Submitted:
22 December 2025
Posted:
26 December 2025
You are already at the latest version
Abstract
Microbial fermentation is a key biotechnological tool for producing bioactive metabo-lites such as alkaloids, carotenoids, essential oils, and phenolic compounds, among others, with applications in human health, agriculture, and food industries. This re-view comprehensively reviews recent information on the synthesis of valuable com-pounds and enzymes through fermentation processes. Here, we discuss the advantages of the different types of fermentation, such as submerged and solid-state fermentation, in optimizing metabolite production by bacteria, fungi, and yeast. The role of microbial metabolism, enzymatic activity, and fermentation conditions in enhancing the bioa-vailability and functionality of these compounds is discussed. Integrating fermentation with emerging biotechnologies, including metabolic engineering, further enhances yields and specificity. The potential of microbial-derived bioactive compounds in de-veloping functional foods, pharmaceuticals, and eco-friendly agricultural solutions positions fermentation as a pivotal strategy for future biotechnological advancements. Therefore, microbial fermentation is a sustainable tool to obtain high-quality metabo-lites from different sources that can be used in agriculture, animal, and human health.
Keywords:
fermentation
; microorganism
; bioactive compounds
; one-health
1. Introduction
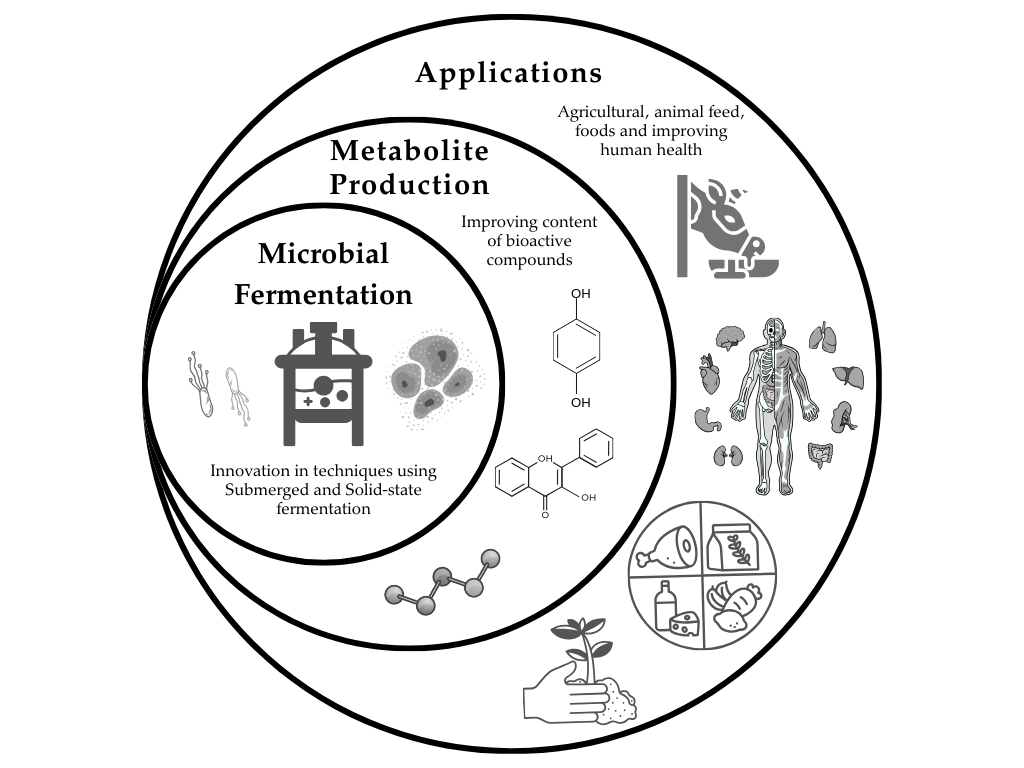
Bioactive compounds are found in plants, fruits, vegetables, and cereals. These compounds, such as alkaloids, antibiotics, mycotoxins, phenolics, pigments, polysaccharides, and vitamins, are among the compounds that microalgae can synthesize or microorganisms can excrete during the fermentation process. These metabolites are linked with promoting benefits for human health, such as antioxidants, anti-tumor, anti-diabetes, cardiovascular protection, and hypoglycemic activities. Therefore, metabolites are a target due to the promising ingredients of different nutraceutical, pharmaceutical, and cosmetic products [1,2,3,4]. Additionally, plant growth factors, biostimulation, elicitation, and the inhibition of phytopathogenic diseases on plants have been studied, as well as the potential activities of metabolites such as polysaccharides, enzymes, and microorganisms in submerged and solid-state fermentations for agricultural uses [5,6,7]. Recently, it has been studied the effect of fermentation of food waste to enhance the diet to improve the health and growth of different animals; in this sense, the use of microorganisms as probiotics on animal models, as well as the metabolites obtained by fermentation of food waste as ingredients on animal feed are demonstrated an enhance of health and growth of goats, pig, among other animals; therefore, the use of fermentation of food waste could be a promising alternative to commercial animal products, contributing to a circular economy by enabling waste materials to be reintegrated into the production cycle and continue generating value [8,9,10,11]. This study highlights the importance of microbial fermentation as an innovative and sustainable alternative to obtain metabolites related to human health, agricultural, and animal uses.
2. Literature Research Methodology
To collect relevant literature on the subject, we performed literature research using the databases Scopus, Web of Science, and Google Scholar using the following keywords: fermentation, “bioactive compounds”, “one-health”, “metabolite production”, “solid-state fermentation”, enzymes, polyphenols, carotenoids, essential oils, polysaccharides, applications, lactic acid bacteria, Lactobacillus, Saccharomyces, Aspergillus, “functional foods”, pharmaceuticals. We mainly included literature from 2015-2025 and only reports published in English. At the end, we included 296 articles in this review paper (Figure 1).
3. Fermentation as a Strategy to Produce Metabolites of Interest
Fermentation is a preservation technique used since ancient times to extend the shelf life of food and improve the bioaccessibility and bioavailability of nutrients from different sources [12]. This process occurs through microorganisms such as bacteria, fungi, and yeast that transform substrates into new ingredients such as textures, fragrances, flavors, enzymes, and bioactive compounds[13,14]. The fermented products are affected by several factors, such as temperature, pH, aeration, agitation, and microorganism-to-substrate ratio; these parameters are monitored and studied to control the fermentation process [14,15]. The fermentation process is divided into submerged fermentation (SF) and solid-state fermentation (SFF), which depend on the substrate and nutrient medium based on the cultivation of microorganisms[14]. These fermentation types have some advantages, which are related to the growth of microorganisms, specificity, and adaptation to fermentation conditions (Figure 2) [16,17]. In this sense, Candida, Galactomyces, Kluyveromyces, Monascus, Pediococcus, Streptococcus, Torulaspora, and Weisella microorganisms have been studied on submerged fermentation; while Lactococcus, Lentimus, Mucor, Thamnidium, Trichoderma, and Yarrowia microorganisms have been used on solid-state fermentation [17,18,19,20,21,22].
3.1. Submerged fermentation
Submerged fermentation is performed in a liquid substrate where microorganisms can adapt to high humidity. This fermentation method is one of the most studied because it is considered technologically easy to scale up to large-scale processes. Less extreme and more economical conditions are required to grow, diffuse, and separate microorganisms and compounds [17,19]. Submerged fermentation can be carried out in various ways, including batch, fed-batch, and continuous fermentation. The fermentation method used needs to be oriented to cover the characteristics of the microorganism and nutrients studied, as well as the infrastructure and separation steps required to obtain product compounds [16,17,23]. This type of fermentation is used in other processes. Also, it has been reported that a wide range of raw materials has been studied to extract different compounds such as flavonoids, antioxidants and antifungal peptides, dietary fibers, volatile compounds, among others of industrial importance [17]. However, submerged fermentation requires a longer fermentation time due to its lower performance. Also, high effluent volumes are produced, and waste needs to be stabilized before being dropped into the environment [24]. The most studied microorganisms on submerged fermentation are bacteria (Bacillus, Bifidobacterium, Lactobacillus, Pediococcus, Streptococcus), yeast (saccharomyces), and some fungi (Cordyceps, Galactomyces, kluyveromyces, Monascus, Picchia, Rhizopus, Torulaspora, Weisella) that can growth on high humidity and transform the carbohydrates into organic acids, alcohols, other compounds, and new ingredients with health benefits, as well as enzymes that are secreted into the liquid medium [17,25].
3.2. Solid-state fermentation
Solid-state fermentation is a technique in which microorganisms grow in the absence, partially or completely, of water and metabolize the nutrients of the solid substrate to transform carbohydrates into other compounds [1]. In recent years, various substrates have been studied for solid-state fermentation. Among these, agricultural biomass is considered an alternative for obtaining metabolites of agricultural, cosmetic, and pharmacological importance using fungi, yeast, and certain bacteria. Also, this solid-state fermentation has been considered cheaper in contrast to submerged fermentation due to processing conditions, simpler equipment, lower sterilization cost, and reduced downstream processing, among others [1,6,20,22]. Likewise, a higher yield of metabolites in shorter times has been obtained under solid-state fermentation compared to submerged fermentation, which could be due to specific growth conditions of microorganisms under stresses conditions, as well as uncompetitive inhibition with other microorganisms to consume the nutrients of substrates [6,12,26].
4. Microbial Metabolism and Fermentation Mechanism
4.1. Metabolic changes in microorganisms
In general, to be considered a “fermentation”, microorganisms must produce alcohol, commonly via pyruvate, converted into ethanol and carbon dioxide; this process is frequently related to yeasts, including Saccharomyces and non-Saccharomyces yeasts [27,28]. Nonetheless, the fermentation process is not exclusive to yeasts; there are several fermentative bacteria, such as Lactic Acid Bacteria, Acetic Acid Bacteria, and native microbiota present in raw materials, which participate in the fermentation of several food and beverage industries [3,29,30,31,32]. In this regard, bacterial fermentation holds special interest due to the increasing demand for fermented produce, which serves as a potential source of rich biomass and bioactive compounds, offering consumer benefits [33,34,35].
During fermentation, the process requires that fermentative bacteria start their metabolic activity, which commonly represents entering a new environment, and bacteria must adapt their genetic content in response to the surrounding conditions [36]. In this regard, there are some factors involved in the process, including temperature, pH, as well as the natural composition of the raw material, which induces the metabolic response of microorganisms [37], dealing with changes in the metabolic pathways, such as amino acids production; Liu, et al. [30] studied the changes in bacterial communities in fermented foods such as Chinese watermelon-soybean paste, revealing that microbial abundance may vary due to fermentation conditions, dealing with the diversity and richness of metabolites in the fermented food, and showing a positive correlation of Bacillus species with the production of all free amino acids and dipeptides, and the presence of 11 major metabolic pathways for the metabolism of tyrosine, arginine and proline, glycerophospholipid, sphingolipid, cofactor biosynthesis, alanine, aspartate, and glutamate; amino sugar and nucleotide sugar metabolism; as well as phenylalanine, tyrosine, and tryptophan biosynthesis. Temperature and time are important factors during of fermentation, which may affect the results and composition of the fermented matrix. Zhao, et al. [38] reported that lactic acid bacteria cells increased their levels at low salt fermentation and low temperature, contributing to acid production during the first 45 days of the process, as well as the increase of l-glutamine, l-ornithine, β-D-fructose, d-arabinose, d-gluconic acid, glycine, l-proline, and guanine as a product of the enrichment of 28 metabolic pathways, mainly for Glycine, serine, and threonine metabolism, and aminoacyl-tRNA biosynthesis. As mentioned before, raw material composition plays an important role in the results of fermentation because it represents the “food” for the bacteria; in this regard, it is crucial to understand the presence of essential nutrients, such as carbon sources, which are not necessarily a high diversity contributes to a better response because there are some microorganisms that show a better grow and productive response in the presence of a sole carbon source and specific amino acids [39].
An example of fermentative bacteria is Lactiplantibacillus plantarum, which is considered a promising probiotic bacterium, given its potential for fermentation and high adaptability, improving the quality and properties of fermented foods [2]. The versatility of this microorganism resides in its genetic content; Lactiplantibacillus plantarum has a genome size of around 3.1 Mbp, which presents diverse genes associated with diverse metabolic functions, such as carbohydrate transport and metabolism, amino acid transport, and metabolism, energy production and conversion, and other genes that contributes to the potential for vitamin production, biotin, alpha- and beta-glucosidase, related to probiotic activity [40,41,42]. The versatility of fermentative bacteria such as Lactobacillus plantarum has been evaluated, reporting its ability to redirect around 44% of its gene expression according to each specific growth habitat, particularly by regulation of metabolic activity, including the metabolism of carbohydrates, pyruvate, energy production, and conversion, transport, and metabolism of amino acids, nucleotides, lipids, and inorganic ions. Pyruvate metabolism is essential for fermentative bacteria; in this regard, growth conditions may deal with marked differences in the gene regulation for its utilization, upregulating the expression of the pyruvate dehydrogenase complex, as well as the stress response against antimicrobial compounds produced during fermentation, demonstrating the metabolic versatility of these kinds of microorganisms and their potential for fermentation processes [43,44].

| Microorganisms | Enzymes | Substrate | Application | Role |
|---|---|---|---|---|
| A. Niger, P. notatum, B. amyloliquefaciencs, B. stearothermophilus. B. licheniformis, S. cerevisiae, A. awamori, Rhizopus oryzae, Gluconacetobacter, Acetobacter xylinus, Komagataeibacter xylinus, Fusarium sp., B. brevis | α-Amylases | Grapes, rice, cereals | Beverages | Juice treatment, low-calorie beer, Clarification of fruit juice. |
| Cellulases | Degrade plant cell walls. In wine production is used to increase yield and quality. | |||
| Esterase | Enhancement of flavor and fragrance in fruit juice. | |||
| Glucoamylases | Convert the starch into maltose and fermentable sugar. Use in sake and light beer production. | |||
| Laccases | Modification of color appearance and wine stabilization. | |||
| Pectinases | Clarification of fruit juice | |||
| Proteases | Improves fermentation of beer. | |||
| Xylanases | Release arabinoxylans and lower oligo saccharides, reducing the muddy appearance and viscosity of the beer. | |||
| Lactobacillus Acidophilus, B. mesentericus, S. boulardii, S. ellipsoideus, P. Ostreatus, S. diastaticus, L. brevis, L. fermentum, R. oryzae, R. oligosporus, L. plantarum, A. oryzae, A. niger | α-Amylases | Wheat, maize, sorghum, millet, rice, soybean | Cereals and legumes | Increase the total starch in barley and peas. Decrease amylose content in rice. Decrease carbohydrates in sorghum. |
| Arabinoxylanases | Decrease insoluble fiber. | |||
| Cellulases | Low crude fiber in Pearl millet Fiber decreases in sorghum and yellow maize. |
|||
| Lipases | Decreased fat in mung beans, pigeon peas, red beans, soy and wild vigna species, maize, and rice. Increase in fat in pearl millet. |
|||
| Polyphenol oxidases | Decrease tannins | |||
| Proteases | Increase in some essential amino acids and IVPD in maize and sorghum. Increase in protein accumulation in pearl millet. |
|||
| Tannases | Decreases tannins in beans, oats, and sorghum. | |||
| Xylanases | Decrease fiber in sorghum | |||
|
Lactobacillus bulgaricus, Lactococcus lactis, L. acidophilus, L. cremoris, L. casei, L. paracasei, L. thermophilus, L. kefiri, L. caucasicus, Penicillium camemberti, P. roqueforti, Acetobacter lovaniensis, Kluyveromyces lactis, S. cerevisiae, A. Niger, A orzyae, B. subtilis, S. boydii, B. subtilis |
Catalases | Milk and casein | Dairy Products | Removes H2O2 and eliminate the effect of volatile sulfhydryl that is responsible for the cooked/off-flavor in ultra-pasteurized milk. |
| Lactases | Lactose hydrolysis, whey hydrolysis. | |||
| Lipases | Cheese flavor. | |||
| Proteases | Protein hydrolysis, milk clotting, low-allergenic infant food formulation, enhanced digestibility and utilization, flavor improvement in milk and cheese. | |||
| Bacillus megaterium, Bacillus subtilis | Amine oxidases | Fish proteins | Aquatic products | Inhibit biogenic amine accumulation, which is responsible for decreasing the quality and safety of fish-fermented products |
| Decarboxylases | Degrade saturated fatty acids, which influence flavor. | |||
| Esterases | Enhance favorable texture (hardness, gumminess, springiness, and chewiness), flavor, and aroma properties. | |||
| Glucosidases | Release aromatic compounds from flavorless precursors. | |||
| Lipases | Contribute to the development of flavor in the products due to the degradation of lipids to free fatty acids. | |||
| Lyases | Produce flavor substances | |||
| Proteases | Can develop different fermentation outcomes, some of which improve the product, while others may not help and might be detrimental. Generating peptides with antioxidant and antibacterial activities. | |||
| Transferases | Produce flavor substances | |||
| L. sakei, L. curvatus, L. plantarum, Leuconostoc carnosum, Leuconostoc gelidium, B. licheniformis, E. faecalis, E. hirae, E. durans, Bacillus subtilis, L. divergens, L. carnis, E. cecorum, B. lentus, T. longibrachiatum, A. niger, A. oryzae, S. aureus | Papain | Meat proteins | Meat | Myofibrillar degradation of as well as collagen proteins helps to tenderize meat |
| Polyphenol oxidases | Improve textural characteristics of meat products. | |||
| Proteases | Tendering tough buffalo and sheep meat. | |||
| Transglutaminases | Modify the texture of meat and meat products |
4.2. Enzymes involved in fermentation
Enzymes are considered environmentally friendly biological catalysts, revolutionizing food preparation methods. Their use is widespread across various global food industries, including dairy, brewing, meat, baking, beverages, cereals, legumes, oils, and fats (Table 1). Incorporating enzymes and microorganisms in food processing is a well-established traditional practice [18,22]. Microbial enzymes have been utilized in various industries, including fuel[49], human health[50], and soil[51], and are crucial to the food industry due to their greater stability than plant and animal enzymes. They can be produced cost-efficiently through fermentation processes, requiring less time and space [47].
Their high consistency also allows for easy process modifications and optimizations that improve vitamins, essential amino acids, proteins, food appearance, flavors, enhanced aroma, and reduced anti-nutrients [22].
4.3. Enzymes used in the food industry
4.3.1. Amylase and glucoamylase
Amylases are generally categorized into two main types based on their enzymatic class: hydrolases (Enzyme Commission, EC 3) and transferases (EC 2). Within the hydrolase class, they are further divided into two primary groups: endoamylases and exoamylases [52].
Glucoamylases, also called saccharifying enzymes, are within the group of exoamylases due to their ability to hydrolyze α-1,4 glycosidic bonds from the non-reducing starch, malto-oligosaccharides, and related substrates, releasing β-D-glucose [53]. They convert the starch to maltose and fermentable sugars. These enzymes are also produced from Saccharomyces cerevisiae during the fermentation with glucose to obtain ethanol. Also, glucoamylases are essential in brewing’s sake and soy sauce and creating light beer. These enzymes break down dextrins, transforming them into fermentable sugars, which result in beer with lower calorie content and reduced alcohol levels [47,53].
On the other hand, α-amylase (EC 3.2.1.1) is part of the family of starch-degrading endoamylase enzymes, which catalyzes the hydrolysis of α-1,4 glycosidic linkages in polysaccharides, producing short-chain dextrins. This enzyme is commonly synthesized by various organisms, including Archaea, fungi, bacteria, and animals [47]. Amylases have diverse applications in food processing, such as brewing, livestock feed, baking, fruit juice production, starch syrups, and starch liquefaction. Specifically, α-amylase breaks starches of flour into fermentable sugars, which are then utilized by yeast during bread production to enhance the bread’s taste and quality. This enzyme also helps slow staling when incorporated into bread-making [21].
4.3.2. Proteases
Proteases are crucial in commercial and industrial applications, and catalyse the hydrolysis of peptide bonds of proteins to peptides [54]. They represent a large and diverse group of hydrolytic enzymes classified by their site of action, enzyme active site structure, and specific reaction mechanisms [55]. Proteases are categorized depending on their action sites and along polypeptide chains into exopeptidases and endopeptidases. Exopeptidases target chain ends, while endopeptidases cleave internal bonds. Endopeptidases are further grouped into six types according to the catalytic residue in their active site: serine endopeptidases, aspartic endopeptidases, cysteine endopeptidases, metalloendopeptidases, glutamic endopeptidases, and threonine endopeptidases [47,56].
Proteases are produced by diverse organisms, including plants, Archaea, fungi, bacteria, and animals, and are widely used in various food industry sectors, such as brewing, dairy, baking, food processing, and animal feed [21]. These microbial enzymes perform diverse biochemical, physiological, and regulatory roles and have been essential in producing traditional fermented foods [54]. The characteristic flavor of fermented products is highly related to proteolysis due to hydrolysates generated precursor compounds, which are related to some important flavor substances during fermented products [57]. Proteases are produced by diverse organisms, including plants, Archaea, fungi, bacteria, and animals. They are widely used in various food industry sectors, such as brewing, dairy, baking, food processing, and animal feed [21]. These microbial enzymes perform diverse biochemical, physiological, and regulatory roles and have been essential in producing traditional fermented foods [54]. The characteristic flavor of fermented products is highly related to proteolysis due to hydrolysates generated precursor compounds, which are related to some important flavor substances during fermented products [57].
4.3.3 Lipases
Lipases (EC 3.1.1.3) are a class of enzymes that catalyze the hydrolysis of long-chain triglycerides. They are widely distributed in animals, plants, and microorganisms. However, microbial lipases find an interesting role in biotechnology, as many of them are stable over a wide range of pH, at elevated temperatures, and in organic solvents. They signify the most important group of biocatalysts for industrial applications [58,59]. These enzymes can modify the properties of lipids by altering the location of fatty acid chains in the glyceride and replacing one or more fatty acids with new ones [58,60].
In the food industry, they are applied to enhance flavors in dairy products, particularly by hydrolyzing milk fats to produce desirable cheese flavors. In bread dough, they hydrolyze triglycerides into diglycerides, monoglycerides, and fatty acids, which improve softness, volume, over-fermentation tolerance, and shelf life [60].
4.3.4. Catalase
Catalase (EC 1.11.1.6) is an oxidoreductase enzyme that reduces reactive oxygen species, particularly hydrogen peroxide, produced during aerobic respiration, thereby serving as an antioxidant and protecting cells from oxidative stress [61]. It is widely used in the food industry, often with enzymes like glucose oxidase for food preservation. In milk processing, catalase removes peroxide; in egg whites, it eliminates glucose; and in baking and food packaging, it helps prevent oxidation and reduces perishability. Its application in cheese production is limited [47].
4.3.5. Cellulases and Xylanases
Cellulases are hydrolytic enzymes that catalyze the cleavage of β-1,4 glycosidic bonds in glucose chains, transforming cellulose into cello-oligosaccharides and glucose via chemical or enzymatic hydrolysis. This category includes endoglucanases (EC 3.2.1.4), exoglucanases (EC 3.2.1.91), and β-glucosidases (EC 3.2.1.21) [62]. Similarly, microorganisms produce xylanases to break down xylans, a key hemicellulose component. Three main enzymes—endoxylanases, exoxylanases, and β-xylosidases—work together to degrade the xylan structure. Specifically, endoxylanases (EC 3.2.1.8) cleave β-1,4 bonds within xylan, while exoxylanases (EC 3.2.1.37) act on the non-reducing ends, producing xylooligosaccharides [63].
In the baking industry, cellulase and xylanase, along with amylases, lipases, and oxidases, enhance dough softness and reduce stickiness by breaking down insoluble cellulose and arabinoxylans in wheat flour into simple sugars, thereby improving flavor and texture in products like bread and cookies [47,64].
These enzymes are also widely used in brewing and fruit juice production, often combined with pectinases and amylases. During brewing, grains such as barley, wheat, and corn undergo mashing and malting to activate enzymes that ferment starch into alcohol. In both brewing and juice processing, cellulases and xylanases, along with pectinases, aid in breaking down cell walls and releasing sugars, nutrients, pigments, and aromatic compounds. These macerating enzymes are essential for producing fruit juices and wine [18,62,63].
4.3.6. Lactases (β-Galactosidase)
Lactase, a hydrolase enzyme known as β-galactosidase, is sourced from plants, animals, and microorganisms. Microbial β-galactosidases are widely used in food technology to hydrolyze lactose in milk and its by-products, particularly beneficial for individuals with lactose intolerance. These enzymes break down β-galactopyranosides (like lactose) and produce galactooligosaccharides (GOS), which offer prebiotic health benefits. Additionally, β-galactosidase aids in producing lactose-based sweeteners from the high-lactose effluents of cheese production [47,65].
4.3.7. Tannases
Tannase or tannin acylhydrolase (EC 3.1.1.20) is an extracellular inducible enzyme that catalyzes the hydrolysis of some tannins and gallic acid esters [66]. Tannases are widely applied in the food, brewing, and pharmaceutical industries. They are found across animal, plant, and microbial sources, with microbial tannases preferred for industrial applications. These enzymes hydrolyze tannins by breaking ester and depside bonds, releasing glucose and gallic acid [67]. In the food industry, tannases improve instant tea by enhancing compound extractability and cold-water solubility. Additionally, tannases prevent haze and undesirable phenolic compounds in beer and wine, improving quality. The enzyme also reduces haze and bitterness in fruit juices. In animal feed, tannases degrade anti-nutritional tannins in agro-industrial by-products, enhancing feed quality [66,68].
4.3.8. Esterases
Esterases are hydrolase enzymes that catalyze the formation and breakdown of ester bonds. Their regio-selectivity, stereospecificity, and stability in organic solvents make them highly valuable for various industrial applications [69]. They are essential in food and alcoholic beverages, mainly for modifying oils and fats in fruit juices and creating flavors and fragrances. A key group, feruloyl esterases, breaks the ester bond between ferulic acid and polysaccharides in plant cell walls. By hydrolyzing lignocellulosic biomass, these enzymes are also crucial for waste management [47].
5. Bioactive Compounds Obtained from Microbial Fermentation
5.1. Carotenoids
Carotenoids are a group of compounds related to the pigments yellow, orange, red, and purple, mainly found in fruits, vegetables, and plants [70]. Carotenoids have a 40-carbon structure with double conjugated bonds and a polyene chain end that varies depending on whether or not an oxygen atom is present; in this sense, carotenoids are classified into two groups, carotenes and xanthophylls [70]. Carotene comprises hydrocarbons, carotenoids such as α-carotene, β-carotene, and lycopene.[70,71]. Xanthophylls are carotenoids with an oxygen atom on the end polyene chain that constitutes a hydroxyl, carbonyl, or aldehyde molecule. Some examples of xanthophylls are lutein, astaxanthin, and fucoxanthin [70]. These compounds have been reported to have a high antioxidant activity that can protect the organisms from oxidative stress caused by biotic and abiotic factors; this effect could be associated with their structure of double conjugated bonds. In this sense, it has been reported that carotenoids are related to potential health effects such as UV-protective, antiproliferative, antidiabetic, anti-inflammatory, and antiatherosclerosis responses.
Carotenoids are a group of compounds related to the pigments yellow, orange, red, and purple, mainly found in fruits, vegetables, and plants [70]. Carotenoids have a 40-carbon structure with double conjugated bonds and a polyene chain end that varies depending on whether or not an oxygen atom; in this sense, carotenoids are classified into two groups, carotenes and xanthophylls [70]. Carotene comprises hydrocarbons, carotenoids such as α-carotene, β-carotene, and lycopene.[70,71]. Xanthophylls are carotenoids characterized by an oxygen atom on the end polyene chain that constitutes a hydroxyl, carbonyl, or aldehyde molecule. Some examples of xanthophylls are lutein, astaxanthin, and fucoxanthin [70].
These compounds have been reported to have a high antioxidant activity that can protect the organisms from oxidative stress caused by biotic and abiotic factors; this effect could be associated with their structure of double conjugated bonds. In this sense, it has been reported that carotenoids are related to potential health effects such as UV-protective, antiproliferative, antidiabetic, anti-inflammatory, and antiatherosclerosis responses [72,73]; therefore, carotenoids are compounds widely used in different industries such as medicine, food, and in pharmacology [74].
Microorganisms have gained popularity as an alternative source to plants to obtain multiple bioactive compounds simultaneously, using cheaper raw materials as carbon sources to reduce the cost of biorefinery [75]. Recently, various microorganisms have been studied for their carotenoid production; among these, photosynthetic bacteria, fungi, marine archaea, and yeast have been utilized to obtain lycopene, ketolases, α-carotene, β-carotene, and other carotenoids through biotechnology strategies [70,74,76,77]. The use of microorganisms as carotenoid producers could be due to the controlled cultivation, high efficiency, and target compounds synthesis, and reduced production period by metabolism modifications [78]. Biotechnology strategies such as metabolic engineering tools for the manipulation of biosynthetic pathways for carotenoid production coupled with the fermentation process followed by separation and purification methods have been widely studied to enhance the production and extraction of carotenoids from microorganisms, where the principal factor studied are pH of culture medium, temperature, process time, moisture content, aeration rate, carbon and nitrogen sources, light, carbon/nitrogen ratio, sonication, chemical supplements, among other parameters [74,79,80]. The use of microorganisms as factories for producing carotenoids has been widely studied due to their diverse color tones, the ability to manipulate them, and low equipment requirements [81]. However, some disadvantages, such as the high production costs, have limited their industrial application. As a result, other biotechnological approaches are being explored to enhance carotenoid production and reduce costs using various microorganisms [75,80,82,83,84].
In this sense, it has been reported that fermentation technology has been used in conjunction with biotechnological manipulations such as metabolic engineering to produce carotenoids with high production yields, efficiency, moderate cost of production, and environmentally friendly in contrast to chemical synthesis and plant or animal sources extraction due to the fast-growing strains and downstream techniques to extract the target products [74,80]. In this sense, microorganisms such as Blakeslea trispora, Chryseobacterium artocarpi CECT 8497, Flavobacterium sp. Cells, Serratia marcescens, and Phaffia rhodozyma, Rhodosporidium toruloides, Sporobolomyces, Sporidioblous, and Xanthophyllomyces Dendrorhous has been used to obtain anthraquinones, astaxanthin, β-carotene, canthaxanthin, flexirubin, lutein, and zeaxanthin by optimized fermentation process [74,77,81,85,86], while other microorganisms as Escherichia Coli, Saccharomyces cerevisiae, Pichia pastoris, and Yarrowia lipolytica has been modified throughout biotechnology strategies as well as different extraction process to obtain carotenoids with high yields (Figure 3) [15,80,84].
On the other hand, the effect of fermentation on carotenoids from different food sources and by-products has been studied [82]. In this sense, orange, sweet potato, carrot, tomato, marigold red paper, goji berries, and marine by-products, among other sources, have been studied to obtain astaxanthin, carotenes (α-carotene and β-carotene), lutein, lycopene, and zeaxanthin [75,82,83,87]. Likewise, fermentation factors such as matrix sources, fermentation conditions, as well as the microorganism studied, have been reported to enhance or maintain the carotenoid content; also, the use of some microorganisms could favor the carotenoid extraction and bioavailability by the production of enzymes that separate the carotenoids from the food matrix, and facilitate the extraction with downstream strategies [82,83].
5.2. Essential oils
Essential oils are bioactive compounds derived from plants composed mainly of low molecular weight volatile compounds such as terpenes and hydrocarbons [88,89]. These substances have attracted the attention of pharmaceutic and food industries due to their therapeutic [90], anticancer [91], antimicrobial [92], antioxidant [93], and food safety properties [94]. Also, essential oils contain compounds with strong odors, and the fragrance industry has also been interested in the obtention of these materials. The production of essential oils can be carried out by direct plant extraction, plant cell cultures, and microbial fermentation [95]. Specifically, the application of microbial fermentation can be classified into biotransformation (fungi or yeast transform compounds based on certain substrates), methods that use waste residues (food waste), and metabolic pathways followed by genetically modified microorganisms (Figure 4).
Besides these methods, there are a few recent reports regarding the de novo synthesis of essential oils by microorganisms. For example, fungal cultures (13 strains belonging to Aspergillus sp., Ceratocystis sp., and Neurospora sp.) produced interesting volatile compounds that can be found in essential oils from plants such as citral and β-citronellol [96], which have been studied for their antifungal [97] and vasorelaxant properties in rats [98].[98] On the other hand, Sen, et al. [99] reported that the interaction of agarwood (Aquilaria malaccensis) callus with the associated fungus Fusarium produced by fermentation led to a significant accumulation of terpenes such as geranyl isovalerate and tetrapentacontane,1,54, dibromo-. Also, the infection of juvenile A. malaccensis plants coupled with associated Fusarium registered the presence of sesquiterpenes such as agarospirol, γ-eudesmol, (−)-aristolene, which were exclusive of the fermentation with this fungus. In this sense, the main issue with the de novo synthesis of essential oil compounds by microorganisms is the accumulation of the obtained metabolites that can result in toxic environments for the organisms. This is the case of geraniol and citronellol production by the fungi genus Ceratocystis, whose removal procedures consisted of integrated bioprocesses that had to be applied to eliminate contaminated substances [100].
Regarding the use of waste residues for obtaining essential oils (or their compounds) through microbial fermentation, [101] proposed a solid-state fermentation process that employs fungi, bacteria, and yeast strains (Aspergillus, Trichoderma, Bacillus, and Saccharomyces) to produce volatile compounds from basil leaf and stem waste. Several substances of importance for human health and the food industry were identified, such as ϒ-Bisabolene, isoprenyl cinnamate, diacetyl, and ethyl isovalerate. These compounds are present in natural essential oils from plants, and they have been studied for antioxidant [102], antimicrobial [103], antifungal [104], and food preservation [105] properties. Another therapeutic compound in essential oils is D-limonene, which can be obtained via the fermentation of olive mill waste by Rhizopus oryzae fungus and Candida tropicalis yeast [106]. The quantification of this substance revealed that R. oryzae produced a higher amount of D-limonene than C. tropicalis in controlled conditions: 87.73 µg/kg and 11.95 µg/kg, respectively. Therefore, agro-industrial waste residues are suitable for producing volatile compounds with health benefits. For example, rice bran oil waste is rich in ferulic acid, a vanillin precursor. Zheng, et al. [107] reported a consecutive bioconversion process from ferulic acid of rice bran oil residue fermented by Aspergillus niger to vanillic acid; and then, this broth was fermented by Pycnoporus cinnabarinus to obtain vanillin. A maximum concentration of this compound (2.8 g/L) with a molar yield of 61.9% at 72 h was registered.
In the case of biotransformation as a method for obtaining essential oils from microbial fermentation, this process is defined as the induction of the microbe to follow a specific metabolic pathway from an introduced precursor [108]. For that, geranic acid has been synthesized from geraniol by Rhodococcus sp., a group of bacteria isolated from soil. This synthesis was controlled by reaching a maximum conversion (54.6%) of geranic acid after 96 h of incubation at 30 °C as the optimal temperature for the reaction [109]. Besides these bacteria, yeasts have been reported to produce volatile compounds via bioconversion. Oda, et al. [110] developed a metabolic pathway by Hansenula and Pichia yeasts using acetyl-CoA to synthesize citronellol, finding that the accumulation of citronellol in the incubating system could intoxicate yeasts; therefore, sodium acetate was added to control the production rate of citronellyl acetate for all strains. In this sense, yeasts can be affected by their metabolites in the production of the fragrance compound 2-phenylethanol (2-PE) from phenylalanine by Saccharomyces cerevisiae, in which the use of hydrophobic polymethylmethacrylate (PMMA) microspheres has been reported to physically remove the accumulation of phenylethanol [111]. This interesting volatile compound has been studied for treating psychiatric disorders such as depression [112].
Based on the above-mentioned, yeasts have been broadly studied to produce compounds of essential oils. However, metabolic engineering has been applied to modify these microorganisms at the genetic level. An increase in geraniol concentration (from 43.19 mg/L to 523.96 mg/L) via synthesis performed by manipulating enzymes GES and Erg20WW in S. cerevisiae was reported [113]. Also, this compound's production was enhanced by modifying the site where the geraniol synthesis occurred in S. cerevisiae peroxisomes, which improved the geraniol titers by 80% [114]. Therefore, S. cerevisiae represents an appropriate “factory” for essential oils generation. Important substances such as limonene have been obtained by orthogonal engineering in which a metabolic pathway was created by genetic modification in this yeast, producing significant amounts of limonene (917.7 mg/L) in fed-batch fermentation [115]. In addition to reported experimental trials, Werner, et al. [116] informed through an in-silico study that modifying gene expression of central metabolic enzymes in S. cerevisiae can increase β-ionone yields up to 4-fold. Most microorganisms used for essential oil production belonged to the fungi kingdom; however, gene modification has been commonly applied in prokaryotic organisms. One of these microorganisms is E. coli, which has been altered to produce volatile compounds such as patchoulol [117], viridiflorol and amorphadiene [118], ionone [119], bisabolol [120], and others.
5.3. Phenolic compounds
Phenolic compounds are bioactive compounds obtained from plants with a benzene ring with one or more hydroxyl groups. These products can be found in various natural foods, such as cereals, vegetables, fruits, and beverages such as wine, coffee, cocoa, and tea [121,122]. Phenolic compounds, including flavonoids, tannins, lignans, and stilbenes, are abundant plant compounds synthesized via the shikimate pathway using L-phenylalanine and L-tyrosine as precursors [123]. These compounds are present in various foods and beverages, contributing to their health benefits and bioactivity. Cocoa and its derivatives are among the most studied food matrices due to their high polyphenol content. Bacterial fermentation of cocoa beans significantly increases the release of conjugated phenolic compounds, such as caffeoyl aspartic acid and p-coumaroyl tyrosine, within 24 hours of fermentation [124]. Using strains like Saccharomyces cerevisiae and Pichia kudriavzevii further enhances phenolic content, showcasing their potential for improving food bioactivity [125]. In microbial fermentation, phenolic acids undergo extensive biotransformation by the gut microbiota, producing smaller bioactive metabolites. Microbial fermentation processes significantly influence protocatechuic and vanillic acids derived from cocoa shells. During colonic fermentation, the gut microbiota transforms these compounds into metabolites such as benzoic, phenylpropanoid, and phenylacetic acids. Additionally, fermentation of cocoa by-products generates other phenolic compounds, such as caffeoyl aspartic acid, p-coumaroyl aspartic acid, clovamide, and p-coumaroyl tyrosine. These transformations are heavily influenced by the dietary fiber content in cocoa shells, which modulates the release and microbial accessibility of phenolic acids [126]. Similarly, coffee pulp fermentation enhances the production of phenolic metabolites such as phenylpropanoic acids and phenyl-γ-valerolactones, while reducing the overall concentration of polyphenols [127]. The fermentation process, driven by microbial activity, modulates these phenolic transformations and generates bioactive compounds like benzoic and phenylacetic acids. Additionally, dietary fiber and alkaloid content, such as caffeine and trigonelline, influence the bioavailability and release of phenolic compounds during fermentation [128].
Cereals naturally contain phenolic compounds predominantly in bound forms, which limits their bioavailability [129,130]. Specific processing techniques, such as microbial fermentation, are required to enhance the release of these antioxidant compounds and increase their free-form levels. Fermentation with ragi tape effectively enhances phenolic content in black glutinous rice. Over 72 hours, free phenolics increased by 49%, and free-conjugated phenolics peaked at 48 hours with an 8% rise. This process significantly improved antioxidant activity, as indicated by reduced IC50 values. Key phenolic compounds, including caffeic and ferulic acids, showed notable increases, highlighting the potential of ragi tape fermentation to boost the antioxidant properties of this rice variety [131]. Solid-state fermentation (SSF) of sorghum grain (SG) enhances sensory characteristics by modifying its polyphenol content. Various microbial strains (Lactiplantibacillus plantarum, Saccharomyces cerevisiae, Rhizopus oryzae, Aspergillus oryzae, and Neurospora sitophila) were used to assess their impact on polyphenols. After SSF, tannin and free phenolic contents were reduced by 56.36% and 23.48%, respectively. Cellulase played a crucial role in degrading tannins and phenolic compounds, while xylanase initially released flavonoids, although microbial consumption altered this effect over time. These findings highlight the potential of SSF to modify the polyphenolic profile and improve SG bioactivity and processing characteristics [132]. In a study on rye flour fermentation, the microbial community of germinated rye was enriched during lactic acid fermentation with increased terpenoid and phenolic compounds. Metabolomic analysis revealed notable changes in phenolic compound profiles, including the accumulation of bioactive polyphenols. Specifically, compounds such as ferulic acid, caffeic acid, and p-coumaric acid were identified post-fermentation, suggesting that sourdough fermentation can enhance the content of bioactive polyphenols, contributing to the health-promoting potential of fermented rye flour [133]. Studies consistently demonstrate that microbial fermentation significantly enhances the phenolic profile and antioxidant activity of cereals. While cereals naturally contain phenolics, a substantial portion exists in bound forms, limiting their bioavailability [134]. Fermentation releases these bound phenolics, as observed in black glutinous rice with ragi tape, sorghum, and sourdough-fermented rye, where free phenolics like caffeic, ferulic, and p-coumaric acids increased, boosting antioxidant activity. This process is attributed to microbial enzymes like feruloyl esterases, which hydrolyze bonds linking phenolic acids to cell walls [135].
Microbial fermentation is a crucial bioprocess for preserving fruits and vegetables. This biological method reduces the risk of contamination by producing antimicrobial compounds like organic acids, ethanol, and bacteriocins. Beyond preservation, fermentation enhances these foods' nutritional value and creates new and desirable tastes and textures[136]. Citrus fruits, including their peels, are abundant sources of bioactive compounds such as phenolic acids, flavanones, and polymethoxylated flavones, alongside carotenoids and ascorbic acid [137,138]. Microbial fermentation of citrus peels, including orange, lemon, and grapefruit, was conducted using Lactobacillus plantarum and Lactobacillus acidophilus as fermentation agents. This process significantly enhanced the phenolic profile by transforming bound phenolics into more bioavailable forms. Key phenolic acids such as ferulic acid, caffeic acid, and p-coumaric acid were notably increased. Additionally, flavonoids like naringenin and hesperidin showed considerable elevation, improving antioxidant activity [139]. In the case of orange peel, SSF significantly increased the release of phenolic compounds. The fermentation process primarily resulted in the formation of flavonoid aglycones, such as naringenin, hesperetin, and nobiletin, replacing the glycoside hesperidin found in unfermented peels. Additionally, the fermented peel showed enhanced antioxidant activity [140].
In chili peppers, microbial fermentation increases both the quantity and diversity of phenolic compounds [141]. Lactic acid bacteria secrete enzymes like amylase, β-glucosidase, and phenolic acid decarboxylase, facilitating the release of bound polyphenols and enhancing antioxidant activity [142]. Caffeic acid, a key phenolic, rises during early fermentation stages but degrades into compounds like vinyl catechol and ethyl catechol as fermentation progresses [143]. Ferulic acid, another significant phenolic found in chili pepper and carrots, can be metabolized into vanillic acid and protocatechuic acid, highlighting the transformative potential of fermentation on polyphenolic profiles [144]. In chili peppers, microbial fermentation increases both the quantity and diversity of phenolic compounds [141]. Lactic acid bacteria secrete enzymes like amylase, β-glucosidase, and phenolic acid decarboxylase, facilitating the release of bound polyphenols and enhancing antioxidant activity [142]. Caffeic acid, a key phenolic, rises during early fermentation stages but degrades into compounds like vinyl catechol and ethyl catechol as fermentation progresses [143]. Ferulic acid, another significant phenolic found in chili pepper and carrots, can be metabolized into vanillic acid and protocatechuic acid, highlighting the transformative potential of fermentation on polyphenolic profiles [144].
Onion by-products are abundant in fiber and bioactive compounds, offering a valuable source for developing various bioproducts, including polyphenols [145]. A study assessed the potential of onion skins for polyphenol release through fermentation by different microorganisms, including bacterial and yeast strains. Results showed that fermentation with specific strains like Lactiplantibacillus plantarum and Saccharomyces cerevisiae increased the content of bioactive compounds, particularly quercetin aglycone, by up to 60% in yellow onion skins [146].
Wild herbs and plants are rich in phenolic compounds, particularly flavonoids [147]. Fermentation with microorganisms such as bacteria is known to increase the bioactive compounds in plants, particularly those with antioxidant and antibacterial properties [148]. For example, a study investigated the fermentation of Dendrobium officinale using brewer's yeast (Saccharomyces cerevisiae) to enhance its bioactive compounds, particularly phenolics. By optimizing fermentation conditions through single-factor and orthogonal experiments, the researchers produced a fermentation broth enriched in these valuable compounds. Notably, the fermentation process yielded four key phenolic acids: gallic acid, protocatechuic acid, catechol, and pentosidine B sesil [149]. Achillea millefolium L. (yarrow) and Origanum majorana L. were studied as sources of phenolic-rich extracts, focusing on their impact on human gut microbiota and microbial metabolism. SSF enhanced the bioavailability of phenolic compounds like rosmarinic acid and caffeoylquinic acids. The microbial metabolism yielded significant metabolites, including phloroglucinol and 3,4-dimethoxyphenylacetic acid [150]. Beverages such as wine, beer, coffee, and tea are substantial sources of polyphenols, the most abundant antioxidants in the human diet [151]. Fermentation is an emerging method used to enhance the phenolic content in beverages, thereby improving their antioxidant properties. For instance, microbial fermentation of tea extracts using Trichoderma reesei, Aspergillus niger, and lactic acid bacteria (LAB) transformed pivotal bioactive compounds. Galloylated catechins were hydrolyzed, and organic acids were decarboxylated, increasing phenolic bioavailability. Additionally, alcohols and ketones accumulated, enhancing the aroma profile [152]. This highlights fermentation as a promising tool for improving both the phenolic composition and sensory qualities of tea beverages. Blueberry pomace, a by-product of juice processing, has shown significant potential as a matrix for phenolic enrichment through SSF. In a recent study, various fungal and LAB strains, including Aspergillus niger (AN), Lactobacillus acidophilus (LA), and Lactobacillus plantarum (LP), were utilized to enhance its polyphenol profile. Fermentation increased the content of key phenolic acids such as gallic acid, caffeic acid, and chlorogenic acid, as well as flavonoids like quercetin. Notably, anthocyanin levels decreased overall, except in pomace fermented by AN, where anthocyanidins showed an upward trend. Enhanced antioxidant activities were observed, with ABTS, DPPH, and FRAP radical scavenging capacities rising by 33.56%, 59.89%, and 87.82%, respectively. Additionally, simulated gastrointestinal digestion revealed improved bioaccessibility of polyphenols, underscoring SSF as an effective method for value-added utilization of blueberry pomace in functional food development [153]. Albino bilberry (Vaccinium myrtillus L.) juice [154] and Chinese rice wine [155] exemplify the impact of fermentation on enhancing phenolic composition and flavor complexity. In albino bilberry juice, fermentation with non-Saccharomyces yeasts resulted in increased phenolic acids (21.8%–42.5%), flavonols (26.8%–47.2%), and flavan-3-ols (4.9%–74.5%), with novel flavonols synthesized during the process. Hanseniaspora uvarum yielded the highest phenolic enhancements, showcasing yeast metabolism's role in antioxidant enrichment. Similarly, in Chinese rice wine brewed from five raw materials, liquid-state fermentation and grain liquefaction influenced flavor compounds and microbial diversity. Buckwheat-fermented wine stood out with the highest ester (27.39 mg/L), amino acid (1.47 mg/mL), and phenolic acid (904.29 mg/L) contents, contributing to its complex sensory profile characterized by honey, floral, and umami notes. Metagenomic sequencing revealed variations in microbial communities, with dominant genera including Saccharomyces, Aspergillus, and Bacillus. Together, these studies emphasize the importance of fermentation parameters and raw material selection in tailoring phenolic profiles, microbial ecology, and sensory attributes in functional beverages.
Fermentation can generate the synthesis of phenolic compounds, further increasing their content and antioxidant capacity [156]. However, the lack of standardized methodologies for analyzing phenolic compounds, the use of various microorganisms, and different extraction methods present opportunities to further improve the understanding and application of microbial fermentation in enhancing phenolic profiles [157,158,159]. Standardizing analytical techniques, selecting the appropriate microorganisms, and extraction methods are essential to maximizing the benefits of fermentation for food quality improvement.
Table 2.
Summary of phenolic compounds enhanced through fermentation in recent studies.
| Food Matrix | Fermentation Treatment | Phenolic Compounds Increased | |
| Cocoa shells | In vitro colonic fermentation | Caffeoyl aspartic acid, p-coumaroyl aspartic acid, clovamide, p-coumaroyl tyrosine | [126] |
| Coffee pulp | In vitro colonic fermentation | Phenylpropanoic acids, phenyl-γ-valerolactones | [127] |
| Black glutinous rice | Ragi tape fermentation | Caffeic acid, ferulic acid |
[131] |
| Sorghum grain | SSF with various microbial strains | Tannins, free phenolics (caffeic, ferulic, p-coumaric acids) | [132] |
| Rye flour | Lactic acid fermentation | Ferulic acid, caffeic acid, p-coumaric acid | [133] |
| Citrus peels (orange, lemon, grapefruit) | Lactobacillus plantarum, Lactobacillus acidophilus | Ferulic acid, caffeic acid, p-coumaric acid, naringenin, hesperidin and nobiletin | [139,140] |
| Chili peppers | Lactic acid bacteria | Caffeic acid, ferulic acid | [141] |
| Onion skins | Lactiplantibacillus plantarum, Saccharomyces cerevisiae | Quercetin aglycone | [146] |
| Dendrobium officinale | Saccharomyces cerevisiae | Gallic acid, protocatechuic acid, catechol, pentosidine B sesil | [149] |
| Achillea millefolium L. and Origanum majorana L. | SSF with various microbial strains | Rosmarinic acid, caffeoylquinic acids, phloroglucinol and 3,4-dimethoxyphenylacetic acid | [150] |
| Tea extracts | Trichoderma reesei, Aspergillus niger, Lactic acid bacteria | Galloylated catechins, organic acids | [152] |
| Blueberry pomace | Various fungal and LAB strains (Aspergillus niger, Lactobacillus acidophilus, Lactobacillus plantarum) | Gallic acid, caffeic acid, chlorogenic acid, quercetin | [153] |
| Albino bilberry juice | Non-Saccharomyces yeasts | Phenolic acids, flavonols, flavan-3-ols | [154] |
| Chinese rice wine | Saccharomyces, Aspergillus, Bacillus | Phenolic acids | [155] |
1 Summary of microbial fermentation treatments applied to various food matrices. The table highlights the phenolic compounds that were enhanced as result of fermentation, showcasing the potential of microbial processes in improving the bioavailability and antioxidant properties of phenolic compounds in food.
5.4. Polysaccharides
The fermentation process induces the production of several types of polysaccharides by microorganisms. Some of these compounds possess properties which are beneficial for human health, such as antioxidant, antitumor, and anti-inflammatory activities [160,161]. Also, they are considered biocompatible and biodegradable materials with high yield and reproducible production [160].
In this sense, a high amount of research can be found in the literature reporting that lactic acid fermentation by bacterial strains (Lactobacillus, Leuconostoc, among others) produces exopolysaccharides. Exopolysaccharides are a specific type of extracellular biopolymers synthesized by bacteria and some fungi, having several health and physicochemical properties [162,163]. For that, Table 3 summarizes different exopolysaccharides with biological activities produced by microbial fermentation.
One of these substances is dextran, a long-chain glycopolymer reported as a functional molecule for medical purposes such as plasma volume expander, wall material for encapsulation, and an alternative to tomography markers [164]. Schmid, et al. [165] stated that the properties of dextran are dependent on its molecular weight (i.e. length of the polymer chain). In that report, dextran polymers with high molecular weights (1.09 x 108 – 1.86 x 108 Da) produced by Liquorilactobacillus hordei exhibited different rheological properties, indicating that the dextran with the longest chain had the best capacity to form films and gels.
Similarly, Esmaeilnejad-Moghadam, et al. [164] found that dextran synthesized by Leuconostoc mesenteroides in milk permeate culture media possessed the lowest molecular weight and increased solubility as compared to dextran produced in broth medium. Besides its techno-functional properties, an appropriate antioxidant activity has been detected (by ABTS and DPPH analyses) in dextran obtained by the fermentation with Leuconostoc pseudomesenteroides isolated from the Juçara palm tree [166]. However, some bacteria cannot produce exopolysaccharides in fermentation conditions. As an example, Lactobacillus fermentum was investigated during the fermentation of longan pulp, finding that the polysaccharides of the fruit changed in composition and chemical structure because of the action of the bacteria. The polysaccharides from the fermented pulp were composed mainly of arabinose (49%), galactose (23%), glucose (14%), and other monosaccharides (~14%), in which immunomodulatory and prebiotic activities were detected [167].
Also, the prebiotic activity has been attributed to fructooligosaccharides that can be synthesized by the enzymes inulosucrase and endoinulinase present in bacteria such as Leuconostoc citreum and Aspergillus niger, respectively. In this study, sucrose was more effective than inulin as a substrate for the chain reaction to produce these prebiotic compounds [168]. On the other hand, the production of functional polysaccharides by fungi has been commonly studied in fermentation conditions involving Tremella spp. The basidiospore fermentation by Tremella aurantialba in tofu wastewater has been proposed as a method for obtaining adequate yields of polysaccharides (~15.02 g/L) whose composition was based on monosaccharides such as glucose and mannose [169]. Similarly, Ge, et al. [170] examined Tremella fuciformis in fermentation conditions, finding that this fungus produced macromolecular polysaccharides (yield ≈ 9.0 g/L) which were composed of xylose, mannose and galactose residues and having antioxidant properties determined by the scavenging capacity of superoxide anions and hydroxyl radicals analyses. Besides the antioxidant activity, neuroprotective [171], immunomodulatory [172], and antitumor [173] activities have been reported for polysaccharides produced by Tremella spp.
Regarding the fermentation by yeasts, Chen, et al. [174] addressed the biological properties of polysaccharides obtained by the fermentation of a flower Dendrobium officinale carried out by Saccharomyces cerevisiae and Wickerhamomyces anomalous. Both yeasts produced four polysaccharides (comprised of mannose and glucose, having the following ratios 3.31:1, 5.56:1, 2.40:1, and 3.29:1) that were isolated, possessing an enhanced antioxidant property; however, the anti-inflammatory activity was inadequate for the proposed experimental conditions.
Also, the production of endopolysaccharides by yeast has been reported for the fermentation of biodiesel-derived crude glycerol, in which yeast strains belonging to Debaryomyces sp., Naganishia uzbekistanensis, Rhodotorula sp., and Yarrowia lipolytica generated biopolymers with potential for pharmaceutical applications [175].
Furthermore, bacteria can produce polysaccharides of interest alongside other microorganisms. In this sense, the fermentation of okara (residue from soybean) by lactic acid bacteria Lactobacillus bulgaricus and fungi Neurospora crassa produced polysaccharides (composed mainly of galacturonic acid, galactose, and arabinose) with sugar blood regulation, glucose adsorption delaying and prebiotic properties [176]. In another study, yeast Saccharomyces cerevisiae and bacteria Bacillus subtilis enhanced the polysaccharides obtained by the fermentation of wheat bran, indicating that the isolated products (composed mostly of galactose, xylose, and galacturonic acid) possessed suitable antioxidant properties (measured by DPPH test) [177]. Therefore, the combination of bacteria with other microorganisms, such as fungi and yeasts can result in the enhancement of polysaccharides produced by fermentation of different sources.
Table 3.
Exopolysaccharides synthesized by microbial fermentation and their health properties
| Microorganism | Specie | Exopolysaccharide | Health properties | Ref. |
|---|---|---|---|---|
| Bacteria | Acetobacter xylinum | Levan | Antioxidant, anti-inflammatory | [178] |
| Bacteria | Lactobacillus plantarum | Glucose and galactose residues | Antioxidant | [179] |
| Bacteria | Paenibacillus polymyxa | Heteroglycan formed by (1→4) and (1→6) hexose residues | Antioxidant, immunomodulatory, mitogenic, allergenic, anti-inflammatory | [180] |
| Bacteria | Escherichia coli (modified with a Leuconostoc citreum gene) | Alternan | Encapsulation capability | [181] |
| Bacteria | Bacillus sp. isolated from fermented pickles | Glucose and galactose residues | Antioxidant | [182] |
| Fungi | Polyporus umbellatus | 3 polysaccharides composed by mannose, galactose and glucose (molar ratios: 43.6:2.5:1.0; 17.7:3.1:1.0 and 4.6:2.6:1.0) | Antioxidant, immunological, cellular aging delaying, DNA damage protecting | [183] |
6. Applications of Bioactive compounds obtained from microbial fermentation
6.1. Animal feed
The use of fermented feed is gaining attention as a sustainable approach to improving livestock health while addressing environmental concerns. Fermented feed is a biologically modified product resulting from microbial fermentation that transforms feed components into microbial proteins, bioactive peptides, amino acids, and beneficial probiotics [184]. This process enhances the digestibility of complex carbohydrates, proteins, and fibers, improving nutrient availability and reducing the impact of anti-nutritional factors such as phytates and protease inhibitors [185,186]. Fermented feed is gaining attention as a sustainable approach to improving livestock health while addressing environmental concerns. Fermented feed is a biologically modified product resulting from microbial fermentation that transforms feed components into microbial proteins, bioactive peptides, amino acids, and beneficial probiotics [184]. This process enhances the digestibility of complex carbohydrates, proteins, and fibers, improving nutrient availability and reducing the impact of anti-nutritional factors such as phytates and protease inhibitors [185,186].
Gut microbiota is crucial in livestock health and productivity [187]. Fermented feed has been shown to positively modulate gut microbiota composition by increasing the abundance and diversity of beneficial microorganisms. This promotes optimal nutrient absorption, gastrointestinal health, and immune function while reducing disease incidence and oxidative stress [188,189]. This modulation of gut microbiota through fermented feed is increasingly recognized as a sustainable alternative to antibiotics, improving animal welfare and production efficiency [190].[190]. It also mitigates oxidative stress by protecting the gastrointestinal tract, aiding recovery, and boosting stress resilience, supporting efficient and sustainable livestock production [188].
The growing interest in fermented feed stems from its demonstrated benefits across various livestock species. For example, in Bamei piglets, the inclusion of Lactobacillus plantarum and Bacillus subtilis-fermented feed led to higher weight gain, better feed efficiency, and improved immunity, as evidenced by elevated levels of immunoglobulins and reduced inflammatory markers. The fermented feed also positively impacted the gut microbiota, enhancing the diversity of beneficial bacteria such as Lactobacillus and Prevotellaceae, which are crucial for maintaining intestinal health and optimizing nutrient absorption [191]. Similarly, fermenting the plant-based fraction of their feed with Lactobacillus and Bacillus subtilis significantly improved growth performance, meat quality, and nutrient utilization in broiler chickens. Additional benefits included reduced cholesterol content and better feed conversion ratios, showcasing its potential to enhance poultry production efficiency and product quality [184]. Similar benefits have been observed in aquaculture, where fermented feeds improve nutrient digestibility, enhance immune responses, and promote growth. These feeds support metabolic and digestive enzyme activity by providing proteins and micronutrients while reducing anti-nutritional factors, contributing to healthier and more sustainable aquaculture production [192]. Furthermore, the microbial fermentation of Psophocarpus tetragonolobus (winged bean tubers) has shown promising results in ruminant diets, enhancing their nutritional value, improving feed degradability, and optimizing volatile fatty acid profiles, with increased propionic acid levels and energy efficiency. This highlights the potential of fermented tuber pellets as a sustainable alternative to traditional feed ingredients like corn meal, supporting both animal health and productivity [193]. In poultry, a study on Xuefeng black-bone chickens demonstrated that the combination of microbial fermented feed and ginseng polysaccharides using (Saccharomyces cerevisiae, Bacillus subtilis, Lactobacillus plantarum, and Enterococcus faecium) further enhanced growth performance, feed efficiency, and immune function [194]. These studies demonstrate the broad applicability of microbial fermentation in improving livestock health and productivity across species such as swine, poultry, ruminants, and fish. By supporting sustainable and efficient production systems, fermented feeds represent a vital tool in advancing eco-friendly livestock management.
6.2. Agricultural use
Microbial products offer a sustainable alternative to traditional agricultural chemicals and fertilizers by enhancing crop yields and improving soil health. Companies increasingly use microorganisms as biocontrol agents and biofertilizers through carrier-based inoculants, which enrich the soil by producing essential nutrients through their metabolic activities. Building upon these benefits, recent studies have highlighted the critical role of microbial metabolites in promoting plant growth and enhancing disease resistance. This supports plant growth by enhancing nutrient availability, such as nitrogen, phosphorus, and potassium, while improving soil properties and boosting beneficial bacteria. These applications not only decrease reliance on chemical inputs but also contribute to soil biodiversity and health, which is crucial for maintaining long-term agricultural sustainability [195,196,197].
Recent studies have highlighted the pivotal role of microbial metabolites in plant growth promotion and disease resistance. Plant growth-promoting rhizobacteria produce metabolites such as auxins, cytokinins, and gibberellins, which enhance nutrient uptake, stimulate plant growth, and improve disease resistance by inhibiting pathogens and inducing systemic resistance [198,199]. Plants further influence their microbiomes by secreting specific compounds that recruit beneficial microbes to suppress pathogens and enhance plant immunity [200]. These insights demonstrate the potential of microbial metabolites to support resilient cropping systems while reducing dependency on chemical inputs.
In addition to promoting disease resistance, microbial metabolites also play a key role in helping plants cope with environmental stresses such as drought, salinity, and temperature extremes by enhancing plant physiology, boosting antioxidants, and improving water retention, ultimately increasing productivity under stress [201,202]. Exopolysaccharides secreted by soil bacteria improve soil aggregation and water retention, promoting plant hydration during drought by enhancing water infiltration [203]. Under salinity stress, osmoprotectants like trehalose and proline produced by microbes accumulate in plant cells to maintain osmotic balance and protect cellular components from salt-induced damage [204]. Microbial compounds also enhance plant antioxidant systems, scavenging reactive oxygen species to prevent oxidative damage and preserve cellular integrity [205]. In addition, some metabolites induce the expression of stress-responsive genes, enabling better adaptation and tolerance to abiotic stresses [206].
Given their benefits, the adoption of microbial metabolites in agriculture represents a sustainable shift from traditional chemical fertilizers and pesticides, promoting eco-friendly farming practices. Microbial inoculants derived from beneficial bacteria and fungi, combined with renewable feedstocks in microbial fermentation, enhance soil fertility, support plant health, and reduce reliance on chemical inputs, contributing to sustainability by minimizing dependence on non-renewable resources [207]. Furthermore, microbial biotechnology bolsters soil biodiversity and health, with managed soil microbiomes significantly improving crop productivity and resilience [208].
Recent advancements in biotechnology, including the development of engineered microbes, have enhanced the production of microbial metabolites, providing sustainable alternatives to traditional fertilizers and pesticides by efficiently converting renewable feedstocks into biofertilizers and biopesticides [209]. These biotechnological advancements not only enhance microbial metabolite production but also significantly contribute to improving agricultural efficiency. By optimizing fermentation conditions and selecting suitable microorganisms, the yield and effectiveness of these metabolites have been enhanced. For example, engineered microbes are capable of converting renewable feedstocks into high-value biofertilizers and biopesticides, which support eco-friendly agricultural practices [210]. Additionally, the use of genetic engineering techniques has led to the development of tailored microbial strains that produce specific metabolites at higher concentrations, improving agricultural efficiency [211]. Microorganisms significantly important in agriculture have long been recognized as an effective and eco-friendly alternative in modern farming, reducing the dependence on synthetic fertilizers and pesticides [212]. Furthermore, biotechnological advancements in microbial applications support sustainable agriculture by minimizing reliance on non-renewable resources, as they utilize renewable feedstocks in fermentation processes. As these technological advances continue to evolve, they further drive the adoption of microbial-based practices, ultimately ensuring a more sustainable and productive agricultural system [213].
6.3 Human health
The fermentations stand out as a promising alternative for improving health due to its content of bioactive compounds, which are linked to the metabolic and biotransformation activities carried out by microorganisms [214]. The transformation of the natural components of the raw material during fermentation results in the production of enzymes that cause the transformation of complex compounds into simple biomolecules, giving rise to compounds with biological activity beneficial to human health (Figure 5) [136,215].
Dairy fermentations such as milk, yogurts, cheeses, creams, and ice creams contain probiotic bacteria, and some meats, sausages, bread, and cereal products [216]. The necessary amount of these viable microorganisms in a food to exert health benefits varies between 10⁸ and 10⁹ colony-forming units (CFU) per day. Their effectiveness depends on the type of microorganism and the physiological conditions of the consumer [217]. The mechanisms of action associated with the health effects of these microorganisms include competitive inhibition of pathogenic bacteria proliferation (through alterations in pH levels and a reduction in oxygen availability) and non-competitive inhibition via the production of bacteriocins [218]; synthesis of essential micronutrients (vitamins, amino acids and enzymes) enhance the bioavailability of compounds [219]; stimulation of the host immune system by promoting the production of interleukin-10 (IL-10) and immunoglobulin A (IgA) antibodies [220]. They also play a crucial role in protecting intestinal epithelial cells against inflammation and associated disorders by regulating the production of antibodies, lymphocytes, interleukins, cytokines, and chemokines [221]. Additionally, probiotic microorganisms present in fermented foods have been reported to produce the enzymes β-galactosidase and lactase, which help combat lactose intolerance by converting glucose and galactose into short-chain fatty acids [222].
On the other hand, the fermentation of whey protein has gained significant attention in recent years, as it enables the enzymatic hydrolysis of peptides that can act as bioactive compounds. This process is made possible by the activity of proteases, which generate low molecular weight peptides (˂10 kDa) with specific compositions that allow them to enter the cell nucleus and interact directly with DNA-associated proteins regulating gene expression [223,224]. These bioactive peptides have been linked to the positive regulation of cell proliferation in human cell cultures and the inhibition of growth in various types of cancer cells. Additionally, they have demonstrated immune system-like functions, such as promoting lymphocyte proliferation, antibody production, and cytokine regulation [225]. Furthermore, they stimulate the phagocytic capacity of macrophages and inhibit the secretion of specific cytokines. Antidiabetic, antihypertensive, and xanthine oxidase inhibitor activities are also reported by these bioactive molecules [226].
Following this, kefir is a fermented lactic acid derivative with characteristics similar to yogurt. Its main components, such as lactic acid bacteria, organic acids, polysaccharides, and bioactive peptides, have significant health benefits [227].[227]. Several studies have demonstrated its antioxidant, anti-inflammatory, anti-hypertensive, anticancer, and antidiabetic properties. Additionally, kefir has been associated with therapeutic effects on bone health, the immune system, cognitive function, and the gut microbiota [228,229,230,231,232].
Continuing with lactic fermentations, but now in solid-state form, bioactive peptides from various cheese varieties have been shown to provide health benefits by acting as antioxidants, antihypertensives, and antidiabetics [233]. For example, in a study where individuals with hypertension consumed 30 g/day of Italian cheese (Grana Padano), a significant reduction in blood pressure (both systolic and diastolic) was observed after two months [234]. In another study, six different types of cheese were evaluated, including Gouda, which showed the best results in antioxidant activity, ECA inhibitory activity, and DPP-IV enzyme inhibition. The study concluded that consuming 10–20 g of Gouda cheese, as part of a balanced diet, could be sufficient to obtain health benefits [235]. Another important compound in this type of lactic fermentation is conjugated linoleic acid. Reports have shown an increase in these bioactive levels during cheese maturation, which could enhance its potential health benefits, including antidiabetic, anticancer, anti-atherosclerotic, and antihypertensive effects [236].
On the other hand, soy fermentation is an important source of bioactive peptides with significant health benefits. For example, peptides obtained from tempeh fermentation have demonstrated antioxidant, anti-inflammatory, and antihypertensive effects [237,238]. Saponins and phytosterols, which are bioactive compounds naturally found in soy, have gained interest due to their medicinal properties [239]; regarding to[239]; regarding saponins, the presence of these compounds in some fermented foods has been associated with obesity prevention, positive regulation of the immune system, antiviral effects, and antitumor properties [240]. At the same time, sterols can inhibit cholesterol absorption in the intestine, potentially reducing the risk of cardiovascular diseases [241]. Another important compound in soy is gamma-aminobutyric acid (GABA), which acts as a neurotransmitter in the central nervous system; its health benefits include lowering blood pressure, promoting relaxation, and improving mood [241]. Fermented food soy has been reported to be a rich source of GABA due to the production of enzymes during fermentation, which facilitates its transformation through the action of glutamic acid decarboxylase [241]. Soy is also naturally rich in vitamins, minerals, and fiber, which are crucial for human growth and proper metabolism [238]. In addition to this, another important aspect of soy is its high content of allergenic proteins and other allergens, such as Gly m 1, P28, and P34, which, during fermentation processes (particularly those involving Rhizopus spp., Aspergillus oryzae, and Bacillus subtilis), can reduce the allergenicity of these proteins, decreasing the symptoms associated with their reactivity [242]. Also, bioactive compounds such as phenolic acids, flavonoids, and isoflavones have been identified in soy-derived fermentations [243]. Some of these fermented products originate from Meju, a dry soybean block fermented with fungi and Bacillus sp., which has been attributed with anticancer potential due to the presence of trypsin inhibitors, isoflavones, vitamin E, and unsaturated fatty acids [244,245]. Additionally, Doenjang extracts have been linked to the activation of the enzyme glutathione S-transferase and the increased activity of natural killer cells [246].
It is important to mention that isoflavones are the main compounds responsible for the anticancer potential of fermented soy products, as they have demonstrated beneficial therapeutic effects in cell lines of various types of cancer, including stomach, colon, lymphoma, pancreas, prostate, breast, and neuroblastoma [245]. Furthermore, these compounds have been associated with cardiovascular health benefits, increasing HDL cholesterol levels while reducing LDL cholesterol and triglyceride levels [247]. On the other hand, the anti-inflammatory effects of soy fermentation have been widely studied. It has been found that consuming foods such as miso and soy sauce reduces serum levels of IL-6 and IL-18 and inflammatory markers, including high-sensitivity C-reactive protein (hs-CRP) [248]. Some studies have also linked isoflavones to neuroprotection, promoting neuronal regeneration and enhancing existing neuronal functions. Therefore, soy fermentations represent an important source of bioactive compounds, particularly genistein, with great potential in preventing various diseases [249].
Fermentation has also been used to develop natural cosmetics with potential benefits for skin health. In this regard, studies on Camellia sinensis var. Assamica has shown that the activity of enzymes produced during the fermentation process can promote the release of bioactive compounds with moisturizing properties (such as amino acids and sugars); these compounds may help improve skin hydration and support its barrier function [250]. Additionally, an increase in the content of compounds like gallic acid and epigallocatechin gallate (EGCG) has been observed in the extract of Camellia sinensis tea, which could enhance its antioxidant and anti-inflammatory properties, which may help protect the skin from damage caused by free radicals and UV radiation, delaying skin aging and contributing to cancer prevention [251].
On the other hand, solid-state fermentation of phenols has been reported to improve the nutritional quality and antioxidant properties of various legumes and cereals (rice, wheat bran, corn, oats, rye, and millet). This process involves enzymes such as amylases, proteases, and lipases, which hydrolyze polysaccharides, proteins, and lipids into products with lower toxicity, improved texture, flavor, and aroma, as well as a reduction in antinutritional compounds such as phytic acid, tannins, and gas-producing compounds [237].
According to several studies, a wide variety of phenolic compounds have also been identified in liquid fermentation, such as wine, including catechins, p-coumaric acid, resveratrol, rutin, quercetin, myricetin, anthocyanins, tannins, and flavan-3-ols, as well as several phenolic acids such as caffeic, ellagic, syringic, vanillic, and ferulic acids [252]. Additionally, the fermentation process of red wine leads to the formation of compounds like melatonin and hydroxytyrosol, which have been strongly associated with the numerous health benefits of this beverage [253,254]. The bioactive compounds in red wine have been extensively studied in both in vitro and in vivo investigations, demonstrating a wide range of health benefits, including antioxidant, antibacterial, anti-inflammatory, anticancer, and antidiabetic properties [255,256,257,258,259]. Other notable effects include antithrombotic, antidepressant, and neuroprotective activities, microbiota regulation, and anti-obesity effects, as they influence adipose tissue metabolism, hypocholesterolemia, and endothelial function [260,261,262,263].
Vinegar is a highly valued fermented product obtained from fruits such as grapes, apples, pomegranates, and cranberries through a fermentation process that can occur via two different pathways: alcoholic and acetic [264]. The microorganisms commonly used in this process belong to the Acetobacter and Komagataeibacter species, which have been associated with increased bioactive compounds such as polyphenols and organic acids [3]. Among these, acetic acid is considered the most important component of vinegar. In this context, apple cider vinegar has been shown to provide various health benefits, including improvements in cognitive, liver, and reproductive function, and therapeutic effects in diabetes [265,266,267,268,269]. It also exhibits antioxidant, antimicrobial, anti-inflammatory, and anti-obesity properties. Another significant effect studied is its ability to combat hypercholesterolemia by improving total cholesterol levels, triglycerides, LDL cholesterol, total cholesterol/HDL-C ratio, and LDL-C/HDL-C ratio [270]. These effects have been linked to bioactive compounds such as catechins, p-hydroxybenzoic acid, gallic acid, caffeic acid, p-coumaric acid, and chlorogenic acid [271].
Kombucha is a liquid fermented that is notable for its potential health benefits, including antioxidant, antitumor, hepatoprotective, and antidiabetic properties. It is prepared by fermenting black or green tea (derived from the Camellia sinensis plant) using a symbiotic culture of yeasts, acetic acid bacteria, and lactic acid bacteria [272,273]. This product contains bioactive compounds such as catechins, theaflavins, thearubigins, and vitamins B1, B6, B12, and C, which are associated with its reported health benefits [274,275]. In this context, in vivo studies have demonstrated the antidiabetic potential of kombucha, showing effects such as reduced blood glucose levels, increased plasma insulin, and modulation of gluconeogenic and glycolytic enzyme activity in experimentally induced diabetic rats [276]. Additionally, this beverage has been shown to inhibit the activity of enzymes like α-amylase and α-glucosidase, leading to a slower rate of blood glucose absorption [277]. The antiproliferative activity of kombucha has also been evaluated in various cancer cell lines, including MCF-7 (breast cancer), A549 (lung cancer), and HCT8, HCT 116, and CACO-2 (colon cancer) [272,278]. Furthermore, studies suggest it may exhibit selectivity toward cancer cells while sparing normal lung cells [279]. On the other hand, this type of fermentation has also been shown to reduce the levels of pro-inflammatory markers, including nitric oxide (NO), tumor necrosis factor (TNF), and interleukin-6 (IL-6) in macrophages activated by LPS. Additionally, it has been found to inhibit the enzyme 15-lipoxygenase (15-LOX), a key mediator in inflammatory processes [278,280].
Another important liquid fermentation is black tea, made by a unique microbial fermentation process involving some basic functional microorganisms of the genera Aspergillus, Bacillus, Candida, Cyberlindnera, Klebsiella, Lactobacillus, Penicillium, Rasamsonia, among others. Compounds such as alkaloids, polyphenols, polysaccharides, and volatile compounds have been identified in this beverage, where caffeine stands out as one of the most abundant alkaloids and catechin in the group of polyphenols [281,282]. Multiple in vivo studies have shown that black tea has health benefits, for example, by reducing blood glucose levels and lowering the risk of diabetes by up to 45% in people with regular consumption of black tea (2-3 g per day) [283]. It has also been associated with significant weight and fat loss, improving the lipid profile and reducing hyperlipidemia in humans [284]. It has also been shown to ameliorate chemically induced colitis in mice by modulating the gut microbiota, to decrease insulin resistance and chronic kidney disease in rats by modulating insulin signaling and increasing Nrf2 expression [285], and to exert a hepatoprotective effect by modulating hepatic oxidative stress, inflammatory response and gut microbiota dysfunction in mice [286], and that it possesses potent lipid-lowering activity in high-fat zebrafish model [287].[287]. Additionally, in some in vitro and in vivo studies, it showed potential to prevent the onset of cardio and neurodegenerative diseases associated with its interaction with proteins of the signaling pathways of these diseases, so this fermentation has potential for the prevention and treatment of chronic diseases [288,289].
5. Perspectives
The use of microorganisms as potential biofactories of bioactive compounds using biotechnological strategies such as fermentation, coupled with other techniques such as metabolic engineering tools, is on the rise due to advantages such as reduced use of chemicals and water, and ease of cultivation and separation of compounds. Also, bioactive compounds obtained from microorganisms are considered to have a natural origin, and the commercial market has become more popular in contrast to chemical synthesis [15,31,75,81]. Likewise, the bioactive compounds such as carotenoids, essential oils, phenolic compounds, and polysaccharides, among others, obtained through microbial fermentation have been used for nutraceutical enrichment of animal feed [290], as well as the production of protein-rich animal feed using different sources such as agro-industrial and food wastes [291]. In recent years, microbial fermentation has been explored as an alternative to using agrochemicals to protect crops and reduce plant diseases caused by bacteria, fungi, nematodes, and yeast. Biostimulants have been developed to enhance the growth, yield, and defense of various crops [292,293]. In human health, microbial fermentation has been studied for the extraction of bioactive compounds as an alternative to traditional chemical extraction for the enrichment of food or for use as a nutraceutical because these compounds have been linked to the proper functioning of the organism and the reduction of non-communicable diseases [294,295,296].
Despite the use of microorganisms to obtain bioactive compounds using biotechnology strategies, research needs to continue developing efficient processes to release and enhance the extraction of the bioactive compounds because fermentation parameters such as fermentation time, solid-to-liquid ratio, temperature, pH, microorganism used, carbon source, a downstream technique to separate the molecules of interest need to be optimized to reduce the high cost of the technologies, to guarantee an appropriate bioprocess to high yield, selectivity, and avoid microbial contamination during fermentation process [1,22,81,160].
6. Conclusions
Bioactive compounds, including alkaloids, phenolic compounds, pigments, vitamins, and others, are of interest in many industries for their potential as chemopreventive agents against many chronic diseases, such as cancer, cardiovascular diseases, and diabetes. They can also prevent aging-related diseases. These metabolites are mainly found in plants, and for their use, the extraction techniques often produce high quantities of toxic waste from solvents such as hexane, methanol, and acetonitrile, among others. Microbial fermentation is a sustainable tool for obtaining high-quality metabolites from various plant and animal matrices and producing them through biotechnology to achieve a higher yield of these compounds. These metabolites can be used in agriculture, as well as in animal and human health. The potential use of these metabolites nowadays relies on the biotechnological techniques implemented to produce each group of metabolites.
Author Contributions
Conceptualization, Supervision, Writing – Original draft, Writing – Review & Editing, E.P.G.-G., L.A.C.B.; Investigation, Writing – Original draft, V.E.Z.-S.; Writing – Original draft, E.G.A., M.S.C.M., J.A.M.F., A.H.A.G.; Writing – Review & Editing, J.B.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
V.E.Z.-S. would like to thank the Secretaría de Ciencia, Humanidades, Tecnología e Innovación (SECIHTI) for the doctoral scholarship.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Martins, S.; et al. Bioactive phenolic compounds: Production and extraction by solid-state fermentation. A review. Biotechnology Advances 2011, 29, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, B.; et al. The Impacts of Lactiplantibacillus plantarum on the Functional Properties of Fermented Foods: A Review of Current Knowledge. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Yassunaka Hata, N.N.; et al. Role of Acetic Acid Bacteria in Food and Beverages. Food Technol Biotechnol 2023, 61, 85–103. [Google Scholar] [CrossRef]
- Han, X.; et al. Differential analysis of metabolites from mulberry (Morus abla L. cv. Longsang 1) juices fermented with Pichia kudriavzevii and Weissella confusa alone or in combination. Journal of Future Foods 2025, 5, 183–192. [Google Scholar] [CrossRef]
- Orts, J.M.; et al. Polyurethane waste valorization: A Two-Phase process using Ozonization and Rhodococcus pyridinivorans fermentation for biofertilizer production. Bioresource Technology 2025, 416, 131814. [Google Scholar] [CrossRef]
- Solano Porras, R.C.; et al. Solid-state fermentation of green waste for the production of biostimulants to enhance lettuce (Lactuca sativa L.) cultivation under water stress: Closing the organic waste cycle. Chemosphere 2025, 370, 143919. [Google Scholar] [CrossRef]
- Wu, Y.; et al. A plant growth-promoting bacterium supports cadmium detoxification of rice by inducing phenylpropanoid and flavonoid biosynthesis. Journal of Hazardous Materials 2025, 484, 136795. [Google Scholar] [CrossRef] [PubMed]
- Fathy, E.-H.; et al. Effect of Using Yeast, Fibrolytic Enzymes and Their Mixture on In Vitro Ruminal Fermentation Characteristics. Egyptian Journal of Veterinary Sciences 2024, 56, 483–491. [Google Scholar] [CrossRef]
- Kaewwongsa, W.; et al. Effects of replacement soybean meal with fermented cassava pulp by Saccharomyces cerevisiae in DIETS on Rumen fermentation in growing goats. Scientific Reports 2024, 14, 28124. [Google Scholar] [CrossRef]
- Kamal, M.; et al. Effects of and on fermentation in the rumen of Hu sheep. Journal of the Science of Food and Agriculture 2025, 105, 498–506. [Google Scholar] [CrossRef]
- Pi, C.-C.; et al. Synergistic fermentation of Cordyceps militaris and herbal substrates boosts grower pig antioxidant and immune function. BMC Veterinary Research 2024, 20, 531. [Google Scholar] [CrossRef]
- Lee, H.Y.; et al. Comparison of primary and secondary metabolites and antioxidant activities by solid-state fermentation of Apios americana Medikus with different fungi. Food Chemistry 2024, 461, 140808. [Google Scholar] [CrossRef]
- Oluwafemi Ayodeji, A.; et al. Fermented Pulse-Based Food Products in Developing Nations as Functional Foods and Ingredients. In Functional Food; Maria Chavarri, H., Ed.; IntechOpen: Rijeka, 2017; p. Ch. 5. [Google Scholar]
- Zhang, J.; et al. Recent Developments in Fermented Cereals on Nutritional Constituents and Potential Health Benefits. Foods 2022, 11. [Google Scholar] [CrossRef]
- Wang, J.; et al. Strategies for the efficient biosynthesis of β-carotene through microbial fermentation. World Journal of Microbiology and Biotechnology 2024, 40, 160. [Google Scholar] [CrossRef]
- Lübeck, M.; Lübeck, P.S. Fungal Cell Factories for Efficient and Sustainable Production of Proteins and Peptides. Microorganisms 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Martí-Quijal, F.J.; et al. Obtaining Antioxidants and Natural Preservatives from Food By-Products through Fermentation: A Review. Fermentation 2021, 7. [Google Scholar] [CrossRef]
- Al-Maqtari, Q.A.; et al. Microbial enzymes produced by fermentation and their applications in the food industry-A review. International Journal of Agriculture Innovations and Research 2019, 8, 2319–1473. [Google Scholar]
- Chai, K.F.; et al. Bioactive peptides from food fermentation: A comprehensive review of their sources, bioactivities, applications, and future development. Comprehensive Reviews in Food Science and Food Safety 2020, 19, 3825–3885. [Google Scholar] [CrossRef]
- Feng, X.; et al. Effect of Solid-State Fermentation on Plant-Sourced Proteins: A Review. Food Reviews International 2023, 1–38. [Google Scholar] [CrossRef]
- Okpara, M.O. Microbial enzymes and their applications in food industry: a mini-review. Advances in Enzyme Research 2022, 10, 23–47. [Google Scholar] [CrossRef]
- Sharma, R.; et al. Microbial Fermentation and Its Role in Quality Improvement of Fermented Foods. Fermentation 2020, 6. [Google Scholar] [CrossRef]
- De Villa, R.; et al. Impact of solid-state fermentation on factors and mechanisms influencing the bioactive compounds of grains and processing by-products. Critical Reviews in Food Science and Nutrition 2023, 63, 5388–5413. [Google Scholar] [CrossRef] [PubMed]
- Doriya, K.; et al. Chapter Six - Solid-State Fermentation vs Submerged Fermentation for the Production of l-Asparaginase. In Advances in Food and Nutrition Research; Kim, S.-K., Toldrá, F., Eds.; Academic Press, 2016; Volume 78, pp. 115–135. [Google Scholar]
- Tang, Z.; et al. A Review on Fruit and Vegetable Fermented Beverage-Benefits of Microbes and Beneficial Effects. Food Reviews International 2023, 39, 4835–4872. [Google Scholar] [CrossRef]
- Dong, J.-W.; et al. Solid-state fermentation of Aspergillus sydowii G12, an approach to produce isocorydine. Process Biochemistry 2024, 147, 147–151. [Google Scholar] [CrossRef]
- Maicas, S. The Role of Yeasts in Fermentation Processes. Microorganisms 2020, 8. [Google Scholar] [CrossRef]
- Zhang, S.; et al. Exploring community succession and metabolic changes in corn gluten meal-bran mixed wastes during fermentation. Journal of Environmental Management 2024, 366, 121684. [Google Scholar] [CrossRef]
- Antolak, H.; et al. Kombucha Tea—A Double Power of Bioactive Compounds from Tea and Symbiotic Culture of Bacteria and Yeasts (SCOBY). Antioxidants 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; et al. Exploring the changes in microorganisms and metabolites during the fermentation process of traditional Chinese watermelon–soybean paste. LWT 2023, 188, 115471. [Google Scholar] [CrossRef]
- Saud, S.; et al. The consequences of fermentation metabolism on the qualitative qualities and biological activity of fermented fruit and vegetable juices. Food Chemistry: X 2024, 21, 101209. [Google Scholar] [CrossRef]
- Wang, Y.; et al. Investigation of volatile compounds during fermentation of Elaeagnus moorcroftii Wall.ex schlecht. juice by Bifidobacterium animalis subsp. lactis HN-3 and Lacticaseibacillus paracasei YL-29. Food Chemistry: X 2024, 21, 101171. [Google Scholar] [CrossRef]
- He, T.; et al. Studies on the Changes of Fermentation Metabolites and the Protective Effect of Fermented Edible Grass on Stress Injury Induced by Acetaminophen in HepG2 Cells. Foods 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Letizia, F.; et al. Antioxidative Properties of Fermented Soymilk Using Lactiplantibacillus plantarum LP95. Antioxidants 2023, 12. [Google Scholar] [CrossRef]
- Sharma, H.; et al. Impact of lactic acid bacteria and their metabolites on the techno-functional properties and health benefits of fermented dairy products. Critical Reviews in Food Science and Nutrition 2023, 63, 4819–4841. [Google Scholar] [CrossRef]
- Feng, R.; et al. Fermentation trip: amazing microbes, amazing metabolisms. Annals of Microbiology 2018, 68, 717–729. [Google Scholar] [CrossRef]
- Popova-Krumova, P.; et al. Lactic Acid Production by Lactiplantibacillus plantarum AC 11S—Kinetics and Modeling. Microorganisms 2024, 12. [Google Scholar] [CrossRef]
- Zhao, Z.; et al. Metabolites changes of a low-temperature and low-salt fermented Chinese kohlrabi during fermentation based on non-targeted metabolomic analysis. Frontiers in Sustainable Food Systems 2023, 7. [Google Scholar] [CrossRef]
- Cheng, T.; et al. Improved production of andrimid in Erwinia persicina BST187 strain by fermentation optimization. BMC Microbiology 2023, 23, 268. [Google Scholar] [CrossRef]
- Idrees, M.; et al. Draft genome sequence of Lactiplantibacillus plantarum subsp. plantarum strain HF43, a human gut-associated potential probiotic. Microbiology Resource Announcements 2023, 12, e00945-00922. [Google Scholar] [CrossRef]
- Karaseva, O.; et al. Whole Genome Sequencing of the Novel Probiotic Strain Lactiplantibacillus plantarum FCa3L. Microorganisms 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Suryavanshi, M.V.; et al. Draft genome sequence of Lactobacillus plantarum strains E2C2 and E2C5 isolated from human stool culture. Standards in Genomic Sciences 2017, 12, 15. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; et al. How Lactobacillus plantarum shapes its transcriptome in response to contrasting habitats. Environmental Microbiology 2018, 20, 3700–3716. [Google Scholar] [CrossRef]
- Reverón, I.; et al. Transcriptomic Evidence of Molecular Mechanisms Underlying the Response of Lactobacillus plantarum WCFS1 to Hydroxytyrosol. Antioxidants 2020, 9. [Google Scholar] [CrossRef]
- Achi, O.K.; Asamudo, N.U. Cereal-Based Fermented Foods of Africa as Functional Foods. In Bioactive Molecules in Food; Mérillon, J.-M., Ramawat, K.G., Eds.; Springer International Publishing: Cham, 2018; pp. 1–32. [Google Scholar]
- Claus, H.; Mojsov, K. Enzymes for Wine Fermentation: Current and Perspective Applications. Fermentation 2018, 4. [Google Scholar] [CrossRef]
- Raveendran, S.; et al. Applications of Microbial Enzymes in Food Industry. Food Technol Biotechnol 2018, 56, 16–30. [Google Scholar] [CrossRef]
- Xu, Y.; et al. Technological roles of microorganisms in fish fermentation: a review. Critical Reviews in Food Science and Nutrition 2021, 61, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Thapa, S.; et al. Biochemical Characteristics of Microbial Enzymes and Their Significance from Industrial Perspectives. Molecular Biotechnology 2019, 61, 579–601. [Google Scholar] [CrossRef] [PubMed]
- Darbandi, A.; et al. Application of microbial enzymes in medicine and industry: current status and future perspectives. Future microbiology 2024, 19, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Althuri, A.; et al. Microbial Enzymes and Lignocellulosic Fuel Production; 2017; pp. 135–170. [Google Scholar]
- Ahmad, M.; et al. An overview of the enzyme: Amylase and its industrial potentials. Bayero Journal of Pure and Applied Sciences 2019, 12, 352–358. [Google Scholar] [CrossRef]
- Wayllace, N.M.; et al. Microbial glucoamylases: structural and functional properties and biotechnological uses. World Journal of Microbiology and Biotechnology 2023, 39, 293. [Google Scholar] [CrossRef]
- dos Santos Aguilar, J.G.; Sato, H.H. Microbial proteases: Production and application in obtaining protein hydrolysates. Food Research International 2018, 103, 253–262. [Google Scholar] [CrossRef]
- Ward, O.P. Proteases; Comprehensive Biotechnology, 2019. [Google Scholar]
- Christensen, L.F.; et al. Extracellular microbial proteases with specificity for plant proteins in food fermentation. International Journal of Food Microbiology 2022, 381, 109889. [Google Scholar] [CrossRef]
- Jokanovic, M.; et al. Proteolysis and texture profile of traditional dry-fermented sausage as affected by primary processing method. Scientific journal" Meat Technology" 2017, 58, 103–109. [Google Scholar]
- Mehta, A.; et al. 6 - The lipases and their applications with emphasis on food industry. In Microbial Biotechnology in Food and Health; Ray, R.C., Ed.; Academic Press, 2021; pp. 143–164. [Google Scholar]
- Salgado, C.A.; et al. Microbial lipases: Propitious biocatalysts for the food industry. Food Bioscience 2022, 45, 101509. [Google Scholar] [CrossRef]
- Negi, S. Lipases: A Promising Tool for Food Industry. In Green Bio-processes: Enzymes in Industrial Food Processing; Parameswaran, B., Varjani, S., Raveendran, S., Eds.; Springer Singapore: Singapore, 2019; pp. 181–198. [Google Scholar]
- Kaushal, J.; et al. Catalase enzyme: Application in bioremediation and food industry. Biocatalysis and Agricultural Biotechnology 2018, 16, 192–199. [Google Scholar] [CrossRef]
- Bajaj, P.; Mahajan, R. Cellulase and xylanase synergism in industrial biotechnology. Applied Microbiology and Biotechnology 2019, 103, 8711–8724. [Google Scholar] [CrossRef]
- Alokika; Singh, B. Production, characteristics, and biotechnological applications of microbial xylanases. Applied Microbiology and Biotechnology 2019, 103, 8763–8784. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.S.; et al. Chapter 1 - Microbial Enzymes—An Overview. In Advances in Enzyme Technology; Singh, R.S., Singhania, R.R., Pandey, A., Larroche, C., Eds.; Elsevier, 2019; pp. 1–40. [Google Scholar]
- Xavier, J.R.; et al. β-galactosidase: Biotechnological applications in food processing. Journal of Food Biochemistry 2018, 42, e12564. [Google Scholar] [CrossRef]
- Kumar, S.S.; et al. Tannase and Its Applications in Food Processing. In Green Bio-processes: Enzymes in Industrial Food Processing; Parameswaran, B., Varjani, S., Raveendran, S., Eds.; Springer Singapore: Singapore, 2019; pp. 357–381. [Google Scholar]
- Sahu, A.; Parihar, D.K. Exploring the Versatile Role of Tannase Enzyme in Food Industry. Chhattisgarh Journal of Science and Technology 2021. [Google Scholar] [CrossRef]
- Yang, H.; et al. The production and application of enzymes related to the quality of fruit wine. Critical Reviews in Food Science and Nutrition 2021, 61, 1605–1615. [Google Scholar] [CrossRef]
- Sharma, D.; et al. Immobilization and applications of esterases. Biocatalysis and Biotransformation 2022, 40, 153–168. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids as natural functional pigments. Journal of Natural Medicines 2020, 74, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Riaz, M.; et al. Chemistry of Carotenoids. In Carotenoids: Structure and Function in the Human Body; Zia-Ul-Haq, M., Dewanjee, S., Riaz, M., Eds.; Springer International Publishing: Cham, 2021; pp. 43–76. [Google Scholar]
- Foong, L.C.; et al. Recent development in the production strategies of microbial carotenoids. World Journal of Microbiology and Biotechnology 2021, 37, 12. [Google Scholar] [CrossRef]
- Saini, R.K.; et al. An updated review on use of tomato pomace and crustacean processing waste to recover commercially vital carotenoids. Food Research International 2018, 108, 516–529. [Google Scholar] [CrossRef] [PubMed]
- Devi, M.; et al. Bacteria as a source of biopigments and their potential applications. Journal of Microbiological Methods 2024, 219, 106907. [Google Scholar] [CrossRef]
- Ayadi, I.; et al. Microbial bioactive compounds from oleaginous yeast culture: insights into molecular docking interactions and toxicity prediction. Biomass Conversion and Biorefinery 2025, 15, 923–941. [Google Scholar] [CrossRef]
- Alcaíno, J.; et al. Carotenoid Distribution in Nature. In Carotenoids in Nature: Biosynthesis, Regulation and Function; Stange, C., Ed.; Springer International Publishing: Cham, 2016; pp. 3–33. [Google Scholar]
- Li, Z.; et al. New strategies to study in depth the metabolic mechanism of astaxanthin biosynthesis in Phaffia rhodozyma. Critical Reviews in Biotechnology 2024, 1–19. [Google Scholar] [CrossRef]
- Saubenova, M.; et al. Production of Carotenoids by Microorganisms. Fermentation 2024, 10. [Google Scholar] [CrossRef]
- Celedón, R.S.; Díaz, L.B. Natural Pigments of Bacterial Origin and Their Possible Biomedical Applications. Microorganisms 2021, 9. [Google Scholar] [CrossRef]
- Paul, D.; et al. Yeast Carotenoids: Cost-Effective Fermentation Strategies for Health Care Applications. Fermentation 2023, 9. [Google Scholar] [CrossRef]
- Sharma, C.; et al. Microbial-Derived Carotenoids and Their Health Benefits. Microbiology Research 2024, 15, 1670–1689. [Google Scholar] [CrossRef]
- Mapelli-Brahm, P.; et al. The impact of fermentation processes on the production, retention and bioavailability of carotenoids: An overview. Trends in Food Science & Technology 2020, 99, 389–401. [Google Scholar] [CrossRef]
- Wang, L.; et al. Biotechnology advances in β-carotene production by microorganisms. Trends in Food Science & Technology 2021, 111, 322–332. [Google Scholar] [CrossRef]
- Yamada, R.; et al. Improving carotenoid production in recombinant yeast, Saccharomyces cerevisiae, using ultrasound-irradiated two-phase extractive fermentation. Engineering in Life Sciences 2022, 22, 4–12. [Google Scholar] [CrossRef]
- Aruldass, C.A.; et al. Current perspective of yellowish-orange pigments from microorganisms- a review. Journal of Cleaner Production 2018, 180, 168–182. [Google Scholar] [CrossRef]
- Chavez-Parga, M.D.C.; et al. Optimization of Zeaxanthin Production by Immobilized Flavobacterium sp. Cells in Fluidized Bed Bioreactor. Advances in Microbiology 2012, Vol.02(No.04), 7. [Google Scholar] [CrossRef]
- Jafari, R.; et al. Optimization and identification of astaxanthin esters from shrimp waste using microbial fermentation method. Biomass Conversion and Biorefinery 2024, 14, 22893–22908. [Google Scholar] [CrossRef]
- Baptista-Silva, S.; et al. The progress of essential oils as potential therapeutic agents: a review. Journal of Essential Oil Research 2020, 32, 279–295. [Google Scholar] [CrossRef]
- Pateiro, M.; et al. Application of essential oils as antimicrobial agents against spoilage and pathogenic microorganisms in meat products. International Journal of Food Microbiology 2021, 337, 108966–108966. [Google Scholar] [CrossRef]
- Herrera-Calderon, O.; et al. Chemical constituents, in vitro antioxidant activity and in silico study on NADPH Oxidase of Allium sativum L. (garlic) essential oil. Antioxidants 2021, 10, 1844–1844. [Google Scholar] [CrossRef]
- Nirmala, M.J.; et al. Anticancer and antibacterial effects of a clove bud essential oil-based nanoscale emulsion system. International Journal of Nanomedicine 2019, 14, 6439–6450. [Google Scholar] [CrossRef] [PubMed]
- Gedikoğlu, A.; et al. Evaluation of Thymus vulgaris and Thymbra spicata essential oils and plant extracts for chemical composition, antioxidant, and antimicrobial properties. Food Science & Nutrition 2019, 7, 1704–1714. [Google Scholar] [CrossRef]
- Değirmenci, H.; Erkurt, H. Relationship between volatile components, antimicrobial and antioxidant properties of the essential oil, hydrosol and extracts of Citrus aurantium L. flowers. Journal of Infection and Public Health 2020, 13, 58–67. [Google Scholar] [CrossRef]
- Perumal, A.B.; et al. Application of essential oils in packaging films for the preservation of fruits and vegetables: A review. Food Chemistry 2022, 375, 131810–131810. [Google Scholar] [CrossRef]
- Ghasemy-Piranloo, F.; et al. Comparison for the production of essential oil by conventional, novel and biotechnology methods. Journal of Essential Oil Research 2022, 34, 455–478. [Google Scholar] [CrossRef]
- Heffernan, D.; et al. Screening of volatile organic compounds (VOCs) from liquid fungal cultures using ambient mass spectrometry. Analytical and Bioanalytical Chemistry 2023, 415, 4615–4627. [Google Scholar] [CrossRef] [PubMed]
- Iraji, A.; et al. Screening the antifungal activities of monoterpenes and their isomers against Candida species. Journal of Applied Microbiology 2020, 129, 1541–1551. [Google Scholar] [CrossRef] [PubMed]
- Demirel, S. Geraniol and β-citronellol participate in the vasorelaxant effects of Rosa damascena Miller essential oil on the rat thoracic aorta. Fitoterapia 2022, 161, 105243–105243. [Google Scholar] [CrossRef]
- Sen, S.; et al. Chemometric analysis reveals links in the formation of fragrant bio-molecules during agarwood (Aquilaria malaccensis) and fungal interactions. Scientific Reports 2017, 7, 44406–44406. [Google Scholar] [CrossRef]
- Bluemke, W.; Schrader, J. Integrated bioprocess for enhanced production of natural flavors and fragrances by Ceratocystis moniliformis. Biomolecular Engineering 2001, 17, 137–142. [Google Scholar] [CrossRef]
- Mahmoud, E.; et al. Production of flavors from agro waste of Ocimumbasilicum L. by different microorganisms using solid state fermentation. Egyptian Journal of Chemistry 2022, 65, 259–273. [Google Scholar] [CrossRef]
- Khojali, I.O.; Mohammed, Y.M. Bisabolene compound extracted from Cassia fistula and docked as antioxidant and vitamin E alternative predicted drug design. GSC Advanced Research and Reviews 2023, 15, 069–075. [Google Scholar] [CrossRef]
- Ahmed, A.F.; et al. Antioxidant activity and total phenolic content of essential oils and extracts of sweet basil (Ocimum basilicum L.) plants. Food Science and Human Wellness 2019, 8, 299–305. [Google Scholar] [CrossRef]
- Ling, L.; et al. Antifungal activity of diacetyl, a volatile organic compound, on Trichoderma lixii F2 isolated from postharvest Lanzhou lily bulbs. Food Bioscience 2023, 52, 102365–102365. [Google Scholar] [CrossRef]
- Ion, V.A.; et al. Physico-chemical parameters and antioxidant activity of Romanian Sea Buckthorn Berries. Revista de Chimie 2019, 70, 4187–4192. [Google Scholar] [CrossRef]
- Guneser, O.; et al. Production of flavor compounds from olive mill waste by Rhizopus oryzae and Candida tropicalis. Brazilian Journal of Microbiology 2017, 48, 275–285. [Google Scholar] [CrossRef]
- Zheng, L.; et al. Production of vanillin from waste residue of rice bran oil by Aspergillus niger and Pycnoporus cinnabarinus. Bioresource Technology 2007, 98, 1115–1119. [Google Scholar] [CrossRef]
- Ndao, A.; Adjallé, K. Overview of the Biotransformation of Limonene and α-Pinene from Wood and Citrus Residues by Microorganisms. Waste 2023, 1, 841–859. [Google Scholar] [CrossRef]
- Chatterjee, T. Biotransformation of geraniol by Rhodococcus sp. strain GR3. Biotechnology and Applied Biochemistry 2004, 39, 303–306. [Google Scholar] [CrossRef]
- Oda, S.; et al. Synthesis of citronellyl acetate via a transacetylation to citronellol from Acetyl Coenzyme A produced from glucose and acetate in growing yeasts. Chemistry Letters 2001, 30, 500–501. [Google Scholar] [CrossRef]
- Achmon, Y.; et al. Hydrophobic microspheres for in situ removal of 2-phenylethanol from yeast fermentation. Journal of Microencapsulation 2011, 28, 628–638. [Google Scholar] [CrossRef]
- Ueno, H.; et al. Anti-depressive-like effect of 2-phenylethanol inhalation in mice. Biomedicine & Pharmacotherapy 2019, 111, 1499–1506. [Google Scholar] [CrossRef]
- Jiang, G.-Z.; et al. Manipulation of GES and ERG20 for geraniol overproduction in Saccharomyces cerevisiae. Metabolic Engineering 2017, 41, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Gerke, J.; et al. Production of the fragrance geraniol in peroxisomes of a product-tolerant baker’s yeast. Frontiers in Bioengineering and Biotechnology 2020, 8, 582052–582052. [Google Scholar] [CrossRef]
- Cheng, S.; et al. Orthogonal engineering of biosynthetic pathway for efficient production of limonene in Saccharomyces cerevisiae. ACS Synthetic Biology 2019, 8, 968–975. [Google Scholar] [CrossRef] [PubMed]
- Werner, N.; et al. Protein engineering of carotenoid cleavage dioxygenases to optimize β-ionone biosynthesis in yeast cell factories. Food Chemistry 2019, 299, 125089–125089. [Google Scholar] [CrossRef]
- Zhou, L.; et al. Enhancement of patchoulol production in Escherichia coli via multiple engineering strategies. Journal of Agricultural and Food Chemistry 2021, 69, 7572–7580. [Google Scholar] [CrossRef]
- Shukal, S.; et al. Systematic engineering for high-yield production of viridiflorol and amorphadiene in auxotrophic Escherichia coli. Metabolic Engineering 2019, 55, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Lukin, I.; et al. Recovery of natural α-Ionone from fermentation broth. Journal of Agricultural and Food Chemistry 2019, 67, 13412–13419. [Google Scholar] [CrossRef]
- Han, G.H.; et al. Fermentative production and direct extraction of (−)-α-bisabolol in metabolically engineered Escherichia coli. Microbial Cell Factories 2016, 15, 185–185. [Google Scholar] [CrossRef]
- Mahfuz, S.; et al. Phenolic compounds as natural feed additives in poultry and swine diets: a review. Journal of Animal Science and Biotechnology 2021, 12, 48. [Google Scholar] [CrossRef]
- Papuc, C.; et al. Plant Polyphenols as Antioxidant and Antibacterial Agents for Shelf-Life Extension of Meat and Meat Products: Classification, Structures, Sources, and Action Mechanisms. Comprehensive Reviews in Food Science and Food Safety 2017, 16, 1243–1268. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnology Reports 2019, 24, e00370. [Google Scholar] [CrossRef]
- Febrianto, N.A.; Zhu, F. Changes in the Composition of Methylxanthines, Polyphenols, and Volatiles and Sensory Profiles of Cocoa Beans from the Sul 1 Genotype Affected by Fermentation. Journal of Agricultural and Food Chemistry 2020, 68, 8658–8675. [Google Scholar] [CrossRef]
- Chagas Junior, G.C.A.; et al. Chemical implications and time reduction of on-farm cocoa fermentation by Saccharomyces cerevisiae and Pichia kudriavzevii. Food Chemistry 2021, 338, 127834. [Google Scholar] [CrossRef]
- Cañas, S.; et al. Transformations of phenolic compounds in cocoa shell during in vitro colonic fermentation. Current Research in Food Science 2024, 9, 100930. [Google Scholar] [CrossRef] [PubMed]
- Cañas, S.; et al. Microbial catabolism of coffee pulp (poly)phenols during in vitro colonic fermentation. Food Chemistry 2025, 463, 141354. [Google Scholar] [CrossRef] [PubMed]
- Myo, H.; et al. Changes in Bioactive Compounds of Coffee Pulp through Fermentation-Based Biotransformation Using Lactobacillus plantarum TISTR 543 and Its Antioxidant Activities. Fermentation 2021, 7. [Google Scholar] [CrossRef]
- Zeng, Z.; et al. The Profile and Bioaccessibility of Phenolic Compounds in Cereals Influenced by Improved Extrusion Cooking Treatment. PLOS ONE 2016, 11, e0161086. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; et al. A review on insoluble-bound phenolics in plant-based food matrix and their contribution to human health with future perspectives. Trends in Food Science & Technology 2020, 105, 347–362. [Google Scholar] [CrossRef]
- Azkia, M.N.; et al. Enhancement of phenolic profile and antioxidant activity of black glutinous rice (Oryza sativa var. glutinosa) due to tape fermentation. Arabian Journal of Chemistry 2023, 16, 105275. [Google Scholar] [CrossRef]
- Zhang, D.; et al. Changing the polyphenol composition and enhancing the enzyme activity of sorghum grain by solid-state fermentation with different microbial strains. Journal of the Science of Food and Agriculture 2024, 104. [Google Scholar] [CrossRef]
- Mancino, W.; et al. Hierarchical Effects of Lactic Fermentation and Grain Germination on the Microbial and Metabolomic Profile of Rye Doughs. Foods 2023, 12. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; et al. Bound phenolics in foods, a review. Food Chemistry 2014, 152, 46–55. [Google Scholar] [CrossRef]
- Wang, Y.-L.; et al. The release and catabolism of ferulic acid in plant cell wall by rumen microbes: A review. Animal Nutrition 2022, 9, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Marco, M.L.; et al. Health benefits of fermented foods: microbiota and beyond. Current Opinion in Biotechnology 2017, 44, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Borghi, S.M.; Pavanelli, W.R. Antioxidant Compounds and Health Benefits of Citrus Fruits. Antioxidants 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; et al. Phenolic composition, antioxidant potential and health benefits of citrus peel. Food Research International 2020, 132, 109114. [Google Scholar] [CrossRef]
- Tang, R.; et al. Biotransformation of citrus fruits phenolic profiles by mixed probiotics in vitro anaerobic fermentation. LWT 2022, 160, 113087. [Google Scholar] [CrossRef]
- Thai Nguyen, H.; et al. Optimising the recovery of phenolic compounds and antioxidant activity from orange peels through solid-state fermentation. Natural product research 2024, 1–10. [Google Scholar] [CrossRef]
- Liu, Z.; et al. A systematic review on fermented chili pepper products: Sensorial quality, health benefits, fermentation microbiomes, and metabolic pathways. Trends in Food Science & Technology 2023, 141, 104189. [Google Scholar] [CrossRef]
- Muñoz, R.; et al. Chapter 4 - Biotransformation of Phenolics by Lactobacillus plantarum in Fermented Foods. In Fermented Foods in Health and Disease Prevention; Frias, J., Martinez-Villaluenga, C., Peñas, E., Eds.; Academic Press: Boston, 2017; pp. 63–83. [Google Scholar]
- Li, M.; et al. Exploring the phytochemicals and inhibitory effects against α-glucosidase and dipeptidyl peptidase-IV in Chinese pickled chili pepper: Insights into mechanisms by molecular docking analysis. LWT 2022, 162, 113467. [Google Scholar] [CrossRef]
- Dong, R.; et al. Release and metabolism of bound polyphenols from carrot dietary fiber and their potential activity in in vitro digestion and colonic fermentation. Food & Function 2020, 11, 6652–6665. [Google Scholar] [CrossRef]
- Sagar, N.A.; Pareek, S. Dough rheology, antioxidants, textural, physicochemical characteristics, and sensory quality of pizza base enriched with onion (Allium cepa L.) skin powder. Scientific Reports 2020, 10, 18669. [Google Scholar] [CrossRef]
- Ramires, F.A.; et al. Liquid submerged fermentation by selected microbial strains for onion skins valorization and its effects on polyphenols. World Journal of Microbiology and Biotechnology 2023, 39, 258. [Google Scholar] [CrossRef] [PubMed]
- Slimestad, R.; et al. Flavonoids and other phenolics in herbs commonly used in Norwegian commercial kitchens. Food Chemistry 2020, 309, 125678. [Google Scholar] [CrossRef]
- Sinaga, D.P.; et al. Fermentation Process Effect to Enhance Antioxidant and Antibacterial Activity of Phenolic Compounds and Its Possible Application to Galactomannan Polysaccharides: A Review. IOP Conference Series: Earth and Environmental Science 2022, 1097, 012027. [Google Scholar] [CrossRef]
- Yu, G.; et al. Improvement of antioxidant activity and active ingredient of Dendrobium officinale via microbial fermentation. Frontiers in Microbiology 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Jalao, I.; et al. Modulation of Gut Microbiota Composition and Microbial Phenolic Catabolism of Phenolic Compounds from Achillea millefolium L. and Origanum majorana L. Journal of Agricultural and Food Chemistry 2025, 73, 478–494. [Google Scholar] [CrossRef]
- Pinto, T.; Vilela, A. Healthy Drinks with Lovely Colors: Phenolic Compounds as Constituents of Functional Beverages. Beverages 2021, 7. [Google Scholar] [CrossRef]
- Guan, X.; et al. Variation of taste and odor compounds in tea beverage after microbial fermentation by HPLC–MS, GC×GC–O–MS, GC–MS, and sensory evaluation. Journal of Food Composition and Analysis 2024, 128, 106075. [Google Scholar] [CrossRef]
- Tian, Z.-X.; et al. Effects of six different microbial strains on polyphenol profiles, antioxidant activity, and bioaccessibility of blueberry pomace with solid-state fermentation. Frontiers in Nutrition 2023, 10. [Google Scholar] [CrossRef]
- Liu, S.; et al. Comparison of phenolic profiles of albino bilberry (Vaccinium myrtillus L.) wines fermented by non-Saccharomyces yeasts. Food Bioscience 2023, 55, 102980. [Google Scholar] [CrossRef]
- Shen, C.; et al. The dynamic of physicochemical properties, volatile compounds and microbial community during the fermentation of Chinese rice wine with diverse cereals. Food Research International 2024, 198, 115319. [Google Scholar] [CrossRef]
- Leonard, W.; et al. Fermentation transforms the phenolic profiles and bioactivities of plant-based foods. Biotechnology Advances 2021, 49, 107763. [Google Scholar] [CrossRef]
- Kafkas, E.; et al. Advanced Analytical Methods for Phenolics in Fruits. Journal of Food Quality 2018, 2018, 1–6. [Google Scholar] [CrossRef]
- Ozturk, T.; et al. Impact of Lactic Acid Bacteria Fermentation on (Poly)Phenolic Profile and In Vitro Antioxidant and Anti-Inflammatory Properties of Herbal Infusions. Antioxidants 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Lama-Muñoz, A.; Contreras, M.D. Extraction Systems and Analytical Techniques for Food Phenolic Compounds: A Review. Foods 2022, 11. [Google Scholar] [CrossRef]
- Prateeksha; et al. Microbial polysaccharides: An emerging family of natural biomaterials for cancer therapy and diagnostics. Seminars in Cancer Biology 2022, 86, 706–731. [Google Scholar] [CrossRef]
- Shanmugam, M.; Abirami, R.G. Microbial polysaccharides - chemistry and applications. Journal of Biologically Active Products from Nature 2019, 9, 73–78. [Google Scholar] [CrossRef]
- Guérin, M.; et al. Lactic acid bacterial production of exopolysaccharides from fruit and vegetables and associated benefits. Fermentation 2020, 6, 115–115. [Google Scholar] [CrossRef]
- Zikmanis, P.; et al. Extracellular polysaccharides produced by bacteria of the Leuconostoc genus. World Journal of Microbiology and Biotechnology 2020, 36, 161–161. [Google Scholar] [CrossRef]
- Esmaeilnejad-Moghadam, B.; et al. Low molecular weight dextran production by Leuconostoc mesenteroides strains: Optimization of a new culture medium and the rheological assessments. Bioactive Carbohydrates and Dietary Fibre 2019, 18, 100181–100181. [Google Scholar] [CrossRef]
- Schmid, J.; et al. Analysis of structural and functional differences of glucans produced by the natively released dextransucrase of Liquorilactobacillus hordei TMW 1.1822. Applied Biochemistry and Biotechnology 2021, 193, 96–110. [Google Scholar] [CrossRef]
- Farinazzo, F.S.; et al. Characterization and antioxidant activity of an exopolysaccharide produced by Leuconostoc pseudomesenteroides JF17 from juçara fruits (Euterpe edulis Martius). Process Biochemistry 2020, 91, 141–148. [Google Scholar] [CrossRef]
- Huang, F.; et al. Physicochemical and biological properties of longan pulp polysaccharides modified by Lactobacillus fermentum fermentation. International Journal of Biological Macromolecules 2019, 125, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Vallejo-García, L.C.; et al. Enzymatic process yielding a diversity of inulin-type microbial fructooligosaccharides. Journal of Agricultural and Food Chemistry 2019, 67, 10392–10400. [Google Scholar] [CrossRef]
- Sun, T.; et al. High-efficiency production of Tremella aurantialba polysaccharide through basidiospore fermentation. Bioresource Technology 2020, 318, 124268–124268. [Google Scholar] [CrossRef]
- Ge, X.; et al. Production, structure, and bioactivity of polysaccharide isolated from Tremella fuciformis XY. International Journal of Biological Macromolecules 2020, 148, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; et al. Studies on the purification of polysaccharides separated from Tremella fuciformis and their neuroprotective effect. Molecular Medicine Reports 2016, 13, 3985–3992. [Google Scholar] [CrossRef]
- Zhou, Y.; et al. Immunomodulatory effect of Tremella polysaccharides against cyclophosphamide-induced immunosuppression in mice. Molecules 2018, 23, 239–239. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; et al. A review on the production, structure, bioactivities and applications of Tremella polysaccharides. International Journal of Immunopathology and Pharmacology 2021, 35, 20587384211000541–20587384211000541. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; et al. Effect of yeast fermentation on the physicochemical properties and bioactivities of polysaccharides of Dendrobium officinale. Foods 2022, 12, 150–150. [Google Scholar] [CrossRef]
- Filippousi, R.; et al. Isolation, identification and screening of yeasts towards their ability to assimilate biodiesel-derived crude glycerol: microbial production of polyols, endopolysaccharides and lipid. Journal of Applied Microbiology 2019, 127, 1080–1100. [Google Scholar] [CrossRef]
- Lin, D.; et al. Study on the functional properties and structural characteristics of soybean soluble polysaccharides by mixed bacteria fermentation and microwave treatment. International Journal of Biological Macromolecules 2020, 157, 561–568. [Google Scholar] [CrossRef]
- Chen, Q.; et al. Characterization and antioxidant activity of wheat bran polysaccharides modified by Saccharomyces cerevisiae and Bacillus subtilis fermentation. Journal of Cereal Science 2021, 97, 103157–103157. [Google Scholar] [CrossRef]
- Srikanth, R.; et al. Antioxidant and anti-inflammatory levan produced from Acetobacter xylinum NCIM2526 and its statistical optimization. Carbohydrate Polymers 2015, 123, 8–16. [Google Scholar] [CrossRef]
- Imran, M.Y.M.; et al. Statistical optimization of exopolysaccharide production by Lactobacillus plantarum NTMI05 and NTMI20. International Journal of Biological Macromolecules 2016, 93, 731–745. [Google Scholar] [CrossRef]
- Yegorenkova, I.V.; et al. Immunomodulatory activity of exopolysaccharide from the rhizobacterium Paenibacillus polymyxa CCM 1465. Archives of Microbiology 2018, 200, 1471–1480. [Google Scholar] [CrossRef]
- Wangpaiboon, K.; et al. An α-1,6-and α-1,3-linked glucan produced by Leuconostoc citreum ABK-1 alternansucrase with nanoparticle and film-forming properties. Scientific Reports 2018, 8, 8340–8340. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; et al. Isolation and characterization of an antioxidant exopolysaccharide produced by Bacillus sp. S-1 from Sichuan Pickles. Carbohydrate Polymers 2019, 204, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.-k.; et al. Three exopolysaccharides from the liquid fermentation of Polyporus umbellatus and their bioactivities. International Journal of Biological Macromolecules 2019, 132, 629–640. [Google Scholar] [CrossRef]
- Sun, H.; et al. Effects of Fermenting the Plant Fraction of a Complete Feed on the Growth Performance, Nutrient Utilization, Antioxidant Functions, Meat Quality, and Intestinal Microbiota of Broilers. Animals 2022, 12. [Google Scholar] [CrossRef]
- Parmar, A.B.; et al. A solid state fermentation, its role in animal nutrition: A review. 2019, 4626–4633. [Google Scholar]
- Yafetto, L.; et al. Valorization of agro-industrial wastes into animal feed through microbial fermentation: A review of the global and Ghanaian case. Heliyon 2023, 9. [Google Scholar] [CrossRef]
- Marková, K.; et al. Are there consistent effects of gut microbiota composition on performance, productivity and condition in poultry? Poultry Science 2024, 103, 103752. [Google Scholar] [CrossRef]
- Lian, X.; et al. The Effects of Unconventional Feed Fermentation on Intestinal Oxidative Stress in Animals. Antioxidants 2024, 13. [Google Scholar] [CrossRef]
- Predescu, N.C.; et al. Fermented Feed in Broiler Diets Reduces the Antinutritional Factors, Improves Productive Performances and Modulates Gut Microbiome—A Review. Agriculture 2024, 14. [Google Scholar] [CrossRef]
- Sun, H.; et al. Progress in Fermented Unconventional Feed Application in Monogastric Animal Production in China. Fermentation 2023, 9. [Google Scholar] [CrossRef]
- Zhang, M.; et al. Effects of Probiotic-Fermented Feed on the Growth Profile, Immune Functions, and Intestinal Microbiota of Bamei Piglets. Animals 2024, 14, 647. [Google Scholar] [CrossRef]
- Siddik, S.; et al. Fermentation in aquafeed processing: Achieving sustainability in feeds for global aquaculture production. Reviews in Aquaculture 2024, 16. [Google Scholar] [CrossRef]
- Srichompoo, P.; et al. Effect of Replacing Corn Meal with Winged Bean Tuber (Psophocarpus tetragonolobus) Pellet on Gas Production, Ruminal Fermentation, and Degradability Using In Vitro Gas Technique. Animals 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; et al. Synergism of fermented feed and ginseng polysaccharide on growth performance, intestinal development, and immunity of Xuefeng black-bone chickens. BMC Veterinary Research 2024, 20, 13. [Google Scholar] [CrossRef]
- Nadarajah, K.; Abdul Rahman, N.S. The Microbial Connection to Sustainable Agriculture. Plants 2023, 12. [Google Scholar] [CrossRef]
- Trivedi, P.; et al. Tiny Microbes, Big Yields: Enhancing food crop production with biological solutions. Microbial Biotechnology 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; et al. Enhancing Soil Health and Plant Growth through Microbial Fertilizers: Mechanisms, Benefits, and Sustainable Agricultural Practices. Agronomy 2024, 14. [Google Scholar] [CrossRef]
- Chen, Q.; et al. Mechanisms and Impact of Rhizosphere Microbial Metabolites on Crop Health, Traits, Functional Components: A Comprehensive Review. Molecules 2024, 29. [Google Scholar] [CrossRef]
- Danchin, A. Exploring overlooked growth-promoting mechanisms by plant-associated bacteria. Sustainable Microbiology 2024, 1, qvae011. [Google Scholar] [CrossRef]
- Pang, Z.; et al. Linking Plant Secondary Metabolites and Plant Microbiomes: A Review. Frontiers in Plant Science 2021, 12. [Google Scholar] [CrossRef]
- Iqbal, B.; et al. Advancing environmental sustainability through microbial reprogramming in growth improvement, stress alleviation, and phytoremediation. Plant Stress 2023, 10, 100283. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; et al. Metabolomics-guided utilization of beneficial microbes for climate-resilient crops. Current Opinion in Chemical Biology 2024, 79, 102427. [Google Scholar] [CrossRef]
- Morcillo, R.J.L.; Manzanera, M. The Effects of Plant-Associated Bacterial Exopolysaccharides on Plant Abiotic Stress Tolerance. Metabolites 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, U.; et al. Understanding the roles of osmolytes for acclimatizing plants to changing environment: a review of potential mechanism. Plant Signaling & Behavior 2021, 16, e1913306. [Google Scholar] [CrossRef]
- Koza, N.A.; et al. Microorganisms in Plant Growth and Development: Roles in Abiotic Stress Tolerance and Secondary Metabolites Secretion. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Shaffique, S.; et al. Research Progress in the Field of Microbial Mitigation of Drought Stress in Plants. Frontiers in Plant Science 2022, 13, 870626. [Google Scholar] [CrossRef]
- O'Callaghan, M.; et al. Soil microbial inoculants for sustainable agriculture: Limitations and opportunities. Soil Use and Management 2022, 38. [Google Scholar] [CrossRef]
- Bertola, M.; et al. Improvement of Soil Microbial Diversity through Sustainable Agricultural Practices and Its Evaluation by -Omics Approaches: A Perspective for the Environment, Food Quality and Human Safety. Microorganisms 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; et al. Microbial Utilization of Next-Generation Feedstocks for the Biomanufacturing of Value-Added Chemicals and Food Ingredients. Frontiers in Bioengineering and Biotechnology 2022, 10, 874612. [Google Scholar] [CrossRef]
- Samoraj, M.; et al. Advancements in algal biorefineries for sustainable agriculture: Biofuels, high-value products, and environmental solutions. Biocatalysis and Agricultural Biotechnology 2024, 58, 103224. [Google Scholar] [CrossRef]
- Peña-Castro, J.M.; et al. Engineering the Metabolic Landscape of Microorganisms for Lignocellulosic Conversion. Microorganisms 2023, 11. [Google Scholar] [CrossRef]
- Upadhayay, V.K.; et al. Microbial Advancement in Agriculture; 2024; pp. 95–125. [Google Scholar]
- Ezeorba, T.P.C.; et al. Recent advances in biotechnological valorization of agro-food wastes (AFW): Optimizing integrated approaches for sustainable biorefinery and circular bioeconomy. Bioresource Technology Reports 2024, 26, 101823. [Google Scholar] [CrossRef]
- Tamang, J.P.; et al. Functional Properties of Microorganisms in Fermented Foods. Front Microbiol 2016, 7, 578. [Google Scholar] [CrossRef]
- Cuvas-Limon, R.B.; et al. Spontaneously fermented traditional beverages as a source of bioactive compounds: an overview. Crit Rev Food Sci Nutr 2021, 61, 2984–3006. [Google Scholar] [CrossRef]
- Terpou, A.; et al. Probiotics in Food Systems: Significance and Emerging Strategies Towards Improved Viability and Delivery of Enhanced Beneficial Value. Nutrients 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E.; et al. Shared mechanisms among probiotic taxa: implications for general probiotic claims. Curr Opin Biotechnol 2018, 49, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Lopetuso, L.R.; et al. Bacteriocins and Bacteriophages: Therapeutic Weapons for Gastrointestinal Diseases? Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- George Kerry, R.; et al. Benefaction of probiotics for human health: A review. Journal of Food and Drug Analysis 2018, 26, 927–939. [Google Scholar] [CrossRef]
- Kober, M.M.; Bowe, W.P. The effect of probiotics on immune regulation, acne, and photoaging. Int J Womens Dermatol 2015, 1, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Sivamaruthi, B.S. A comprehensive review on clinical outcome of probiotic and synbiotic therapy for inflammatory bowel diseases. Asian Pacific Journal of Tropical Biomedicine 2018, 8, 179–186. [Google Scholar] [CrossRef]
- den Besten, G.; et al. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J Lipid Res 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Bellaver, E.H.; Kempka, A.P. Potential of milk-derived bioactive peptides as antidiabetic, antihypertensive, and xanthine oxidase inhibitors: a comprehensive bibliometric analysis and updated review. Amino Acids 2023, 55, 1829–1855. [Google Scholar] [CrossRef]
- Khavinson, V.K.; et al. Peptide Regulation of Gene Expression: A Systematic Review. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Gill, H.S.; et al. Immunoregulatory peptides in bovine milk. Br J Nutr 2000, 84 Suppl 1, S111–117. [Google Scholar] [CrossRef]
- Mann, B.; et al. Chapter 14 - Bioactive Peptides from Whey Proteins. In Whey Proteins; Deeth, H.C., Bansal, N., Eds.; Academic Press, 2019; pp. 519–547. [Google Scholar]
- Deveci, G.; et al. Certain Fermented Foods and Their Possible Health Effects with a Focus on Bioactive Compounds and Microorganisms. Fermentation 2023, 9, 923. [Google Scholar] [CrossRef]
- Erdogan, F.S.; et al. The effect of kefir produced from natural kefir grains on the intestinal microbial populations and antioxidant capacities of Balb/c mice. Food Res Int 2019, 115, 408–413. [Google Scholar] [CrossRef]
- BELLIKCI-KOYU, E.; et al. Effects of Regular Kefir Consumption on Gut Microbiota in Patients with Metabolic Syndrome: A Parallel-Group, Randomized, Controlled Study. Nutrients 2019, 11, 2089. [Google Scholar] [CrossRef]
- Karaffová, V.; et al. Differences in Immune Response and Biochemical Parameters of Mice Fed by Kefir Milk and Lacticaseibacillus paracasei Isolated from the Kefir Grains. Microorganisms 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Hamet, M.F.; et al. Oral administration of kefiran exerts a bifidogenic effect on BALB/c mice intestinal microbiota. Benef Microbes 2016, 7, 237–246. [Google Scholar] [CrossRef]
- Malta, S.M.; et al. Identification of bioactive peptides from a Brazilian kefir sample, and their anti-Alzheimer potential in Drosophila melanogaster. Sci Rep 2022, 12, 11065. [Google Scholar] [CrossRef] [PubMed]
- Shafique, B.; et al. Proteolysis and therapeutic potential of bioactive peptides derived from Cheddar cheese. Food Science & Nutrition 2023, 11, 4948–4963. [Google Scholar] [CrossRef]
- Álvarez Ramos, L.; et al. Antioxidant and antihypertensive activity of Gouda cheese at different stages of ripening. Food Chemistry: X 2022, 14, 100284. [Google Scholar] [CrossRef] [PubMed]
- Helal, A.; Tagliazucchi, D. Peptidomics Profile, Bioactive Peptides Identification and Biological Activities of Six Different Cheese Varieties. Biology 2023, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Martín-Del-Campo, S.T.; et al. Production of Antioxidant and ACEI Peptides from Cheese Whey Discarded from Mexican White Cheese Production. Antioxidants (Basel) 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Bhanja Dey, T.; et al. Antioxidant phenolics and their microbial production by submerged and solid state fermentation process: A review. Trends in Food Science & Technology 2016, 53, 60–74. [Google Scholar] [CrossRef]
- Harahap, I.A.; et al. Fermented soy products: A review of bioactives for health from fermentation to functionality. Compr Rev Food Sci Food Saf 2025, 24, e70080. [Google Scholar] [CrossRef]
- Qiao, Y.; et al. Fermented soybean foods: A review of their functional components, mechanism of action and factors influencing their health benefits. Food Research International 2022, 158, 111575. [Google Scholar] [CrossRef] [PubMed]
- Oleszek, M.; Oleszek, W. Saponins in Food. In Handbook of Dietary Phytochemicals; Xiao, J., Sarker, S.D., Asakawa, Y., Eds.; Springer Singapore: Singapore, 2021; pp. 1501–1540. [Google Scholar]
- Swallah, M.S.; et al. The Pros and Cons of Soybean Bioactive Compounds: An Overview. Food Reviews International 2023, 39, 5104–5131. [Google Scholar] [CrossRef]
- Rahim, M.H.A.; et al. Roles of fermented plant-, dairy- and meat-based foods in the modulation of allergic responses. Food Science and Human Wellness 2023, 12, 691–701. [Google Scholar] [CrossRef]
- Shukla, S.; et al. Total phenolic content, antioxidant, tyrosinase and α-glucosidase inhibitory activities of water soluble extracts of noble starter culture Doenjang, a Korean fermented soybean sauce variety. Food Control 2016, 59, 854–861. [Google Scholar] [CrossRef]
- Wang, D.; et al. A novel method for highly efficient biotransformation and separation of isoflavone aglycones from soybean with high-speed counter-current chromatography. Industrial Crops and Products 2019, 129, 224–230. [Google Scholar] [CrossRef]
- Shin, D.; Jeong, D. Korean traditional fermented soybean products: Jang. Journal of Ethnic Foods 2015, 2, 2–7. [Google Scholar] [CrossRef]
- do Prado, F.G.; et al. Fermented Soy Products and Their Potential Health Benefits: A Review. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Singh, B.P.; et al. Soybean bioactive molecules: current trend and future prospective. In Bioactive molecules in food; Springer, 2019; pp. 267–294. [Google Scholar]
- Yang, X.; et al. Associations between intake of dietary fermented soy food and concentrations of inflammatory markers: a cross-sectional study in Japanese workers. J Med Invest 2018, 65, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Dias, G.P.; et al. The role of dietary polyphenols on adult hippocampal neurogenesis: molecular mechanisms and behavioural effects on depression and anxiety. Oxid Med Cell Longev 2012, 2012, 541971. [Google Scholar] [CrossRef]
- Makhamrueang, N.; et al. Beneficial Bio-Extract of Camellia sinensis var. assamica Fermented with a Combination of Probiotics as a Potential Ingredient for Skin Care. Cosmetics 2023, 10, 85. [Google Scholar] [CrossRef]
- Kim, E.; et al. Skin Protective Effect of Epigallocatechin Gallate. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef]
- Garaguso, I.; Nardini, M. Polyphenols content, phenolics profile and antioxidant activity of organic red wines produced without sulfur dioxide/sulfites addition in comparison to conventional red wines. Food Chem 2015, 179, 336–342. [Google Scholar] [CrossRef]
- Viegas, O.; et al. Simultaneous determination of melatonin and trans-resveratrol in wine by dispersive liquid–liquid microextraction followed by HPLC-FLD. Food Chemistry 2021, 339, 128091. [Google Scholar] [CrossRef]
- Álvarez-Fernández, M.A.; et al. Determination of hydroxytyrosol produced by winemaking yeasts during alcoholic fermentation using a validated UHPLC-HRMS method. Food Chem 2018, 242, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, I.; et al. The Pro-Oxidant Activity of Red Wine Polyphenols Induces an Adaptive Antioxidant Response in Human Erythrocytes. Antioxidants (Basel) 2021, 10. [Google Scholar] [CrossRef]
- Chalons, P.; et al. Red Wine Extract Disrupts Th17 Lymphocyte Differentiation in a Colorectal Cancer Context. Mol Nutr Food Res 2020, 64, e1901286. [Google Scholar] [CrossRef] [PubMed]
- Mahjabeen, W.; et al. Role of resveratrol supplementation in regulation of glucose hemostasis, inflammation and oxidative stress in patients with diabetes mellitus type 2: A randomized, placebo-controlled trial. Complement Ther Med 2022, 66, 102819. [Google Scholar] [CrossRef]
- Wu, H.; et al. The Cytotoxicity Effect of Resveratrol: Cell Cycle Arrest and Induced Apoptosis of Breast Cancer 4T1 Cells. Toxins (Basel) 2019, 11. [Google Scholar] [CrossRef]
- Chitimus, D.M.; et al. Melatonin's Impact on Antioxidative and Anti-Inflammatory Reprogramming in Homeostasis and Disease. Biomolecules 2020, 10. [Google Scholar] [CrossRef]
- Wang, P.; et al. Resveratrol reduces obesity in high-fat diet-fed mice via modulating the composition and metabolic function of the gut microbiota. Free Radic Biol Med 2020, 156, 83–98. [Google Scholar] [CrossRef]
- Wei, R.M.; et al. Resveratrol ameliorates maternal separation-induced anxiety- and depression-like behaviors and reduces Sirt1-NF-kB signaling-mediated neuroinflammation. Front Behav Neurosci 2023, 17, 1172091. [Google Scholar] [CrossRef] [PubMed]
- Iban-Arias, R.; et al. Role of Polyphenol-Derived Phenolic Acid in Mitigation of Inflammasome-Mediated Anxiety and Depression. Biomedicines 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Inchingolo, A.D.; et al. Benefits and Implications of Resveratrol Supplementation on Microbiota Modulations: A Systematic Review of the Literature. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Ousaaid, D.; et al. Fruits Vinegar: Quality Characteristics, Phytochemistry, and Functionality. Molecules 2021, 27. [Google Scholar] [CrossRef]
- Ousaaid, D.; et al. Beneficial Effects of Apple Vinegar on Hyperglycemia and Hyperlipidemia in Hypercaloric-Fed Rats. J Diabetes Res 2020, 2020, 9284987. [Google Scholar] [CrossRef]
- Halima, B.H.; et al. Apple Cider Vinegar Attenuates Oxidative Stress and Reduces the Risk of Obesity in High-Fat-Fed Male Wistar Rats. J Med Food 2018, 21, 70–80. [Google Scholar] [CrossRef]
- Yagnik, D.; et al. Antimicrobial activity of apple cider vinegar against Escherichia coli, Staphylococcus aureus and Candida albicans; downregulating cytokine and microbial protein expression. Scientific Reports 2018, 8, 1732. [Google Scholar] [CrossRef]
- Tripathi, S.; et al. Ameliorative effects of apple cider vinegar on neurological complications via regulation of oxidative stress markers. Journal of Food Biochemistry 2020, 44, e13504. [Google Scholar] [CrossRef]
- Shams, F.; et al. Effect of apple vinegar on folliculogenesis and ovarian kisspeptin in a high-fat diet-induced nonalcoholic fatty liver disease in rat. BMC Endocr Disord 2022, 22, 330. [Google Scholar] [CrossRef]
- Hadi, A.; et al. The effect of apple cider vinegar on lipid profiles and glycemic parameters: a systematic review and meta-analysis of randomized clinical trials. BMC Complement Med Ther 2021, 21, 179. [Google Scholar] [CrossRef]
- Bakir, S.; et al. Fruit Antioxidants during Vinegar Processing: Changes in Content and in Vitro Bio-Accessibility. Int J Mol Sci 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Diez-Ozaeta, I.; Astiazaran, O.J. Recent advances in Kombucha tea: Microbial consortium, chemical parameters, health implications and biocellulose production. Int J Food Microbiol 2022, 377, 109783. [Google Scholar] [CrossRef] [PubMed]
- Ojo, A.O.; de Smidt, O. Microbial Composition, Bioactive Compounds, Potential Benefits and Risks Associated with Kombucha: A Concise Review. Fermentation 2023, 9, 472. [Google Scholar] [CrossRef]
- Valduga, A.T.; et al. Chemistry, pharmacology and new trends in traditional functional and medicinal beverages. Food Research International 2019, 120, 478–503. [Google Scholar] [CrossRef]
- Villarreal-Soto, S.A.; et al. Understanding Kombucha Tea Fermentation: A Review. J Food Sci 2018, 83, 580–588. [Google Scholar] [CrossRef]
- Zubaidah, E.; et al. Comparison of in vivo antidiabetes activity of snake fruit Kombucha, black tea Kombucha and metformin. Biocatalysis and Agricultural Biotechnology 2019, 17, 465–469. [Google Scholar] [CrossRef]
- Watawana, M.I.; et al. Value-added Tea (Camellia sinesis) as a functional food using the Kombucha ‘Tea Fungus’. Warasan Khana Witthayasat Maha Witthayalai Chiang Mai 2018, 45, 136–146. [Google Scholar]
- Villarreal-Soto, S.A.; et al. Impact of fermentation conditions on the production of bioactive compounds with anticancer, anti-inflammatory and antioxidant properties in kombucha tea extracts. Process Biochemistry 2019, 83, 44–54. [Google Scholar] [CrossRef]
- Cardoso, R.R.; et al. Kombuchas from green and black teas have different phenolic profile, which impacts their antioxidant capacities, antibacterial and antiproliferative activities. Food Research International 2020, 128, 108782. [Google Scholar] [CrossRef]
- Vázquez-Cabral, B.D.; et al. Oak kombucha protects against oxidative stress and inflammatory processes. Chem Biol Interact 2017, 272, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.-z.; et al. Microbial bioconversion of the chemical components in dark tea. Food Chemistry 2020, 312, 126043. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.-J.; et al. State-of-the-art review of dark tea: From chemistry to health benefits. Trends in Food Science & Technology 2021, 109, 126–138. [Google Scholar] [CrossRef]
- Chen, Y.; et al. Tea consumption and risk of diabetes in the Chinese population: a multi-centre, cross-sectional study. Br J Nutr 2020, 123, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; et al. Theabrownin from Pu-erh tea attenuates hypercholesterolemia via modulation of gut microbiota and bile acid metabolism. Nature Communications 2019, 10, 4971. [Google Scholar] [CrossRef]
- Du, H.; et al. Fu Brick Tea Alleviates Chronic Kidney Disease of Rats with High Fat Diet Consumption through Attenuating Insulin Resistance in Skeletal Muscle. J Agric Food Chem 2019, 67, 2839–2847. [Google Scholar] [CrossRef]
- Zhang, X.; et al. Consumption of post-fermented Jing-Wei Fuzhuan brick tea alleviates liver dysfunction and intestinal microbiota dysbiosis in high fructose diet-fed mice. RSC Adv 2019, 9, 17501–17513. [Google Scholar] [CrossRef]
- Xiao, Y.; et al. The biochemical characteristics of a novel fermented loose tea by Eurotium cristatum (MF800948) and its hypolipidemic activity in a zebrafish model. LWT 2020, 117, 108629. [Google Scholar] [CrossRef]
- Yu, Y.; et al. Pu-erh tea extract induces the degradation of FET family proteins involved in the pathogenesis of amyotrophic lateral sclerosis. Biomed Res Int 2014, 2014, 254680. [Google Scholar] [CrossRef]
- Lu, Y.; et al. New Acylglycosides Flavones from Fuzhuan Brick Tea and Simulation Analysis of Their Bioactive Effects. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Conceição, A.A.; et al. Nutraceutical Enrichment of Animal Feed by Filamentous Fungi Fermentation. Fermentation 2022, 8. [Google Scholar] [CrossRef]
- Yafetto, L.; et al. Valorization of agro-industrial wastes into animal feed through microbial fermentation: A review of the global and Ghanaian case. Heliyon 2023, 9, e14814. [Google Scholar] [CrossRef]
- Hao, P.F.; et al. Agriculture organic wastes fermentation CO2 enrichment in greenhouse and the fermentation residues improve growth, yield and fruit quality in tomato. JOURNAL OF CLEANER PRODUCTION 2020, 275. [Google Scholar] [CrossRef]
- Porras, R.C.S.; et al. Breaking New Ground: Exploring the Promising Role of Solid-State Fermentation in Harnessing Natural Biostimulants for Sustainable Agriculture. PROCESSES 2023, 11. [Google Scholar] [CrossRef]
- Bergsma, S.; et al. Lactic acid bacteria as bioprotective agents: A mini-review on biotechnological-and human health-based facets. AIMS ENVIRONMENTAL SCIENCE 2023, 10, 206–225. [Google Scholar] [CrossRef]
- Luo, J.W.; et al. In vitro fermentation of pineapple-whey protein fermentation product on human intestinal microbiota derived from fecal microbiota transplant donors. LWT-FOOD SCIENCE AND TECHNOLOGY 2024, 191. [Google Scholar] [CrossRef]
- Vilela, A. The Importance of Yeasts on Fermentation Quality and Human Health-Promoting Compounds. FERMENTATION-BASEL 2019, 5. [Google Scholar] [CrossRef]
Figure 1.
Literature research strategy.
Figure 2.
Submerged and solid-state fermentation advantages and microorganisms studied.
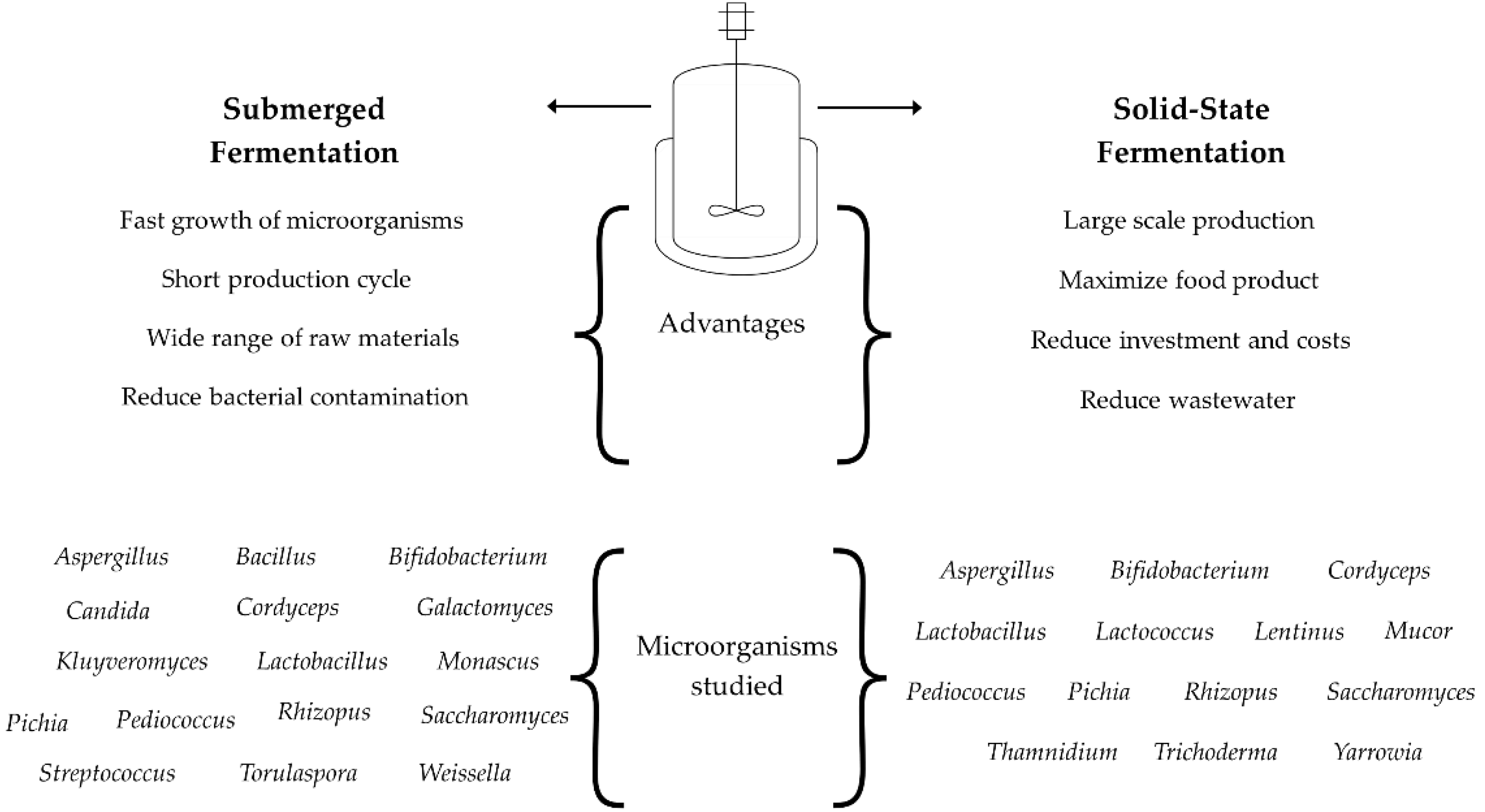
Figure 3.
Carotenoids obtained by the fermentation process of microorganisms
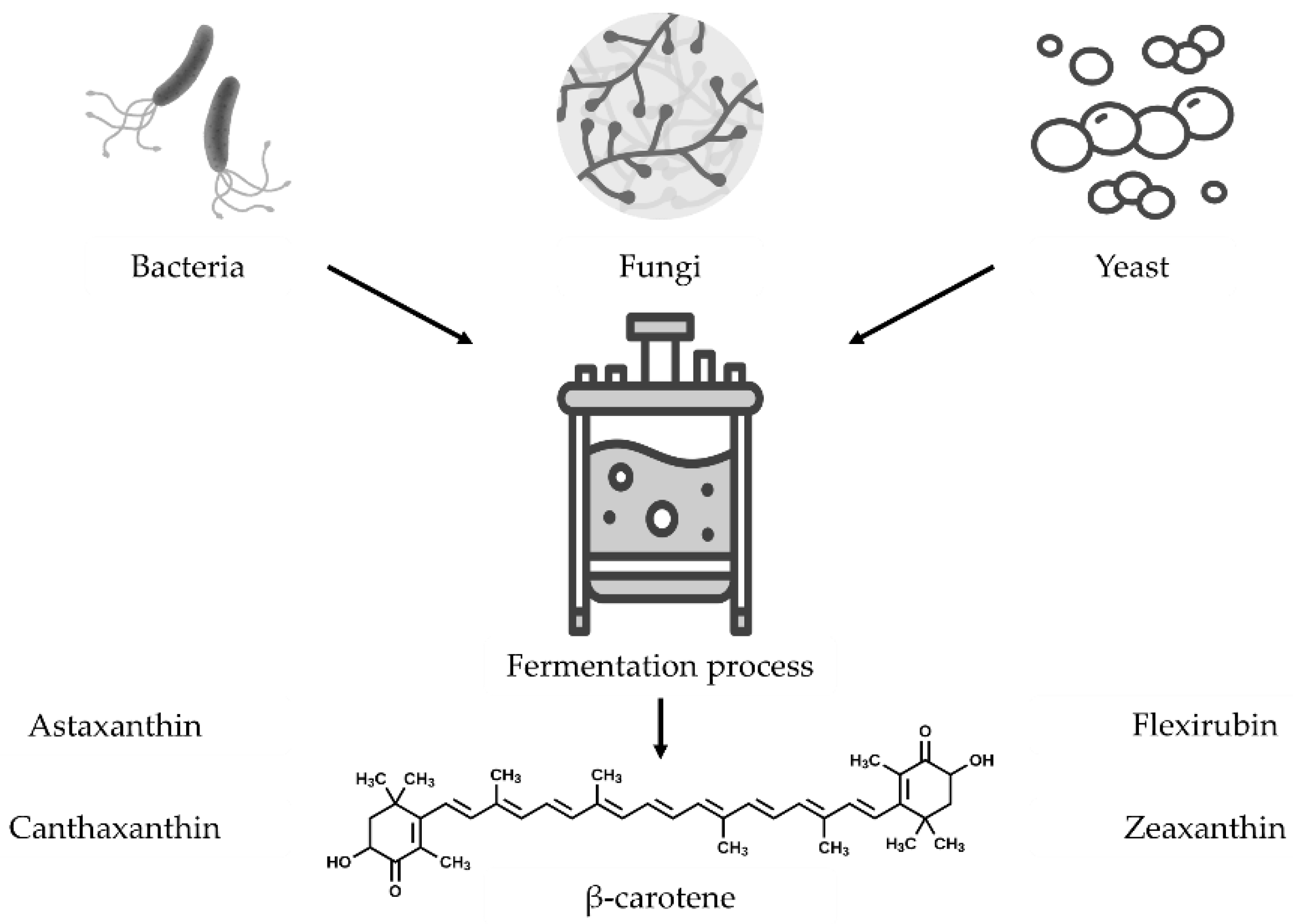
Figure 4.
Methods reported in the literature to produce essential oils by microbial fermentation.
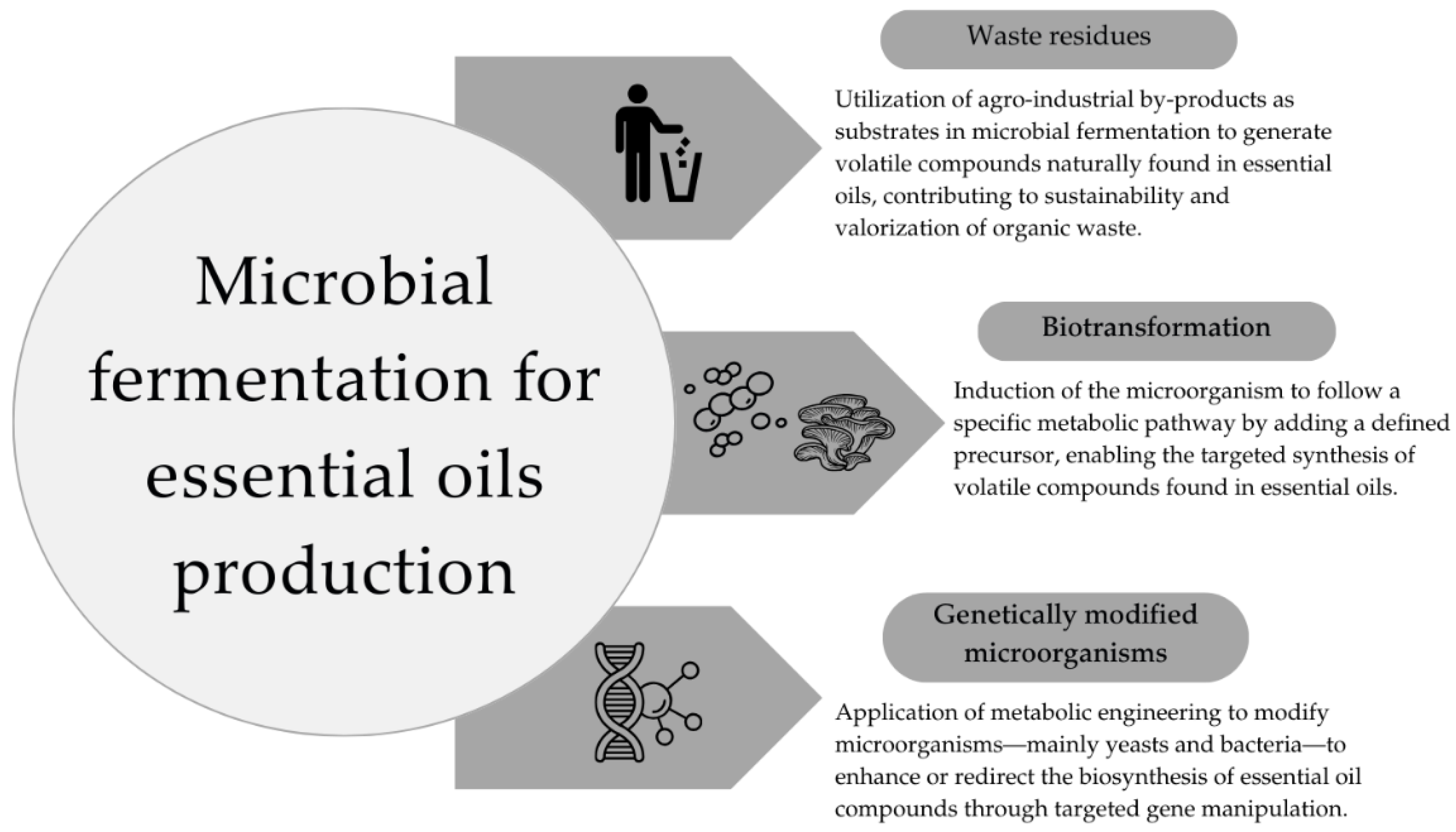
Figure 5.
Principal human health applications of bioactive compounds from microbial fermentation. Created by Biorender.com.
Figure 5.
Principal human health applications of bioactive compounds from microbial fermentation. Created by Biorender.com.
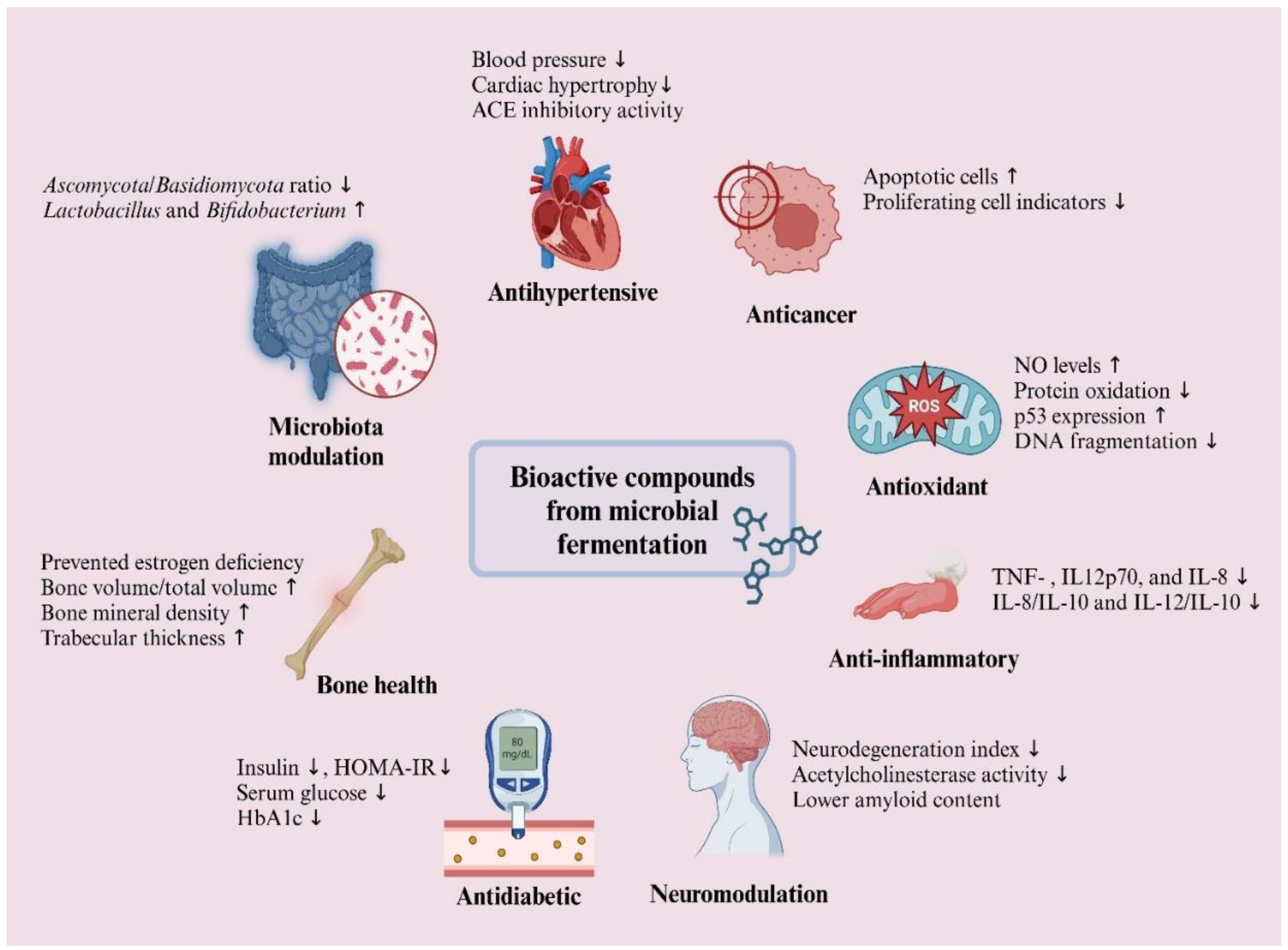
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



