Submitted:
31 December 2025
Posted:
01 January 2026
You are already at the latest version
Abstract
Vector-borne plant diseases represent a complex phytosanitary challenge. Mathematical models serve as a key tool for analyzing integrated management strategies, enabling more effective control of these pests. A dynamical system is presented to model the infection of Tahiti lime (Citrus x latifolia) with the bacterium Candidatus Liberibacter asiaticus (CLas), transmitted mainly by infected adults of the psyllid D. citri, which causes the citrus greening—Huanglongbing (HLB). The proposed model is based on the D. citri-HLB pathosystem, basic interactions between bacteria, vector psyllid hosts, trees and a vector parasitoid wasp. It consists of nine ordinary differential equations that model the rates of change of the numbers of infected and uninfected vector nymphs and adult females, of infected and uninfected trees of high and low productivity, and of the parasitoid Tamarixia radiata, recommended for the biological control of D. citri. The parameters of the model are identified from extant literature or otherwise estimated, in both cases being adjusted to Colombian conditions. A mathematical analysis of a simplified model is carried out, and simulations are conducted to demonstrate the effect of different types of control easures.
Keywords:
1. Introduction
- The extinction equilibrium —locally asymptotically stable when the basic reproduction number of D. citri, , is smaller than 1 (in which case it is the only steady state), and unstable when it is larger than 1;
- an equilibrium with D. citri nymphs and adults but no T. radiata that exists only when ; and
- a coexistence equilibrium of vectors and parasitoids.
2. Materials and Methods
- := number of D. citri stage 3–4–5 nymphs not carrying bacteria CLas (i.e., uninfected nymphs);
- := number of D. citri stage 3–4–5 nymphs carrying bacteria CLas (i.e., infected/infectious nymphs);
- := number of D. citri adults that do not transmit the HLB.
- := number of D. citri adults that transmit the HLB.
- := number of citrus trees with high sprouting and fruit production, and free of HLB
- := number of citrus trees with high sprouting and fruit production, and infected with HLB
- := number of citrus trees with low sprouting and fruit production, and free of HLB.
- := number of citrus trees with low sprouting and fruit production, and infected with HLB
- := number of adult female T. radiata
- The per capita “oviposition" rate of adult D. citri, , is actually the product of four factors: their natural per capita egg-laying (= oviposition) rate, the probability of survival from egg to nymph-1 stage, the probability of survival from nymph-1 to nymph-2 stage, and the probability of survival from nymph-2 to nymph-3 stage. Hence the quotation marks around ‘oviposition’.
- The per capita oviposition rate of adult D. citri is the same whether they are healthy or infected; thus the generation of new nymphs does not depend on whether they originate from a healthy or an infected adult.
- Nymphs that hatch on a healthy tree are considered healthy, while those that hatch on an HLB-infected tree are considered infected, regardless of the HLB-infection status of the adult parent. This is justified because, as far as we know, HLB is not transmitted vertically from adult female moths to their offspring to daughter.
- Because oviposition and nymph development take place only on citrus tree shoots, we assume that the availability or scarcity of the latter impacts the natural oviposition rate through a factor that is an increasing function of the total number of shoots of the corresponding infection status, with values between 0 and 1.
- Because D. citri nymphs are immobile, they do not have logistic per capita mortality driven by intra-species competition.
- Because the livelihood of adult D. citri is intimately tied to the availability of citrus tree shoots to feed on, logistic mortality from intra-species competition is assumed to occur at a rate that decreases with increasing availability of shoots of any infection status.
- Because adult D. citri who get infected with HLB from feeding on shoots from an infected tree have too short a life-span to develop a pathogen load high enough to further transmit infection to uninfected trees when feeding on their shoots, we do not consider such feeding as a source of infected adults since we only want the compartment to contain infectious adults and not infected-but-not-infectious.
- Because T. radiata deposes its eggs only in D. citri nymphs —only one egg per nymph— and this results in the death of the nymph, the per capita T. radiata-induced mortality rate of D. citri nymphs is assumed to be proportional to the number of T. radiata wasps.
- D. citri adults are considered infected only if they originate from infected nymphs.
- D. citri nymphs may die due to natural causes, to the effect of agrochemical products used for their control, or to parasitism by T. radiata.
- D. citri adults may die due to natural causes or as a result of agrochemical control.
- We do not include the planting of new trees. Therefore, the total tree population can only decrease.
- Trees are classified into high-fitness —producing a commercially acceptable amount of fruit— and low-fitness —with very low and commercially negligible production.
- Irrespective of their fitness, trees can develop HLB only if an infected D. citri adult feeds on them.
- The only way for a high-fitness tree to transition to low fitness is through neglect, irrespective of whether it is healthy or HLB-infected.
- A healthy low-fitness tree can recover and transition to high fitness with proper care.
- An HLB-infected low-fitness tree cannot recover and regain high fitness, because of the disease.
- Irrespective of their fitness, healthy trees produce healthy shoots and HLB-infected trees produce infected shoots.
- HLB-infected trees, whether of high or low fitness, may be eradicated as a control measure.
- T. radiata wasps experience natural mortality and may also die from the effect of agrochemicals applied for the control of D. citri.

| Symbol | Description |
|---|---|
| maximal per capita production rate of D. citri 3rd-instar nymphs (per adult) | |
| average number of shoots per high-sprouting uninfected tree | |
| average number of shoots per high-sprouting infected tree | |
| average number of shoots per low-sprouting uninfected tree | |
| average number of shoots per low-sprouting infected tree | |
| natural per capita mortality rate of stages 3–5 nymphs | |
| natural per capita mortality rate of adult D. citri | |
| per capita mortality rate of low-productivity uninfected trees | |
| per capita mortality rate of low-productivity infected trees | |
| per capita mortality rate of T. radiata | |
| per capita transition rate of stage 3–5 nymphs to adulthood | |
| per capita oviposition rate of T. radiata | |
| per capita mortality rate of D. citri nymphs from agrochemicals | |
| per capita mortality rate of adult D. citri from agrochemicals | |
| per capita mortality rate of T. radiata from agrochemicals | |
| intra-specific competition constant in adult D. citri mortality per sprout | |
| per capita infection rate of trees by adult D. citri when all of them are infected | |
| per capita transition rate of low-productivity trees to high productivity | |
| per capita transition rate of uninfected trees from high to low productivity | |
| per capita transition rate of infected trees from high to low productivity | |
| per capita eradication rate of high-productivity trees with HLB | |
| per capita eradication rate of low-productivity trees with HLB | |
| per capita release rate of T. radiata | |
| number of nymphs in stages 3–5 resulting in 50% reduction of T. radiata | |
| fraction of adult D. citri to keep tree infection rate continuous at |
3. Results
3.1. Mathematical Analysis of a Reduced Model
3.1.1. Well-Posedness
3.1.2. Steady States of the System
3.1.3. Local Stability of Equilibrium Points
3.1.4. Global Stability of the Trivial Equilibrium
- Since is equivalent to
-
Also, because we have .Use monotonicity bounds:Also
3.2. Simulations
3.2.1. Reduced System
| Parameter | Value | Parameter | Value |
|---|---|---|---|
| 1/32 | 1/6 | ||
| 1/6.11 | 0.04 | ||
| 2 | 200 |
| Variable | Initial Value |
|---|---|
| 30 | |
| 0 | |
| 200 | |
| 100 | |
| 0.2 | |
| 150 | |
| , | 0 |
| 1,000 |
3.2.2. Full System
- No transition between low-productivity and high-productivity trees (or vice versa)
- No tree eradication
- Effectiveness of agrochemical decays linearly to zero 30 days after application
| Parameter | Value | Source |
|---|---|---|
| 0.995 | Estimated value from [11] | |
| 42 | Estimated value from [12] | |
| 32 | Assumed from [12] | |
| 21 | Assumed from [12] | |
| 21/2 | Assumed from [12] | |
| 0.02 | Estimated value from [12] | |
| 1/(48.25) | Estimated value from [13] | |
| 0.04 | Estimated value from [14] | |
| 1/(10*365) | Estimated value from [15] | |
| 1/(5*365) | Estimated value from [16] | |
| 1/9 | Estimated value from [13] | |
| 25 | Estimated value from [17] | |
| Assumed | ||
| [18] | ||
| 0 | Assumed | |
| , | 0 | Assumed |
| , | 0 | Assumed |
| 200 | Assumed | |
| Assumed |
| Variable | Initial Value |
|---|---|
| 192 | |
| 214 | |
| 100 | |
| 43 | |
| 96 | |
| 107 | |
| 100 | |
| 42 | |
| 0 |
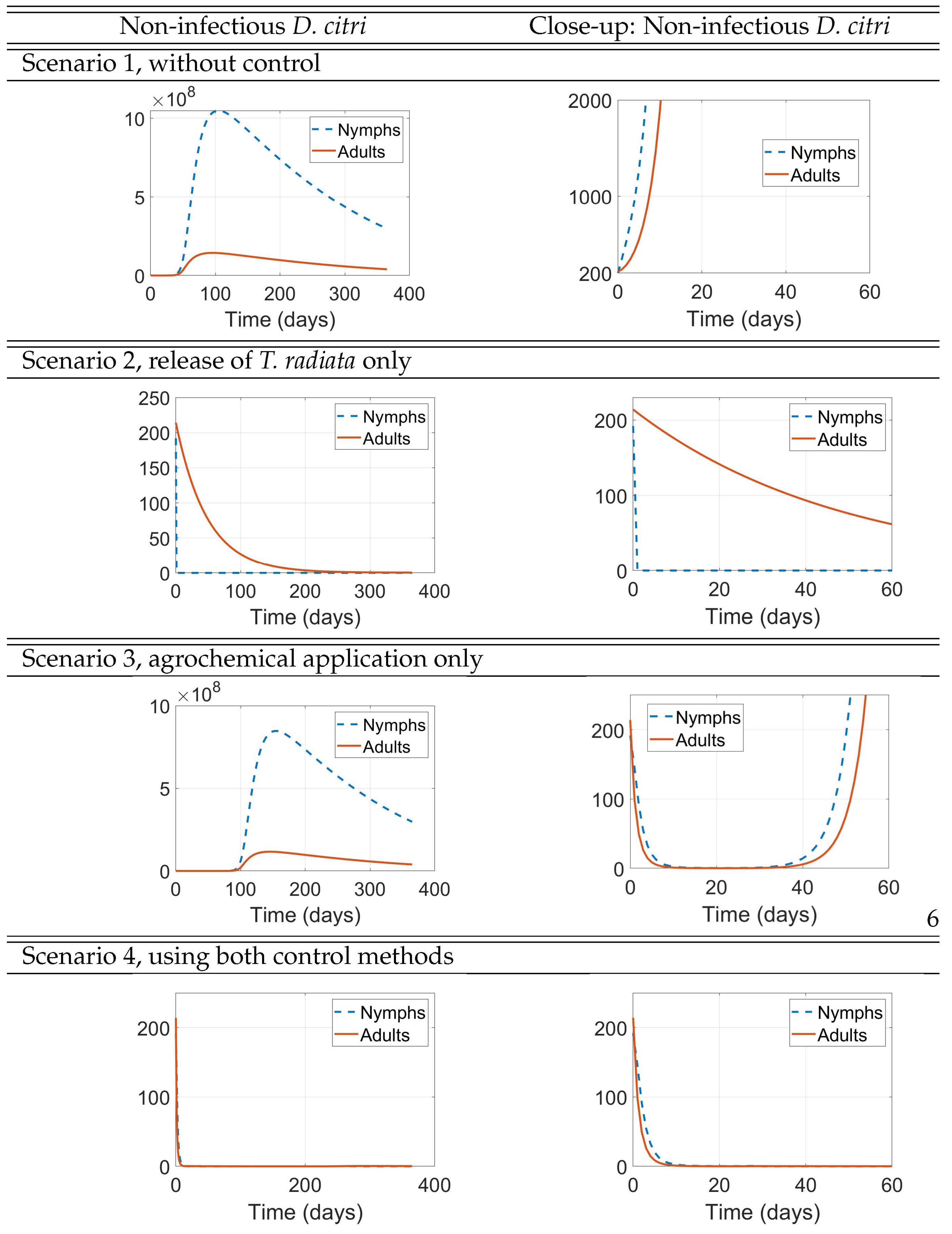
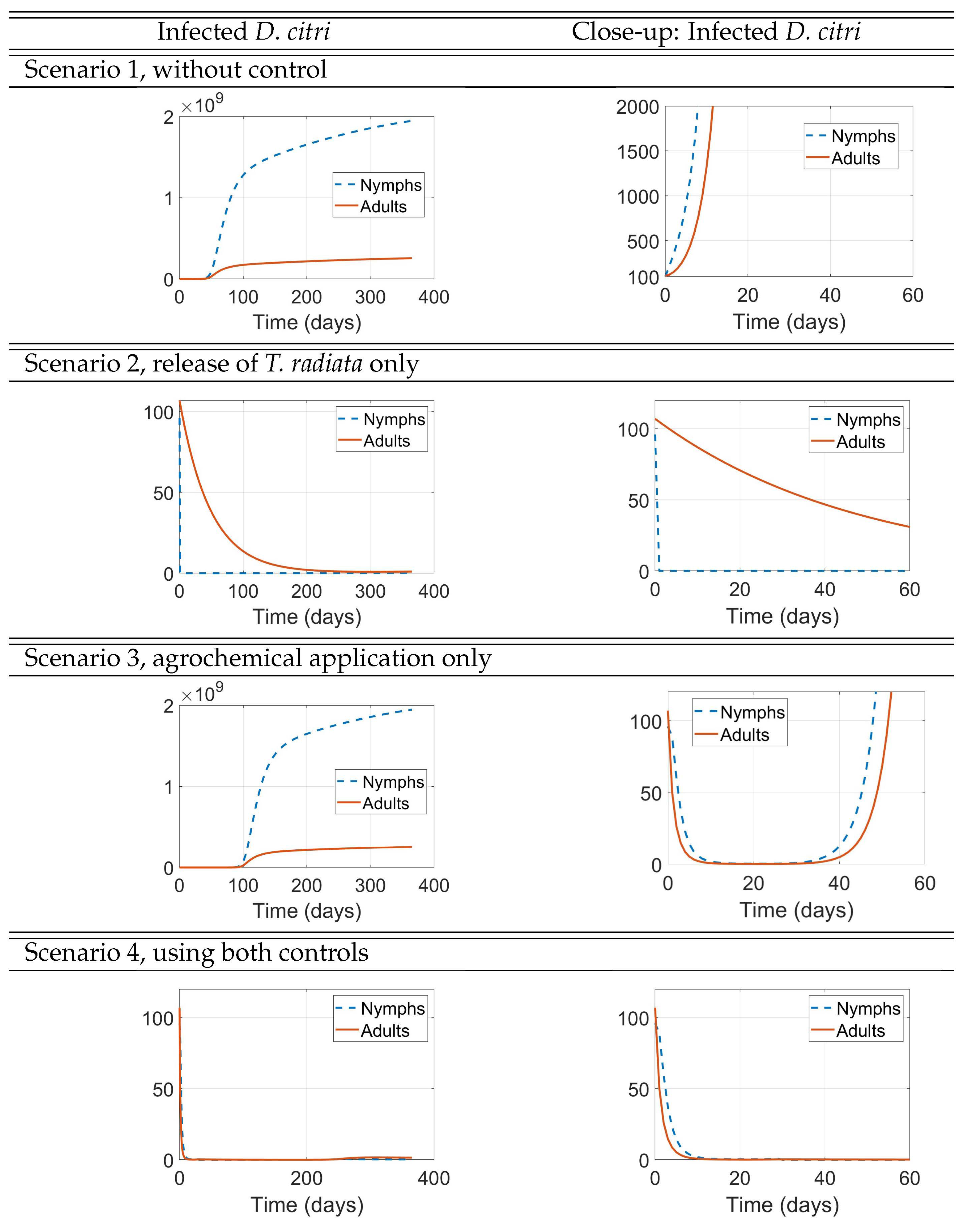
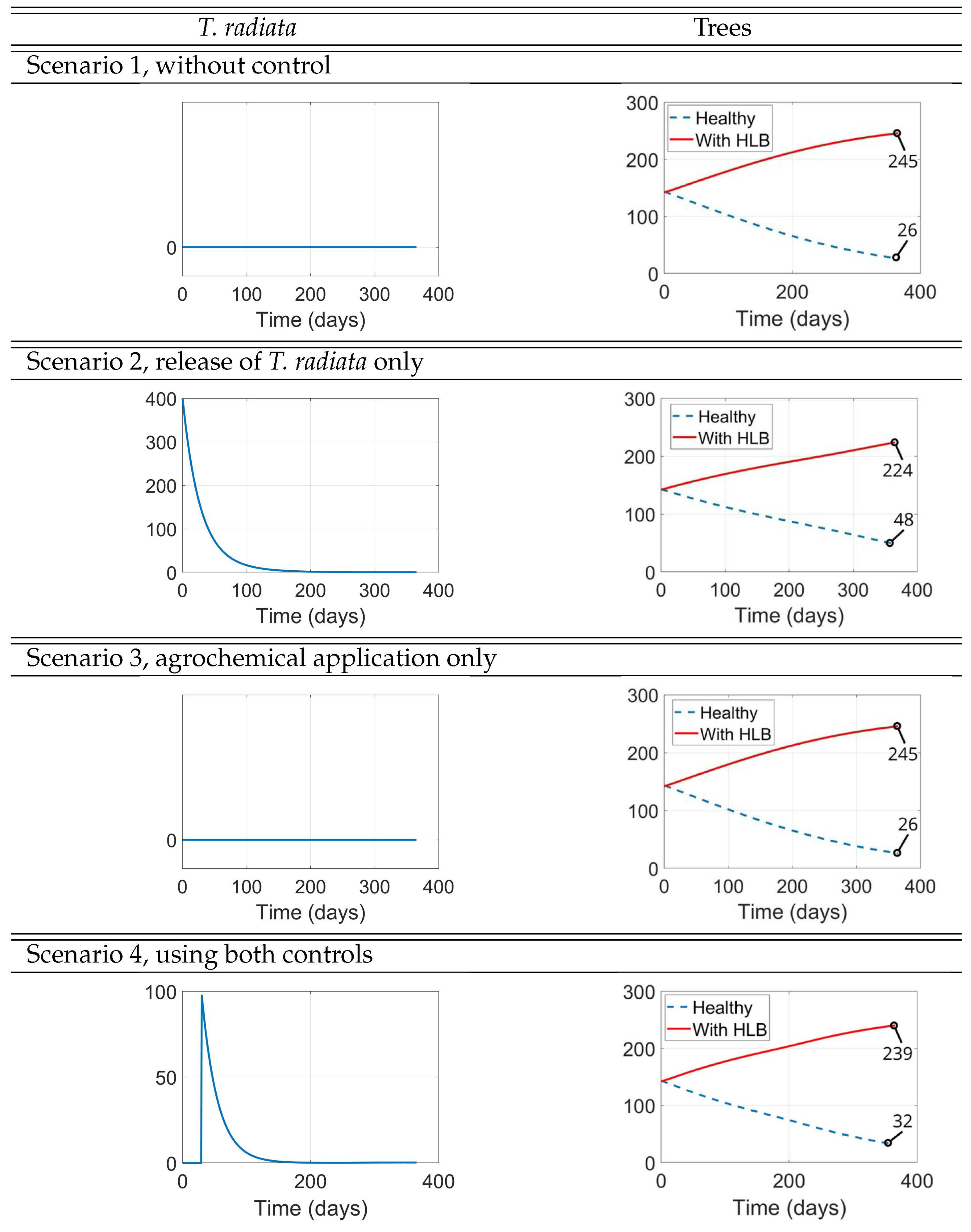
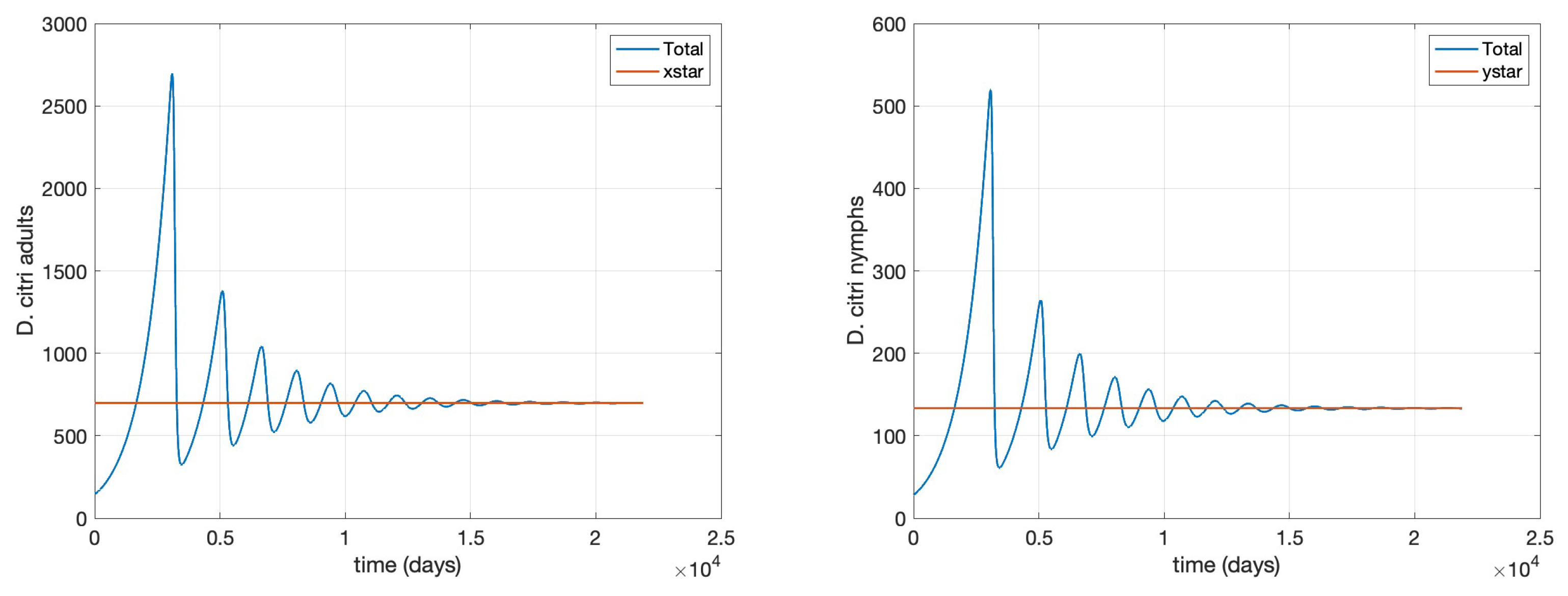
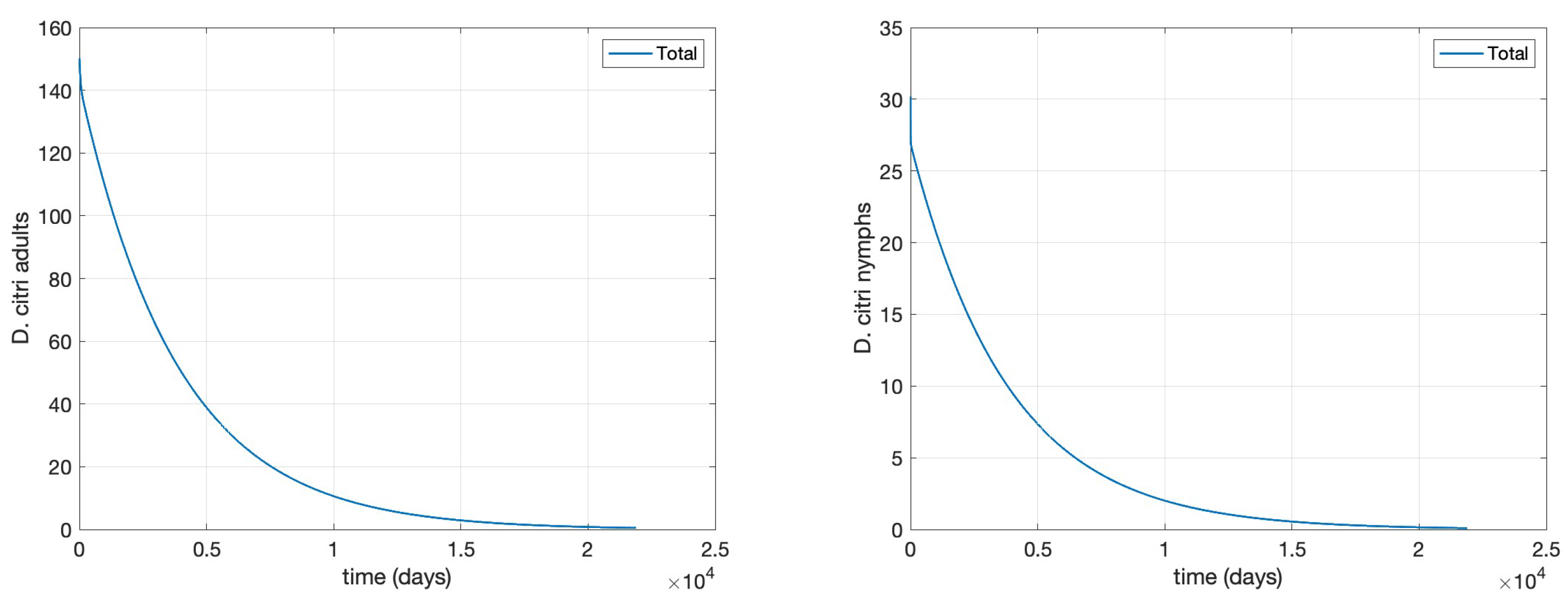
- Scenario 1, No control: (i.e., no management interventions)
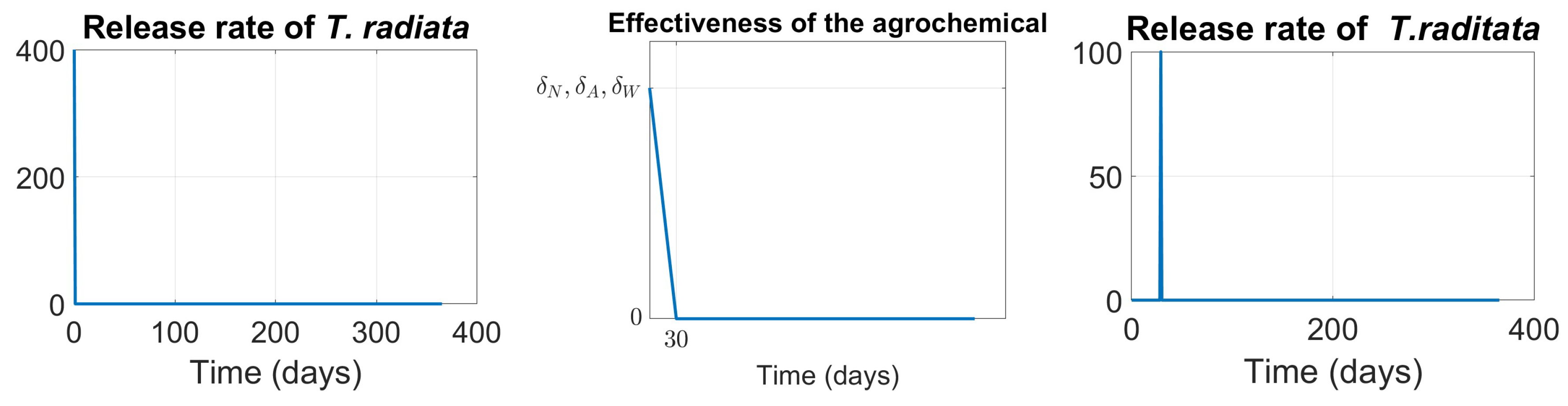
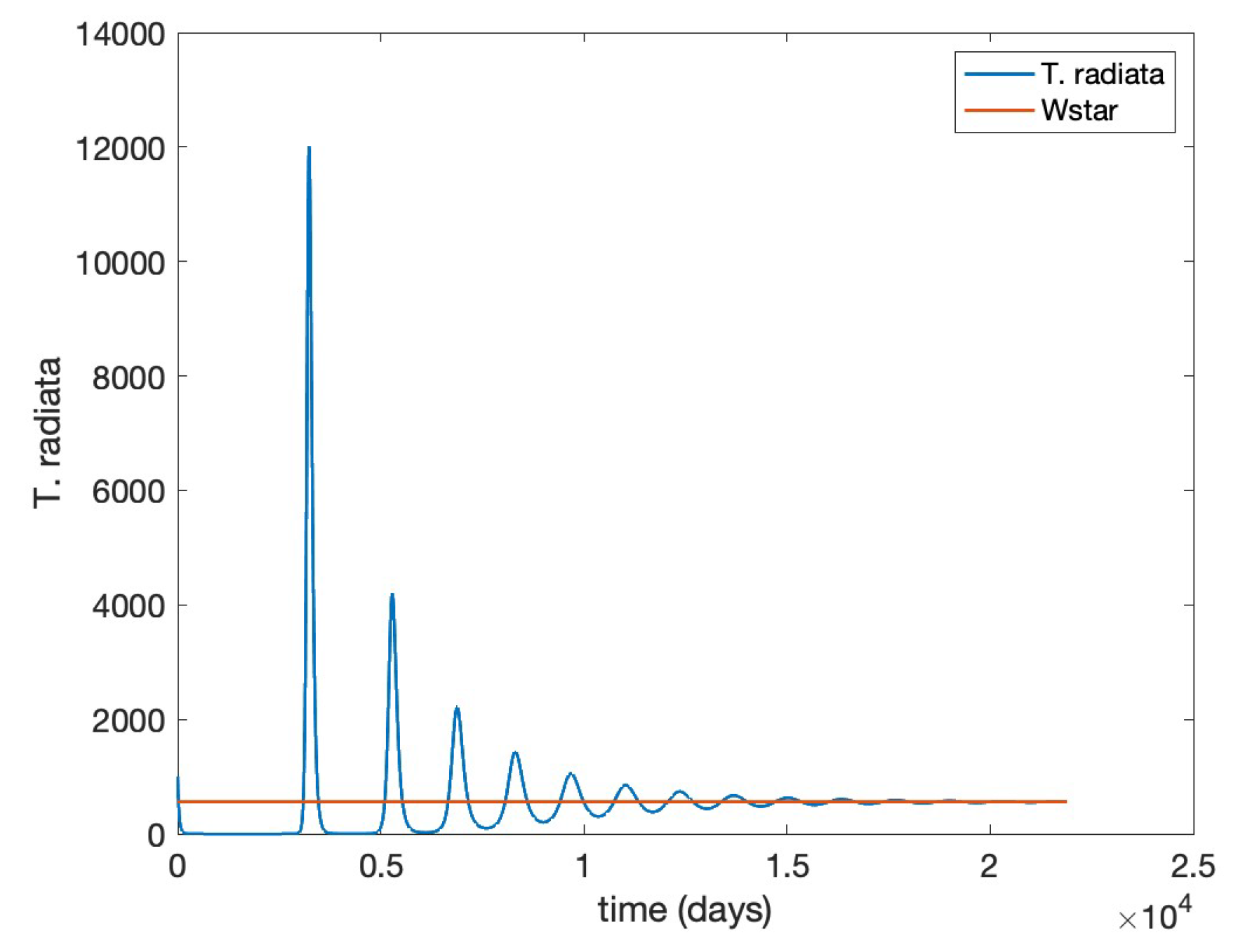
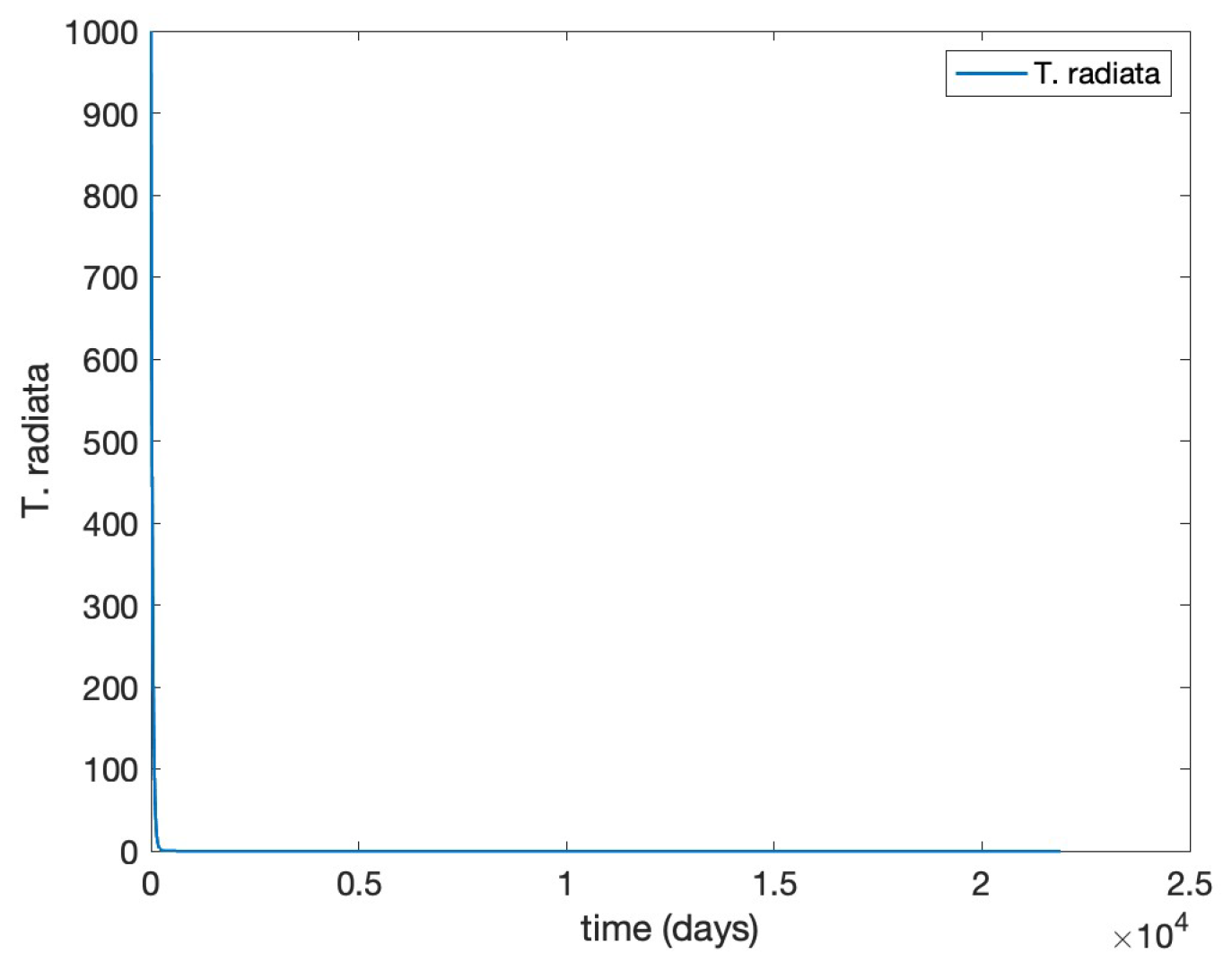
- Scenario 2,T. radiatarelease only: introduction of 400 females at the initial time, taking into account as a reference the prescribed release rates (individuals/ha, between male and female) [15], as shown in Figure .
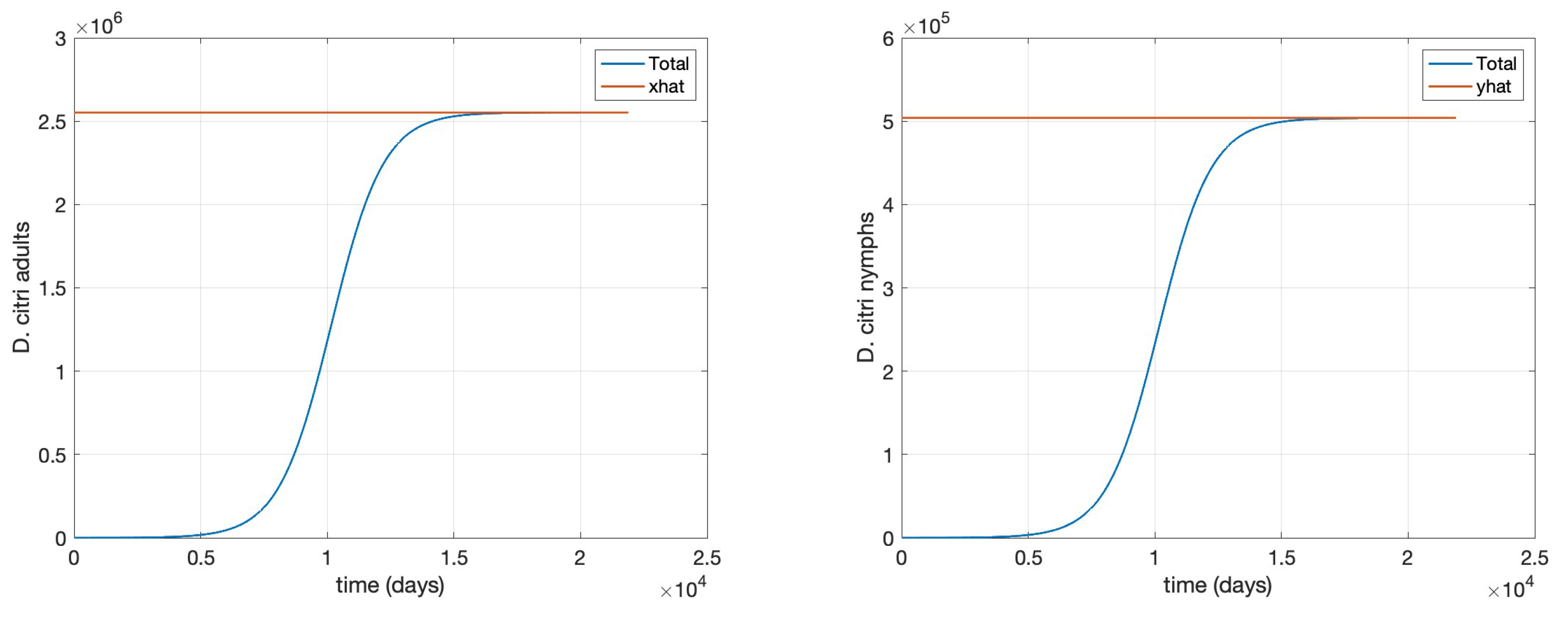
- Scenario 3, Agrochemical application only: (, , ) shown in Figure .
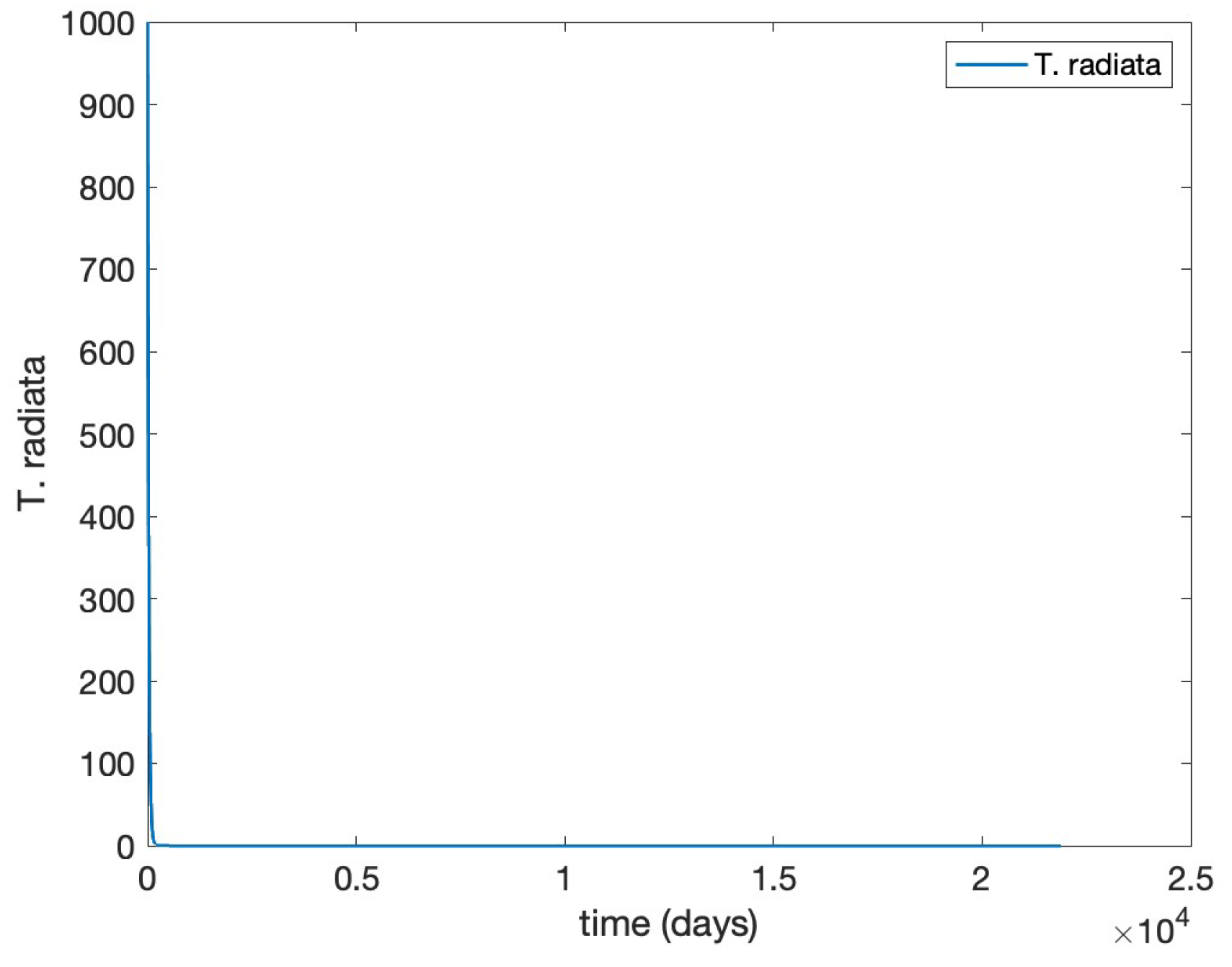
- Scenario 4, Combined control: pairing initial agrochemical application (see Figure ) with T. radiata release of 100 females at day 30 (see Figure ).
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HLB | Huanglongbin |
| CLas | Candidatus Liberibacter asiaticus |
References
- Aguirre, A.; Giancola, S.; Gochez, A. Manejo del complejo HLB-vector en regiones citrícolas afectadas en centro y sudamérica. Technical report, FONTAGRO, 2025.
- Gottwald, T.R. Current epidemiological understanding of citrus huanglongbing. Annual Review of Phytopathology 2010, 48, 119–139. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wu, F.; Duan, Y.; Singerman, A.; Guan, Z. Citrus greening: Management strategies and their economic impact. HortScience 2020, 55, 604–612. [Google Scholar] [CrossRef]
- Villar-Luna, H.; Santos-Cervantes, M.E.; Rodríguez-Negrete, E.A.; Méndez-Lozano, J.; Leyva-López, N.E. Economic and Social Impact of Huanglongbing on the Mexico Citrus Industry: A Review and Future Perspectives, 2024. [CrossRef]
- de Agricultura, M.; (ICA), I.C.A. PLAN DE ACCIÓN NACIONALPARA EL HLB DE LOS CÍTRICOS 2019-2022. In Proceedings of the Mesa temática nacional del huamglomgbinb de los cítricos, 2019, pp. 1–49.
- Kondo, T.; Riaño, N.M.; Pérez-Artiles, L.; Martínez, M.F.; Ríos-Rojas, L.; Fuentes, M.P.; Guarín, J.H.; Carabalí-Muñoz, A.; Orduz-Rodríguez, J.O.; Urquiza, G.P.C.; et al. Avances en el conocimiento y recomendaciones para el manejo integrado de la enfermedad Huanglongbing (HLB) en los cultivos de cítricos en Colombia; Corporación Colombiana de Investigación Agropecuaria (Agrosavia). 2024. [Google Scholar] [CrossRef]
- Chen, X.; Wong, S.W.; Stansly, P.A. Functional Response of Tamarixia radiata (Hymenoptera: Eulophidae) to Densities of Its Host, Diaphorina citri (Hemiptera: Psylloidea). Annals of the Entomological Society of America 2016, 109, 432–437. [Google Scholar] [CrossRef]
- Ammar, E.D.; Ramos, J.E.; Hall, D.G.; Dawson, W.O.; Shatters, R.G. Acquisition, replication and inoculation of Candidatus Liberibacter asiaticus following various acquisition periods on huanglongbing-infected citrus by nymphs and adults of the asian citrus psyllid. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Stansly, P.A. Biology of Tamarixia radiata (Hymenoptera: Eulophidae), parasitoid of the citrus greening disease vector Diaphorina citri (Hemiptera: Psylloidea): A mini review. Florida Entomologist 2014, 97, 1404–1413. [Google Scholar] [CrossRef]
- George, J.; Ammar, E.D.; Hall, D.G.; Shatters, R.G.; Lapointe, S.L. Prolonged phloem ingestion by Diaphorina citri nymphs compared to adults is correlated with increased acquisition of citrus greening pathogen. Scientific Reports 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Camilo, G.G.C. Fluctuación poblacional y tabla de vida Diaphorina citri Kuwayama (Hemiptera: Liviidae) en cultivares de cítricos en las condiciones del municipio de Zona Bananera, Magdalena. 2022. [Google Scholar]
- Parra-Fuentes, M.; Pérez-Artiles, L.; Arias-Castro, J.H.; Martínez-Romero, H. Estimation of the reproductive and population parameters of Diaphorina citri (Hemiptera: Liviidae). Revista Ciência Agronômica 2025, 56, 1–15. [Google Scholar] [CrossRef]
- García, Y.; Ramos, P.Y.; Sotelo, P.A.; Kondo, T. Biología de Diaphorina citri (Hemiptera: Liviidae) bajo condiciones de invernadero en Palmira, Colombia. Revista Colombiana de Entomología 2016, 42, 36–42. [Google Scholar] [CrossRef]
- Quilici, S.; Joulain, H.; Manikom, R. Étude de la fécondité de Tamarixia radiata (Waterston, 1922) (Hymenoptera: Eulophidae), ectoparasitoïde primaire du psylle asiatique Kuwayama (Homoptera: Psyllidae), vecteur du greening des agrumes. Fruits 1992, 47, 184–194. [Google Scholar]
- Riaño, N.M.; Martínez, M.F.; Orduz-Rodríguez, J.O.; Ríos-Rojas, L.; Galé, Y.L.; Hernández, M.J.Y.; Muñoz, A.C.; Kondo, T.; Muñoz, M.C.G.; González, J.L.; et al. Modelo productivo de lima ácida Tahití (Citrus × latifolia Tanaka ex Q. Jiménez) para Colombia; Agrosavia. 2020. [Google Scholar] [CrossRef]
- Halbert, S.E.; Manjunath, K.L. Asian citrus Psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: a literature review and assesment of risk in Florida. Florida Entomologist 2004, 87, 330–353. [Google Scholar] [CrossRef]
- Gmez-Torres, M.L.; Nava, D.E.; Parra, J.R.P. Life table of tamarixia radiata (Hymenoptera: Eulophidae) on diaphorina citri (Hemiptera: Psyllidae) at different temperatures. Journal of Economic Entomology 2012, 105, 338–343. [Google Scholar] [CrossRef]
- Martini, X.; Willett, D.S.; Kuhns, E.H.; Stelinski, L.L. Disruption of Vector Host Preference with Plant Volatiles May Reduce Spread of Insect-Transmitted Plant Pathogens. Journal of Chemical Ecology 2016, 42, 357–367. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.



