Submitted:
31 December 2025
Posted:
02 January 2026
You are already at the latest version
Abstract
The TGF-β superfamily, including BMP and TGF-β pathways, regulates fundamental cellular processes such as fate specification, tissue patterning, and stem cell homeostasis across metazoans. The BMP pathway across these diverse animal systems share a conserved intracellular SMAD signaling architecture. The quantitative dynamics of SMAD signaling and regulatory strategies governing pathway activity show a wide range of variation across developmental and stem-cell systems. In this review, we summarize insights from six major biological contexts — Drosophila embryo, germline stem cells, and the larval and pupal wing discs; the Danio rerio (zebrafish) embryo; and human pluripotent stem cells (hPSCs) — to compare how BMP and TGF-β signals are measured, manipulated, modeled, and integrated. We begin by outlining the canonical TGF-β/BMP signaling pathway and gradient formation across these species. We then compare the mechanism that shapes these signaling profiles from ligand diffusion, extracellular modulation and intracellular feedback through genetic, chemical, and mechanical perturbations. Finally, we highlight how endogenous pathway activity is measured through quantitative imaging of the pSmad/pMad activity, and how this approach contributes to the development of a full range of mechanistic and computational models across the presented systems. These insights reveal unifying design principles and performance objectives that govern BMP–SMAD signaling across species and cell types.
Keywords:
Developmental mechanisms
; Drosophila
; Zebrafish
; pluripotent stem cell
; systems biology
; BMP Pathway
Introduction
The Transforming Growth Factor β (TGF-β) superfamily of growth factors regulates homeostasis, differentiation, and regeneration, and is highly conserved throughout the animal kingdom. Ligands from the TGF-β superfamily include TGF-β isoforms, Bone-Morphogenetic Proteins (BMPs), Growth and Differentiation Factors (GDFs), activin, and nodal. The focus of this review is TGF-β isoforms in human pluripotent stem cells (hPSCs), Bone Morphogenetic Proteins (BMPs), and their orthologs in Drosophila, which regulate pattern formation and morphogenesis across several length and timescales. Particularly, we aim to review the quantitative advances in understanding the BMP-Smad pathway by primarily focusing on the work performed in six model systems: the zebrafish embryo, hPSCs, and Drosophila germline stem cells, embryo, and larval and pupal wing discs (Figure 1).
The TGF-β/BMP pathway regulates a broad range of developmental processes across species. In Drosophila it patterns the dorsal/ventral axis in the embryo, defines the anterior-posterior boundary in the larval wing disc, contributes to vein differentiation in the pupal wing and regulates maintenance and differentiation in germline stem cells. Likewise, in its vertebrate counterparts (Zebrafish and human pluripotent stem cells), this pathway governs embryonic axis formation, lineage specification and cell-fate decisions [1]. The core members of the TGF-β/BMP pathway include the TGF-β/BMP ligand, the Type-I and Type-II receptors, and the intracellular receptor-regulated (R-SMAD) and co-receptor-regulated (Co-SMADs) SMADs. In the canonical SMAD pathway, the extracellular BMP ligand (BMP2, BMP4, and BMP7) binds to the Type-I/Type-II transmembrane serine/threonine kinase receptors, which trigger intracellular phosphorylation of R-SMAD. R-SMAD then interacts with Co-SMAD to form a trimeric transcription factor that regulates several downstream target genes. Activin can also function as a ligand for the Type II receptors, to activate the pathway independent of the Type I receptor [2].
The structure and function of the TGF-β/BMP receptors and ligands are highly conserved; as such, human BMP ligands can replace their orthologs in Drosophila [3,4]. The homologs of the BMP ligands (BMP2, BMP4), Type I receptors (ALK2, ALK3, ALK6), Type II receptors (BMPR2, ACVR2), receptor-regulated Smads (R-Smad: Smad 1/5/9), and Co-receptor-regulated Smad (Co-Smad: Smad4) are prevalent across the vertebrate systems, including zebrafish, mice, and humans (Figure 2). The orthologs of BMP components in Drosophila include the ligands Decapentaplegic (Dpp; BMP2/4 ortholog) and Glass Bottom Boat (Scw/Gbb; BMP5/6/7/8 ortholog), the Type-I receptors Thickveins (Tkv), Saxophone (Sax) and Baboon (Bab), the Type-II receptors Punt (Punt) and Wishful Thinking (Wit), and the intracellular Smads, Mothers against Dpp (Mad; Smad1/5/9 ortholog) and Medea (Med; Smad4 ortholog (Figure 2). The signaling pathway recruits several extra- and intra-cellular mechanisms to distinctly regulate its downstream target genes in a tissue-specific, context-dependent manner across cell length scales ranging from microns in adult Drosophila germline stem cells to millimeters in the zebrafish embryo and time scales ranging from minutes to days in the Drosophila embryo and pupal wing, respectively (Table 1).
BMPs are tightly controlled during development and in homeostasis, since dysregulation can result in developmental defects or disease. Regulation commonly occurs through inhibitory Smads (I-Smads), Phosphatases (PPases), and E3 ubiquitin ligases, which are also conserved across these six model systems (Figure 2). The I-Smads (Smad6/7; Drosophila ortholog Daughters against Dpp, Dad) introduce negative feedback to the pathway topology as they are transcriptionally upregulated by the BMP signal transduction and terminate BMP signaling by interacting with the activated receptors or R-Smads [26]. Phosphatases regulate the BMP pathway through dephosphorylation of receptors or other downstream components. This dephosphorylation deactivates transcriptional controls to terminate the signaling cascade. Key phosphatases, such as CTDNEP1 (C-terminal Domain Nuclear Envelope Phosphatase 1; Dullard (Dd) in Drosophila), are also conserved across species and are suspected to regulate the pathway through the dephosphorylation of Type I receptors or R-SMADs; however, it is worth noting that these phosphatases are not specific to the BMP pathway and also dephosphorylate other components in the Wnt pathway [27,28,29]. Likewise, E3 ubiquitin ligases regulate this pathway by promoting the proteolysis of Smads [27,30]. This includes the SMURF family of E3 ubiquitin ligases, which are major negative regulators of the BMP pathway in hiPCS, zebrafish embryo, and Drosophila [30,31,32]. The complexity of these interactions underscores the importance of continued research into the mechanisms of pSmad regulation to fully elucidate the intricacies of TGF-β/BMP signaling.
The mechanism through which extracellular BMP gradients are formed is highly context-dependent, including combinatorial signal perception and regulation through BMP inhibitors, enabling adaptation to specific developmental contexts, ranging from short-range signaling in Drosophila female germline stem cells to long-range signaling in the Drosophila wing disc [33,34,35]. Despite the diversity of extracellular cues that activate the TGF-β/BMP pathway, the intracellular SMAD module that interprets these signals is remarkably conserved, functioning as a dynamic decoder that transforms spatial gradients and temporal fluctuations into precise transcriptional outputs. There is growing evidence from both experimental and computational studies that SMAD signaling is a dynamic process, including dynamic signal processing [32,33,36], which provides a more complex function that cannot be captured through typically quantitative approaches that measure BMP concentration. Schmierer et al. demonstrated that rapid and continuous shuttling of Smad proteins between the cytoplasm and nucleus enables cells to effectively integrate signaling over time, filters noise, and allows cells to detect transient BMP inputs [37]. Findings in human embryonic stem cells (hESCs) demonstrate that rapid changes in BMP concentration are key drivers of self-organized patterning through their effects on pSMAD dynamics and downstream WNT/NODAL activation [38]. These insights indicate that the robustness and versatility of BMP/TGF-β signaling arise not only from classical inhibitory controls but also from the dynamic signal-processing capabilities, which allow conserved intracellular machinery to generate diverse developmental outcomes as a modifiable signaling module.
Here, we summarize the recent quantitative understanding of the mechanisms that confer diverse modalities in the TGF-β/BMP pathway tuned for dynamic performance across model systems. These include the vertebrate model systems, zebrafish and mammals, and the invertebrate Drosophila systems, germline stem cells, embryo, wing disc, and the adult wing, which each function under different time and scale constraints (Table 1). First, we review the mechanism of gradient formation and highlight the similarities and differences in gradient diffusion lengths, signaling schemes and underlying feedback loops across the six model systems (Section I). Second, we examine how BMP activity is interpreted through a tiered transcriptional response of downstream target genes (Section II). Third, we highlight that pSmad has conventionally been used as a proxy for extracellular TGF-β/BMP gradients and evaluated the quantitative live imaging approaches to record endogenous expression levels (Section III). Finally, we perform a meta-analysis of the mechanistic and computational models of the pathway across the chosen model systems (Section VI). We emphasize how these elements unify design principles and performance objectives for the BMP-Smad pathway across species and cell types (Section V). Finally, we discuss performance objectives of BMP signaling across diverse biological contexts, and how experimental data and computational models can be integrated to better understand the versatile dynamics of this pathway (Section V).
Mechanisms of Gradient Formation Across Development
Morphogens provide positional information to cells by spatially varying its concentration across tissues. In the TGF-β/BMP pathway, these gradients arise through a combination of ligand diffusion, extracellular modulation, receptor interactions, and feedback control. Although the core signaling machinery is highly conserved, the specific mechanisms that generate BMP gradients differ substantially across developmental contexts, reflecting variations in tissue architecture, regulatory proteins, and signaling dynamics. Here’s a summary for the six systems of interest in this review:
Drosophila blastoderm embryo
In Drosophila, the mechanism behind Dpp gradient formation changes during different stages of development. In the Drosophila blastoderm-stage embryo, BMP ligands Dpp and Scw are expressed broadly; however, their spatial activity is modulated by extracellular inhibitors (Short gastrulation (Sog) and Twisted gastrulation (Tsg), which bind to Dpp/Scw and prevent receptor activation in lateral regions. These ligand-inhibitor complexes diffuse dorsally, where the metalloprotease Tolloid (Tld) cleaves Sog, releasing active Dpp/Scw ligands near the dorsal midline. This facilitated diffusion mechanism, more commonly known as “shuttling,” concentrates BMP ligands dorsally. In early embryogenesis in Drosophila, BMP establishes a sharply localized dorsal signaling domain through a combination of diffusion, binding, and proteolytic regulation. One proposed mechanism of gradient formation through Sog/Tsg shuttling and Tolloid-mediated cleavage was successfully mimicked in vivo behavior, which demonstrated that canonical pMad gradient formation in the early embryo requires the presence of BMP antagonists (Sog and Tsg) and the metalloprotease, Tolloid [24,39,40]. Using a diffusion–degradation model, it was demonstrated that ligand diffusion alone cannot account for the sharpness of the signaling domain and that BMP initially accumulates at the dorsal midline before gradually intensifying and broadening both in amplitude and spatial extent [8]. Analysis of this model demonstrated that it successfully captured the progressive growth of the signaling domain but did not reproduce the sharpening and contraction of the pMad signal towards the dorsal midline observed in vivo. Based upon this, Umulis et al. (2006), advocated that incorporating feedback regulation and receptor mediated degradation along with shuttling was necessary to achieve the rapid sharpening and spatial bi-stability seen in the system [41]. Specifically the concepts of spatial bistability and intracellular positive feedback was adopted from Wang & Ferguson (2005), whose experimental results showed the dependence of spatial patterning upon this positive feedback [42]. In a follow-up study by Umulis et al., 3D modeling of the Drosophila embryo further showed the consistency between the suggested feedback regulation mechanism and the experimental data [43].
Additionally, the Drosophila embryo also exhibits scale invariance. Dorsal surface patterning by Dpp/BMP scales between closely related species and between individuals within a species; specifically, the ratio of pMad pattern width-to-embryo length is constant between Drosophila melanogaster, the larger Drosophila virilis, and the smaller Drosophila busckii, as well as among individual embryos of differing sizes within each species [43].
Drosophila Larval Wing Disc
During larval wing disc development, Dpp is expressed down the middle of the imaginal disc, which allows it to form a long range gradient [44]. This Dpp gradient facilitates the formation of precursors of the longitudinal veins (LVs) in the wing disc along the anteroposterior axis of the wing disc [8]. The major competing hypotheses regarding the long-range Dpp gradient formation in Drosophila wing discs are through receptor-mediated transcytosis, active transport mechanisms involving cytonemes, and restricted extracellular diffusion (including hindered diffusion)[45,46]. According to the receptor-mediated transcytosis hypothesis, long-range distribution and transport of Dpp occur through repeated receptor-mediated uptake and secretion.
Receptor-mediated transcytosis was previously a leading hypothesis in the Dpp gradient formation in the wing disc; however, it more recent years, it has since been argued that this mechanism cannot account for rapid Dpp diffusion and may have a limited role in Dpp transport [46]. Another proposed alternate mechanism of Dpp movement involves cytoneme-mediated active transport, which involves cellular projections that directly connect the ligand-receiving to ligand-producing cells. The cytonemes are suspected to directly transport the receptors bound to Dpp between cells. Notably, the role of cytonemes in Dpp gradient formation in wing discs has not been conclusively established, but more recent work has shown direct imaging of cytonemes in the adjacent air sac primordium (ASP) [47]. Considering a synthesis of recent studies, restricted extracellular diffusion is suspected to be the major mechanism behind Dpp gradient formation in the larval wing disc, but cytoneme-mediated transport may play a role as well. During restricted-extracellular diffusion, extracellular matrix proteins, such as Heparan Sulfate Proteoglycans (HPSGs) act as ligand sinks upon interacting with Dpp to facilitate Dpp cellular endocytosis [45]. A direct investigation between receptor-mediated transcytosis and restricted-extracellular diffusion, further supported this argument by demonstrating that long-range distribution of Dpp occurs through restricted extracellular diffusion when utilizing computational modeling, gain-of-function and loss-of-function Tkv receptor mutant clones [45].
To better demonstrate this phenomenon, other gradient formation models attempt to combine extracellular diffusion and receptor/matrix interactions to explain the mechanism of the Dpp gradient formation through the wing disc through temporal regulation of gene expression. In one particular model, Dpp levels are highest near the anterior-posterior axis and reduced laterally, due to the gradient formation [48]. This model further explores a hypothetical scenario in which Dpp levels are made uniform across the tissue. Under this assumption, lateral cells, which normally receive lower Dpp, are predicted to experience increased proliferation due to their lower response threshold, whereas medial cells, which are normally exposed to high Dpp levels, will experience less proliferation. However, this model fails to accurately portray in vivo behavior where proliferation is uniform across the wing disc, despite the Dpp gradient signaling. The Dpp gradient in Drosophila wing disc also exhibits dynamic scaling, as characterized in [49], who proposed the Expansion-Repression (ER) model, where a secreted inhibitor Pentagone (Pent) acts as a rapidly diffusing "expander" molecule of the Dpp gradient [49]. This expansion provides the necessary scaling. However, Zhu et al. (2020) challenged Pent’s role as the primary global expander due to its limited spatial range [50]. They instead proposed a pseudo source-sink model, demonstrating that scaling is driven primarily by the Dpp-mediated feedback downregulation of its own receptors, rather than by a secreted expander. Another mechanism has been suggested by [51], who propose that Dpp gradient scaling is driven by a tunable "recycling gear," where the feedback regulator Pent modulates receptor binding kinetics to favor the re-exocytosis of internalized ligand over its lysosomal degradation. By progressively increasing the fraction of Dpp returned to the extracellular space, this mechanism actively extends the morphogen’s effective range to match the growing tissue size.
Drosophila Pupal Wing
Unlike Dpp signaling in the Drosophila embryonic and larval development, pupal wing Dpp signaling lacks a continuous 2D gradient and instead transitions between planar and localized 3D Dpp distributions (18). Instead, it relies on localized ligand distribution through restricted extracellular diffusion from discrete, vein-associated sources, where the ligand is locally captured in crossvein territories through shuttling [9,20,52]. During this stage of development, Dpp signaling drives the formation of the longitudinal veins (LVs) and the crossveins (CVs) [8]. While LV precursor cells are first defined during larval development, the patterning of the CVs occurs during the pupal stages [8]. At this stage, Dpp expression expands into the LVs where it acts locally to refine LV fate and at a long range to direct CV formation [8]. Formation of the posterior crossvein (PCV) is a result of Dpp expression in adjacent LVs; whereas, the anterior crossvein (ACV) is formed when Dpp is expressed in a stripe intersecting that region [8].
Dpp signaling in the pupal wing regulates tissue proliferation following a transition from lateral diffusion within the LVs to interplanar diffusion between epithelial layers (8,20,21,53). There are 3 main stages during pupal development (first apposition, inflation, and second apposition), which take place eight hours, ten to twenty hours, and twenty hours after pupal formation (APF), respectively. During the first apposition, Dpp is expressed in the LV primordia cells (LV2-5). Dpp maintains a tissue-wide gradient through lateral diffusion during inflation (~18 hr APF) and regulates tissue proliferation by inhibiting brk expression [21,53]. During second apposition (~24 hr APF), Dpp signaling becomes localized for LV formation, diffuses vertically between the dorsal and ventral epithelia, and undergoes long-range diffusion from adjacent LVs into the PCV region to facilitate PCV formation and the formation of the 3D tissue architecture (8,21,53).
Paralleling the facilitated transport mechanisms described in the Drosophila embryo, extracellular regulators including Sog and Tolloid-related proteases, shape Dpp signaling in the pupal wing [8]. However, the Tolloid-related protease involved in PCV formation in the pupal wing, Tolloid-related (Tlr), exhibits slower Sog cleavage kinetics (fivefold) than its embryonic counterpart Tolloid (Tld) and performs a non-redundant, tissue-specific function [33,54]. Transient Dpp expression is thought to be stabilized by Dpp signaling in the anterior lateral region of the pupal wing through the positive feedback factor, crossveinless-2 (CV-2) [8,55]. CV-2 is suspected to promote local Dpp signaling by facilitating ligand retention and spatial redistribution [8].
While quantitative models describing the Dpp gradient have been developed for the Drosophila embryo and larval wing, no quantitative models currently exist that describe Dpp signaling in the pupal wing, likely due to the discrete and transient nature of Dpp sources at this stage. Instead, Dpp signaling in the pupal wing has been primarily described through conceptual models involving restricted extracellular diffusion and shuttling ligand redistribution mediated by extracellular regulators such as Sog and Tolloid-family proteases. Additionally, it is suspected that their is a feedback mechanism between LV and PCV formation that facilitates the vectorial transport of Dpp from LVs to the PCV. This existence of feedback is supported by disrupted BMP signaling when LV Tkv receptors are constitutively active, and increased LV Dpp motility and improper vein formation when disrupted [53].
Drosophila Germline Stem Cells
Dpp signaling is required for the self-renewal of germline stem cells (GSCs); however, it needs to be downregulated to allow GSCs to differentiate into Cystoblasts [22]. The GSCs are at the apical tip of the germarium in the niche, which is composed of the Terminal Filament cells (TFCs), Cap cells (CpCs), and Anterior Escort Cells (AECs). JAK/STAT signaling in the CpCs [56] and Wnt signaling in the AECs [57] produce Dpp, such that the Dpp originating in the CpCs is the major source of the ligand in GSCs [13]. The CpCs being at the apical tip of the germarium ensures a higher level of Dpp is available to the GSCs than the Cystoblasts, which are the differentiated daughter cells of the GSCs [13]. Several intracellular feedback mechanisms, including the positive feedback mediated through Fused (Fu) [58,59,60], and the negative feedback mediated through Daughters against Dpp (Dad) [60,61,62] ensure Dpp signaling is upregulated in GSCs and downregulated in the differentiated daughter cells located one cell diameter away to precisely control stem cell differentiation on a length-scale of 2-cell diameters. In addition to these feedback mechanisms, the GSCs form cytoneme projections in response to BMP signaling which attenuates BMP signal transduction enabling dynamic modulation of BMP signaling after GSC division [13].
Zebrafish
During zebrafish embryogenesis, secreted BMP2 and BMP7 homo- and heterodimers are secreted by a minority of cells [63]. The extracellular regulatory network sharpens dimeric BMP ligands to establish a gradient across the zebrafish embryo, characterized by high ligand concentration on the ventral side and progressively lower levels toward the dorsal side, across which they interact with and recruit cell membrane receptors into signaling complexes that transduce the spatially patterned Smad activation within cells. A diverse set of extracellular regulators plays a crucial role in regulating this gradient, including antagonists Chordin (homolog of Drosophila Sog), Noggin, and Tsg, which bind BMP ligands and limit their diffusion; and modulators such as Tolloid (Tld), Crossveinless-2 (CV2), and Sizzled influence ligand stability, degradation, and shuttling [64,65,66,67]. These regulators contribute to counter-gradients, intracellular shuttling, and transcriptional mechanisms, enhancing the overall robustness of gradient formation. More recent characterization of this gradient through quantification and the development of a computational model has indicated gradient formation via a source-sink mechanism is the most likely way the gradient forms[63,68] . A critical aspect of this patterning is scale invariance: the ability to maintain proportional patterning when the size of the embryo changes. In zebrafish, the BMP signaling gradient maintains scaling despite experimental reductions in embryo size of up to 30%, a robustness achieved by active feedback regulating the stability of the Chordin sink [36]. Li et al. have since generated a model that utilizes complex three-dimensional virtual embryos and simulated advection-diffusion-reaction using partial differential equations (PDEs) to more closely mimic the geometry of the system, which further supported the source-sink mechanism [69]. However, limitations of this model derive from limited accuracy in experimentally quantified gene expression domains due to their significant impact on model outputs.
Human Stem Cells
In vitro models such as micropatterned hESC/hiPSC colonies mimic peri-gastrulation–like patterning using BMP4 stimulation. When colonies are stimulated with uniform BMP4, they self-organize into a radial gradient of SMAD1/5/8 activity [70]. Inhibitory signals restrict BMP4 activities to the colony periphery and generate an Activin/Nodal gradient toward the interior that patterns mesendodermal fates. On the other hand, BMP-SMAD signaling exhibits an asymmetric gradient in the post-implantation amniotic sac embryoid, which is suspected to play a role in tissue patterning. During pre-gastrulation, there is a prominent population of nuclear pSMAD1/5 at amniotic poles, and the nuclear pSMAD1/5 is present in the tissues before CDX2, which is a transcription factor for differentiation and proliferation of cells [71]. Quantitative computational models, including reaction-diffusion frameworks [72], agent-based models focused on gene regulatory networks (GRN) and cell-cell interaction [73], and cross-signaling models of the BMP-Wnt-Nodal cascade model [74,75], have shown how colony geometry, inhibitor feedback, and signaling dynamics collectively shape the BMP-SMAD gradient. These modeling studies demonstrate that the patterning in hiPSCs, along with other developmental systems, arises from a pattern from a self-organized BMP/Smad signaling gradient, and both the formation and interpretation can be accurately captured and predicted through computational approaches. Furthermore, these modeling studies can be used to improve differentiation efficiency toward mesodermal and endothelial cells [76,77].
Figure 3.
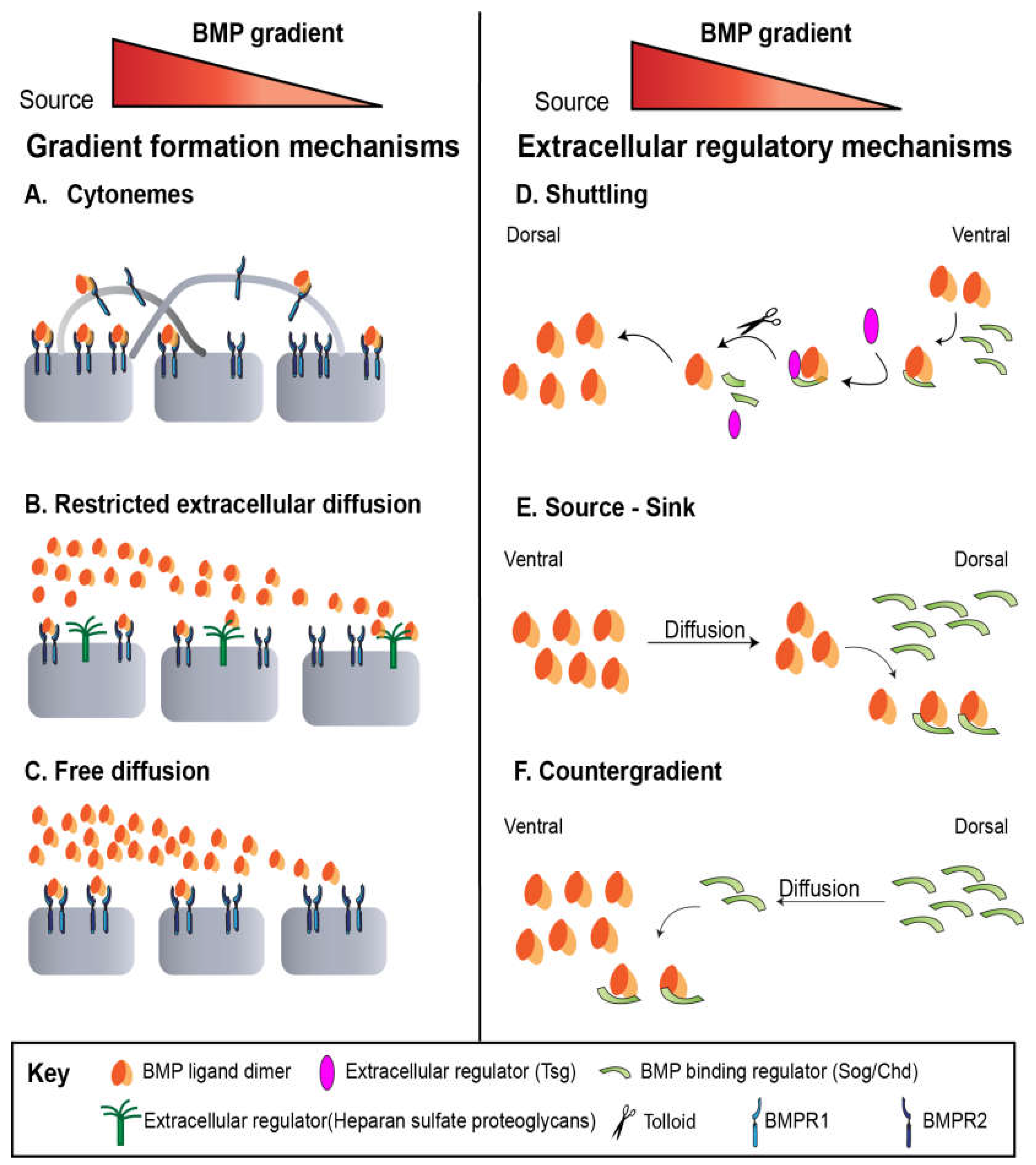
Summary of proposed mechanisms of gradient formation across systems. The concentration of BMP decreases from left to right, as shown by the top figure. (A) Gradient formation by cytonemes: Cytonemes are thin, cellular projections that can emanate from the cell towards the morphogen source. (B) Gradient formation by restricted extracellular diffusion: Extracellular proteins in the matrix regulate and restrict free diffusion. (C) Gradient formation by free diffusion: The BMP ligand diffuses from regions of high concentration to low concentration. (D) Shuttling mechanism, as employed in Drosophila embryo and pupal wing; BMP regulator, Sog concentrates BMP towards the dorsal midline. (E) Source-sink mechanism, as employed in zebrafish; BMP regulator Chordin acts as a dorsal sink for the freely diffusing BMP. (F) A counter gradient, as proposed in zebrafish, where Chordin diffuses to regulate BMP.
Figure 3.
Summary of proposed mechanisms of gradient formation across systems. The concentration of BMP decreases from left to right, as shown by the top figure. (A) Gradient formation by cytonemes: Cytonemes are thin, cellular projections that can emanate from the cell towards the morphogen source. (B) Gradient formation by restricted extracellular diffusion: Extracellular proteins in the matrix regulate and restrict free diffusion. (C) Gradient formation by free diffusion: The BMP ligand diffuses from regions of high concentration to low concentration. (D) Shuttling mechanism, as employed in Drosophila embryo and pupal wing; BMP regulator, Sog concentrates BMP towards the dorsal midline. (E) Source-sink mechanism, as employed in zebrafish; BMP regulator Chordin acts as a dorsal sink for the freely diffusing BMP. (F) A counter gradient, as proposed in zebrafish, where Chordin diffuses to regulate BMP.
Interpretation of BMP Signals via Transcriptional Regulation
Despite differences in ligand–receptor composition and extracellular modulators, the intracellular Smad-mediated signaling logic downstream of BMP remains remarkably conserved. Understanding how this conserved signaling machinery translates intercellular BMP gradients into precise target gene expression patterns provides a critical link between molecular signaling mechanisms and systems-level developmental outcomes. Across all these systems, the BMP activity is interpreted through a tiered transcriptional response of downstream target genes. This network includes both direct activators and negative regulators that collectively shape the signaling outcome.
Drosophila embryo development
BMP signaling in Drosophila embryos directs downstream target genes that have been classified into three types—Type I, II, and III—based on their threshold sensitivities to BMP signal strength and their spatial expression patterning [78]. Several studies have illustrated these distinctions by examining how spatial domain of gene expression shifts in wild-type and mutant embryos [8,24,43,79].
Type I genes are induced only under high BMP signaling at the dorsal midline. Type II genes require moderate BMP input and are expressed in an intermediate band. Type III genes are the most sensitive and respond to low levels of BMP signaling. These genes are activated early and often maintain broad expression after the BMP signal refines to a narrow dorsal stripe of 3-4 cells, suggesting a memory mechanism may be at play [17]. Consistent with this, Brantley et al. (2025, preprint) observed a gradual, spatially broad accumulation of zen protein before gastrulation, forming a broad distribution exceeding the width of the Med-GFP gradient across the dorsal surface. Zen may act as an early “sensitizer” that helps cells respond to low BMP, broadening or accelerating the transcriptional response [18]. These findings illustrate the spatiotemporal control of BMP signaling required for precise patterning in early Drosophila embryos.
Another study revealed that BMP signaling modulates the frequency of transcriptional bursts—periods when a gene is actively transcribed—rather than the speed of transcription once active by measuring ush (II) and hnt (III) in the embryo. In lower BMP regions, both genes showed reduced promoter occupancy and burst frequency due to a lower rate of promoter activation, while the transcriptional loading rate and burst duration remained unchanged [80].
Drosophila Larval Wing Disc
In the larval wing disc, the pMad–Medea complex drives activation of target genes such as brinker (brk), optomotor-blind (omb), daughters against dpp (dad), and spalt major (sal). Omb and sal are transcription factors that promote the transcription of genes involved in wing formation and patterning, whose expression scales with BMP signal strength. When there is little to no Dpp bound to the receptors, brk is expressed and represses dad by blocking its transcription. At low BMP levels, omb continues to be transcribed even though dad is suppressed, which does not occur when Dpp is absent [44], guiding proper adult wing shape and vein formation.
Dad, an inhibitory Smad, is directly induced by BMP signaling and suppresses further R-Smad phosphorylation through inhibition of the Type I receptor, forming a negative feedback loop that prevents cells from prolonged or excessive signaling. Brk acts as a transcriptional repressor whose expression is inversely proportional to Dpp signaling. Under high levels of BMP, the pMad-Medea complex recruits the co-repressor Schnurri (Shn) to silence the locus of brk, allowing transcription of dad, omb, and sal [13].
Beyond molecular feedback regulation, several systems-level models have been developed to capture how feedback control stabilizes the Dpp gradient and maintains robust tissue growth. Feedback-control models incorporate negative regulators to stabilize the Dpp gradient and promote robustness during tissue growth. Two main frameworks, growth equalization and Vestigial (Vg) feed-forward models, both implement transcriptional feedback loops that balance proliferation across the wing disc. In the growth equalization model, Brk acts as a growth suppressor that is inversely regulated by Dpp activity. This model predicts that Dpp activity is highest in medial cells and declines laterally. The growth equalization model also predicts that if Dpp signaling were artificially uniform across the tissue, lateral proliferation would increase due to reduced Brk expression. However, this prediction contradicts in vivo findings that demonstrate Brk levels are normally high in lateral regions, where they suppress Dpp/BMP pathway activity and reduce proliferation [44].The Vg feed-forward model postulates that coordinated input from the Dpp, Wg, and Vg morphogens creates a feed-forward loop that recruits adjacent non-wing cells to become proliferating wing cells. In this model, Dpp and Wg induce Vg expression; Vg then cooperates with Yorkies to reinforce Vg expression in adjacent cells, thereby expanding wing cell development through a positive feedback loop [44].
Drosophila Pupal Wing
The core mechanisms of the BMP signaling pathway described in the Drosophila larval wing disc are conserved during the pupal stage of development; however, the downstream transcriptional targets differ. As a direct output of the pMad-Medea complex, Dad is the only conserved downstream transcriptional target between the larval and pupal stage [6,7]. The other direct downstream target of the BMP pathway during pupal development is the BMP-responsive extracellular regulator, Crossveinless-2 (CV-2) [8,9]. CV-2 is an extracellular regulator of the BMP pathway that is suspected to act in a biphasic manner where it promotes BMP signaling at low Dpp levels and inhibits BMP activity at high Dpp levels [52]. The biphasic manner of CV-2 is supported by findings where ectopic BMP signaling resulted in high levels of CV-2, and mutations in CV-2 prevented local BMP signaling in the PCV during pupal development [52].
Other downstream targets of the BMP pathway during pupal development, occur indirectly though crosstalk with the EGFR/MAPK and Notch pathways. These indirect downstream targets of the BMP pathway facilitate vein differentiation through the expression of Rhomboid (rho), Star (s), and argos (aos), and the repression of blistered/Srf (bs) [10,11,12] in the EGFR/MAPK pathway. EGFR signaling is amplified by Rho and Star expression, which are believed to be integral in vein formation, since they are highly expressed in L3 and L4 proteins, where Dpp levels are amplified in the pupal wing [10]. Aos is an inhibitors that interacts with the EGFR pathway through a negative feedback loop [10]. During pupal development, the EGFR pathway repressed bs expression, which is lost in the proveins and retained in the interveins [10,12]. Inactivation of the EGFR pathway disrupts vein formation in the pupal wing, in particular, loss of rho and star block distal vein formation and have little effect on the LVs and CVs [10]. The Knirps/Knirps-like transcription factors (Kni/Knrl) induce EGFR activation in the L2 provein through rho transcription, by transducing BMP pathway positional information. Development of the CVs in the pupal wing is first initiated by BMP signaling but is maintained until adulthood through the EGFR pathway upon rho and star transcription, likely caused by BMP pathway cues. Ectopic vein formation is a result of pathway crosstalk between the EGFR, BMP, and Notch pathways, where hyperactivity of rho and star interact with Dpp to activate BMP signaling, which then activates the Notch pathway through its ligands, Delta and Serrate [10]. Loss of one these pathways reduce activity of the other pathways, demonstrating their interaction [10].
Drosophila Germline Stem Cells
In the Drosophila ovary, BMP signaling in the germline stem cell niche maintains GSC identity through differential gene expression. This is achieved through repression of bag of marbles (bam), a driver of germline differentiation. Asymmetric division moves the differentiated daughter cystoblast (CB) away from the niche, reducing BMP exposure and hence pMad levels; with the loss of BMP-mediated repression, bam is derepressed in the CB, initiating differentiation. Genetic perturbations that maintain high pMad in daughter cells prevent bam expression and block differentiation, underscoring the necessity of down-modulating BMP after niche exit [81,82]. The sharp pMad-bam axis is maintained by intermediaries including a serine/threonine kinase Fused (Fu), along with E3 ligase Smurf which targets BMP receptor Thickveins for ubiquitination in the CBs [58].
Further, Wilcockson and Ashe (2021) performed RNA-Seq on genetically expanded GSC populations—flies carrying constitutively active tkv together with bam knockdown—allowing comparison of “high Dpp” and “low Dpp” GS [13]. In the high Dpp condition, pMad⁺ cells were observed throughout the germarium, reflecting global activation of the self-renewal program. In the low Dpp condition, bam knockdown limited differentiation, which again led to an abnormal increase in pMad⁺ cells across the germarium. This work identified ~300 Dpp-responsive genes, including dad, Rfx, and futsch, thereby broadening the catalog of BMP targets in germline stem cell regulation.
Intracellular regulation, mediated by phosphatases and feedback loops, sharpens this transition to establish pMad asymmetry. The GSC division cycle has a duration of 12-15 h during which the GSC and preCB have a shared cytoplasm. The distribution of pMad in the G1/S phase of cell division is asymmetric, such that pMad expression in the preCB is less than the pMad expression in the GSC. Mad phosphatase, Dullard, dephosphorylates Mad at the nuclear pores which in conjunction with higher BMP activation in the GSCs than CBs establishes pMad asymmetry. Further Daughters against Dpp (Dad), upregulated by BMP signaling within the niche, limits receptor-mediated phosphorylation of R-Smads and targets Type I receptor Tkv for ubiquitination [13,62]. This negative feedback likely enforces the steep signaling boundary between GSCs and CBs, minimizing “leakage” of stemness signals into differentiating daughters. This network ensures that only cells in proximity to the niche retain stem cell identity, while even modest decreases in BMP signaling in daughters reliably trigger bam expression and differentiation. This spatially gated, robust switch safeguards the balance between self-renewal and differentiation.
Several modeling works have focused on simulating the bistable nature of the GSC/CB differentiation. Pargett et al., (2014), developed a model of BMP signaling in the germarium to identify the role of Brat as a differentiation factor [83]. Their simulations suggested that Pum–Nos-mediated degradation of Brat in GSCs preserves self-renewal, while in CBs, the absence of this repression allows Brat to antagonize BMP signaling and promote differentiation. Xia et al. (2012) modeled a positive feedback loop between Fused (Fu) and the BMP pathway, showing that Fu degradation within GSCs sustains BMP signaling to maintain stemness, whereas Fu accumulation in CBs facilitates pathway downregulation and differentiation [59]. More recently, Shaikh and Reeves (2024) developed a biologically-informed mathematical model of multi-compartment GSC division to investigate the dynamic roles Dad and Fused play in determining cell fate . Their analysis revealed that Dad optimally tunes BMP signal transduction to maintain GSC homeostasis, and in Dad knockout mutants, GSCs were more likely to divide symmetrically. Their work identifies the synergistic role of Dad and Fused, rendering robustness to stem cell division.
Zebrafish
In the developing zebrafish embryo, cells have been shown to interpret the BMP gradient through concentration threshold [14]. Distinct levels of phosphorylated Smad5 (pSmad5) activate specific sets of target genes, creating sharply defined expression domains across the embryo. Using transcriptional profiling combined with single-cell pSmad5 measurements, Greenfeld et al. identified over fifty primary BMP target genes and demonstrated that cells decode BMP signaling based on absolute pSmad5 concentration cutoffs, rather than relative position or temporal history of exposure [14].
Similar to the Drosophila embryo target genes, these BMP target genes can be grouped into three categories according to their activation thresholds and spatial expression profiles along the dorsoventral (DV) axis. High-threshold genes, such as sizzled (szl) and tp63, are expressed exclusively in the most ventral regions where BMP signaling is strongest. Intermediate-threshold genes, including foxi1 and gata2a, are activated in the ventral-animal domains at moderate pSmad5 levels. Finally, low-threshold genes such as bambia (or bambi) respond to weaker pSmad5 activity closer to the dorsal side. Together, these categories represent discrete transcriptional responses corresponding to specific concentration steps within the BMP gradient, suggesting that pSmad5 functions as a digital readout gated by defined concentration thresholds [14,84].
Several of these BMP target genes also participate in feedback mechanisms that fine-tune the pathway. bambia (low-threshold) encodes a transmembrane pseudoreceptor that attenuates BMP signaling, acting as a negative regulator of the pathway in the already low-activity area [85]. Similarly, szl (high-threshold) encodes a secreted inhibitor that prevents Chordin (Chd) cleavage by Tolloid (Tld), thereby maintaining the BMP gradient; szl also possesses both BMP and Wnt inhibitory activity [65,86]. At the same time, high BMP activity represses chd transcription, confining its expression to the dorsal regions. Since these feedback loops become active after gastrulation onset, they serve primarily to stabilize and regulate the established BMP signaling domains rather than to initiate them. A computational model of this system from Tuazon et al. in 2020 is one of the only ones to consider transcription, which makes the network considerably more complex [87]. Their model strongly supported the source-sink mechanism under comparison with wild-type and a variety of mutant lines [87].
Beyond this threshold mechanism, BMP target gene expression is refined through combinatorial and temporal inputs from other signaling pathways. Crosstalk with FGF and Nodal signaling helps sharpen domain boundaries; inhibition of either pathway collapses the diversity of BMP target expression into uniform ventralized patterns [68,87,88]. In parallel, extracellular modulators such as Chd, Bmp1a, and Tld sculpt the BMP gradient by restricting Chordin diffusion and establishing localized ligand sinks. Together, these interactions ensure the precision and robustness of BMP-mediated dorsoventral patterning in the zebrafish embryo.
Human Pluripotent Stem Cells
In human pluripotent stem cells, including human embryonic stem cells (hESCs) and human induced pluripotent stem cells (hiPSCs), some of the most immediate and conserved BMP targets are the Inhibitor of Differentiation genes ID1–ID3 [16,89,90]. ID proteins interfere with factors that promote neural identity, preventing premature activation of the neural program and helping the cells remain undifferentiated [90,91,92,93,94]. In both mouse and human pluripotent systems, ID proteins serve as key mediators translating SMAD signaling into transcriptional and epigenetic control of pluripotency and lineage bias [95]. ETV2 and ETS2 are important transcription factors that work with the BMP pathway to regulate hiPSCs differentiation into mesodermal and endothelial cells [96,97].
BMP signaling also activates a multilayered negative-feedback network that tightly regulates the duration and intensity of the pathway at both intracellular and extracellular levels. The inhibitory SMADs, SMAD6 and SMAD7 are transcriptionally induced by BMP–SMAD complexes and subsequently and directly inhibit receptor activation by competing with R-Smads or by recruiting E3 ubiquitin ligases (SMURF1/2) to promote receptor and R-SMAD degradation [98,99,100,101].
Similarly, to the above and to zebrafish, BAMBI (BMP and activin membrane-bound inhibitor), a kinase-deficient pseudoreceptor, is upregulated by BMP signaling and inhibits further receptor complex formation, adding another layer of intracellular negative feedback [102,103,104]. At the extracellular level, BMP signaling also induces several secreted antagonists, including NOGGIN, CHORDIN, and follistatin [105,106,107]. These factors sequester BMP ligands and restrict their diffusion by competitively binding them, thereby establishing a complementary feedback circuit that modulates BMP availability at the ligand–receptor interface [108,109].
Phosphorylated Smad as a Proxy for BMP Signaling Activity - Quantitative measurements
Across all the model organisms described, Phosphorylated Smad1/5/(8/9) (pSmad) in vertebrates and phosphorylated Mad (pMad) in Drosophila serve as the primary quantitative readouts of BMP pathway [110,111,112]. Their nuclear localization provides a robust and experimentally accessible proxy for ligand–receptor engagement, signal transduction, and transcriptional output. Because secreted Dpp/BMP ligands are difficult to directly visualize due to their complex extracellular regulation interactions, intracellular Mad/Smad protein phosphorylation (pMad/pSmad) is used as a functional reporter of pathway activation). Immuno-fluorescent staining followed by the confocal microscopy enables quantification of nuclear pMad/pSmad intensity, which reflects the spatial distribution of BMP signaling across tissue [36,113]. The relative fluorescent intensities provide a robust and reproducible readout of BMP pathway activity without requiring absolute molecular calibration.
In zebrafish and Drosophila embryos, pSmad/pMad immunostaining is done at various stages of development. In zebrafish embryos, analysis relies almost exclusively on fixed immunostaining, due to rapid gradient formation (3-9 hours post-fertilization) that governs the dorsoventral patterning. Within this limited time frame, the embryo is developing from hundreds to thousands of cells, which makes live tracking and imaging of BMP signaling technically challenging. Although live imaging for other transcription factors have been targeted, pSmad immunostaining remains the most robust and widely favored BMP-quantification method in zebrafish embryos [36,114,115,116].
In Drosophila, pMad immunostaining with pMad1/5 antibody is broadly applied across germline stem cells (GSC-preCB), ovaries, third instar larvae, pupal wing, and wing discs to approximate BMP signal transduction in both wild-type and mutant contexts [28,59,117,118]. Immunostaining ovaries dissected from 3-5 day old adult flies with pSmad1/5 antibody has facilitated quantification of pMad asymmetry during the G1/S phase of GSC division [28]. Likewise, in the embryos, the nuclear pMad domain typically spans 5–6 cell diameters, and that in sog mutants, uniform distribution of dpp in the dorsal region resulted in a broadened pMad pattern spanning 10–13 cells diameters [39,40,42].
More recently, live imaging approaches have begun to complement fixed pMad measurements. In Drosophila embryos, nuclear accumulation of Smad4/Medea has been quantified in vivo, and reveals a strong correlation between pMad levels and nuclear Med-GFP intensity [17]. These findings suggest that nuclear Medea can be used to serve as a live proxy for BMP activity [17,18].
Because pSmad/pMad patterns encode both spatial and temporal features of BMP signaling, nearly all computational efforts—across intracellular, tissue-level, and multi-scale modeling frameworks—aim to reproduce or predict these dynamics. Thus, pSmad/pMad is not only a measurement tool but also the central benchmark for linking experimental datasets with diverse modeling frameworks to validate mechanistic and data-driven models of BMP–SMAD regulation, as discussed in earlier sections.
Computational Modelling Approaches that Drive Mechanical Insights and Cross-Species Insights
Building on the system-specific models reviewed above, including pattern-formation models, bistability and gene-regulatory network (GRN) models, feedback-control frameworks, and source–sink transport models, the computational approach can be summarized into several major categories: Intracellular ODE models capture receptor activation, Smad phosphorylation, nucleocytoplasmic shuttling, and feedback loops, explaining gene-regulatory mechanisms such as bistability in stem cell systems and signal amplification or buffering observed across species [28,59,60,83,117]. PDE models based on the reaction–diffusion equation extend these insights to the tissue scale, showing how extracellular transport, inhibitor gradients, ligand shuttling, and transcriptionally induced modulators shape the pSmad/pMad patterns underlying dorsoventral patterning, germline maintenance, wing growth control, and self-organization in human pluripotent stem cells [41,42,43,63,68,69,72]. Complementing these frameworks, agent-based and GRN models integrate cell–cell interactions, spatial neighborhood effects, and transcriptional networks to explain how local signaling generates colony-scale or tissue-scale pSmad/pMad heterogeneity [73]. Stochastic models highlight how intrinsic noise in ligand–receptor binding or transcription contributes to variability in pSmad/pMad readouts [83,119]. Multi-scale and mechanochemical models combine intracellular biochemistry with tissue geometry, cell flow, or morphogenesis, reflecting the complex environments in which BMP gradients are interpreted [43,69,75]. To place these system-specific models in a unifying framework, Figure 6 summarizes the dominant mechanisms of BMP gradient formation, SMAD signal transduction, and gene regulation across all reviewed systems, and Table 2 catalogs the corresponding computational models.
These modeling strategies not only uncover the mechanistic principles driving BMP–SMAD patterning arises from the interplay of feedback regulation, extracellular transport, tissue geometry, and noise, producing robust and self-organized signaling landscapes across diverse biological systems but also provide a common analytical language for comparing signaling behavior across the six systems in this review. By enabling parameter inference, model transfer, and cross-species prediction, computational approaches facilitate a translational perspective in which conserved pathway logic can be distinguished from system-specific regulatory features. This perspective is exemplified by the “Digital Cousins” framework, which demonstrates how a single mechanistic model, appropriately parameterized, can capture BMP signaling behavior across biologically distinct systems and reveal shared design constraints despite anatomical and molecular differences [120]. These integration approaches reveal how a conserved BMP–SMAD signaling module gives rise to diverse developmental outcomes across organisms, tissues, and scales.
Figure 7.
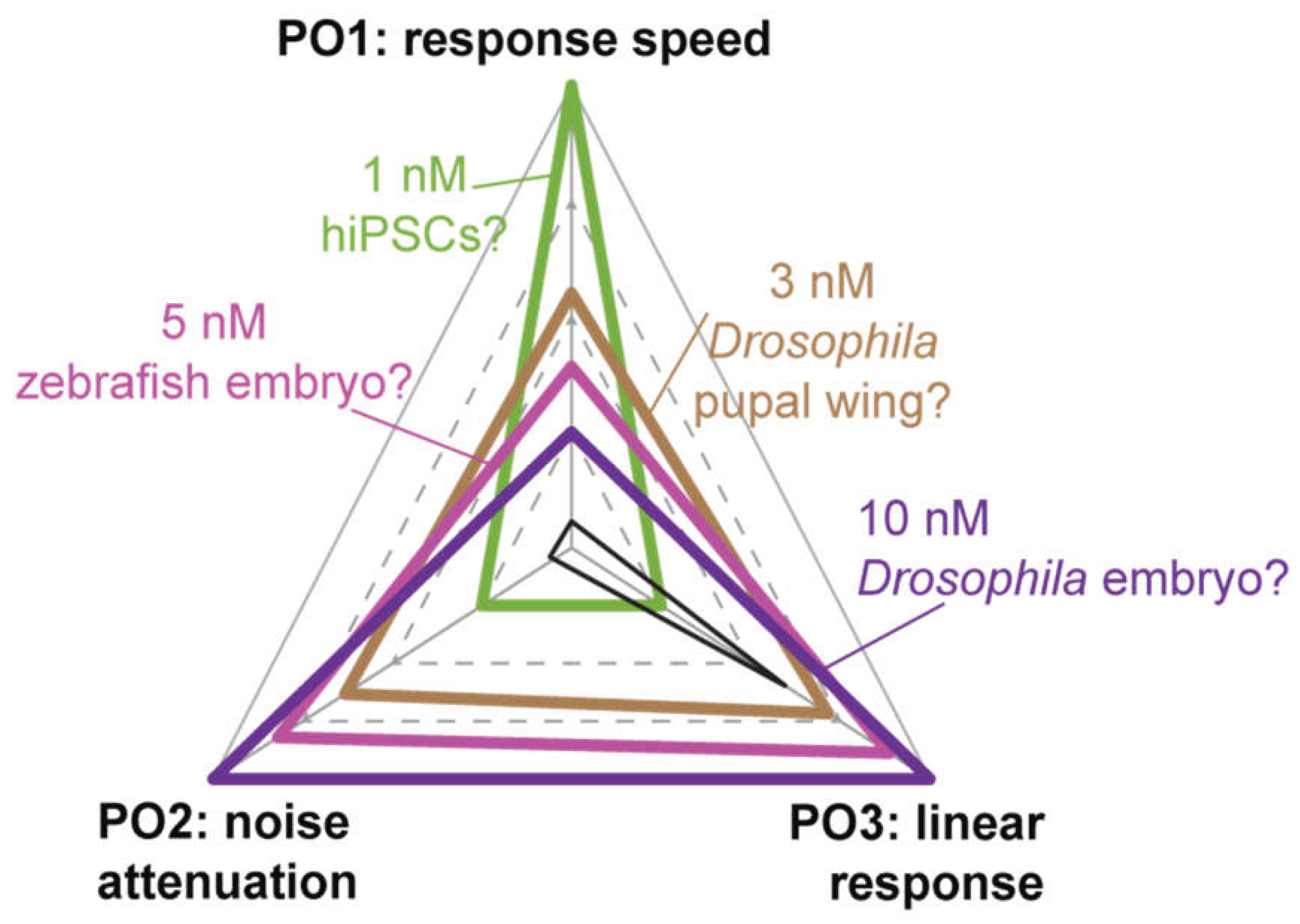
Simulation results from Shaikh et al. (2024) show how the balance of the POs changes when Phosphatase concentration is varied from 1 nM to 10 nM. In general, increasing Phosphatase concentration decreases response time and increases linear sensitivity while sacrificing noise reduction. Embryo systems potentially favor response speed and linearity, while differentiating organs and stem cells favor noise reduction. The “utopian ideal”, in which all three POs are optimized, is shown in black.
Figure 7.
Simulation results from Shaikh et al. (2024) show how the balance of the POs changes when Phosphatase concentration is varied from 1 nM to 10 nM. In general, increasing Phosphatase concentration decreases response time and increases linear sensitivity while sacrificing noise reduction. Embryo systems potentially favor response speed and linearity, while differentiating organs and stem cells favor noise reduction. The “utopian ideal”, in which all three POs are optimized, is shown in black.
Pathway level Performance Objectives in System-Specific Contexts
Despite the highly conserved nature of the BMP/Smad pathway across various organisms, its functional behavior varies strikingly across developmental and cellular contexts, particularly in response speed, noise filtering capacity, and output sensitivity to pathway inputs [60]. These properties have been proposed to represent competing performance objectives (POs) that are optimized differently in each system, depending on context-specific factors such as the nuclear import rate of the Smad complex and the intracellular concentrations of Smad proteins and phosphatases. Importantly, this diversity in pathway behavior does not arise from rewiring the core signaling network or altering the biochemical functions of its components, as these features are highly conserved throughout the animal kingdom. Instead, tunable system-level behaviors emerge from variations in non-conserved parameters (NCPs), such as the concentrations of pathway components, with cellular phosphatase levels exerting especially strong influence in a computational model of the Smad pathways [37]. These system-level behaviors can be quantitatively characterized by three minimal performance objectives: (1) Response speed, indicated by the rise time (trise), defined as the duration taken by the Smad signaling complex to reach 95% of its final steady state; (2) Noise filtering, indicated by the Noise Attenuation Ratio (NAR), defined as the ratio of the coefficient of variation of the response signal to that of the input signal; and (3) Output sensitivity, indicated by the steady-state sensitivity coefficient, defined as the ratio of the fractional change in steady-state output to that of the input.
The balance among these performance objectives is system-specific, with each developmental or cellular context emphasizing different trade-offs. For example, in Drosophila germline stem cells, using the GSC cell-division model, Shaikh and Reeves (2024) analyzed rise time, a key performance metric for response time in both wild-type and dad knockout (dadKO) GSCs. They found that the rise time of Fused/Smurf in the preCB is higher in dad knocked-out simulations than in wild-type (59,60,83). Because precise control of differentiation must occur within the division time frame, the rise time of Fused is expected to be tightly regulated. In dadKO, the concentration profile takes significantly longer to respond and reach a steady state, which is in agreement with delayed differentiation observed in dad mutants [22]. Furthermore, it has been hypothesized that Dad confers robustness to GSC signaling by enabling adaptation to environmental and intrinsic variability, both across germaria and among GSCs within the same germarium. Fused/Smurf makes the system robust by ensuring that the CBs have a low value of pMad. In dadKO ovaries, the pMad levels in GSCs would be higher, which has been hypothesized to make the pMad levels in CB higher. This loss of noise attenuation is consistent with the delayed differentiation phenotype and reduced GSC number [22,62]. Similarly, a hypomorphic Dullard allele and a Dullard RNAi line exhibit similar levels of pMad throughout the G1/S phase as opposed to an asymmetric distribution of pMad in the wild-type tissues [28]. This highlights the effect of the phosphatase, Dullard, in conferring robustness in GSCs.
BMP signaling in the early Drosophila embryo also prioritizes rapid activation. In a two-three-hour old Drosophila embryo, BMP signaling patterns the dorsal half of the embryo [121,122]. Initial Smad signaling is broad and weak on the dorsal 25-30% of the embryo. About 30 minutes later, the Smad signal intensifies and sharpens to a narrow domain in the dorsal-most 5-10% of the embryo, which illustrates the rapid response time required in the embryonic Drosophila Smad network [8,17,123,124].
Later in Drosophila development, BMP controls robust and highly precise cell differentiation during vein differentiation in the pupal wing. Yan et al. (2009) demonstrated that synchronized activation of BMP and EGFR signaling arises from a bistable positive feedback loop driven by mutual activation of ligand production [101]. The bistable regime requires that both BMP and EGFR signaling act on similar timescales of tens of minutes. Furthermore, single pulses of constitutively active Type I BMP receptor Thickveins resulted in stimulation of both BMP and EGFR signaling activity, but prolonged ectopic stimulation decreases ectopic vein formation. This suggests the presence of a negative feedback mechanism operating on longer, hour-scale timescales that limits the spatial spread of vein differentiation. Precise free vein patterning, therefore, requires fine-tuning of the BMP response time, fast enough to engage positive feedback for vein induction, yet sufficiently delayed allowing longer-term negative feedback to suppress extraneous and noisy differentiation.
In zebrafish, the dorsoventral axis is patterned during embryogenesis, more specifically, late blastula and early gastrula stages, of the embryo within 3.5-8 hours post fertilization (HPF) [125,126,127]. The BMP signaling gradient forms with remarkable speed, establishing a consistent nuclear-pSmad profile that can be immunostained, visualized, and quantified as the overall output of the signaling network. This rapid and reproducible gradient formation reflects a system optimized for fast response and temporal precision. Quantitative analyses across mutant lines have revealed how such fast patterning is coupled with developmental robustness, as the network maintains consistent outputs despite perturbations [36]. Another common method of assessing BMP activity is to categorize the 24-HPF embryo phenotype. BMP drives ventral tissue differentiation, and phenotypes have been categorized with overactive BMP lines as V1 to V4, with increasing ventralization, and lines with diminished BMP activity as C1 to C5, being dorsalized [128,129]. At the same time, the signaling gradient develops rapidly across multiple layers of signaling. The mechanisms of this cascade that provide robustness and noise attenuation have proven difficult to untangle. Greenfeld and Mullins (2021) found that BMP gene activation relied more heavily on threshold levels rather than a strict gradient shape or exposure duration [14]. Larson et. al. (2025) further suggest that noise attenuation arises from the BMP heterodimer-receptor interactions and low-pass filtering behavior downstream, uncovered with stochastic signaling [119]. Consistent with this, Shaikh and Reeves (2024) found that trade-offs in performance objectives between systems highlighted response time in rapidly developing systems such as the zebrafish embryo, related especially to downstream phosphatase levels [60].
Discussion & Open Questions
In Drosophila, zebrafish, and hPSCs, there is a remarkable conservation of core components of the SMAD pathway, contrasted with a striking diversity in regulatory strategies. By comparing the roles of BMP and its orthologs, we can understand how different organisms have adapted this fundamental signaling network to regulate cell differentiation through comparison of the functional roles of BMP and its orthologs in diverse systems. For instance, in Drosophila embryos BMP gradient formation depends on a 'shuttling' mechanism involving Sog and Tld [24], while the wing disc primarily relies on restricted extracellular diffusion to spread Dpp [45]. Whereas zebrafish establish their BMP gradient via a 'source-sink’, and feedback loops, crosstalks with other signaling pathways and extracellular modulators help stabilize and refine the gradient [63,68]. Moreover, the interpretation of this signal is not only determined by spatial gradients but is strongly influenced by signaling dynamics and precisely tailored to the biological context. For instance, the Drosophila embryo decodes through a dynamic and temporally integrated response to determine downstream gene expression, rather than instantaneous concentration alone. The dynamic decoding activating a tiered (Type I-III) gene response to specify multiple cells fates across the dorsoventral axis [17,39,79,80,130], with receptor–ligand binding kinetics and feedback regulation contributing to the robustness of BMP signaling [41,42]. In contrast, the GSC niche uses the pathway as a switch: high BMP signaling represses the differentiation gene bam to maintain stemness, a steep transition that is spatially defined by asymmetric division and sharpened by negative feedback from the inhibitory Smad, Dad.
These diverse interpretive strategies all showcase the use of complex feedback networks to ensure the robustness of developmental patterning, while simultaneously having functional behavior that depends on cellular and tissue-level context. This context-dependency means each system must balance competing Performance Objectives (POs) [60]. For instance, the Drosophila GSC niche optimizes for noise filtering to create a sharp, binary "off" signal, potentially at the cost of a slower response. In contrast, the early Drosophila and zebrafish embryos prioritize rapid response times to meet tight developmental schedules, achieving stability and precision through means such as threshold-based interpretation and extracellular motifs [8,88,123,124,131]. Identifying how these trade-offs are balanced is a key knowledge gap. However, attempts to quantitatively formalize the trade-offs among multiple competing objectives across model systems, not to mention specifically manipulating these competing objectives across different model systems, are still lacking, hindering the discovery of generalized biological rules [132]. Bridging this gap will demand precise, quantitative access to the Smad pathway's inputs and outputs. This is essential for measuring key network response dynamics, such as response time and noise attenuation, and for mapping the quantitative relationship between BMP signaling and nuclear pSmad activity, the essential conserved intracellular proxy, including the underlying biophysical interactions.
The BMP/Smad pathway is an ideal system to pioneer this quantitative, cross-species approach. Modeling has already proven invaluable for describing gradient formation, resolving conflicting hypotheses like long-range Dpp transport, and understanding how complex, emergent properties arise. The pathway's power as a model system arises from a central dichotomy: its core topology and protein functions are highly conserved, while the protein concentrations and the parameters they control, like dephosphorylation and nuclear transport rates, are non-conserved and highly context-dependent [3,133,134,135]. This presents an opportunity to leverage this conserved architecture to build a predictive, cross-species computational model. Such a framework would overcome the limitations of siloed, ad-hoc studies [136]. By integrating experimental data from Drosophila, zebrafish, and hPSCs, a cross-species model could predict differences in the balance of PO trade-offs and identify the specific non-conserved parameters that matter most. Validation of this model, along with a unified toolkit of characterized BMP tools, would allow discoveries in one system to immediately and directly inform our knowledge of others, enabling a more holistic understanding of the regulatory features that drive context-specific cell behavior.
Computational models of the BMP–Smad signaling pathway offer powerful tools for cross-species optimization and translational prediction. As quantitative datasets accumulate across Drosophila, zebrafish, and human pluripotent stem cells systems, it is becoming feasible to build multi-objective models that identify conserved design principles while capturing species-specific constraints. Such “cross-species digital cousins” can help determine which aspects of signaling dynamics, like gradient shape, noise buffering, and temporal encoding, are functionally conserved and therefore predictive in translational contexts [120]. These models also create a path toward computational drug screening that leverages variation in signaling pathways and mechanisms to evaluate pathway perturbations across multiple biological backgrounds before moving into human systems, especially in cancer treatment. While emerging AI and machine-learning approaches—including physics-informed neural networks, generative models of signaling dynamics, and AI-accelerated parameter inference—are transforming the scale at which these models can be trained, optimized, and validated. Together, these advances point toward a future in which mechanistic modeling and AI are tightly integrated to provide predictive, cross-species, and ultimately clinically actionable.
References
- Donovan, P.J.; Gearhart, J. The end of the beginning for pluripotent stem cells. Nature 2001, 414, 92–97. [Google Scholar] [CrossRef]
- Itoh, F.; Asao, H.; Sugamura, K.; Heldin, C.; Dijke, P.T.; Itoh, S. Promoting bone morphogenetic protein signaling through negative regulation of inhibitory Smads. EMBO J. 2001, 20, 4132–4142. [Google Scholar] [CrossRef]
- Padgett, R.W.; Wozney, J.M.; Gelbart, W.M. Human BMP sequences can confer normal dorsal-ventral patterning in the Drosophila embryo. Proc. Natl. Acad. Sci. 1993, 90, 2905–2909. [Google Scholar] [CrossRef] [PubMed]
- Raftery, L.A.; Sutherland, D.J. TGF-β Family Signal Transduction in Drosophila Development: From Mad to Smads. Dev. Biol. 1999, 210, 251–268. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, C.; Zuñiga, A.; Hanna, P.; Hodar, C.; Gonzalez, M.; Cambiazo, V. Target genes of Dpp/BMP signaling pathway revealed by transcriptome profiling in the early D. melanogaster embryo. Gene 2016, 591, 191–200. [Google Scholar] [CrossRef]
- Minami, M.; Kinoshita, N.; Kamoshida, Y.; Tanimoto, H.; Tabata, T. brinker is a target of Dpp in Drosophila that negatively regulates Dpp-dependent genes. Nature 1999, 398, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Sturtevant, M.A.; Biehs, B.; François, V.; Padgett, R.W.; Blackman, R.K.; Bier, E. The Drosophila decapentaplegic and short gastrulation genes function antagonistically during adult wing vein development. Development 1996, 122, 4033–4044. [Google Scholar] [CrossRef]
- O’Connor, MB; Umulis, D; Othmer, HG; Blair, SS. Shaping BMP morphogen gradients in the Drosophila embryo and pupal wing. Development 2006, 133(2), 183–93. [Google Scholar] [CrossRef]
- Matsuda, S.; Shimmi, O. Directional transport and active retention of Dpp/BMP create wing vein patterns in Drosophila. Dev. Biol. 2012, 366, 153–162. [Google Scholar] [CrossRef]
- Blair, S.S. Wing Vein Patterning in Drosophila and the Analysis of Intercellular Signaling. Annu. Rev. Cell Dev. Biol. 2007, 23, 293–319. [Google Scholar] [CrossRef]
- Wasserman, J.D.; Urban, S.; Freeman, M. A family of rhomboid-like genes: Drosophila rhomboid-1 and roughoid/rhomboid-3 cooperate to activate EGF receptor signaling. Genes Dev. 2000, 14, 1651–1663. [Google Scholar] [CrossRef]
- Montagne, J.; Groppe, J.; Guillemin, K.; Krasnow, M.A.; Gehring, W.J.; Affolter, M. The Drosophila Serum Response Factor gene is required for the formation of intervein tissue of the wing and is allelic to blistered. Development 1996, 122, 2589–2597. [Google Scholar] [CrossRef]
- Wilcockson, S.G.; Ashe, H.L. Drosophila Ovarian Germline Stem Cell Cytocensor Projections Dynamically Receive and Attenuate BMP Signaling. Dev. Cell 2019, 50, 296–312.e5. [Google Scholar] [CrossRef] [PubMed]
- Greenfeld, H.; Lin, J.; Mullins, M.C. The BMP signaling gradient is interpreted through concentration thresholds in dorsal–ventral axial patterning. PLOS Biol. 2021, 19, e3001059. [Google Scholar] [CrossRef] [PubMed]
- Rogers, K.W.; ElGamacy, M.; Jordan, B.M.; Müller, P. Optogenetic investigation of BMP target gene expression diversity. eLife 2020, 9. [Google Scholar] [CrossRef]
- Sánchez-Duffhues, G.; Hiepen, C. Human iPSCs as Model Systems for BMP-Related Rare Diseases. Cells 2023, 12, 2200. [Google Scholar] [CrossRef] [PubMed]
- Al Asafen, H.; Beseli, A.; Chen, H.-Y.; Hiremath, S.; Williams, C.M.; Reeves, G.T. Dynamics of BMP signaling and stable gene expression in the early Drosophila embryo. Biol. Open 2024, 13. [Google Scholar] [CrossRef]
- Brantley, SE; Janssen, J; Chao, A; Vergassola, M; Blythe, SA; Di Talia, S. Rapid transcriptional response to a dynamic morphogen by time integration [Internet]. Developmental Biology. Available from. 2025. http://biorxiv.org/lookup/doi/10.1101/2025.06.09.658715.
- A Teleman, A.; Cohen, S.M. Dpp Gradient Formation in the Drosophila Wing Imaginal Disc. Cell 2000, 103, 971–980. [Google Scholar] [CrossRef]
- Ralston, A.; Blair, S.S. Long-range Dpp signaling is regulated to restrict BMP signaling to a crossvein competent zone. Dev. Biol. 2005, 280, 187–200. [Google Scholar] [CrossRef]
- Gui, J.; Huang, Y.; Montanari, M.; Toddie-Moore, D.; Kikushima, K.; Nix, S.; Ishimoto, Y.; Shimmi, O. Coupling between dynamic 3D tissue architecture and BMP morphogen signaling during Drosophila wing morphogenesis. Proc. Natl. Acad. Sci. 2019, 116, 4352–4361. [Google Scholar] [CrossRef]
- Xie, T.; Spradling, A.C. decapentaplegic Is Essential for the Maintenance and Division of Germline Stem Cells in the Drosophila Ovary. Cell 1998, 94, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Madamanchi, A.; Mullins, M.C.; Umulis, D.M. Diversity and robustness of bone morphogenetic protein pattern formation. Development 2021, 148. [Google Scholar] [CrossRef]
- Mizutani, C.M.; Nie, Q.; Wan, F.Y.; Zhang, Y.-T.; Vilmos, P.; Sousa-Neves, R.; Bier, E.; Marsh, J.L.; Lander, A.D. Formation of the BMP Activity Gradient in the Drosophila Embryo. Dev. Cell 2005, 8, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Loh, K.M.; Chen, A.; Koh, P.W.; Deng, T.Z.; Sinha, R.; Tsai, J.M.; Barkal, A.A.; Shen, K.Y.; Jain, R.; Morganti, R.M.; et al. Mapping the Pairwise Choices Leading from Pluripotency to Human Bone, Heart, and Other Mesoderm Cell Types. Cell 2016, 166, 451–467. [Google Scholar] [CrossRef]
- Kamiya, N.; Ye, L.; Kobayashi, T.; Mochida, Y.; Yamauchi, M.; Kronenberg, H.M.; Feng, J.Q.; Mishina, Y. BMP signaling negatively regulates bone mass through sclerostin by inhibiting the canonical Wnt pathway. Development 2008, 135, 3801–3811. [Google Scholar] [CrossRef] [PubMed]
- Urrutia, H; Aleman, A; Eivers, E. Drosophila Dullard functions as a Mad phosphatase to terminate BMP signaling. Sci Rep. 2016, 6(1), 32269. [Google Scholar] [CrossRef]
- Sardi, J.; Bener, M.B.; Simao, T.; Descoteaux, A.E.; Slepchenko, B.M.; Inaba, M. Mad dephosphorylation at the nuclear pore is essential for asymmetric stem cell division. Proc. Natl. Acad. Sci. 2021, 118. [Google Scholar] [CrossRef]
- Tanaka, S.S.; Nakane, A.; Yamaguchi, Y.L.; Terabayashi, T.; Abe, T.; Nakao, K.; Asashima, M.; Steiner, K.A.; Tam, P.P.L.; Nishinakamura, R. Dullard/Ctdnep1 Modulates WNT Signalling Activity for the Formation of Primordial Germ Cells in the Mouse Embryo. PLOS ONE 2013, 8, e57428. [Google Scholar] [CrossRef]
- Podos, S.D.; Hanson, K.K.; Wang, Y.-C.; Ferguson, E.L. The DSmurf Ubiquitin-Protein Ligase Restricts BMP Signaling Spatially and Temporally during Drosophila Embryogenesis. Dev. Cell 2001, 1, 567–578. [Google Scholar] [CrossRef]
- Piacentino, M.L.; E Bronner, M. Intracellular attenuation of BMP signaling via CKIP-1/Smurf1 is essential during neural crest induction. PLOS Biol. 2018, 16, e2004425. [Google Scholar] [CrossRef]
- Murakami, K.; Etlinger, J.D. Role of SMURF1 ubiquitin ligase in BMP receptor trafficking and signaling. Cell. Signal. 2019, 54, 139–149. [Google Scholar] [CrossRef]
- Akiyama, T.; A Raftery, L.; A Wharton, K. Bone morphogenetic protein signaling: the pathway and its regulation. Genetics 2023, 226. [Google Scholar] [CrossRef]
- Antebi, Y.E.; Linton, J.M.; Klumpe, H.; Bintu, B.; Gong, M.; Su, C.; McCardell, R.; Elowitz, M.B. Combinatorial Signal Perception in the BMP Pathway. Cell 2017, 170, 1184–1196.e24. [Google Scholar] [CrossRef]
- Klumpe, H.E.; Langley, M.A.; Linton, J.M.; Su, C.J.; Antebi, Y.E.; Elowitz, M.B. The context-dependent, combinatorial logic of BMP signaling. Cell Syst. 2022, 13, 388–407.e10. [Google Scholar] [CrossRef]
- Zinski, J.; Tuazon, F.; Huang, Y.; Mullins, M.; Umulis.
- Schmierer, B.; Tournier, A.L.; Bates, P.A.; Hill, C.S. Mathematical modeling identifies Smad nucleocytoplasmic shuttling as a dynamic signal-interpreting system. Proc. Natl. Acad. Sci. 2008, 105, 6608–6613. [Google Scholar] [CrossRef] [PubMed]
- Heemskerk, I.; Burt, K.; Miller, M.; Chhabra, S.; Guerra, M.C.; Liu, L.; Warmflash, A. Rapid changes in morphogen concentration control self-organized patterning in human embryonic stem cells. eLife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.J.; Shimmi, O.; Vilmos, P.; Petryk, A.; Kim, H.; Gaudenz, K.; Hermanson, S.; Ekker, S.C.; O'COnnor, M.B.; Marsh, J.L. Twisted gastrulation is a conserved extracellular BMP antagonist. Nature 2001, 410, 479–483. [Google Scholar] [CrossRef]
- Dorfman, R.; Shilo, B.-Z. Biphasic activation of the BMP pathway patterns the Drosophila embryonic dorsal region. Development 2001, 128, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Umulis, D.M.; Serpe, M.; O’connor, M.B.; Othmer, H.G. Robust, bistable patterning of the dorsal surface of the Drosophila embryo. Proc. Natl. Acad. Sci. 2006, 103, 11613–11618. [Google Scholar] [CrossRef]
- Wang, Y.-C.; Ferguson, E.L. Spatial bistability of Dpp–receptor interactions during Drosophila dorsal–ventral patterning. Nature 2005, 434, 229–234. [Google Scholar] [CrossRef]
- Umulis, D.M.; Shimmi, O.; O'COnnor, M.B.; Othmer, H.G. Organism-Scale Modeling of Early Drosophila Patterning via Bone Morphogenetic Proteins. Dev. Cell 2010, 18, 260–274. [Google Scholar] [CrossRef]
- Matsuda, S.; Affolter, M. Is Drosophila Dpp/BMP morphogen spreading required for wing patterning and growth? BioEssays 2023, 45, e2200218. [Google Scholar] [CrossRef] [PubMed]
- Schwank, G.; Dalessi, S.; Yang, S.-F.; Yagi, R.; de Lachapelle, A.M.; Affolter, M.; Bergmann, S.; Basler, K. Formation of the Long Range Dpp Morphogen Gradient. PLOS Biol. 2011, 9, e1001111. [Google Scholar] [CrossRef]
- Restrepo, S.; Zartman, J.J.; Basler, K. Coordination of Patterning and Growth by the Morphogen DPP. Curr. Biol. 2014, 24, R245–R255. [Google Scholar] [CrossRef]
- Fancher, S.; Mugler, A. Diffusion vs. direct transport in the precision of morphogen readout. eLife 2020, 9. [Google Scholar] [CrossRef]
- Matsuda, S.; Harmansa, S.; Affolter, M. BMP morphogen gradients in flies. Cytokine Growth Factor Rev. 2016, 27, 119–127. [Google Scholar] [CrossRef]
- Ben-Zvi, D.; Barkai, N. Scaling of morphogen gradients by an expansion-repression integral feedback control. Proc. Natl. Acad. Sci. 2010, 107, 6924–6929. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Qiu, Y.; Chen, W.; Nie, Q.; Lander, A.D. Scaling a Dpp Morphogen Gradient through Feedback Control of Receptors and Co-receptors. Dev. Cell 2020, 53, 724–739.e14. [Google Scholar] [CrossRef] [PubMed]
- Romanova-Michaelides, M.; Hadjivasiliou, Z.; Aguilar-Hidalgo, D.; Basagiannis, D.; Seum, C.; Dubois, M.; Jülicher, F.; Gonzalez-Gaitan, M. Morphogen gradient scaling by recycling of intracellular Dpp. Nature 2021, 602, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Serpe, M.; Umulis, D.; Ralston, A.; Chen, J.; Olson, D.J.; Avanesov, A.; Othmer, H.; O'COnnor, M.B.; Blair, S.S. The BMP-Binding Protein Crossveinless 2 Is a Short-Range, Concentration-Dependent, Biphasic Modulator of BMP Signaling in Drosophila. Dev. Cell 2008, 14, 940–953. [Google Scholar] [CrossRef]
- Antson, H.; Tõnissoo, T.; Shimmi, O. The developing wing crossvein of Drosophila melanogaster: a fascinating model for signaling and morphogenesis. Fly 2022, 16, 118–127. [Google Scholar] [CrossRef]
- Serpe, M.; Ralston, A.; Blair, S.S.; O'Connor, M.B. Matching catalytic activity to developmental function: Tolloid-related processes Sog in order to help specify the posterior crossvein in theDrosophilawing. Development 2005, 132, 2645–2656. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, S; Schaefer, JV; Mii, Y; Hori, Y; Bieli, D; Taira, M; et al. Asymmetric requirement of Dpp/BMP morphogen dispersal in the Drosophila wing disc. Nat Commun. 2021, 12(1), 6435. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, Z.; Cai, Y. The JAK/STAT pathway positively regulates DPP signaling in the Drosophila germline stem cell niche. J. Cell Biol. 2008, 180, 721–728. [Google Scholar] [CrossRef]
- Wang, X.; Page-McCaw, A. Wnt6 maintains anterior escort cells as an integral component of the germline stem cell niche. Development 2018, 145. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Jia, S.; Huang, S.; Wang, H.; Zhu, Y.; Mu, Y.; Kan, L.; Zheng, W.; Wu, D.; Li, X.; et al. The Fused/Smurf Complex Controls the Fate of Drosophila Germline Stem Cells by Generating a Gradient BMP Response. Cell 2010, 143, 978–990. [Google Scholar] [CrossRef]
- Xia, L.; Zheng, X.; Zheng, W.; Zhang, G.; Wang, H.; Tao, Y.; Chen, D. The Niche-Dependent Feedback Loop Generates a BMP Activity Gradient to Determine the Germline Stem Cell Fate. Curr. Biol. 2012, 22, 515–521. [Google Scholar] [CrossRef]
- Shaikh, R.; Larson, N.J.; Kam, J.; Hanjaya-Putra, D.; Zartman, J.; Umulis, D.M.; Li, L.; Reeves, G.T. Optimal performance objectives in the highly conserved bone morphogenetic protein signaling pathway. npj Syst. Biol. Appl. 2024, 10, 1–13. [Google Scholar] [CrossRef]
- Casanueva, M.O.; Ferguson, E.L. Germline stem cell number in theDrosophilaovary is regulated by redundant mechanisms that control Dpp signaling. Development 2004, 131, 1881–1890. [Google Scholar] [CrossRef]
- Li, W.; Li, W.; Zou, L.; Ji, S.; Li, C.; Liu, K.; Zhang, G.; Sun, Q.; Xiao, F.; Chen, D. Membrane targeting of inhibitory Smads through palmitoylation controls TGF-β/BMP signaling. Proc. Natl. Acad. Sci. 2017, 114, 13206–13211. [Google Scholar] [CrossRef] [PubMed]
- Pomreinke, AP; Soh, GH; Rogers, KW; Bergmann, JK; Bläßle, AJ; Müller, P. Dynamics of BMP signaling and distribution during zebrafish dorsal-ventral patterning. eLife 2017, 6, e25861. [Google Scholar] [CrossRef]
- Connors, S.A.; Trout, J.; Ekker, M.; Mullins, M.C. The role of tolloid/mini fin in dorsoventral pattern formation of the zebrafish embryo. Development 1999, 126, 3119–3130. [Google Scholar] [CrossRef]
- Muraoka, O.; Shimizu, T.; Yabe, T.; Nojima, H.; Bae, Y.-K.; Hashimoto, H.; Hibi, M. Sizzled controls dorso-ventral polarity by repressing cleavage of the Chordin protein. Nat. Cell Biol. 2006, 8, 329–340. [Google Scholar] [CrossRef]
- Yabe, T.; Shimizu, T.; Muraoka, O.; Bae, Y.-K.; Hirata, T.; Nojima, H.; Kawakami, A.; Hirano, T.; Hibi, M. Ogon/Secreted Frizzled functions as a negative feedback regulator of Bmp signaling. Development 2003, 130, 2705–2716. [Google Scholar] [CrossRef]
- Rentzsch, F.; Zhang, J.; Kramer, C.; Sebald, W.; Hammerschmidt, M. Crossveinless 2 is an essential positive feedback regulator of Bmp signaling during zebrafish gastrulation. Development 2006, 133, 801–811. [Google Scholar] [CrossRef]
- Zinski, J.; Bu, Y.; Wang, X.; Dou, W.; Umulis, D.; Mullins, M.C. Systems biology derived source-sink mechanism of BMP gradient formation. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, X.; Mullins, M.C.; Umulis, D.M. Evaluation of BMP-mediated patterning in a 3D mathematical model of the zebrafish blastula embryo. J. Math. Biol. 2019, 80, 505–520. [Google Scholar] [CrossRef] [PubMed]
- Warmflash, A.; Sorre, B.; Etoc, F.; Siggia, E.D.; Brivanlou, A.H. A method to recapitulate early embryonic spatial patterning in human embryonic stem cells. Nat. Methods 2014, 11, 847–854. [Google Scholar] [CrossRef]
- Shao, Y.; Taniguchi, K.; Townshend, R.F.; Miki, T.; Gumucio, D.L.; Fu, J. A pluripotent stem cell-based model for post-implantation human amniotic sac development. Nat. Commun. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Etoc, F.; Metzger, J.; Ruzo, A.; Kirst, C.; Yoney, A.; Ozair, M.Z.; Brivanlou, A.H.; Siggia, E.D. A Balance between Secreted Inhibitors and Edge Sensing Controls Gastruloid Self-Organization. Dev. Cell 2016, 39, 302–315. [Google Scholar] [CrossRef]
- Kaul, H.; Werschler, N.; Jones, R.D.; Siu, M.M.; Tewary, M.; Hagner, A.; Ostblom, J.; Aguilar-Hidalgo, D.; Zandstra, P.W. Virtual cells in a virtual microenvironment recapitulate early development-like patterns in human pluripotent stem cell colonies. Stem Cell Rep. 2022, 18, 377–393. [Google Scholar] [CrossRef]
- Chhabra, S.; Liu, L.; Goh, R.; Kong, X.; Warmflash, A. Dissecting the dynamics of signaling events in the BMP, WNT, and NODAL cascade during self-organized fate patterning in human gastruloids. PLOS Biol. 2019, 17, e3000498. [Google Scholar] [CrossRef]
- Teague, S.; Primavera, G.; Chen, B.; Liu, Z.-Y.; Yao, L.; Freeburne, E.; Khan, H.; Jo, K.; Johnson, C.; Heemskerk, I. Time-integrated BMP signaling determines fate in a stem cell model for early human development. Nat. Commun. 2024, 15, 1–18. [Google Scholar] [CrossRef]
- Hanjaya-Putra, D.; Gerecht, S. Vascular engineering using human embryonic stem cells. Biotechnol. Prog. 2009, 25, 2–9. [Google Scholar] [CrossRef]
- Kusuma, S.; Shen, Y.-I.; Hanjaya-Putra, D.; Mali, P.; Cheng, L.; Gerecht, S. Self-organized vascular networks from human pluripotent stem cells in a synthetic matrix. Proc. Natl. Acad. Sci. 2013, 110, 12601–12606. [Google Scholar] [CrossRef]
- Ashe, H.L.; Mannervik, M.; Levine, M. Dpp signaling thresholds in the dorsal ectoderm of the Drosophila embryo. Development 2000, 127, 3305–3312. [Google Scholar] [CrossRef] [PubMed]
- Chahda, J.S.; Ambrosi, P.; Mizutani, C.M. The nested embryonic dorsal domains of BMP-target genes are not scaled to size during the evolution of Drosophila species. J. Exp. Zoöl. Part B: Mol. Dev. Evol. 2022, 340, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, C; Bowles, JR; Minchington, TG; Sutcliffe, C; Upadhyai, P; Rattray, M; et al. Modulation of the Promoter Activation Rate Dictates the Transcriptional Response to Graded BMP Signaling Levels in the Drosophila Embryo. Dev Cell 2020, 54(6), 727–741.e7. [Google Scholar] [PubMed]
- Song, X.; Wong, M.D.; Kawase, E.; Xi, R.; Ding, B.C.; McCarthy, J.J.; Xie, T. Bmp signals from niche cells directly repress transcription of a differentiation-promoting gene,bag of marbles, in germline stem cells in theDrosophilaovary. Development 2004, 131, 1353–1364. [Google Scholar] [CrossRef]
- Chen, D.; McKearin, D. Dpp Signaling Silences bam Transcription Directly to Establish Asymmetric Divisions of Germline Stem Cells. Curr. Biol. 2003, 13, 1786–1791. [Google Scholar] [CrossRef]
- Pargett, M.; Rundell, A.E.; Buzzard, G.T.; Umulis, D.M. Model-Based Analysis for Qualitative Data: An Application in Drosophila Germline Stem Cell Regulation. PLOS Comput. Biol. 2014, 10, e1003498. [Google Scholar] [CrossRef] [PubMed]
- Zinski, J.; Tajer, B.; Mullins, M.C. TGF-β Family Signaling in Early Vertebrate Development. Cold Spring Harb. Perspect. Biol. 2017, 10, a033274. [Google Scholar] [CrossRef]
- Grotewold, L.; Rüther, U. The Wnt antagonist Dickkopf-1 is regulated by Bmp signaling and c-Jun and modulates programmed cell death. EMBO J. 2002, 21, 966–975. [Google Scholar] [CrossRef]
- Lee, H.X.; Ambrosio, A.L.; Reversade, B.; De Robertis, E. Embryonic Dorsal-Ventral Signaling: Secreted Frizzled-Related Proteins as Inhibitors of Tolloid Proteinases. Cell 2006, 124, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Tuazon, F.B.; Wang, X.; Andrade, J.L.; Umulis, D.; Mullins, M.C. Proteolytic Restriction of Chordin Range Underlies BMP Gradient Formation. Cell Rep. 2020, 32, 108039–108039. [Google Scholar] [CrossRef]
- Ramel, M.-C.; Hill, C.S. The ventral to dorsal BMP activity gradient in the early zebrafish embryo is determined by graded expression of BMP ligands. Dev. Biol. 2013, 378, 170–182. [Google Scholar] [CrossRef]
- Hollnagel, A.; Oehlmann, V.; Heymer, J.; Rüther, U.; Nordheim, A. Id Genes Are Direct Targets of Bone Morphogenetic Protein Induction in Embryonic Stem Cells. J. Biol. Chem. 1999, 274, 19838–19845. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Hsiao, E.C.; Sami, S.; Lancero, M.; Schlieve, C.R.; Nguyen, T.; Yano, K.; Nagahashi, A.; Ikeya, M.; Matsumoto, Y.; et al. BMP-SMAD-ID promotes reprogramming to pluripotency by inhibiting p16/INK4A-dependent senescence. Proc. Natl. Acad. Sci. 2016, 113, 13057–13062. [Google Scholar] [CrossRef]
- Jen, Y.; Weintraub, H.; Benezra, R. Overexpression of Id protein inhibits the muscle differentiation program: in vivo association of Id with E2A proteins. Genes Dev. 1992, 6, 1466–1479. [Google Scholar] [CrossRef]
- Melnikova, I.N.; A Christy, B. Muscle cell differentiation is inhibited by the helix-loop-helix protein Id3. 1996, 7, 1067–79. [Google Scholar]
- Korchynskyi, O.; Dijke, P.T. Identification and Functional Characterization of Distinct Critically Important Bone Morphogenetic Protein-specific Response Elements in the Id1 Promoter. J. Biol. Chem. 2002, 277, 4883–4891. [Google Scholar] [CrossRef]
- Chen, T.; Wang, F.; Wu, M.; Wang, Z.Z. Development of Hematopoietic Stem and Progenitor Cells From Human Pluripotent Stem Cells. J. Cell. Biochem. 2015, 116, 1179–1189. [Google Scholar] [CrossRef] [PubMed]
- Fei, T.; Xia, K.; Li, Z.; Zhou, B.; Zhu, S.; Chen, H.; Zhang, J.; Chen, Z.; Xiao, H.; Han, J.-D.J.; et al. Genome-wide mapping of SMAD target genes reveals the role of BMP signaling in embryonic stem cell fate determination. Genome Res. 2009, 20, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Graham, F.; Knopp, J.; Patzke, C.; Hanjaya-Putra, D. Robust Differentiation of Human Pluripotent Stem Cells into Lymphatic Endothelial Cells Using Transcription Factors. Cells Tissues Organs 2024, 213, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, M.; James, D.; Ding, B.-S.; Nolan, D.; Geng, F.; Butler, J.M.; Schachterle, W.; Pulijaal, V.R.; Mathew, S.; Chasen, S.T.; et al. Efficient Direct Reprogramming of Mature Amniotic Cells into Endothelial Cells by ETS Factors and TGFβ Suppression. Cell 2012, 151, 559–575. [Google Scholar] [CrossRef]
- Imamura, T.; Takase, M.; Nishihara, A.; Oeda, E.; Hanai, J.-I.; Kawabata, M.; Miyazono, K. Smad6 inhibits signalling by the TGF-β superfamily. Nature 1997, 389, 622–626. [Google Scholar] [CrossRef]
- Ishida, W; Hamamoto, T; Kusanagi, K; Yagi, K; Kawabata, M; Takehara, K; et al. Smad6 Is a Smad1/5-induced Smad Inhibitor. J Biol Chem. 2000, 275(9), 6075–9. [Google Scholar] [CrossRef]
- Nakao, A; Afrakhte, M; Morn, A; Nakayama, T; Christian, JL; Heuchel, R; et al. Identification of Smad7, a TGFβ-inducible antagonist of TGF-β signalling. Nature 1997, 389(6651), 631–5. [Google Scholar] [CrossRef]
- Yan, X; Liu, Z; Chen, Y. Regulation of TGF-β signaling by Smad7. Acta Biochim Biophys Sin. 2009, 41(4), 263–72. [Google Scholar] [CrossRef]
- Onichtchouk, D; Chen, YG; Dosch, R; Gawantka, V; Delius, H; Massague’, J; et al. Silencing of TGF-β signalling by the pseudoreceptor BAMBI. Nature 1999, 401(6752), 480–5. [Google Scholar] [CrossRef]
- Grotewold, L.; Plum, M.; Dildrop, R.; Peters, T.; Rüther, U. Bambi is coexpressed with Bmp-4 during mouse embryogenesis. Mech. Dev. 2001, 100, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Guillot, N.; Kollins, D.; Gilbert, V.; Xavier, S.; Chen, J.; Gentle, M.; Reddy, A.; Bottinger, E.; Jiang, R.; Rastaldi, M.P.; et al. BAMBI Regulates Angiogenesis and Endothelial Homeostasis through Modulation of Alternative TGFβ Signaling. PLOS ONE 2012, 7, e39406. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, L.B.; De Jesús-Escobar, J.M.; Harland, R.M. The Spemann Organizer Signal noggin Binds and Inactivates Bone Morphogenetic Protein 4. Cell 1996, 86, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Piccolo, S.; Sasai, Y.; Lu, B.; De Robertis, E.M. Dorsoventral Patterning in Xenopus: Inhibition of Ventral Signals by Direct Binding of Chordin to BMP-4. Cell 1996, 86, 589–598. [Google Scholar] [CrossRef]
- Fainsod, A.; Deißler, K.; Yelin, R.; Marom, K.; Epstein, M.; Pillemer, G.; Steinbeisser, H.; Blum, M. The dorsalizing and neural inducing gene follistatin is an antagonist of BMP-4. Mech. Dev. 1997, 63, 39–50. [Google Scholar] [CrossRef]
- Gazzerro, E; Canalis, E. Bone morphogenetic proteins and their antagonists. Rev Endocr Metab Disord 2007, 7(1–2), 51–65. [Google Scholar] [CrossRef]
- Rider, C.C.; Mulloy, B. Bone morphogenetic protein and growth differentiation factor cytokine families and their protein antagonists. Biochem. J. 2010, 429, 1–12. [Google Scholar] [CrossRef]
- Kurata, T.; Nakabayashi, J.; Yamamoto, T.S.; Mochii, M.; Ueno, N. Visualization of endogenous BMP signaling during Xenopus development. Differentiation 2001, 67, 33–40. [Google Scholar] [CrossRef]
- Patterson, SE; Bird, NC; Devoto, SH. BMP regulation of myogenesis in zebrafish. Dev Dyn. 2010, 239(3), 806–17. [Google Scholar] [CrossRef]
- Ohta, S.; Mansour, S.L.; Schoenwolf, G.C. BMP/SMAD signaling regulates the cell behaviors that drive the initial dorsal-specific regional morphogenesis of the otocyst. Dev. Biol. 2010, 347, 369–381. [Google Scholar] [CrossRef]
- Kane, N.S.; Vora, M.; Padgett, R.W.; Li, Y. bantam microRNA is a negative regulator of the Drosophila decapentaplegic pathway. Fly 2018, 12, 105–117. [Google Scholar] [CrossRef]
- Laux, D.W.; Febbo, J.A.; Roman, B.L. Dynamic analysis of BMP-responsive smad activity in live zebrafish embryos. Dev. Dyn. 2011, 240, 682–694. [Google Scholar] [CrossRef] [PubMed]
- Gramann, A.K.; Venkatesan, A.M.; Guerin, M.; Ceol, C.J. Regulation of zebrafish melanocyte development by ligand-dependent BMP signaling. eLife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Mouillesseaux, K.P.; Wiley, D.S.; Saunders, L.; Wylie, L.A.; Kushner, E.J.; Chong, D.C.; Citrin, K.M.; Barber, A.T.; Park, Y.; Kim, J.-D.; et al. Notch regulates BMP responsiveness and lateral branching in vessel networks via SMAD6. Nat. Commun. 2016, 7, 13247. [Google Scholar] [CrossRef] [PubMed]
- Wilcockson, S.G.; Ashe, H.L. Live imaging of the Drosophila ovarian germline stem cell niche. STAR Protoc. 2021, 2, 100371. [Google Scholar] [CrossRef]
- O'KEefe, D.D.; Thomas, S.; Edgar, B.A.; Buttitta, L. Temporal regulation of Dpp signaling output in the Drosophila wing. Dev. Dyn. 2014, 243, 818–832. [Google Scholar] [CrossRef]
- Larson, NJ; Madamanchi, A; Li, L; Umulis, DM. Stochastic Modeling of BMP Heterodimer-Receptor Interactions Shows Emergence of Low-Pass Filtering Behavior [Internet]. Developmental Biology. Available from. 2025. http://biorxiv.org/lookup/doi/10.1101/2025.10.17.677951.
- Li, L.; Mdluli, T.; Buzzard, G.; Umulis, D. Digital cousins: Simultaneous optimization of one model for BMP signaling in distant relatives reveals essential core. Comput. Struct. Biotechnol. J. 2025, 27, 3729–3741. [Google Scholar] [CrossRef]
- Padgett, R.W.; Johnston, R.D.S.; Gelbart, W.M. A transcript from a Drosophila pattern gene predicts a protein homologous to the transforming growth factor-β family. Nature 1987, 325, 81–84. [Google Scholar] [CrossRef]
- Arora, K.; Levine, M.S.; O'COnnor, M.B. The screw gene encodes a ubiquitously expressed member of the TGF-beta family required for specification of dorsal cell fates in the Drosophila embryo. Genes Dev. 1994, 8, 2588–2601. [Google Scholar] [CrossRef]
- Raftery, AE; Zheng, Y. Discussion: Performance of Bayesian Model Averaging. J Am Stat Assoc. 2003, 98(464), 931–8. [Google Scholar] [CrossRef]
- Gavin-Smyth, J.; Wang, Y.-C.; Butler, I.; Ferguson, E.L. A Genetic Network Conferring Canalization to a Bistable Patterning System in Drosophila. Curr. Biol. 2013, 23, 2296–2302. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Handin, R.I. Identification and Characterization of Zebrafish Tissue Factor and Its Role in Embryonic Blood Vessel Development. Blood 2007, 110, 3706–3706. [Google Scholar] [CrossRef]
- Langdon, Y.G.; Mullins, M.C. Maternal and Zygotic Control of Zebrafish Dorsoventral Axial Patterning. Annu. Rev. Genet. 2011, 45, 357–377. [Google Scholar] [CrossRef]
- Pinheiro, D.; Heisenberg.
- Mullins, M.C.; Hammerschmidt, M.; Kane, D.A.; Odenthal, J.; Brand, M.; van Eeden, F.J.M.; Furutani-Seiki, M.; Granato, M.; Haffter, P.; Heisenberg, C.-P.; et al. Genes establishing dorsoventral pattern formation in the zebrafish embryo: the ventral specifying genes. Development 1996, 123, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, Y.; Lee, K.-H.; Zon, L.; Hammerschmidt, M.; Schulte-Merker, S. The molecular nature of zebrafish swirl: BMP2 function is essential during early dorsoventral patterning. Development 1997, 124, 4457–4466. [Google Scholar] [CrossRef] [PubMed]
- Miles, W.O.; Jaffray, E.; Campbell, S.G.; Takeda, S.; Bayston, L.J.; Basu, S.P.; Li, M.; Raftery, L.A.; Ashe, M.P.; Hay, R.T.; et al. Medea SUMOylation restricts the signaling range of the Dpp morphogen in the Drosophila embryo. Genes Dev. 2008, 22, 2578–2590. [Google Scholar] [CrossRef]
- Tucker, J.A.; Mintzer, K.A.; Mullins, M.C. The BMP Signaling Gradient Patterns Dorsoventral Tissues in a Temporally Progressive Manner along the Anteroposterior Axis. Dev. Cell 2008, 14, 108–119. [Google Scholar] [CrossRef]
- Holford, M.; Normark, B.B. Integrating the Life Sciences to Jumpstart the Next Decade of Discovery. Integr. Comp. Biol. 2021, 61, 1984–1990. [Google Scholar] [CrossRef]
- Attisano, L; Tuen Lee-Hoeflich, S. The Smads. Genome Biol. 2001, 2(8), reviews3010.1. [Google Scholar] [CrossRef] [PubMed]
- Holley, S.A.; Jackson, P.D.; Sasai, Y.; Lu, B.; De Robertis, E.M.; Hoffmann, F.M.; Ferguson, E.L. A conserved system for dorsal-ventral patterning in insects and vertebrates involving sog and chordin. Nature 1995, 376, 249–253. [Google Scholar] [CrossRef] [PubMed]
- A Holley, S.; Neul, J.L.; Attisano, L.; Wrana, J.L.; Sasai, Y.; O'COnnor, M.B.; De Robertis, E.M.; Ferguson, E.L. The Xenopus Dorsalizing Factor noggin Ventralizes Drosophila Embryos by Preventing DPP from Activating Its Receptor. Cell 1996, 86, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Frick, C.L.; Yarka, C.; Nunns, H.; Goentoro, L. Sensing relative signal in the Tgf-β/Smad pathway. Proc. Natl. Acad. Sci. 2017, 114, E2975–E2982. [Google Scholar] [CrossRef]
Figure 1.
Spatial patterning of BMP ligands is a common theme across development from Drosophila to human systems. The cells that produce BMP ligands are in parenthesis for each model system and illustrated in maroon. Examples include patterning in the Drosophila embryo (Dorsal half), larval (a stripe of cells situated at the boundary between the anterior and posterior compartments) and pupal wing anlage (wing veins), and the adult female germline stem cells (cap cells); Zebrafish embryo (ventral) and the human embryo (extraembryonic mesoderm). The figure highlights the diversity in length and timescales across developmental contexts.
Figure 1.
Spatial patterning of BMP ligands is a common theme across development from Drosophila to human systems. The cells that produce BMP ligands are in parenthesis for each model system and illustrated in maroon. Examples include patterning in the Drosophila embryo (Dorsal half), larval (a stripe of cells situated at the boundary between the anterior and posterior compartments) and pupal wing anlage (wing veins), and the adult female germline stem cells (cap cells); Zebrafish embryo (ventral) and the human embryo (extraembryonic mesoderm). The figure highlights the diversity in length and timescales across developmental contexts.
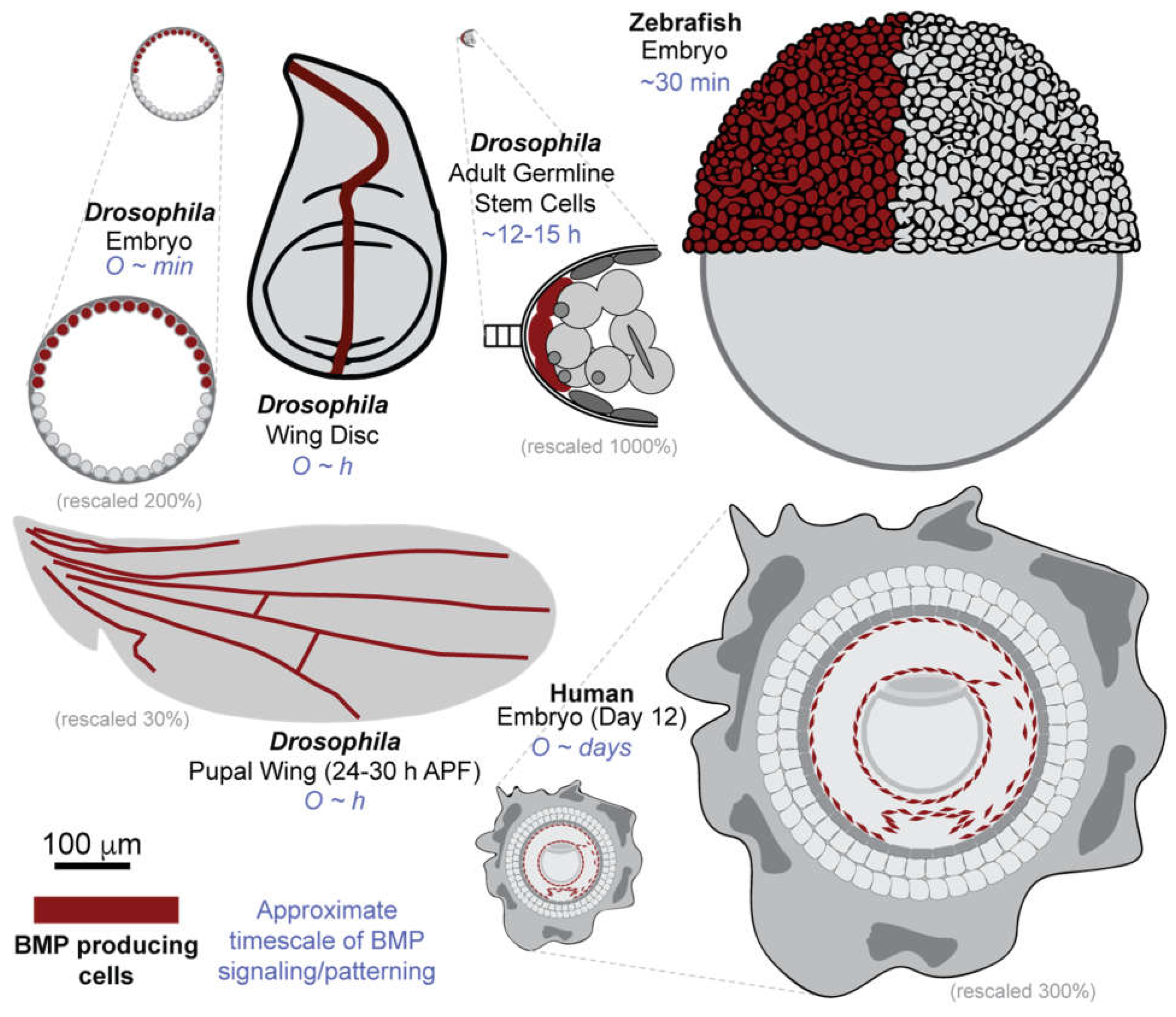
Figure 2.
The core conserved network of BMP signaling. This figure illustrates the canonical BMP signal transduction pathway topology and the core molecular players across three species: Drosophila melanogaster, Danio rerio, and Homo sapiens.
Figure 2.
The core conserved network of BMP signaling. This figure illustrates the canonical BMP signal transduction pathway topology and the core molecular players across three species: Drosophila melanogaster, Danio rerio, and Homo sapiens.
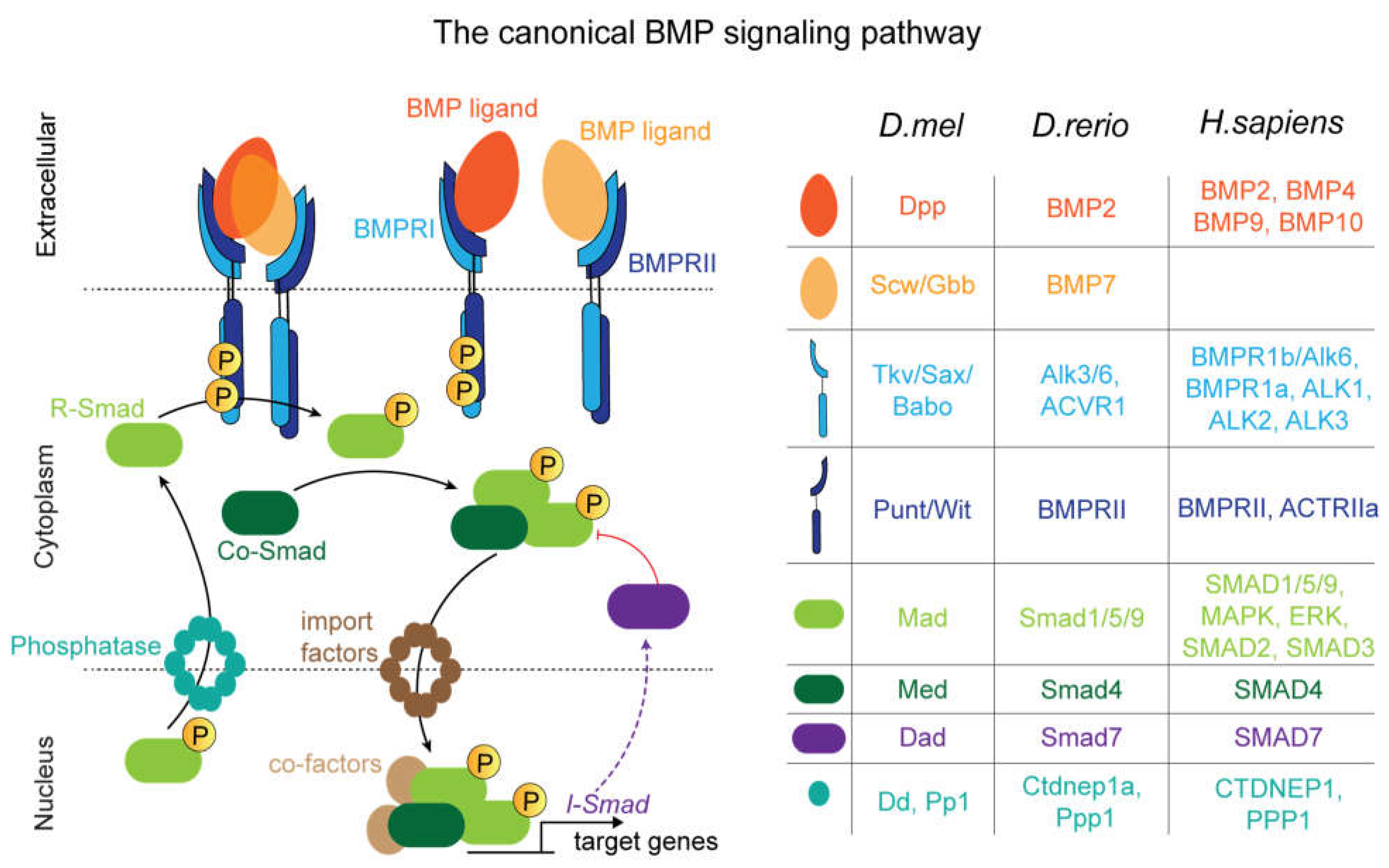
Figure 6.
Overview of existing mathematical models of BMP gradient formation, signal transduction and gene expression across the reviewed model systems. Dashed lines indicate that all the mathematical models listed for a given model system incorporate a common mechanistic component. This demonstrates that, despite spanning diverse systems, all of the models have shared mechanistic foundations.
Figure 6.
Overview of existing mathematical models of BMP gradient formation, signal transduction and gene expression across the reviewed model systems. Dashed lines indicate that all the mathematical models listed for a given model system incorporate a common mechanistic component. This demonstrates that, despite spanning diverse systems, all of the models have shared mechanistic foundations.
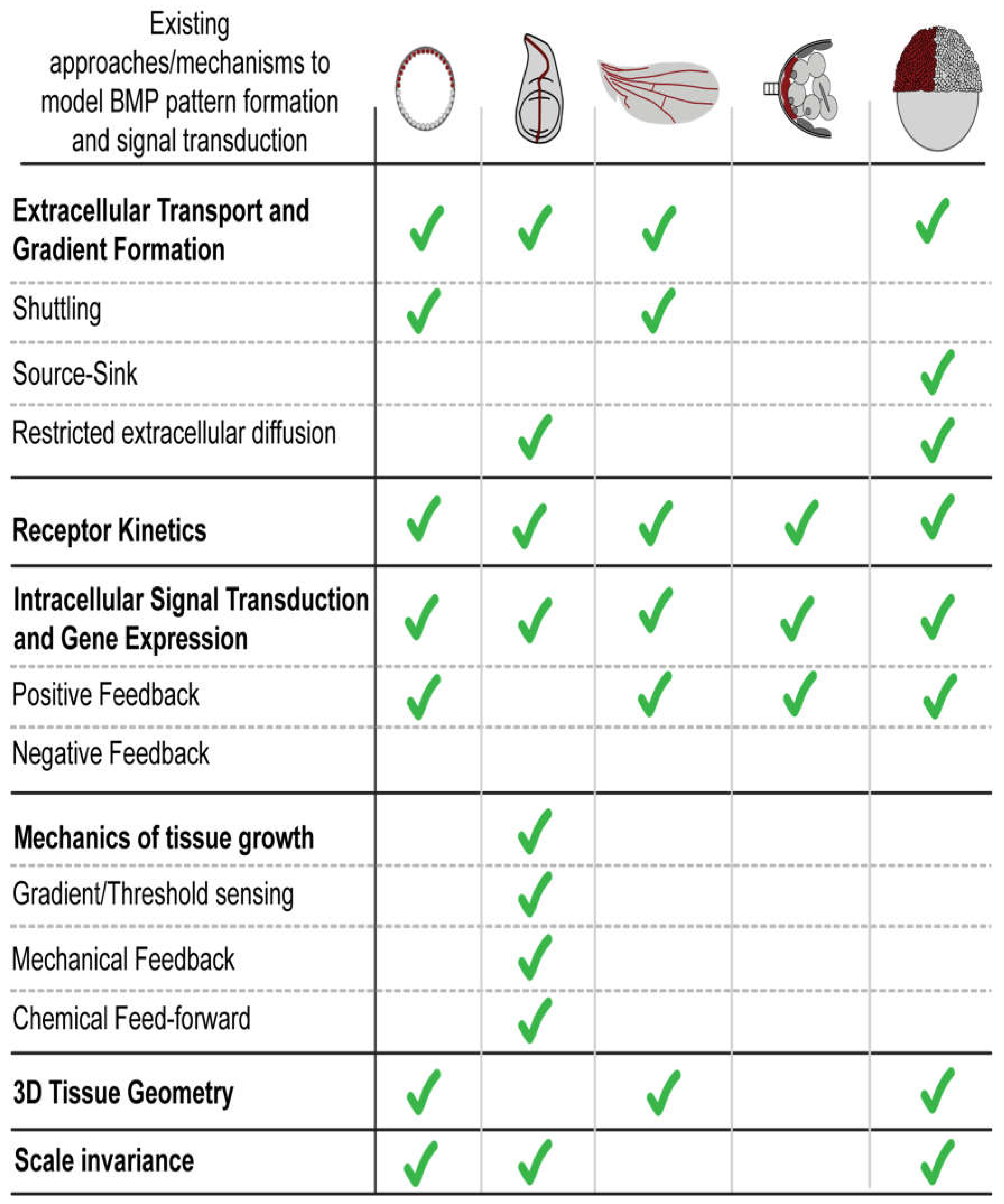

Table 1.
Dynamics and functional outputs of SMAD network across biological systems. *For in vitro human pluripotent stem cell systems, BMP is applied uniformly and does not form a spatial gradient; hence, reported time scales reflect transcriptional endpoints rather than resolved signaling kinetics.
Table 1.
Dynamics and functional outputs of SMAD network across biological systems. *For in vitro human pluripotent stem cell systems, BMP is applied uniformly and does not form a spatial gradient; hence, reported time scales reflect transcriptional endpoints rather than resolved signaling kinetics.
| BMP pathway component and properties | Drosophila: Blastoderm Embryo |
Drosophila: Larval Wing Disc |
Drosophila: Pupal Wing |
Drosophila: Germline stem cells |
Zebrafish: Embryo |
Human: Endothelial cells differentiated from hPSCs |
|---|---|---|---|---|---|---|
| Downstream target/ differentiation marker | Type I target: zen, Race,& hnt Type II target: rho, tup,& ush Type III target: pnr [5] |
Brinker (brk), optomotor-blind (omb), daughters against dpp (dad), and spalt major (sal) [6,7] |
Dad [6,7] & Crossveinless-2 (CV-2) (BMP-responsive extracellular regulator) [8,9] EGFR/MAPK outputs (vein differentiation): Rhomboid (rho), Star (s), and argos (aos) (indirect) [10,11], and blistered/Srf (bs) (repressed) [12] |
dad, Rfx, & futsch, bag of marbles (bam) [13] |
highest threshold: sizzled (szl) and tp63 Intermediate threshold: foxi1 and gata2a Low threshold: bambia [14,15] |
ID1, ID2, ID3 SMAD6/7 [16] |
| BMP Gradient Formation Time Scale | Tens of minutes after nuclear cycle 13 [17,18] | < 4 hours [19] | Hours (~18-30 h AP) [20,21] | Minutes (rapidly) [13,22] | Hours [23] | Not applicable* (uniform BMP signaling) |
| Gene expression time scale | ~30 minutes [24] | Not directly measured/TBD | Hours (Overlaps with gradient formation) [20] | Hours (not directly measured) (8) | Minutes to hours [14,15] |
≤ 24 hours (endpoint measured; onset not resolved) [25] |
| Gradient length scale | 5-6 cell diameters (35 μm) [24] |
20-40 cell diameters (100 μm) [23] | Short-range: vein restricted cell-scale, ~1-3 cell diameters (~5-15 μm) [10,20] Long-range: Transport to crossveins (~50-100 μms) [15,21] |
1-2 cell diameters (5 μm) (13,22,23) | 25+ cell diameters (~ 700 μm) [23] |
Not applicable* (uniform BMP signaling) |
Table 2.
provides a detailed listing of the specific models associated with each system.
| System | Modeling Approach/Type | Author(s) | Key Finding or Purpose |
|---|---|---|---|
| Drosophila Embryo | Shuttling and receptor mediated degradation | Mizutani et al. (2005) | Showed that the sharp pMad peak requires the Sog/Tsg shuttling mechanism and Tolloid-mediated cleavage. |
| Positive feedback | Wang & Ferguson (2005) | Proposed that positive feedback and bistable dynamics reinforce and maintain sharp signaling boundaries. | |
| Receptor kinetics and feedback | Umulis et al. (2006) | Showed that slow, reversible receptor-ligand binding kinetics provide robustness against changes in receptor levels. | |
| 3D geometry | Umulis et al. (2010) | Argued that robustness must be evaluated by its effect on threshold and downstream gene expression. | |
| DrosophilaWing Disc | Restricted Diffusion | Schwank et. al. (2011) | Investigated competing hypotheses to show that Dpp transport occurs via restricted extracellular diffusion. |
| Gradient | Lawrence, P. A., & Struhl, G. (1996). | Proposes that the slope of the Dpp gradient drives cell proliferation if the slope is steeper than a certain threshold. | |
| Threshold (Gene Expression) | Schwank, G et al.(2008) | Proposes that cells proliferate if Dpp signaling is above a certain threshold level | |
| Temporal | Wartlick, O et al. (2011) | Proposed that cells respond to a relative increase (not absolute level) in Dpp to trigger uniform proliferation. | |
| Growth Equalization Model | Schwank, G. et al.(2008) & Schwank, G. et al.(2011) | Modeled how the target gene brk acts as a growth suppressor to balance proliferation across the disc. | |
| Mechanical Model | Aegerter-Wilmsen et al.(2007),Hufnagel,L. et al. (2007) | Modeled how physical tissue forces (compression/stretching) provide feedback to ensure uniform proliferation. | |
| Vg Feed-Forward Model | Zecca, M., & Struhl, G. (2021). | Modeled how Dpp, Wg, and Vg interact in a feed-forward loop to coordinate and expand the wing tissue. | |
| Expansion- Repression model | Ben-Zvi & Barkai (2010) | Modeled how Pentagone (Pent) acts as a rapidly diffusing "expander" molecule of the Dpp gradient. | |
| Pseudo source sink | Zhu et. al (2020) | Modeled how Dpp-mediated feedback downregulation of its own receptors drives scaling. | |
| Recycling | Romanova-Michaelides et al. (2021) | Modeled how Dpp gradient scaling is driven by a tunable “recycling gear” mechanism where the feedback regulator Pentagone modulates receptor binding to favor ligand re-exocytosis. | |
| Drosophila Pupal Wing | Gradient Formation | Gui et al. (2019) | Describes Dpp gradient formation mechanism during first apposition, inflation, and second apposition of pupal wing development to create the 3D architecture of the adult wing. |
| Receptor- Mediated Degradation & Shuttling | O’Connor (2006) | Discusses long-range Dpp transport through Sog/Ts2 shuttling mechanism and Tolkin protein-mediated cleavage. Proposed PCV formation is driven by Dpp/Gbb heterodimers. | |
| Positive Feedback Loop | H. Antson et al. (2022), O’Connor (2006) | Discusses positive-feedback loop of the Sog/CV-2 mediated transport and its proposed role in releasing Dpp:Gbb heterodimers in LVs to facilitate PCV formation. It also discusses the ability of this feedback loop to create sharp step-gradients for spatial bistability. | |
| DrosophilaGSC Niche | Bistability / Differentiation | Pargett et al. (2014) | Modeled the GSC/CB switch to identify Brat as a key differentiation factor that antagonizes BMP. |
| Positive Feedback Loop | Xia et al. (2012) | Modeled how Fused (Fu) dynamics create a positive feedback loop to maintain stemness (GSC) or allow differentiation (CB). | |
| Multi-compartment GSC division | Shaikh & Reeves (2024) | Modeled how Dad and Fused work synergistically to ensure robust GSC division and homeostasis. | |
| Zebrafish embryo | Mechanistic/ Gradient Formation | Zinski et. al. (2017) & Pomrienke et. al. (2017) | Tested theorized gradient models against mutant data, providing strong support for the source-sink mechanism. |
| 3D Simulation (Advection Diffusion Reaction) | Li et. al. (2019) | Developed a more geometrically accurate 3D model that also supported the source-sink mechanism. | |
| Stochastic Receptor Model | Larson et. al. (2025, preprint) | Modeled receptor combinations for signal transduction, showing emergent low-pass filter mechanism arising from heterodimer-heterotetramer requirement. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



