Submitted:
03 January 2026
Posted:
05 January 2026
You are already at the latest version
Abstract
Extended-spectrum β-lactamase–producing E. coli (ESBL-EC) threatens public health by driving widespread antimicrobial resistance transmission in environmental and agricultural settings. This study examined the prevalence, genetic determinants, and phylogenetic relationships of ESBL-EC isolated from municipal wastewater treatment plants (WWTPs) and farm environments in southeastern North Carolina. A cross-sectional survey was conducted between May and September 2025 across two WWTPs and two farms (cattle and poultry). We sampled influent and effluent wastewater, plus fecal and water specimens collected from chickens, ducks, and cattle. Antimicrobial susceptibility testing was performed using Kirby–Bauer disk diffusion method against nine drugs, while PCR and sequencing were used for genotypic characterization. Phylogenetic analysis assessed genetic relatedness among isolates. ESBL-EC was detected in 27.4% (n = 124) of 452 samples, with the highest prevalence in chickens (31.5%), followed by WWTP influents (28.2%), ducks (18.5%), and cattle (12.1%). Dominant resistance genes included blaCMY-2 (71.8%), blaCTX-M-1 and blaOXA (54% each), and blaSHV (29.8%). Co-occurrence of blaCMY-2 with blaCTX-M-1 and blaOXA was observed in poultry isolates. Phylogenetic analysis revealed clonal relatedness between poultry and cattle isolates. These findings highlight poultry as a key reservoir and emphasize the need for One Health surveillance to mitigate cross-reservoir transmission of resistant E. coli.
Keywords:
Escherichia coli
; beta-lactamases
; drug resistance
; bacterial
; wastewater
; agriculture
; poultry
; cattle
; phylogeny
; One Health
1. Introduction
Escherichia coli is a versatile bacterial species that plays essential roles in both human and animal health, including environmental systems [1,2,3]. While many strains are harmless commensals that support digestion and synthesize vital nutrients, others have acquired virulence and antimicrobial resistance (AMR) determinants that enable them to cause significant disease, including urinary tract infections, sepsis, and neonatal meningitis [2,4,5]. Because E. coli is shed in large numbers in the feces of humans and animals, it is widely used as an indicator organism for fecal contamination in aquatic and terrestrial environments [6,7]. Municipal wastewater discharge, agricultural runoff, and direct deposition from wildlife are key pathways through which E. coli enters environmental systems [8]. Studies show that bacteria can persist even after conventional wastewater treatment [9,10].
Beyond its value as a fecal indicator, E. coli serves as a sentinel organism for monitoring AMR in the environment [11,12,13]. Its ubiquity, genomic plasticity, and ability to acquire and exchange resistance genes across human, animal, and environmental interfaces make it a cornerstone organism in One Health AMR surveillance [14,15]. Among the most concerning resistance phenotypes are extended-spectrum β-lactamase–producing E. coli (ESBL-EC) [3,16]. ESBL EC—encoded predominantly by genes such as blaCTX-M, blaTEM, blaSHV, and blaCMY-2—hydrolyze a broad range of β-lactam antibiotics, including third-generation cephalosporins [16,17]. These enzymes severely restrict treatment options and are associated with increased morbidity, mortality, and healthcare costs [17]. Global surveillance indicates that over 40% of E. coli isolates are now resistant to third-generation cephalosporins, underscoring the expanding healthcare challenge posed by ESBL-producing strains [18,19].
In the United States, CDC has reported rising ESBL-EC infections in both healthcare and community settings [20]. Parallel increases have been documented in environmental reservoirs, reflecting contributions from wastewater, hospital effluents, and animal agriculture—sectors where antimicrobial use and human activity create conditions favorable for the emergence and dissemination of resistant bacteria [21]. Wastewater treatment plants (WWTPs) are particularly critical points in this transmission network, as they collect waste from households, hospitals, and industries, making them important indicators of community-level resistance trends [8,21,22]. Similarly, poultry and cattle operations have been identified as hotspots for AMR due to routine antibiotic use and the high density of animals, which facilitate horizontal gene transfer and the amplification of resistant strains [3,14,23,24].
Adopting a One Health perspective aligns with global recommendations that emphasize coordinated AMR surveillance across human, animal, and environmental domains [22,25]. Integrated monitoring provides a more comprehensive understanding of how resistance genes persist, move between ecosystems, and potentially re-enter human populations [26,27]. Despite the recognized importance of environmental reservoirs in AMR dissemination, several gaps remain [28]. Few studies integrate wastewater and livestock settings within the same surveillance framework, and even fewer examine the genetic relatedness of isolates from these systems [29,30].
In southeastern North Carolina, limited data exists on the occurrence and molecular characteristics of ESBL-EC across municipal wastewater and agricultural environments. Consequently, the degree to which environmental strains contribute to cross-reservoir movement of ESBL genes—and whether they mirror clinically relevant resistance patterns—remains poorly understood [31]. This knowledge gap limits the ability of regional public health systems to assess risk, identify transmission pathways, and implement targeted interventions. Environmental and agricultural reservoirs may play an underrecognized role in maintaining and disseminating ESBL determinants, yet the extent of overlap between wastewater and livestock sources remains unclear [32]. An integrated assessment across these settings is therefore essential for understanding how resistant E. coli circulate and whether similar genetic lineages or resistance genes occur across human, animal, and environmental interfaces [3,14].
Generating locally relevant data is especially important in regions where wastewater systems and livestock operations coexist yet remain understudied in the context of ESBL-EC. Therefore, the aim of this study was to generate baseline data on ESBL-EC circulating in municipal wastewater treatment plants and livestock farm environments in southeastern North Carolina. Using culture-based detection, antimicrobial susceptibility testing, molecular characterization of ESBL-associated genes, and phylogenetic analysis, we examined the occurrence, distribution, and genetic relatedness of ESBL-EC across these systems. By comparing isolates from wastewater influent and effluent with those from poultry and cattle environments, this study evaluated the extent to which these settings may function as interconnected reservoirs of AMR and contribute to regional transmission dynamics.
2. Materials and Methods
2.1. Study Design and Sampling Sites
A cross-sectional study was conducted from May to September 2025 in Fayetteville, North Carolina. Environmental samples were collected from two WWTPs, A and B, and two agricultural sites: a beef cattle farm and a mixed-species backyard poultry farm housing chickens and ducks. At both WWTPs, influent, effluent, and sludge samples were collected. Environmental sponge swabs were also obtained from high-contact surfaces (ropes, walls, and poles) to assess biofilm-associated contamination. One of the wastewater sampling events was conducted shortly after a storm event. Influent to the WWTPs originated from hospitals, residential households, and industrial sources and did not include direct animal inputs. Agricultural sampling included fecal samples, soil, lake water, drinking water, and trough swabs from the cattle farm and fecal samples from chickens and ducks, drinking water, and trough swabs from the poultry farm. All sites were sampled twice monthly to capture temporal and environmental variability.
2.2. Sample Collection and Processing
All samples were collected aseptically in sterile containers, immediately placed on ice, and transported to the laboratory for processing within 6 hours. Sponge swabs from wastewater facilities were immersed in 20 mL of buffered peptone water (BPW) and incubated at 37°C for 6 hours. Fecal and soil samples were homogenized in 9 mL BPW and incubated similarly. Water samples were processed by plate spreading or membrane filtration depending on clarity. All procedures were conducted under BSL-2 laboratory conditions.
2.3. Isolation and Identification of E. coli
Samples were plated onto MacConkey agar and MacConkey agar supplemented with 1 µg/mL cefotaxime to isolate presumptive E. coli and potential ESBL-producing strains. Plates were incubated at 37°C for 24 hours. All colonies were enumerated for effluent samples. For influent, sludge, fecal, and soil, only plates with 13–300 colony forming units (CFU) were counted. Representative colonies were selected for confirmation. Presumptive E. coli colonies were streaked onto Eosin Methylene Blue (EMB) agar. Colonies exhibiting a characteristic metallic green sheen were confirmed as E. coli. Pure colonies were sub-cultured on tryptone soy agar for downstream analysis.
2.4. Antimicrobial Susceptibility Testing
Phenotypic susceptibility was assessed using the Kirby–Bauer disk diffusion method on Mueller–Hinton agar following CLSI guidelines [33]. The antimicrobial susceptibility panel included β-lactams (ceftazidime, cefotaxime, ampicillin, and penicillin) as well as agents from other classes, namely tetracycline, streptomycin, trimethoprim-sulfamethoxazole, chloramphenicol, and ciprofloxacin. ESBL production was confirmed using the CLSI combination disk test with ceftazidime and ceftazidime–clavulanic acid (CAZ/CLA), and cefotaxime and cefotaxime–clavulanic acid (CTX/CLA). An increase of ≥5 mm in zone diameter for CAZ/CLA or CTX/CLA relative to ceftazidime or cefotaxime alone indicated ESBL positivity.
2.5. Genotypic Detection of Resistance Genes Using PCR
DNA was extracted using the boiling method, in which a loopful of bacterial biomass was suspended in sterile water, boiled for 15 minutes, centrifuged briefly, and the supernatant collected as template DNA. Singleplex PCR assays were performed to detect blaTEM, blaCMY-2, and sul2, while a multiplex PCR assay targeted the blaSHV, blaOXA-1, and blaCTX-M-1 gene families. Reactions were prepared with Phusion Master Mix and primers at 10 µM, following validated cycling protocols as previously described [34]. PCR amplification was carried out under the following thermal cycling parameters: an initial denaturation step at 94°C for 10 minutes, followed by 30 cycles consisting of denaturation at 94 °C for 30 seconds, annealing for 35 seconds at gene-specific temperatures (60.1°C for blaTEM, 58°C for blaSHV, 60.1°C for blaOXA-1, 57.8°C for blaCTX-M-1, and 57°C for blaCMY-2), and extension at 72°C for 1 minute. A final extension was performed at 72°C for 9 minutes. For singleplex PCR reactions (25 µl total volume), primers were added at a final concentration of 0.2 µM using a 10 µM primer stock, and 1 µl of DNA lysate was used as the template. Expected amplicon sizes ranged from 286 to 1143 bp. PCR products were separated in a 1% agarose gel at 80 V for 1 h 20 min and visualized in UV light after staining with SBYR safe DNA gel stain (Invitrogen, Carlsbad CA, USA) for 3min. A 1 kb DNA ladder (ThermoFisher Scientific, USA) was used as a molecular size marker. All PCR experiments were carried out using SimpliAmpTM Thermal cycler (ThermoFisher Scientific, USA).
2.6. DNA Extraction, Library Preparation and Whole-Genome Sequencing
A subset of phenotypically confirmed generic E. coli and ESBL-EC isolates (selected based on gene profiles, source type, and resistance patterns) underwent whole-genome sequencing (WGS). Genomic DNA was extracted from selected isolates using the DNeasy PowerLyzer Microbial Kit (Qiagen, Germantown, MD, USA) following the manufacturer’s Spin Wash method instructions. Bacterial colonies were collected from agar plates and resuspended in Power Bead Solution with SL Solution, followed by mechanical lysis using the Bead Mill 24. Lysates were centrifuged, and the supernatant was treated sequentially with IRS and SB Solutions before being applied to spin columns. Columns were washed twice with CB Solution, and DNA was eluted in 50 µL EB Solution. Extracted DNA was quantified through Nanodrop, prepared using standard library preparation kits, and sequenced on an Illumina platform generating paired-end reads.
Genomic libraries with an insert size of approximately 350 bp were prepared using the Illumina DNA Prep Kit and Illumina DNA/RNA UD Indexes (Illumina, San Diego, CA, USA). The pooled library was loaded into an Illumina NextSeq 1000/2000 P1 Reagent Kit (600 cycles) and sequenced on the NextSeq 2000 platform to generate paired-end 250 bp reads.
Sequencing quality was assessed using the MicroRunQC workflow, a custom GalaxyTrakr instance for the GenomeTrakr surveillance network (protocols.io). Output included quality assessments of raw reads (Illumina paired-end FASTQ files) and draft de novo assemblies, as well as multilocus sequence typing for each isolate. Summary metrics included sequence quality, number of contigs, estimated genome size, sequence type (ST), and other standard read and assembly statistics (e.g., median insert size, mean read length, and N50).
2.7. Bioinformatics and Phylogenetic Analysis
Raw reads were trimmed and assembled using SPAdes [35]. Genomes were annotated using Prokka [36]. Single nucleotide polymorphisms (SNPs) were identified using Snp dists, and a core genome SNP alignment was generated. A maximum-likelihood phylogenetic tree was constructed using RAxML to assess genetic relatedness among isolates across wastewater and agricultural environments [37].
2.8. Data Analysis
Prevalence, colony counts, and proportions of ESBL-positive isolates were calculated for each site and sample type. AST results were interpreted according to CLSI breakpoints [38]. PCR data were recorded as presence/absence of each gene. WGS and SNP-based phylogenetic results were integrated to assess clonality and cross-environment relatedness. WGS data for bacterial pathogens that met QC criteria, along with associated metadata, were submitted to NCBI for further analysis within the NCBI Pathogen Detection system. All isolates were assigned unique Sequence Read Archive (SRA) and BioSample accessions under the organizing BioProject PRJNA293225. Descriptive statistics were generated using Microsoft Excel, R for statistical analysis (vs 4.5.2) and phylogenetic outputs were visualized using iTOL version 7 [39].
3. Results
3.1. Prevalence of E. coli in Wastewater, Poultry, Cattle and Farm Environment
Overall, 452 samples were collected, including wastewater (n = 168), animal feces (n = 128), farm water (n = 126), and farm soil (n = 30). Sampling was conducted at two municipal wastewater treatment plants—WWTP-A (n = 88) and WWTP-B (n = 80)—as well as a commercial cattle farm (n = 129) and backyard poultry farm (n = 155) in Fayetteville. The overall prevalence of E. coli from all sources was 33% (n = 149) out of which 45% (n = 67) were obtained from a backyard poultry farm and 23.5% (n = 35) from a cattle farm. Out of 67 E. coli isolates from the poultry farm, 56.7% (n = 38) were from chicken feces; 38.8% (n = 26) from duck feces, while 4.5% (n = 3) were from water samples. Out of 35 E. coli isolates from the cattle farm, 60% (n = 21) were from feces; 25.7% (n = 9) from soil, while 14.3% (n = 5) were from water samples. Forty-seven E. coli isolates were obtained from both WWTPs out of which 61.7% (n = 29) were from WWTP-A while 38.3% (n = 18) were from WWTP-B. Of these, 63.8% (n=30) originated from influent (untreated) samples while 36.2% (n=17) were from effluent (treated) samples.
3.2. Detection of ESBL-EC Isolates
Of 452 samples, 27.4% (n = 124) were confirmed as ESBL producers by phenotypic characterization. Prevalence of ESBL-EC was highest among poultry samples (54.8%, n = 68), followed by WWTP-A (17.7%, n = 22), cattle samples (13.7%, n = 17), and WWTP-B (13.7%, n = 17). More than half (62.9%) of these isolates originated from animal feces, followed by WWTP samples (Figure 1).
3.3. Antimicrobial Susceptibility Testing of ESBL-EC Isolates
Of the 124 ESBL-EC isolates examined, 82.3% (n = 102) were multidrug-resistant (MDR). The majority of MDR isolates were recovered from poultry farms (91.2%, 62/68), followed by wastewater samples at WWTP-A (95.5%, 21/22) and WWTP-B (70.6%, 12/17). The lowest proportion was observed among isolates from cattle farms (41.2%, 7/17), as illustrated in Figure 2. Antibiotic resistance among ESBL-EC isolates was highest to penicillin (97.6%, n = 121), ampicillin (96.0%, n = 119), and cefotaxime (96.0%, n = 119). Moderate resistance was observed to tetracycline (67.7%, n = 84) and ceftazidime (58.9%, n = 73), while lower resistance frequencies were noted for sulfamethoxazole + trimethoprim (34.7%, n = 43), streptomycin (33.9%, n = 42), ciprofloxacin (25.0%, n = 31), and chloramphenicol (15.3%, n = 19) as shown in Table 1.
3.4. PCR-Based Detection of Resistance Genes
PCR analysis of 124 ESBL-EC isolates detected several resistance genes, including blaCMY2, blaCTX-M-1, blaOXA, blaSHV, blaTEM, qnrS, and sul2. The majority of these were identified in poultry farm samples, particularly from chicken and duck feces. Among them, sul2 was the most frequent, detected in 57 isolates, followed by blaCMY2 in 51 isolates and blaOXA in 40 isolates (Figure 3a). The prevalence of blaCMY2 was highest in poultry, followed by isolates from WWTP-A, cattle farms, and WWTP-B. Additionally, blaCMY, blaSHV, blaTEM, qnrS, and sul2 genes were detected in effluent wastewater samples from WWTP-A (Figure 3b).
3.5. WGS-Based Detection of Resistance Genes
WGS identified 48 distinct AMR determinants across 55 E. coli isolates, of which 24 (43.6%) were classified as ESBL-EC. The most prevalent determinants are presented in Figure 3c. The mdf(A) gene which encodes a transporter protein that contributes to multidrug resistance, was detected in 38 isolates. Fifteen different β-lactamase gene variants were identified, including blaACT-16, blaCTX-M-1, blaCTX-M-15, blaCTX-M-32, blaCTX-M-55, blaCTX-M-124, blaMIR-1, blaOXA-1, blaOXA-344, blaOXA-396, blaOXY-1, blaPAO, blaSHV-106, blaTEM-1A, and blaTEM-1B. Among these, the blaCTX-M variants were the classical markers of ESBL-EC. The blaCMY gene, which encodes an AmpC-type β-lactamase, was detected in four distinct variants: blaCMY-2, blaCMY-80, blaCMY-110, and blaCMY-152. Aminoglycosides were the most detected with seven different variants (aadA5, aac(3)-Ila, aac(6)-Ib-cr, aph(3')-Ia, aph(3'')-Ib, aph(6)-Id) and ant(3'')-Ia).
Two ESBL-EC isolates exhibited the aac(6)-Ib-cr gene responsible for the reduction in ciprofloxacin activity. The ant(3'')-Ia gene encodes an aminoglycoside nucleotidyltransferase enzyme that inactivates certain aminoglycoside antibiotics, was detected in eight ESBL-EC isolates from the poultry farm. This was followed by the detection of folate pathway antagonist resistance genes, represented by five variants (sul1, sul2, dfrA1, dfrA14, and dfrA17). In addition, quinolone resistance genes—often linked to chromosomal mutations and classified by the World Health Organization as critically important antimicrobials—were identified in seven variants (aac(6')-Ib-cr, qnrB1, qnrB17, qnrB19, qnrS1, oqxA and oqxB). Other AMR determinants detected in E. coli isolates were phenicol resistance (cat1, catB3, catB7, floR), fosfomycin resistance (fosA, fosA7), and tetracycline resistance (tetA, tetB, tetJ).
3.6. Plasmid Replicons Detected in E. coli Isolates
Thirty-eight different replicon types were observed among 55 E. coli isolates from all sources (Figure 4a). The most prevalent replicon types were IncFIB(AP001918) (n = 39, 70.9%), IncFIA (n = 18, 32.7%), IncFIC(FII) (n = 18, 32.7%), ColRNAI (n = 18, 32.7%), IncI-1 (n = 16, 29.1%), IncFII (n = 15, 27.3%) and Col440I (n = 12, 21.8%). The IncFIB(AP001918) were prevalent in isolates from cattle (n=20) and poultry (n=17) farms (Figure 4a). Six replicon types including IncFIB(AP001918), ColRNAI, IncFIA, IncFII, IncFIB(K) and IncX1 were common to all isolate sources.
3.7. Plasmid-Mediated Antimicrobial Resistance
Plasmid analysis revealed that ESBL-EC isolates harbored diverse plasmid replicon types carrying clinically relevant AMR genes (Figure 4b-f). In isolate 5T1ECE recovered from WWTP-A, two plasmids were identified: an IncQ1 plasmid carrying aph(6)-Id, aph(3ʺ)-Ib, and sul2 genes on contig 63, and a Col440I plasmid harboring the quinolone resistance gene qnrB19 on contig 68. The cattle-associated isolate 20T1ECE carried an IncFIC(FII) plasmid encoding the ESBL gene blaCTX-M-32, along with plasmid transfer genes (traJ, traT), virulence-associated genes (anr), and insertion sequences ISKpn26 and IS26. In the chicken-derived isolate 40T1ECE, an IncI1 plasmid was detected carrying tetA and blaCTX-M-1, together with insertion sequences ISSbo1 and ISEc9. Additionally, isolate RFIG1ECE from WWTP-B contained an IncQ1 plasmid encoding aph(6)-Id, aph(3ʺ)-Ib, and sul2.
3.8. Multi-Locus Sequence Typing of E. coli Isolates
MLST analysis revealed that 55 E. coli isolates were distributed across 31 known sequence types (STs). Four isolates could not be assigned to any known ST, and notably, ST 348 was shared between isolates (n=2) from the cattle farm and poultry farm. The most prevalent sequence types were ST602 (n=9) and ST2063 (n=3), detected in isolates from poultry farm samples. Other sequence types included ST119 (n=2), ST278 (n=2), ST645 (n=2), ST1704 (n=2), ST2073 (n=2), and ST2387 (n=2), all originating from cattle farm isolates. Additional sequence types detected included ST636 (n=2), while several others were observed as singletons, particularly among wastewater isolates (Supplementary data 1).
3.9. Phylogenetic Analysis of E. coli and ESBL-EC Isolates
Overall, 55 isolates were included in a single-nucleotide polymorphism (SNP)–based maximum-likelihood phylogenetic analysis (Figure 5). The E. coli and ESBL-EC isolates from wastewater and farm environments exhibited substantial genetic diversity, with limited evidence of genetic relatedness. Clonal relationships were observed only among isolates originating from the same or closely related sources. Analysis of the core-genome SNP matrix from 55 E. coli genomes identified a single clonal pair with a pairwise SNP distance of zero (Figure 5). Phylogenetic clustering was consistent with MLST, with clonally related isolates grouping by sequence type. Two isolates classified as sequence type ST348, exhibiting a pairwise SNP distance of zero, were identified in both cattle and poultry farm environments, suggesting possible cross-reservoir transmission.
4. Discussion
The emergence and dissemination of ESBL-EC pose a critical challenge to both human and animal health, driven by the interplay between antimicrobial use in livestock, environmental contamination, and bacterial adaptation [1,2,3,40]. The findings of this study, demonstrating the presence of ESBL-EC across diverse farm and wastewater sources, highlight the complexity of AMR ecology and the role of multiple reservoirs in maintaining and spreading resistant strains. The substantial genetic and resistome diversity observed underscores that resistance is not confined to a single lineage but rather distributed across multiple independent lineages, emphasizing the need for integrated, One Health–oriented approaches to surveillance, prevention, and control [14,15,25,41,42].
The detection of ESBL-EC in approximately one in four samples from this present study underscores the substantial presence of AMR bacteria across both animal and environmental reservoirs. The notably high prevalence of ESBL-EC in poultry samples, exceeding fifty percent, aligns with previous reports identifying backyard poultry production as a hotspot for the emergence and spread of these resistant bacteria, likely driven by the use of antibiotics in feed and therapeutic treatments [43,44,45]. Our study reports a relatively low prevalence of ESBL-EC in beef cattle, which contrasts with the substantially higher prevalences reported in non-organic dairy calves in the Netherlands (32.9%), beef cattle in Nigeria (45.4%), and dairy cattle in East Tennessee (47.5%) [3,46,47].
The prevalences observed in this study are comparable to those reported from other urban wastewater systems in North Carolina, including Chapel Hill (12.3%) and Raleigh (18.7%) suggesting that the occurrence of ESBL-EC in treated wastewater is not uncommon [30]. These findings highlight the capacity of WWTPs to receive, concentrate, and potentially disseminate resistant bacteria originating from human, animal, and environmental sources [9,48,49]. One wastewater sampling event was conducted shortly after a storm, a factor that may have influenced the detection of ESBL-EC in effluent despite the absence of direct animal inputs to the treatment plants. Extreme weather events, including heavy rainfall and flooding, can enhance the mobilization and redistribution of AMR bacteria from wastewater and other environmental reservoirs into surface waters, increasing opportunities for human exposure and highlighting the importance of incorporating environmental context into AMR surveillance efforts [54,55]. The persistence of ESBL-EC in these systems raises concerns about downstream environmental release and exposure risks, emphasizing the need for continued wastewater surveillance to monitor the spread of AMR [41,49,50].
Notably, more than half of the ESBL-EC isolates originated from animal feces, reinforcing the role of livestock as a primary source of AMR in the community [3,30,46,47]. The presence of ESBL-EC in WWTP samples further suggests that wastewater may act as a conduit for resistant bacteria, facilitating their spread into natural water systems and potentially impacting human health [41,42,49]. These findings emphasize the need for integrated surveillance and control strategies targeting both animal production systems and environmental sources to mitigate the dissemination of ESBL-EC [42,51].
The diversity and breadth of AMR determinants identified in these E. coli isolates reflect the complex resistome circulating within farm and wastewater environments. The widespread presence of mdf(A), which contributes to multidrug efflux, highlights the potential for co-selection of resistance even in the absence of direct antimicrobial pressure [49,52]. The variety of β-lactamase genes, particularly the blaCTX-M variants, underscores the ongoing circulation of classical ESBL determinants, while the detection of AmpC-type blaCMY genes suggests additional mechanisms contributing to β-lactam resistance [42,46].
The detection of blaSHV and blaCMY-2 in treated wastewater effluent indicates a potential microbiological hazard for environmental dissemination and human exposure, particularly where treated effluent or sludge is applied to agricultural land, underscoring the need for targeted risk-assessment studies [53]. The prevalence of aminoglycoside, folate pathway antagonist, and quinolone resistance genes—including those linked to critically important antibiotics—signals a high potential for therapeutic challenges and limited treatment options [51,56,57]. Notably, the presence of aac(6')-Ib-cr and qnr variants indicates the co-occurrence of resistance to multiple drug classes, enhancing the risk of cross-resistance [14,56,58]. The identification of resistance determinants to phenicols, fosfomycin, and tetracyclines further illustrates the extensive resistome diversity and the potential for horizontal gene transfer [59,60,61,62]. No carbapenem-resistance genes were detected in this study.
Plasmid replicon analysis revealed a predominance of IncF-family plasmids, particularly IncFIB(AP001918), which was the most prevalent replicon type and was mainly detected in isolates from cattle and poultry farms. IncF plasmids are well recognized for their role in disseminating ESBL genes and other AMR determinants among Enterobacterales, especially within animal production systems [63,64]. The frequent detection of IncFIA, IncFIC(FII), and IncFII further highlights the importance of this plasmid backbone in maintaining and spreading resistance traits across diverse ecological niches [63,65,66]. The presence of IncI-1 and Col440I plasmids, which are commonly associated with food-producing animals and environmental reservoirs, underscores their contribution to resistance gene mobility [3,67]. Notably, six replicon types were shared across all sampling sources, suggesting widespread circulation of common plasmid platforms at the human–animal–environment interface [63].
The detection of diverse plasmid incompatibility groups carrying AMR genes across ESBL-EC isolates underscores the critical role of plasmids in mediating resistance dissemination at the human–animal–environment interface. The presence of small, mobilizable plasmids such as IncQ1 in both wastewater and agricultural isolates is particularly concerning, given their broad host range and ability to facilitate rapid horizontal transfer of resistance determinants, including aminoglycoside and sulfonamide resistance genes [63,68]. These findings suggest that WWTPs and livestock environments may act as interconnected reservoirs that promote persistence and amplification of plasmid-mediated resistance.
MLST revealed genetic diversity among the ESBL-EC isolates, distributed across 31 distinct sequence types, indicating the circulation of multiple lineages across environmental and agricultural reservoirs [3,64,69]. The predominance of ST602 among poultry isolates highlights this lineage as an emerging clone of One Health concern. Reports of CTX-M–producing E. coli ST602 with a broad resistome in South American wild birds indicate its ability to persist across diverse hosts and ecosystems [70]. Its presence in poultry production systems raises concerns about amplification in intensive farming and dissemination via the food chain or environmental pathways, underscoring the need for coordinated genomic surveillance across animal, environmental, and wildlife reservoirs [70,71].
The observed genetic diversity among E. coli and ESBL-EC isolates underscores the complex and heterogeneous nature of these populations across wastewater and farm environments. The limited occurrence of clonal relationships suggests that the dissemination of ESBL-EC is driven more by diverse, independent lineages rather than widespread clonal expansion [53,72]. However, the presence of ST348 in both cattle and poultry isolates suggests cross-reservoir transmission or shared ecological drivers. Similar cross-host circulation of MDR ST348 has been reported in horses and humans, underscoring its capacity to disseminate across species and highlighting the need for One Health surveillance [73]. The presence of numerous singleton STs, particularly among wastewater isolates, further highlights the high genetic heterogeneity of E. coli circulating in municipal wastewater systems, which integrate inputs from human, animal, and environmental sources [42,49].
These findings highlight plasmids as critical drivers of ESBL persistence and cross-reservoir dissemination across agricultural and wastewater environments. The diversity of plasmid backbones underscores the complexity of resistance transmission and reinforces the need for integrated One Health surveillance and stewardship to limit the spread of MDR E. coli.
This study has several limitations. The number of farms and WWTPs included was limited, which restricts the generalizability of our findings to broader settings. Additionally, due to funding constraints, only a subset of isolates underwent whole-genome sequencing, limiting the resolution of our genetic and resistome analyses across the full collection. Despite these constraints, the study provides valuable insights into the prevalence, diversity, and genomic characteristics of ESBL-EC in farm and environmental reservoirs, offering a foundation for future, more comprehensive investigations.
5. Conclusions
This study demonstrates the widespread presence and genetic diversity of ESBL-EC across animal and environmental reservoirs, highlighting the interconnectedness of farms and wastewater systems in the dissemination of AMR. The co-occurrence of multiple resistance determinants and evidence of cross-reservoir transmission underscore the complexity of AMR ecology and the urgent need to address these reservoirs collectively. These findings contribute to a deeper understanding of the drivers of ESBL-EC dissemination and provide a foundation for informed surveillance and intervention strategies. To mitigate the spread of ESBL-EC, integrated surveillance across livestock, wastewater, and environmental sources should be prioritized, adopting a One Health framework. Future studies should expand sampling to include a broader range of farms and treatment plants to improve representativeness and generalizability. Comprehensive WGS of larger isolate collections will enhance understanding of resistance gene dissemination, clonal relationships, and potential transmission pathways. Additionally, implementing targeted antimicrobial stewardship programs in animal production and improving wastewater management practices could reduce the emergence and environmental release of MDR bacteria.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Table S1. ESBL-EC strains used for AMR analysis and MLST; Supplementary Table S2. ESBL-EC strains used for Plasmid analysis.
Author Contributions
I.T conceptualized the research idea. J.D and I.T were the principal investigators, collected data, isolated the organism, performed antibiotic sensitivity testing and wrote the original draft. D.J, S.W, D.S, K.E performed PCR and gel electrophoresis. Y.W and G.D extracted DNA for genomic sequencing and contributed to the original draft. L.H generated the whole genome sequence profile for all the isolates and contributed to the original draft. R.L facilitated access for sample collection. M.K.A. and M.S analyzed the data, interpreted the data, performed bioinformatics analysis and wrote the second draft of the manuscript. R.L, M.S and M.K.A revised the article critically including important intellectual content. All authors read and approved the final manuscript.
Funding
This work was supported by the Department of Biological and Forensic Sciences, Fayetteville State University, North Carolina, USA through the New Faculty Seed Grant program.
Acknowledgments
The authors sincerely thank the management teams of the wastewater treatment plants, as well as the poultry and cattle farm managers, for granting access and facilitating sample collection. We are grateful to the Department of Biological and Forensic Sciences, Fayetteville State University, North Carolina for providing the seed grant that supported this research. We also extend our appreciation to the members of the Thakur Molecular Laboratory at North Carolina State University for their technical assistance and valuable contributions to the success of this study.
Conflicts of Interest
The authors declare no competing interests.
References
- Braz, S; Melchior, K; Moreira, CG; Farfan, MJ; Toro, CS. Escherichia coli as a Multifaceted Pathogenic and Versatile Bacterium. 2020, 1–9. [Google Scholar] [CrossRef]
- Ramos, S; Silva, V; de Lurdes Enes Dapkevicius, M; Caniça, M; Tejedor-Junco, MT; Igrejas, G; et al. Escherichia coli as commensal and pathogenic bacteria among food-producing animals: Health implications of extended spectrum β-lactamase (ESBL) production. Animals 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Aworh, MK; Ekeng, E; Nilsson, P; Egyir, B; Owusu-Nyantakyi, C; Hendriksen, RS. Extended-Spectrum ß-Lactamase-Producing Escherichia coli Among Humans, Beef Cattle, and Abattoir Environments in Nigeria. Front Cell Infect Microbiol 2022, 12, 1–11. [Google Scholar] [CrossRef]
- Pakbin, B; Allahyari, S; Amani, Z; Brück, WM; Mahmoudi, R. Prevalence, Phylogroups and Antimicrobial Susceptibility of Escherichia coli Isolates from Food Products. Antibiotics 2021, 10, 1–11. [Google Scholar] [CrossRef]
- Yousefipour, M; Rezatofighi, SE; Ardakani, MR. Detection and characterization of hybrid uropathogenic Escherichia coli strains among E. coli isolates causing community-acquired urinary tract infection. J Med Microbiol. 2023, 72, 1–11. [Google Scholar] [CrossRef]
- Zaghloul, A; Saber, M; Gadow, S; Awad, F. Biological indicators for pollution detection in terrestrial and aquatic ecosystems. Bull Natl Res Cent. 2020, 44, 1–11. [Google Scholar] [CrossRef]
- Paruch, L; Paruch, AM. An Overview of Microbial Source Tracking Using Host-Specific Genetic Markers to Identify Origins of Fecal Contamination in Different Water Environments. Water 2022, 14, 1–18. [Google Scholar] [CrossRef]
- Kusi, J; Ojewole, CO; Ojewole, AE. Antimicrobial Resistance Development Pathways in Surface Waters and Public Health Implications. Antibiotics 2022, 11, 1–22. [Google Scholar] [CrossRef]
- Yu, D; Ryu, K; Zhi, S; Otto, SJG; Neumann, NF. Naturalized Escherichia coli in Wastewater and the Co-evolution of Bacterial Resistance to Water Treatment and Antibiotics. Front Microbiol 2022. [Google Scholar] [CrossRef]
- Graham, JP; Amato, HK; Mendizabal-Cabrera, R; Alvarez, D; Ramay, BM. Waterborne urinary tract infections: Have we overlooked an important source of exposure? Am J Trop Med Hyg. 2021, 105, 12–7. [Google Scholar] [CrossRef]
- Anjum, MF; Schmitt, H; Bo, S; Stehling, EG; Boerlin, P; Topp, E; et al. The potential of using E. coli as an indicator for the surveillance of antimicrobial resistance (AMR) in the environment. Curr Opin Microbiol. 2021, 62, 152–8. [Google Scholar]
- Mkilima, T. Wastewater as a Sentinel for Emerging Antimicrobial Resistance Threats: A Review. Environ Qual Manag. 2025, 35, 1–19. [Google Scholar]
- Gregova, G; Kmet, V; Szaboova, T. New Insight on Antibiotic Resistance and Virulence of Escherichia coli from Municipal and Animal Wastewater. Antibiotics 2021, 10, 1–11. [Google Scholar] [CrossRef]
- Aworh, MK; Kwaga, J; Hendriksen, R; Okolocha, E; Thakur, S. Genetic relatedness of multidrug resistant Escherichia coli isolated from humans, chickens and poultry environments. Antimicrob Resist Infect Control 2021, 10, 1–13. [Google Scholar]
- Fatokun, O; Selvaraja, M; Anuar, H; Zetty, T; Tengku, M. Antimicrobial resistance at the human – animal – environment interface: A focus on antimicrobial-resistant Escherichia coli transmission dynamics, clinical implications, and future directions. Int J One Heal 2024, 10, 161–71. [Google Scholar]
- Addae-nuku, DS; Kotey, FCN; Dayie, NTKD; Osei, M; Tette, EMA; Debrah, P; et al. Multidrug-Resistant Bacteria in Hospital Wastewater of the Korle Bu Teaching Hospital in Accra, Ghana. Environ Health Insights 2022, 16, 1–9. [Google Scholar]
- Husna, A; Rahman, MM; Badruzzaman, ATM; Sikder, MH; Islam, MR; Rahman, MT; et al. Extended-Spectrum β-Lactamases (ESBL): Challenges and Opportunities. Biomedicines 2023, 11, 1–22. [Google Scholar]
- Mandujano-hern, A; Ver, A; Paz-gonz, AD; Herrera-mayorga, V; Mario, S; Lara-ram, EE; et al. The Global Rise of ESBL-Producing Escherichia coli in the Livestock Sector: A Five-Year Overview. Animals 2024, 14, 1–23. [Google Scholar]
- World Health Organization (WHO). Global antibiotic resistance surveillance report 2025. 2025. [Google Scholar]
- Duffy, N; Karlsson, M; Reses, HE; Daniels, J; Stanton, RA; Janelle, SJ; et al. Epidemiology of extended-spectrum β-lactamase–producing Enterobacterales in five US sites participating in the Emerging Infections Program, 2017. Infect Control Hosp Epidemiol 2022, 43, 1586–94. [Google Scholar] [CrossRef]
- Cheney, C; Johnson, JD; Marie, JPS; Gacosta, KYM; Drumm, NBD; Jones, GD. coli and metagenomic analysis of source wastewater samples. Microbiol Spectr 2024, 12. [Google Scholar]
- Tiwari, A; Kurittu, P; Al-Mustapha, AI; Heljanko, V; Johansson, V; Thakali, O; et al. Wastewater surveillance of antibiotic-resistant bacterial pathogens: A systematic review. Front Microbiol 2022, 1–19. [Google Scholar] [CrossRef]
- Aworh, MK; Thakur, S; Gensler, C; Harrell, E; Harden, L; Fedorka-Cray, PJ; et al. Characteristics of antimicrobial resistance in Escherichia coli isolated from retail meat products in North Carolina. PLoS One 2024, 19, e0294099. [Google Scholar] [CrossRef]
- Xin, H; Gao, M; Wang, X; Qiu, T; Guo, Y; Zhang, L. Animal farms are hot spots for airborne antimicrobial resistance. Sci Total Environ 2022, 158050. [Google Scholar] [CrossRef]
- La Rosa, MC; Maugeri, A; Favara, G; La Mastra, C; Magnano San Lio, R; Barchitta, M; et al. The Impact of Wastewater on Antimicrobial Resistance: A Scoping Review of Transmission Pathways and Contributing Factors. Antibiotics 2025, 14, 1–29. [Google Scholar] [CrossRef]
- Zhang, T; Nickerson, R; Zhang, W; Peng, X; Shang, Y; Zhou, Y; et al. The impacts of animal agriculture on One Health—Bacterial zoonosis, antimicrobial resistance, and beyond. One Heal 2024, 18, 100748. [Google Scholar] [CrossRef]
- Al-khalaifah, H; Rahman, MH. A One-Health Perspective of Antimicrobial Resistance ( AMR ): Human, Animals and Environmental Health. Life 2025, 15, 1–24. [Google Scholar] [CrossRef]
- Patra, M; Dubey, SK. Antimicrobial Resistance Transmission in Environmental Matrices: Current Prospects and Future Directions. Rev Environ Contam Toxicol. 2025, 263, 1–56. [Google Scholar] [CrossRef]
- Djordjevic, SP; Jarocki, VM; Seemann, T; Cummins, ML; Watt, AE; Drigo, B; et al. Genomic surveillance for antimicrobial resistance — a One Health perspective. Nat Rev Genet. 2024, 25, 142–57. [Google Scholar] [CrossRef]
- Appling, KC; Sobsey, MD; Durso, LM; Fisher, MB. Environmental monitoring of antimicrobial resistant bacteria in North Carolina water and wastewater using the WHO Tricycle protocol in combination with membrane filtration and compartment bag test methods for detecting and quantifying ESBL E. coli. PLOS Water 2023, 2, e0000117. [Google Scholar] [CrossRef]
- Skandalis, N; Papafotis, D; Miller, S; Lee, B; Theologidis, I; Luna, B. Environmental Spread of Antibiotic Resistance. Antibiotics 2021, 10, 1–14. [Google Scholar] [CrossRef]
- Zhao, D; Paco, J; Wojnarowski, K; Cholewi, P. Antibiotic Resistance Genes in Food Animal Production: Environmental Implications and One Health Challenges. Environments 2025, 12, 1–25. [Google Scholar] [CrossRef]
- CLSI. M100, Performance Standards for Antimicrobial Susceptibility Testing. 2022.
- Ogutu, JO; Zhang, Q; Huang, Y; Yan, H; Su, L; Gao, B; et al. Development of a multiplex PCR system and its application in detection of blaSHV, blaTEM, blaCTX-M-1, blaCTX-M-9 and blaOXA-1 group genes in clinical Klebsiella pneumoniae and Escherichia coli strains. J Antibiot (Tokyo) 2015, 68, 725–33. [Google Scholar] [CrossRef]
- Bankevich, A; Nurk, S; Antipov, D; Gurevich, AA; Dvorkin, M; Kulikov, AS; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 2012, 19, 455–77. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–9. [Google Scholar] [CrossRef]
- Kozlov, AM; Darriba, D; Flouri, T; Morel, B; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–5. [Google Scholar] [CrossRef]
- CLSI. M100, Performance Standards for Antimicrobial Susceptibility Testing. 2022.
- Letunic, I; Bork, P. Interactive tree of life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–6. [Google Scholar] [CrossRef]
- Aworh, MK; Kwaga, J; Okolocha, E; Harden, L; Hull, D; Hendriksen, RS; et al. Extended-spectrum ß-lactamase-producing Escherichia coli among humans, chickens and poultry environments in Abuja, Nigeria. One Heal Outlook 2020, 2, 8. [Google Scholar] [CrossRef]
- Martak, D; Henriot, CP; Hocquet, D. Environment, animals, and food as reservoirs of antibiotic-resistant bacteria for humans: One health or more? Infect Dis Now 2024, 54, 104895. [Google Scholar] [CrossRef]
- Milenkov, M; Proux, C; Rasolofoarison, TL; Rakotomalala, FA; Rasoanandrasana, S; Rahajamanana, VL; et al. Implementation of the WHO Tricycle protocol for surveillance of extended-spectrum β-lactamase producing Escherichia coli in humans, chickens, and the environment in Madagascar: a prospective genomic epidemiology study. The Lancet Microbe 2024, 5, 100850. [Google Scholar] [CrossRef]
- Saeed, MA; Saqlain, M; Waheed, U; Ehtisham-ul-Haque, S; Khan, AU; Rehman A ur; et al. Cross-Sectional Study for Detection and Risk Factor Analysis of ESBL-Producing Avian Pathogenic Escherichia coli Associated with Backyard Chickens in Pakistan. Antibiotics 2023, 12, 1–15. [Google Scholar] [CrossRef]
- Aliyu, AB; Jalila, A; Saleha, AA; Zunita, Z. ESBL Producing E. coli in Chickens and Poultry Farms Environment in Selangor, Malaysia: A Cross-Sectional Study on Their Occurrence and Associated Risk Factors With Environment and Public Health Importance. Zoonoses Public Health 2024, 71, 962–71. [Google Scholar] [CrossRef]
- Widodo, A; Khairullah, AR; Effendi, MH; Moses, IB; Agustin, ALD. Extended-spectrum β-lactamase-producing Escherichia coli from poultry: A review. Vet World 2024, 17, 2017–27. [Google Scholar] [CrossRef]
- Gonggrijp, MA; Velthuis, AGJ; Heuvelink, AE; van den Heuvel, KWH; ter Bogt-Kappert, CC; Buter, GJ; et al. Prevalence of extended-spectrum and AmpC β-lactamase-producing Escherichia coli in young calves on Dutch dairy farms. J Dairy Sci. 2023, 106, 4257–65. [Google Scholar] [CrossRef]
- Gelalcha, BD; Gelgie, AE; Kerro Dego, O. Prevalence and antimicrobial resistance profiles of extended-spectrum beta-lactamase-producing Escherichia coli in East Tennessee dairy farms. Front Vet Sci 2023, 1–14. [Google Scholar] [CrossRef]
- Aguilar-salazar, A; Martínez-Vázquez, A; Aguilera-arreola, G; De Luna-santillana, EDJ; Escobedo-bonilla, CM; Lara-ram, E; et al. Prevalence of ESKAPE Bacteria in Surface Water and Wastewater Sources: Multidrug Resistance and Molecular Characterization, an Updated Review. Water 2023, 15, 1–23. [Google Scholar] [CrossRef]
- Marutescu, LG; Popa, M; Gheorghe-Barbu, I; Barbu, IC; Rodríguez-Molina, D; Berglund, F; et al. Wastewater treatment plants, an “escape gate” for ESCAPE pathogens. Front Microbiol 2023, 1–15. [Google Scholar] [CrossRef]
- Dawangpa, A; Lertwatcharasarakul, P; Ramasoota, P; Boonsoongnern, A; Ratanavanichrojn, N; Sanguankiat, A; et al. Genotypic and phenotypic situation of antimicrobial drug resistance of Escherichia coli in water and manure between biogas and non-biogas swine farms in central Thailand. J Environ Manage 2021;279 July 2020, 111659. [Google Scholar] [CrossRef]
- Furlan, JPR; Stehling, EG. Multiple sequence types, virulence determinants and antimicrobial resistance genes in multidrug- and colistin-resistant Escherichia coli from agricultural and non-agricultural soils. Environ Pollut 2021. [Google Scholar] [CrossRef]
- Nolan, TM; Martin, NA; Reynolds, LJ; Sala-Comorera, L; O’Hare, GMP; O’Sullivan, JJ; et al. Agricultural and urban practices are correlated to changes in the resistome of riverine systems. Sci Total Environ 2024, 172261. [Google Scholar] [CrossRef]
- Pino-Hurtado, MS; Fernández-Fernández, R; Rubio-Tomás, L; Marañón-Clemente, I; Álvarez-Gómez, T; García-Mora, DJ; et al. Characterization of Carbapenem-Resistant and ESBL-Producing Enterobacterales in Wastewater and Sludge Environments from Northern Spain. Appl Sci. 2025, 15, 1–18. [Google Scholar] [CrossRef]
- Dickin, SK; Schuster-Wallace, CJ; Qadir, M; Pizzacalla, K. A review of health risks and pathways for exposure to wastewater Use in Agriculture. Environ Health Perspect. 2016, 124, 900–9. [Google Scholar] [CrossRef]
- Aziz, F; Wang, X; Mahmood, MQ; Juliette, OK. Assessing human health risks associated with wastewater flooding. Environ Impact Assess Rev. 2025, 108031. [Google Scholar] [CrossRef]
- Aworh, MK; Kwaga, JKP; Hendriksen, RS; Okolocha, EC; Harrell, E; Thakur, S. Quinolone-resistant Escherichia coli at the interface between humans, poultry and their shared environment- a potential public health risk. One Heal Outlook 2023, 5. [Google Scholar]
- WHO. WHO | WHO list of Critically Important Antimicrobials (CIA). 2019. [Google Scholar]
- Kanokudom, S; Assawakongkarat, T; Akeda, Y; Ratthawongjirakul, P; Chuanchuen, R; Chaichanawongsaroj, Id N. Rapid detection of extended spectrum β-lactamase producing Escherichia coli isolated from fresh pork meat and pig cecum samples using multiplex recombinase polymerase amplification and lateral flow strip analysis. 2021. [Google Scholar] [CrossRef]
- Abdelrahim, A; Harrell, E; Fedorka-Cray, PJ; Jacob, M; Thakur, S. Phenotypic and Genotypic Characterizations of Antimicrobial-Resistant Escherichia coli Isolates from Diverse Retail Meat Samples in North Carolina During 2018-2019. Foodborne Pathog Dis. 2024, 21, 211–9. [Google Scholar] [CrossRef]
- Surleac, M; Barbu, IC; Paraschiv, S; Popa, LI; Gheorghe, I; Marutescu, L; et al. Whole genome sequencing snapshot of multidrug resistant Klebsiella pneumoniae strains from hospitals and receiving wastewater treatment plants in Southern Romania. PLoS One 2020, 15, 1–17. [Google Scholar]
- Abdallah, K; Tayh, G; Maamar, E; Mosbah, A; Abbes, O; Fliss, I; et al. Genotypic Characterisation and Risk Assessment of Virulent ESBL-Producing E. coli in Chicken Meat in Tunisia: Insights from Multi-Omics Machine Learning Perspective. Microbiol Res (Pavia) 2025, 16, 1–25. [Google Scholar] [CrossRef]
- Hossain, AKMZ; Chowdhury, AMMA. Understanding the Evolution and Transmission Dynamics of Antibiotic Resistance Genes: A Comprehensive Review. J Basic Microbiol 2024, 64, 1–7. [Google Scholar] [CrossRef]
- Zhang, S; Yang, J; Abbas, M; Yang, Q; Li, Q; Liu, M; et al. Threats across boundaries: the spread of ESBL-positive Enterobacteriaceae bacteria and its challenge to the “one health” concept. Front Microbiol 2025. [Google Scholar] [CrossRef]
- Martínez-Álvarez, SA; Asencio-Egea, MÁ; Huertas-Vaquero, M; Cardona-Cabrera, T; Zarazaga, M; Höfle, U; et al. Genomic Epidemiology of ESBL- and Carbapenemase-Producing Enterobacterales in a Spanish Hospital: Exploring the Clinical–Environmental Interface. Microorganisms 2025, 13, 1–24. [Google Scholar] [CrossRef]
- Hirabayashi, A; Yahara, K; Mitsuhashi, S; Nakagawa, S; Imanishi, T; Ha, VTT; et al. Plasmid analysis of NDM metallo-β-lactamaseproducing Enterobacterales isolated in Vietnam. PLoS One 2021, 16 7 July, 1–16. [Google Scholar]
- Gelalcha, BD; Mohammed, RI; Gelgie, AE; Kerro Dego, O. Molecular epidemiology and pathogenomics of extended-spectrum beta-lactamase producing- Escherichia coli and - Klebsiella pneumoniae isolates from bulk tank milk in Tennessee, USA. Front Microbiol 2023, 1–14. [Google Scholar] [CrossRef]
- Juraschek, K; Malekzadah, J; Malorny, B; Käsbohrer, A; Schwarz, S; Meemken, D; et al. Characterization of qnrB-carrying plasmids from ESBL- and non-ESBL-producing Escherichia coli. BMC Genomics 2022, 23, 1–16. [Google Scholar] [CrossRef]
- Raufu, IA; Ahmed, OA; Aremu, A; Ameh, JA; Timme, RE; Hendriksen, RS; et al. Antimicrobial and Genomic Characterization of Salmonella Nigeria from Pigs and Poultry in Ilorin, North-central, Nigeria. J Infect Dev Ctries 2021, 15, 1899–909. [Google Scholar] [CrossRef]
- Levent, G; Schlochtermeier, A; Ives, SE; Norman, KN; Lawhon, SD; Loneragan, GH; et al. High-Resolution Genomic Comparisons within Salmonella enterica Serotypes Derived from Beef Feedlot Cattle: Parsing the Roles of Cattle Source, Pen, Animal, Sample Type, and Production Period. Appl Environ Microbiol 2021, 87, 1–20. [Google Scholar] [CrossRef]
- Dalazen, G; Fuentes-Castillo, D; Pedroso, LG; Fontana, H; Sano, E; Cardoso, B; et al. CTX-M-producing Escherichia coli ST602 carrying a wide resistome in South American wild birds: Another pandemic clone of One Health concern. One Heal 2023, 100586. [Google Scholar] [CrossRef]
- Borges, CA; Tarlton, NJ; Riley, LW. Escherichia coli from Commercial Broiler and Backyard Chickens Share Sequence Types, Antimicrobial Resistance Profiles, and Resistance Genes with Human Extraintestinal Pathogenic Escherichia coli. Foodborne Pathog Dis. 2019, 16, 813–22. [Google Scholar] [CrossRef]
- Ojer-Usoz, E; González, D; Vitas, AI. Clonal diversity of ESBL-producing Escherichia coli isolated from environmental, human and food samples. Int J Environ Res Public Health 2017, 14, 10–2. [Google Scholar] [CrossRef]
- Da Roza, FT; Couto, N; Carneiro, C; Cunha, E; Rosa, T; Magalhães, M; et al. Commonality of Multidrug-Resistant Klebsiella pneumoniae ST348 Isolates in Horses and Humans in Portugal. Front Microbiol 2019. [Google Scholar]
Figure 1.
Proportion of ESBL-EC among positive E. coli isolates from municipal wastewater treatment plants, cattle, poultry and farm environment. The proportion of E. coli positive isolates are plotted as bars on the primary axis while the proportion of ESBL-EC in percentages are plotted as a line graph on the secondary axis.
Figure 1.
Proportion of ESBL-EC among positive E. coli isolates from municipal wastewater treatment plants, cattle, poultry and farm environment. The proportion of E. coli positive isolates are plotted as bars on the primary axis while the proportion of ESBL-EC in percentages are plotted as a line graph on the secondary axis.
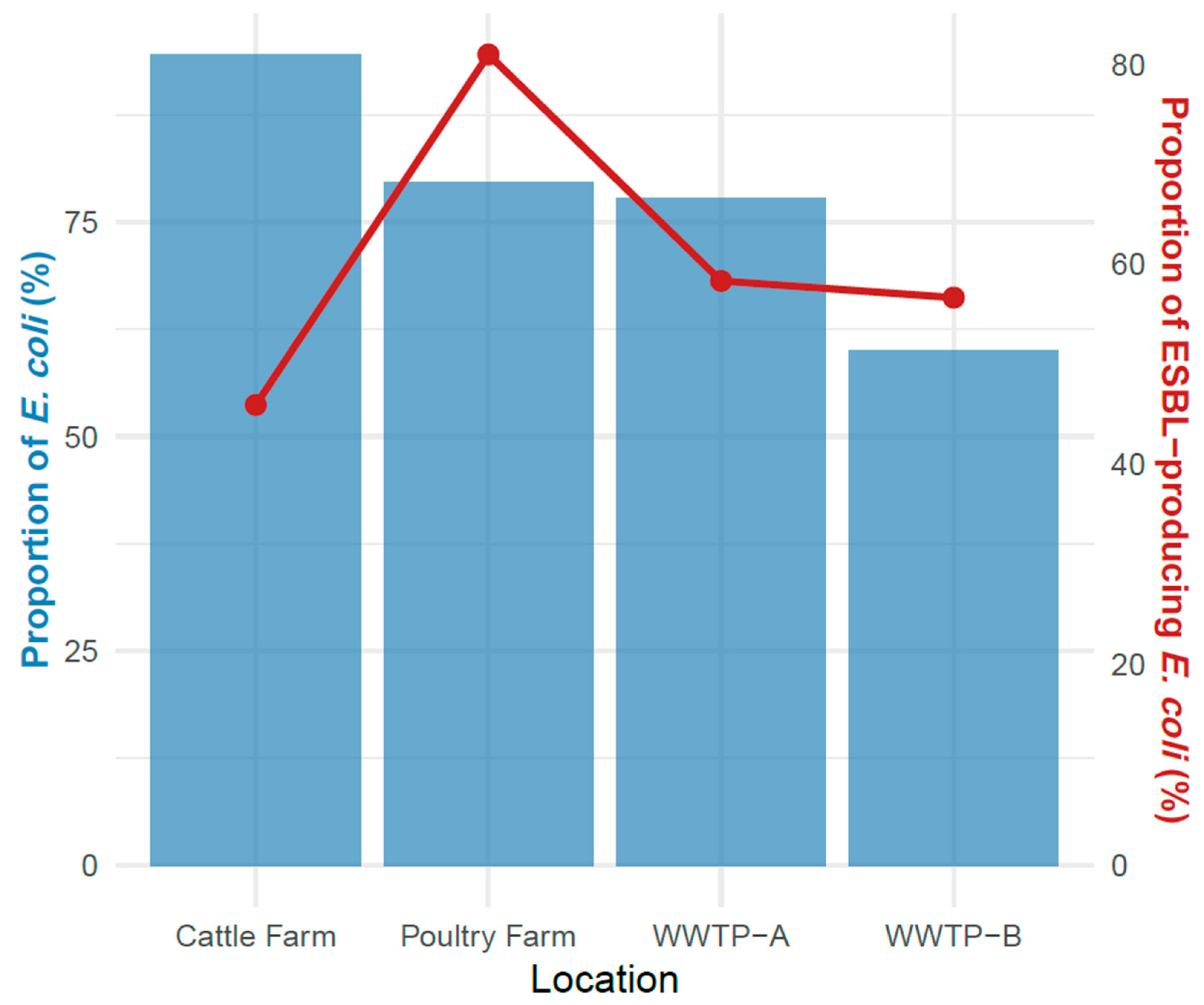
Figure 2.
Prevalence of MDR ESBL-EC isolated from farms and municipal wastewater samples, Fayetteville, NC, 2025. Bars represent the proportion of MDR ESBL-EC isolates from each isolation source.
Figure 2.
Prevalence of MDR ESBL-EC isolated from farms and municipal wastewater samples, Fayetteville, NC, 2025. Bars represent the proportion of MDR ESBL-EC isolates from each isolation source.
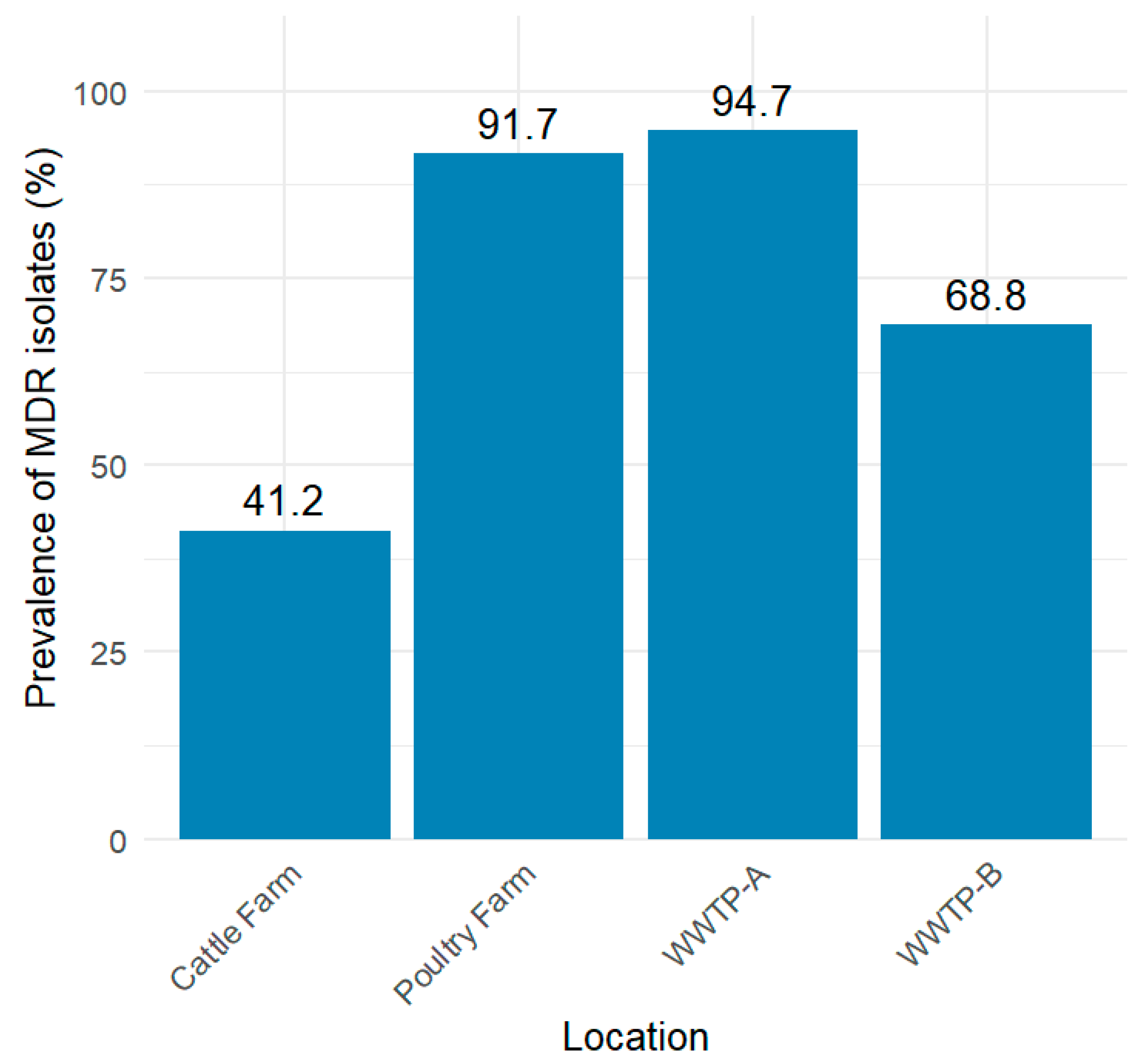
Figure 3.
a. Distribution of Resistance Genes Detected by PCR in ESBL-EC Isolates from Farms and Municipal Wastewater.
Figure 3.
a. Distribution of Resistance Genes Detected by PCR in ESBL-EC Isolates from Farms and Municipal Wastewater.
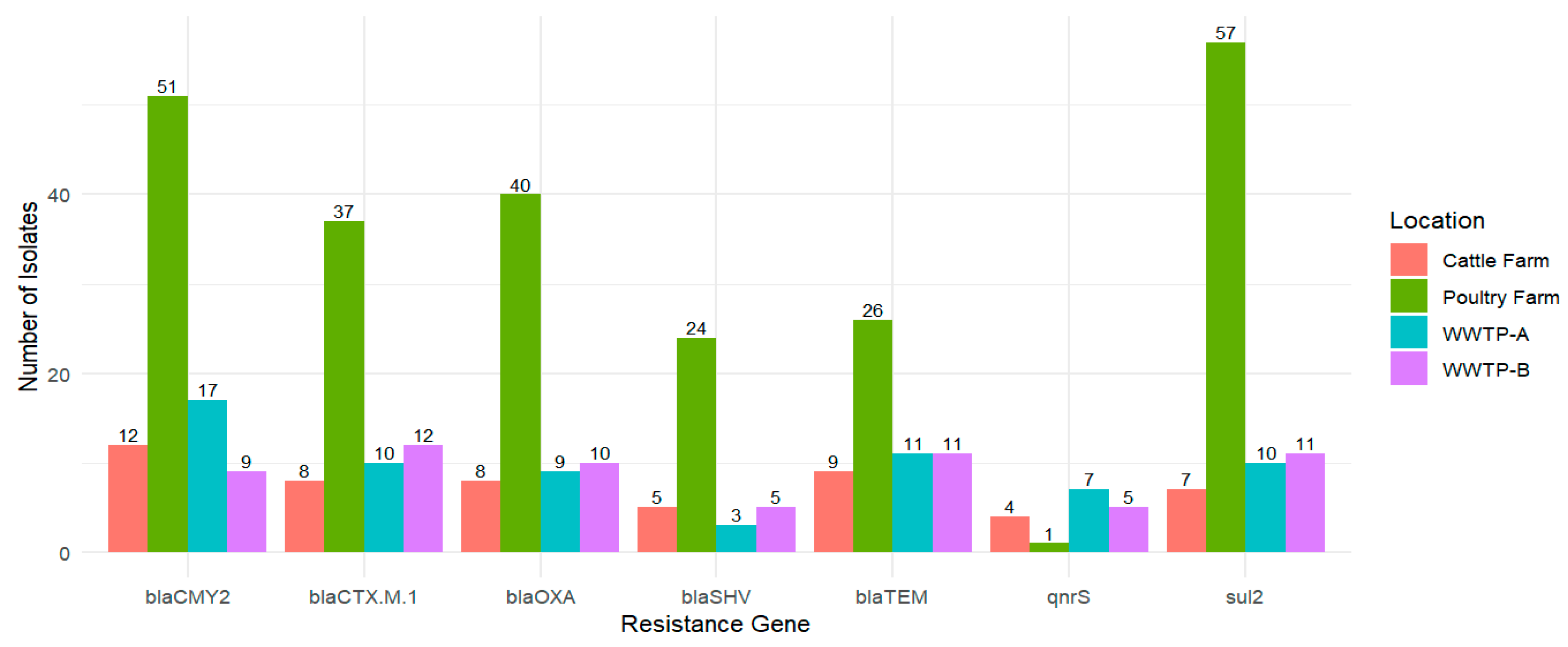
Figure 3.
b. Sample Type and Source-Specific Detection of Resistance Genes in ESBL-EC Isolates by PCR.
Figure 3.
b. Sample Type and Source-Specific Detection of Resistance Genes in ESBL-EC Isolates by PCR.
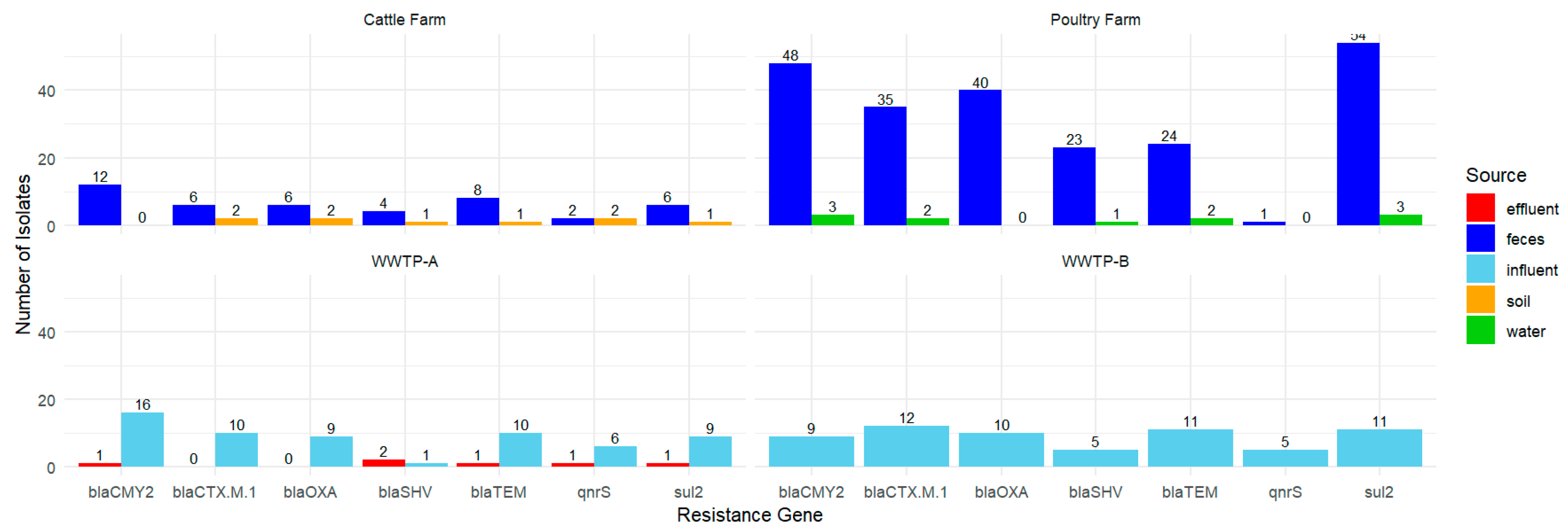
Figure 3.
c. Distribution of Resistance Genes Detected by WGS in ESBL-EC Isolates from Farms and Municipal Wastewater.
Figure 3.
c. Distribution of Resistance Genes Detected by WGS in ESBL-EC Isolates from Farms and Municipal Wastewater.
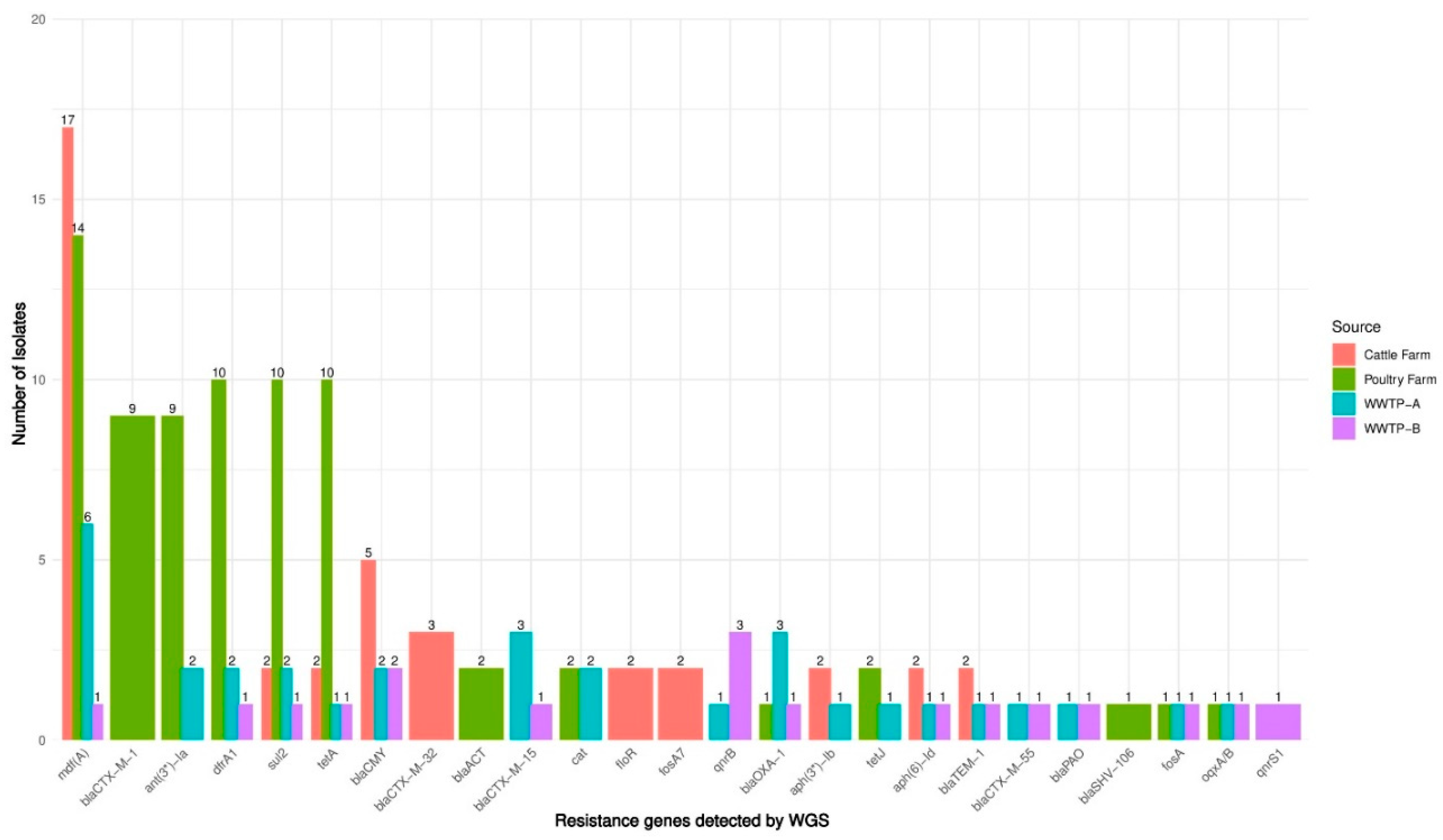
Figure 4.
a. Distribution of Resistance Genes Detected by WGS in ESBL-EC Isolates from Farms and Municipal Wastewater.
Figure 4.
a. Distribution of Resistance Genes Detected by WGS in ESBL-EC Isolates from Farms and Municipal Wastewater.
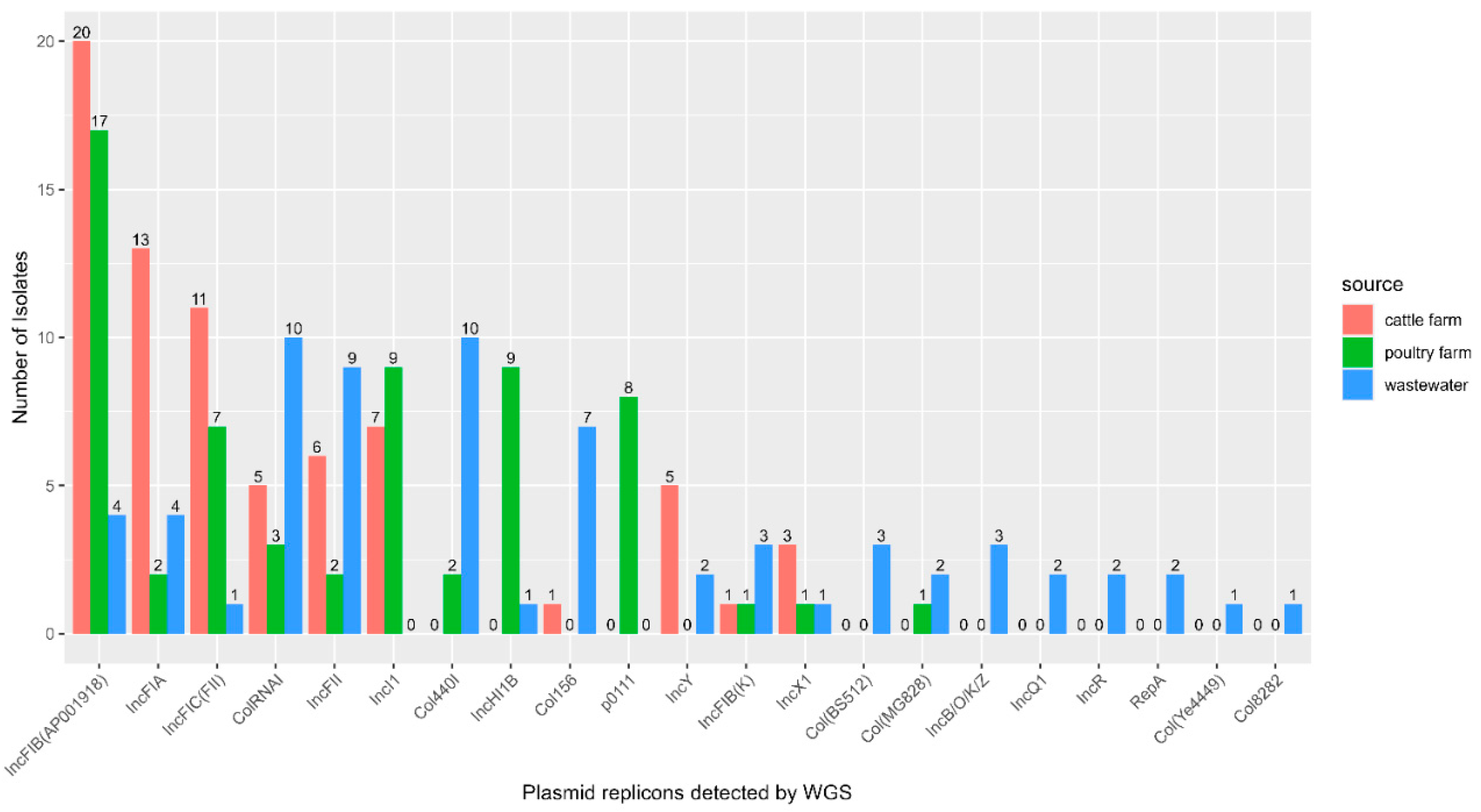
Figure 4.
b-f. Plasmid-mediated AMR determinants identified among ESBL-EC isolates from wastewater and farm environments. Isolate 5T1ECE from WWTP-A harbored an IncQ1 plasmid carrying aph(6)-Id, aph(3″)-Ib, and sul2 (green) on contig 63, as well as a Col440I plasmid carrying qnrB19 (green) on contig 68. Isolate 20T1ECE from cattle feces carried an IncFIC(FII) plasmid encoding blaCTX-M-32 (green), virulence-associated genes (traJ, traT, anr) in light blue color, and insertion sequences ISKpn26 and IS26 (yellow). Isolate 40T1ECE from chicken feces harbored an IncI1 plasmid carrying tetA and blaCTX-M-1 (green), along with insertion sequences ISSbo1 and ISEc9 (yellow). Isolate RFIG1ECE from WWTP-B contained an IncQ1 plasmid encoding aph(6)-Id, aph(3″)-Ib, and sul2 (green).
Figure 4.
b-f. Plasmid-mediated AMR determinants identified among ESBL-EC isolates from wastewater and farm environments. Isolate 5T1ECE from WWTP-A harbored an IncQ1 plasmid carrying aph(6)-Id, aph(3″)-Ib, and sul2 (green) on contig 63, as well as a Col440I plasmid carrying qnrB19 (green) on contig 68. Isolate 20T1ECE from cattle feces carried an IncFIC(FII) plasmid encoding blaCTX-M-32 (green), virulence-associated genes (traJ, traT, anr) in light blue color, and insertion sequences ISKpn26 and IS26 (yellow). Isolate 40T1ECE from chicken feces harbored an IncI1 plasmid carrying tetA and blaCTX-M-1 (green), along with insertion sequences ISSbo1 and ISEc9 (yellow). Isolate RFIG1ECE from WWTP-B contained an IncQ1 plasmid encoding aph(6)-Id, aph(3″)-Ib, and sul2 (green).
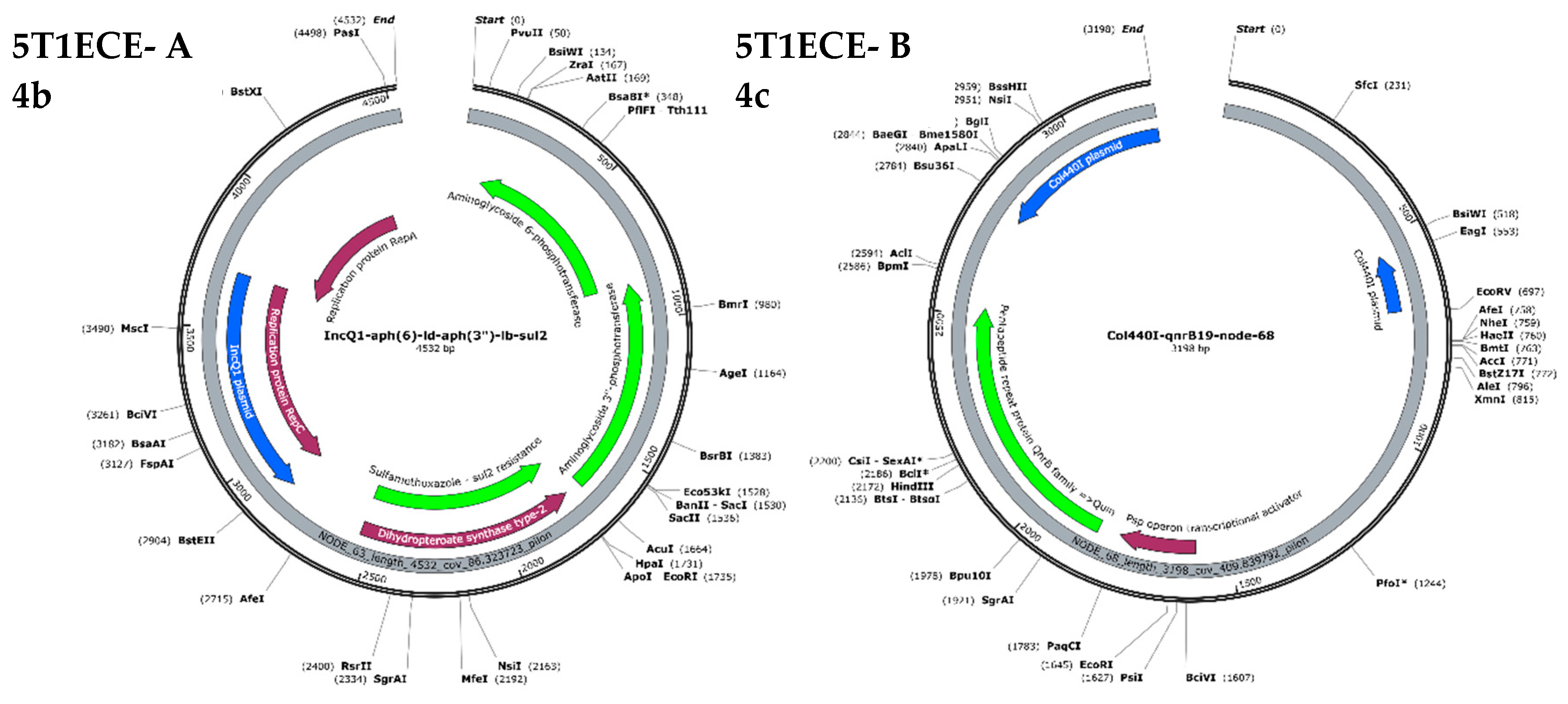
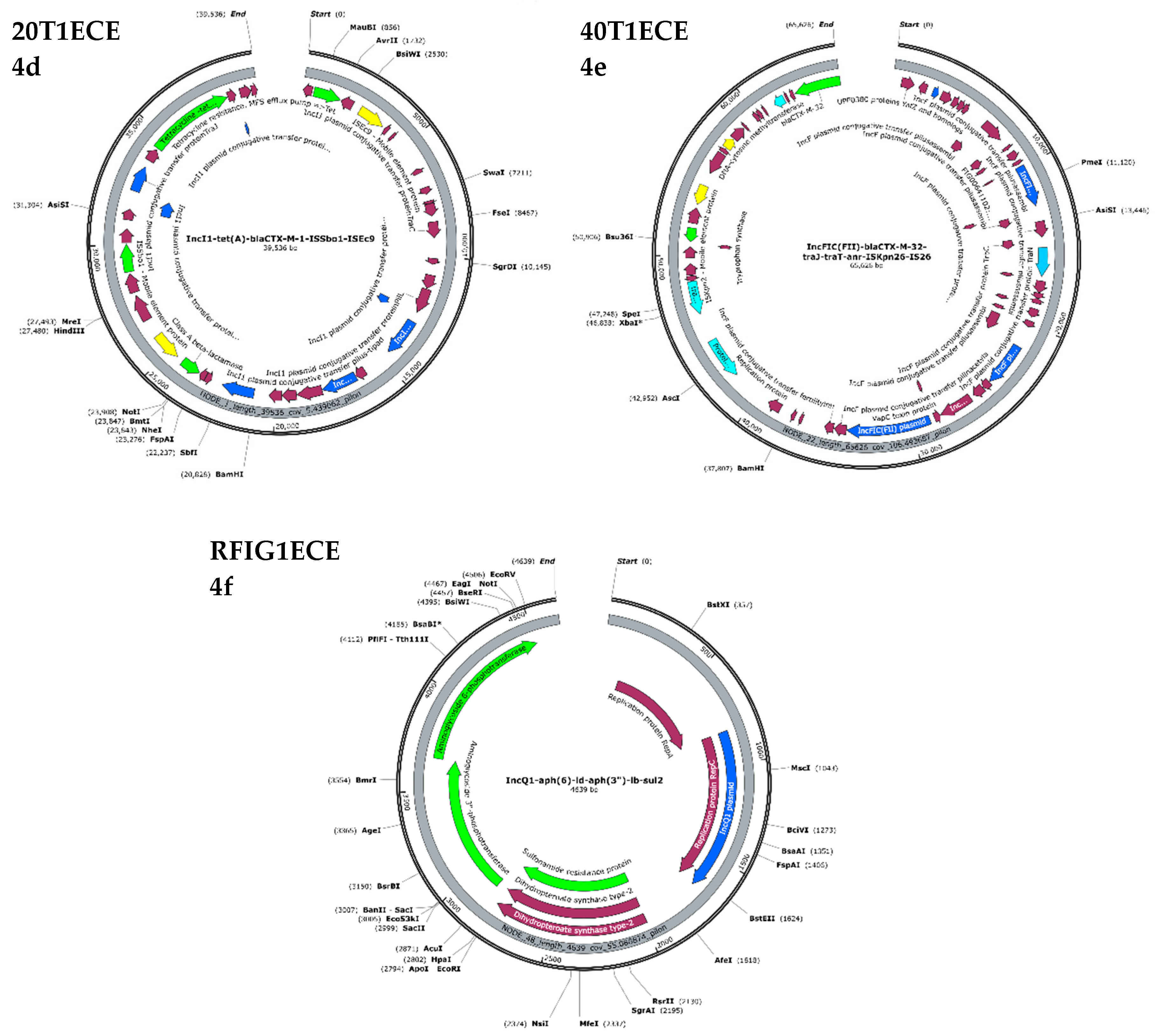
Figure 5.
SNP-based phylogeny of isolates from municipal wastewater treatment plants, cattle, and poultry farms in Fayetteville, NC. SNP-based maximum likelihood phylogeny of E. coli and ESBL-EC isolates visualized in iterative Tree of life tool (iTol). Clustering of isolates was found to be following the core genome and SNP- based phylogenies. The clustering of isolates belonging to the same sequence type was consistent. Shown for each isolate is the source, sample type, AMR genes and plasmid replicons.
Figure 5.
SNP-based phylogeny of isolates from municipal wastewater treatment plants, cattle, and poultry farms in Fayetteville, NC. SNP-based maximum likelihood phylogeny of E. coli and ESBL-EC isolates visualized in iterative Tree of life tool (iTol). Clustering of isolates was found to be following the core genome and SNP- based phylogenies. The clustering of isolates belonging to the same sequence type was consistent. Shown for each isolate is the source, sample type, AMR genes and plasmid replicons.
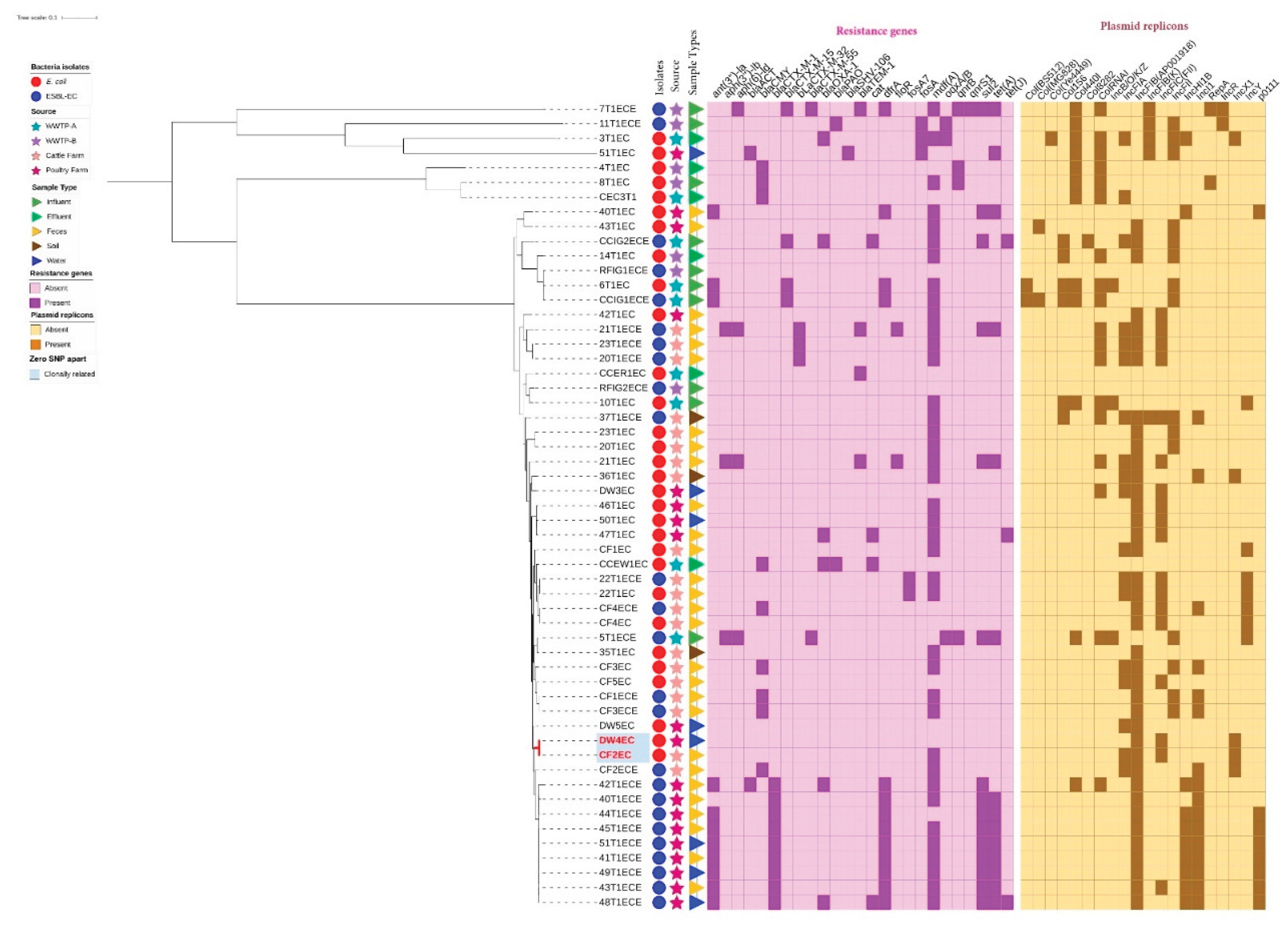

Table 1.
Antimicrobial resistance profiles of ESBL-EC isolates from cattle farm, poultry farm, and municipal wastewater treatment plants in Fayetteville, NC 2025.
Table 1.
Antimicrobial resistance profiles of ESBL-EC isolates from cattle farm, poultry farm, and municipal wastewater treatment plants in Fayetteville, NC 2025.
| Drug Class |
Antimicrobial drugs |
Resistance breakpoint (mm) |
Total (n = 124; %) |
Cattle Farm (n = 17; %) |
Poultry Farm (n = 68; %) |
WWTP-A (n = 22; %) |
WWTP-B (n = 17; %) |
| Tetracycline | Tetracycline (30 µg) | ≤11 | 84 (67.7) | 3 (17.6) | 61 (89.7) | 11 (50.0) | 9 (52.9) |
| Folate pathway antagonists |
Trimethoprim/Sulfamethoxazole (5 µg /300 µg) | ≤15 | 43 (34.7) | 0 (0.0) | 24 (35.3) | 10 (45.5) | 9 (52.9) |
| Penicillins | Ampicillin (10 µg) | ≤13 | 119 (96.0) | 16 (94.1) | 65 (95.6) | 21 (95.5) | 17 (100.0) |
| Penicillin (10 µg) | ≤ 14 | 121 (97.6) | 17 (100.0) | 65 (95.6) | 22 (100.0) | 17 (100.0) | |
| Quinolones | Ciprofloxacin (5 µg) | ≤21 | 31 (25.0) | 4 (23.5) | 3 (4.4) | 14 (63.6) | 10 (58.8) |
| Aminoglycosides | Streptomycin (10 µg) | ≤11 | 42 (33.9) | 1 (5.9) | 24 (35.3) | 10 (45.5) | 7 (41.2) |
| Phenicols | Chloramphenicol (30 µg) | ≤12 | 19 (15.3) | 2 (11.8) | 5 (7.4) | 8 (36.4) | 4 (23.5) |
| Cephalosporins | Cefotaxime (30 µg) | ≤22 | 119 (96.0) | 16 (94.1) | 65 (95.6) | 21 (95.5) | 17 (100.0) |
| Ceftazidime (30 µg) | ≤17 | 73 (58.9) | 13 (76.5) | 35 (51.4) | 16 (72.7) | 9 (52.9) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



