavity initiates Tat translocase assembly. J Cell Biol 205, 51–65. [CrossRef] [PubMed]
- Habersetzer, J., Moore, K., Cherry, J., Buchanan, G., Stansfeld, P. J., and Palmer, T. (2017) Substrate-triggered position switching of TatA and TatB during Tat transport in Escherichia coli. Open Biol 7. [CrossRef] [PubMed]
- Werner, M.-H., Mehner-Breitfeld, D., and Brüser, T. (2024) A larger TatBC complex associates with TatA clusters for transport of folded proteins across the bacterial cytoplasmic membrane. Sci Rep 14, 13754. [CrossRef] [PubMed]
- Barrett, C. M., Mangels, D., and Robinson, C. (2005) Mutations in subunits of the Escherichia coli twin-arginine translocase block function via differing effects on translocation activity or Tat complex structure. J Mol Biol 347, 453–463. [CrossRef]
- Celedon, J. M., and Cline, K. (2012) Stoichiometry for binding and transport by the twin arginine translocation system. J Cell Biol 197, 523–534. [CrossRef] [PubMed]
- Fröbel, J., Rose, P., Lausberg, F., Blümmel, A.-S., Freudl, R., and Müller, M. (2012) Transmembrane insertion of twin-arginine signal peptides is driven by TatC and regulated by TatB. Nat Commun 3, 1311. [CrossRef] [PubMed]
- Gerard, F., and Cline, K. (2006) Efficient twin arginine translocation (Tat) Pathway transport of a precursor protein covalently anchored to its initial cpTatC binding site. J Biol Chem 281, 6130–6135. [CrossRef]
- Bageshwar, U. K., Whitaker, N., Liang, F. C., and Musser, S. M. (2009) Interconvertibility of lipid- and translocon-bound forms of the bacterial Tat precursor pre-SufI. Mol Microbiol 74, 209–226. [CrossRef] [PubMed]
- Ramasamy, S., Abrol, R., Suloway, C. J. M., and Clemons, W. M. (2013) The glove-like structure of the conserved membrane protein TatC provides insight into signal sequence recognition in twin-arginine translocation. Structure 21, 777–788. [CrossRef] [PubMed]
- Taubert, J., Hou, B., Risselada, H. J., Mehner, D., Lünsdorf, H., Grubmüller, H., and Brüser, T. (2015) TatBC-independent TatA/Tat substrate interactions contribute to transport efficiency. PLoS ONE 10, e0119761. [CrossRef] [PubMed]
- Rathmann, C., Schlösser, A. S., Schiller, J., Bogdanov, M., and Brüser, T. (2017) Tat transport in Escherichia coli requires zwitterionic phosphatidylethanolamine but no specific negatively charged phospholipid. FEBS Lett 591, 2848–2858. [CrossRef] [PubMed]
- Fröbel, J., Rose, P., and Müller, M. (2011) Early contacts between substrate proteins and TatA translocase component in twin-arginine translocation. J Biol Chem 286, 43679–43689. [CrossRef] [PubMed]
- Blaudeck, N., Kreutzenbeck, P., Müller, M., Sprenger, G. A., and Freudl, R. (2005) Isolation and characterization of bifunctional Escherichia coli TatA mutant proteins that allow efficient Tat-dependent protein translocation in the absence of TatB. J Biol Chem 280, 3426–3432. [CrossRef] [PubMed]
- Taubert, J., and Brüser, T. (2014) Twin-arginine translocation-arresting protein regions contact TatA and TatB. Biol Chem 395, 827–836. [CrossRef] [PubMed]
- Klösgen, R. B., Brock, I. W., Herrmann, R. G., and Robinson, C. (1992) Proton gradient-driven import of the 16 kDa oxygen-evolving complex protein as the full precursor protein by isolated thylakoids. Plant Mol Biol 18, 1031–1034. [CrossRef]
- Cline, K., Ettinger, W. F., and Theg, S. M. (1992) Protein-specific energy requirements for protein transport across or into thylakoid membranes. Two lumenal proteins are transported in the absence of ATP. J Biol Chem 267, 2688–2696. [PubMed]
- Braun, N. A., Davis, A. W., and Theg, S. M. (2007) The chloroplast Tat pathway utilizes the transmembrane electric potential as an energy source. Biophys J 93, 1993–1998. [CrossRef]
- Theg, S. M., Cline, K., Finazzi, G., and Wollman, F. A. (2005) The energetics of the chloroplast Tat protein transport pathway revisited. Trends Plant Sci 10, 153–154. [CrossRef]
- Bageshwar, U. K., and Musser, S. M. (2007) Two electrical potential-dependent steps are required for transport by the Escherichia coli Tat machinery. J Cell Biol 179, 87–99. [CrossRef] [PubMed]
- Buchanan, G., Leeuw, E., Stanley, N. R., Wexler, M., Berks, B. C., Sargent, F., and Palmer, T. (2002) Functional complexity of the twin-arginine translocase TatC component revealed by site-directed mutagenesis. Mol Microbiol 43, 1457–1470. [CrossRef]
- Maurer, C., Panahandeh, S., Jungkamp, A.-C., Moser, M., and Müller, M. (2010) TatB Functions as an Oligomeric Binding Site for Folded Tat Precursor Proteins. Mol Biol Cell 21, 4151–4161.
- Berghöfer, J., and Klösgen, R. B. (1999) Two distinct translocation intermediates can be distinguished during protein transport by the TAT (Dph) pathway across the thylakoid membrane. FEBS Lett 460, 328–332.
- Gerard, F., and Cline, K. (2007) The thylakoid proton gradient promotes an advanced stage of signal peptide binding deep within the Tat pathway receptor complex. J Biol Chem.
- Lindenstrauss, U., and Brüser, T. (2009) Tat transport of linker-containing proteins in Escherichia coli. FEMS Microbiol Lett 295, 135–140. [CrossRef] [PubMed]
- Cline, K., and McCaffery, M. (2007) Evidence for a dynamic and transient pathway through the TAT protein transport machinery. EMBO J. 26, 3039–3049.
- Aldridge, C., Storm, A., Cline, K., and Dabney-Smith, C. (2012) The chloroplast twin arginine transport (Tat) component, Tha4, undergoes conformational changes leading to Tat protein transport. J Biol Chem 287, 34752–34763. [CrossRef] [PubMed]
- Varadarajan, A., Oswald, F., Lill, H., Peterman, E. J., and Bollen, Y. J. M. (2020) Rapid diffusion of large TatA complexes detected using single particle tracking microscopy. BioRxiv, 2020.05.14.095463. [CrossRef]
- Hao, B., Zhou, W., and Theg, S. M. (2022) Hydrophobic mismatch is a key factor in protein transport across lipid bilayer membranes via the Tat pathway. J Biol Chem 298, 101991. [CrossRef] [PubMed]
- Fan, E., Jakob, M., and Klösgen, R. B. (2010) One signal is enough: Stepwise transport of two distinct passenger proteins by the Tat pathway across the thylakoid membrane. Biochem Biophys Res Commun 398, 438–443. [CrossRef]
- Dabney-Smith, C., Mori, H., and Cline, K. (2006) Oligomers of Tha4 organize at the thylakoid Tat translocase during protein transport. J Biol Chem 281, 5476–5483. [CrossRef]
- Gohlke, U., Pullan, L., McDevitt, C. A., Porcelli, I., Leeuw, E. de, Palmer, T., Saibil, H. R., and Berks, B. C. (2005) The TatA component of the twin-arginine protein transport system forms channel complexes of variable diameter. Proc Natl Acad Sci U S A 102, 10482–10486. [CrossRef] [PubMed]
- Berthelmann, F., Mehner, D., Richter, S., Lindenstrauss, U., Lünsdorf, H., Hause, G., and Brüser, T. (2008) Recombinant expression of tatABC and tatAC results in the formation of interacting cytoplasmic TatA tubes in Escherichia coli. J Biol Chem 283, 25281–25289. [CrossRef] [PubMed]
- Jormakka, M., Törnroth, S., Byrne, B., and Iwata, S. (2002) Molecular basis of proton motive force generation: structure of formate dehydrogenase-N. Science 295, 1863–1868. [CrossRef] [PubMed]
- Abramson, J., Adler, J., Dunger, J., Evans, R., Green, T., Pritzel, A., Ronneberger, O., Willmore, L., Ballard, A. J., Bambrick, J., Bodenstein, S. W., Evans, D. A., Hung, C.-C., O'Neill, M., Reiman, D., Tunyasuvunakool, K., Wu, Z., Žemgulytė, A., Arvaniti, E., Beattie, C., Bertolli, O., Bridgland, A., Cherepanov, A., Congreve, M., Cowen-Rivers, A. I., Cowie, A., Figurnov, M., Fuchs, F. B., Gladman, H., Jain, R., Khan, Y. A., Low, C. M. R., Perlin, K., Potapenko, A., Savy, P., Singh, S., Stecula, A., Thillaisundaram, A., Tong, C., Yakneen, S., Zhong, E. D., Zielinski, M., Žídek, A., Bapst, V., Kohli, P., Jaderberg, M., Hassabis, D., and Jumper, J. M. (2024) Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 630, 493–500. [CrossRef] [PubMed]
- Hou, B., and Brüser, T. (2011) The Tat-dependent protein translocation pathway. Biomol Concepts 2, 507–523. [CrossRef] [PubMed]
- Hicks, M. G., Leeuw, E. de, Porcelli, I., Buchanan, G., Berks, B. C., and Palmer, T. (2003) The Escherichia coli twin-arginine translocase: conserved residues of TatA and TatB family components involved in protein transport. FEBS Lett 539, 61–67. [CrossRef]
- Walther, T. H., Grage, S. L., Roth, N., and Ulrich, A. S. (2010) Membrane alignment of the pore-forming component TatA(d) of the twin-arginine translocase from Bacillus subtilis resolved by solid-state NMR spectroscopy. J Am Chem Soc 132, 15945–15956. [CrossRef] [PubMed]
- Stockwald, E. R., Steger, L. M. E., Vollmer, S., Gottselig, C., Grage, S. L., Bürck, J., Afonin, S., Fröbel, J., Blümmel, A.-S., Setzler, J., Wenzel, W., Walther, T. H., and Ulrich, A. S. (2022) Length matters: Functional flip of the short TatA transmembrane helix. Biophys J 122, 2125–2146. [CrossRef] [PubMed]
- Richter, S., Lindenstrauss, U., Lücke, C., Bayliss, R., and Brüser, T. (2007) Functional Tat transport of unstructured, small, hydrophilic proteins. J Biol Chem 282, 33257–33264. [CrossRef] [PubMed]
- Alder, N. N., and Theg, S. M. (2003) Energetics of protein transport across biological membranes. a study of the thylakoid ΔpH-dependent/cpTat pathway. Cell 112, 231–242. [CrossRef]
- Fincher, V., McCaffery, M., and Cline, K. (1998) Evidence for a loop mechanism of protein transport by the thylakoid ΔpH pathway. FEBS Lett 423, 66–70. [CrossRef]
- Hou, B., Frielingsdorf, S., and Klösgen, R. B. (2006) Unassisted membrane insertion as the initial step in ΔpH/Tat-dependent protein transport. J Mol Biol 355, 957–967. [CrossRef]
- Richter, S., and Brüser, T. (2005) Targeting of unfolded PhoA to the TAT translocon of Escherichia coli. J Biol Chem 280, 42723–42730. [CrossRef] [PubMed]
- Di Cola, A., and Robinson, C. (2005) Large-scale translocation reversal within the thylakoid Tat system in vivo. J Cell Biol 171, 281–289. [CrossRef]
- Whitaker, N., Bageshwar, U. K., and Musser, S. M. (2012) Kinetics of precursor interactions with the bacterial Tat translocase detected by real-time FRET. J Biol Chem 287, 11252–11260. [CrossRef] [PubMed]
- Alcock, F., Baker, M. A. B., Greene, N. P., Palmer, T., Wallace, M. I., and Berks, B. C. (2013) Live cell imaging shows reversible assembly of the TatA component of the twin-arginine protein transport system. Proc Natl Acad Sci U S A 110, E3650-9. [CrossRef] [PubMed]
- Shanmugham, A., Wong Fong Sang, H. W., Bollen, Y. J., and Lill, H. (2006) Membrane binding of twin arginine preproteins as an early step in translocation. Biochemistry 45, 2243–2249. [CrossRef]
- Pop, O. I., Westermann, M., Volkmer-Engert, R., Schulz, D., Lemke, C., Schreiber, S., Gerlach, R., Wetzker, R., and Müller, J. P. (2003) Sequence-specific binding of prePhoD to soluble TatAd indicates protein-mediated targeting of the Tat export in Bacillus subtilis. J Biol Chem 278, 38428–38436. [CrossRef]
- Schreiber, S., Stengel, R., Westermann, M., Volkmer-Engert, R., Pop, O. I., and Müller, J. P. (2006) Affinity of TatCd for TatAd elucidates its receptor function in the Bacillus subtilis Tat translocase system. J Biol Chem 281, 19977–19984. [CrossRef]
- Keersmaeker, S. de, van Mellaert, L., Lammertyn, E., Vrancken, K., Anne, J., and Geukens, N. (2005) Functional analysis of TatA and TatB in Streptomyces lividans. Biochem Biophys Res Commun 335, 973–982. [CrossRef] [PubMed]
- Dilks, K., Gimenez, M. I., and Pohlschröder, M. (2005) Genetic and biochemical analysis of the twin-arginine translocation pathway in halophilic archaea. J Bacteriol 187, 8104–8113. [CrossRef]
- Frielingsdorf, S., Jakob, M., and Klösgen, R. B. (2008) A stromal pool of TatA promotes Tat-dependent protein transport across the thylakoid membrane. J Biol Chem 283, 33838–33845. [CrossRef] [PubMed]
- Pettersson, P., Patrick, J., Jakob, M., Jacobs, M., Klösgen, R. B., Wennmalm, S., and Mäler, L. (2021) Soluble TatA forms oligomers that interact with membranes: Structure and insertion studies of a versatile protein transporter. Biochim Biophys Acta 1863, 183529. [CrossRef] [PubMed]
- Keersmaeker, S. de, van Mellaert, L., Schaerlaekens, K., van Dessel, W., Vrancken, K., Lammertyn, E., Anne, J., and Geukens, N. (2005) Structural organization of the twin-arginine translocation system in Streptomyces lividans. FEBS Lett 579, 797–802. [CrossRef] [PubMed]
- Thomas, J. R., and Bolhuis, A. (2006) The tatC gene cluster is essential for viability in halophilic archaea. FEMS Microbiol Lett 256, 44–49. [CrossRef] [PubMed]
- Goddard, T. D., Huang, C. C., Meng, E. C., Pettersen, E. F., Couch, G. S., Morris, J. H., and Ferrin, T. E. (2018) UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Protein Sci 27, 14–25. [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



