
Submitted:
26 December 2025
Posted:
29 December 2025
You are already at the latest version
Abstract
Methanogens, also known as methanogenic archaea, are among the most ancient and widespread microorganisms, despite their particular requirements for growth. These oxygen-sensitive microorganisms have impacted climate and biogeochemical cycles throughout Earth’s history, although their specific roles in the long-term carbon cycle remain little explored. Methanogens evolved early during Earth’s history, likely during the Archaean Eon, in layered benthic microbial communities called microbial mats. These ancient mats, when lithified, form microbialites that represent some of the earliest evidence of life in the fossil record dating back > 3.5 Gy. Contemporary microbial mats experience a wide range of fluctuating conditions, including dramatic diel shifts in oxygen, sulfide, redox, temperature, salinity and pH. Methanogens are an integral part of marine and freshwater microbial mats and have been identified in the oxic zone of these sedimentary ecosystems; however, their adaptations to apparently unfavorable conditions and their role in long-term CO2 sequestration through precipitation of carbonate are unclear. Furthermore, the importance and coevolution of methanogens and microbial mats may explain the global role these organisms had on Earth’s major climate events during the Archean and Proterozoic eons, notably in the ending of icehouse periods and recovery of mats following mass extinctions – often in conditions with low or no oxygen. In addition to an important role in the evolution of our planet, methanogens may also produce biosignatures that are relevant for astrobiology research [and space exploration]. This review will discuss the diversity, physiology, and ecology of methanogens in order to clarify their role in biogeochemical processes through geologic time.
Keywords:
methanogens
; archaea
; anaerobes
; microbial mats
; early life
; exobiology
; climate regulation
; mass extinctions
1. The Biogeochemistry of Methanogens
1.1. Introduction
Methanogens are strict anaerobes with highly specialized biochemistry. All methanogens belong to the domain Archaea and produce methane as an end-product of their metabolism. Methanogenesis serves as their primary energy-conserving pathway, an oxygen-independent process at the limits of what is thermodynamically possible [1,2,3,4]. In addition to methane, some species also produce carbon dioxide as an end-product, contributing significantly to greenhouse gas emissions and thereby regulating the Earth’s temperature [2,5], and potentially that of other planets [6].
For most of geological history, prokaryotes have dominated our planet [7] filling a diverse array of ecological niches [8,9] Methanogens are a prime, yet understudied example of an ancient and versatile group of microorganisms, that thrive in diverse environments, sometimes under poly-extreme conditions [3,10]. Their metabolic versatility, unique biogeochemistry, and interactions with other microorganisms, such as fermenters, syntrophs, methane oxidizers, and sulfate-reducers, may have facilitated microbial life to develop throughout Earth’s history [11,12]. Furthermore, their exemplary ecological success through geologic time and space (i.e., distribution across diverse and extreme conditions) suggests that methanogens could survive conditions elsewhere in the solar system and possibly beyond, making them relevant model organisms for astrobiology [10,13].
1.2. Methanogenic Habitats
Methanogens are characterized by the production of methane (CH4) as the obligatory product of their oxygen-independent metabolism [3]. Methanogens are found in nearly all anoxic environments on Earth, and many anaerobic niches, including deep sea, freshwater, and terrestrial sediments, marshes, swamps, permafrost soils, geothermal springs, hydrothermal vents, the rumen of cows and intestines of other mammals (including humans), arthropods, and man-made environments such as landfills, (anaerobic) digesters, and rice paddies [3,10,14,15,16,17,18,19,20,21,22]. Some methanogens have also been found in oxic environments as well [23], such as the surface of biofilms and microbial mats [4,23,24], the upper oxic layer of water bodies [25,26,27], mangroves [28], and oxygenated soils [29]. Survival of methanogens has also been assessed under a variety of exoplanetary, artificial, and space conditions, including Martian soil analogues [30,31], under simulated conditions of ocean worlds (such as Enceladus) [13], on the outside of spacecraft during interplanetary travel [10], and in clean rooms [32].
1.3. Physicochemical Boundaries of Methanogenesis
Methanogens have a unique biochemistry, which makes them well-suited for life in extreme niches [10]. As such, methanogens have been isolated from environments with a wide range of physicochemical and thermochemical conditions (Table 1), including: temperature [33,34], pH [3,35], salinity [16], and pressure gradients [31,34,36,37,38]. Some methanogens can also withstand desiccation [31], freeze/thaw cycles [33], and survive high doses of ionizing and UV radiation [39]. Methanogens are found in a wide range of extreme environments and tolerate multiple extreme conditions, making them versatile polyextremophiles [4,10,14,16]. For example, barophilic and thermophilic methanogens, members of the Methanopyrus genus, Methanocaldococcus jannaschii, and Methanothermococcus thermolithotrophicus, have been isolated from hydrothermal vent sites, with Methanopyrus kandleri strain 116 being the most heat-tolerant microorganism cultured, growing at temperatures up to 122oC and pressures up to 20 MPa [34]. On the opposite extreme, psychrophilic methanogens are abundant in wetlands and arctic permafrost [3,10] and can thrive in sub-atmospheric pressure [40]. Soda and saline lakes host alkaliphilic and halophilic methanogens belonging to the genera Methanohalobium, Methanohalophilus and Methanosalsum [16].
Methanogens are widely distributed and well adapted to thrive in microbial mat ecosystems [4,41,42,43], where they could be exposed to high and fluctuating concentrations of O2 despite the traditional belief that they are obligate anaerobes or oxygen-sensitive microorganisms [24]. Their unique biology allows them to adapt and thrive in a range of challenging environments. For example, hydrogenotrophic methanogens, under pH and thermal stress, low H2, iron, phosphate, and trace metals can split their metabolic investments to maintain homeostasis or add biomass, a process referred to as ‘uncoupling’ [13]. Some methanogens can undergo reverse reactions of some steps in methanogenesis to maximize their energy yield when substrates for respiration are sparse [15,44,45]. Such mechanisms may have played a role in their evolution through conditions in Earth past, such as through glaciations and the oxygenation of Earth’s atmosphere (see Section 2).

Table 1.
Growth limits for methanogens and examples of species that grow at these limits. Definitions of extremes, in terms of the known limits for life [8,46] and the limits that have been identified for methanogens. The geologic relevance highlights some of the environments on early Earth, in the present, and on other planets where these conditions could be/have been found.
Table 1.
Growth limits for methanogens and examples of species that grow at these limits. Definitions of extremes, in terms of the known limits for life [8,46] and the limits that have been identified for methanogens. The geologic relevance highlights some of the environments on early Earth, in the present, and on other planets where these conditions could be/have been found.
| Environmental condition | Extremophile type (and growth definition(s)) | Growth limits for Methanogens | Examples of species found within extreme |
Geologic relevance |
|---|---|---|---|---|
| Temperature | 1)Hyperthermophile (>80oC) 2)Thermophile (60-80oC) 3)Psychrophile (<15oC) |
1 & 2) 122oC [2,34]. 3) -2.5oC [47]. |
1 & 2) Methanopyrus kandleri strain 116, Methanopyrus genus, Methanocaldococcus jannaschii, and Methanothermococcus thermolithotrophicus [34]. 3) Candidatus Methanoflorens stordalenmirensis (freeze/thawing environments) [33], Methanococcoides burtonii [47]. |
1) Hydrothermal vents (modern day [34], potential vents of Europa and Enceladus [13], Hot springs/geothermal springs [20]. 2) Archean Oceans [48]. 3) Permafrost and ice; modern day and during glaciation events, like at the last glacial maxima; deep sediments [33,49], Terrestrial surface of Mars [36], Oceans of Europa and Enceladus [10,46]. |
| Radiation | 1) Exposure to UV 2) Ionizing radiation |
1) 24 hours of 254 nm UV radiation [50]. | 1) Methanococcus maripaludis and Methanobacterium formicicum [50]. 2) Methanoscarcina spp. [50]. |
1) High UV radiation during the Archean [46]; surface of Mars [39]. 2) Ionizing radiation from Ur decay in nuclear geysers, possible location for the origin of life [51]. |
| Light | 1) Exposure to visible light (380-750 nm) | 1) Unknown limit; growth inhibition, particularly in the blue region of the spectrum (≈370–430 nm) [52]. Reduced sensitivity in association with photosynthetic bacteria [53]. |
1) Methanosarcina and Methanocella spp. (observed in soil) [53]; Methanosarcina barkeri in a cocultured biofilm with Synechocystis spp. strain PCC6803 [23]. | 1) Surface environments on Earth (from ancient to modern day). |
| Salinity | 1) Halophile (2-5M NaCl) | 1) Moderate/extreme halophiles growth 2-3M [16]. |
1) Methanosarcinales spp., Methanohalobium, Methanohalophilus and Methanosalsum spp., Methanonatronarchaeum thermophilum and Candidatus Methanohalarchaeum thermophilum [16,38]. | 1) Archean oceans; 1.5–2 times modern salinity [54]; Saline lakes [16]. |
| Pressure | 1) Survivability to low pressure 2) Barophile (obligate high pressure)/ Piezophile (adaptation to high pressure) |
1) Low pressure 6mbar – 143 mbar [37]. 2 & 3) 200 bar [34], 500 bar [38]. |
1) Methanothermobacter wolfeii, Methanosarcina barkeri, Methanobacterium formicicum, Methanococcus maripaludis [31,36,37]. 2) Methanopyrus kandleri strain 116, Methanopyrus genus, Methanocaldococcus jannaschii, and Methanothermococcus thermolithotrophicus [34,37]. |
1) Terrestrial surface of Mars [37]. 2) Hydrothermal vents (modern day [34], and possible location for the origin of life [55]; Deep Marine Sediments [56]. |
| pH | 1) Alkaliphile 2) Acidophile |
1) pH > 10.2 [3,35]. 2) pH < 3 [3]. |
1) Methanohalobium, Methanohalophilus and Methanosalsum [35]. | 1) Soda lakes [35]. |
| Hydrogen concentration | 1) High pH2 2) Low pH2 3) Fluctuations in H2 concentration |
1) > 6x10-2 bar [57]*. 2 & 3) As low as 0.1 Pa pH2 [58]; fluctuating H2 (from low pH2 to overpressure) [14]. |
1) Species name unknown. Plausibly, early hydrogenotrophs. 2 & 3) Methanobacterium bryantii, Methanoculleus bourgensis MAB1, and Methanosarcina barkeri [58]; Methanobrevibacter spp., Methanomicrobium spp., and Methanosarcinales [14]. |
1) Earth’s secondary atmosphere [57]. 2) Titan [59]. 3) Cow rumen [14]; deep biosphere [58]. |
| Oxygen | 1) Aerotolerant (tolerates some (high to low) amount of O2) | 1) Unknown limit; exposure up to 300 μM O2 in lithifying microbial mat [60]. | 1) Methanosarcina barkeri [23]; Methanogens in microbial mats [24]. Class II methanogens [61]. | 1) Euxinic oceans of Proterozoic [48]; Modern day (and ancient) biofilms and microbial mats [23,24]. |
| Metals | 1) Arsenic 2) Cadmium |
1) Unknown limit; Methane production by methanogens observed under high arsenic concentrations (0.8-1.5 mM) in microbial mats [62,63]. 2) Unknown limit; 10 μM of CdCl2 activates the rate of methanogenesis. ≥ 100 μM of CdCl2 inhibits cell growth [64]. |
1) Methanomassiliicoccus spp. (Chen et al., 2023) 2) Methanosarcina acetivorans [64]. Other divalent metals (Co, Zn, Cu, Fe) also activate the rate of methanogenesis at 10 μM. |
1 & 2) Early oceans [65], rice paddies [66], modern day anoxic microbial mats in the Atacama, Chile [62,63]. |
* pH2 of Earth’s atmosphere at the time of early methanogen evolution.
1.3.1. Temperature Extremes
Temperature on Earth through time and space shows huge variations ranging from extreme cold or psychrophilic conditions for life (growth at < 15 °C) to thermophilic (60-80 °C), and hyperthermophilic (> 80oC) conditions [8]. These temperature extremes not only exist in specific environments (e.g., modern-day hydrothermal vents [34,55]. Arctic and Antarctic permafrost and water (ice) [33,67], and deep and high-altitude sediments [49,68], but also as widespread environmental conditions of the past and present, such as during glaciations (e.g., “Snowball Earth”) [69,70], through mass extinctions (rapid warming/cooling) [71], and in the Archean oceans [54]. Such extremes in temperature are also relevant for potential life in other locations in our solar system, such as on the Martian surface and in the oceans of Europa, Enceladus, and Titan [10,46] and beyond (exoplanets).
Methanogenesis, like all metabolisms, is a temperature-dependent process and therefore both substrate use and community compositions of methanogens are dependent on temperature [2]. For example, at temperatures higher than 30oC, methanogens tend to utilize H2 to produce CH4 rather than acetate as a growth substrate at lower temperatures [2]. Adaptations of methanogens to become psychrophiles, thermophiles, and hyperthermophiles are associated with modifications at the tRNA and protein level [10]. Psychrophilic Methanococcoides burtonii displays increased flexibility of polynucleotides at low temperatures associated with tRNA due to modifications with dihydrouridine [72]. The same study found tRNA modifications (e.g., methylation of nucleosides) in a thermophilic species of archaeon, Stetteria hydrogenophila, which can stabilize tRNA to accommodate growth under high temperatures [72]. At the protein level, Methanococcoides burtonii displays cold shock proteins such as Elongation Factor 2, which is unstable at higher temperatures [47]. Similarly, a so-called “Dead box” RNA helicase gene is expressed in this species at 4oC [73], a process which is also used by the thermophile, Thermococcus kodakarensis, when experiencing “cold” stress at 60oC [74]. Modifications are made to another cold shock protein (Csp), where the CspA homolog domain helps cells cope with cold stress [75]. Similar to adaptations in bacteria, all archaea must maintain membrane fluidity in cold environments by regulating the unsaturation of membrane lipids [76]. Low permeability and liquid crystalline phases of isoprenoid ether lipid membranes at temperatures ranging from 0-100oC are characteristic of archaeal membranes. They allow for stability at a wider range of temperatures than that found in bacteria [77]. Still, there is less membrane lipid variation observed in thermophilic methanogens than in bacteria [77].
1.3.2. UV and Ionizing Radiation
The surface of Mars experiences high doses of UV radiation from the sun, which may restrict extant life to the subsurface and could erase surface biosignatures of ancient life over geologic time [78]. Life on the Archean Earth also experienced higher levels of UV radiation due to the lack of an ozone layer in the upper atmosphere [79]. This condition favors life to develop and evolve in microbial mats [4]. Exposure to UV radiation leads to the limited survival of Methanococcus maripaludis and Methanobacterium formicicum under simulated Mars conditions [39]. While the exact mechanisms by which methanogens adapt to UV stress are unknown, DNA repair mechanisms could play a role [39]. Physical barriers can protect cells from UV radiation, including FeS, which was abundant on early Earth [48], in layers of soil and sediment surrounding the cells [39], through exoplymeric substances (EPS), and in photopigments in microbial mats containing methanogens [4,69]. Methanoscarcina spp. survives radiation and desiccation [50] by deploying enzymes and DNA repair mechanisms to protect against oxidative stress [50].
1.3.3. Visible Light
The sun’s luminosity has increased through geological time, while short-wavelength radiation has decreased [80]. Light that reaches the Earth’s surface is a critical environmental factor that regulates a wide range of biological processes. At the molecular level, visible light (wavelengths: from 380 to 750 nm) is perceived by sensory photoreceptor proteins that detect specific wavelengths and convert light signals into biochemical responses [81,82]. These photoreceptors span a broad region of the electromagnetic spectrum, ranging from near-ultraviolet (near-UV) to near-infrared (NIR) wavelengths [82].
In methanogens, visible light from both solar and artificial sources negatively affects growth, morphology, and physiology, and in some species, it can even trigger cell lysis and death [52]. Conversely, the absence of visible light has long been considered a critical parameter for the optimal cultivation of multiple strains of methanogens isolated from diverse environments [83,84,85].
Recent mesocosm experiments, conducted in artificial ponds containing environmental sediments, showed that methane emissions by methanogens are two- to threefold higher at night compared to daytime levels [86,87,88]. A decade-long monitoring of microbial diversity and functional activity demonstrated that sunlight modulates the production rates of methane and carbon dioxide, indicating sunlight-dependent metabolic changes in methanogens [88]; however, it is still unclear whether this effect is due to temperature fluctuations or direct exposure to sunlight.
Despite their sensitivity to visible light, methanogens coexist with oxygenic photosynthetic bacteria in various natural habitats, including microbial mats, soil crusts, and the aerobic epilimnion of oligotrophic lakes [23,24,53,89,90,91]. The coexistence of methanogens and phototrophs may be more common and widespread in nature than previously recognized [23,92,93]. It is likely that anoxygenic photosynthetic bacteria act as photosensitizers, promoting the CO2-to-CH4 conversion in association with anaerobic methanogens when cocultured in anoxic biofilms [23]. Taken together, these observations suggest that visible light exerts a far more complex influence on methanogens than previously understood, not only constraining their physiology but also enabling novel ecological interactions.
1.3.4. Salinity
Methanogens in the Archean would have evolved in an ocean 1.5-2 times more saline [54] than the average of 35g.kg-1 today [94]. Higher salinity would have affected not only the ecophysiology and evolution of methanogens but also could have aided in the warming of the early oceans [94].
There are two primary mechanisms archaea utilize to maintain osmotic balance in halophilic environments. The first is a so-called ‘salt out’ approach in which an organic osmotic solute, such a glycine betaine, dimethylsulfoniopropionate, ecotine, or hydroxyctine is utilized internally by the cell to maintain lower cellular concentrations of NaCl than in its surroundings [16,43,95,96]. These compatible solutes provide osmotic protection for the cell through the removal of salts without interfering with biochemical processes [16,97]. The second is a ‘salt in’ method, through high intracellular concentrations of K+, which is employed by species of the genus Methanonatronarchaeum [38,95]. This genus contains extremely halophilic methanogens species Methanonatronarchaeum thermophilum and Candidatus Methanohalarchaeum thermophilum, which are novel in terms of their metabolism, utilizing C1 compounds as an electron acceptor and H2 as an electron donor for methanogenesis [16]. These species have abundant UspA family stress response proteins, their membrane lipids contain pleckstrin homology domains, and archaeal histones may reinforce the stability of RNA, DNA, and proteins – all of which may aid in their adaptations to high salt concentrations [16].
1.3.5. Pressure
Studies simulating low-pressure conditions on Mars reveal the survival of methanogens in 6-400 mbar [36,37]. The physiological and physiochemical adaptations of methanogens in low-pressure environments are unknown, due to the lack of relevance in Earth environments. Survival in low-pressure environments is in part attributed to the rigidity and structure of archaeal lipid membranes [36]. Some species, like Methanoscarcina are flexible in their electron donor choice and grow in multicellular clusters, which makes them more resistant to desiccation and possibly low gas pressures [36].
High pressure, or barophilic conditions, are better studied in the context of the Earth’s biosphere, for example, in the deep-sea or in oceanic and terrestrial subsurface sediments [56]. Barophilic pressure conditions often coincide with other extremes, such as hyperthermophilic conditions at hydrothermal vents [34,38,56]. Morphological adaptations to high temperatures are thus present in methanogens growing at high pressures [38,56]. Some species, like Methanococcus thermolithotrophicus, typically coccoidal cells, have been observed to grow in an elongated shape under high hydrostatic pressure [38].
1.3.6. Hydrogen Concentration
Low and high partial pressure of hydrogen (pH2) is found in a variety of environments on Earth and is highly relevant in astrobiology, such as the low pH2 atmosphere of Saturn’s moon Titan [59]. A high pH2 was characteristic of Earth’s early secondary atmosphere, with a pH2 of over 6x10-2 bar [57]; conditions under which early methanogens evolved. The evolution of nitrogen fixation over 3.2 billion years ago [98] may have contributed to an abundance of H2 [99], which could have supported early methanogenesis.
At the community level, methanogens adapt well to fluctuations in H2 found in the rumens of cows and sheep, which change from low concentration H2 to supersaturated conditions [14,100]. Some of the species found in these communities, like Methanosarcinales, lack cytochromes and are associated with short doubling times and mesophilic temperatures [14]. Lithotrophic methanogenesis (i.e., use of H2/CO2; see Section 1.4.1.3) is most common in gut communities [14]. Trace metals, such as nickel used in cytochromes, may aid in adaptation to low H2 conditions [58].
1.3.7. Oxygen Exposure
Methanogens are considered oxygen-sensitive microorganisms and obligate anaerobes, but are exposed to high levels of oxygen in modern-day microbial mats [4,24] and potentially in oxic environments such as surface waters, including those of the Proterozoic eon. Presence of O2 in the environment expanded the redox states of elements and allowed for greater variation in electron donor/acceptor availability (i.e., aerobic metabolism) [101]. Exposure to O2 in biofilms [23], microbial mats [24], and other terrestrial and aquatic systems [25,26,27,29] may challenge the doctrine that methanogens are strict obligate anaerobes: some species are clearly more aerotolerant than previously thought [102]. Ye and colleagues (2024) observed enhanced methanogenesis in lab-grown cocultured biofilms of a cyanobacterium, Synechocystis PCC6803, and a methanogenic archaeaon, Methanosarcina barkeri. During dark cycles, syntropic methanogenesis was driven by inorganic products (H2/CO2) produced through respiration by cyanobacteria, while M. barkeri reduced the pH2, enhancing Synechocystis PCC6803 growth [23]. During the day, active photosynthesis oxygenated the biofilm, triggering the generation of reactive oxygen species (ROS) by M. barkeri, which oxidized methyl radicals, eventually resulting in the abiotic production of CH4 [23,103]. The ROS produced in the presence of O2 can also be used as an organic substrate for CH4 production by M. barkeri [23]. Additionally, diel fluctuations in metabolic activity by M. barkeri established Fe-redox cycling, which enhanced growth [23].
1.3.8. Metals
The abundance and redox state of elements have changed radically through Earth’s evolution, predominantly due to the oxygenation of the oceans and atmosphere [65]. Methanogens can transform the redox state of metals in processes that lead to their solubilization or biomineralization [104]. This dynamic interaction is a response to their need for certain elements to carry out their physiological functions [105]. A considerable number of metals are indispensable for cellular function, given their well-established biological roles. These include Fe, Na, K, Ca, Cu, Zn, Mn, Mo, Co, and Mg [106]. Collectively, these elements constitute the basis of cellular metabolism and are fundamental to processes such as enzymatic catalysis, biological polymer folding, osmoregulation, membrane transport, and signal transduction [107]. Consequently, intracellular biochemistry can produce a “chemical fossil record” that reflects the geochemical conditions of the early Earth [108]. The use of metals can be viewed as a primitive metabolic strategy that has influenced the evolution of protocells since the very origin of life [109]. Indeed, all classes of biological molecules, from nucleic acids to proteins and antibiotics, interact with metal ions, which explains why living cells invest considerable effort in tightly regulating their intracellular concentrations [107].
There is strong scientific evidence that metals (both essential and non-essential) play a crucial role in the global carbon cycle, particularly in the production of methane and the consumption of CO2 [64,110,111,112,113,114,115]. All methanogens require Fe, Ca, Co and Ni for optimal growth, while some also need Zn, Se, Mo and/or W [111,116,117]. Iron can serve as a carrier or reactant in electron transfer, especially in the form of iron-sulfur ([Fe-S]) clusters [105]. The greater reliance on [Fe-S] clusters in methanogens has been suggested to result from increased bioavailability of Fe and S in anoxic early Earth environments (see Section 2.1) [118]. Nickel is an important cofactor present in methyl coenzyme M reductase (Mcr); the final enzyme in the methanogenesis pathway, and is a key component of the cofactor F430 [111]. One hypothesis suggests that the decline in nickel availability around 2.7 billion years ago contributed to the reduction of atmospheric methane levels, thereby weakening the methane greenhouse and paving the way for the Great Oxidation Event (see Section 2.2) [119,120]. Cobalt is found in corrinoid cofactors (cobalamides), which are present within methyltransferases required for the transfer of the methyl group from methyl-tetrahydromethanopterin (methyl-H₄MPT) to coenzyme M (CoM) [121]. Calcium and magnesium are required at low concentrations for methanogenic growth [122,123], but at high concentrations disrupt cellular processes, such as quorum sensing (i.e., cell-to-cell communication) [124] and electron transport [125]. Zinc is present in the heterodisulfide reductase (Hdr), where it plays a structural and catalytic role in the reduction of CoM-S-S-CoB, thereby recycling the cofactors required for the activity of the methyl-coenzyme M reductase enzyme [126]. Molybdenum is involved in catalyzing the first step of hydrogenotrophic methanogenesis through formylmethanofuran dehydrogenase, as well as the Wood-Ljungdahl pathway of CO2 fixation. Similarly, the oxidation of formate to CO2 and hydrogen (H2) is catalyzed by formate dehydrogenase, which is Mo-dependent [127,128]. Under conditions of Mo limitation, tungsten can substitute for molybdenum due to its similar chemical properties [129].
Methanogens exhibit distinct responses to heavy metals. Although some of these are required for growth, others are well-known for their inhibitory effects on these microorganisms. Heavy metals are generally defined as metallic elements with a high atomic weight and a density at least five times greater than that of water [130]. The term “heavy metals” refers to both essential and nonessential trace elements, which may exert toxic effects depending on their intrinsic properties, chemical speciation, and concentration [131]. This definition classifies cadmium, copper, chromium, mercury, nickel, lead, and zinc as heavy metals. The abundance of these heavy elements in the dissolved state may have supported the evolution of methanogens early in Earth’s history [65]. It should be noted that the term “heavy metals” includes certain metalloids, such as arsenic and selenium, based on the historical assumption that heaviness correlates with toxicity [130]. As a consequence of such long-standing interactions, microorganisms are equipped at least with one As-detoxifying mechanism to prevent its toxicity [132]. This phenomenon may reflect the central role of arsenic toxicity in the evolution and survival of life throughout Earth’s history [133,134,135]. Methylation of arsenic, a common detoxification pathway, has been documented in methanogens [66]. Although arsenic can inhibit aceticlastic and hydrogenotrophic methanogenesis [136,137], recent reports indicate that arsenic can also promote aceticlastic methane production at low concentrations (10µM) [138]. The effect of As on methane production seems site-specific. For example, in permanently anoxic microbial mats in the Atacama, Chile, methane was produced in the presence of 0.8-1.5 mM total arsenic [62,63]. A similar stimulating effect has been observed with Cd, which can increase methanogenesis rates up to nine-fold in pure cultures at concentrations of 10 µM [139,140]. In contrast, metals such as copper (Cu) and chromium (Cr) inhibit methanogenesis, although they do not necessarily lead to a significant reduction in biomass [111,141].
1.4. Evolution and Diversity of Methanogenic Pathways
As mentioned, methanogenesis is one of the oldest metabolisms on our planet, with estimates placing the onset no later than 3.8 to 3.5 Gya [6,48,142,143] in Earth’s secondary, pH2-rich atmosphere [48,94]. Phylogenetic analyses suggest that methanogenesis may have evolved earlier, possibly around 4.1 to 3.8 Gya [7], during the late heavy bombardment [144]. They diverged within the kingdom Euryarchaeota ~3.51 Gya [143] based on estimates of the date of horizontal gene transfer (HGT) events between methanogens and cyanobacteria, which were calibrated to chemo- and microfossil evidence [143]. There is limited geochemical evidence of methanogenesis before their divergence within Euryarcheota in the Archean. To date, the only geochemical evidence that places the evolution of methanogenesis during the Archean is CH4-rich fluid inclusions in metamorphosed rocks from the Pilbara craton, dated back to ~3.5 Gy [142] and confirmed by Missbach and colleagues [145], as well as from isotopic signatures of 13C from ~2.7 Ga-old kerogens [146]. Lipid biomarkers of archaeal membranes have been identified in the rock record dating back to 2.7 Gy [147]. However, certain abiotic processes could produce similar isotopic signatures, and the biogenicity of methane in Archean samples remains unresolved [62].
1.4.1. Methanogenic Pathways
Methanogens are a diverse group of microorganisms that belong to several different taxa within domain Archaea [10,143,148]. Methanogens produce CH4 anaerobically [3], however, they are not the only microorganisms to produce this gas [103].
There are three primary pathways associated with methanogenesis, also called the Wolfe-cycle (see Section 1.4.1), but methanogens are often metabolically flexible (i.e., mixotrophic), with some capable of utilizing more than one energy-generating pathway. For example, Methanoscarcina acetivorans is a model organism for aceticlastic methanogenesis, but can also grow on certain C1 compounds, such as methanol, methylamines, and methyl sulfides [15]. While methanogenesis lies on the thermodynamic threshold of life (ΔG0’ = −20 kJ mol−1) [3,149,150], the energetic yield can be modified by combining methane production with other metabolic reactions. Coupling of one of the primary methanogenic pathways with other microbial metabolisms increases the overall energy yield, such as in syntrophic methanogenesis (e.g., with cable bacteria) ([11,151,152], interspecies hydrogen transfer [153,154], and reverse methanogenesis coupled to sulfate reduction (anaerobic methane oxidation) [154,155]. Energetic yield of the various reactions varies, depending on temperature and actual concentration of the reactants and products [2,150,155].
1.4.1.1. Aceticlastic Methanogenesis
Aceticlastic (also referred to as acetoclastic) methanogenesis is perhaps the most common form of methane production in the environment [156,157], as acetate is likely abundant in many habitats [2,15,156]. The aceticlastic pathway is a chemoorganoheterotrophic process of methane production that utilizes acetate (ΔG0’ = -11.0 kJ mol−1) [150] or acetate-like compounds, such as pyruvate or propionate (Equation (1)) [3,150,158,159]. For detailed information on the thermodynamics for various growth substates, see [15,150].
CH3COOH → CH4 + CO2
1.4.1.2. Methylotrophic Methanogenesis
C1 compounds (e.g., methanol, methylamines, methyl sulfides and thiols, methyl halogens, and methylated aromatics) get reduced through the methylotrophic pathway (Equation (2)) (another mode of chemoorganoheterotrophic growth) [3,150,158,159]. The energy yield of these compounds depends largely on the number of methyl groups available; for example, the ΔG0’ of methanol is -290.0 kJ mol−1, dimethylamine is -167.0 kJ mol−1, and trimethylamine is -510.9 kJ mol−1 [2,150].
4CH3OH → 3CH4 + CO2 + 2H2O
1.4.1.3. Lithotrophic Methanogenesis
Lithotrophic methanogenesis (i.e., hydrogenotrophic methanogenesis) is a chemolithoautotrophic pathway where CO2 is reduced (from CO2(g) or HCO3-/CO2 (aq), CaCO3 (S), carbon monoxide, formate, or some alcohols paired with CO2) using H2 as an electron source (Equation (3)) [150,160,161,162]. In some cases, calcium carbonate is used as a source of electron acceptor (i.e., as source of CO2) [160]. The ΔG0’ of equation 3, using H2(g) as an electron donor, is -193.0 kJ mol−1 [2,150].
CO2 + 4H2 → CH4 + 2H2O
In terms of their evolution, hydrogenotrophic pathway are likely more ancient than aceticlastic and methylotrophic methanogenesis [48]. While methylotrophic methanogenesis was likely next to evolve [48]. Hydrogenotrophic methanogenesis may have been supported by the abundance of H2 in an early anoxic atmosphere. Estimates for Archean atmospheric hydrogen concentrations range from a conservative 1000 ppm [48] to 10,000 ppm [5] or even 300,000 ppm [163], which are all well above modern levels of 530 ppb [164]. Lithotrophic methanogens also share hydrogenases with the Last Universal Common Ancestor (LUCA), which suggests LUCA likely utilized H2 as part of its metabolism [48,165,166].
2. Signatures of Methanogens Through Geologic Time
The ecophysiological background of methanogens reviewed above is instrumental in understanding the impact that these microbes have had on the evolution of our planet. From early Earth to modern time, methanogens have altered Earth’s biosphere, atmosphere, and lithosphere. While there are challenges associated with biogeochemical evidence of methanogenesis in deep time (i.e., through the Archean and Proterozoic eons), indicators of methanogen activity in the rock record include depleted δ13C isotopes -27‰ to -38‰ [167], of which the activity of biological methane oxidation (both anaerobic and aerobic) can alter these signatures [48], further fractionating 13C isotopes [146]. In some cases, the isotopic fractionation effect deviates, e.g., hyperthermophilic methanogenesis in environmental extremes of high heat (~122oC) and hydrostatic pressure (> 40 MPa), yields a fractionation of < −12‰ [34]. Methanogen-specific lipids have been used as an indicator of methanogens in sedimentary structures, both ancient and modern [142,168,169]. It should be noted that abiotic fractionation can also produce a negative δ13C-methane value. In Fischer-Tropsch synthesis reactions, the methane that forms can have isotopic C values as low as -45‰, depending on the iron mineral used in the reaction [170].
2.1. The Archean Eon (4-2.5 Gy)
The atmosphere of the Archean was defined by pervasive anoxia. Methane was delivered to early Earth’s atmosphere by volcanism (which released H2, CH4, CO2, and NH3), metamorphic reactions such as serpentinization (Equation (4)) [171] and Fischer-Tropsch synthesis reactions, which often concur in hydrothermal vents. (Equation (5)) [170], and through impacts with comets and asteroids [6,172], but in addition, and largely, through biogenic CH4 production [6,48,142,167].
6Fe2SiO4 + 7H2O → 3Fe3Si2O5(OH)4 + Fe3O4 + H2
(2n+1)H2 + nCO → CnH2n+2 + nH2O
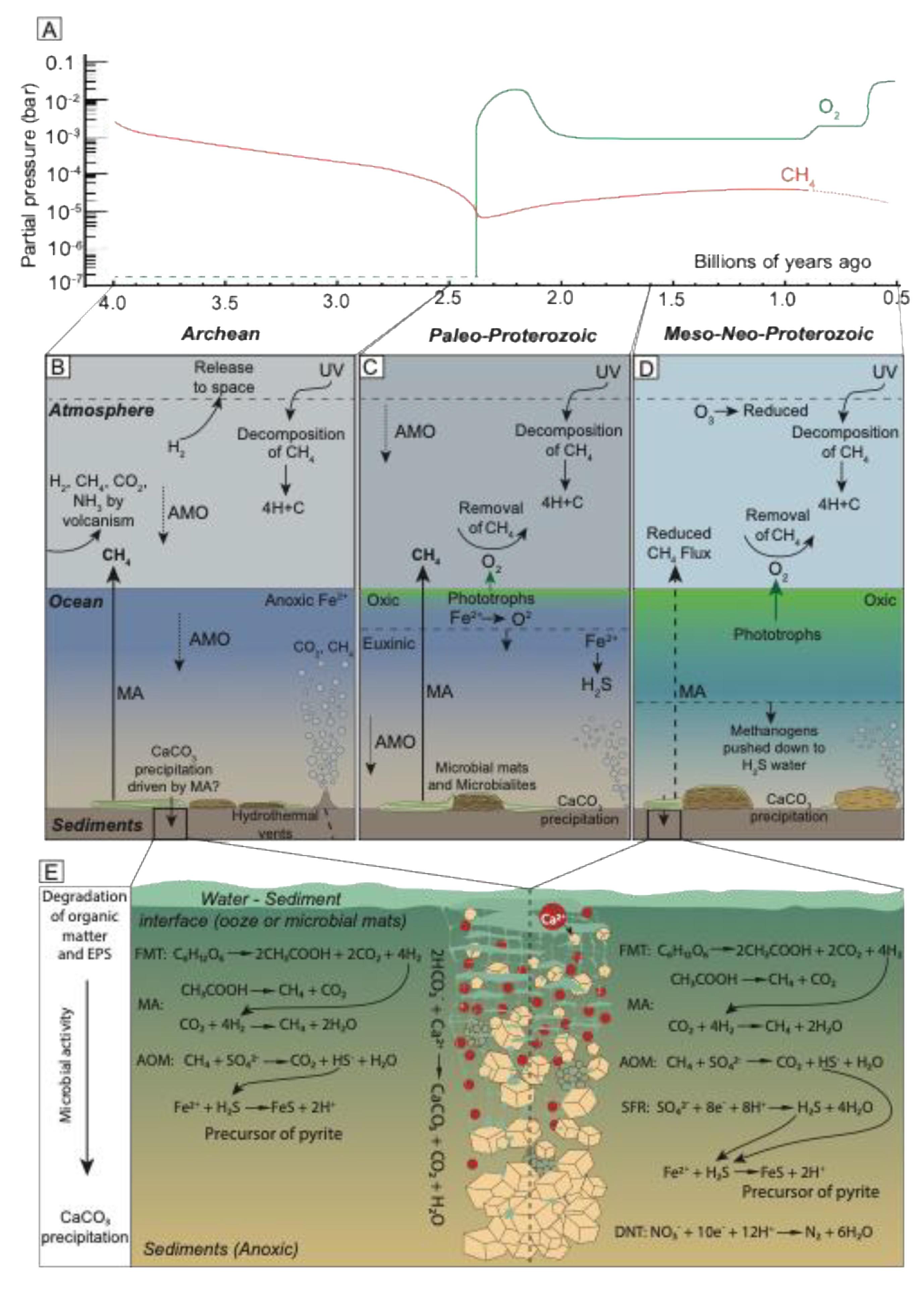
Large amounts of microbially-produced methane in the atmosphere (100-15,000 times present concentrations) (Figure 1A) [48] made up a major component of the carbon cycle and contributed to a greenhouse effect along with high atmospheric pCO2 (0.6 bar) [57]. This obviously would have supported early methanogenesis [173]. The high concentration of greenhouse gases kept global temperatures elevated [48,174], despite 20-25% lower solar radiation than today [48]. Secondary ion mass spectrometric analysis of kerogens recovered from 3.4 Gy Apex chert of Northwestern Western Australia revealed isotopic signatures consistent with methanogenesis, suggesting that this metabolism was likely an important process in the Archean biosphere [167]. This is consistent with the earliest evidence of methanogenesis from hydrothermal deposits dating back to 3.5 Gy [142].
2.1.1. Sulfur Cycling
The Archaean oceans contained high levels of iron sulfides (FeS and FeS2), hydrogen sulfide (H2S), zero-valent sulfur (So), and possibly thiosulfate (S2O32-) [48,175]. All these reduced sulfur compounds can fulfill the nutritional requirements for methanogens [48,176,177]. Sulfur fractionation occurs during enzymatic So-reduction with H2 [178]. Low sulfate and thus the absence of sulfate-reducing microbes in the early Archean [179,180] as well as a lesser degree of δ34S fractionation observed in the fossil record from that Eon [178], has been used to corroborate an early (i.e., Paleoarchean) presence of assimilatory sulfur reduction by methanogens [48]. Furthermore, proteins of lithotrophic methanogenic species that contain Fe-S clusters are thought to be among the earliest inorganic catalysts, as they can self-aggregate in high concentrations, like those found in the iron and sulfide-rich oceans [48].
In addition to inorganic sulfur, organic sulfur compounds made up an important part of the cycling of this element during the Archean as well [181]. Both biotic and abiotic reactions produce low-molecular-weight organic sulfur compounds [43]. Of these organosulfur compounds, methanethiol (CH3SH) and dimethylsulfide ((CH3)2S) are important sources of electrons during methane generation in microbial mats [42,43]. Vast amounts of methylated sulfides were produced by volcanoes and hydrothermal vents during the Hadean and Archean [182], and could thus have supported the evolution of methylotrophic methanogens early during Earth’s history. A reconstruction of microbial metabolisms in microbialites of the Dresser Formation, Pilbara, Australia, suggests that sulfate- and arsenate-reducing microbes may have competed with methanogens for metabolic substrates [183], a notion is based on the presence of sulfate and arsenic in these 3.5 Gy-old microbial sediments. Demethylation of arsenic is not only a detoxification mechanism, but in some cases, it can also support methane production, thus generating energy in methylotrophic methanogens [184].
2.1.2. Environmental Conditions and Photochemistry
Methanogens in the Archean would have experienced several ‘extreme’ conditions, like increased temperatures [54], higher amounts of UV surface radiation [79], and higher salinity in the oceans [54]. This may have been ‘normal’ for methanogens that had evolved at the time. Some localized environments may have hosted methanogens and could have been potential locations for the origin of life, such as hyperthermophilic conditions at hydrothermal vents [55] or exposure to ionizing radiation in nuclear geysers [51]. The average global ocean temperatures were in the range of 55 to 85oC and could have supported thermophiles as potentially the most ancient extremophilic mode of life [54]. This is further supported by some studies that suggest that LUCA could have been a thermophilic microbe [166,185]. Alternatively, studies focusing on hydrogen and inorganic phosphate (δ18OP) isotopes report a much lower ocean temperature of 26 to 40oC [186,187]. Under this scenario, or if it evolved in a non-marine system, LUCA may have been a mesophile. A mesophilic origin of LUCA is also supported by the thermoreduction hypothesis [188]. Increased ocean temperatures could have a greater potential for methane production from hydrogenotrophic and mixotrophic methanogens [189]; however, an ocean temperature of 40oC could have potentially been (near) optimal for methane production from both aceticlastic and hydrogenotrophic species [2,189].
Exposure to UV radiation at the surface of the planet was greater during the Archean due to the lack of ozone in the upper atmosphere [79]. Physical barriers to this radiation, perhaps protected microbes in the ocean, such as the high abundance of iron [39]. A variety of photopigments in microbial mats and microbialites could have quenched this damaging high-energy radiation [4]. Salinity reconstructions of the Archean oceans suggest that salinity was 1.5 to 2 times higher than it is at present [54], which would have supported halophilic species.
The impact methanogens had on the Archean carbon cycle not only affected the increased global temperature at the time but had a lasting effect on the evolution of Earths secondary and tertiary atmosphere as methane likely supported the irreversible oxidation of the atmosphere, oceans, and terrestrial systems in the Proterozoic [48]. Methane was removed in the Archean atmosphere by photochemical breakdown of CH4 into 4H and C by UV radiation, providing a source of OH radicals in the upper atmosphere [48,190] (Equation (6)).
CH4 + uv → C + 4H
While it is still debated, anaerobic methane oxidation (possibly by methanogens and other heterotrophic acetogens [15,191]) coupled to sulfate-, arsenate-, nitrite- or iron reduction [183,192] could have played a role in the removal of methane in the late Archean [146] (Figure 1B). Methanogens may also have been critical in the formation of an organic haze. When the atmospheric ratio of CH4:CO2 exceeds 0.1, a photochemical reaction of these gases using energy from UV radiation produces a variety of n-alkanes [193]. Atmospheric photolysis of CH4 and N2 formed HCN and NH4(OH) [194], adding an essential nutrient for life to the global oceans. This Archean organic atmospheric haze was not as thick as, e.g., the current haze on Saturn’s moon Titan (which has a CH4:CO2 ratio > 1) and may have had a climate-stabilizing effect on our planet [195].
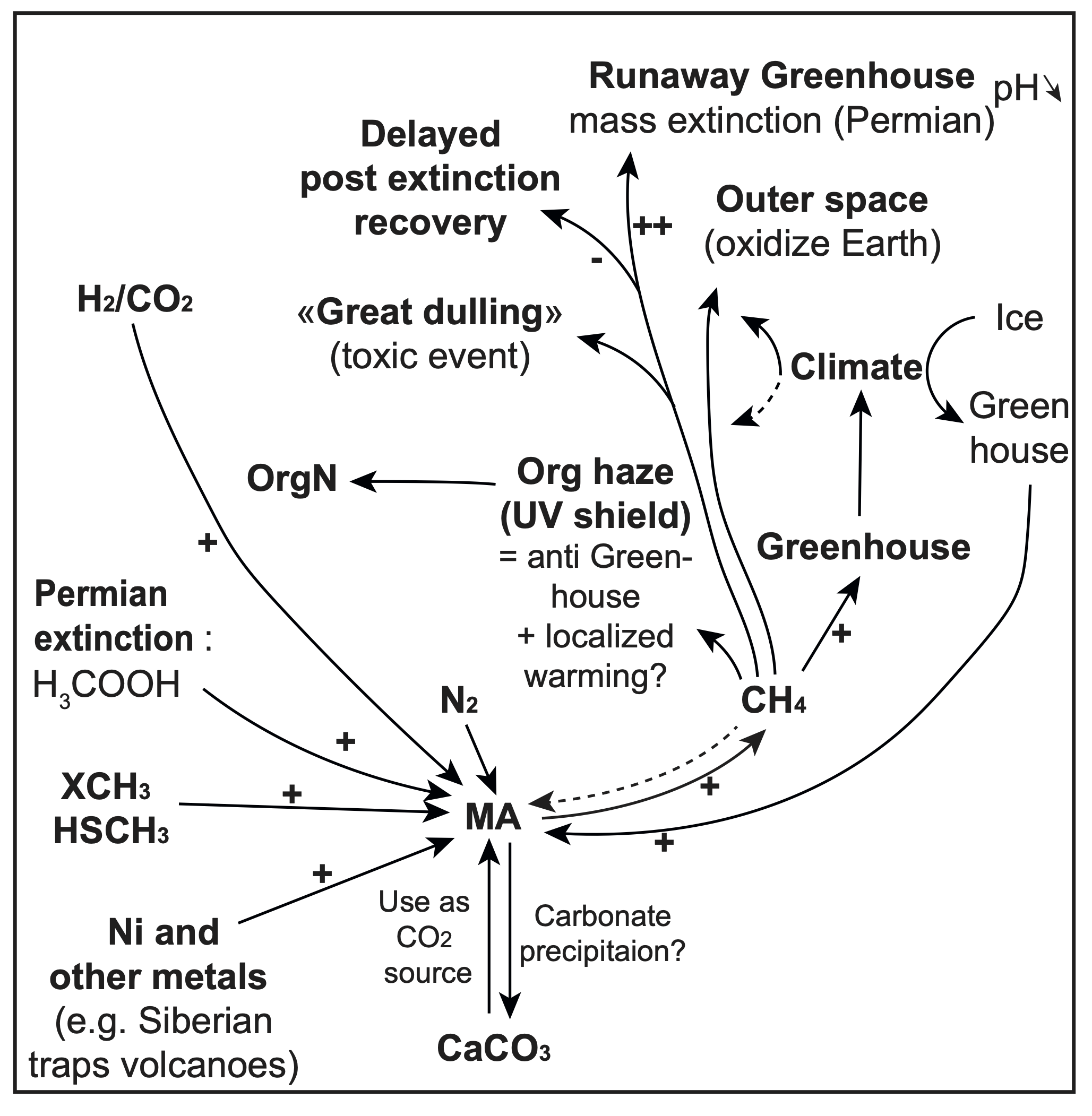
Figure 1.
Methane in the carbon cycle during the Archean and Proterozoic Eons. (A) Atmospheric CH4 and O2 concentrations from ~4.0 Gy to 0.5 Gy, adapted from [57] (B) The role of CH4 in the iron-rich oceans of the Archean, when there was a large flux of CH4 from methanogenesis (MA) in the oceans; sinks of methane included photochemical breakdown in the upper atmosphere and possible anaerobic oxidation of methane (AOM). The CH4 production exceeded removal, which resulted in a strong greenhouse effect and warming characteristic of the Archean atmosphere. Arrows show the cycling of CH4, including the flux of CH4 to the atmosphere by MA and the removal and decomposition of CH4. (C) CH4 and the carbon cycle in the Paleo-Proterozoic. Oxygen produced by cyanobacterial photosynthesis emerges as a novel oxidant for microbial removal (sink) of CH4 from the atmosphere. (D) Atmospheric CH4 is further reduced during the Meso-Neo-Proterozoic due to increased removal by oxic and anoxic methane oxidation, i.e., reverse methanogenesis coupled to sulfate and/or arsenate reduction, as well as through photochemical reaction of CH4 with O3 (through OH.). (E) CH4 production by syntrophic microbial processes through carbon degradation in sediments during the Archean and Meso-Neo-Proterozoic Eons. H2, and organic products (such as CH3COOH), from fermentation (FMT) could have supported MA in sediments. Anaerobic oxidation of methane (AOM) by sulfate reduction (SRF; shown here, but possibly also respiration using other electron donors such as nitrate, selenate, or arsenate (see Section 1.3.8) could have contributed to the removal of CH4 in sediments and promoted mineral formation (such as (arseno)pyrite). Active MA combined with other microbial processes [196] can drive carbonate mineral precipitation (such as CaCO3) in sediments and microbial mats.
Figure 1.
Methane in the carbon cycle during the Archean and Proterozoic Eons. (A) Atmospheric CH4 and O2 concentrations from ~4.0 Gy to 0.5 Gy, adapted from [57] (B) The role of CH4 in the iron-rich oceans of the Archean, when there was a large flux of CH4 from methanogenesis (MA) in the oceans; sinks of methane included photochemical breakdown in the upper atmosphere and possible anaerobic oxidation of methane (AOM). The CH4 production exceeded removal, which resulted in a strong greenhouse effect and warming characteristic of the Archean atmosphere. Arrows show the cycling of CH4, including the flux of CH4 to the atmosphere by MA and the removal and decomposition of CH4. (C) CH4 and the carbon cycle in the Paleo-Proterozoic. Oxygen produced by cyanobacterial photosynthesis emerges as a novel oxidant for microbial removal (sink) of CH4 from the atmosphere. (D) Atmospheric CH4 is further reduced during the Meso-Neo-Proterozoic due to increased removal by oxic and anoxic methane oxidation, i.e., reverse methanogenesis coupled to sulfate and/or arsenate reduction, as well as through photochemical reaction of CH4 with O3 (through OH.). (E) CH4 production by syntrophic microbial processes through carbon degradation in sediments during the Archean and Meso-Neo-Proterozoic Eons. H2, and organic products (such as CH3COOH), from fermentation (FMT) could have supported MA in sediments. Anaerobic oxidation of methane (AOM) by sulfate reduction (SRF; shown here, but possibly also respiration using other electron donors such as nitrate, selenate, or arsenate (see Section 1.3.8) could have contributed to the removal of CH4 in sediments and promoted mineral formation (such as (arseno)pyrite). Active MA combined with other microbial processes [196] can drive carbonate mineral precipitation (such as CaCO3) in sediments and microbial mats.
2.1.3. Methanogens in Early Hydrothermal Vents
Hydrothermal vents were important sites for the evolution and diversification of early life [55,183,197,198,199]. The oldest fossilized hydrothermal vents date back 440 My [200], but these sites must have existed much longer, possibly 4.3 Gya [201]. Vents (fluid) of the Archean were likely highly alkaline (pH 8.5 to >12) [202] and high temperature (~290 to 350oC) [183,198], vastly different than the black smokers (350oC to >450oC; pH 2-3) and white carbonate chimneys (50-90oC; pH 9-11) found in today’s oceans [55]. Similarly, the physicochemical gradient of vent fluid, in their interaction with the surrounding neutral to acidic seawater, was the reverse of what is observed today, where vent fluids interact with a more alkaline ocean [198]. Still, modern-day vent systems may provide relics of ancient biogeochemical interactions (for further reading, see [55]).
Archean vent fluids were rich in iron and sulfide [183,203], CO2 and CaCO3 [198], and SiO2 [198]. While the Fe content of ancient vent fluid and, likewise, a hydrothermal contribution to Archean banded iron formation is debated [198,204], serpentinization in vent fluids supplied abundant H2, C1-4 intermediates, and metals which could have supported hydrogenotrophic methanogens, acetogens, and sulfate reducers as early metabolisms [166,199,205,206]. For example, transition metals Co, Ni, and Fe from vents could have been a source of early cofactors in the acetyl-CoA (Wood-Ljungdahl) pathway found in modern-day methanogens, acetogens [205,207], and possibly in LUCA [166,199]. Additionally, some methanogens can fix N2 using dinitrogenase reductase [208]. Such nitrogenases are ancient and were likely important for early biochemistry at hydrothermal vents [199]. Mo may have also been abundant in Peridotite-hosted alkaline systems [209]. Hydrothermal activity in the Archean could have also been a source of oxidized compounds, such as SO42-, which could have expanded element cycles and the potential for microbial metabolisms (i.e, sulfate reduction) [183,202].
2.1.4. Microbial Mats of the Archean
Microbial mats are semi-closed sedimentary biofilms comprising bacteria and archaea, in a matrix made up of extracellular polymeric substances [4]. Microbial mats in the Archean were likely dominated by H2S –including methanogens and perhaps methane oxidizers, as well as other early heterotrophs [4,63,183,210,211,212]. However, it is thought that the earliest microbial mats may have been chemolithotrophic rather than autotrophic [4,63,183]. Microbial mats could have been among the earliest stable habitats [212], and the combination of a complex geochemical composition, a high cell density, and a close proximity of organisms would have made them ideal sites for the evolution of early life [4,63,212] and the diversification of metabolisms [183]. At the onset of the Neoarchean, cyanobacteria in microbial mats combined two photosystems [213] and the oxygenation of Earth’s oceans, atmosphere, and protocontinents took off [214]. Although the cyanobacteria date back ~ 2.8 Gy, their photosynthesis can also be supported by reduced sulfur species, and thus their first appearance can not be used as evidence for oxygen production. In fact, anoxygenic photosynthesis (e.g., using reduced sulfur as an electron donor) likely prevailed for tens of millions of years in these phototrophs [215]. Microbially-induced sedimentary systems [216] and stromatolites [217] record the preservation of microbial mats in the fossil record. The earliest evidence for this is found in the Pilbara dating back ~ 3.5 Gy [210,218,219]. Even though the true nature of the 3.7 Gy-old stromatolitic structures remains to date under discussion [220,221,222], these fossilized lithified microbial mats, which include stromatolites, thrombolites, dendrolites, and leiolites, are the main systems that provide insight into the biogeochemistry of the Archean world [4,217,223].
2.2. The Proterozoic
2.2.1. The Great Oxygenation Event and the Paleoproterozoic (2.5 to 1.6 Gy)
The evolution of oxygenic photosynthesis ~2.5 Gya spurred the permanent oxidation of the planet (Figure 1A) and led to greater oxidation states of the sulfur cycle [48,175] and expansion to the carbon cycle through the evolution of new metabolic pathways [48]. The abundance of methane left over from the Archean likely contributed to the irreversible, but slow, oxidation of the atmosphere, terrestrial environments, and Earth’s oceans [179]. Anoxia in the atmosphere was prolonged due to icehouse conditions [224]. Reduced atmospheric CO2 could have contributed to quicker removal (through oxidation) of methane by O2 [224] (Figure 1C). This relatively rapid removal of CH4 likely contributed greatly to/was an important factor in the reduction in global temperatures [48,174]. The oxidation of sulfide increased the sulfate concentration in the oceans and set the stage for the evolution of sulfur metabolisms 2.3-2.4 Gya [175].
2.2.2. The Boring Billion (1.8 to 0.8 Gy)
The onset of oxidation of reducing pre-GOE conditions resulted in euxinic, stratified oceans with oxygenated surface waters and sulfidic, anoxic bottom waters [48]. Anoxic oceans contained an abundance of hydrogen sulfide, which some methanogens used as a sulfur source [48]. Reactions between the oceanic ferrous iron (Fe(II)) and atmospheric O2 lead to the precipitation of iron sulfide minerals, like pyrite. This limited the availability of Fe(II) and other trace metals, such as nickel [225], which are essential for methanogen growth [58]. Some species of methanogens can reductively dissolve nickel and iron sulfide minerals, making them more bioavailable in euxinic conditions [112]. This capacity could have been used to survive in the anoxic zone of the oceans during the Boring Billion, which would have supported a continuing methane flux to the atmosphere [174]. However, methane concentrations were presumably reduced to less than 10 ppm in the mid Proterozoic [174] from Archean levels in the magnitude of 102 to 103 ppm [48,190], either through removal of large amounts of CH4 from the atmospheric reservoir and/or a reduction in CH4 flux to the atmosphere [174].
Sulfate reduction-supported anaerobic oxidation of methane could have limited the impact of methanogenesis on the global climate [174], in which CH4 is removed via anaerobic methane oxidation (Figure 1E) (Equation (7)):
CH4 + 2H2O + OH- → HCO3- + 4H2
Sulfate-reducing bacteria (SRB) consume H2 (Equation (8)) produced during methanogenesis, lowering the availability of this for methanogens [45]:
4H2 + SO42- → HS- + OH- + 3H2O
A similar role in anaerobic methane oxidation has been attributed to arsenate, iron, manganese, and nitrate reducers (see Section 1.3.8). Anaerobic methane oxidation thus created a sink for methane in the Proterozoic oceans (Figure 1D), removing most of this gas before it could escape into the atmosphere [174]. Simultaneously, methane removal by UV (Equation (6)) slowed down when oxygenation (and consequently O3 production) of the atmosphere commenced. These two events combined may help explain the steady but severely reduced temperature during the Boring Billion [226].
2.2.3. Methanogens Through Glaciations
2.2.3.1. The Ending of Marinoan “Snowball” Earth (650 – 635 Mya)
The continual decrease in the greenhouse gas concentration, including CO2 and CH4, through the mid-Proterozoic and Neoproterozoic eras triggered extensive low latitude glaciation, and perhaps the most extensive ice age of the planet [69,70]. Deglaciation and recovery of primary production was part of a feedback loop, which resulted in the reestablishment of strong euxinic oceanic conditions [69,70]. This was the result of extensive continental weathering following the accumulation of atmospheric CO2 during the glacial period [69]. A large flux of CH4 into the atmosphere played an important role in deglaciation [69] from active methanogenesis that may have utilized methyl sulfide compounds from seawater [69,70], volcanic activity [227], and the release of methane from methane clathrates [69,228]. Though still debated, methanogenesis likely played a key role in the formation of these methane clathrates [228]. Methanogens that produce CH4 under low temperature and/or high pressure are known to contribute to clathrate formation [229]. The production and consecutive release of methane from clathrates to the atmosphere has also been invoked later in the more recent events like the melting of the Laurentide–Cordilleran ice sheet [230].
2.2.4. Methanogens Through Mass Extinctions
Widespread and regional anoxic and euxinic ocean conditions have been characteristic of multiple extinction events , such as those found in Western Tethys and Panthalassa at the end Triassic mass extinction (~201.6 Mya) [206,231] and during the Permian mass extinction (252Mya) [232]. A rise in global temperature following rapid release of greenhouse gases (CO2 and CH4) [71,206,231], via volcanism (e.g., the Siberian Traps) [233], destabilization of methane reservoirs (e.g., methane clathrates/hydrates and permafrost) [206,232], and/or microbial sources (i.e., methanogenesis) [206]. These anoxic reservoirs could have fueled methanogen activity, creating a positive feedback loop that further contributed to a global rise in temperature, which in turn led to melting and release of additional methane. The End Triassic mass extinction experienced increases in acetate and H2 availability and a decrease in ocean sulfate concentrations that may have contributed to increased methane flux from methanogen activity in the oceans [206]. Likewise, following the Permian mass extinction, a euxinic photic zone and low ocean oxygen concentrations led to a buildup of organic material in sediments [234]. The degradation of this organic matter was likely both a substrate for methanogenesis and a source of acetate, which could have supported the evolution of the modern-day high efficiency aceticlastic pathway [234] in Methanoscarcina spp. [235]. Increased CH4 and H2S fluxes from the oceans during the Permian reduced atmospheric O2 and O3 by ~14% and consequently decreased CH4 oxidation in the atmosphere [206]. Under lower atmospheric O2 conditions like this, aerobic and anaerobic methane oxidation may play a critical role in the removal of CH4, in environments with and without oxygen, respectively [236]. Similar conditions may have been present during other mass extinction events [237], even though methanogenic activity during these times is not discussed extensively in the literature.
2.2.4.1. Resurgence of Methanogens and Microbialites Following Mass Extinctions
Methanogens often thrive following mass extinction events, significantly delaying ecosystem recovery, notably in marine environments. By disrupting the global carbon cycle, methanogens can create persistent toxic conditions, resulting in “dead zones”. Consequently, re-establishment of complex life is difficult due to warming and anoxic effects due to CH4, slowing the return of diverse ecosystems [238].
There is often an increase in the abundance of microbialites following mass extinction events [239] due to a carbonate-saturated, alkaline ocean, increased availability of nutrients (e.g., nitrogen-fixing cyanobacteria, weathering) [240,241], and a lack of competition with decimated eukaryote populations (i.e., decreased bioturbation and grazing pressure) [239,242,243]. Following the Permian-Triassic extinction event, the abundance of organic carbon combined with dysoxic conditions likely supported sulfate-reduction [244] and methanogenesis [234], and possibly anoxygenic photosynthesis by Gakhumella-like cyanobacteria [245]. However, early Triassic oceans may have had a low oceanic sulfate concentration [246], which would have profited methanogens [234]. Importantly, all three anaerobic metabolisms favor lithification of mats, potentially forming stromatolites [4,196]. A similar resurgence of microbialites under anoxic or dysoxic conditions, possibly supported in part by methanogens, occurred, e.g., during the Early Silurian, following the Late Ordovician Extinction Event [247] and the end-Devonian mass extinction [248]
2.3. Past, Present, and Future – Methanogens in Climate Change
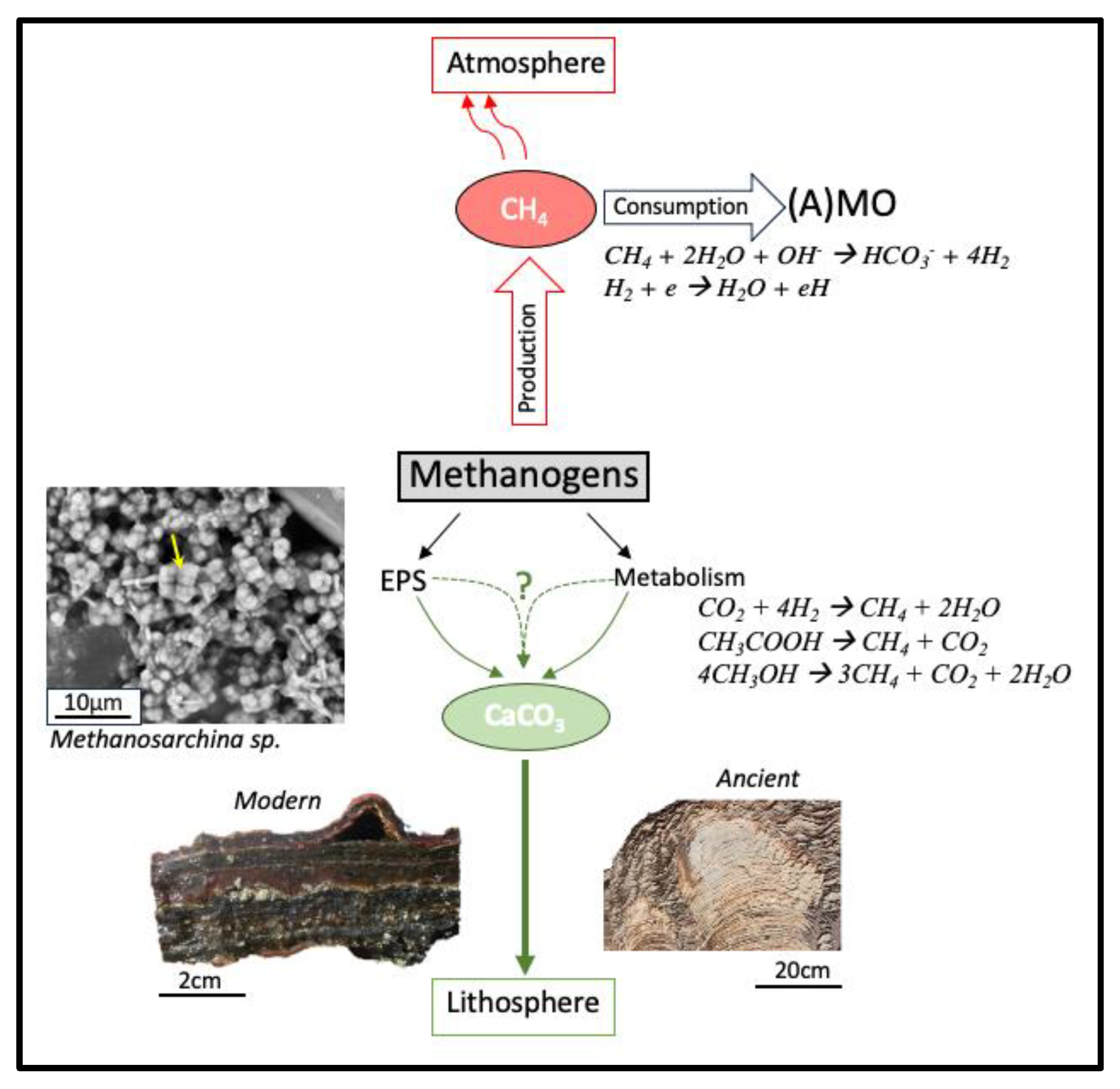
Methanogens still play an essential role in the modern-day climate, producing over 70% of biogenic methane emissions annually [249,250]. Although methane has a lower abundance in the atmosphere (~1.4 to 1.9 ppm; [48,251] than CO2, at ~360 to 390 ppm [252,253], it is about 30 times more potent as a greenhouse gas [3]. Methanogens process in excess of 500 Tg C y-1 through their metabolism in the production of CH4 and biomass [254]. However, methanogens may play an additional role in the global carbon cycle by mediating precipitation and dissolution of carbonate minerals [255], thus impacting long-term storage of atmospheric CO2 (Figure 2) [162]. This makes their specific role in the carbon cycle more complex than previously thought.
Lithotrophic metabolisms are believed to promote the precipitation of carbonate minerals [255] (Equation (3)) where removal of CO2 favors the production of CO32- ions, increasing pH (Equation (9)) in the surrounding environment. This could promote the precipitation of carbonates, such as CaCO3 [4,196] (Equation (10), net Equation (11)).
HCO3- → CO2 + OH-
HCO3- + Ca2+ → CaCO3 + H+ and H+ + OH- → H2O
2HCO3- + 4H2 + Ca2+ → CaCO3 + CH4 + 3H2O
However, other types of methanogenesis have different effects on the CaCO3 budget. Methanogens produce exoplymeric substance (EPS) [256], which could also aid in the process of mineralization [196,217]. On the other hand, aceticlastic (Equation (1)) and methylotrophic (Equation (2)) species may promote the dissolution of CaCO3 minerals through a decrease in pH, where CO2 is produced as a product of the metabolisms (Equations (1) and (2)). The increase in pH would favor HCO3- ions (Equation (12)) and promote the dissolution of CaCO3 minerals (Equation (13); net (for aceticlastic methanogenesis) Equation (14)). In which case, carbonate minerals could serve as a potential carbon source for methanogen growth [162].
CO2 + OH- →HCO3-
H2O → H+ + OH- and CaCO3 + H+ → HCO3- + Ca2+
CH3COOH + CaCO3 + H2 → 2HCO3- + CH4 + Ca2+
This dual role that methanogens play in the global carbon cycle and their metabolic and biogeochemical role in CaCO3 production remains unstudied, leaving gaps in our understanding of the capacity of microbes to regulate planetary climates. Currently, studies of methanogen-driven carbonate mineralization are limited to wastewater [256,257,258], carbonates in authigenic marine sediments [259,260], and Mn carbonates formed as a product of the methanogenic pathway [261]. Methanogens are abundant in many sediments, including microbial mats [24,43], which makes mat ecosystems prime locations to study methanogenic CaCO3 precipitation [42,262]. Methanogens occupy a unique niche within mats, relying mostly on non-competitive methylated compounds for growth, such as methylamines and methylsulfides are not used by SRB [43,263,264].
Figure 2.
The multiple roles of methanogens in the modern-day carbon cycle. Processes shown in red contribute to greenhouse gas production. Part of the methane is removed through microbial methane oxidation (A)MO and photochemical reactions, where e is a generic electron donor (e.g., arsenate or sulfate) that is reduced to eH. Processes depicted in green sequester CO2, storing it long-term as carbonates. The size of the arrows relates to the relative biogeochemical importance of the process. The metabolic activity of methanogens may locally alter pH and produce copious amounts of EPS (seen as a veil surrounding the cells in the microphotograph (yellow arrow)). These EPS bind cations (e.g., Ca, Mg) and act as nucleation site, while elevated pH enhances the carbonate precipitation in microbial mats [196]. This contribution by methanogens may ultimately produce microbialites [183].
Figure 2.
The multiple roles of methanogens in the modern-day carbon cycle. Processes shown in red contribute to greenhouse gas production. Part of the methane is removed through microbial methane oxidation (A)MO and photochemical reactions, where e is a generic electron donor (e.g., arsenate or sulfate) that is reduced to eH. Processes depicted in green sequester CO2, storing it long-term as carbonates. The size of the arrows relates to the relative biogeochemical importance of the process. The metabolic activity of methanogens may locally alter pH and produce copious amounts of EPS (seen as a veil surrounding the cells in the microphotograph (yellow arrow)). These EPS bind cations (e.g., Ca, Mg) and act as nucleation site, while elevated pH enhances the carbonate precipitation in microbial mats [196]. This contribution by methanogens may ultimately produce microbialites [183].
3. Methanogens in Microbial Mats as Biosignatures for Life
Organisms in microbial mats must cope with a range of not only extreme, but also fluctuating conditions [4]. Sulfide and oxygen gradients in these systems fluctuate over diel cycles [4,196,217], with O2 peaking during the early afternoon, along with peak photosynthesis by cyanobacteria, reaching daily peaks of > 600% O2 saturation in lithifying mats [4,60]. In contrast, peak sulfide concentrations are found near the mat surface, peaking at the end of the night, associated with sulfate reduction activity [4,196].
While the modern-day diversity of archaea in microbial mats is limited [262,265], methanogens have been identified (via their metabolic activity) in the anoxic but also the oxic zone of microbial mats [24,42,43,266,267]. Microbial mats are often found in extreme environments; in hypersaline conditions, when the salinity exceeds 180 PSU, methanogens can outcompete other anaerobic heterotrophs, notably SRB [60]. Desiccation impacts microbial mat communities through wetting and drying cycles [50] that enhance compatible solute production. As noted above, several of these compounds are precursors to methylamines and methylsulfides used by methanogens [42,267]. Desiccation has been proposed as a survival strategy in methanogens [50]. These authors reported that the viability of methanosarcinales was greatly enhanced by EPS, and that desiccated organisms had a higher resistance to high temperature and oxidative conditions. These properties may prove useful in extraterrestrial environments, such as Mars (See Section 4) [37].
3.1. The Evolution of Microbial Mats and Stromatolites
Microbial mats are organosedimentary systems made up of complex microbial communities embedded in an exopolymeric matrix [217]. Some mat communities facilitate microbially induced CaCO3 precipitation and produce lithifying mats [4,196,255]. The precipitation potential is determined by the combined metabolic activities of the entire microbial community and by the environmental (physicochemical) conditions [4,268]. Complete element cycling, notably of C, O, and S, is very efficient in these semi-closed, self-sustaining [255]. Microbial mats could have been among the first complex microbial communities to evolve on Earth [212], in shallow ponds near hot springs [183,269], appearing in the fossil record as stromatolites as early as ~3.5-3.7 Gya [220,222]. A variety of available chemical elements supporting a range of metabolisms, combined with a close proximity of an assortment of microorganisms [270] characteristic of mats, could have been instrumental in the diversification of microbial life.
Both lithified and non-lithified (soft) mats biofilm systems were more abundant during the Archean and Proterozoic eons, but in modern times have been forced to environments with extreme conditions likely due to competition with eukaryotes [4,271]. The presence of methanogenic activity in the oldest microbial mats and stromatolites has been suggested, mainly based on carbon isotopic evidence and metal availability[62,183,272,273]. Methanogens are still involved in the formation of modern microbialites [255], although actual their role in the lithification process remains largely unknown.
3.2. The Conundrum of Methanogenesis and Oxygen in Mats
It is yet unclear how methanogens cope with the fluctuating conditions in microbial mats. Notably, the presence of supersaturated oxygen concentrations during the early afternoon creates a challenge for these anaerobic organisms [24] due to the oxygen-sensitivity of methyl-coenzyme M reductase [274]. Although methanogens possess biochemical properties, such as ether-linked membrane lipids [275] or pseudomurein cell walls or proteinaceous sheaths [276], that make these microbes more resistant to extreme conditions (e.g., pH, temperature, mechanical stress, salinity), other survival mechanisms are employed. Methanogens do not form endospores but survive in a dormant state followed by rapid recovery [277]. Such a state of reduced activity is accomplished during desiccation. Additionally, there are specific solutions to oxygen-related stress: although glutathione has not been found in methanogens, enzymes like superoxide dismutase, catalase, and peroxidase cope directly with reactive oxygen species, and repair mechanisms of oxygen damage (e.g., disulfide reductases) also exist [102,278]. Other physiological adaptations include modified electron transport chains, production of protein-rich EPS, formation of cell clusters, polyphosphate accumulation, or a metabolic shift to methylated substrate use [50,102,276]. Quorum sensing has been reported in some methanogens[102,124], which hypothetically could allow for cyanobacteria, methanotrophs, and methanogens to live in close proximity while metabolically active (Figure 3). Such a model was proposed previously for the coexistence of sulfate-reducers, sulfide-oxidizers, and cyanobacteria [279]. Last but not least, other ecological adaptations may enable methanogens to thrive in oxic conditions. These include formation of consortia [280], syntrophism [281], and interspecies transfer of electrons [282].
4. Methanogens in the Search for Extraterrestrial Life
Methanogens are of prime interest for possible life on exoplanets. The primary product of their metabolism, methane, is an ideal gas for detection in remote atmospheres due to the unique absorption spectrum and short photochemical lifespan [6]. To date, methane has been detected on several planetary bodies in our solar system that may be fit for supporting methanogenesis-like life, including Mars [283], Saturn’s moon Enceladus [284], and Titan [285]. The detection of methane on other planets and its short residency time in planetary atmospheres suggests sources for an active methane flux on these planetary bodies, which on Earth, primarily is of biological origin [6]. The potential of methane, through the greenhouse effect, to warm planetary atmospheres may expand the possibility for life beyond the “Goldilocks” zone [10].
Many studies have assessed methanogen survival under Mars-like conditions [30,36,37,39,286] and their potential habitability on ice worlds like Enceladus [13]. The extreme environments many methanogens are found are used as analogs for extraterrestrial bodies [10,35,168]. For example, soda lakes are studied for their potential as analogs to Mars’s ancient ocean [35,287]. Similarly, hydrothermal vents are expected to exist on other worlds, like Enceladus, where products of serpentinization reactions and silica-rich particles have been detected [13]. Thermophilic methanogens may be able to survive at hydrothermal vent sites on Enceladus, and the predicted temperature and pressure associated with this ecological extreme [13]. Survival of methanogens to conditions on Mars (e.g., low pressure at the surface (6 mbar), desiccated soil, high UV radiation at the surface, and freezing surface conditions and permafrost due to lack of an atmosphere) [30,36,37,39,78,286] has supported the idea of potential subsurface life on the planet [36], or more broadly, a subsurface source for methane [288]. Microbial mats themselves are useful biosignatures due to their self-sustainability, resistance to extremes, and abundance on early Earth [168].
5. Conclusions
Methanogens were among the earliest microorganisms to evolve on Earth and continue to thrive in a wide range of both seemingly ‘non-extreme’ and ‘extreme’ environments today, several of which are similar to those throughout Earth’s history (Figure 4). Microbial mats are a prime example of ecosystems that evolved with our planet (or, arguably, changed our planet’s properties) and were present in abundance during the Archean and Proterozoic eons. Methanogens were likely part of the early stromatolite communities and continued to be ubiquitous in microbial mat ecosystems through time [212]. The transition from strictly oxic conditions during the day and anoxic ones during the night may have supported methanogens to be well-adapted to a wide range of sometimes fluctuating conditions (e.g., diel fluctuations of O2, sulfide, pH, and light gradients) [4]. The lack of a proper understanding of the ecophysiological capabilities and flexibilities (e.g., EPS properties, stress responses, viral-host relationships, cell-to-cell communication) is a major gap in the understanding of the role and evolution of methanogens and their impact on our planet through geologic time. Their elementary metabolism (e.g., respiration of H2/CO2), efficient use of a minimal energy yield, and the simplicity of their main metabolic product (i.e., CH4) – which is observable remotely and supports a diversity of photochemical reactions – make methanogens the ultimate candidates for astrobiological studies.
Funding
This work for this paper received no external funding.
Acknowledgements
PV acknowledges support from NASA Exobiology NNH22ZDA001N-WXO grant. PT received support from the National Science Foundation Graduate Research Fellowship Grant No. ####. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation. PB acknowledges support of AAPG-1-HARMI-ANR. This work was also supported by Start Up funds provided to MGS-M (#MCB4171510) by the Department of Molecular and Cell Biology (MCB) and the College of Liberal Arts and Sciences (CLAS) at the University of Connecticut (UConn)
Conflicts of Interest
The authors declare no conflicts of interest
References
- Megonigal, J.P.; Mines, M.E.; Visscher, P.T. Anaerobic Metabolism: Linkages to Trace Gases and Aerobic Processes. Biogeochemistry 2004, 8, 317–424.
- Conrad, R. Complexity of Temperature Dependence in Methanogenic Microbial Environments. Front Microbiol 2023, 14. [CrossRef]
- Buan, N.R. Methanogens: Pushing the Boundaries of Biology. Emerg Top Life Sci 2018, 2, 629–646. [CrossRef]
- Visscher, P.T.; Gallagher, K.L.; Bouton, A.; Vennin, E.; Thomazo, C.; Iii, R.A.W.; Burns, B.P. Treatise on Invertebrate Paleontology, Part B, Vol. 2, Chapter 3: Microbial Mats. Kansas Univ Press 2022, 163, 24–56.
- Kadoya, S.; Catling, D.C. Constraints on Hydrogen Levels in the Archean Atmosphere Based on Detrital Magnetite. Geochim Cosmochim Acta 2019, 262, 207–219. [CrossRef]
- Thompson, M.A.; Krissansen-Totton, J.; Wogan, N.; Telus, M.; Fortney, J.J. The Case and Context for Atmospheric Methane as an Exoplanet Biosignature. Proc Natl Acad Sci U S A 2022, 119. [CrossRef]
- Battistuzzi, F.U.; Feijao, A.; Hedges, S.B. A Genomic Timescale of Prokaryote Evolution: Insights into the Origin of Methanogenesis, Phototrophy, and the Colonization of Land. BMC Evol Biol 2004, 4, 1–14. [CrossRef]
- Rothschild, L.; Nature, R.M.-; 2001, undefined Life in Extreme Environments. nature.com 2001.
- Lyons, T.W.; Tino, C.J.; Fournier, G.P.; Anderson, R.E.; Leavitt, W.D.; Konhauser, K.O.; Stüeken, E.E. Co-evolution of Early Earth Environments and Microbial Life. Nature Reviews Microbiology 2024 22:9 2024, 22, 572–586. [CrossRef]
- Taubner, R.-S.; Schleper, C.; Firneis, M.G.; K-M Rittmann, S.R.; Klenk, H.-P.; W Adams, M.W.; Garrett, R.A. Assessing the Ecophysiology of Methanogens in the Context of Recent Astrobiological and Planetological Studies. Life 2015, 5, 1652–1686. [CrossRef]
- Jung, H.; Yu, H.; Lee, C. Direct Interspecies Electron Transfer Enables Anaerobic Oxidation of Sulfide to Elemental Sulfur Coupled with CO2-Reducing Methanogenesis. iScience 2023, 26, 107504. [CrossRef]
- Yin, Q.; Gu, M.; Hermanowicz, S.W.; Hu, H.; Wu, G. Potential Interactions between Syntrophic Bacteria and Methanogens via Type IV Pili and Quorum-Sensing Systems. Environ Int 2020, 138, 105650. [CrossRef]
- Taubner, R.; Pappenreiter, P.; Zwicker, J.; Bach, W.; Peckmann, J.; Paulik, C.; Firneis, M.; Schleper, C.; Rittman, S. Biological Methane Production under Putative Enceladus-like Conditions. Nat Commun 2018, 748. [CrossRef]
- Morgavi, D.P.; Forano, E.; Martin, C.; Newbold, C.J. Microbial Ecosystem and Methanogenesis in Ruminants. Animal 2010, 4, 1024–1036. [CrossRef]
- Ferry, J.G. Methanosarcina Acetivorans: A Model for Mechanistic Understanding of Aceticlastic and Reverse Methanogenesis. Front Microbiol 2020, 11, 545389. [CrossRef]
- Sorokin, Di.Y.; Makarova, K.S.; Abbas, B.; Ferrer, M.; Golyshin, P.N.; Galinski, E.A.; Ciordia, S.; Mena, M.C.; Merkel, A.Y.; Wolf, Y.I.; et al. Discovery of Extremely Halophilic, Methyl-Reducing Euryarchaea Provides Insights into the Evolutionary Origin of Methanogenesis. Nat Microbiol 2017, 2. [CrossRef]
- Protasov, E.; Nonoh, J.O.; Kästle Silva, J.M.; Mies, U.S.; Hervé, V.; Dietrich, C.; Lang, K.; Mikulski, L.; Platt, K.; Poehlein, A.; et al. Diversity and Taxonomic Revision of Methanogens and Other Archaea in the Intestinal Tract of Terrestrial Arthropods. Front Microbiol 2023, 14, 1281628. [CrossRef]
- Wen, X.; Yang, S.; Horn, F.; Winkel, M.; Wagner, D.; Liebner, S. Global Biogeographic Analysis of Methanogenic Archaea Identifies Community-Shaping Environmental Factors of Natural Environments. Front Microbiol 2017, 8, 280269. [CrossRef]
- Merkel, A.Y.; Podosokorskaya, O.A.; Sokolova, T.G.; Bonch-Osmolovskaya, E.A. Diversity of Methanogenic Archaea from the 2012 Terrestrial Hot Spring (Valley of Geysers, Kamchatka). Microbiology (Russian Federation) 2016, 85, 342–349. [CrossRef]
- Wang, J.; Qu, Y.N.; Evans, P.N.; Guo, Q.; Zhou, F.; Nie, M.; Jin, Q.; Zhang, Y.; Zhai, X.; Zhou, M.; et al. Evidence for Nontraditional Mcr-Containing Archaea Contributing to Biological Methanogenesis in Geothermal Springs. Sci Adv 2023, 9. doi:10.1126/SCIADV.ADG6004/SUPPL_FILE/SCIADV.ADG6004_SUPPLEMENTARY_DATA_S1_TO_S4.ZIP. [CrossRef]
- Schulz, S.; Conrad, R. Influence of Temperature on Pathways to Methane Production in the Permanently Cold Profundal Sediment of Lake Constance. FEMS Microbiol Ecol 1996, 20, 1–14. [CrossRef]
- Simankova, M. V.; Kotsyurbenko, O.R.; Lueders, T.; Nozhevnikova, A.N.; Wagner, B.; Conrad, R.; Friedrich, M.W. Isolation and Characterization of New Strains of Methanogens from Cold Terrestrial Habitats. Syst Appl Microbiol 2003, 26, 312–318. [CrossRef]
- Ye, J.; Zhuang, M.; Hong, M.; Zhang, D.; Ren, G.; Hu, A.; Yang, C.; He, Z.; Zhou, S. Methanogenesis in the Presence of Oxygenic Photosynthetic Bacteria May Contribute to Global Methane Cycle. Nat Commun 2024, 15, 1–11. [CrossRef]
- Buckley, D.H.; Baumgartner, L.K.; Visscher, P.T. Vertical Distribution of Methane Metabolism in Microbial Mats of the Great Sippewissett Salt Marsh. Environ Microbiol 2008, 10, 967–977. [CrossRef]
- Bogard, M.J.; Del Giorgio, P.A.; Boutet, L.; Carolina, M.; Chaves, G.; Prairie, Y.T.; Merante, A.; Derry, A.M. Oxic Water Column Methanogenesis as a Major Component of Aquatic CH 4 Fluxes. Nat Commun 2014, 5, 5350. [CrossRef]
- Patel, L.; Singh, R.; Thottathil, S.D. Contribution of Photosynthesis-Driven Oxic Methane Production to the Methane Cycling of a Tropical River Network. ACS ES&T Water 2024, 4, 2836–2847. [CrossRef]
- Cynar, F.J.; Yayanos, A.A. Enrichment and Characterization of a Methanogenic Bacterium from the Oxic Upper Layer of the Ocean. Curr Microbiol 1991, 23, 89–96. [CrossRef]
- Zhang, C.J.; Zhang, C.J.; Pan, J.; Liu, Y.; Duan, C.H.; Duan, C.H.; Li, M. Genomic and Transcriptomic Insights into Methanogenesis Potential of Novel Methanogens from Mangrove Sediments. Microbiome 2020, 8, 1–12. doi:10.1186/S40168-020-00876-Z/FIGURES/4. [CrossRef]
- Angle, J.C.; Morin, T.H.; Solden, L.M.; Narrow, A.B.; Sith, G.J.; Borton, M.A.; Rey-Sanchez, C.; Daly, R.A.; Mirfenderesgi, G.; Hoyt, D.W.; et al. Methanogenesis in Oxygenated Soils Is a Substantial Fraction of Wetland Methane Emissions. Nat Commun 2017, 8, 1567. [CrossRef]
- Kendrick, M.G.; Kral, T.A. Survival of Methanogens during Desiccation: Implications for Life on Mars. Astrobiology 2006, 6, 546–551. [CrossRef]
- Kral, T.A.; Travis Aitheide, S. Methanogen Survival Following Exposure to Desiccation, Low Pressure and Martian Regolith Analogs. Planet Space Sci 2013, 89, 167–171. [CrossRef]
- Moissl-Eichinger, C. Archaea in Artificial Environments: Their Presence in Global Spacecraft Clean Rooms and Impact on Planetary Protection. ISME J 2011, 5, 209–219. [CrossRef]
- Mondav, R.; Woodcroft, B.J.; Kim, E.H.; Mccalley, C.K.; Hodgkins, S.B.; Crill, P.M.; Chanton, J.; Hurst, G.B.; Verberkmoes, N.C.; Saleska, S.R.; et al. Discovery of a Novel Methanogen Prevalent in Thawing Permafrost. Nature Communications 2014 5:1 2014, 5, 1–7. [CrossRef]
- Takai, K.; Nakamura, K.; Toki, T.; Tsunogai, U.; Miyazaki, M.; Miyazaki, J.; Hirayama, H.; Nakagawa, S.; Nunoura, T.; Horikoshi, K. Cell Proliferation at 122 °C and Isotopically Heavy CH 4 Production by a Hyperthermophilic Methanogen under High-Pressure Cultivation. 2008. Epub2008Jul29. [CrossRef]
- Oremland, R.S.; Marsh, L.; DesMarais, D.J. Methanogenesis in Big Soda Lake, Nevada: An Alkaline, Moderately Hypersaline Desert Lake. Appl Environ Microbiol 1982, 43, 462. [CrossRef]
- Mickol, R.L.; Kral, T.A. Low Pressure Tolerance by Methanogens in an Aqueous Environment: Implications for Subsurface Life on Mars. Origins of Life and Evolution of Biospheres 2017, 47, 511–532. doi:10.1007/S11084- 016-9519-9/FIGURES/4. [CrossRef]
- Kral, T.A.; Altheide, T.S.; Lueders, A.E.; Schuerger, A.C. Low Pressure and Desiccation Effects on Methanogens: Implications for Life on Mars. Planet Space Sci 2011, 59, 264–270. [CrossRef]
- Bernhardt, G.; Jaenicke, R.; Ludemann, H.; Konig, H. High Pressure Enhances the Growth Rate of the Thermophilic Archaebacterium Methanococcus Thermolithotrophicus without Extending Its Temperature Range. Appl Environ Microbiol 1988, 54, 1258–1261. [CrossRef]
- Sinha, N.; Kral, T.A. Effect of UVC Radiation on Hydrated and Desiccated Cultures of Slightly Halophilic and Non-Halophilic Methanogenic Archaea: Implications for Life on Mars. Microorganisms 2018, 6, 43. [CrossRef]
- Harris, R.L.; Schuerger, A.C. Hydrogenotrophic Methanogenesis at 7–12 Mbar by Methanosarcina Barkeri under Simulated Martian Atmospheric Conditions. Sci Rep 2025, 15, 1–15. [CrossRef]
- Wong, H.L.; Visscher, P.T.; White, R.A.; Smith, D.L.; Patterson, M.M.; Burns, B.P. Dynamics of Archaea at Fine Spatial Scales in Shark Bay Mat Microbiomes. Sci Rep 2017, 7, 1–12. doi:10.1038/SREP46160;TECHMETA=23,45;SUBJMETA=158,26,326,631,704,855;KWRD=ARCHAEA,MICROBIAL+ECOLOGY. [CrossRef]
- Visscher, P.T.; Baumgartner, L.K.; Buckley, D.H.; Rogers, D.R.; Hogan, M.E.; Raleigh, C.D.; Turk, K.A.; Des Marais, D.J. Dimethyl Sulphide and Methanethiol Formation in Microbial Mats: Potential Pathways for Biogenic Signatures. Environ Microbiol 2003, 5, 296–308. [CrossRef]
- Visscher, P.T.; Quist, P.; Van Gemerden, H. Methylated Sulfur Compounds in Microbial Mats: In Situ Concentrations and Metabolism by a Colorless Sulfur Bacterium. Appl Environ Microbiol 1991, 57, 1758–1763. [CrossRef]
- Timmers, P.H.A.; Welte, C.U.; Koehorst, J.J.; Plugge, C.M.; Jetten, M.S.M.; Stams, A.J.M. Reverse Methanogenesis and Respiration in Methanotrophic Archaea. Archaea 2017, 2017, 1654237. [CrossRef]
- Boetius, A.; Ravenschlag, K.; Schubert, C.J.; Rickert2, D.; Widdel, F.; Gieseke, A.; Amann, R.; Jùrgensen, B.B.; Witte, U.; Pfannkuche2, O. A Marine Microbial Consortium Apparently Mediating Anaerobic Oxidation of Methane. Nature 2000, 407. [CrossRef]
- Merino, N.; Aronson, H.S.; Bojanova, D.P.; Feyhl-Buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the Extremes: Extremophiles and the Limits of Life in a Planetary Context. Front Microbiol 2019, 10. [CrossRef]
- Thomas, T.; Kumar, N.; Cavicchioli, R. Effects of Ribosomes and Intracellular Solutes on Activities and Stabilities of Elongation Factor 2 Proteins from Psychrotolerant and Thermophilic Methanogens. J Bacteriol 2001, 183, 1974–1982. [CrossRef]
- Liu, Y.; Beer, L.L.; Whitman, W.B. Methanogens: A Window into Ancient Sulfur Metabolism. Trends Microbiol 2012, 20, 251–258. [CrossRef]
- Nozhevnikova, A.N.; Zepp, K.; Vazquez, F.; Zehnder, A.J.B.; Holliger, C. Evidence for the Existence of Psychrophilic Methanogenic Communities in Anoxic Sediments of Deep Lakes. Appl Environ Microbiol 2003, 69, 1832–1835. doi:10.1128/AEM.69.3.1832-1835.2003/ASSET/B7324751-4850-4993-95E5-369B8A71FD77/ASSETS/GRAPHIC/AM0331479004.JPEG.
- Anderson, K.L.; Apolinario, E.E.; Sowers, K.R. Desiccation as a Long-Term Survival Mechanism for the Archaeon Methanosarcina Barkeri. Appl Environ Microbiol 2012, 78, 1473. [CrossRef]
- Ebisuzaki, T.; Maruyama, S. Nuclear Geyser Model of the Origin of Life: Driving Force to Promote the Synthesis of Building Blocks of Life. 2016. [CrossRef]
- Olson, K.D.; McMahon, C.W.; Wolfe, R.S. Light Sensitivity of Methanogenic Archaebacteria. Appl Environ Microbiol 1991, 57, 2683–2686. [CrossRef]
- Angel, R.; Matthies, D.; Conrad, R. Activation of Methanogenesis in Arid Biological Soil Crusts Despite the Presence of Oxygen. PLoS One 2011, 6, e20453. [CrossRef]
- Knauth, L.P. Temperature and Salinity History of the Precambrian Ocean: Implications for the Course of Microbial Evolution. Palaeogeogr Palaeoclimatol Palaeoecol 2005, 219, 53–69. [CrossRef]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal Vents and the Origin of Life. Nat Rev Microbiol 2008, 6, 805–814. [CrossRef]
- Horikochi, K. Barophiles: Deep-Sea Microorganisms Adapted to an Extreme Environment. Curr Opin Microbiol 1998, 1, 291–295. [CrossRef]
- Catling, D.C.; Zahnle, K.J. The Archean Atmosphere. Sci Adv 2020, 6. https://doi.org/10.1126/SCIADV.AAX1420/ASSET/6320EA09-4E49-402D-9855-C6D7C27118CD/ASSETS/GRAPHIC/AAX1420-F5.JPEG.
- Neubeck, A.; Sjöberg, S.; Price, A.; Callac, N.; Schnürer, A. Effect of Nickel Levels on Hydrogen Partial Pressure and Methane Production in Methanogens. PLoS One 2016, 11, e0168357. [CrossRef]
- Strobel, D.F. Photochemistry of the Reducing Atmospheres of Jupiter, Saturn and Titan. Int Rev Phys Chem 1983, 3, 145–176. [CrossRef]
- Visscher, P.T.; Dupraz, C.; Braissant, O.; Gallagher, K.L.; Glunk, C.; Casillas, L.; Reed, R.E.S. Biogeochemistry of Carbon Cycling in Hypersaline Mats: Linking the Present to the Past through Biosignatures. 2010, 443–468. [CrossRef]
- Lyu, Z.; Lu, Y. Metabolic Shift at the Class Level Sheds Light on Adaptation of Methanogens to Oxidative Environments. ISME Journal 2018, 12, 411–423. https://doi.org/10.1038/ISMEJ.2017.173;TECHMETA=45;SUBJMETA=158,2526,26,326,631,855;KWRD=ARCHAEAL+GENOMICS,MICROBIAL+ECOLOGY. [CrossRef]
- Lepot, K. Signatures of Early Microbial Life from the Archean (4 to 2.5 Ga) Eon. Earth Sci Rev 2020, 209, 103296. [CrossRef]
- Visscher, P.T.; Gallagher, K.L.; Bouton, A.; Farias, M.E.; Kurth, D.; Sancho-Tomás, M.; Philippot, P.; Somogyi, A.; Medjoubi, K.; Vennin, E.; et al. Modern Arsenotrophic Microbial Mats Provide an Analogue for Life in the Anoxic Archean. Communications Earth & Environment 2020 1:1 2020, 1, 1–10. [CrossRef]
- Lira-Silva, E.; Santiago-Martínez, M.G.; Hernández-Juárez, V.; García-Contreras, R.; Moreno-Sánchez, R.; Jasso-Chávez, R. Activation of Methanogenesis by Cadmium in the Marine Archaeon Methanosarcina Acetivorans. PLoS One 2012, 7, e48779. [CrossRef]
- Anbar, A.D.; Knoll, A.H. Proterozoic Ocean Chemistry and Evolution: A Bioinorganic Bridge? Science (1979) 2002, 297, 1137–1142. [CrossRef]
- Chen, C.; Li, L.; Huang, K.; Zhang, J.; Xie, W.Y.; Lu, Y.; Dong, X.; Zhao, F.J. Sulfate-Reducing Bacteria and Methanogens Are Involved in Arsenic Methylation and Demethylation in Paddy Soils. ISME Journal 2019, 13, 2523–2535. [CrossRef]
- Karr, E.A.; Sattley, W.M.; Rice, M.R.; Jung, D.O.; Madigan, M.T.; Achenbach, L.A. Diversity and Distribution of Sulfate-Reducing Bacteria in Permanently Frozen Lake Fryxell, McMurdo Dry Valleys, Antarctica. Appl Environ Microbiol 2005, 71, 6353–6359. [CrossRef]
- Zhang, G.; Jiang, N.; Liu, X.; Dong, X. Methanogenesis from Methanol at Low Temperatures by a Novel Psychrophilic Methanogen, “Methanolobus Psychrophilus” Sp. Nov., Prevalent in Zoige Wetland of the Tibetan Plateau. Appl Environ Microbiol 2008, 74, 6114–6120. https://doi.org/10.1128/AEM.01146-08/SUPPL_FILE/SUPPLEMENT_MATERIAL.ZIP.
- Zhao, Z.; Shen, B.; Zhu, J.M.; Lang, X.; Wu, G.; Tan, D.; Pei, H.; Huang, T.; Ning, M.; Ma, H. Active Methanogenesis during the Melting of Marinoan Snowball Earth. Nat Commun 2021, 12. [CrossRef]
- Peng, Y.; Bao, H.; Jiang, G.; Crockford, P.; Feng, D.; Xiao, S.; Kaufman, A.J.; Wang, J. A Transient Peak in Marine Sulfate after the 635-Ma Snowball Earth. Proc Natl Acad Sci U S A 2022, 119. [CrossRef]
- Song, H.; Kemp, D.B.; Tian, L.; Chu, D.; Song, H.; Dai, X. Thresholds of Temperature Change for Mass Extinctions. Nat Commun 2021, 12, 1–8. https://doi.org/10.1038/S41467-021-25019-2;SUBJMETA=158,181,2462,414,631,670,704;KWRD=BIODIVERSITY,PALAEOECOLOGY,PALAEONTOLOGY.
- Noon, K.R.; Guymon, R.; Crain, P.F.; McCloskey, J.A.; Thomm, M.; Lim, J.; Cavicchioli, R. Influence of Temperature on TRNA Modification in Archaea: Methanococcoides Burtonii (Optimum Growth Temperature [Topt], 23 °C) and Stetteria Hydrogenophila (Topt, 95 °C). J Bacteriol 2003, 185, 5483–5490. https://doi.org/10.1128/JB.185.18.5483-5490.2003/ASSET/84D10F3D-7305-46A9-9F70-323BE4B94A4B/ASSETS/GRAPHIC/JB1830355003.JPEG.
- Lim, J.; Thomas, T.; Cavicchioli, R. Low Temperature Regulated DEAD-Box RNA Helicase from the Antarctic Archaeon, Methanococcoides Burtonii. J Mol Biol 2000, 297, 553–567. [CrossRef]
- Shimada, Y.; Fukuda, W.; Akada, Y.; Ishida, M.; Nakayama, J.; Imanaka, T.; Fujiwara, S. Property of Cold Inducible DEAD-Box RNA Helicase in Hyperthermophilic Archaea. Biochem Biophys Res Commun 2009, 389, 622–627. [CrossRef]
- Saunders, N.F.W.; Thomas, T.; Curmi, P.M.G.; Mattick, J.S.; Kuczek, E.; Slade, R.; Davis, J.; Franzmann, P.D.; Boone, D.; Rusterholtz, K.; et al. Mechanisms of Thermal Adaptation Revealed from the Genomes of the Antarctic. Genome Res 2003, 13, 1580–1588. [CrossRef]
- Nichols, D.S.; Miller, M.R.; Davies, N.W.; Goodchild, A.; Raftery, M.; Cavicchioli, R. Cold Adaptation in the Antarctic Archaeon Methanococcoides Burtonii Involves Membrane Lipid Unsaturation. J Bacteriol 2004, 186, 8508–8515. [CrossRef]
- Koga, Y. Thermal Adaptation of the Archaeal and Bacterial Lipid Membranes. Archaea 2012, 2012, 789652. [CrossRef]
- Cheptsov, V.S.; Vorobyova, E.A.; Osipov, G.A.; Manucharova, N.A.; Polyanskaya, L.M.; Gorlenko, M. V.; Pavlov, A.K.; Rosanova, M.S.; Lomasov, V.N. Microbial Activity in Martian Analog Soils after Ionizing Radiation: Implications for the Preservation of Subsurface Life on Mars. AIMS Microbiol 2018, 4, 541–562. [CrossRef]
- Cockell, C.S. Biological Effects of High Ultraviolet Radiation on Early Earth—a Theoretical Evaluation. J Theor Biol 1998, 193, 717–729. [CrossRef]
- Zahnle, K.J.; Walker, J.C.G. The Evolution of Solar Ultraviolet Luminosity. Reviews of Geophysics 1982, 20, 280–292. [CrossRef]
- Ohlendorf, R.; Möglich, A. Light-Regulated Gene Expression in Bacteria: Fundamentals, Advances, and Perspectives. Front Bioeng Biotechnol 2022, 10. [CrossRef]
- Elahi, Y.; Baker, M.A.B. Light Control in Microbial Systems. International Journal of Molecular Sciences 2024, Vol. 25, Page 4001 2024, 25, 4001. [CrossRef]
- Katayama, T.; Kamagata, Y. Cultivation of Methanogens. 2015, 177–195. [CrossRef]
- Paula, F.S.; Chin, J.P.; Schnürer, A.; Müller, B.; Manesiotis, P.; Waters, N.; Macintosh, K.A.; Quinn, J.P.; Connolly, J.; Abram, F.; et al. The Potential for Polyphosphate Metabolism in Archaea and Anaerobic Polyphosphate Formation in Methanosarcina Mazei. Sci Rep 2019, 9, 1–12. [CrossRef]
- Mullaymeri, A.; Payr, M.; Wunderer, M.; Eva Maria, E.M.; Wagner, A.O. Shaken Not Stirred - Effect of Different Mixing Modes during the Cultivation of Methanogenic Pure Cultures. Curr Res Microb Sci 2025, 8, 100386. [CrossRef]
- Jeffrey, L.C.; Maher, D.T.; Johnston, S.G.; Kelaher, B.P.; Steven, A.; Tait, D.R. Wetland Methane Emissions Dominated by Plant-Mediated Fluxes: Contrasting Emissions Pathways and Seasons within a Shallow Freshwater Subtropical Wetland. Limnol Oceanogr 2019, 64, 1895–1912. [CrossRef]
- Zhang, M.; Xiao, Q.; Zhang, Z.; Gao, Y.; Zhao, J.; Pu, Y.; Wang, W.; Xiao, W.; Liu, S.; Lee, X. Methane Flux Dynamics in a Submerged Aquatic Vegetation Zone in a Subtropical Lake. Science of The Total Environment 2019, 672, 400–409. [CrossRef]
- Zhu, Y.; Purdy, K.J.; Eyice, Ö.; Shen, L.; Harpenslager, S.F.; Yvon-Durocher, G.; Dumbrell, A.J.; Trimmer, M. Disproportionate Increase in Freshwater Methane Emissions Induced by Experimental Warming. Nat Clim Chang 2020, 10, 685–690. [CrossRef]
- Bolhuis, H.; Cretoiu, M.S.; Stal, L.J. Molecular Ecology of Microbial Mats. FEMS Microbiol Ecol 2014, 90, 335–350. [CrossRef]
- Grossart, H.P.; Frindte, K.; Dziallas, C.; Eckert, W.; Tang, K.W. Microbial Methane Production in Oxygenated Water Column of an Oligotrophic Lake. Proc Natl Acad Sci U S A 2011, 108, 19657–19661. [CrossRef]
- Wang, S.; Sun, P.; Liu, J.; Xu, Y.; Dolfing, J.; Wu, Y. Distribution of Methanogenic and Methanotrophic Consortia at Soil-Water Interfaces in Rice Paddies across Climate Zones. iScience 2023, 26, 105851. [CrossRef]
- Huang, L.; Liu, X.; Rensing, C.; Yuan, Y.; Zhou, S.; Nealson, K.H. Light-Independent Anaerobic Microbial Oxidation of Manganese Driven by an Electrosyntrophic Coculture. ISME Journal 2023, 17, 163–171. [CrossRef]
- Kalathil, S.; Rahaman, M.; Lam, E.; Augustin, T.L.; Greer, H.F.; Reisner, E. Solar-Driven Methanogenesis through Microbial Ecosystem Engineering on Carbon Nitride. Angewandte Chemie International Edition 2024, 63, e202409192. [CrossRef]
- Olson, S.; Jansen, M.F.; Abbot, D.S.; Halevy, I.; Goldblatt, C. The Effect of Ocean Salinity on Climate and Its Implications for Earth’s Habitability. Geophys Res Lett 2022, 49, e2021GL095748. [CrossRef]
- Bardavid, R.E.; Oren, A. The Amino Acid Composition of Proteins from Anaerobic Halophilic Bacteria of the Order Halanaerobiales. Extremophiles 2012, 16, 567–572. [CrossRef]
- Welsh, D.T. Ecological Significance of Compatible Solute Accumulation by Micro- Organisms: From Single Cells to Global Climate. FEMS Microbiol Rev 2000, 24, 263–290. [CrossRef]
- Empadinhas, N.; da Costa, M.S. Osmoadaptation Mechanisms in Prokaryotes: Distribution of Compatible Solutes. Int Microbiol 2008, 11, 151–161. [CrossRef]
- Stüeken, E.E.; Buick, R.; Guy, B.M.; Koehler, M.C. Isotopic Evidence for Biological Nitrogen Fixation by Molybdenum-Nitrogenase from 3.2 Gyr. Nature 2015, 520, 666–669. https://doi.org/10.1038/NATURE14180;TECHMETA=140,58;SUBJMETA=158,181,47,631,704;KWRD=BIOGEOCHEMISTRY,EVOLUTION.
- Hoehler, T.M.; Bebout, B.M.; Des Marais, D.J. The Role of Microbial Mats in the Production of Reduced Gases on the Early Earth. Nature 2001, 412, 324–327. [CrossRef]
- Wang, M.; Ungerfeld, E.M.; Wang, R.; Zhou, C.S.; Basang, Z.Z.; Ao, S.M.; Tan, Z.L. Supersaturation of Dissolved Hydrogen and Methane in Rumen of Tibetan Sheep. Front Microbiol 2016, 7. [CrossRef]
- Catling, D.C. The Great Oxidation Event Transition. Treatise on Geochemistry: Second Edition 2014, 6, 177–195. [CrossRef]
- Jasso-Chávez, R.; Santiago-Martínez, M.G.; Lira-Silva, E.; Pineda, E.; Zepeda-Rodríguez, A.; Belmont-Díaz, J.; Encalada, R.; Saavedra, E.; Moreno-Sánchez, R. Air-Adapted Methanosarcina Acetivorans Shows High Methane Production and Develops Resistance against Oxygen Stress. PLoS One 2015, 10, e0117331. [CrossRef]
- Ernst, L.; Steinfeld, B.; Barayeu, U.; Klintzsch, T.; Kurth, M.; Grimm, D.; Dick, T.P.; Rebelein, J.G.; Bischofs, I.B.; Keppler, F. Methane Formation Driven by Reactive Oxygen Species across All Living Organisms. Nature 2022 603:7901 2022, 603, 482–487. [CrossRef]
- Eliani-Russak, E.; Tik, Z.; Uzi-Gavrilov, S.; Meijler, M.M.; Sivan, O. The Reduction of Environmentally Abundant Iron Oxides by the Methanogen Methanosarcina Barkeri. Front Microbiol 2023, 14, 1197299. [CrossRef]
- Glass, J.B.; Orphan, V.J. Trace Metal Requirements for Microbial Enzymes Involved in the Production and Consumption of Methane and Nitrous Oxide. Front Microbiol 2012, 3, 61. [CrossRef]
- Rossetto, D.; Mansy, S.S. Metals Are Integral to Life as We Know It. Front Cell Dev Biol 2022, 10, 864830. [CrossRef]
- Capdevila, D.A.; Edmonds, K.A.; Giedroc, D.P. Metallochaperones and Metalloregulation in Bacteria. Essays Biochem 2017, 61, 177–200. [CrossRef]
- Summons, R.E.; Welander, P. V.; Gold, D.A. Lipid Biomarkers: Molecular Tools for Illuminating the History of Microbial Life. Nat Rev Microbiol 2022, 20, 174–185. [CrossRef]
- Mulkidjanian, A.Y.; Bychkov, A.Y.; Dibrova, D. V.; Galperin, M.Y.; Koonin, E. V. Origin of First Cells at Terrestrial, Anoxic Geothermal Fields. Proc Natl Acad Sci U S A 2012, 109, E821–E830. https://doi.org/10.1073/PNAS.1117774109/SUPPL_FILE/SAPP.PDF. [CrossRef]
- Ünal, B.; Perry, V.R.; Sheth, M.; Gomez-Alvarez, V.; Chin, K.J.; Nüsslein, K. Trace Elements Affect Methanogenic Activity and Diversity in Enrichments from Subsurface Coal Bed Produced Water. Front Microbiol 2012, 3, 175. [CrossRef]
- Abdel Azim, A.; Rittmann, S.K.M.R.; Fino, D.; Bochmann, G. The Physiological Effect of Heavy Metals and Volatile Fatty Acids on Methanococcus Maripaludis S2. Biotechnol Biofuels 2018, 11, 1–16. doi:10.1186/S13068-018-1302-X/TABLES/2. [CrossRef]
- Spietz, R.L.; Payne, D.; Boyd, E.S. Methanogens Acquire and Bioaccumulate Nickel during Reductive Dissolution of Nickelian Pyrite. Appl Environ Microbiol 2023, 89. https://doi.org/10.1128/AEM.00991-23/SUPPL_FILE/AEM.00991-23-S0002.XLS.
- Glass, J.B.; Chen, S.; Dawson, K.S.; Horton, D.R.; Vogt, S.; Ingall, E.D.; Twining, B.S.; Orphan, V.J. Trace Metal Imaging of Sulfate-Reducing Bacteria and Methanogenic Archaea at Single-Cell Resolution by Synchrotron X-Ray Fluorescence Imaging. Geomicrobiol J 2018, 35, 81–89. [CrossRef]
- Holmes, D.E.; Tang, H.; Woodard, T.; Liang, D.; Zhou, J.; Liu, X.; Lovley, D.R. Cytochrome-Mediated Direct Electron Uptake from Metallic Iron by Methanosarcina Acetivorans. mLife 2022, 1, 443–447. https://doi.org/10.1002/MLF2.12044;JOURNAL:JOURNAL:2770100X;WGROUP:STRING:PUBLICATION.
- Wagner, T.; Ermler, U.; Shima, S. The Methanogenic CO2 Reducing-and-Fixing Enzyme Is Bifunctional and Contains 46 [4Fe-4S] Clusters. Science (1979) 2016, 354, 114–117. https://doi.org/10.1126/SCIENCE.AAF9284/SUPPL_FILE/WAGNER.SM.PDF.
- Demirel, B.; Scherer, P. Trace Element Requirements of Agricultural Biogas Digesters during Biological Conversion of Renewable Biomass to Methane. Biomass Bioenergy 2011, 35, 992–998. [CrossRef]
- Rother, M.; Krzycki, J.A. Selenocysteine, Pyrrolysine, and the Unique Energy Metabolism of Methanogenic Archaea. Archaea 2010, 2010, 453642. [CrossRef]
- Johnson, C.; England, A.; Munro-Ehrlich, M.; Colman, D.R.; DuBois, J.L.; Boyd, E.S. Pathways of Iron and Sulfur Acquisition, Cofactor Assembly, Destination, and Storage in Diverse Archaeal Methanogens and Alkanotrophs. J Bacteriol 2021, 203, 117–138. https://doi.org/10.1128/JB.00117-21/SUPPL_FILE/JB.00117-21-S0005.XLSX.
- Bini, E. Archaeal Transformation of Metals in the Environment. FEMS Microbiol Ecol 2010, 73, 1–16. [CrossRef]
- Konhauser, K.O.; Pecoits, E.; Lalonde, S. V.; Papineau, D.; Nisbet, E.G.; Barley, M.E.; Arndt, N.T.; Zahnle, K.; Kamber, B.S. Oceanic Nickel Depletion and a Methanogen Famine before the Great Oxidation Event. Nature 2009, 458, 750–753. https://doi.org/10.1038/NATURE07858;KWRD=SCIENCE.
- Gottschalk, G.; Thauer, R.K. The Na+-Translocating Methyltransferase Complex from Methanogenic Archaea. Biochimica et Biophysica Acta (BBA) - Bioenergetics 2001, 1505, 28–36. [CrossRef]
- Boone, D.R.; Mah, R.A. Effects of Calcium, Magnesium, PH, and Extent of Growth on the Morphology of Methanosarcina Mazei S-6. Appl Environ Microbiol 1987, 53, 1699–1700. [CrossRef]
- Vanček, M.; Vidová, M.; Majerník, A.I.; Šmigáň, P. Methanogenesis Is Ca2+ Dependent in Methanothermobacter Thermautotrophicus Strain DeltaH. FEMS Microbiol Lett 2006, 258, 269–273. [CrossRef]
- Zhang, G.; Zhang, F.; Ding, G.; Li, J.; Guo, X.; Zhu, J.; Zhou, L.; Cai, S.; Liu, X.; Luo, Y.; et al. Acyl Homoserine Lactone-Based Quorum Sensing in a Methanogenic Archaeon. ISME Journal 2012, 6, 1336–1344. https://doi.org/10.1038/ISMEJ.2011.203;SUBJMETA.
- Yepez-Ceron, O.D.; McCarthy, P.; Patterson, S.; Casey, E.; Dereli, R.K. The Role of Calcium in Anaerobic Treatment: The Good, the Bad and the Ugly. Water Res X 2025, 28, 100381. [CrossRef]
- Thauer, R.K.; Kaster, A.K.; Goenrich, M.; Schick, M.; Hiromoto, T.; Shima, S. Hydrogenases from Methanogenic Archaea, Nickel, a Novel Cofactor, and H2 Storage. Annu Rev Biochem 2010, 79, 507–536. [CrossRef]
- Leimkühler, S. Metal-Containing Formate Dehydrogenases, a Personal View. Molecules 2023, Vol. 28, Page 5338 2023, 28, 5338. [CrossRef]
- Mota, C.S.; Rivas, M.G.; Brondino, C.D.; Moura, I.; Moura, J.J.G.; González, P.J.; Cerqueira, N.M.F.S.A. The Mechanism of Formate Oxidation by Metal-Dependent Formate Dehydrogenases. J Biol Inorg Chem 2011, 16, 1255–1268. [CrossRef]
- Payne, D.; Keller, L.M.; Larson, J.; Bothner, B.; Colman, D.R.; Boyd, E.S. Alternative Sources of Molybdenum for Methanococcus Maripaludis and Their Implication for the Evolution of Molybdoenzymes. Communications Biology 2024 7:1 2024, 7, 1–13. [CrossRef]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy Metals Toxicity and the Environment. EXS 2012, 101, 133. [CrossRef]
- Marcovecchio JE, SE Botté, R.F. Heavy Metals, Major Metals, Trace Elements. In Handbook of Water AnalysisEdition; Nollet, L.M.L., Ed.; CRC Press, Taylor & Francis Group LCC, Boca Ratón: Florida (USA), 2007; pp. 273–210.
- Slyemi, D.; Bonnefoy, V. How Prokaryotes Deal with Arsenic†. Environ Microbiol Rep 2011, 4, no-no. [CrossRef]
- Van Lis, R.; Nitschke, W.; Duval, S.; Schoepp-Cothenet, B. Arsenics as Bioenergetic Substrates. Biochim Biophys Acta Bioenerg 2013, 1827, 176–188.
- Fekih, I. Ben; Zhang, C.; Li, Y.P.; Zhao, Y.; Alwathnani, H.A.; Saquib, Q.; Rensing, C.; Cervantes, C. Distribution of Arsenic Resistance Genes in Prokaryotes. Front Microbiol 2018, 9, 2473. [CrossRef]
- Fru, E.C.; Somogyi, A.; El Albani, A.; Medjoubi, K.; Aubineau, J.; Robbins, L.J.; Lalonde, S. V.; Konhauser, K.O. The Rise of Oxygen-Driven Arsenic Cycling at ca. 2.48 Ga. Geology 2019, 47, 243–246. [CrossRef]
- Sierra-Alvarez, R.; Cortinas, I.; Yenal, U.; Field, J.A. Methanogenic Inhibition by Arsenic Compounds. Appl Environ Microbiol 2004, 70, 5688. [CrossRef]
- Rodriguez-Freire, L.; Moore, S.E.; Sierra-Alvarez, R.; Field, J.A. Adaptation of a Methanogenic Consortium to Arsenite Inhibition. Water Air Soil Pollut 2015, 226, 414. [CrossRef]
- Liang, Y.; Yan, Y.; Shi, L.; Wang, M.; Yuan, X.; Wang, S.; Ye, L.; Yan, Z. Molecular Basis of Thioredoxin-Dependent Arsenic Transformation in Methanogenic Archaea. Environ Sci Technol 2025, 59, 443–453. https://doi.org/10.1021/ACS.EST.4C06611/ASSET/IMAGES/LARGE/ES4C06611_0005.JPEG.
- Lira-Silva, E.; Santiago-Martínez, M.G.; Hernández-Juárez, V.; García-Contreras, R.; Moreno-Sánchez, R.; Jasso-Chávez, R. Activation of Methanogenesis by Cadmium in the Marine Archaeon Methanosarcina Acetivorans. PLoS One 2012, 7, e48779. [CrossRef]
- Lira-Silva, E.; Santiago-Martínez, M.G.; García-Contreras, R.; Zepeda-Rodríguez, A.; Marín-Hernández, A.; Moreno-Sánchez, R.; Jasso-Chávez, R. Cd2+ Resistance Mechanisms in Methanosarcina Acetivorans Involve the Increase in the Coenzyme M Content and Induction of Biofilm Synthesis. Environ Microbiol Rep 2013, 5, 799–808. [CrossRef]
- Singh, R.; Dong, H.; Liu, D.; Zhao, L.; Marts, A.R.; Farquhar, E.; Tierney, D.L.; Almquist, C.B.; Briggs, B.R. Reduction of Hexavalent Chromium by the Thermophilic Methanogen Methanothermobacter Thermautotrophicus. Geochim Cosmochim Acta 2014, 148, 442. [CrossRef]
- Ueno, Y.; Yamada, K.; Yoshida, N.; Maruyama, S.; Isozaki, Y. Evidence from Fluid Inclusions for Microbial Methanogenesis in the Early Archaean Era. Nature 2006 440:7083 2006, 440, 516–519. [CrossRef]
- Wolfe, J.M.; Fournier, G.P. Horizontal Gene Transfer Constrains the Timing of Methanogen Evolution. Nat Ecol Evol 2018, 2, 897–903. [CrossRef]
- Lowe, D.R.; Byerly, G.R. The Terrestrial Record of Late Heavy Bombardment. New Astronomy Reviews 2018, 81, 39–61. [CrossRef]
- Mißbach, H.; Duda, J.P.; van den Kerkhof, A.M.; Lüders, V.; Pack, A.; Reitner, J.; Thiel, V. Ingredients for Microbial Life Preserved in 3.5 Billion-Year-Old Fluid Inclusions. Nat Commun 2021, 12, 1–11. https://doi.org/10.1038/S41467-021-21323-Z;TECHMETA=133,140,58;SUBJMETA=209,213,2151,47,704;KWRD=BIOGEOCHEMISTRY,GEOCHEMISTRY,GEOLOGY.
- Hinrichs, K.U. Microbial Fixation of Methane Carbon at 2.7 Ga: Was an Anaerobic Mechanism Possible? Geochemistry, Geophysics, Geosystems 2002, 3, 1–10. [CrossRef]
- Brocks, J.J.; Logan, G.A.; Buick, R.; Summons, R.E. Archean Molecular Fossils and the Early Rise of Eukaryotes. Science (1979) 1999, 285, 1033–1036. [CrossRef]
- Kohtz, A.J.; Petrosian, N.; Krukenberg, V.; Jay, Z.J.; Pilhofer, M.; Hatzenpichler, R. Cultivation and Visualization of a Methanogen of the Phylum Thermoproteota. Nature 2024 632:8027 2024, 632, 1118–1123. [CrossRef]
- Huang, Y.; Igarashi, K.; Liu, L.; Mayumi, D.; Ujiie, T.; Fu, L.; Yang, M.; Lu, Y.; Cheng, L.; Kato, S.; et al. Methanol Transfer Supports Metabolic Syntrophy between Bacteria and Archaea. Nature 2025, 639, 190–195. https://doi.org/10.1038/S41586-024-08491-W;TECHMETA.
- Cozannet, M.; Le Guellec, S.; Alain, K. A Variety of Substrates for Methanogenesis. Case Studies in Chemical and Environmental Engineering 2023, 8, 100533. [CrossRef]
- Hamilton, J.J.; Calixto Contreras, M.; Reed, J.L. Thermodynamics and H2 Transfer in a Methanogenic, Syntrophic Community. PLoS Comput Biol 2015, 11, e1004364. [CrossRef]
- Kato, S.; Hashimoto, K.; Watanabe, K. Methanogenesis Facilitated by Electric Syntrophy via (Semi)Conductive Iron-Oxide Minerals. Environ Microbiol 2012, 14, 1646–1654. [CrossRef]
- Ishii, ichi; Kosaka, T.; Hori, K.; Hotta, Y.; Watanabe, K. Coaggregation Facilitates Interspecies Hydrogen Transfer between Pelotomaculum Thermopropionicum and Methanothermobacter Thermautotrophicus. Appl Environ Microbiol 2005, 71, 7838–7845. [CrossRef]
- Meulepas, R.J.W.; Jagersma, C.G.; Khadem, A.F.; Stams, A.J.M.; Lens, P.N.L. Effect of Methanogenic Substrates on Anaerobic Oxidation of Methane and Sulfate Reduction by an Anaerobic Methanotrophic Enrichment. APPLIED MICROBIAL AND CELL PHYSIOLOGY 2010, 87, 1499–1506. [CrossRef]
- Stams, A.J.M.; Plugge, C.M. Electron Transfer in Syntrophic Communities of Anaerobic Bacteria and Archaea. Nat Rev Microbiol 2009, 7, 568–577. https://doi.org/10.1038/NRMICRO2166;KWRD.
- Oremland, R.S.; Marsh, L.M.; Polcin, S. Methane Production and Simultaneous Sulphate Reduction in Anoxic, Salt Marsh Sediments. Nature 1982, 296, 143–145. https://doi.org/10.1038/296143A0;KWRD.
- Cappenberg, T.E.; Prins, R.A. Interrelations between Sulfate-Reducing and Methane-Producing Bacteria in Bottom Deposits of a Fresh-Water Lake. III. Experiments with 14C-Labeled Substrates. Antonie Van Leeuwenhoek 1974, 40, 457–469. https://doi.org/10.1007/BF00399358/METRICS.
- Ferry, J.G. Fundamentals of Methanogenic Pathways That Are Key to the Biomethanation of Complex Biomass. Curr Opin Biotechnol 2011, 22, 351. [CrossRef]
- Sorokin, D.Y.; Banciu, H.L.; Muyzer, G. Functional Microbiology of Soda Lakes. Curr Opin Microbiol 2015, 25, 88–96. [CrossRef]
- Wormald, R.M.; Hopwood, J.; Humphreys, P.N.; Mayes, W.; Gomes, H.I.; Rout, S.P. Methanogenesis from Mineral Carbonates, a Potential Indicator for Life on Mars. Geosciences (Switzerland) 2022, 12, 138. [CrossRef]
- Kral, T.A.; Birch, W.; Lavender, L.E.; Virden, B.T. Potential Use of Highly Insoluble Carbonates as Carbon Sources by Methanogens in the Subsurface of Mars. 2014. [CrossRef]
- Fiore, N.A.; Kohtz, A.J.; Miller, D.N.; Antony-Babu, S.; Pan, D.; Lahey, C.; Huang, X.; Lu, Y.; Buan, N.R.; Weber, K.A. Microbial Methane Production from Calcium Carbonate at Moderately Alkaline PH. Commun Earth Environ 2025, 6, 1–13. [CrossRef]
- Tian, F.; Toon, O.B.; Pavlov, A.A.; De Sterck, H. A Hydrogen-Rich Early Earth Atmosphere. Science (1979) 2005, 308, 1014–1017. [CrossRef]
- Novelli, P.C.; Lang, P.M.; Masarie, K.A.; Hurst, D.F.; Myers, R.; Elkins, J.W. Molecular Hydrogen in the Troposphere: Global Distribution and Budget. Journal of Geophysical Research: Atmospheres 1999, 104, 30427–30444. [CrossRef]
- Mei, R.; Kaneko, M.; Imachi, H.; Nobu, M.K. The Origin and Evolution of Methanogenesis and Archaea Are Intertwined. PNAS Nexus 2023, 2, 1–10. [CrossRef]
- Weiss, M.C.; Soussa, F.L.; Mrnjavac, N.; Neukirchen, S.; Roettger, M.; Nelson-Sathi, S.; Martin, W.F. The Physiology and Habitat of the Last Universal Common Ancestor. Nat Microbiol 2016, 1, 16116. [CrossRef]
- Schopf, J.W.; Kitajima, K.; Spicuzza, M.J.; Kudryavtsev, A.B.; Valley, J.W. SIMS Analyses of the Oldest Known Assemblage of Microfossils Document Their Taxon-Correlated Carbon Isotope Compositions. Proc Natl Acad Sci U S A 2018, 115, 53–58. [CrossRef]
- Foster, J.S.; Mobberley, J.M. Past, Present, and Future: Microbial Mats as Models for Astrobiological Research. 2010, 563–582. [CrossRef]
- Xia, L.; Cao, J.; Hu, W.; Zhi, D.; Tang, Y.; Li, E.; He, W. Coupling of Paleoenvironment and Biogeochemistry of Deep-Time Alkaline Lakes: A Lipid Biomarker Perspective. Earth Sci Rev 2021, 213, 103499. [CrossRef]
- Zhao, Z.; Feng, Q.; Lu, H.; Peng, P.; Zhang, T.; Hsu, C.S. Stable Carbon Isotopic Fractionation and Hydrocarbon Generation Mechanism of CO2 Fischer–Tropsch-Type Synthesis under Hydrothermal Conditions. Energy & Fuels 2021, 35, 11909–11919. [CrossRef]
- Rezaee, R. Quantifying Natural Hydrogen Generation Rates and Volumetric Potential in Onshore Serpentinization. Geosciences (Basel) 2025, 3, 112. [CrossRef]
- Kasting, J.F.; Siefert, J.L. Life and the Evolution of Earth’s Atmosphere. Science (1979) 2002, 296, 1066–1068. [CrossRef]
- Ding, Y.; Wang, M.; Du, X.; He, X.; Xu, T.; Liu, X.; Song, F. The Impact of Elevated CO2 on Methanogen Abundance and Methane Emissions in Terrestrial Ecosystems: A Meta-Analysis. iScience 2024, 27, 111504. [CrossRef]
- Olson, S.L.; Reinhard, C.T.; Lyons, T.W. Limited Role for Methane in the Mid-Proterozoic Greenhouse. Proc Natl Acad Sci U S A 2016, 113, 11447–11452. [CrossRef]
- Canfield, D.E.; Raiswell, R. The Evolution of the Sulfur Cycle. Am J Sci 1999, 299, 697–723. [CrossRef]
- Steward, K.F.; Payne, D.; Kincannon, W.; Johnson, C.; Lensing, M.; Fausset, H.; Németh, B.; Shepard, E.M.; Broderick, W.E.; Broderick, J.B.; et al. Proteomic Analysis of Methanococcus Voltae Grown in the Presence of Mineral and Nonmineral Sources of Iron and Sulfur. Microbiol Spectr 2022, 10. [CrossRef]
- Payne, D.; Spietz, R.L.; Boyd, E.S. Reductive Dissolution of Pyrite by Methanogenic Archaea. ISME J 2021, 15, 3498–3507. [CrossRef]
- Canfield, D.E. Biogeochemistry of Sulfur Isotopes. Rev Mineral Geochem 2001, 43, 607–636. [CrossRef]
- Canfield, D.E.; Habicht, K.S.; Thamdrup, B. The Archean Sulfur Cycle and the Early History of Atmospheric Oxygen. Science (1979) 2000, 288, 658–661. https://doi.org/10.1126/SCIENCE.288.5466.658/SUPPL_FILE/CANFIELD.XHTML.
- Fakhraee, M.; Crockford, P.W.; Bauer, K.W.; Pasquier, V.; Sugiyama, I.; Katsev, S.; Raven, M.R.; Gomes, M.; Philippot, P.; Crowe, S.A.; et al. The History of Earth’s Sulfur Cycle. Nat Rev Earth Environ 2025, 6, 106–125. [CrossRef]
- Fakhraee, M.; Katsev, S. Organic Sulfur Was Integral to the Archean Sulfur Cycle. Nat Commun 2019, 10, 1–8. [CrossRef]
- Reeves, E.P.; McDermott, J.M.; Seewald, J.S. The Origin of Methanethiol in Midocean Ridge Hydrothermal Fluids. Proceedings of the National Academy of Sciences 2014, 111, 5474–5479. [CrossRef]
- Baumgartner, R.J.; Van Kranendonk, M.J.; Caruso, S.; Campbell, K.A.; Dobson, M.J.; Teece, B.L.; Verrall, M.; Homann, M.; Lalonde, S.; Visscher, P.T. Pyritic Stromatolites from the Paleoarchean Dresser Formation, Pilbara Craton: Resolving Biogenicity and Hydrothermally Influenced Ecosystem Dynamics. Geobiology 2024, 22. [CrossRef]
- Xiao, X.; Wang, Y.; Huang, F.; Yang, Y.; Long, Y.; Fang, C.; Hu, L. Uncover Co-Evolution of Arsenic Transformation and Methanogens in Landfills. J Hazard Mater 2025, 497, 139623. [CrossRef]
- Moody, E.R.R.; Álvarez-Carretero, S.; Mahendrarajah, T.A.; Clark, J.W.; Betts, H.C.; Dombrowski, N.; Szánthó, L.L.; Boyle, R.A.; Daines, S.; Chen, X.; et al. The Nature of the Last Universal Common Ancestor and Its Impact on the Early Earth System. Nature Ecology & Evolution 2024 8:9 2024, 8, 1654–1666. [CrossRef]
- Hren, M.T.; Tice, M.M.; Chamberlain, C.P. Oxygen and Hydrogen Isotope Evidence for a Temperate Climate 3.42 Billion Years Ago. Nature 2009 462:7270 2009, 462, 205–208. [CrossRef]
- Blake, R.E.; Chang, S.J.; Lepland, A. Phosphate Oxygen Isotopic Evidence for a Temperate and Biologically Active Archaean Ocean. Nature 2010 464:7291 2010, 464, 1029–1032. [CrossRef]
- Penny, D.; Poole, A. The Nature of the Last Universal Common Ancestor. Current Opinions in Genetics & Development 1999, 9, 672–677.
- Liu, R.; Gong, H.; Xu, Y.; Cai, C.; Hua, Y.; Li, L.; Dai, L.; Dai, X. The Transition Temperature (42 °C) from Mesophilic to Thermophilic Micro-Organisms Enhances Biomethane Potential of Corn Stover. Science of The Total Environment 2021, 759, 143549. [CrossRef]
- Catling, D.C.; Zahnle, K.J.; McKay, C.P. Biogenic Methane, Hydrogen Escape, and the Irreversible Oxidation of Early Earth. Science (1979) 2001, 293, 839–843. [CrossRef]
- Yang, S.; Lv, Y.; Liu, X.; Wang, Y.; Fan, Q.; Yanf, Z.; Boon, N.; Wang, F.; Xiao, X.; Zhang, Y. Genomic and Enzymatic Evidence of Acetogenesis by Anaerobic Methanotrophic Archaea. Nat Commun 2020, 11, 3941. [CrossRef]
- Ettwig, K.F.; Zhu, B.; Speth, D.; Keltjens, J.T.; Jetten, M.S.M.; Kartal, B. Archaea Catalyze Iron-Dependent Anaerobic Oxidation of Methane. Proceedings of the National Academy of Sciences 2016, 113, 12792–12796. [CrossRef]
- Pavlov, A.A.; Kasting, J.F.; Eigenbrode, J.L.; Freeman, K.H. Organic Haze in Earth’s Early Atmosphere: Source of Low-13C Late Archean Kerogens? | Geology | GeoScienceWorld. Geology 2001, 29, 1003–1006.
- Tian, F.; Kasting, J.F.; Zahnle, K. Revisiting HCN Formation in Earth’s Early Atmosphere. Earth Planet Sci Lett 2011, 308, 417–423. [CrossRef]
- Wolf, E.T.; Toon, O.B. Fractal Organic Hazes Provided an Ultraviolet Shield for Early Earth. Science (1979) 2010, 328, 1266–1268. [CrossRef]
- Dupraz, C.; Visscher, P.T. Microbial Lithification in Marine Stromatolites and Hypersaline Mats. Trends Microbiol 2005, 13, 429–438. [CrossRef]
- Colman, D.R.; Templeton, A.S.; Spear, J.R.; Boyd, E.S. Microbial Ecology of Serpentinite-Hosted Ecosystems. ISME J 2025, 19, 29. [CrossRef]
- Shibuya, T.; Komiya, T.; Nakamura, K.; Takai, K.; Maruyama, S. Highly Alkaline, High-Temperature Hydrothermal Fluids in the Early Archean Ocean. Precambrian Res 2010, 182, 230–238. [CrossRef]
- Preiner, M.; Xavier, J.C.; Sousa, F.L.; Zimorski, V.; Neubeck, A.; Lang, S.Q.; Chris Greenwell, H.; Kleinermanns, K.; Tüysüz, H.; McCollom, T.M.; et al. Serpentinization: Connecting Geochemistry, Ancient Metabolism and Industrial Hydrogenation. Life (Basel) 2018, 8. [CrossRef]
- Georgieva, M.N.; Little, C.T.S.; Bailey, R.J.; Ball, A.D.; Glover, A.G. Microbial-Tubeworm Associations in a 440 Million Year Old Hydrothermal Vent Community. Proceedings of the Royal Society B 2018, 285. [CrossRef]
- Dodd, M.S.; Papineau, D.; Grenne, T.; Slack, J.F.; Rittner, M.; Pirajno, F.; O’Neil, J.; Little, C.T.S. Evidence for Early Life in Earth’s Oldest Hydrothermal Vent Precipitates. Nature 2017, 543, 60–64. [CrossRef]
- Aoyama, S.; Nishizawa, M.; Miyazaki, J.; Shibuya, T.; Ueno, Y.; Takai, K. Recycled Archean Sulfur in the Mantle Wedge of the Mariana Forearc and Microbial Sulfate Reduction within an Extremely Alkaline Serpentine Seamount. Earth Planet Sci Lett 2018, 491, 109–120. [CrossRef]
- Tosca, N.J.; Tutolo, B.M. Hydrothermal Vent Fluid-Seawater Mixing and the Origins of Archean Iron Formation. Geochim Cosmochim Acta 2023, 352, 51–68. [CrossRef]
- Bjerrum, C.J.; Canfield, D.E. Ocean Productivity before about 1.9 Gyr Ago Limited by Phosphorus Adsorption onto Iron Oxides. Nature 2002, 417, 159–162. [CrossRef]
- Martin, W.; Russell, M.J. On the Origin of Biochemistry at an Alkaline Hydrothermal Vent. Philos Trans R Soc Lond B Biol Sci 2007, 362, 1887–1926. [CrossRef]
- Wang, Y.; Fu, X.; Wang, J.; Wei, H.; Zeng, Y.; Ahmed, M.S.; Mansour, A. Methane Emissions during Marine Environmental Perturbations before the End-Triassic Mass Extinction. Facies 2025, 71, 1–12. [CrossRef]
- Mrnjavac, N.; Schwander, L.; Brabender, M.; Martin, W.F. Chemical Antiquity in Metabolism. Acc Chem Res 2024, 57, 2267–2278. [CrossRef]
- Bae, H.S.; Morrison, E.; Chanton, J.P.; Ogram, A. Methanogens Are Major Contributors to Nitrogen Fixation in Soils of the Florida Everglades. Appl Environ Microbiol 2018, 84. [CrossRef]
- Evans, G.N.; Ji, S.; Kaçar, B.; Anbar, A.D.; Seyfried, W.E. Transition Metals in Alkaline Lost City Vent Fluids Are Sufficient for Early-Life Metabolisms. Geochim Cosmochim Acta 2024, 385, 61–73. [CrossRef]
- Noffke, N.; Christian, D.; Wacey, D.; Hazen, R.M. Microbially Induced Sedimentary Structures Recording an Ancient Ecosystem in the ca. 3.48 Billion-Year-Old Dresser Formation, Pilbara, Western Australia. Astrobiology 2013, 13, 1103–1124. [CrossRef]
- Gutierrez-Preciado, A.; Saghai, A.; Moreira, D.; Zivanovic, Y.; Dschamps, P.; Lopez-Garcia, P. Functional Shifts in Microbial Mats Recapitulate Early Earth Metabolic Transitions. Nat Ecol Evol 2018, 2, 1700–1708. [CrossRef]
- Nisbet, E.G.; Fowler, C.M.R. Archaean Metabolic Evolution of Microbial Mats. Proc R Soc Lond B Biol Sci 1999, 266, 2375–2382. [CrossRef]
- Blank, C.E.; SÁnchez-Baracaldo, P. Timing of Morphological and Ecological Innovations in the Cyanobacteria - A Key to Understanding the Rise in Atmospheric Oxygen. Geobiology 2010, 8, 1–23. [CrossRef]
- Knoll, A.H.; Nowak, M.A. The Timetable of Evolution. Sci Adv 2017, 3. [CrossRef]
- Hamilton, T.L.; Bryant, D.A.; Macalady, J.L. The Role of Biology in Planetary Evolution: Cyanobacterial Primary Production in Low-Oxygen Proterozoic Oceans. Environ Microbiol 2016, 18, 325–340. [CrossRef]
- Noffke, N.; Gerdes, G.; Klenke, T.; Krumbein, W.E. Microbially Induced Sedimentary Structures: A New Category within the Classification of Primary Sedimentary Structures. Journal of Sedimentary Research 2001, 71, 649–656. [CrossRef]
- Dupraz, C.; Reid, P.R.; Braissant, O.; Decho, A.W.; Normal, S.R.; Visscher, P.T. Processes of Carbonate Precipitation in Modern Microbial Mats. Earth Sci Rev 2009, 96, 141–162. [CrossRef]
- Allwood, A.C.; Walter, M.R.; Kamber, B.S.; Marshall, C.P.; Burch, I.W. Stromatolite Reef from the Early Archaean Era of Australia. Nature 2006, 441, 714–718. [CrossRef]
- Allwood, A.C.; Walter, M.R.; Burch, I.W.; Kamber, B.S. 3.43 Billion-Year-Old Stromatolite Reef from the Pilbara Craton of Western Australia: Ecosystem-Scale Insights to Early Life on Earth. Precambrian Res 2007, 158, 198–227. [CrossRef]
- Nutman, A.P.; Bennett, V.C.; Friend, C.R.L.; Van Kranendonk, M.J.; Chivas, A.R. Rapid Emergence of Life Shown by Discovery of 3,700-Million-Year-Old Microbial Structures. Nature 2016 537:7621 2016, 537, 535–538. [CrossRef]
- Van Kranendonk, M.J.; Nutman, A.P.; Friend, C.R.L.; Bennett, V.C. A Review of 3.7 Ga Stromatolites from the Isua Supracrustal Belt, West Greenland. Earth Sci Rev 2025, 262, 105034. [CrossRef]
- Allwood, A.C.; Rosing, M.T.; Flannery, D.T.; Hurowitz, J.A.; Heirwegh, C.M. Reassessing Evidence of Life in 3,700-Million-Year-Old Rocks of Greenland. Nature 2018, 563, 241–244. [CrossRef]
- Allwood, A.C.; Grotzinger, J.P.; Knoll, A.H.; Burch, I.W.; Anderson, M.S.; Coleman, M.L.; Kanik, I. Controls on Development and Diversity of Early Archean Stromatolites. Proceedings of the National Academy of Sciences 2009, 106, 9548–9555. [CrossRef]
- Watanabe, Y.; Tajika, E.; Ozaki, K. Biogeochemical Transformations after the Emergence of Oxygenic Photosynthesis and Conditions for the First Rise of Atmospheric Oxygen. Geobiology 2023, 21, 537–555. [CrossRef]
- Anbar, A.D. Oceans. Elements and Evolution. 2008, 322, 1481–1483, doi:doi:10.1126/science.1163100.
- Mukherjee, I.; Large, R.R.; Corkrey, R.; Danyushevsky, L. V. The Boring Billion, a Slingshot for Complex Life on Earth. Sci Rep 2018, 8, 1–7. [CrossRef]
- Stott, L.D.; Harazin, K.M.; Quintana Krupinski, N.B. Hydrothermal Carbon Release to the Ocean and Atmosphere from the Eastern Equatorial Pacific during the Last Glacial Termination. Environmental Research Letters 2019, 14. [CrossRef]
- Buffett, B.A. Clathrate Hydrates. Annu Rev Earth Planet Sci 2000, 28, 477–507. [CrossRef]
- Huard, D.J.E.; Johnson, A.M.; Fan, Z.; Kenney, L.G.; Xu, M.; Drori, R.; Gumbart, J.C.; Dai, S.; Lieberman, R.L.; Glass, J.B. Molecular Basis for Inhibition of Methane Clathrate Growth by a Deep Subsurface Bacterial Protein. PNAS Nexus 2023, 2. [CrossRef]
- Weitemeyer, K.A.; Buffett, B.A. Accumulation and Release of Methane from Clathrates below the Laurentide and Cordilleran Ice Sheets. Glob Planet Change 2006, 53, 176–187. [CrossRef]
- Petryshyn, V.A.; Greene, S.E.; Farnsworth, A.; Lunt, D.J.; Kelley, A.; Gammariello, R.; Ibarra, Y.; Bottjer, D.J.; Tripati, A.; Corsetti, F.A. The Role of Temperature in the Initiation of the End-Triassic Mass Extinction. Earth Sci Rev 2020, 208, 103266. [CrossRef]
- Brand, U.; Blamey, N.; Garbelli, C.; Griesshaber, E.; Posenato, R.; Angiolini, L.; Azmy, K.; Farabegoli, E.; Came, R. Methane Hydrate: Killer Cause of Earth’s Greatest Mass Extinction. Palaeoworld 2016, 25, 496–507. [CrossRef]
- Burgess, S.D.; Black, B.A. The Anatomy and Lethality of the Siberian Traps Large Igneous Province. Annu Rev Earth Planet Sci 2025, 53, 567–594. doi:10.1146/ANNUREV-EARTH-040722-105544/CITE/REFWORKS. [CrossRef]
- Rothman, D.H.; Fournier, G.P.; French, K.L.; Alm, E.J.; Boyle, E.A.; Cao, C.; Summons, R.E. Methanogenic Burst in the End-Permian Carbon Cycle. Proceedings of the National Academy of Sciences 2014, 111, 5462–5467. [CrossRef]
- Fournier, G.P.; Gogarten, J.P. Evolution of Acetoclastic Methanogenesis in Methanosarcina via Horizontal Gene Transfer from Cellulolytic Clostridia. J Bacteriol 2008, 190, 1124–1127. [CrossRef]
- Strock, K.E.; Krewson, R.B.; Hayes, N.M.; Deemer, B.R. Oxidation Is a Potentially Significant Methane Sink in Land-Terminating Glacial Runoff. Sci Rep 2024, 14, 1–9. [CrossRef]
- Luo, G.; Liu, D.; Yang, H. Microbes in Mass Extinction: An Accomplice or a Savior? Natl Sci Rev 2023, 11, 2024. [CrossRef]
- Lau, K. V.; Maher, K.; Altiner, D.; Kelley, B.M.; Kump, L.R.; Lehrmann, D.J.; Silva-Tamayo, J.C.; Weaver, K.L.; Yu, M.; Payne, J.L. Marine Anoxia and Delayed Earth System Recovery after the End-Permian Extinction. Proc Natl Acad Sci U S A 2016, 113, 2360–2365. https://doi.org/10.1073/PNAS.1515080113;WEBSITE:WEBSITE:PNAS-SITE;WGROUP:STRING:PUBLICATION. [CrossRef]
- Mata, S.A.; Bottjer, D.J. Microbes and Mass Extinctions: Paleoenvironmental Distribution of Microbialites during Times of Biotic Crisis. Geobiology 2012, 10, 3–24. [CrossRef]
- Shubert, J.; Bottjer, D.J. Early Triassic Stromatolites as Post-Mass Extinction Disaster Forms | Geology | GeoScienceWorld. Geology 1992, 10, 883–886.
- Kirton, J.M.C.; Woods, A.D. Stromatolites from the Lower Triassic Virgin Limestone at Blue Diamond, NV USA: The Role of Dysoxia, Enhanced Calcification and Nutrient Availability in the Growth of Post-Extinction Microbialites. Glob Planet Change 2021, 198, 103429. [CrossRef]
- Baud, A.; Cirilli, S.; Marcoux, J. BIOTIC RESPONSE TO MASS EXTINCTION: THE LOWERMOST TRIASSIC MICROBIALITES.
- Vennin, E.; Olivier, N.; Brayard, A.; Bour, I.; Thomazo, C.; Escarguel, G.; Fara, E.; Bylund, K.G.; Jenks, J.F.; Stephen, D.A.; et al. Microbial Deposits in the Aftermath of the End-Permian Mass Extinction: A Diverging Case from the Mineral Mountains (Utah, USA). Sedimentology 2015, 62, 753–792. [CrossRef]
- Schobben, M.; Stebbins, A.; Ghaderi, A.; Strauss, H.; Korn, D.; Korte, C. Flourishing Ocean Drives the End-Permian Marine Mass Extinction. Proc Natl Acad Sci U S A 2015, 112, 10298–10303. [CrossRef]
- Wu, S.; Chen, Z.Q.; Fang, Y.; Pei, Y.; Yang, H.; Ogg, J. A Permian-Triassic Boundary Microbialite Deposit from the Eastern Yangtze Platform (Jiangxi Province, South China): Geobiologic Features, Ecosystem Composition and Redox Conditions. Palaeogeogr Palaeoclimatol Palaeoecol 2017, 486, 58–73. [CrossRef]
- Luo, G.; Kump, L.R.; Wang, Y.; Tong, J.; Arthur, M.A.; Yang, H.; Huang, J.; Yin, H.; Xie, S. Isotopic Evidence for an Anomalously Low Oceanic Sulfate Concentration Following End-Permian Mass Extinction. Earth Planet Sci Lett 2010, 300, 101–111. [CrossRef]
- Sheehan, P.M.; Harris, M.T. Microbialite Resurgence after the Late Ordovician Extinction. Nature 2004, 430, 75–78. [CrossRef]
- Yao, L.; Aretz, M.; Chen, J.; Webb, G.E.; Wang, X. Global Microbial Carbonate Proliferation after the End-Devonian Mass Extinction: Mainly Controlled by Demise of Skeletal Bioconstructors. Sci Rep 2016, 6, 1–9. [CrossRef]
- Liu, Y.; Whitman, W.B. Metabolic, Phylogenetic, and Ecological Diversity of the Methanogenic Archaea. Ann N Y Acad Sci 2008, 1125, 171–189. [CrossRef]
- Ehhalt, D.H. The Atmospheric Cycle of Methane. Tellus A: Dynamic Meteorology and Oceanography 1974, 26, 58. [CrossRef]
- Skeie, R.B.; Hodnebrog, Ø.; Myhre, G. Trends in Atmospheric Methane Concentrations since 1990 Were Driven and Modified by Anthropogenic Emissions. Commun Earth Environ 2023, 4, 1–14. [CrossRef]
- Veron, J.E.N.; Hoegh-Guldberg, O.; Lenton, T.M.; Lough, J.M.; Obura, D.O.; Pearce-Kelly, P.; Sheppard, C.R.C.; Spalding, M.; Stafford-Smith, M.G.; Rogers, A.D. The Coral Reef Crisis: The Critical Importance of .
- Ehleringer, J.R.; Cerling, T.E. Atmospheric CO2 and the Ratio of Intercellular to Ambient CO2 Concentrations in Plants. Tree Physiol 1993, 15, 105–111. [CrossRef]
- Dlugokencky, E.J.; Nisbet, E.G.; Fisher, R.; Lowry, D. Global Atmospheric Methane: Budget, Changes and Dangers. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences 2011, 369, 2058–2072. [CrossRef]
- Visscher, P.T.; Stolz, J.F. Microbial Mats as Bioreactors: Populations, Processes, and Products. Paleogeography, Palaeoclimatology, Palaeoecology 2005, 1, 87–100. [CrossRef]
- Veiga, M.C.; Jain, M.K.; Wu, W.M.; Hollingsworth, R.I.; Zeikus, J.G. Composition and Role of Extracellular Polymers in Methanogenic Granules. Appl Environ Microbiol 1997, 63, 403–407. [CrossRef]
- Su, F.; Yang, Y.Y. Microbially Induced Carbonate Precipitation via Methanogenesis Pathway by a Microbial Consortium Enriched from Activated Anaerobic Sludge. J Appl Microbiol 2021, 131, 236–256. [CrossRef]
- Sowers, K.R.; Baron, S.F.; Ferry, J.G. Methanosarcina Acetivorans Sp. Nov., an Acetotrophic Methane-Producing Bacterium Isolated from Marine Sediments . Appl Environ Microbiol 1984, 47, 971–978. [CrossRef]
- Loyd, S.J.; Marissa N. Smirnoff Progressive Formation of Authigenic Carbonate with Depth in Siliciclastic Marine Sediments Including Substantial Formation in Sediments Experiencing. Chem Geol 2022, 120775. [CrossRef]
- Beckmann, S.; Farag, I.F.; Zhao, R.; Christman, G.D.; Prouty, N.G.; Biddle, J.F. Expanding the Repertoire of Electron Acceptors for the Anaerobic Oxidation of Methane in Carbonates in the Atlantic and Pacific Ocean. ISME J 2021, 2523–2536. [CrossRef]
- Cai, S.; Wei, G.; Xian, H.; Zhu, J.; Xu, Y.G. Methanogen-Mediated Precipitation of Mn Carbonates at the Expense of Mn Oxides. Geophys Res Lett 2024, 51. [CrossRef]
- Ramírez-Arenas, P.J.; Latisnere-Barragán, H.; García-Maldonado, J.Q.; López-Cortés, A. Highly Diverse–Low Abundance Methanogenic Communities in Hypersaline Microbial Mats of Guerrero Negro B.C.S., Assessed through Microcosm Experiments. PLoS One 2024, 19, e0303004. [CrossRef]
- Oremland, R.S.; Polcin, S. Methanogenesis and Sulfate Reduction: Competitive and Noncompetitive Substrates in Estuarine Sediments. Appl Environ Microbiol 1982, 44, 1270. [CrossRef]
- Mitterer, R.M. Methanogenesis and Sulfate Reduction in Marine Sediments: A New Model. Earth Planet Sci Lett 2010, 295, 358–366. [CrossRef]
- Vogt, J.C.; Abed, R.M.M.; Albach, D.C.; Palinska, K.A. Bacterial and Archaeal Diversity in Hypersaline Cyanobacterial Mats Along a Transect in the Intertidal Flats of the Sultanate of Oman. Microb Ecol 2018, 75, 331–347. https://doi.org/10.1007/S00248-017-1040-9/FIGURES/6. [CrossRef]
- Visscher, P. Cellular Origin, Life in Extreme Habitats and Astrobiology ; 2010; Vol. 14;.
- Visscher, P.T.; Taylor, B.F.; Kiene, R.P. Microbial Consumption of Dimethyl Sulfide and Methanethiol in Coastal Marine Sediments. FEMS Microbiol Ecol 1995, 18, 145–154. [CrossRef]
- Visscher, P.T.; Reid, R.P.; Bebout, B.M.; Hoeft, S.E.; Macintyre, I.G.; Thompson, J.A. Formation of Lithified Micritic Laminae in Modern Marine Stromatolites (Bahamas); the Role of Sulfur Cycling. American Mineralogist 1998, 83, 1482–1493. [CrossRef]
- Djokic, T.; VanKranendonk, M.J.; Campbel, K.A.; Walter, M.R.; Ward, C.R. Earliest Signs of Life on Land Preserved in ca. 3.5 Ga Hot Spring Deposits. Nat Commun 2017, 8, 1–9. [CrossRef]
- Baumgartner, L.K.; Dupraz, C.; Buckley, D.H.; Spear, J.R.; Pace, N.R.; Visscher, P.T. Microbial Species Richness and Metabolic Activities in Hypersaline Microbial Mats: Insight into Biosignature Formation Through Lithification. https://home.liebertpub.com/ast 2009, 9, 861–874. [CrossRef]
- Walter, M.R.; Heys, G.R. Links between the Rise of the Metazoa and the Decline of Stromatolites. Precambrian Res 1985, 29, 149–174. [CrossRef]
- Hohl, S. V.; Bian, X.; Viehmann, S.; Yang, S.C.; Raad, R.J.; Meister, P.; John, S.G. A Novel Biomarker for Deep-Time Methanogenesis – Perspectives from Nickel Isotope Fractionation in Modern Microbialites. Earth Planet Sci Lett 2025, 666, 119492. [CrossRef]
- Coffey, J.M.; Flannery, D.T.; Walter, M.R.; George, S.C. Sedimentology, Stratigraphy and Geochemistry of a Stromatolite Biofacies in the 2.72 Ga Tumbiana Formation, Fortescue Group, Western Australia. Precambrian Res 2013, 236, 282–296. [CrossRef]
- Scheller, S.; Goenrich, M.; Thauer, R.K.; Jaun, B. Methyl-Coenzyme M Reductase from Methanogenic Archaea: Isotope Effects on the Formation and Anaerobic Oxidation of Methane. 2013. [CrossRef]
- Van de Vossenberg, J.L.C.M.; Driessen, A.J.M.; Konings, W.N. The Essence of Being Extremophilic: The Role of the Unique Archaeal Membrane Lipids. Extremophiles 1998, 2, 163–170. [CrossRef]
- Myers, T.; Dykstra, C.M. Teaching Old Dogs New Tricks: Genetic Engineering Methanogens. Appl Environ Microbiol 2024, 90. [CrossRef]
- Conrad, R. Methane Production in Soil Environments—Anaerobic Biogeochemistry and Microbial Life between Flooding and Desiccation. Microorganisms 2020, Vol. 8, Page 881 2020, 8, 881. [CrossRef]
- Horne, A.J.; Lessner, D.J. Assessment of the Oxidant Tolerance of Methanosarcina Acetivorans. FEMS Microbiol Lett 2013, 343, 13–19. [CrossRef]
- Decho, A.W.; Norman, R.S.; Visscher, P.T. Quorum Sensing in Natural Environments: Emerging Views from Microbial Mats. Trends Microbiol 2010, 18, 73–80. [CrossRef]
- Guerrero-Cruz, S.; Cremers, G.; van Alen, T.A.; Op den Camp, H.J.M.; Jetten, M.S.M.; Rasigraf, O.; Vaksmaa, A. Response of the Anaerobic Methanotroph “ Candidatus Methanoperedens Nitroreducens” to Oxygen Stress. Appl Environ Microbiol 2018, 84. [CrossRef]
- Magalhães, C.P.; Duarte, M.S.; Pereira, M.A.; Stams, A.J.M.; Cavaleiro, A.J. Facultative Anaerobic Bacteria Enable Syntrophic Fatty Acids Degradation under Micro-Aerobic Conditions. Bioresour Technol 2025, 417, 131829. [CrossRef]
- Zhao, Y.-C.; Chong, S.+; Zhao, X.-M.; Du, J.-X.; Zou, L.; Yong, Y.-C. Unnatural Direct Interspecies Electron Transfer Enabled by Living Cell-Cell Click Chemistry. Angewandte Chemie 2024, 136, e202402318. [CrossRef]
- Formisano, V.; Atreya, S.; Encrenaz, T.; Ignatiev, N.; Giuranna, M. Detection of Methane in the Atmosphere of Mars. Science (1979) 2004, 306, 1758–1761. [CrossRef]
- Waite, J.H.; Combi, M.R.; Ip, W.; Thomas, E.C.; McNutt, R.L.; Kasprzak, W.; Yelle, R.; Luhmann, J.; Niemann, H.; Gell, D.; et al. Cassini Ion and Neutral Mass Spectrometer: Enceladus Plume Composition and Structure. Science 2006, 311, 1419–1422. [CrossRef]
- Niemann, H.B.; Atreya, S.K.; Bauer, S.J.; Carignan, G.R.; Demick, J.E.; Frost, R.L.; Gautier, D.; Haberman, J.A.; Harpold, D.N.; Hunten, D.M.; et al. The Abundances of Constituents of Titan’s Atmosphere from the GCMS Instrument on the Huygens Probe. Nature 2005, 438, 779–784. [CrossRef]
- Mickol, R.L.; Takagi, Y.A.; Kral, T.A. Survival of Non-Psychrophilic Methanogens Exposed to Martian Diurnal and 48-h Temperature Cycles. Planet Space Sci 2018, 157, 63–71. [CrossRef]
- Hurowitz, J.A.; Catling, D.C.; Fischer, W.W. High Carbonate Alkalinity Lakes on Mars and Their Potential Role in an Origin of Life Beyond Earth LAKES ON MARS. ElEmEnts 2023, 19, 37–44. [CrossRef]
- Yung, Y.L.; Chen, P.; Nealson, K.; Atreya, S.; Beckett, P.; Blank, J.G.; Ehlmann, B.; Eiler, J.; Etiope, G.; Ferry, J.G.; et al. Methane on Mars and Habitability: Challenges and Responses. Astrobiology 2018, 18, 1221–1242. [CrossRef]
Figure 3.
Hypothetical interactions between cyanobacteria, methanogens and methane oxidizers in the oxic layer of a microbial mat. Quorum sensing (red arrows) between consortia of cyanobacteria, sulfate reducers, and sulfide-oxidizing bacteria was proposed to allow for maximum metabolic potential under unfavorable conditions [279]. Methanogens (MA) may facilitate quorum sensing in a similar manner between methane oxidizers (MO), and/or cyanobacteria [4]. Modified from [279].
Figure 3.
Hypothetical interactions between cyanobacteria, methanogens and methane oxidizers in the oxic layer of a microbial mat. Quorum sensing (red arrows) between consortia of cyanobacteria, sulfate reducers, and sulfide-oxidizing bacteria was proposed to allow for maximum metabolic potential under unfavorable conditions [279]. Methanogens (MA) may facilitate quorum sensing in a similar manner between methane oxidizers (MO), and/or cyanobacteria [4]. Modified from [279].
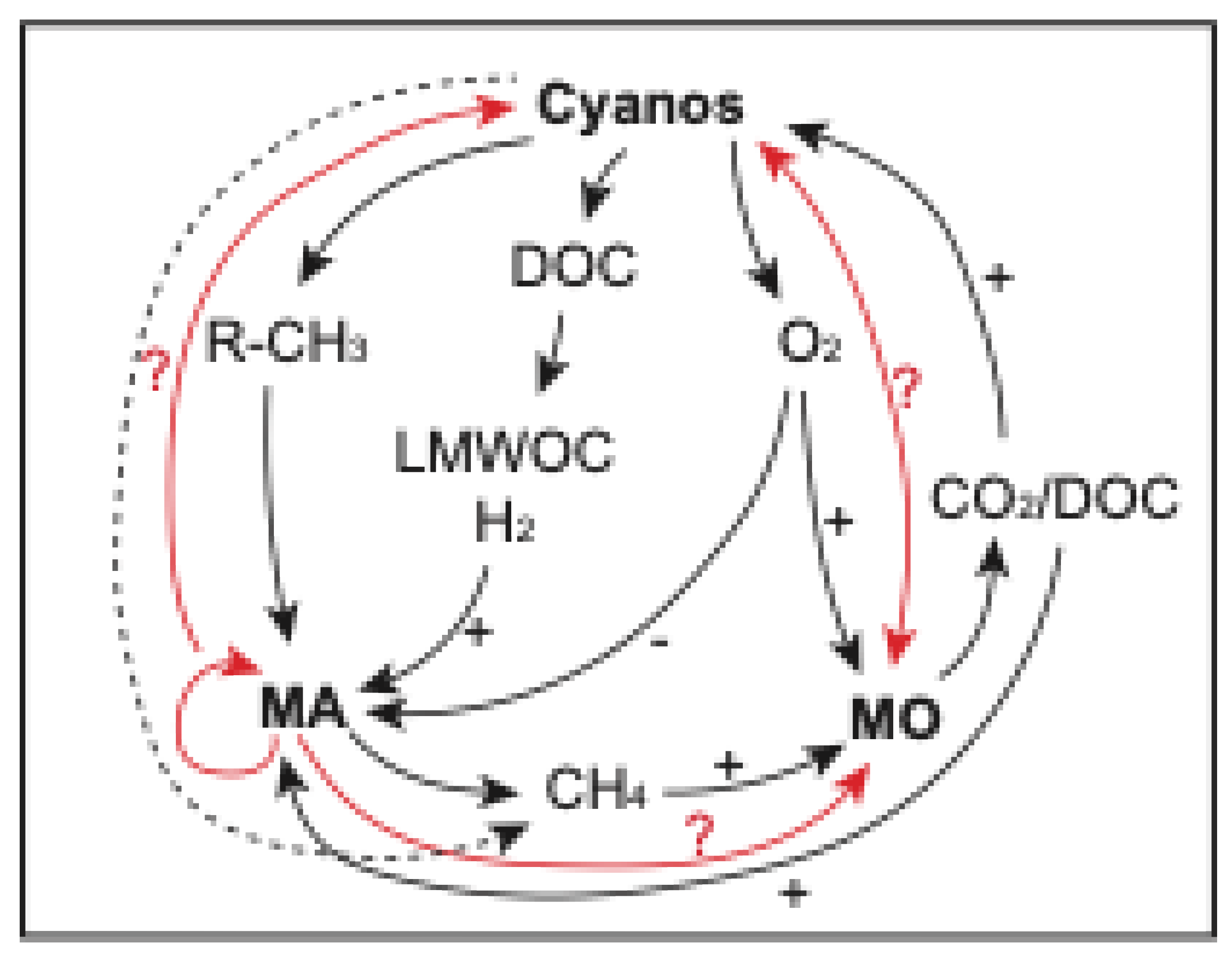
Figure 4.
Summary of the various roles of methanogens in biogeochemical processes, including positive and negative feedback, through geologic time that impacted climate. The major processes methanogens participate in include: i) consumption of organic carbon (e.g., XCH3, or methylated compounds), producing biomass and CH4 and fractionating stable isotopes of carbon; ii) consumption of H2/CO2 (e.g., from Earth’s early atmosphere), potentially changing the redox balance; iii) precipitation of minerals (e.g., carbonates); iv) use and fractionation of trace elements (e.g., Ni) that stimulated their growth and metabolic activity; v) the release of CH4 to the environment, stimulating methane-consuming microbes; vi) the release of CH4 to the atmosphere, acting as a greenhouse gas, contributing to organic haze formation that yield organic nitrogen, thereby regulating climate and consequently, play a role in beginning and/or end of mass extinctions and the ecological recovery after these icehouse events. Note that the strong isotopic fractionation of carbon and nickel by methanogens can be used to reconstruct early ecosystems on Earth. The production of methane and fractionation of carbon is a potential biosignature that can be used in the search for life beyond our planet.
Figure 4.
Summary of the various roles of methanogens in biogeochemical processes, including positive and negative feedback, through geologic time that impacted climate. The major processes methanogens participate in include: i) consumption of organic carbon (e.g., XCH3, or methylated compounds), producing biomass and CH4 and fractionating stable isotopes of carbon; ii) consumption of H2/CO2 (e.g., from Earth’s early atmosphere), potentially changing the redox balance; iii) precipitation of minerals (e.g., carbonates); iv) use and fractionation of trace elements (e.g., Ni) that stimulated their growth and metabolic activity; v) the release of CH4 to the environment, stimulating methane-consuming microbes; vi) the release of CH4 to the atmosphere, acting as a greenhouse gas, contributing to organic haze formation that yield organic nitrogen, thereby regulating climate and consequently, play a role in beginning and/or end of mass extinctions and the ecological recovery after these icehouse events. Note that the strong isotopic fractionation of carbon and nickel by methanogens can be used to reconstruct early ecosystems on Earth. The production of methane and fractionation of carbon is a potential biosignature that can be used in the search for life beyond our planet.
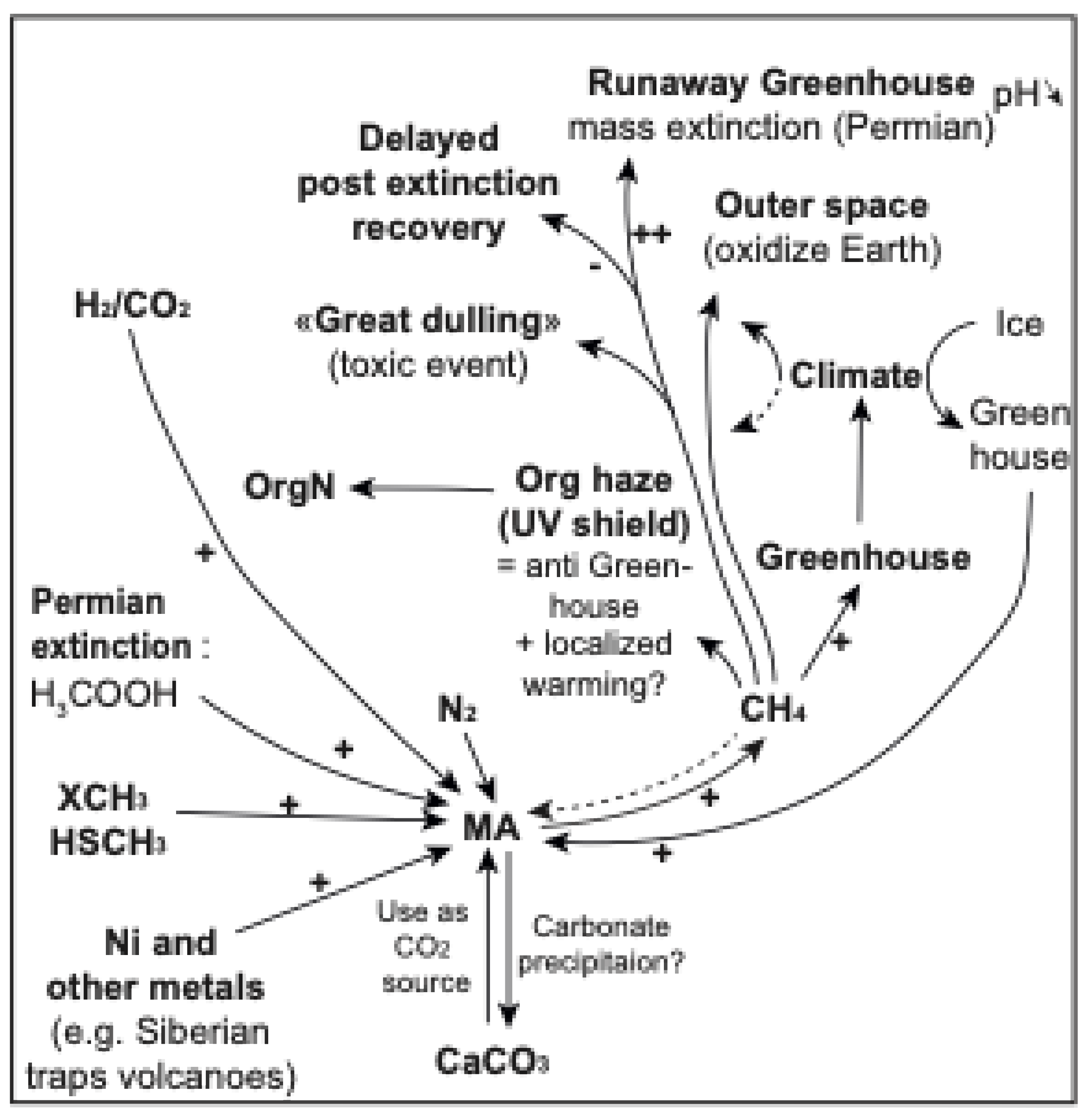
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



