
Submitted:
30 December 2025
Posted:
31 December 2025
You are already at the latest version
Abstract
Nutraceuticals, bioactive compounds derived from foods, are increasingly investigated as interventions to promote healthy ageing. Multi-ingredient formulations may offer additive or synergistic benefits by targeting multiple ageing pathways while using low doses of each component for improved safety. However, their efficacy in mammals remains poorly understood. Here, we compared the effects of a continuous multi-ingredient nutraceutical intervention with two short-courses of senolytic regimen in naturally aged male C57Bl/6J mice. Importantly, these mice were overweight following a switch to soaked food at 20 months, a protocol that increased caloric intake and likely induced metabolic stress. This context frames the study as a model of ‘rescue’ from premature ageing rather than extension of maximum lifespan. Mice were assigned to either control, nutraceutical (12 pro-longevity natural compounds), or senolytic (Navitoclax plus BAM15) groups at 20 months of age. Lifespan and healthspan indicators were assessed longitudinally. Both interventions improved survival compared to controls (median lifespan +18–21%) and mitigated frailty progression, but with distinct patterns: nutraceutical benefits accumulated gradually, whereas senolytic effects were transient. Cognitive performance was preserved in nutraceutical-treated mice and improved shortly after senolytic treatment. In vitro, the nutraceutical lacked senolytic activity but exhibited senostatic effects, reducing nuclear size, ROS release, and IL-6 secretion in senescent fibroblasts. These findings suggest that multi-ingredient nutraceuticals can restore healthspan compromised by metabolic stress and deliver benefits comparable to senolytics when administered continuously, potentially through senostatic mechanisms. Combining senolytics to reduce senescent burden with long-term nutraceutical treatment may offer a safe, accessible strategy to optimise healthspan, particularly in the context of modern human ageing, which often occurs under conditions of caloric excess and metabolic syndrome.
Keywords:
aging
; cellular senescence
; senotherapeutics
; senolytics
; geroprotectors
; healthspan
; frailty
1. Introduction
Nutraceuticals are bioactive compounds derived from foods and plants and are increasingly investigated as interventions to promote healthy ageing.
Unlike conventional pharmaceuticals, nutraceuticals are typically consumed as dietary supplements and aim to modulate biological processes associated with ageing, including those described as the ‘hallmarks of aging’ [1] such as chronic inflammation, mitochondrial dysfunction, disabled macroautophgy and cellular senescence.
A growing body of evidence from studies across multiple model organisms suggests that certain nutraceuticals can improve healthspan and, in some cases, extend lifespan [2,3,4,5,6,7,8,9]. Examples include polyphenols such as fisetin and pterostilbene, amino acid derivatives such as glycine, and metabolic intermediates such as alpha-ketoglutarate. Some of these compounds have progressed to early-phase clinical trials, including fisetin [10], pterostilbene [11] or calcium alpha-ketoglutarate [12]. However, robust evidence from well-controlled studies in mammals remains limited.
Ageing is driven by complex, interconnected biological processes at cellular, tissue and organismal levels [1]. Among these, cellular senescence plays a central role in age-related pathologies and frailty. Senotherapeutic strategies, including senolytics, which selectively eliminate senescent cells, and senostatics, which suppress senescence-associated phenotypes, have shown remarkable success in preclinical models [13,14,15].
Some nutraceuticals exhibit senotherapeutic properties. For example, fisetin has been investigated as a senolytic in several clinical trials [16]. Beyond direct senescence targeting, interventions that modulate ageing hallmarks can exert senostatic effects indirectly; for instance, improving mitochondrial function may reduce reactive oxygen species (ROS) production and senescence-associated secretory phenotype (SASP), while enhancing genome stability may limit persistent DNA damage responses.
Multi-ingredient nutraceutical formulations may allow simultaneous modulation of complementary processes, potentially producing additive or even synergistic effects, and may have senostatic function in long-term [17]. Importantly, these formulations often use low concentrations of each ingredient, which may reduce toxicity and improve safety compared to high-dose single-compound interventions [e.g., 18], which is a critical consideration for long-term use. Yet, systematic evaluation of multi-ingredient combinations remains scarce, leaving a gap in understanding whether such approaches can rival existing strategies.
Both senolytics and senostatic interventions can alleviate ageing phenotypes in vivo broadly, but their patterns of benefit may differ. We have previously demonstrated that short-course senolytic regimens using navitoclax or dasatinib plus quercetin, and continuous senostatic intervention with metformin, both alleviated whole-body irradiation-induced premature ageing in mice [19]. The effects of senolytics was seen already after only one month, but that of metformin become clearer after 3-4 months of intervention. This direct comparison revealed that a single-course senolytic therapy and continuous senostatic intervention produced broadly comparable benefits in reducing frailty and preserving cognitive function, highlighting that different strategies can achieve similar outcomes when targeting senescence-related pathways.
Building on these findings, the present study aimed to determine whether a continuous multi-ingredient nutraceutical intervention specifically design to target the hallmarks of aging [20], combining low doses of 12 ingredients: pterostilbene [8], glucosamine sulfate [3], fisetin [7], glycine [2], lithium aspartate [5], calcium alpha-ketoglutarate [9], magnesium malate [21], vitamin C (ascorbic acid) [22], L-theanine [23], hyaluronic acid [24], Rhodiola rosea root extract [25], and ginger root extract [26], could achieve comparable benefits to senolytic treatment in naturally aged mice. Specifically, we assessed effects on lifespan and healthspan indicators, including frailty and cognitive performance, to evaluate whether nutraceutical combinations can rival pharmacological senescence-targeting approaches.
Importantly, this study was conducted in naturally aged male C57Bl/6J mice that became overweight following a switch to soaked food at 20 months [27], a protocol intended to ensure uniform nutraceutical delivery. This context frames our findings within a model of metabolic stress rather than optimised ageing conditions. Interventions that restore lifespan under such stress may have an important translational relevance to human ageing, which is often characterised by caloric excess and obesity rather than caloric restriction.
2. Material and Methods
2.1. Animals
Male C57Bl/6J mice (post-weaning) were obtained from Charles River Laboratories (UK). Animals were housed in individually ventilated cages (56 × 38 × 18 cm; North Kent Plastics, UK) in groups of three, with sawdust bedding, shredded paper, and cardboard tunnels for enrichment. Environmental conditions were maintained at 20 ± 2 °C, 45–65% humidity, under a 12 h light/12 h dark cycle. Standard CRM-P rodent diet (SDS diets) and water were provided ad libitum.
At 18 months of age, mice underwent baseline phenotyping for frailty, cognitive performance, and body weight. Frailty scores were ranked from low to high, cognitive performance from high to low, and body weight from high to low. Individual ranks were averaged, and mice were allocated accordingly to three experimental groups to minimise baseline variability: Control (C), multi-ingredient nutraceutical (NUT), and senolytic (SEN). Interventions began at 20 months of age, and the study was terminated at 30 months of age. Frailty and cognition were assessed every two months, at least four weeks after the last drug treatment, to avoid acute effects of gavage or injection.
All procedures were approved by the UK Home Office (Project Licence PBDAFDFB0) and complied with the Animals (Scientific Procedures) Act 1986 and ARRIVE guidelines. Mice were monitored daily for health and welfare. Humane endpoints included severe distress and persistent weight loss (Supplementary Table S1).
2.2. Interventions
The multi-ingredient nutraceutical was produced by NOVOS Labs (composition and human-equivalent doses listed in Supplementary Table S2). The formulation contained 12 bioactive compounds: pterostilbene, glucosamine sulfate, fisetin, glycine, lithium aspartate, calcium alpha-ketoglutarate, magnesium malate, vitamin C (ascorbic acid), L-theanine, hyaluronic acid, Rhodiola rosea root extract, and ginger root extract.
The multi-ingredient nutraceutical was incorporated into soaked food prepared by grinding CRM-P pellets, mixing with the nutraceutical, and adding sterile water (1:2 ratio) to achieve a porridge-like consistency. The dose was determined from a preliminary dose-finding study to approximate human-equivalent intake scaled for mice.
As a control, fresh soaked food, with the multi-ingredient nutraceutical (for NUT group) or without (for SEN and C groups) was provided daily throughout the intervention period starting at 20 months of age.
Senolytic treatment followed previously described protocols [28]. Briefly, SEN-group mice received oral gavage of navitoclax (0.5 mg/kg body weight in 10% Polyethylene Glycol (PEG) 400 for five consecutive days, combined with the mitochondrial uncoupler BAM15 (2.5 mg/kg body weight in 40% PEG400) administered by injection. All other mice received vehicle gavage and injection. Navitoclax (A3007-APE-100 mg) was purchased from Stratech; BAM15 (Cat. 5737) from Biotechne; and PEG400 (8074851000) from Merck. Two months after the first course, a second five-day course was administered. During the first course, BAM15 (or vehicle) was delivered intraperitoneally, which likely resulted in accidental puncture of seminal vesicles in some animals, leading to infection-related deaths. Enlarged seminal vesicles are a common, non-malignant pathology in aged male C57Bl/6J mice [29]. These fluid-filled structures can occupy a substantial portion of the lower abdominal cavity, creating a high risk during blind intraperitoneal injections. To mitigate this risk, the second course used subcutaneous injection.
2.3. Ageing Phenotype Assessments
Frailty was assessed using a 30-parameter scoring index [30], adapted from Whitehead et al., 2014 [31], covering integument, musculoskeletal, sensory, and general health domains. Each parameter was scored as 0 (normal), 0.5 (mild deficit), or 1 (severe deficit), and averaged to generate a frailty index score.
Forelimb grip strength was measured using BIO-GS3 BIOSEP apparatus. Each mouse performed three trials, and the mean value was normalised to reference strength values from adult males.
Cognitive function was evaluated using a spontaneous alternation Y-Maze test [30], during the dark phase under dim light. The Y-maze consisted of three arms (40cm long, 5cm wide, and 10cm high), made of dark grey plastic. Mice were placed in arm 1 and allowed to freely explore for 8 minutes. During the 8 minutes, the order of maze arm entry was manually recorded. Spontaneous alternation was defined as entry into three different arms consecutively. Alternation frequency was calculated as: (Number of alternations) ÷ (Total arm entries − 2)(number of spontaneous alternations, divided by the total number of arm entries minus two). Mice that spent the majority of time in the home arm were excluded (n = 2).
2.3. Cell Culture
MRC5 human lung fibroblasts or human dermal fibroblasts were cultured in Dulbecco’s Modified Eagles Medium (DMEM, Sigma Aldrich D5671), supplemented with 10% heat-inactivated Foetal Bovine Serum (FBS, Sigma Aldrich F9665), 100U/ml Penicillin and 100mg/ml Streptomycin (Sigma Alrich P4333), and 2mM L-glutamine (Sigma Aldrich G7513) in 5% CO2 at 37oC.
2.4. Cell Viability Assays
For individual ingredients, senescent MRC5 cells were generated by 20 Gy X-ray irradiation in situ and cultured for 10 days before treatment. Non-senescent cells were plated 24 hours prior to exposure. Serial dilutions were prepared for dose-response curves using appropriate solvents (DMSO, ethanol, or water, with the solvent percentage equal across all dilutions; Supplementary Table S3). Cells were treated for 72 hours, stained with crystal violet (0.2% in 1% ethanol), washed, dried, solubilized in 1% SDS, and absorbance measured at 590 nm (FLUOstar Omega, BMG).
For the multi-ingredient nutraceutical, senescent and non-senescent fibroblasts were co-cultured after stable transduction with pSLIEW and mCherry respectively [28]. Senescent cells were irradiated (20 Gy) 10 days prior, and non-senescent cells were added 24 hours before treatment. Cultures were treated for 72 hours with nutraceutical stock solutions prepared in ultrapure water/ethanol (see ‘Senostatic Measures’ section). Imaging was performed pre- and post-treatment (Leica DMi8). To determine cell size while also quantifying the number of cells per field, cell area was quantified using ImageJ, and divided by the number of cells, to give average cell size per field. Reduction in senescent cell number indicated senolytic activity, and decreased senescent cell size suggested senostatic effects (Rapamycin served as positive control; Supplementary Figure S1).
2.5. Senostatic Measures
Senescent MRC5 cells were treated with filter-sterilized multi-ingredient nutraceutical for 72 hours. 1 packet of the multi-ingredient nutraceutical provided by the Movos Labs (12.9g) was dissolved in 1l of ultrapure water (Milli-Q) or ethanol, and the treatment doses chosen were 1/10 dilution in water, and 1/100 dilution in ethanol, based on the results on their effects on cell sizes (Figure 4).
Nuclear size was assessed by DAPI staining and ImageJ analysis. Reactive oxygen species (ROS) production was measured as hydrogen peroxide release rates using Amplex Red (AR) assay (5 µM AR, 2 U/ml HRP; 1 h incubation in the culture media), followed by fluorescence reading (Ex 544 nm/Em 590 nm) using a Fluostar Omega (BMG Labtech). Background fluorescence was subtracted using blanks, and H₂O₂ standards by using known amounts H₂O₂ were used for calibration. Data was normalised to cell number.
IL-6 secretion was quantified using DuoSet Human IL-6 ELISA (Biotechne DY206) following manufacturer’s protocol. Media was collected after 24 hours of serum-free incubation post-treatment. Absorbance was measured at 450 nm and 540 nm using a Fluostar Omega (BMG Labtech), and the concentrations were calculated from standard curves.
2.6. Statistics
All statistics and graphics were performed using GraphPad Prism 10.3. One-way analysis of variance, and nonlinear regression analysis tests were performed using GraphPad Prism 10.3. Survival was assessed using Kaplan-Meyer curves and log-rank analysis.
3. Results
3.1. Dose Optimization for the Multi-Ingredient Nutraceutical
To determine an appropriate dose for long-term administration, food intake was measured in 20 young male C57Bl/6J mice for 8 days on standard dry pellets and for 7 days after switching to soaked food. The switch increased daily intake from 2.6 g to 5.3 g (Supplementary Figure S2a). This value was used to calculate amounts of the multi-ingredient nutraceutical required to achieve 1×, 5×, 10×, and 25× the recommended human daily dose per kg body weight (BW). Four mice per group were fed these doses for 14 days, whereas controls received soaked food without the multi-ingredient nutraceutical. BW was monitored daily (Supplementary Figure S2b). Statistical analysis found significant effects of time and individual animal, but not dose (p = 0.312). After 14 days, no differences were observed between groups in BW, grip strength, or body temperature (data not shown). Following necropsy, increased fat depots in all groups were observed, likely due to higher caloric intake from soaked food, but no pathological changes were found. Liver, spleen, and kidney weights were similar across groups. These findings indicate that the multi-ingredient nutraceutical was well tolerated up to 25× the human-equivalent dose. Based on the recommended human-to-mouse conversion factor [32], a dose of 12.3× human daily dose per kg BW was selected for the main study. The composition of the multi-ingredient nutraceutical at this dose for a 40 g mouse is shown in Supplementary Table S4.
3.2. Comparison Between Senolytic and the Multi-Ingredient Nutraceutical Interventions
3.2.1. Survival
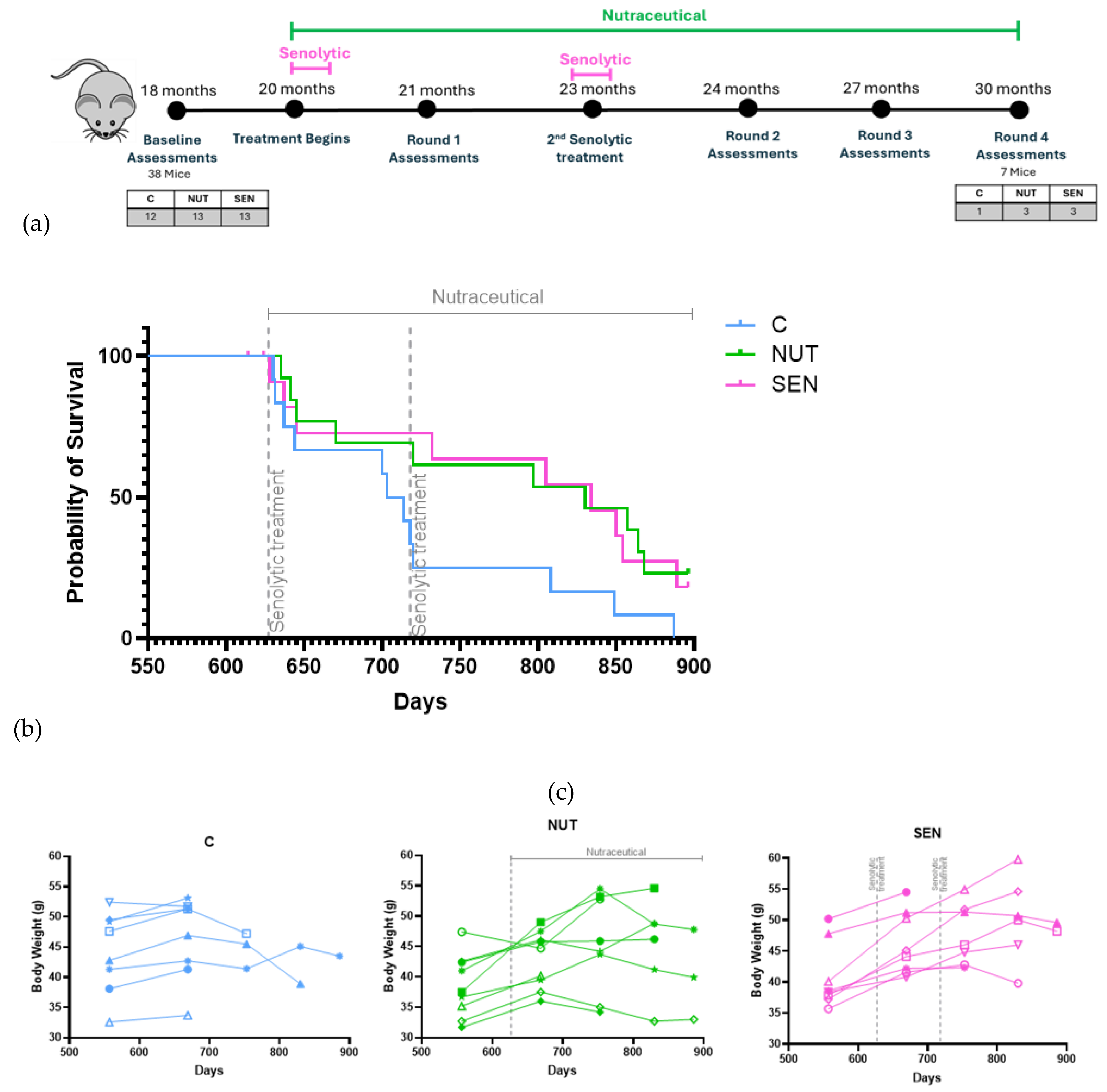
At 18 months, mice were ranked by combined frailty, neuromuscular performance, cognitive score, and BW to minimise baseline variability, as there is substantial variation between mice by this age. Groups were assigned as Control (C), the multi-ingredient nutraceutical (NUT), and senolytic (SEN) (experimental overview: Figure 1a).
All mice received soaked food from 20 months onward; NUT group received food supplemented with the 0.87% w/w multi-ingredient nutraceutical (4.338 g per 500 g food) until study endpoint (30 months). SEN group received navitoclax (0.5 mg/kg BW, gavage) and BAM15 (2.5 mg/kg BW, injection) for 5 consecutive days at 20 and 23 months. On the same days, the mice in groups C and NUT received vehicle gavage and injection. Deaths before treatment initiation (two in SEN group) were censored. Kaplan–Meier analysis showed significant survival improvement for both interventions (Figure 1b). Median survival [95% CI] increased from 703 [637, 720] days in controls to 830 [645, 868] days in N and 834 [637, 889] days in SEN. Notably, the median lifespan of controls (703 days) was ~20% shorter than historical norms for C57Bl/6J (~900 days) [33]. Given that the soaked food protocol doubled caloric intake (2.6 g/day to 5.3 g/day) and increased body weight by 5–10 g, metabolic stress may have accelerated mortality independent of intrinsic ageing. It is well-established that obesity reduces lifespan and increases risks of a variety of age- related conditions [e.g., 34-36]. Therefore, the observed survival benefit should be interpreted as restoring lifespan toward the expected baseline rather than exceeding species maximum.
A cluster of deaths occurred within two weeks of the first senolytic course, affecting all groups. Postmortem findings indicated abdominal infections consistent with seminal vesicle perforation during IP injections. When these deaths were classified as procedure-related and censored from survival analysis to avoid biasing ageing outcomes, consistent with Kaplan–Meier guidelines for excluding deaths unrelated to biological ageing, median lifespans were 719 days for controls, 860.5 days for N, and 852 days for SEN (Supplementary Figure S4), corresponding to 19.7% and 18.5% increases over controls.
3.2.2. Body Weight
Male mice typically reach peak BW at 20–22 months, followed by ~30% decline by 30 months [37]. Retention of body mass predicts survival in ageing mice [37,38,39]. Before the first assessment (21 months), BW increased in most mice, likely due to higher intake of soaked food. After 21 months, BW in controls plateaued or declined, whereas treated mice (NUT and SEN) continued gaining until 24–27 months, suggesting delayed age-related weight loss. Thus, the longevity gains by N and SEN interventions appeared to have been reflected in longitudinal BW data (Figure 1c).
Figure 1.
Effects of a multi-ingredient nutraceutical (NUT) and a senolytic (SEN) on survival in male mice. (a) Design of the experiment. (b) Kaplan-Meier survival curves. Log-rank tests for the difference to controls resulted in p=0.045 (NUT) and p=0.043 (SEN), respectively. (c) Body weights of individual mice. Left (blue): controls (C), middle (Green): The multi-ingredient nutraceutical (NUT), right (pink): Senolytic (SEN) groups.
Figure 1.
Effects of a multi-ingredient nutraceutical (NUT) and a senolytic (SEN) on survival in male mice. (a) Design of the experiment. (b) Kaplan-Meier survival curves. Log-rank tests for the difference to controls resulted in p=0.045 (NUT) and p=0.043 (SEN), respectively. (c) Body weights of individual mice. Left (blue): controls (C), middle (Green): The multi-ingredient nutraceutical (NUT), right (pink): Senolytic (SEN) groups.
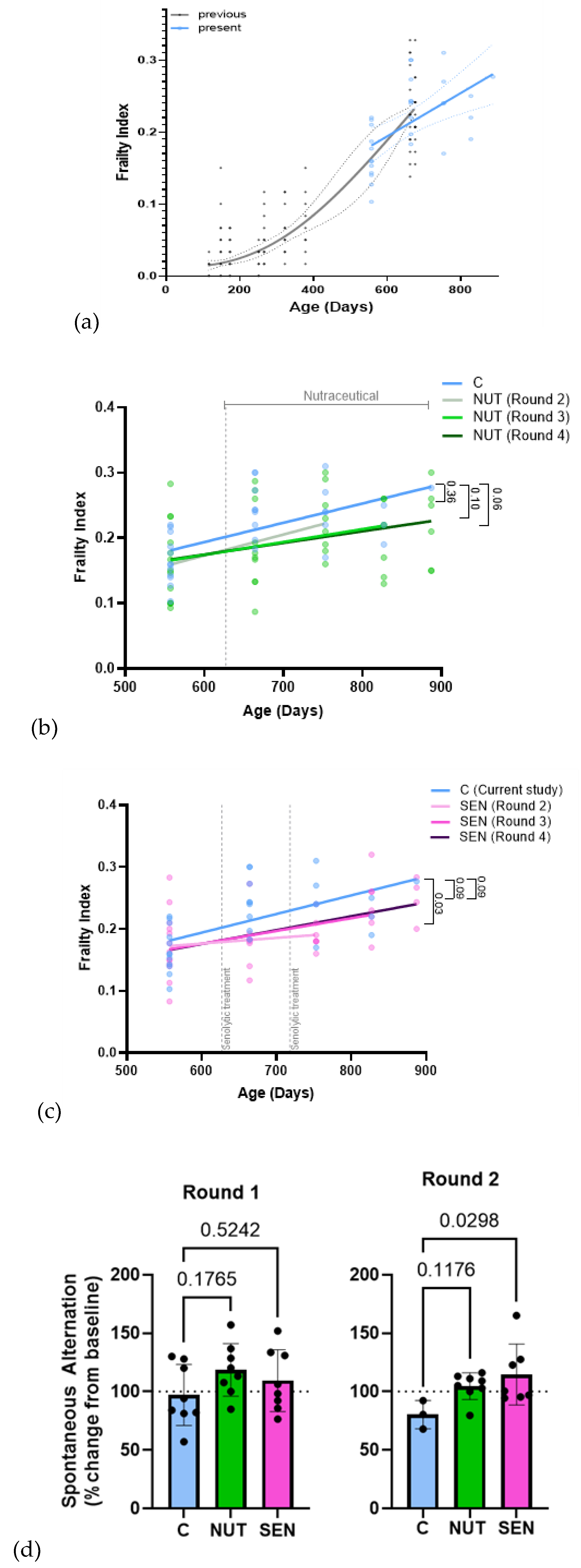
3.2.3. Frailty and Cognition
Frailty indices in controls matched ageing profiles for male C57Bl/6J mice that we previously published [30] (Figure 2a). Both interventions attenuated frailty progression but with distinct patterns: NUT showed gradual improvement over time, approaching statistical significance at 30 months compared with controls (Figure 2b), whereas SEN produced a marked benefit shortly after the second treatment (assessed at 24 months) that diminished thereafter (Figure 2c).
Similarly, SEN improved short-term memory (spontaneous alternation in Y-maze) at 24 months (round 2 assessment) (Figure 2d), while NUT maintained memory performance over time. Baseline ranking resulted in lower initial memory scores for NUT, which improved to control levels but not beyond (Supplementary Figure S3). Limited sample size in the control group prevented comparisons at 27 and 30 months.
Overall, NUT and SEN exhibited distinct trajectories of benefit.
Figure 2.
Health span effects. (a) Comparison between frailty indices in present (blue) and previous [27] (black) controls. All animals were male C57Bl/6J mice. Individual frailty indices (dots), regression lines (solid lines) and 95% confidence intervals (dotted lines) are shown. (b) and (c) Frailty indices in the multi-ingredient neutraceutical (NUT, (b)) and senolytic (SEN, (c)) groups in comparison to controls. Frailty indices for each animal (dots) and linear regression lines from baseline to every assessment time point are indicated to illustrate the kinetic developments. p values for the differences in regression slopes to that of control are shown. (d) Spontaneous alternation in a Y-maze 1 month following the first and second round of senolytic treatment as percentage change from each mouse’s pre-treatment performance. Percentage changes of individual mice are illustrated as dots. Data are mean ± SD, n≥3. ANOVA p values for differences between controls and colour-coded treatment group are indicated.
Figure 2.
Health span effects. (a) Comparison between frailty indices in present (blue) and previous [27] (black) controls. All animals were male C57Bl/6J mice. Individual frailty indices (dots), regression lines (solid lines) and 95% confidence intervals (dotted lines) are shown. (b) and (c) Frailty indices in the multi-ingredient neutraceutical (NUT, (b)) and senolytic (SEN, (c)) groups in comparison to controls. Frailty indices for each animal (dots) and linear regression lines from baseline to every assessment time point are indicated to illustrate the kinetic developments. p values for the differences in regression slopes to that of control are shown. (d) Spontaneous alternation in a Y-maze 1 month following the first and second round of senolytic treatment as percentage change from each mouse’s pre-treatment performance. Percentage changes of individual mice are illustrated as dots. Data are mean ± SD, n≥3. ANOVA p values for differences between controls and colour-coded treatment group are indicated.
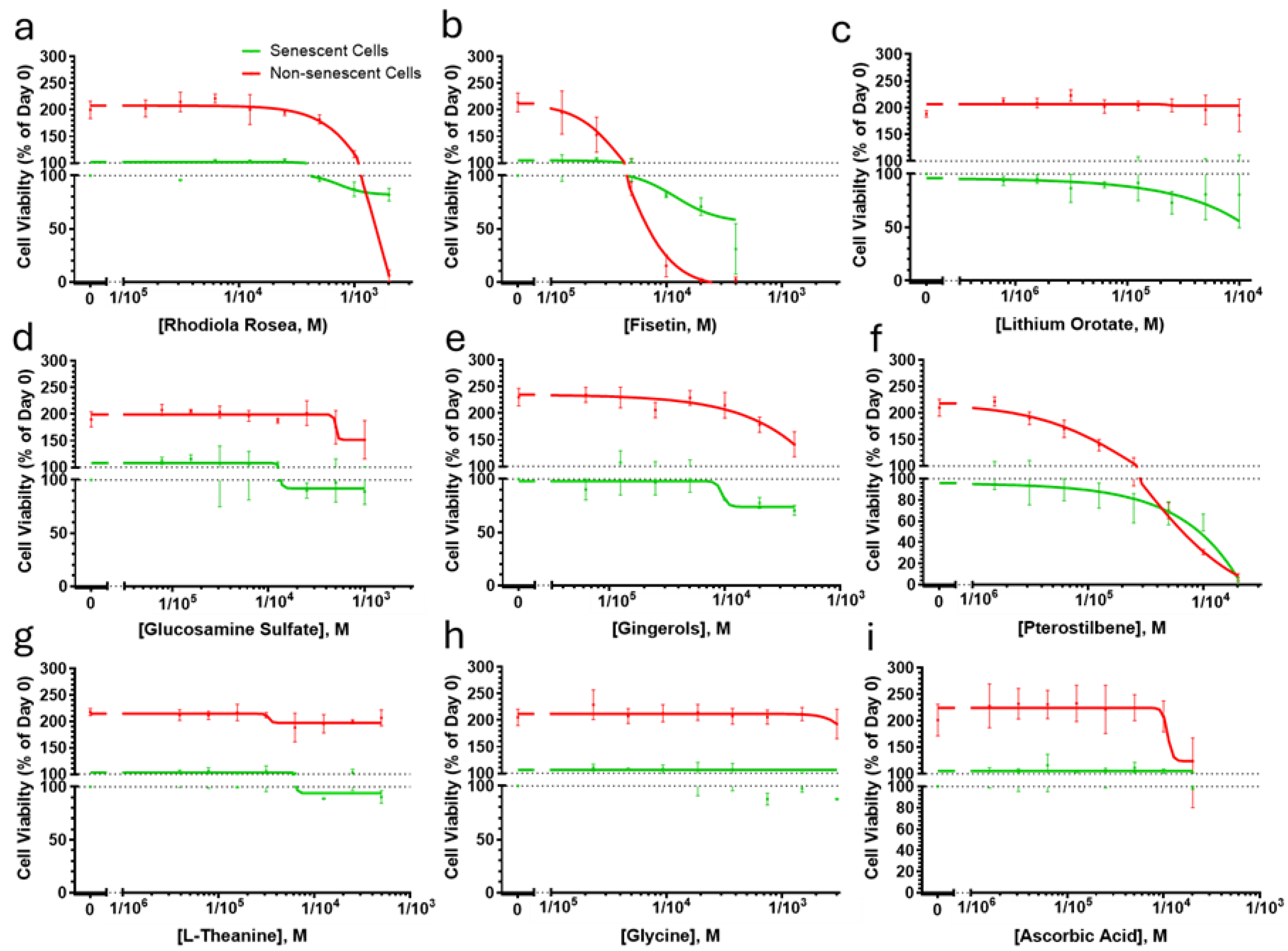
3.3. Senotheapeutic Properties of the Multi-Ingredient Nutraceutical at a Cellular Level In Vitro
3.3.1. Senolytic Activity
Comparing the trajectories of frailty indices changes, the multi-ingredient nutraceutical is unlikely to function as senolytic. Indeed, none of the nutraceutical’s individual ingredients (Figure 3a–i), nor the complete formulation (Figs 4c,d), showed senolytic activity in human fibroblasts across a wide concentration range, except calcium alpha-ketoglutarate, which was excluded due to poor solubility. Fisetin, previously reported as a weak senolytic in ERCC1-/- fibroblasts and IMR90 human fibroblasts [7], did not exhibit senolytic effects in MRC5 fibroblasts in this study.
Figure 3.
The individual ingredients of the multi-ingredient nutraceutical do not act as senolytic in human fibroblasts. Senescent (orange) and non-senescent (green) human fibroblasts were treated for 3 days with the indicated concentrations of its active ingredients (a-i) and cell viability was measured using crystal violet. Titration curves are presented as moving averages ± SEM from between 2 and 4 experiments per ingredient.
Figure 3.
The individual ingredients of the multi-ingredient nutraceutical do not act as senolytic in human fibroblasts. Senescent (orange) and non-senescent (green) human fibroblasts were treated for 3 days with the indicated concentrations of its active ingredients (a-i) and cell viability was measured using crystal violet. Titration curves are presented as moving averages ± SEM from between 2 and 4 experiments per ingredient.
3.3.2. Senostatic Indicators
Although senolytic activity was absent, the multi-ingredient nutraceutical influenced senostatic markers. Cell size can be indicative of a senostatic activity [40] as illustrated with the senostatic effects of rapamycin (Supplementary Figure S1). In co-culture assays, increasing concentrations reduced senescent cell size (Figure 4e,f) without preferential elimination (Figure 4c,d), regardless of solvent (ethanol (Figure 4c,e) or water; Figure 4d,f)). Fisetin alone also reduced cell size at 1–10 µM (Figure 4g).
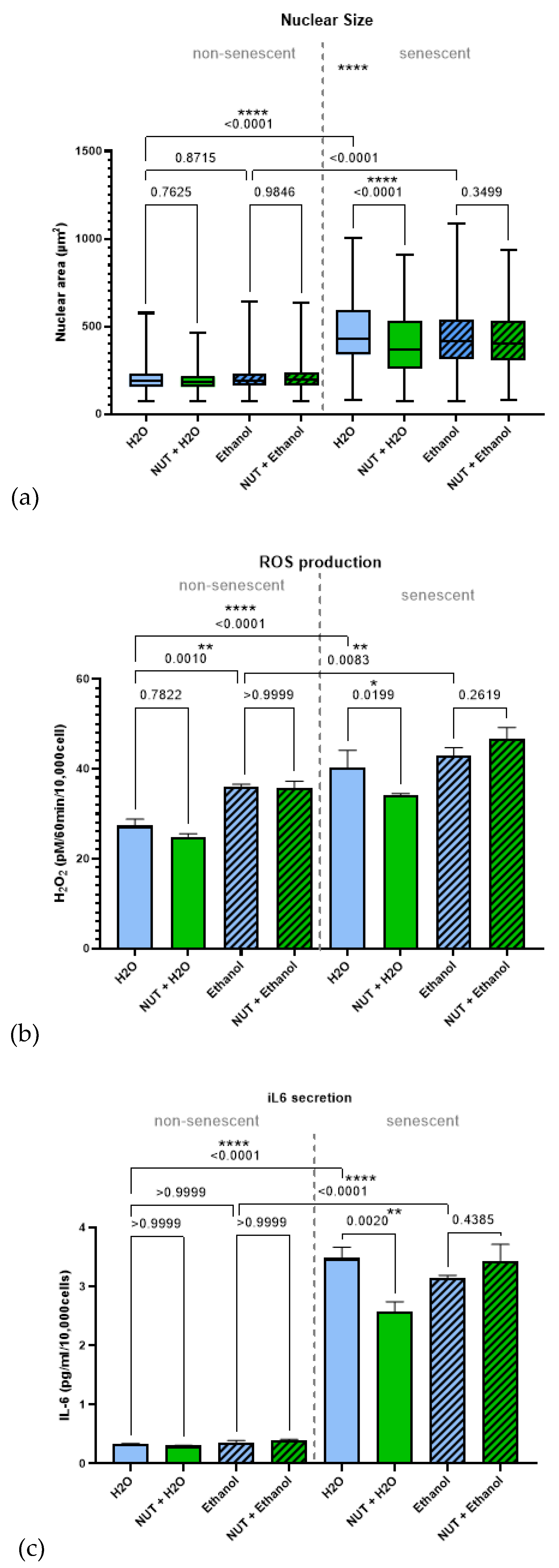
Further assays confirmed senostatic effects: treatment with the water-dissolved multi-ingredient nutraceutical (1/10 dilution; chosen based on cell size data) for 72 hrs significantly decreased nuclear size, ROS production, and IL-6 secretion in senescent fibroblasts (Figure 5a–c), although the ethanol-dissolved multi-ingredient nutraceutical did not (1/100 dilution; chosen based on cell size data). Non-senescent cells were unaffected by either preparation of the multi-ingredient nutraceutical. Ethanol vehicle controls showed higher ROS production compared to the one in water even in non-senescent cells (Figure 5b), suggesting solvent-related adverse effectsas a possible confounding factor for the ethanol-dissolved multi-ingredient nutraceutical treatment. Collectively, these findings indicate that the multi-ingredient nutraceutical, when dissolved in water, exerts senostatic effects at a cellular level in vitro.
Figure 4.
The multi-ingredient nutraceutical and its component fisetin affect cell size in senescent human fibroblasts. (a) Representative micrographs of the senescent/non-senescent human fibroblast co-culture before (day 0) and after (day 3) treatment with the multi-ingredient nutraceutical at 1/100 dilution in ethanol. Senescent cells are labelled in green and non-senescent in red. (b) Higher magnification images of three senescent cells from (a) at start (top) and end (bottom) of treatment with their respective areas indicated. (c) Effect of the complete multi-ingredient nutraceutical dissolved in ethanol on numbers of senescent (orange) and non-senescent (blue) cells. (d) Effect of the complete multi-ingredient nutraceutical dissolved in H2O on numbers of senescent (orange) and non-senescent (blue) cells. (e) Effect of the complete multi-ingredient nutraceutical dissolved in ethanol on average sizes of senescent (orange) and non-senescent (blue) cells. (f) Effect of the complete multi-ingredient nutraceutical dissolved in H2O on average size of senescent (orange) and non-senescent (blue) cells. (g) Effect of Fisetin diluted in DMSO on average size of senescent cells. Data are mean ± SEM from 3 technical repeats.
Figure 4.
The multi-ingredient nutraceutical and its component fisetin affect cell size in senescent human fibroblasts. (a) Representative micrographs of the senescent/non-senescent human fibroblast co-culture before (day 0) and after (day 3) treatment with the multi-ingredient nutraceutical at 1/100 dilution in ethanol. Senescent cells are labelled in green and non-senescent in red. (b) Higher magnification images of three senescent cells from (a) at start (top) and end (bottom) of treatment with their respective areas indicated. (c) Effect of the complete multi-ingredient nutraceutical dissolved in ethanol on numbers of senescent (orange) and non-senescent (blue) cells. (d) Effect of the complete multi-ingredient nutraceutical dissolved in H2O on numbers of senescent (orange) and non-senescent (blue) cells. (e) Effect of the complete multi-ingredient nutraceutical dissolved in ethanol on average sizes of senescent (orange) and non-senescent (blue) cells. (f) Effect of the complete multi-ingredient nutraceutical dissolved in H2O on average size of senescent (orange) and non-senescent (blue) cells. (g) Effect of Fisetin diluted in DMSO on average size of senescent cells. Data are mean ± SEM from 3 technical repeats.
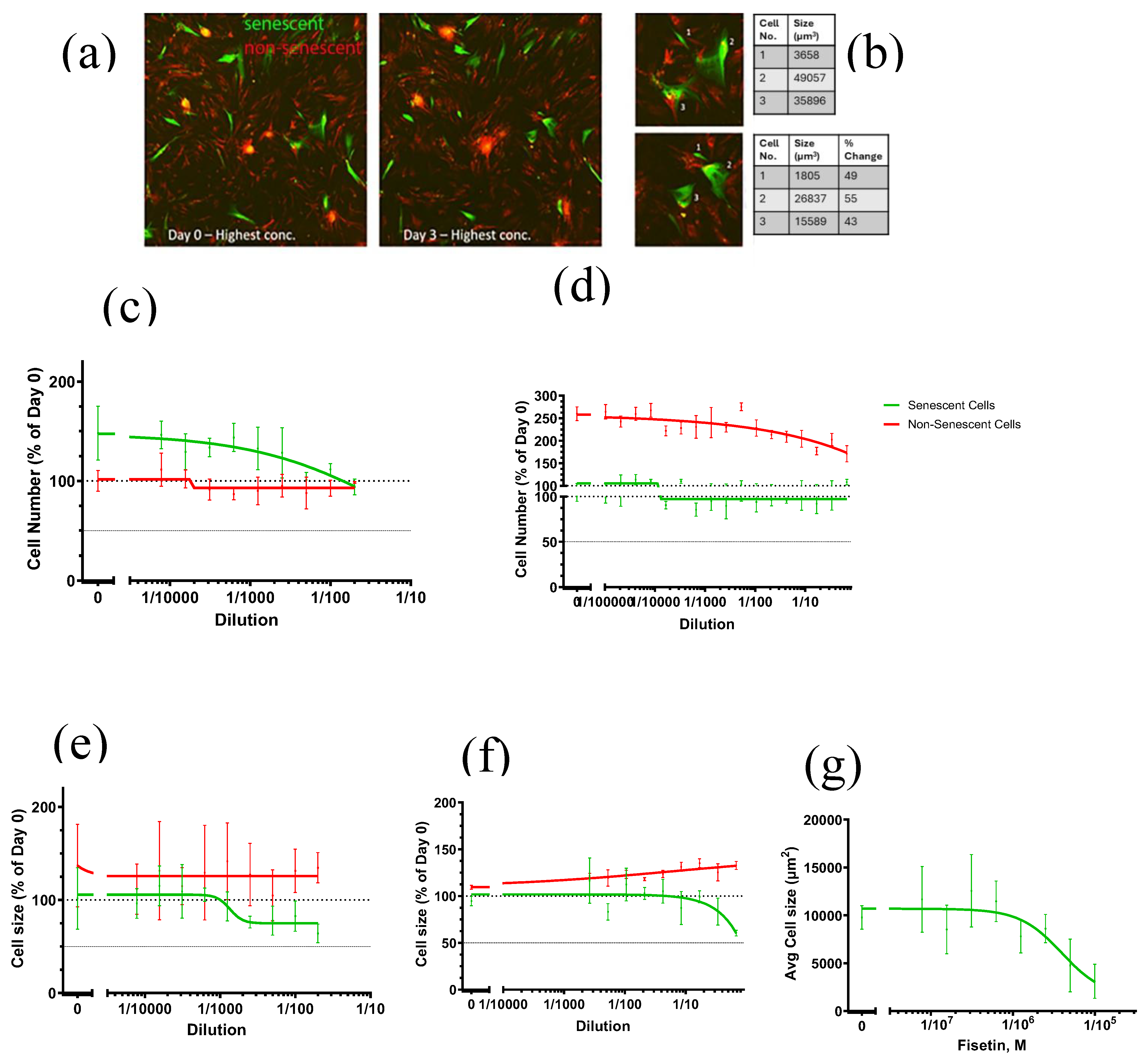
Figure 5.
The multi-ingredient nutraceutical exhibits a senostatic effect on human fibroblasts. Human fibroblasts were irradiated with 20Gy X-ray irradiation and cultured for 10 days to allow senescence development. Cells were then treated for 3 days with either vehicle control (H2O or Ethanol) or the multi-ingredient nutraceutical which was dissolved in 1/10 in H2O, or 1/100 in Ethanol. Cells treated with Ethanol are indicated by stripped pattern on graph. (a) Average nuclear size. (b) ROS production. (c) IL6 secretion. Data are mean ± SEM from 3 technical repeats. ANOVA p values of individual comparisons are indicated.
Figure 5.
The multi-ingredient nutraceutical exhibits a senostatic effect on human fibroblasts. Human fibroblasts were irradiated with 20Gy X-ray irradiation and cultured for 10 days to allow senescence development. Cells were then treated for 3 days with either vehicle control (H2O or Ethanol) or the multi-ingredient nutraceutical which was dissolved in 1/10 in H2O, or 1/100 in Ethanol. Cells treated with Ethanol are indicated by stripped pattern on graph. (a) Average nuclear size. (b) ROS production. (c) IL6 secretion. Data are mean ± SEM from 3 technical repeats. ANOVA p values of individual comparisons are indicated.
4. Discussions
The shortened control lifespan (703, or 719 days if likely procedure-related deaths were censored) compared to historical norms (~900 days) warrants explicit discussion. We hypothesise that the soaked food protocol, which increased caloric intake and body weight, induced obesity and metabolic stress [27], creating a premature ageing phenotype. Under these conditions, the multi-ingredient nutraceutical and navitoclax/BAM15 did not extend maximum lifespan but acted to enhance metabolic resilience, rescuing survival compromised by environmental stress. In addition, we acknowledge an experimental design limitation which resulted in likely procedure-related deaths by intraperitoneal injections during the first round of senolytic treatment. Blind Intraperitoneal injection in aged males with enlarged seminal vesicles carries substantial risk. Future studies should employ safer administration routes, such as oral gavage or subcutaneous injection, to minimise technical mortality. According to survival analysis standards, censoring the deaths which were unrelated to intrinsic ageing processes could be justified, and when censored, median lifespans were 719 days for controls, 860.5 days for N, and 852 days for SEN (Supplementary Figure S4), corresponding to 19.7% and 18.5% increases over controls. Thus, relative treatment effects appear robust despite this confounder.
Healthspan outcomes differed by intervention type and schedule. Continuous multi-ingredient nutraceutical intervention slowed frailty progression over time, approaching significance after 10 months, delayed age-dependent weight loss, and preserved short-term memory, while controls declined. In contrast, senolytic benefits were most pronounced shortly after the second treatment (24 months) but diminished thereafter. This feature contrasts with our previous findings where a single senolytic course, either first-generation senolytic (navitoclax or dasatinib+quecetin) [19] or the navitoclax/BAM15 combination used here [28], produced sustained improvements in prematurely ageing mice after sublethal whole-body irradiation. Irradiation accelerates senescence burden, doubling the rate of frailty progression [30], but one senolytic course normalised the frailty progression for at least a year [19,28]. Naturally aged mice, that are expected to have ongoing senescent cell accumulation, may require repeated senolytic dosing for durable benefit. Our approach, reducing navitoclax dose by two orders of magnitude with BAM15 co-treatment while maintaining efficacy, may offer a promising strategy for safer senolytic therapy.
The multi-ingredient nutraceutical showed no senolytic activity in vitro, either as a whole or as individual components, including fisetin, which has been reported as a weak senolytic in ERCC1−/− and IMR90 cells [7]. Our intervention delivered fisetin at ~18 mg/kg BW, far below doses (100–500 mg/kg) previously linked to senolytic effects or lifespan extension [7,41]. Nonetheless, comparable oral doses (~20 mg/kg) have shown cognitive benefits [42] and antiviral protection [43], suggesting possible contribution to observed outcomes. Importantly, the multi-ingredient nutraceutical exhibited senostatic properties in vitro: treatment reduced nuclear size, ROS release, and IL-6 secretion in senescent fibroblasts, without affecting non-senescent cells. These effects align with in vivo observations where continuous intervention with the multi-ingredient nutraceutical slowed frailty progression, resembling patterns seen with metformin, a known senostatic, in our prior work [19].
Unlike senolytics, which may produce rapid effects by clearing senescent cells, senostatics may exert gradual effects by modulating ageing hallmarks such as mitochondrial dysfunction and chronic inflammation. This may reduce the generation of new senescent cells and limit harmful bystander signaling from existing ones. While our mechanistic analysis was restricted to in vitro cellular assays, it is plausible that long-term nutraceutical intervention also exerts systemic senostatic effects, for example by enhancing immune-mediated clearance of senescent cells. In addition, other systemic benefits, such as modulation of hormonal signaling or gut microbiome composition, may contribute to improved healthspan independently of senostatic mechanisms. These broader effects warrant investigation. Taken together, combining senolytics to reduce existing senescent burden with sustained senostatic therapy, such as multi-ingredient nutraceuticals, may represent an effective and safe strategy to optimise healthspan, which could be explored for the long-term efficacy.
Our study was limited to male mice due to funding constraints. Nevertheless, it is, to our knowledge, the first to directly compare healthspan and lifespan effects of a defined multi-ingredient nutraceutical with a senolytic regimen. Both interventions, initiated at advanced age in overweight animals, improved survival and showed promise for frailty and cognitive health. This ‘rescue model’ may have an important relevance to modern human ageing than conventional longevity paradigms. Most humans do not live under optimised, pathogen-free, calorically restricted conditions but they experience metabolic stress, obesity, and caloric excess. Interventions that restore normal lifespan and healthspan under such conditions can have immediate translational and public health utility. These findings highlight the need to optimise treatment schedules and provide a strong rationale for mechanistic studies on multi-ingredient nutraceuticals.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Figures S1-S4, and Supplementary Tables S1-S4.
Author Contributions
Conceptualisation, T.vZ., D.B. and S.M.; methodology and investigation, C.B., E.F., and E.L; writing—original draft preparation, T.vZ. and S.M.; writing—review and editing, D.B. and S.M.; supervision, T.vZ. and S.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Medical Research Council UK grant No MR/X50290X/1 to T. vZ. and by a donation from NOVOS Labs to S.M.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee of Newcastle University (Animal Welfare and Ethical Review Body (AWERB) ID 786, November 2019).
Conflicts of Interest
Authors S.M. and T.vZ. are named inventors on a patent describing the combination of BH3 mimetics and mitochondrial uncouplers as senolytics (WO2022053800A1). D.B. is an employee of NOVOS Labs.The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef] [PubMed]
- Brind, J.; Miller, R.A.; Strong, R.; Harrison, D.E.; Macchiarini, F. Dietary Glycine Supplementation Extends Lifespan of Genetically Heterogeneous Mice. Faseb J 2018, 32. [Google Scholar] [CrossRef]
- Shintani, T.; Kosuge, Y.; Ashida, H. Glucosamine Extends the Lifespan of Caenorhabditis elegans via Autophagy Induction. J Appl Glycosci 2018, 65, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; et al. Resveratrol Attenuates Oxidative Stress and Extends Life Span in the Annual Fish Nothobranchius guentheri. Rejuvenation Res 2015, 18, 225–233. [Google Scholar] [CrossRef]
- Toricelli, M.; Evangelista, S.R.; Buck, H.S.; Viel, T.A. Microdose Lithium Treatment Reduced Inflammatory Factors and Neurodegeneration in Organotypic Hippocampal Culture of Old SAMP-8 Mice. Cell Mol Neurobiol 2021, 41, 1509–1520. [Google Scholar] [CrossRef]
- Prasath, G.S.; Pillai, S.I.; Subramanian, S.P. Fisetin improves glucose homeostasis through the inhibition of gluconeogenic enzymes in hepatic tissues of streptozotocin induced diabetic rats. Eur J Pharmacol 2014, 740, 248–254. [Google Scholar] [CrossRef]
- Yousefzadeh, M.J.; et al. Fisetin is a senotherapeutic that extends health and lifespan. Ebiomedicine 2018, 36, 18–28. [Google Scholar] [CrossRef]
- Yang, Q.; et al. Pterostilbene, an active constituent of blueberries, enhances innate immune activation and restricts enterovirus D68 infection. Front Immunol 2023, 14, 1118933. [Google Scholar] [CrossRef]
- Demidenko, O.; et al. Rejuvant®, a potential life-extending compound formulation with alpha-ketoglutarate and vitamins, conferred an average 8 year reduction in biological aging, after an average of 7 months of use, in the TruAge DNA methylation test. Aging (Albany NY) 2021, 13(22), 24485–24499. [Google Scholar] [CrossRef]
- Verdoorn, B.P.; et al. Fisetin for COVID-19 in skilled nursing facilities: Senolytic trials in the COVID era. J Am Geriatr Soc 2021, 69, 3023–3033. [Google Scholar] [CrossRef]
- Dellinger, R.W.; et al. Repeat dose NRPT (nicotinamide riboside and pterostilbene) increases NAD(+) levels in humans safely and sustainably: a randomized, double-blind, placebo-controlled study. NPJ Aging Mech Dis 2017, 3, 17. [Google Scholar] [CrossRef] [PubMed]
- Sandalova, E.; et al. Alpha-ketoglutarate supplementation and BiologicaL agE in middle-aged adults (ABLE)-intervention study protocol. Geroscience 2023, 45, 2897–2907. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; et al. Past and Future Directions for Research on Cellular Senescence. Cold Spring Harb Perspect Med 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, J.L.; Tchkonia, T. Senolytic drugs: from discovery to translation. J Int Med 2020, 288, 518–536. [Google Scholar] [CrossRef]
- Short, S.; et al. Senolytics and senostatics as adjuvant tumour therapy. Ebiomed 2019, 41, 683–692. [Google Scholar] [CrossRef]
- Tavenier, J.; et al. Fisetin as a senotherapeutic agent: Evidence and perspectives for age-related diseases. Mech Age Dev 2024, 222, 111995. [Google Scholar] [CrossRef]
- Alum, E.U.; et al. Targeting Cellular Senescence for Healthy Aging: Advances in Senolytics and Senomorphics. Drug Des Dev Therapy 2025, 19, 8489–8522. [Google Scholar] [CrossRef]
- Hartmann, A.; et al. High-Dose Polyphenol-Rich Nutrition Improves Lipid and Inflammation Profiles and Can Trigger Apoptotic Signaling in Healthy Older Adults (the ErdBEHR Study). Aging Biology 2024, 2. [Google Scholar] [CrossRef]
- Fielder, E.; et al. Short senolytic or senostatic interventions rescue progression of radiation-induced frailty and premature ageing in mice. Elife 2022, 11. [Google Scholar] [CrossRef]
- Punzo, A.; et al. Promising Effects of Novel Supplement Formulas in Preventing Skin Aging in 3D Human Keratinocytes. Nutrients 2024, 16, 2770. [Google Scholar] [CrossRef]
- Edwards, C.B.; Copes, N.; Brito, A.G.; Canfield, J.; Bradshaw, P.C. Malate and fumarate extend lifespan in Caenorhabditis elegans. PLoS One 2013, 8, e58345. [Google Scholar] [CrossRef] [PubMed]
- Massie, H.R.; Aiello, V.R.; Doherty, T.J. Dietary vitamin C improves the survival of mice. Gerontology 1984, 30, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Zarse, K.; Jabin, S.; Ristow, M. L-Theanine extends lifespan of adult Caenorhabditis elegans. Eur J Nutr 2012, 51, 765–768. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; et al. Effect of Sodium Hyaluronate on Antioxidant and Anti-Ageing Activities in Caenorhabditis elegans. Foods 2023, 12. [Google Scholar] [CrossRef]
- Schriner, S.E.; et al. Extension of Drosophila lifespan by Rhodiola rosea through a mechanism independent from dietary restriction. PLoS One 2013, 8, e63886. [Google Scholar] [CrossRef]
- Xu, T.; et al. Longevity-promoting properties of ginger extract in Caenorhabditis elegans via the insulin/IGF-1 signaling pathway. Food Funct 2022, 13, 9893–9903. [Google Scholar] [CrossRef]
- Desmarchelier, C.; et al. Diet-induced obesity in ad libitum-fed mice: food texture overrides the effect of macronutrient composition. Br J Nutr 2013, 109(8), 1518-27–27. [Google Scholar] [CrossRef]
- Fielder, E.P.; et al. Mild Uncoupling of Mitochondria Synergistically Enhances Senolytic Specificity and Sensitivity of BH3 Mimetics. Aging Biol 2024, 1(1), 20240022. [Google Scholar] [CrossRef]
- Finch, C.E.; Girgis, F.G. Enlarged seminal vesicles of senescent C57BL-6J mice. J Gerontol. 1974, 29(2), 134–138. [Google Scholar] [CrossRef]
- Fielder, E.; et al. Sublethal whole-body irradiation causes progressive premature frailty in mice. Mech Ageing Dev 2019, 180, 63–69. [Google Scholar] [CrossRef]
- Whitehead, J.C.; et al. A clinical frailty index in aging mice: comparisons with frailty index data in humans. J Gerontol A Biol Sci Med Sci 2014, 69, 621–632. [Google Scholar] [CrossRef]
- Nair, A.B.; et al. A simple practice guide for dose conversion between animals and human. J basic clin pharm 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Pabis, K.; et al. The impact of short-lived controls on the interpretation of lifespan experiments and progress in geroscience – Through the lens of the “900-day rule”. Ageing Res Rev 2024, 101, 102512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; et al. Obesity-induced oxidative stress, accelerated functional decline with age and increased mortality in mice. Arc Biochem Biophys 2015, 576, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; et al. Life-long consumption of high level of fruits and vegetables reduces tumor incidence and extends median lifespan in mice. Front Nutr 2023, 10, 1286792. [Google Scholar] [CrossRef]
- Hussain, A.; Cho, J.S.; Kim, J.S.; Lee, Y.I. Protective Effects of Polyphenol Enriched Complex Plants Extract on Metabolic Dysfunctions Associated with Obesity and Related Nonalcoholic Fatty Liver Diseases in High Fat Diet-Induced C57BL/6 Mice. Molecules 2021, 26(2), 302. [Google Scholar] [CrossRef]
- Cameron, K.M.; Miwa, S.; Walker, C.; von Zglinicki, T. Male mice retain a metabolic memory of improved glucose tolerance induced during adult onset, short-term dietary restriction. Longev Healthspan 2012, 1, 3. [Google Scholar] [CrossRef]
- Liao, C.Y.; et al. Fat maintenance is a predictor of the murine lifespan response to dietary restriction. Aging Cell 2011, 10, 629–639. [Google Scholar] [CrossRef]
- Jurk, D.; et al. Chronic inflammation induces telomere dysfunction and accelerates ageing in mice. Nat Commun 2014, 2, 4172. [Google Scholar] [CrossRef]
- Chadha, Y.; Khurana, A.; Schmoller, K.M. Eukaryotic cell size regulation and its implications for cellular function and dysfunction. Physiol Rev 2024, 104(4), 1679–1717. [Google Scholar] [CrossRef]
- Mahoney, S.A.; et al. Intermittent supplementation with fisetin improves arterial function in old mice by decreasing cellular senescence. Aging Cell 2024, 23, e14060. [Google Scholar] [CrossRef]
- Maher, P.; et al. Flavonoid fisetin promotes ERK-dependent long-term potentiation and enhances memory. Proceedings of the National Academy of Sciences 2006, 103. [Google Scholar] [CrossRef]
- Camell, C.D.; et al. Senolytics reduce coronavirus-related mortality in old mice. Science 2021, 6552. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



