
Submitted:
22 December 2025
Posted:
26 December 2025
You are already at the latest version
Abstract
Tropical Andean forests are biodiversity hotspots that have been transformed by anthropogenic activities, making ecosystem regeneration and restoration essential for their recovery. This study evaluated floristic composition, forest structure, and diversity in three land cover types within tropical Andean ecosystems: riparian forest (RF), natural regeneration (RN), and ecological restoration areas (RE). Vegetation was inventoried using standardized plots, recording species composition, diameter, and height. Basal area, size class distribution, and vertical structure were estimated. The Shan-non-Wiener and Simpson indices were evaluated. RF exhibited greater structural com-plexity, a larger basal area, and defined vertical stratification, indicating advanced suc-cessional stages and functional stability. NR showed the highest diversity values and a predominance of individuals in lower diameter and height classes, reflecting active re-cruitment and intermediate successional stages. Segment E exhibited lower diversity and intermediate structural development, consistent with shorter recovery periods and limitations in restoration design. Overall, the integration of floristic, structural, and diversity attributes indicates distinct successional trajectories, conditioned by land-use history, disturbance intensity, and environmental heterogeneity. These findings high-light the great potential for natural regeneration under reduced anthropogenic pressure and emphasize the need to integrate passive and active restoration strategies to enhance biodiversity and resilience in Andean tropical forests.
Keywords:
biodiversity recovery
; floristic composition
; forest structure
; successional gradient
; tropical andes
1. Introduction
Biological diversity is fundamental to the stability, functioning, and long-term persistence of terrestrial ecosystems. It underpins key ecological processes and provides ecosystem services essential for human well-being.[1]. In this sense, understanding the structure and composition of biological communities is crucial for designing effective conservation strategies and guiding ecosystem management, especially in regions where biodiversity loss is accelerating as a result of anthropogenic pressures.[2]. Floristic inventories and vegetation studies play a central role, as they provide baseline information on species composition, diversity patterns, and structural attributes of ecosystems, contributing to a better understanding of ecological processes and the definition of conservation priorities.[3].
The tropical Andean ecosystems of Colombia are recognized as one of the most important centers of biodiversity on the planet, characterized by high levels of species richness and endemism [4,5]. This extraordinary diversity is the result of a complex geological and evolutionary history, coupled with marked environmental heterogeneity generated by topography, climatic variability, and biogeographic connectivity [6]. However, these ecosystems have also been intensely transformed by human activities, as they form the basis of Colombia's agricultural, urban, and cultural development. Consequently, a significant proportion of Andean forests have been fragmented or replaced, leading to an accelerated reduction in forest cover and biodiversity, primarily due to deforestation, land-use change, and the expansion of the agricultural frontier. Landscape fragmentation is currently considered one of the main drivers of biodiversity loss in the Andes [7].
Given this scenario of high disturbance, the study of ecological succession becomes especially relevant for understanding ecosystem resilience, recovery trajectories, and long-term dynamics. Secondary succession involves progressive changes in species composition, structural complexity, and functional processes, which can eventually lead to conditions similar to those before the disturbance or to the formation of distinct ecological states [8]. Analyzing these successional trajectories is fundamental to assessing the potential of natural regeneration as a passive restoration mechanism, as well as the effectiveness of active ecological restoration strategies, particularly in tropical montane forests, where recovery processes can be slow and highly dependent on the environmental context and land use history[9].
Despite the ecological importance of Andean montane forests, comparative studies that comprehensively assess floristic composition, diversity, and vegetation structure along successional gradients are still limited [10,11], While riparian vegetation has been widely recognized as a key component influencing ecosystem integrity and hydrological functioning in tropical landscapes, it has often been addressed from functional or integrative assessment frameworks rather than through detailed analyzes of plant community composition and structure[12]. Consequently, riparian forests are still underrepresented in comparative studies that include conserved reference ecosystems, areas undergoing natural regeneration, and sites subject to active ecological restoration, despite their role as biodiversity reservoirs, ecological corridors, and valuable reference systems for restoration planning..
In this context, the present study contributes to the knowledge of floristic composition, diversity, and structural attributes of different vegetation cover types along a successional gradient in the Las Piedras River basin, located in Popayán, Cauca, in southwestern Colombia. This landscape is a heterogeneous mosaic, shaped by historical land use and current management practices, and includes preserved riparian forests (RF), areas of natural regeneration(NR), and sites undergoing active ecological restoration (ER) [13]. The objective of this study was to compare floristic diversity and vegetation structure among these cover types, as well as to evaluate their successional potential and ecological functionality. The results aim to contribute to the understanding of forest recovery processes in Andean montane ecosystems and provide input for the design of conservation and restoration strategies in fragmented tropical landscapes.
2. Materials and Methods
2.1. Área de Estudio
The study was conducted in the Las Piedras River basin, located in the intertropical equatorial Andes in southwestern Colombia, within the municipality of Popayan, Cauca (2°21′35″ N, 76°33′10″ W). The basin covers approximately 58 km² and spans an altitudinal range from 1980 to 3820 m a.s.l., encompassing sub-Andean and Andean forest formations (Figure 1).
The landscape is predominantly mountainous, with steep slopes in the upper basin (35–98%), moderate slopes in the middle zone (16–35%), and gentle slopes in the lower zone (3–15%). The climate corresponds to equatorial montane conditions, with mean annual temperatures ranging from 10.4 to 18.4 °C and a pronounced seasonality in precipitation, characterized by a wet period from October to May and a relatively dry season from June to September.
Soils are mainly Andisols derived from volcanic ash, with loamy to clay-loam textures, good drainage, weak structure, and strongly acidic conditions. These environmental characteristics, combined with pronounced topographic heterogeneity, promote high plant diversity and the development of a mosaic of forest covers along the successional gradient [14].
2.2. Vegetation Sampling
Three vegetation cover types representative of a successional gradient within the watershed were evaluated: riparian forest (RF), natural regeneration (NR), and ecological restoration (ER). In each vegetation cover, six linear transects measuring 50 × 4 m (200 m²) were established, resulting in a total sampled area of 0.12 ha per cover type. Transects were positioned to avoid spatial overlap and were separated by a minimum distance of 30 m to reduce spatial autocorrelation.
In each transect, all woody individuals with a diameter at breast height (DBH) ≥ 5 cm were recorded. For each individual, the species, circumference at breast height (CBH), stem height, and total height (m) were recorded, following standard methodologies for forest structure studies [15]. In areas dominated by grasslands, the proportion of individuals per unit area was estimated. Botanical specimens were collected, pressed, and processed using standard herbarium techniques. Voucher specimens were deposited in the Álvaro Fernández Pérez Herbarium (AFP) of the Fundación Universitaria de Popayán.
2.3. Taxonomic Identification
The taxonomic identification of the species was carried out using specialized keys, regional botanical literature, and digital resources, including keys for Rubiaceae and Melastomataceae of Colombia [16,17], as well as online databases such as World Flora Online (WFO), Useful Plants of Colombia (ColPlantA), Tropicos, Plants of the World Online (POWO), additionally, direct comparison was made with specimens deposited in the herbaria AFP (Fundación Universitaria de Popayán) and CAUP (Universidad del Cauca).
2.4. Structural and Diversity Analyses
Vegetation structural attributes were quantified based on the field data. Tree density was calculated as the number of individuals per hectare (individuals ha⁻¹), extrapolated from the sampled area. Basal area (m² ha⁻¹) was estimated as the sum of the cross-sectional area of all recorded stems at DBH [18].
Horizontal structure was analyzed by grouping individuals into diameter classes, while vertical structure was assessed using height classes. Class intervals for both diameter and height were defined using Sturges’ rule to facilitate comparison among vegetation covers. Distributions were expressed as absolute frequencies and as proportions (%) of the total number of individuals per cover type [19].
Alpha diversity was evaluated using the Shannon–Wiener diversity index (H′), which accounts for species richness and evenness, and the Simpson index (λ), which reflects dominance patterns. Lower Simpson values were interpreted as higher diversity [20].
2.5. Data Processing, Software, and Visualization
All statistical analyses and graphical representations were performed using R software version 4.5.1. Data manipulation and calculation of structural and diversity metrics were conducted using the packages dplyr and vegan [21], while figures were produced using ggplot2 [22].
Vegetation profiles illustrating vertical stratification patterns were generated using Python code in Blender version 5.0 [23], based on field-measured height and DBH data. These profiles represent schematic and idealized depictions of the dominant structural characteristics of each vegetation cover, rather than exact spatial reconstructions of individual transects.
Exploratory analyses and cross-validation of diversity indices were additionally supported using PAST software version 2.17 [24].
3. Results
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
3.1. Species Composition
The floristic composition recorded across the studied vegetation covers revealed a clear dominance of a limited number of plant families in the study area: Melastomataceae was the most representative family, with six genus and nine species, followed by Asteraceae (six genus and seven species) and Lauraceae (five genus and six species). Other families with a notable contribution included Clusiaceae (three genus and five species), Escalloniaceae (two genus and three species), as well as Phyllanthaceae and Solanaceae, each represented by two genus and two species.
Most of the remaining families showed low representation, typically with one or two genus and a single species per family, reflecting a common pattern in Andean montane plant communities, where floristic diversity is unevenly distributed among taxonomic groups (Figure 2).
The floristic composition varied markedly among the evaluated vegetation covers (Table 1), reflecting differences in successional stage and land-use history. RF were characterized by the presence of late-successional and forest-associated species such as Aniba perutilis, Cedrela odorata, Juglans neotropica, Quercus humboldtii, and Weinmannia tomentosa, indicating more structurally developed conditions. ER showed a mixed species composition, including taxa shared with riparian forests (e.g., Meriania speciosa, Myrcia popayanensis, Oreopanax incisus) and species typical of secondary or disturbed environments, suggesting intermediate successional stages. In contrast, NR were dominated by pioneer and early-successional species, particularly from Asteraceae and Poaceae, such as Ageratina theifolia, Baccharis sp., Chromolaena ivifolia, and Cenchrus purpureus. Several species occurred in more than one vegetation cover, indicating floristic overlap along the successional gradient.
The relative contribution of plant families varied notably among vegetation covers (Figure 3). RF were dominated by families characteristic of mature Andean forests, particularly Lauraceae, Viburnaceae, Phyllanthaceae, and Myrtaceae, whereas ecological restoration sites ER showed an intermediate pattern, with higher contributions of families such as Gesneriaceae and Piperaceae. In contrast, natural regeneration areas NR were dominated by early-successional and herbaceous families, especially Poaceae, Solanaceae, Asteraceae, and Ericaceae, highlighting clear floristic differentiation along the successional gradient.
3.2. Forest Structure of Land Uses
Forest structural attributes differed among the evaluated plant covers (Table 2). RF and ER showed comparable basal area values, with means of 15.75 ± 1.03 m² ha⁻¹ and 16.33 ± 1.29 m² ha⁻¹, respectively, whereas NR exhibited substantially lower basal area (6.08 ± 1.45 m² ha⁻¹). A similar pattern was observed for tree density, with higher values recorded in RF (1138 ± 6.49 trees ha⁻¹) and ER (1085 ± 5.57 trees ha⁻¹) compared to NR (873 ± 3.87 trees ha⁻¹). These results indicate clear structural differences among vegetation covers, consistent with their contrasting successional conditions.
The vegetation in RF shows a well-defined vertical stratification with a closed canopy reaching approximately 20 m, a developed subcanopy, and a dense understory, indicating a structurally mature forest (Figure 4a). ER exhibits intermediate structural complexity, with a heterogeneous canopy (up to ~15 m), mixed planted and naturally regenerated individuals, and an incipient but continuous understory (Figure 4b). In contrast, NR is characterized by a simpler structure, dominated by low- to medium-height vegetation (≤10 m), a discontinuous canopy, and greater light penetration, typical of early successional stages (Figure 4c). Taken together, these vegetation profiles reflect a clear gradient of increasing structural complexity from NR to ER and RF (Figure 4).
3.3. Diameter Distribution
The diameter class distribution highlights clear differences in the structural development of the studied vegetation covers. In all cases, individuals are predominantly concentrated in the lower diameter classes (DBH < 0.30 m), indicating active regeneration processes and a prevalence of young trees. However, the relative contribution of intermediate and larger diameter classes varies among plant covers, suggesting contrasting successional stages and structural complexity in RF, ER and NR (Table 3).
3.4. Altimetric Distribution
The height class distribution further illustrates differences in vertical structure among vegetation covers. In all covers, individuals were predominantly concentrated in the lower height classes (≤10 m), indicating the prevalence of juvenile and subcanopy strata. RF exhibited a higher proportion of individuals in intermediate and upper height classes, reflecting greater vertical stratification and structural development. ER showed an intermediate pattern, whereas NR were strongly dominated by low-stature individuals, consistent with early successional stages and simpler vertical structure(Table 4).
3.5. Ecological Indices
Alpha diversity differed among vegetation covers as reflected by the Shannon and Simpson indices (Table 5). NR exhibited the highest diversity values (Shannon = 2.89; Simpson = 0.93), followed by RF (Shannon = 2.45; Simpson = 0.89), whereas ER showed comparatively lower diversity (Shannon = 2.03; Simpson = 0.78). These patterns indicate differences in species richness and evenness among covers, providing a quantitative basis for comparing floristic diversity along the successional gradient.
4. Discussion
4.1. Floristic Composition and Forest Structure
The floristic composition of the different vegetation cover types (Table 1; Figure 3) reflects a clear successional gradient associated with the levels of intervention and conservation status. RF showed a greater representation of woody species and families typical of mature Andean montane forests, generally associated with shade-tolerant, slow-growing, and longer-lived species, suggesting floristically more consolidated and functionally stable communities. These characteristics indicate relatively stable environmental conditions and well-established ecological processes, such as microclimatic regulation and niche differentiation [25,26]. In contrast, the NR areas showed high species richness dominated by pioneer and early successional species, as well as by families with ecological strategies geared toward rapid establishment and efficient dispersal, indicating a highly dynamic system in an active phase of colonization and floristic replacement, typical of young secondary forests [27]. Meanwhile, the ER areas exhibited lower species richness and a more homogeneous composition, influenced by the selection of species used in the restoration processes and by the time elapsed since the intervention, resulting in an intermediate floristic and family composition within the successional gradient [28]. Taken together, these patterns suggest that the observed floristic, structural, and functional differences are not random but rather reflect the successional stage of each land cover and the management strategy applied, leading to contrasting recovery trajectories that condition structural complexity, biological diversity, and ecosystem functioning across the landscape.
The structural comparison between the land cover types (Table 2) showed that the RF exhibited the greatest structural complexity, reflected in high basal area values, high density, and a more balanced distribution of individuals across diameter and height classes, indicating an advanced successional stage and greater ecosystem stability. In contrast, the ER showed an intermediate structural condition, dominated by young, growing individuals, characteristic of the initial stages of active restoration, while the natural regeneration (NR) presented a smaller basal area and a high concentration of individuals in lower classes, suggesting a young, expanding forest. This pattern aligns with studies demonstrating that, during tropical secondary succession, basal area progressively increases as a result of diameter growth and biomass accumulation, gradually approaching the values observed in mature forests, in parallel with an increase in the structural and functional complexity of the ecosystem [29]. However, basal area recovery does not occur uniformly, as land-use history can modify the rate and trajectory of succession, leading to contrasting structural patterns even in forests of similar age. In this sense, the intensity and duration of previous land uses influence the convergence toward structures characteristic of mature forests and have direct implications for planning restoration strategies [30].
Taken together, the integration of floristic composition patterns (Table 1; Figure 3) and forest structure (Table 2) confirms that the evaluated land covers are organized along a successional gradient characteristic of Andean tropical forests, where environmental heterogeneity, land-use history, and disturbance dynamics decisively condition the configuration of plant communities. The greater floristic and structural complexity observed in the RF is consistent with communities nearing maturity, typical of Andean montane forests with high microclimatic stability and consolidated ecological processes, while the NR and ER exhibit compositional and structural arrangements characteristic of early and intermediate successional stages, respectively. These differences reflect the close interdependence between structure, composition, and functioning in the tropical Andes, where long-term successional processes define contrasting trajectories of forest development and determine ecosystem complexity, stability, and resilience at the landscape scale [30,31]
4.2. Diversity, Ecological Succession and Implications for Restoration
The calculated ecological indices reveal marked contrasts in diversity and dominance patterns among the evaluated land cover types (Table 4). The NR (Normal Forest) showed the highest diversity values according to the Shannon (H′) and Simpson (λ) indices, indicating a highly heterogeneous community with low dominance, characteristic of secondary forests in intermediate successional stages, where the coexistence of pioneer, early secondary, and shade-tolerant species increases the structural and functional complexity of the system [32,33]. Although the RF (Fairy Forest) recorded the greatest overall floristic richness (Table 1), its diversity values were intermediate, reflecting a greater dominance of species typical of advanced successional stages, associated with canopy closure and an intensification of competition for resources, without implying a reduction in ecosystem stability or functionality [34]. In contrast, the ER areas showed the lowest values of diversity and evenness, consistent with their shorter recovery time and the influence of the restoration design on the initial floristic composition; in these systems, the increase in diversity occurs progressively and depends strongly on landscape connectivity, the availability of propagules and the action of dispersers, factors that condition the convergence towards more diverse and functionally complex communities [35].Together, these patterns indicate that diversity and dominance dynamics are tightly linked to successional stage and management history, but in tropical Andean forests they are further shaped by strong environmental heterogeneity associated with steep elevational gradients, complex topography, and pronounced microclimatic variation, which promote divergent ecological trajectories across the landscape [36].
From an ecological restoration perspective in tropical Andean forests, the integration of the results for floristic composition (Table 1), forest structure (Table 2), diversity (Table 4), and vegetation profiles (Figure 4) demonstrates that each analyzed land cover represents a distinct successional trajectory, strongly influenced by land-use history, environmental heterogeneity, and the disturbance dynamics characteristic of these mountain ecosystems. Riparian forest (RF) is established as a reference ecosystem due to its high degree of structural and functional maturity, playing a key role in microclimatic regulation, biodiversity conservation, and the maintenance of ecological processes at the landscape scale. Natural regeneration (NR) highlights the high potential of passive restoration in the tropical Andes when anthropogenic pressures are reduced, favoring spontaneous processes of recruitment, diversification, and functional recovery. For their part, ecological restoration (ER) areas show that human intervention can accelerate the initial recovery of forests in degraded Andean contexts, provided it is based on adaptive management approaches that integrate species diversity, landscape connectivity, and ecological facilitation processes. Taken together, these results reinforce the need to combine passive and active strategies to restore biodiversity, ecosystem services, and the resilience of tropical mountain forests, priority ecosystems for conservation in the Andean region [37]. Furthermore, evidence from Andean forests indicates that natural regeneration after severe disturbances plays a key role in the progressive recovery of soil ecosystem services at different time scales, while traditional ecological knowledge can enhance these processes by guiding restoration practices more closely aligned with local dynamics, especially in Andean cloud forest ecosystems[38,39].
Taken together, the integration of multiple ecological dimensions in this study demonstrates that, in tropical Andean forests, diversity patterns, forest structure, and ecosystem functioning are determined by closely coupled successional processes operating at multiple spatial and temporal scales. The contrasts observed between land cover types reflect not only differences in disturbance history and management strategies, but also the environmental heterogeneity inherent in Andean mountain landscapes, highlighting the need for locally context-specific restoration approaches that are aligned with the unique ecological dynamics of these ecosystems.
5. Conclusions
The results of this study demonstrate that floristic composition, diversity, and forest structure in tropical Andean forests respond to a successional gradient clearly determined by land-use history, disturbance intensity, and the environmental heterogeneity characteristic of mountainous landscapes. The differences observed between RF, NR, and ER reflect contrasting successional trajectories that are expressed both in species dominance and in the structural and functional complexity of plant communities.
RF is consolidated as a reference ecosystem due to its greater structural and functional maturity, while NR demonstrates the high potential of passive processes for the recovery of diversity and ecosystem functioning when anthropogenic pressures are reduced. ER, for its part, shows that active interventions can accelerate the initial stages of recovery, although its convergence toward more complex states depends on the implementation of adaptive management approaches that integrate species diversity, landscape connectivity, and local ecological processes.
Overall, this study highlights that the effective restoration of tropical Andean forests requires differentiated and contextual strategies, based on an understanding of successional pathways and environmental heterogeneity. The combination of passive and active restoration emerges as a key strategy for strengthening biodiversity, ecosystem services, and the resilience of these mountain ecosystems, which are considered a priority for conservation in the face of global change.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/doi/s1, Figure S1: title; Table S1: title; Video S1: title.
Author Contributions
Conceptualization, VAMV and CACM; methodology, VAMV; software, VAMV and CACM; validation, LGC and DJMP; formal analysis, VAMV; investigation, VAMV and LGC and CACM; resources, LGC, CACM and DJMP; data curation, VAMV, LGC and DJMP; writing—original draft preparation, VAMV; writing—review and editing, VAMV.; visualization, CACM AND DJMP; supervision, DJMP; project administration, VAMV and DJMP; funding acquisition, VAMV, DJMP and LGC. All authors have read and agreed to the published version of the manuscript.reported.
Funding
This research was funded by the Eology program of the Fundación Universitaria de Popayán and the PhD in Environmental Sciences, University of Cauca. The APC was funded by the Doctorate in Environmental Sciences, Universidad del Cauca.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request. No publicly archived datasets were generated or analyzed during this study.
Acknowledgments
We express our gratitude to the Universidad del Cauca and the Fundación Universitaria de Popayán for their institutional support and the in-kind resources that made this research possible.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| TLA | Three letter acronym |
| LD | Linear dichroism |
References
- Hernández-blanco, M.; Costanza, R.; Chen, H.; Jarvis, D.; Montoya, J.; Sangha, K.; Stoeckl, N.; Turner, K.; Hoff, V. Van Ecosystem Health , Ecosystem Services , and the Well-Being of Humans and the Rest of Nature. 0–2. [CrossRef]
- Stevance, A.S.; Bridgewater, P.; Louafi, S.; King, N.; Beard, T.D.; Van Jaarsveld, A.S.; Ofir, Z.; Kohsaka, R.; Jenderedijan, K.; Rosales Benites, M.; et al. The 2019 Review of IPBES and Future Priorities: Reaching beyond Assessment to Enhance Policy Impact. Ecosyst. People 2020, 16, 70–77. [Google Scholar] [CrossRef]
- Kier, G.; Mutke, J.; Dinerstein, E.; Ricketts, T.H. Global Patterns of Plant Diversity and Floristic Knowledge. 2005, 1–10. [CrossRef]
- Rubio, S.P. Objetivos y Estrategias de La Cooperación Española Para La Conservación de La Agrobiodiversidad En La Región Andina Central. 2023. [Google Scholar]
- Comer, P.J.; Valdez, J.; Pereira, H.M.; Acosta-muñoz, C.; Campos, F.; Javier, F.; Garc, B.; Claros, X.; Castro, L.; Dallmeier, F.; et al. Conserving Ecosystem Diversity in the Tropical Andes. 2022. [Google Scholar] [CrossRef]
- Benavides, J.C.; Arenas-Bautista, M.C. The Andes: The Physical Setting. Sustain. Dev. Goals Ser. 2026, Part F1174, 3–17. [Google Scholar] [CrossRef]
- Morales, M.; Armenteras, D. Colombian Andean Cloud Forests Conservation Status, A Multiscalar Analysis. Boletín Científico. Cent. Museos. Mus. Hist. Nat. 2013, 17, 64–72. [Google Scholar]
- Marcelo, D.; Bank, W.; Rodr, N. Biodiversidad Y Actividad Humana : Relaciones En Ecosistemas De Bosque; 2007; ISBN 9789588151984. [Google Scholar]
- Por, P.; Alonso, J.; Delgado, M.; De Biología, P. Incidencia Antrópica En La Composición Florística, Estructural y Comportamiento de Rasgos Funcionales En La Comunidad de Manglar En Cartagena de Indias, Colombia. 2022. [Google Scholar]
- Cuesta, F.; Muriel, P.; Llambí, L.D.; Halloy, S.; Aguirre, N.; Beck, S.; Carilla, J.; Meneses, R.I.; Cuello, S.; Grau, A.; et al. Latitudinal and Altitudinal Patterns of Plant Community Diversity on Mountain Summits across the Tropical Andes. Ecography (Cop.). 2017, 40, 1381–1394. [Google Scholar] [CrossRef]
- Groups, F. Floristic Groups , and Changes in Diversity and Structure of Ecuador. 2021.
- Santos, A.R.; Barbosa, M.A.G.A.; Bolleli, T.; Anjinho, P.S.; Roque, R.; Mauad, F.F. Assessment of Water Ecosystem Integrity (WEI) in a Transitional Brazilian Cerrado–Atlantic Forest Interface. Water (Switzerland) 2023, 15. [Google Scholar] [CrossRef]
- Alfonso, V.; Valencia, M.; Casas, A.F.; Jesús, D.; Pinto, M. Modeling Soil Organic Carbon Dynamics Across Land Uses in Tropical Andean Ecosystems. 2025, 1–16.
- Mondragón Valencia, V.A.; Figueroa Casas, A.; Macias Pinto, D.J.; Rosas-Luis, R. Soil Organic Carbon Storage in Different Land Uses in Tropical Andean Ecosystems and the Socio-Ecological Environment. Earth 2025, 6, 106. [Google Scholar] [CrossRef]
- Freitas, J. Inventarios Forestales Nacionales de América Latina y El Caribe: Hacia La Armonización de La Información Forestal. 2021.
- Gentry, A.H. Diversity and Floristic Composition of Neotropical Dry Forests. Seas. Dry Trop. For. 1995, 146–194. [Google Scholar] [CrossRef]
- Mendoza-Cifuentes, H. Patrones De Riqueza Específica De Las Familias Melastomataceae Y Rubiaceae En La Cordillera Oriental, Colombia, Norte De Los Andes Y Consideraciones Para La Conservación. Colomb. For. 2012, 15, 5. [Google Scholar] [CrossRef]
- López Vargas, L.E.; Becoche Mosquera, J.M.; Macías Pinto, D.J.; Ruiz Montoya, K.; Velasco Reyes, A.; Pineda, S. Estructura Y Composición Florística De La Reserva Forestal - Institución Educativa Cajete, Popayán (Cauca). Luna Azul 2015, 131–151. [Google Scholar] [CrossRef]
- Rangel-Ch, J.O.; Velázquez, A. Metodos de Estudio de La Vegetación. Colomb. Divers. Biot. II 1997, 378. [Google Scholar]
- Magurran, A.E. Diversity Indices and Species Abundance Models. Ecol. Divers. Its Meas. 1988, 7–45. [Google Scholar] [CrossRef]
- Varsos, C.; Patkos, T.; Oulas, A.; Pavloudi, C.; Gougousis, A.; Ijaz, U.Z.; Filiopoulou, I.; Pattakos, N.; Berghe; Vanden, E.; Fernández-Guerra, A.; et al. Optimized R Functions for Analysis of Ecological Community Data Using the R Virtual Laboratory (RvLab). Biodivers. Data J. 2016, 4. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [CrossRef]
- Marín, J.; Baptiste, T.M.G.; Rodero, C.; Williams, S.E.; Niederer, S.A.; García-Fernández, I. SciBlend: Advanced Data Visualization Workflows within Blender. Comput. Graph. 2025, 130, 104264. [Google Scholar] [CrossRef]
- HAMMER, O. PAST : Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol Electron 2001, 4, 9. [Google Scholar]
- Guariguata, M.R.; Ostertag, R. Sucesión Secundaria. 2014.
- de Oliveira Loconte, C. The Volumetric Sustainability of Timber-Based Tropical Forest Management. Environ. Footprints Eco-Design Prod. Process. 2023, Part F1202, 51–87. [Google Scholar] [CrossRef]
- Chazdon, R.L. Tropical Forest Recovery: Legacies of Human Impact and Natural Disturbances. Perspect. Plant Ecol. Evol. Syst. 2003, 6, 51–71. [Google Scholar] [CrossRef]
- Martínez-Garza, C.; Howe, H.F. Restoring Tropical Diversity: Beating the Time Tax on Species Loss. J. Appl. Ecol. 2003, 40, 423–429. [Google Scholar] [CrossRef]
- Matsuo, T.; Martínez-Ramos, M.; Bongers, F.; van der Sande, M.T.; Poorter, L. Forest Structure Drives Changes in Light Heterogeneity during Tropical Secondary Forest Succession. J. Ecol. 2021, 109, 2871–2884. [Google Scholar] [CrossRef] [PubMed Central]
- Jakovac, C.C.; Junqueira, A.B.; Crouzeilles, R.; Peña-Claros, M.; Mesquita, R.C.G.; Bongers, F. The Role of Land-Use History in Driving Successional Pathways and Its Implications for the Restoration of Tropical Forests. Biol. Rev. 2021, 96, 1114–1134. [CrossRef]
- Finegan, B. Pattern and Process in Neotropical Secondary Rain Forests: The First 100 Years of Succession. Trends Ecol. Evol. 1996, 11, 119–124. [Google Scholar] [CrossRef]
- Dent, D.H.; Joseph Wright, S. The Future of Tropical Species in Secondary Forests: A Quantitative Review. Biol. Conserv. 2009, 142, 2833–2843. [Google Scholar] [CrossRef]
- Sanaphre-Villanueva, L.; Dupuy, J.M.; Andrade, J.L.; Reyes-García, C.; Paz, H.; Jackson, P.C. Functional Diversity of Small and Large Trees along Secondary Succession in a Tropical Dry Forest. Forests 2016, 7. [Google Scholar] [CrossRef]
- Larocque, G.R.; Shugart, H.H.; Xi, W.; Holm, J.A. Forest Succession Models. Ecol. For. Manag. Handb. 2024, 166–208. [Google Scholar] [CrossRef]
- Holl, K.D. Restoration of Tropical Forests. Restor. Ecol. New Front. 2012, 103–114. [Google Scholar] [CrossRef]
- Cavieres, L.A.; Llambí, L.D.; Anthelme, F.; Hofstede, R.; Arroyo, M.T.K. High-Andean Vegetation Under Environmental Change: A Continental Synthesis. Annu. Rev. Environ. Resour. 2025, 50, 219–245. [Google Scholar] [CrossRef]
- Crouzeilles, R.; Ferreira, M.S.; Chazdon, R.L.; Lindenmayer, D.B.; Sansevero, J.B.B.; Monteiro, L.; Iribarrem, A.; Latawiec, A.E.; Strassburg, B.B.N. Ecological Restoration Success Is Higher for Natural Regeneration than for Active Restoration in Tropical Forests. Sci. Adv. 2017, 3, 1–7. [Google Scholar] [CrossRef]
- Ortiz, J.; Panichini, M.; Neira, P.; Henríquez-Castillo, C.; Gallardo Jara, R.E.; Rodriguez, R.; Mutis, A.; Ramos, C.; Espejo, W.; Puc-Kauil, R.; et al. How Natural Regeneration After Severe Disturbance Affects Ecosystem Services Provision of Andean Forest Soils at Contrasting Timescales. Forests 2025, 16, 1–22. [Google Scholar] [CrossRef]
- Mariscal, A.; Tigabu, M.; Savadogo, P.; Odén, P.C. Regeneration Status and Role of Traditional Ecological Knowledge for Cloud Forest Ecosystem Restoration in Ecuador. Forests 2022, 13. [Google Scholar] [CrossRef]
Figure 1.
Location of the study area: Coordinates are expressed in UTM (meters), Datum WGS84, Zone 18N.
Figure 1.
Location of the study area: Coordinates are expressed in UTM (meters), Datum WGS84, Zone 18N.
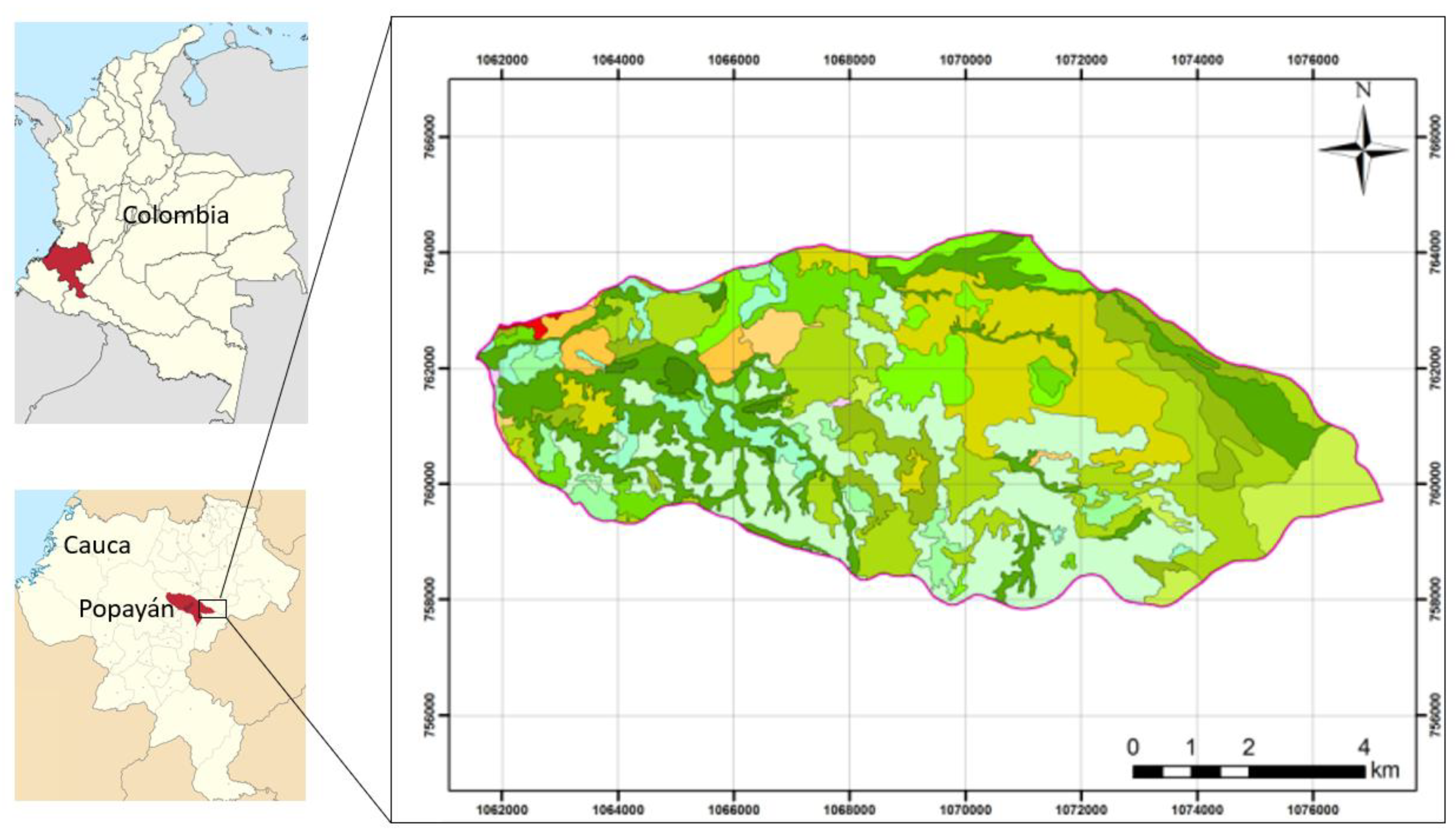
Figure 2.
Representative families by genus and species.
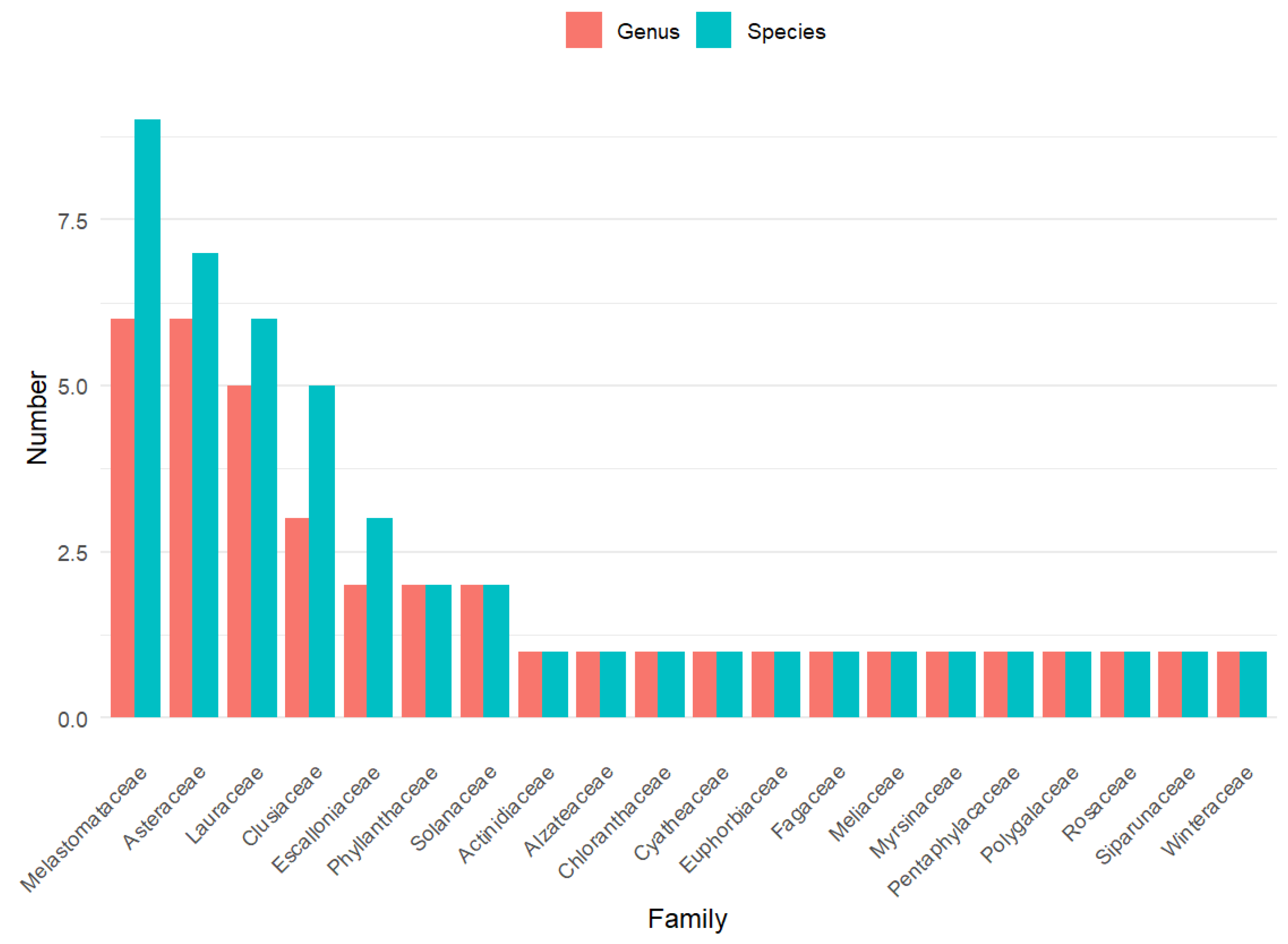
Figure 3.
Representative families by coverage (%)
Figure 4.
Idealized profiles of the studied vegetation covers: (a) RF: 1. Axinaea macrophylla; 2. Cecropia peltata; 3. Andesanthus lepidotus; 4. Quercus humboldtii; 5. Aniba perutilis; 6. Cedrela odorata; 7. Hieronyma macrocarpa; 8. Ocotea oblonga; 9. Miconia notabilis; 10. Palicourea angustifolia; 11. Chaetogastra mollis; 12. Piper crassinervium; 13. Cyathea sp.; 14. Siparuna echinata; 15. Myrcia popayanensis; 16. Hedyosmum cumbalense; 17. Cinchona pubescens; 18. Monnina salicifolia, (b) ER: 1. Alzatea verticillata; 2. Clusia multiflora; 3. Columnea cf. Sanguinea; 4. Hedyosmum cumbalense;5. Oreopanax incisus; 6. Aparisthmium cordatum; 7. Palicourea thyrsifolia; 8. Palicourea angustifolia; 9. Escallonia paniculata; 10. Meriania speciosa; 11. Morella pubescens; 12. Myrcia popayanensis; 13. Myrcianthes rhopaloides; 14. Myrsine coriacea; 15. Piper aduncum, (c) NR: 1. Chusquea sp.; 2. Cortaderia nitida; 3. Holcus lanatus; 4. Lolium multiflorum; 5. Monochaetum sp.; 6. Rhynchospora nervosa; 7. Vaccinium meridionale Sw.; 8. Ageratina theifolia; 9. Baccharis sp.; 10. Bejaria resinosa ; 11. Miconia notabilis; 12. Cinchona pubescens; 13. Chaetogastra grossa; 14. Chaetogastra mollis; 15. Chromolaena ivifolia; 16. Fuchsia caucana; 17. Galium hypocarpium; 18. Gynoxys sp..
Figure 4.
Idealized profiles of the studied vegetation covers: (a) RF: 1. Axinaea macrophylla; 2. Cecropia peltata; 3. Andesanthus lepidotus; 4. Quercus humboldtii; 5. Aniba perutilis; 6. Cedrela odorata; 7. Hieronyma macrocarpa; 8. Ocotea oblonga; 9. Miconia notabilis; 10. Palicourea angustifolia; 11. Chaetogastra mollis; 12. Piper crassinervium; 13. Cyathea sp.; 14. Siparuna echinata; 15. Myrcia popayanensis; 16. Hedyosmum cumbalense; 17. Cinchona pubescens; 18. Monnina salicifolia, (b) ER: 1. Alzatea verticillata; 2. Clusia multiflora; 3. Columnea cf. Sanguinea; 4. Hedyosmum cumbalense;5. Oreopanax incisus; 6. Aparisthmium cordatum; 7. Palicourea thyrsifolia; 8. Palicourea angustifolia; 9. Escallonia paniculata; 10. Meriania speciosa; 11. Morella pubescens; 12. Myrcia popayanensis; 13. Myrcianthes rhopaloides; 14. Myrsine coriacea; 15. Piper aduncum, (c) NR: 1. Chusquea sp.; 2. Cortaderia nitida; 3. Holcus lanatus; 4. Lolium multiflorum; 5. Monochaetum sp.; 6. Rhynchospora nervosa; 7. Vaccinium meridionale Sw.; 8. Ageratina theifolia; 9. Baccharis sp.; 10. Bejaria resinosa ; 11. Miconia notabilis; 12. Cinchona pubescens; 13. Chaetogastra grossa; 14. Chaetogastra mollis; 15. Chromolaena ivifolia; 16. Fuchsia caucana; 17. Galium hypocarpium; 18. Gynoxys sp..


Table 1.
General list of recorded species and their relationship with the studied land cover.
| Family | Species | RF | ER | NR |
| Actinidiaceae | Saurauia scabra (Kunth) D.Dietr. | X | ||
| Alstroemeriaceae | Bomarea patinii Baker | X | X | |
| Alzateaceae | Alzatea verticillata Ruiz & Pav | X | ||
| Araliaceae | Oreopanax incisus (Willd. ex Schult.) Decne. & Planch. | X | X | |
| Asteraceae | Ageratina theifolia (Benth.) R.M.King & H.Rob. | X | ||
| Baccharis sp. | X | |||
| Cirsium vulgare (Savi) Ten | X | |||
| Gynoxys sp. | X | |||
| Smallanthus pyramidalis (Triana) H.Rob | X | X | ||
| Chromolaena ivifolia (L.) R.King & H.Rob. | X | |||
| Chloranthaceae | Hedyosmum cumbalense H.Karst. | X | X | |
| Clusiaceae | Clusia alata Planch. & Triana | X | ||
| Clusia multiflora Kunth | X | |||
| Cunoniaceae | Weinmannia tomentosa L.f. | X | ||
| Cyatheaceae | Cyathea sp. | X | ||
| Cyperaceae | Rhynchospora nervosa (Vahl) Boeckeler | X | ||
| Ericaceae | Bejaria resinosa Mutis ex L.fil. | X | ||
| Vaccinium meridionale Sw. | X | X | ||
| Escalloniaceae | Escallonia myrtilloides L.fil. | X | ||
| Escallonia paniculata (Ruiz & Pav.) Roem. & Schult. | X | |||
| Euphorbiaceae | Aparisthmium cordatum (A.Juss.) Baill | X | ||
| Fabaceae | Inga densiflora Benth. | X | ||
| Fagaceae | Quercus humboldtii Bonpl. | X | ||
| Gesneriaceae | Columnea cf. sanguinea (Pers.) Hanst. | X | ||
| Kohleria warszewiczii (Regel) Hanst. | X | |||
| Juglandaceae | Juglans neotropica Diels | X | ||
| Lauraceae | Aniba perutilis Hemsl. | X | ||
| Nectandra mollis (Kunth) Nees | X | |||
| Ocotea oblonga (Meisn.) Mez | X | |||
| Melastomataceae | Andesanthus lepidotus (Humb. & Bonpl.) P.J.F.Guim. & Michelang. | X | ||
| Axinaea macrophylla Triana | X | |||
| Chaeotogastra grossa (L.fil.) P.J.F.Guim. & Michelang. | X | X | ||
| Chaetogastra mollis (Bonpl.) DC. | X | X | ||
| Meriania nobilis Triana | X | |||
| Meriania speciosa (Bonpl.) Naudin | X | |||
| Miconia notabilis Triana | X | X | ||
| Miconia theaezans (Bonpl.) Cogn. | X | X | ||
| Monochaetum sp. | X | |||
| Meliaceae | Cedrela odorata L.f. | X | ||
| Myricaceae | Morella pubescens (Humb. & Bonpl. ex Willd.) Wilbur | X | ||
| Myrsinaceae | Myrsine coriacea (Sw.) R.Br. ex Roem. & Schult. | X | X | |
| Myrtaceae | Myrcia popayanensis Hieron | X | X | |
| Myrcianthes fragans (Sw.) McVaugh | X | |||
| Myrcianthes rhopaloides (Kunth) Mc Vaugh | X | X | ||
| Onagraceae | Fuchsia caucana P.E.Berry | X | ||
| Pentaphylacaceae | Freziera reticulata Bonpl. | X | ||
| Phyllanthaceae | Hieronyma macrocarpa Müll. Arg. | X | ||
| Phyllanthus salviifolius Kunth | X | |||
| Pinaceae | Pinus radiata D.Don | X | ||
| Piperaceae | Piper aduncum L. | X | ||
| Piper crassinervium Kunth. | X | X | ||
| Poaceae | Cenchrus purpureus (Schumach.) Morrone | X | ||
| Chusquea sp. | X | |||
| Cortaderia nitida (Kunth) Pilg. | X | |||
| Holcus lanatus L. | X | |||
| Lolium multiflorum Lam. | X | |||
| Polygalaceae | Monnina salicifolia Ruiz & Pav. | X | ||
| Primulaceae | Stylogyne sp. | X | ||
| Rosaceae | Rubus bogotensis Kunth | X | ||
| Rubiaceae | Cinchona pubescens Vahl | X | X | |
| Coccocypselum lanceolatum (Ruiz & Pav.) Pers. | X | |||
| Elaeagia pastoensis L.E. Mora | X | |||
| Galium hypocarpium (L.) Endl. ex Griseb. | X | |||
| Ladenbergia oblongifolia (Humb. ex Mutis) L.Andersson | X | |||
| Palicourea thyrsifolia (Ruiz & Pav.) DC. | X | |||
| Palicourea angustifolia Kunth | X | X | ||
| Sapotaceae | Pouteria caimito (Ruiz & Pav.) Radlk. | X | ||
| Siparunaceae | Siparuna echinata (Kunth) A.DC | X | ||
| Solanaceae | Solanum cf. venosum Humb. & Bonpl. ex Dunal | X | ||
| Solanum sp. | X | |||
| Urticaceae | Cecropia peltata L. | X | ||
| Viburnaceae | Viburnum pichinchense Benth. | X | ||
| Viburnum triphylum Benth | X | |||
| Winteraceae | Drimys granadensis L.f. | X |
Table 2.
Forestal structure and to the plant covers.
| Plant cover | |||
| Variables | RF | ER | NR |
| Basal area/ha (m2 ) | 15,75 ± 0,03 | 16,33 ± 0,02 | 6.08 ± 0,04 |
| No. trees/ha (DAP>0,1m) | 1138 ± 0,04 | 1085 ± 0,05 | 873 ± 0,08 |
Values are mean, standard deviation (SD), n = 3 transects per land use. Tree density values (trees ha−1) were obtained by scaling counts from 300 m2 transects to a per-hectare basis. RF: riparian forest; ER: ecological restoration; NR: natural regeneration.
Table 3.
Diameter distributions of vegetation cover in the studied plots. RF, ER, NR.
| Plant cover | Diameter classes | DBH range (m) | Individuals Proportion (%) |
| RF | I | 0,1 – 0,199 | 42 |
| II | 0,2 – 0,299 | 33 | |
| III | 0,3 – 0,399 | 15 | |
| IV | 0,4 – 0,499 | 10 | |
| ER | I | 0,1 – 0,199 | 48 |
| II | 0,2 – 0,299 | 30 | |
| III | 0,3 – 0,399 | 15 | |
| IV | 0,4 – 0,499 | 7 | |
| NR | I | 0,1 – 0,199 | 65 |
| II | 0,2 – 0,299 | 25 | |
| III | 0,3 – 0,399 | 7 | |
| IV | 0,4 – 0,499 | 3 |
Table 4.
Altimetric distributions of vegetation cover in the studied plots. RF, ER, NR.
| Plant cover | Altimetric classes | Height range (m) | Individuals Proportion (%) |
| BR | I | 1 – 4,99 | 34 |
| II | 5 – 9,99 | 40 | |
| III | 10 – 14,99 | 18 | |
| IV | 15 – 19,99 | 8 | |
| RE | I | 1 – 4,99 | 42 |
| II | 5 – 9,99 | 37 | |
| III | 10 – 14,99 | 15 | |
| IV | 15 – 19,99 | 6 | |
| RN | I | 1 – 4,99 | 55 |
| II | 5 – 9,99 | 33 | |
| III | 10 – 14,99 | 9 | |
| IV | 15 – 19,99 | 3 |
Table 5.
Ecological indices by plant cover.
| Plant Cover | Shannon | Simpson |
| RF | 2.447506 | 0.8927708 |
| ER | 2.034597 | 0.7839089 |
| NR | 2.890507 | 0.9334027 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



