
Submitted:
18 January 2026
Posted:
21 January 2026
You are already at the latest version
Abstract
Background: Myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS) is character-ised by dysautonomia, unrefreshing sleep, and post-exertional malaise. Epidemiologicalstudies have identified an association between adverse childhood experiences (ACEs) andME/CFS risk, but the mechanistic links underlying this relationship remain unclear. Hypothesis: We propose that repeated corticotropin-releasing factor-driven stress arisingfrom ACEs induces a maladaptive 'high-tonic/low-phasic' firing mode in the locus co-eruleus. Over time, this leads to mitochondrial strain, adenosine triphosphate shortfall,and vesicular norepinephrine depletion—producing a 'wired-but-tired' state in which neu-rons fire continuously but release inadequate neurotransmitter. An acute trigger, oftenviral infection, then overwhelms this vulnerable system, resulting in persistent neuroin-flammation and impaired glymphatic clearance. Rationale: Four lines of evidence support this hypothesis. First, ACE-related autonomicand inflammatory signatures persist into adulthood and are well documented. Second,preliminary pharmacological observations suggest substrate limitation rather than recep-tor dysfunction. Third, LC neurons are known to be metabolically vulnerable to chronicactivation. Fourth, recent research has established that glymphatic function depends crit-ically on LC quiescence during sleep. Testable Predictions: The hypothesis generates five specific, falsifiable predictions. First,LC neuromelanin MRI signal should be reduced and correlate inversely with ACE scoresand ME/CFS severity. Second, norepinephrine transporter PET binding in the LC regionshould be reduced and correlate with hypervigilance symptoms. Third, cerebrospinal fluidlevels of 3-methoxy-4-hydroxyphenylglycol should be paradoxically low despite autonomicsymptoms suggesting high sympathetic activity. Fourth, heart rate variability and pupil-lometry measures should correlate with functional capacity. Fifth, pharmacological probestudies should differentiate substrate-limited from receptor-limited states. Limitations: Several important limitations must be acknowledged. No direct measure-ments of vesicular norepinephrine content in human LC neurons currently exist. TheACE-ME/CFS association derives from cross-sectional data, and prospective confirmationis lacking. The pharmacological observations that inform this hypothesis are anecdotal ra-ther than derived from controlled studies. Prospective validation is therefore essential.
Keywords:
myalgic encephalomyelitis
; chronic fatigue syndrome
; locus coeruleus
; norepinephrine depletion
; adverse childhood experiences
; dysautonomia
; hypothesis
1. Introduction
1.1. The ME/CFS Enigma
Myalgic encephalomyelitis/chronic fatigue syndrome represents one of the most challenging conditions in modern medicine. Despite affecting millions of people globally and causing profound disability, its pathophysiology remains incompletely understood. Recent research increasingly recognises ME/CFS as a systemic neuroimmunological disease rather than a purely psychological condition (Komaroff and Lipkin, 2021). Nevertheless, mechanistic models capable of explaining its core features remain elusive. These features include post-exertional malaise, unrefreshing sleep, cognitive impairment, orthostatic intolerance, and—perhaps most puzzling—the paradoxical coexistence of hyperarousal and exhaustion that patients often describe as feeling 'wired but tired.'
1.2. The ACE Connection
Epidemiological studies have identified an association between adverse childhood experiences and ME/CFS risk, although large-scale prospective studies are still lacking (Van Houdenhove et al., 2001). Clinical observation reveals that the relevant patterns extend beyond classical acute trauma to include chronic emotional neglect, excessive performance pressure, parentification, and conditional love based on achievement. Patients with these histories often exhibit self-critical perfectionism and high achievement orientation—patterns that are measurable via validated instruments such as the Childhood Trauma Questionnaire and consistent with long-term stress adaptation (Kempke et al., 2013).
The connection between ACEs and ME/CFS has typically been discussed in psychological or behavioural terms. This paper proposes instead a specific neurobiological mechanism: chronic programming of the locus coeruleus–norepinephrine system, leading to metabolic exhaustion that renders the system vulnerable to collapse following an acute trigger.
1.3. Preliminary Pharmacological Observations
Clinical reports describe unusual response patterns to stimulant medications in some ME/CFS patients. Randomised controlled trials have demonstrated modest, short-term improvements in fatigue and executive function in certain subgroups. Young (2013) reported significant improvement in executive functioning with lisdexamfetamine over eight weeks, while Blockmans et al. (2006) found that methylphenidate produced moderate symptom reduction in a subset of patients. Importantly, neither study documented rapid tolerance development.
Anecdotal clinical observations—not derived from controlled studies—suggest a different pattern in some patients: marked initial improvement followed by rapid tolerance requiring dose escalation. If confirmed prospectively, this sequence would be consistent with substrate limitation (specifically, vesicular depletion) rather than receptor dysfunction. This observation is presented as a hypothesis-generating clinical impression requiring controlled validation, not as established evidence.
Such a pattern would be mechanistically plausible given known pharmacology. Amphetamines increase cytosolic monoamine concentration and promote vesicular release through vesicular monoamine transporter interaction and transporter reversal (Eiden and Weihe, 2011). However, vesicle filling requires vacuolar-type ATPase-dependent proton gradient formation and adequate ATP availability, while the conversion of dopamine to norepinephrine via dopamine-β-hydroxylase is energy- and cofactor-dependent (Edwards, 2007). If vesicular stores were chronically depleted, forced release would produce pronounced but unsustainable effects—exactly the pattern some clinicians have observed.
1.4. Scope and Structure
This paper presents a conceptual framework linking adverse childhood experiences to ME/CFS through chronic locus coeruleus dysfunction. The structure proceeds as follows. Section 2 reviews the neurobiological foundations, including LC anatomy, the CRF-LC stress axis, the effects of ACEs on stress system programming, transgenerational epigenetic transmission, LC vulnerability to chronic stress, and the glymphatic connection. Section 3 proposes a specific multi-stage mechanism and differentiates it from existing two-hit models. Section 4 interprets clinical phenomena through the lens of LC depletion. Section 5 evaluates supporting evidence and alternative explanations while acknowledging limitations. Section 6 generates specific, testable predictions with proposed study designs. Section 7 and Section 8 discuss potential disconfirming evidence and broader implications.
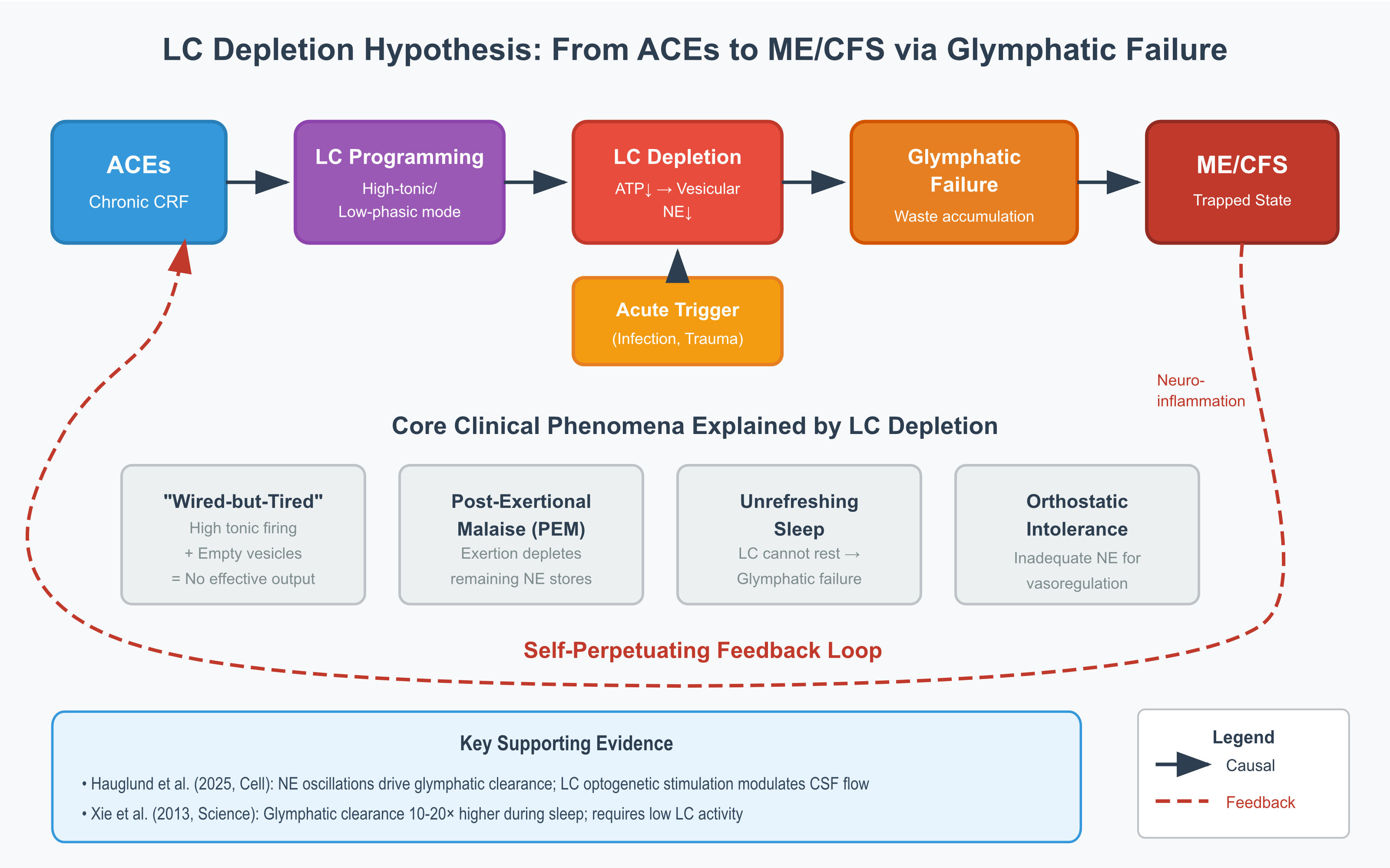
This is a hypothesis paper that synthesises existing data and proposes testable mechanisms. It makes no therapeutic claims. The proposed mechanistic pathway is illustrated in Figure 1.
2. Neurobiological Foundations
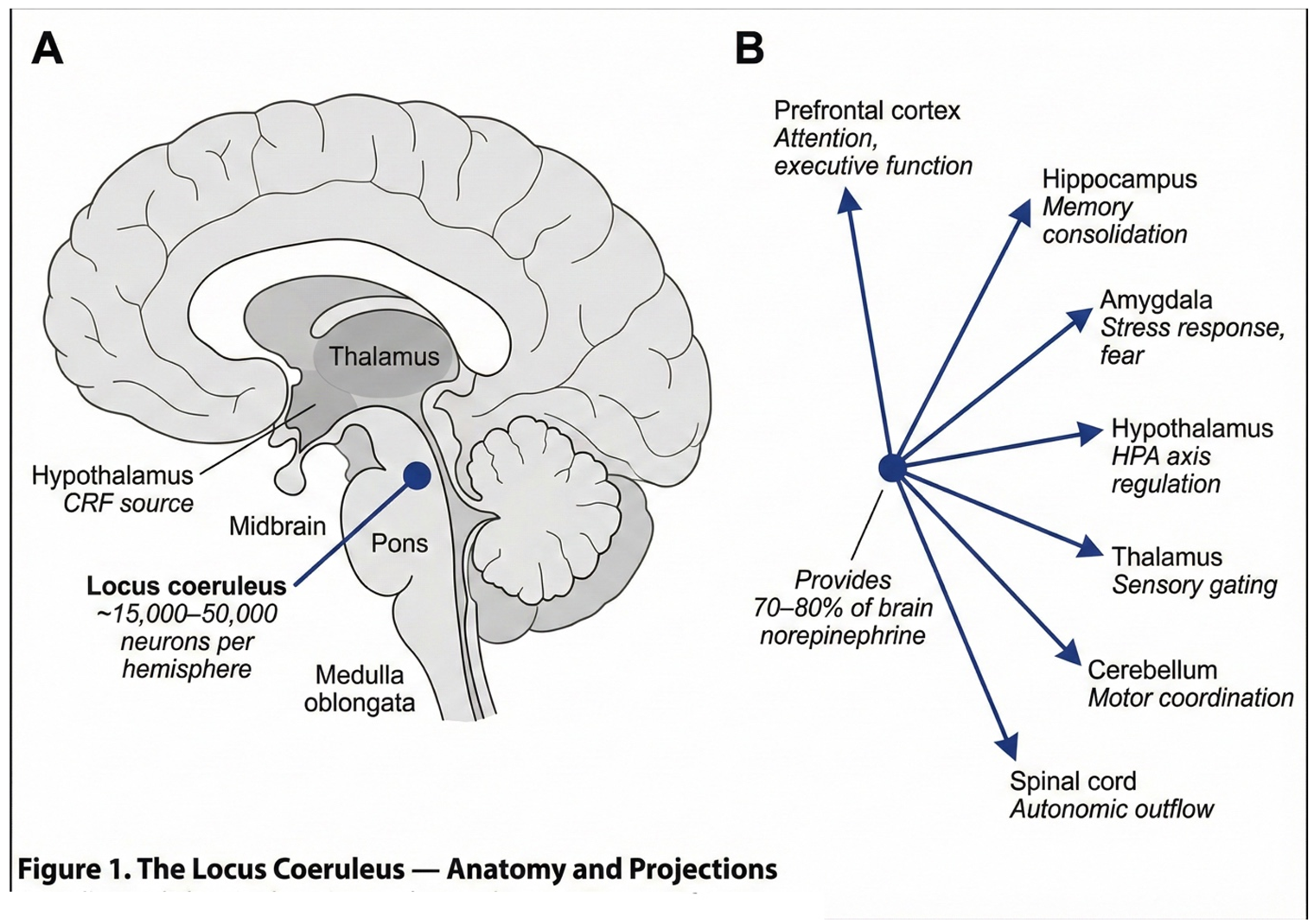
2.1. The Locus Coeruleus: Anatomy and Function
The locus coeruleus is a small, bilateral structure located in the dorsal pons, containing approximately 15,000 to 50,000 neurons per hemisphere—estimates vary across studies (Schwarz and Luo, 2015). Despite its modest size, this nucleus exerts remarkably broad influence over brain function. The LC supplies approximately 70 to 80 percent of the brain's norepinephrine through extensive projections that reach virtually all brain regions (Schwarz and Luo, 2015).
The functional roles of the LC are correspondingly diverse. As the primary brain target of corticotropin-releasing factor, it orchestrates the central stress response (Valentino and Van Bockstaele, 2008). Beyond stress regulation, LC projections modulate arousal and attention, facilitate memory consolidation, and regulate pain perception through descending pathways. The LC also influences cerebrovascular tone and contributes to peripheral immune modulation through its effects on sympathetic outflow.
The relationship between LC activity and cognitive performance follows an inverted-U function, as described by the Yerkes-Dodson relationship. Too-low activity produces drowsiness and inattention, optimal activity enables alert focus, and too-high activity causes hypervigilance and cognitive disorganisation (Aston-Jones and Cohen, 2005). This delicate balance is maintained through two complementary firing modes. Tonic activity consists of steady background firing at 2 to 5 Hz, establishing baseline arousal. Phasic activity involves burst firing exceeding 10 Hz in response to salient stimuli, enabling rapid attentional shifts and adaptive responding. As will become important later, the balance between these modes—and the capacity to shift flexibly between them—appears to be disrupted in ME/CFS.
2.2. The CRF-LC Stress Axis
The locus coeruleus does not operate in isolation; it functions as the central node of a stress-response network that links threat perception to physiological mobilisation. When the amygdala detects potential danger, it triggers the release of corticotropin-releasing factor from the central amygdala and the bed nucleus of the stria terminalis. This CRF acts directly on LC neurons, binding to CRFR1 receptors and producing rapid increases in noradrenergic firing (Valentino and Van Bockstaele, 2008). The resulting surge in norepinephrine heightens arousal, sharpens attention, and prepares the organism for defensive action.
In parallel with this rapid noradrenergic response, the hypothalamic-pituitary-adrenal axis is engaged, producing cortisol release over a longer timescale. The two systems are complementary: the LC-NE system provides immediate mobilisation, while the HPA axis sustains the stress response and facilitates eventual recovery. Under acute, time-limited stress, this dual architecture is highly adaptive.
The problem arises when stress becomes chronic. The CRF-LC axis, designed for episodic activation, was not built to sustain continuous operation. As will be discussed in Section 2.5, prolonged CRF exposure produces cumulative changes in LC physiology that eventually compromise its function—setting the stage for the vulnerability that precedes ME/CFS onset.
2.3. ACEs: Programming the Stress System
The landmark Adverse Childhood Experiences Study, which included over 17,000 participants, revealed a dose-response relationship between childhood adversity and adult health outcomes (Felitti et al., 1998). Each additional ACE increases the risk of cardiovascular disease, autoimmune disorders, chronic pain syndromes, and neuroimmunological conditions. Crucially, these effects persist regardless of adult behaviour, suggesting that the relevant changes involve biological programming rather than simply learned behavioural patterns.
The neurobiological consequences of ACEs have been extensively documented. HPA axis function is characteristically altered, with flattened diurnal cortisol rhythms replacing the normal morning peak and evening nadir (Bunea et al., 2017). Chronic low-grade inflammation becomes evident in elevated circulating levels of C-reactive protein, interleukin-6, and tumour necrosis factor-α (Baumeister et al., 2016). Autonomic regulation is compromised, manifest as reduced heart rate variability. At the molecular level, epigenetic modifications accumulate, including methylation of the NR3C1 glucocorticoid receptor gene—a change that perpetuates stress sensitivity by reducing receptor expression (McGowan et al., 2009; Perroud et al., 2011). Brain structural changes, including reduced hippocampal volume and heightened amygdala reactivity, complete the picture of a nervous system calibrated for threat.
Several important caveats must be acknowledged. The ACE-ME/CFS association derives from cross-sectional data, and prospective confirmation is lacking. Confounders including socioeconomic status, comorbidities, and retrospective reporting bias have not been fully controlled. The model proposed here applies to a hypothesised subgroup with ACE histories; given the heterogeneity of ME/CFS, other pathways almost certainly exist.
2.4. Transgenerational Epigenetic Transmission: The 'ACE-Negative' Paradox
A significant proportion of ME/CFS patients—estimated at 25 to 35 percent in clinical samples—report no significant adverse childhood experiences on standardised instruments such as the Childhood Trauma Questionnaire (Kempke et al., 2013). This 'ACE-negative' subgroup presents a conceptual challenge: if early-life stress programmes the LC-NE system toward vulnerability, how do patients without reported ACEs develop ME/CFS?
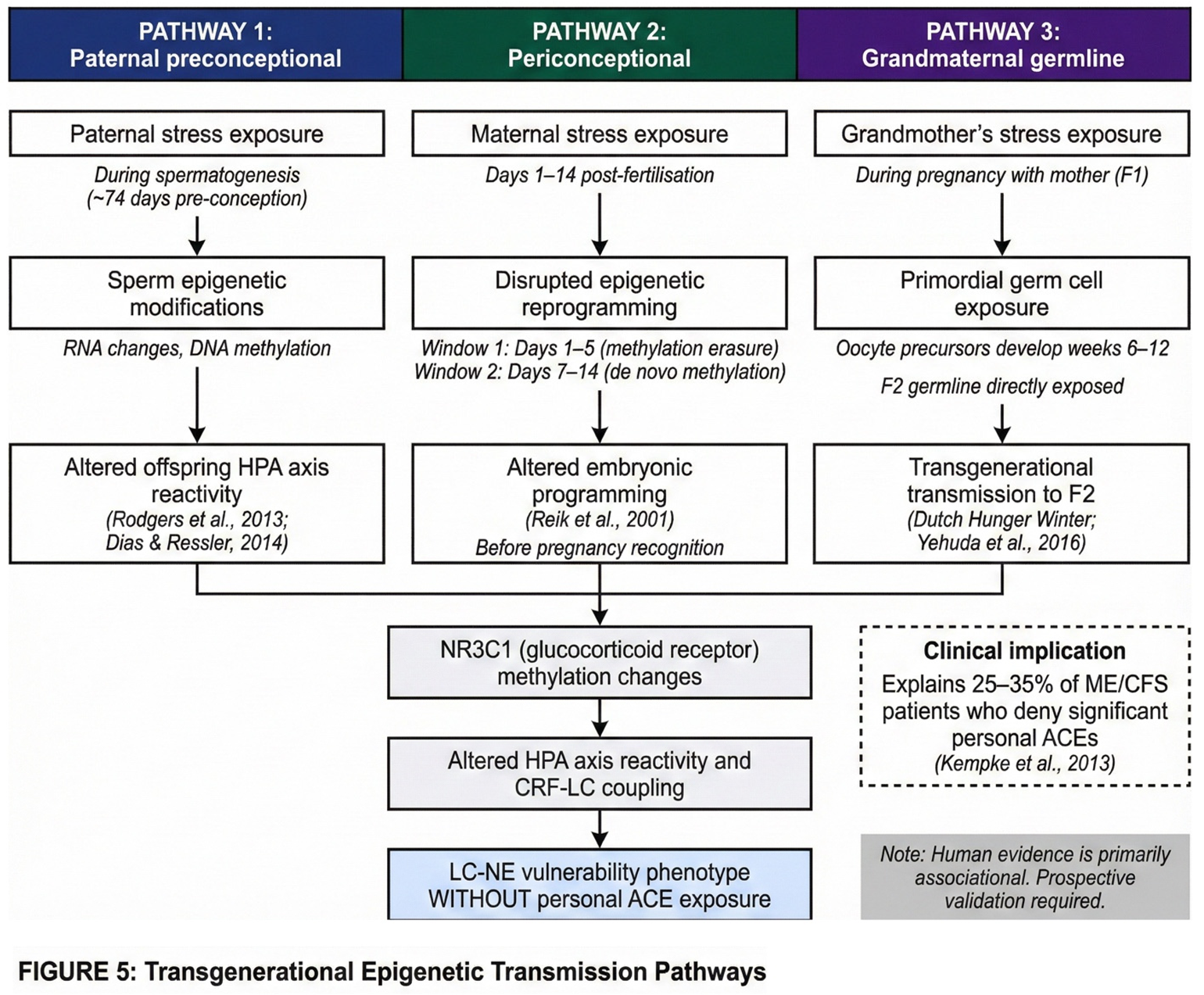
Transgenerational epigenetic transmission offers a mechanistically plausible explanation. Three distinct pathways merit consideration.
The first pathway involves paternal preconceptional stress. Human spermatogenesis requires approximately 74 days (Amann, 2008), and during this period developing spermatocytes are susceptible to epigenetic modification. Animal studies demonstrate that paternal stress during the preconceptional period alters offspring HPA axis reactivity and anxiety-related behaviours through changes in sperm RNA and DNA methylation (Rodgers et al., 2013; Dias and Ressler, 2014). While human evidence remains limited, the biological plausibility is established: chronic paternal stress in the 10 weeks preceding conception could programme offspring stress reactivity without the offspring experiencing any direct adversity.
The second pathway involves periconceptional epigenetic reprogramming. Two critical windows of epigenetic vulnerability occur in the earliest stages of development. The first window, spanning days 1 to 5 post-fertilisation, involves near-complete erasure of parental methylation marks during preimplantation development. The second window, approximately days 7 to 14, involves de novo methylation establishment following implantation (Reik et al., 2001). Maternal stress during these periods—typically occurring before pregnancy recognition—can influence epigenetic reprogramming through altered oviductal and uterine environments (Burkusis et al., 2015). Importantly, these effects occur before any conscious maternal awareness of pregnancy, rendering them invisible to retrospective recall.
The third pathway involves grandmaternal germline exposure. The primordial germ cells that will become a woman's oocytes develop during gestational weeks 6 to 12 of her own foetal development (Sasaki and Matsui, 2008). Consequently, stress experienced by the grandmother during this critical window directly exposes both the developing mother (F1) and her germline precursors (future F2). The Dutch Hunger Winter studies provide compelling epidemiological evidence: grandchildren of women pregnant during the 1944-45 famine exhibited increased adiposity and metabolic dysfunction, even when their mothers showed no such effects (Painter et al., 2008; Lumey et al., 2011). Rachel Yehuda's Holocaust research similarly demonstrated transgenerational transmission of PTSD-related epigenetic signatures at the FKBP5 locus across three generations (Yehuda et al., 2016; Yehuda and Lehrner, 2018).
These transgenerational mechanisms suggest that LC-NE system programming may occur through ancestral stress exposure rather than personal childhood adversity. The glucocorticoid receptor gene methylation documented in ACE-exposed individuals (McGowan et al., 2009) may be transmitted transgenerationally, as demonstrated in Holocaust offspring (Yehuda et al., 2016). This would produce the same downstream effect—altered HPA axis reactivity and CRF-LC coupling—without any personally recalled trauma. These three transgenerational transmission pathways are illustrated in Figure 5.
A critical caveat is necessary here. Transgenerational epigenetic inheritance in humans remains an active and contested area of investigation. While animal evidence is robust, human studies face substantial methodological challenges including confounding by shared postnatal environments, retrospective ascertainment, and the difficulty of distinguishing biological from psychosocial transmission. The evidence in humans is therefore primarily associational rather than causal. The proposed mechanisms are biologically plausible but require prospective validation. Nonetheless, they offer a coherent explanation for why approximately one-third of ME/CFS patients report no personal ACE history yet present with the same neurobiological phenotype as ACE-positive patients.
2.5. LC Vulnerability to Chronic Stress
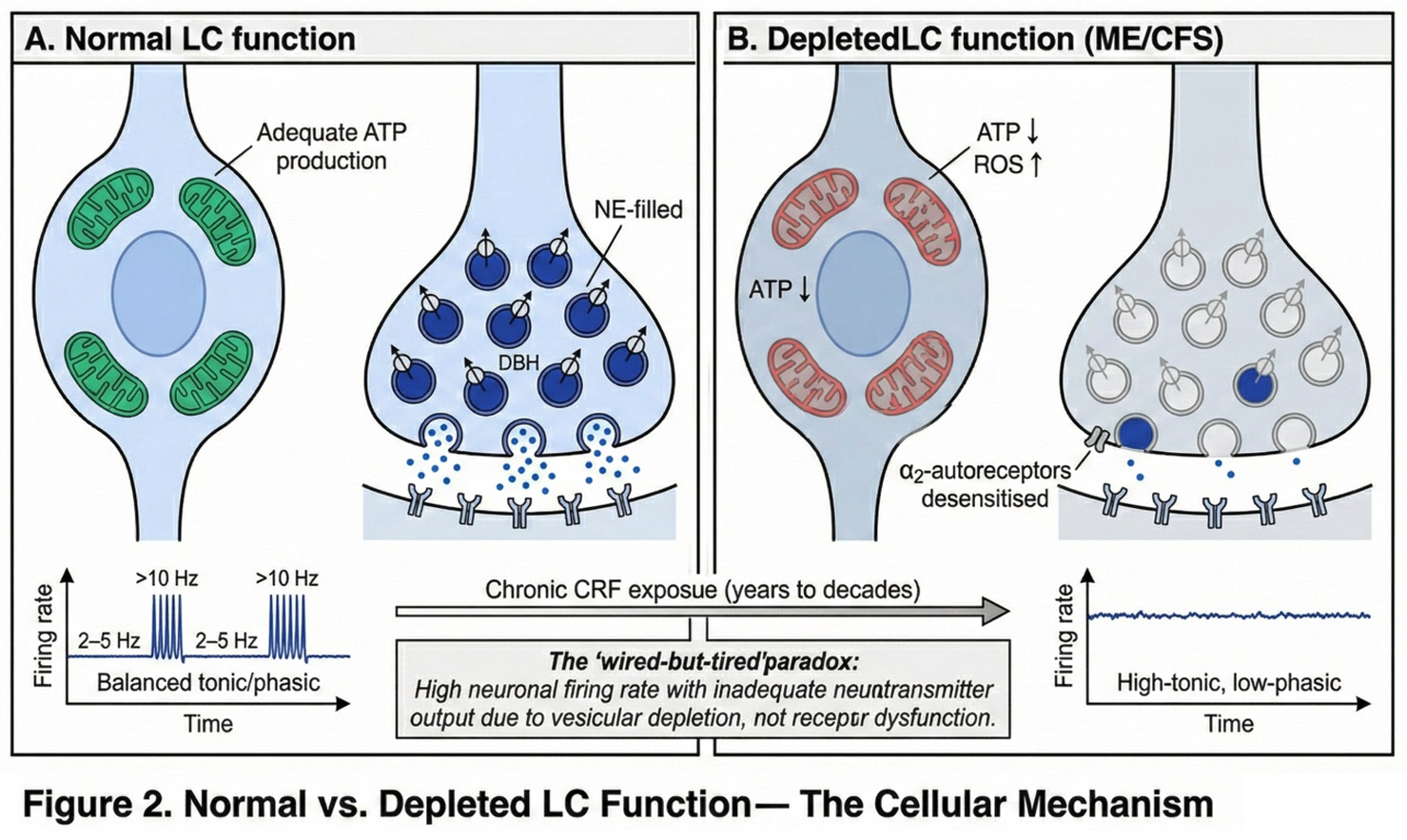
The locus coeruleus possesses several features that make it particularly vulnerable to chronic stress. Understanding these vulnerabilities is essential to the proposed mechanism.
The first vulnerability is metabolic fragility. LC neurons maintain high mitochondrial density, reflecting their substantial energy demands. Constant norepinephrine synthesis requires ATP, both for the conversion of dopamine to norepinephrine via dopamine-β-hydroxylase and for vesicular loading via V-ATPase-dependent proton gradients. Prolonged high firing rates generate oxidative stress as a byproduct of this intense metabolic activity.
The second vulnerability involves autoreceptor desensitisation. Under normal conditions, α2-adrenergic autoreceptors on LC neurons provide inhibitory feedback, dampening norepinephrine release when extracellular concentrations rise. However, with chronic CRF stimulation, these autoreceptors desensitise, resulting in loss of this negative feedback mechanism (Valentino and Van Bockstaele, 2008). The LC thus loses a key regulatory brake on its own activity.
The third vulnerability concerns firing pattern rigidity. Chronic stress shifts the LC from a responsive, flexible firing mode toward a rigid pattern characterised by elevated tonic activity with diminished phasic responses. This 'high-tonic/low-phasic' pattern has been documented in PTSD studies (Bangasser and Valentino, 2020) and manifests clinically as hypervigilance combined with impaired ability to shift attention appropriately.
Taken together, these vulnerabilities create a scenario in which chronic stress progressively degrades LC function. The neurons fire at increasingly high rates due to desensitised autoreceptors, generating oxidative stress and depleting ATP. As ATP becomes limiting, vesicular loading becomes inefficient, and the neurons—though firing rapidly—release progressively less norepinephrine per action potential. This is the cellular basis of the 'wired-but-tired' state. The contrast between normal and depleted LC function is illustrated in Figure 2.
2.6. The Glymphatic Connection
The glymphatic system serves as the brain's waste clearance mechanism (Nedergaard, 2013). During sleep, astrocytes shrink, expanding the extracellular space by approximately 60 percent. Cerebrospinal fluid then flows along perivascular spaces, washing out metabolic waste products. This clearance is substantially enhanced during sleep compared to wakefulness (Xie et al., 2013).
The critical point for the present hypothesis is that effective glymphatic function requires reduced LC activity. Recent research has demonstrated that norepinephrine oscillations during non-rapid eye movement sleep are among the strongest predictors of glymphatic clearance (Hauglund et al., 2025). Optogenetic stimulation of the LC directly modulates cerebrospinal fluid flow, while pharmacological suppression of NE oscillations may attenuate glymphatic clearance, though the magnitude of this effect requires further quantification.
These findings establish a direct causal link between LC activity patterns and brain waste clearance. With chronic LC overactivation, the system cannot achieve the rhythmic NE fluctuations necessary for effective clearance. Waste products accumulate, potentially causing further damage to LC neurons and perpetuating a vicious cycle. The clinical manifestation of this impaired clearance is 'unrefreshing sleep'—the experience of waking as exhausted as one went to bed, regardless of sleep duration.
3. The Proposed Mechanism
3.1. Multi-Stage Model
We propose that the transition from ACE exposure to ME/CFS proceeds through five distinguishable stages, though in practice these stages overlap and interact rather than occurring as discrete phases.
During childhood and adolescence, the LC-NE system adapts to chronic stress through what we term the adaptive response phase. Repeated ACE-related stress produces sustained CRF elevation, which programmes the LC into a state of heightened tonic activity. From the child's perspective, this adaptation is protective: in an unpredictable or threatening environment, constant vigilance confers survival advantage. The neural substrate of this vigilance—elevated LC firing—becomes the organism's baseline operating mode.
Over years to decades, however, this adaptive pattern becomes maladaptive. We refer to this as the chronic maladaptation phase. Sustained high-tonic firing imposes cumulative metabolic costs on LC neurons. The α2-adrenergic autoreceptors that normally provide inhibitory feedback become desensitised, eliminating a key regulatory brake (Valentino and Van Bockstaele, 2008). Mitochondria, operating continuously at high demand, generate increasing oxidative stress and struggle to maintain ATP production. Because vesicle refilling depends on ATP-driven proton gradients via V-ATPase, and because the conversion of dopamine to norepinephrine via dopamine-β-hydroxylase requires ATP and cofactors, vesicular stores become progressively depleted. The result is a paradoxical state—high neuronal firing with inadequate neurotransmitter output—that we term the wired-but-tired phenotype.
The third stage represents a vulnerable equilibrium. The individual continues to function—often at high levels of achievement—but the system operates with no reserve capacity. Vesicles are chronically partially depleted, mitochondria are stressed, glymphatic clearance is impaired, and subclinical neuroinflammation may be present. The person appears healthy and often performs well, driven by the same hypervigilance that depleted the system in the first place. But one more significant stressor will cause the system to collapse.
The fourth stage is the acute trigger. This is typically a viral infection such as Epstein-Barr virus, SARS-CoV-2, or influenza, though it may also be major surgery, physical trauma, severe psychological stress, or physiological transitions such as pregnancy and childbirth. The trigger produces massive LC activation at a time when the system cannot sustain it. Complete vesicular depletion follows. Post-infectious neuroinflammation persists because the pre-damaged system cannot restore homeostasis. The individual does not recover as expected.
The fifth and final stage is the trapped ME/CFS state. The system becomes locked in a pathological equilibrium characterised by high neuronal activity with low norepinephrine output, persistent neuroinflammation, continued glymphatic dysfunction, and progressive mitochondrial failure. Any attempt to increase demand—whether through physical exertion, cognitive effort, or emotional stress—depletes the remaining vesicular stores, producing post-exertional malaise. Orthostatic intolerance reflects the inability to mobilise adequate norepinephrine for cardiovascular regulation. Cognitive impairment reflects the loss of phasic LC responses needed for attentional shifting. Unrefreshing sleep reflects impaired glymphatic clearance. Positive feedback loops trap the system in this state: poor sleep impairs recovery, impaired recovery maintains inflammation, inflammation sustains LC activation, and sustained LC activation prevents the quiescence needed for restorative sleep. This multi-stage progression from vulnerability to collapse is summarised in Figure 3.
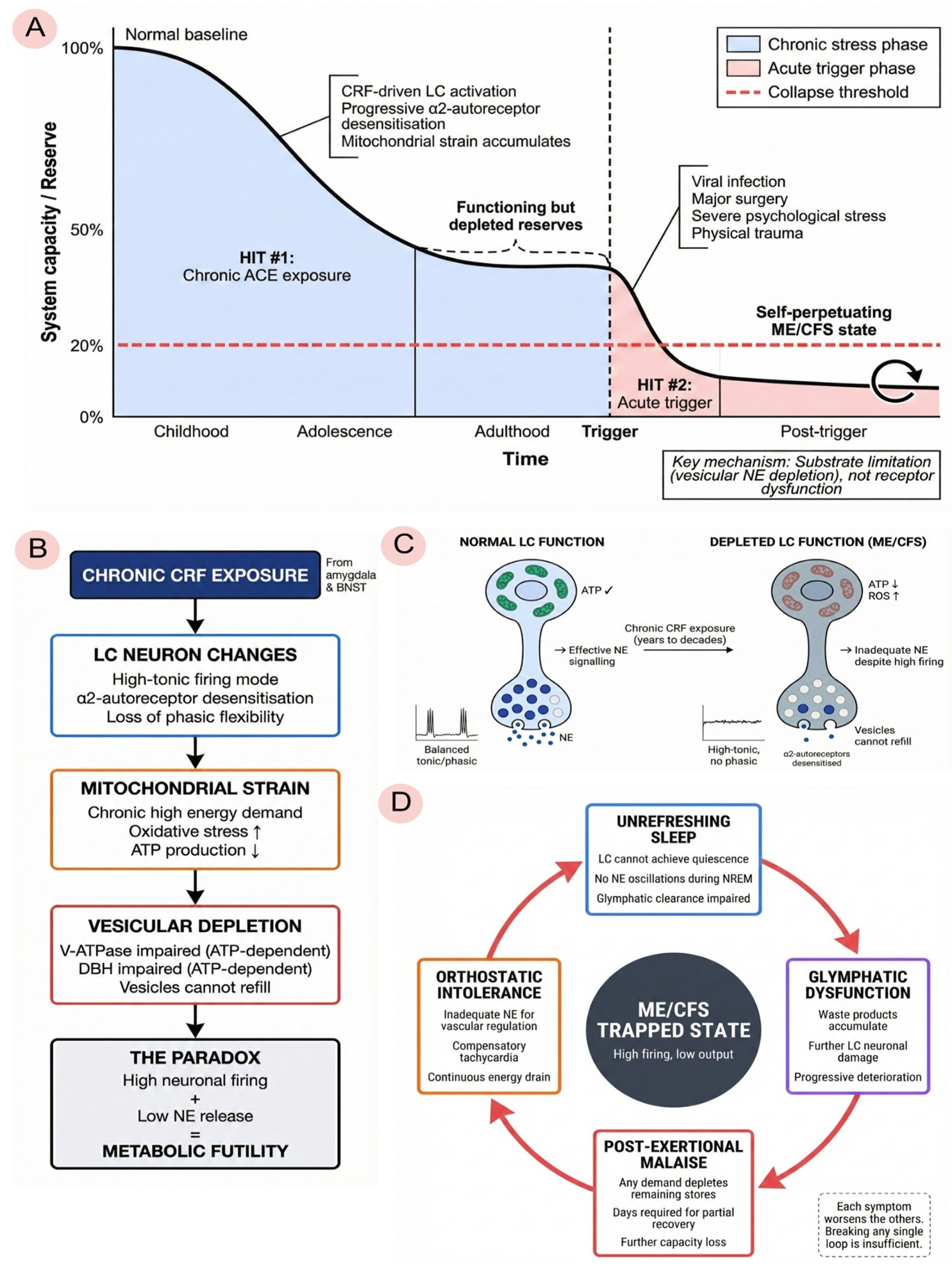
3.2. Differentiation from Existing Two-Hit Models
Two-hit models have been proposed previously in the ME/CFS literature (Komaroff and Lipkin, 2021). The present model differs from these in four important respects.
First, it specifies a precise neuroanatomical target. Rather than invoking a diffuse 'stress response' or generalised autonomic dysfunction, the model identifies the locus coeruleus as the critical vulnerable node. This specificity enables targeted investigation and potential intervention.
Second, it proposes a specific cellular mechanism. The core problem, according to this model, is vesicular norepinephrine depletion secondary to ATP limitation—not receptor dysfunction, not altered sensitivity, but inadequate neurotransmitter available for release despite ongoing neuronal activity. This distinction has important implications for treatment, as interventions targeting receptors would be predicted to fail while interventions supporting mitochondrial function or reducing metabolic demand might succeed.
Third, the model provides a mechanistic link to the sleep pathology that is central to ME/CFS. By connecting LC activity patterns to glymphatic clearance, it explains why sleep is unrefreshing even when sleep architecture appears relatively preserved on standard polysomnography. The problem is not sleep per se but the failure to achieve the LC quiescence necessary for effective waste clearance during sleep.
Fourth, the model generates testable predictions at multiple levels of analysis—cellular, circuit, and systems—enabling empirical evaluation rather than remaining purely theoretical.
4. Clinical Phenomena Through the LC Depletion Lens
4.1. The 'Wired-but-Tired' Paradox
Perhaps the most striking feature of ME/CFS is the coexistence of hyperarousal and exhaustion—what patients often describe as feeling 'wired but tired.' They report hypervigilance, anxiety, and difficulty disengaging from stimulation, yet simultaneously experience profound fatigue, inability to sustain even modest activity, and sleep that fails to restore. This paradox is difficult to explain under models that treat arousal and fatigue as opposite ends of a single continuum.
The LC depletion hypothesis offers a resolution. The 'wired' component reflects persistently elevated tonic LC firing, driven by desensitised α2-autoreceptors that no longer provide inhibitory feedback. The 'tired' component reflects the consequences of this firing: chronically depleted vesicles that cannot release adequate norepinephrine despite high neuronal activity. The system is running at high metabolic cost while producing minimal functional output—a state of metabolic futility that manifests as the subjective experience of exhausted hyperarousal.
4.2. Post-Exertional Malaise
Post-exertional malaise is widely considered the hallmark symptom of ME/CFS. Physical or cognitive exertion produces delayed symptom exacerbation—typically emerging 24 to 72 hours after the triggering activity—that may last days to weeks and is disproportionate to the exertion involved.
The LC depletion model provides a straightforward explanation for this phenomenon. Any exertion requires norepinephrine mobilisation to support attention, cardiovascular adjustment, and motor coordination. In the depleted state, vesicles release whatever content remains. With compromised mitochondria and depleted ATP, the system cannot rapidly resynthesise norepinephrine or refill vesicles. Days are required for even partial recovery—in contrast to healthy individuals, who restore neurotransmitter stores within hours.
The delay between exertion and symptom exacerbation reflects the time course of vesicular depletion and its downstream consequences. The severity and duration of the malaise reflects the degree of depletion and the impaired capacity for recovery.
4.3. Orthostatic Intolerance
Upon standing, many ME/CFS patients experience dizziness, tachycardia, cognitive impairment, and nausea. Postural orthostatic tachycardia syndrome is a common comorbidity, and some patients meet criteria for small fibre neuropathy.
The LC depletion model interprets orthostatic intolerance as a consequence of inadequate norepinephrine availability for cardiovascular regulation. Vasoregulation upon standing requires rapid norepinephrine release to prevent blood pooling in the lower extremities and maintain cerebral perfusion. With inadequate norepinephrine, this compensation fails. The heart rate increases as a secondary compensatory mechanism to maintain cardiac output, producing the characteristic tachycardia. But this compensation is imperfect, and cerebral perfusion often remains inadequate—hence the cognitive symptoms and dizziness.
This interpretation explains the apparent paradox of sympathetic overactivity (manifested as tachycardia) coexisting with reduced effective noradrenergic function. The neurons are firing, but they are not releasing adequate neurotransmitter. Comorbidities such as POTS and small fibre neuropathy may act as moderator variables that amplify this core dysfunction.
4.4. Unrefreshing Sleep
Patients with ME/CFS characteristically report that sleep provides minimal restoration. They often wake feeling as exhausted as when they went to bed, regardless of sleep duration. The relationship between LC activity and glymphatic clearance is illustrated in Figure 4.
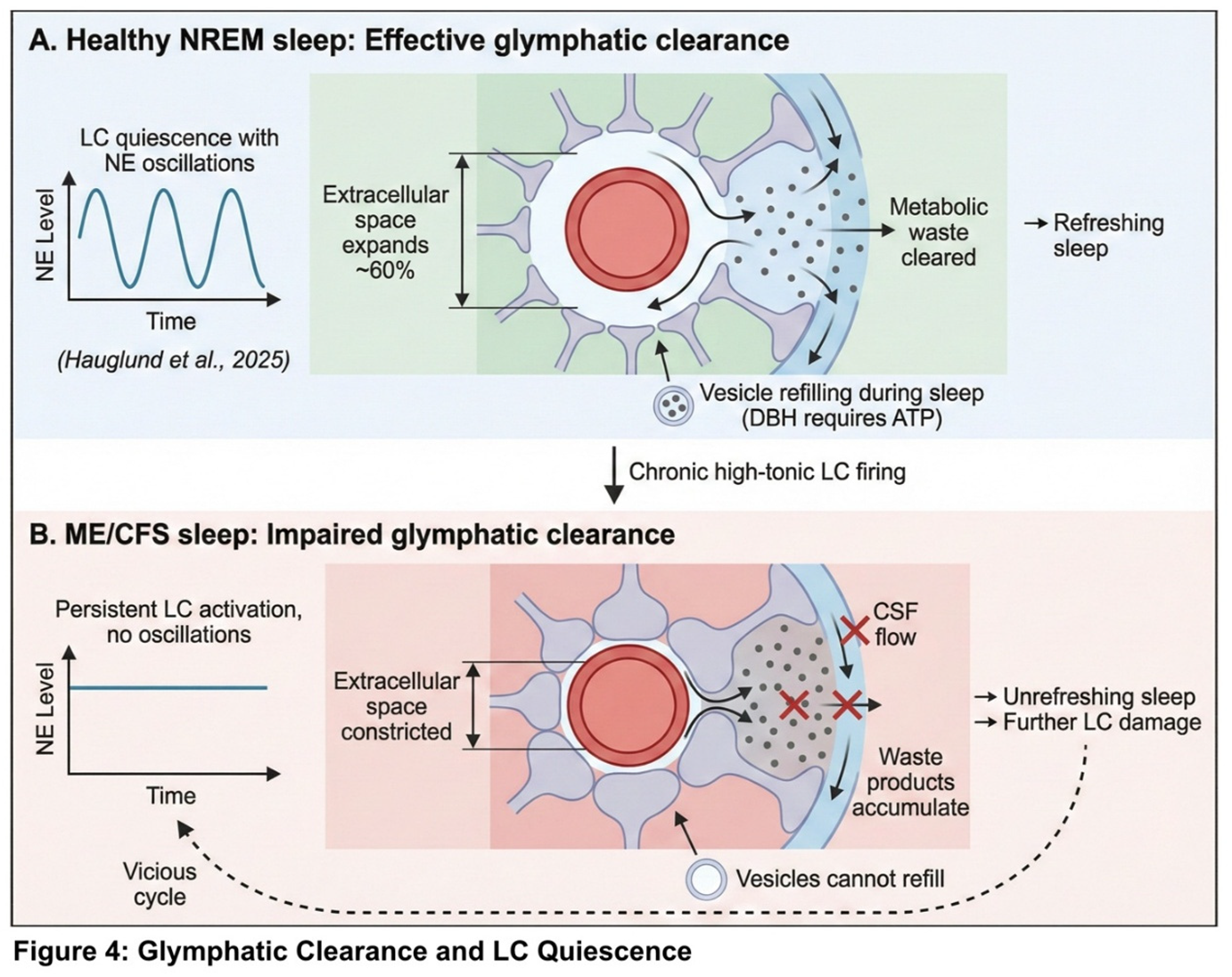
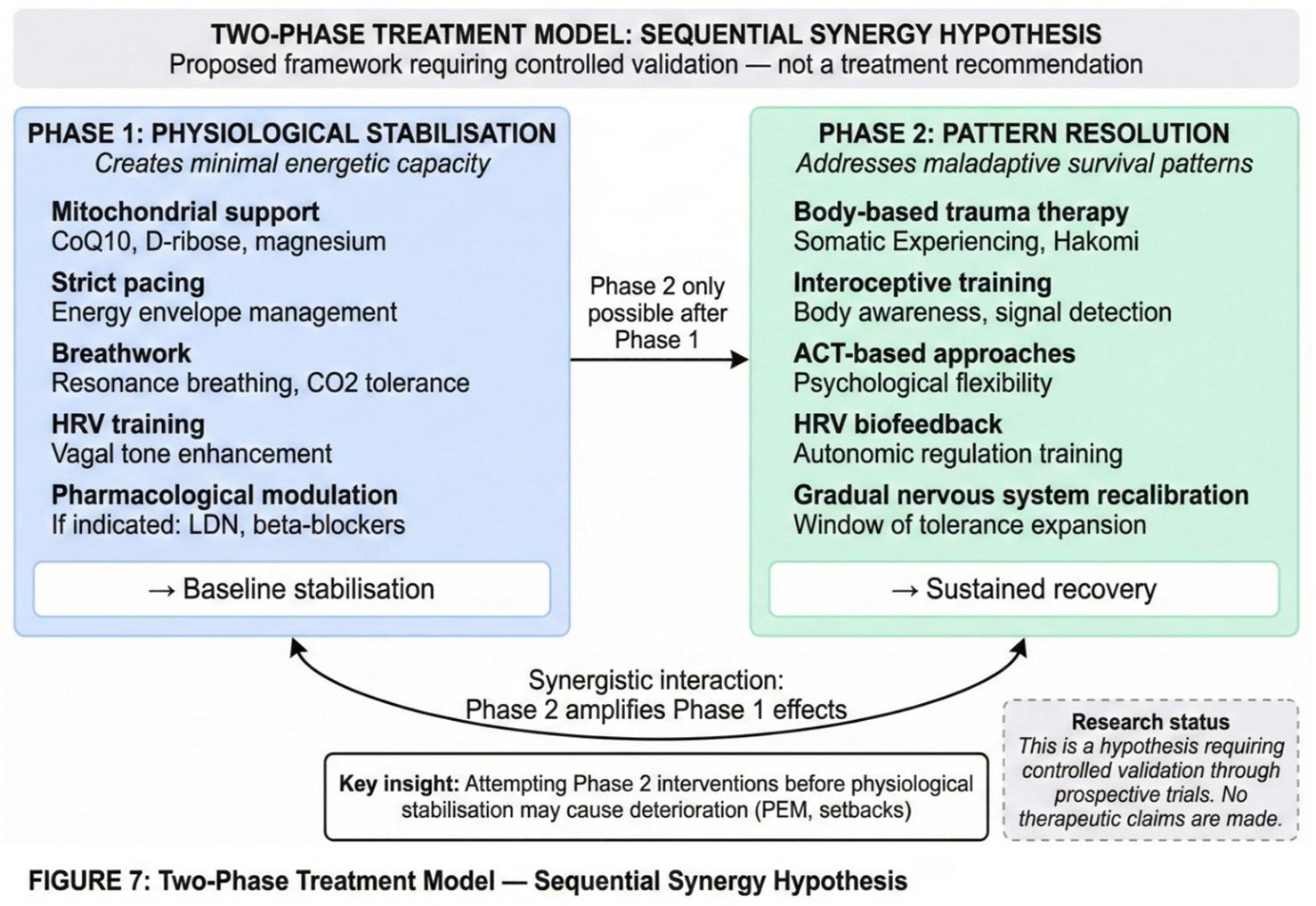
The LC depletion model explains unrefreshing sleep through the glymphatic connection outlined in Section 2.6. Effective waste clearance during sleep requires rhythmic norepinephrine oscillations during NREM sleep, with periods of LC quiescence allowing astrocyte shrinkage and CSF flow (Hauglund et al., 2025). With chronic high-tonic LC firing, the system cannot achieve these oscillations. Waste products accumulate rather than being cleared. Moreover, because dopamine-β-hydroxylase requires ATP, the sleep period cannot be used effectively to refill depleted vesicles. When the morning cortisol surge activates the system, it encounters a neurotransmitter reserve that has not been replenished—hence the sensation of waking unrestored. A proposed two-phase treatment framework, which requires controlled validation, is outlined in Figure 7.
5. Supporting Evidence and Alternative Explanations
5.1. Evidence Supporting the Hypothesis
The LC depletion hypothesis rests on several lines of evidence that, while not definitive, establish biological plausibility and consistency with clinical observations.
The biological plausibility of the core mechanism is well established. LC metabolic vulnerability has been documented, as has CRF-LC stress coupling. Chronic stress is known to produce mitochondrial dysfunction, vesicular monoamine storage is known to require ATP, and recent research has confirmed that glymphatic clearance requires reduced LC activity.
The clinical phenomenology of ME/CFS is consistent with the model's predictions. The hypothesis explains the 'wired-but-tired' paradox, post-exertional malaise, orthostatic intolerance, unrefreshing sleep, cognitive impairment, and the coexistence of hypervigilance and fatigue—all core features of the syndrome.
The dysautonomia profile observed in ME/CFS also fits the model. Reduced heart rate variability is well documented in ME/CFS (Nelson et al., 2019), as is orthostatic intolerance with reduced cerebral blood flow (van Campen et al., 2020). The paradox of apparent sympathetic overactivity coexisting with ineffective central noradrenergic output is precisely what the model predicts: hyperactive neurons with inadequate neurotransmitter release.
Preliminary conference data provide additional support, though these findings await peer review and cannot be considered definitive evidence. Presentations at the IACFS/ME 2025 conference reported findings consistent with the hypothesis, including low brain norepinephrine (Goldstein and Aregawi, 2025, conference abstract), reduced CRH-producing neurons in an autopsy study (Da Silva et al., 2025, conference abstract), and sympathetic dysfunction in a meta-analysis (Hendrix et al., 2025, conference abstract). These presentations are hypothesis-generating and should not be considered evidence on which the present model depends.
5.2. Alternative and Complementary Mechanisms
The LC depletion hypothesis is not mutually exclusive with other proposed mechanisms in ME/CFS. Indeed, given the heterogeneity of the syndrome, multiple pathways likely contribute.
Autoimmune mechanisms have been proposed, including β2-adrenergic receptor autoantibodies (Wirth and Scheibenbogen, 2020). Such autoantibodies could potentially impair noradrenergic signalling through a different mechanism than vesicular depletion, and both mechanisms might operate in different patient subgroups or even in the same patients.
Small fibre neuropathy has been documented in a significant subset of ME/CFS patients. This peripheral nerve pathology could contribute to autonomic dysfunction independently of central LC mechanisms, or could interact with central dysfunction to amplify symptoms.
Mast cell activation, which is increasingly recognised in ME/CFS, could affect LC function through histamine's known effects on noradrenergic neurons. Cerebrovascular dysfunction with reduced cerebral blood flow has been documented and could contribute to cognitive symptoms.
Mitochondrial dysfunction is well established in ME/CFS (Naviaux et al., 2016), though whether this is primary or secondary to other processes remains unclear. The present hypothesis proposes that mitochondrial dysfunction in the LC is a consequence of chronic stress-induced overactivation, but generalised mitochondrial dysfunction from other causes could produce similar downstream effects.
Immune-mediated LC damage from post-viral neuroinflammation has been suggested by neuroimaging studies showing microglial activation in ME/CFS (Nakatomi et al., 2014). Such inflammation could contribute to LC dysfunction through mechanisms distinct from the metabolic exhaustion proposed here.
The present hypothesis emphasises one pathway while acknowledging that ME/CFS is likely heterogeneous, with multiple pathways potentially converging on similar clinical endpoints.
5.3. Limitations and Uncertainties
Several important limitations must be acknowledged.
First, no direct measurements of vesicular norepinephrine content in ME/CFS LC neurons currently exist. The hypothesis rests on indirect evidence and mechanistic plausibility rather than direct demonstration of vesicular depletion.
Second, the ACE-ME/CFS link derives from cross-sectional data, and prospective studies confirming a causal relationship are lacking. Confounders including socioeconomic status, comorbidities, and recall bias have not been fully controlled.
Third, the model's reliance on ACE history presents a methodological limitation. ACE questionnaires cannot capture transgenerational epigenetic transmission. An estimated 25 to 35 percent of ME/CFS patients deny significant personal ACEs (Kempke et al., 2013). These 'ACE-negative' patients may represent underreporting due to recall limitations, forms of adversity not captured by standard ACE instruments, transgenerational epigenetic programming from parental or grandparental stress, or alternative pathogenic pathways not involving early-life stress. The present model accommodates transgenerational transmission (Section 2.4) but acknowledges that ME/CFS is likely aetiologically heterogeneous.
Fourth, the pharmacological observations regarding stimulant tolerance are anecdotal, derived from clinical impressions rather than controlled studies.
Fifth, the conference data cited are preliminary and have not undergone peer review.
Sixth, causal direction is unclear and likely bidirectional. LC dysfunction could cause ME/CFS features, but ME/CFS features could also worsen LC dysfunction through feedback loops.
Seventh, the model may be incomplete, focusing on the LC while potentially missing other critical nodes in the pathophysiology.
6. Testable Predictions and Research Agenda
To enable empirical evaluation of the proposed model, this section specifies falsifiable predictions and corresponding study designs. For each prediction, we describe primary endpoints, relevant covariates, and considerations for power and analysis. All proposed studies would conform to established methodological standards: MRI methods would follow COBIDAS-MRI recommendations; PET acquisition, modelling, and quantification would follow SNMMI procedure standards and QIBA guidance; HRV analysis would follow Task Force recommendations; and observational reporting would follow STROBE guidelines. Site-specific OSF preregistration and local IRB approval would precede any data collection. An overview of the testable predictions and corresponding methodologies is provided in Figure 6.
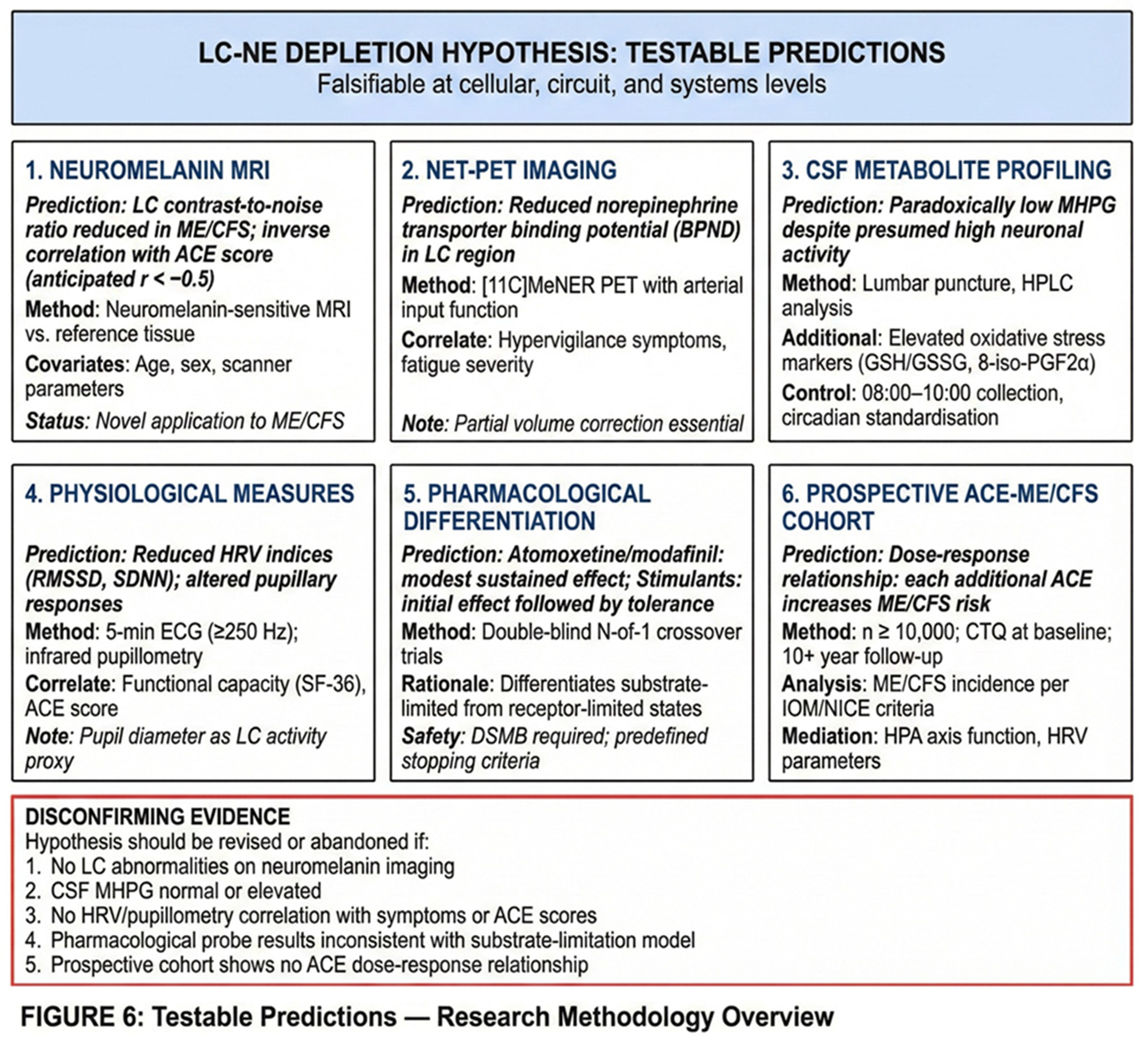
6.1. Neuromelanin-Sensitive MRI of the LC
The first prediction concerns LC structure as assessed by neuromelanin-sensitive MRI. We predict that LC contrast-to-noise ratio will be reduced in ME/CFS patients compared to healthy controls and will correlate inversely with ACE score as measured by the Childhood Trauma Questionnaire. Based on the model's assumptions, we anticipate a correlation of r < −0.5, representing a medium-to-large effect. LC signal should also correlate inversely with ME/CFS symptom severity.
The proposed study design involves cross-sectional comparison of ME/CFS patients (n ≥ 60) with healthy controls (n ≥ 60), stratified by ACE score tertiles. The primary endpoint would be LC contrast-to-noise ratio relative to dorsal pontine reference tissue.
Several interpretive considerations are important. Neuromelanin MRI signal reflects multiple factors including norepinephrine synthesis history, iron content, and age-related changes. Signal reductions therefore cannot be directly equated with acute norepinephrine depletion. Age- and scanner-related partial volume effects would need to be modelled as covariates. Exploratory analyses could include rostro-caudal LC profile analysis to detect regionally selective signal changes.
Relevant covariates include age (modelled non-linearly), sex, scanner and coil type, signal-to-noise ratio, voxel size, and head motion. Power analysis indicates that a pilot study of approximately 20 participants per group would enable variance estimation, with the main study powered for a medium effect size (d ≈ 0.5) at α = 0.05 two-sided. Multiple comparison correction would follow a hierarchical testing strategy with family-wise error control.
Neuromelanin-sensitive MRI is an established technique for LC visualisation (Betts et al., 2019; Priovoulos et al., 2018), but its application to ME/CFS would be novel.
6.2. NET-PET Imaging
The second prediction concerns norepinephrine transporter availability as assessed by positron emission tomography. We predict that NET binding potential will be reduced in the LC region in ME/CFS patients and will correlate with hypervigilance symptoms and fatigue severity.
The proposed protocol would use [11C]MeNER as the primary radioligand, with 90-minute dynamic acquisition and either arterial input function or reference tissue modelling using occipital cortex. Geometric transfer matrix partial volume correction would be applied for the LC region of interest given the structure's small size.
Several interpretive considerations apply. Reduced NET binding can reflect multiple factors: decreased transporter density, increased endogenous norepinephrine occupancy, or partial-volume effects. The latter concern is particularly relevant given the LC's small size. Alternative or complementary PET ligands, such as [18F]-labelled tracers if available, might provide converging evidence. Indeed, converging evidence from neuromelanin MRI, CSF markers, and physiological measures would be essential for confident interpretation (Pietrzak et al., 2013).
Relevant covariates include endogenous norepinephrine levels, medications (SNRIs, atomoxetine, tricyclic antidepressants, and beta-blockers, all requiring appropriate washout periods), smoking status, age, and sex.
6.3. CSF Metabolite Profiling
The third prediction concerns cerebrospinal fluid metabolites. We predict paradoxically low levels of 3-methoxy-4-hydroxyphenylglycol (MHPG), the principal norepinephrine metabolite, despite clinical and physiological signs suggesting high sympathetic activity. We also predict elevated oxidative stress markers (GSH/GSSG ratio, 8-iso-PGF2α) and elevated inflammatory markers.
The proposed design involves comparison of ME/CFS patients (n ≥ 30) with controls, with subgroup analysis by ACE score.
Interpretation of CSF metabolites requires caution given substantial inter-individual variability, posture effects, and diurnal fluctuations. Pre-analytic standardisation is therefore critical. Lumbar puncture should be performed between 08:00 and 10:00 to control for circadian variation. Polypropylene collection tubes should be used to prevent binding. Immediate centrifugation, aliquoting, and storage at −80°C are essential, as is exclusion of samples with visible haemolysis. Analysis would employ robust regression with adjustment for multiple comparisons (Shungu et al., 2012).
Pre-analytics and reporting would follow STROBE and BRISQ principles, with site-level standard operating procedures preregistered on OSF before enrolment.
6.4. Physiological Measures: Heart Rate Variability and Pupillometry
The fourth prediction concerns physiological measures that can serve as non-invasive windows on LC function. We predict that HRV parameters—specifically RMSSD, SDNN, and LF/HF ratio—will show reduced parasympathetic indices in ME/CFS patients. We further predict that pupillometry will reveal reduced pupillary light reflex amplitude and altered spontaneous pupil fluctuations (Murphy et al., 2014; Joshi et al., 2016). Both sets of measures should correlate with functional capacity and ACE score.
The proposed HRV protocol involves 5-minute seated rest followed by 5-minute standing, with ECG sampling at 250 Hz or higher and exclusion of individuals with significant arrhythmias. Both time-domain analysis (RMSSD, SDNN) and frequency-domain analysis (LF, HF, LF/HF) would be performed.
An important methodological note concerns the LF/HF ratio, which has been used as an index of sympathovagal balance but remains controversial. Notably, this ratio becomes uninterpretable when respiratory rate is 6 breaths per minute or lower, as occurs in coherent breathing, because respiratory sinus arrhythmia then falls entirely within the LF band. For this reason, time-domain measures (RMSSD, SDNN) are prioritised as more robust markers of cardiac vagal tone.
The proposed pupillometry protocol involves infrared eye-tracking with measurement of baseline pupil diameter, light reflex amplitude, latency, and return-to-baseline time, as well as a spontaneous pupil fluctuation index. Test-retest reliability would be reported as intraclass correlation coefficient. Constant room illumination (below 5 lux) and screen luminance would be documented.
Pre-registered primary endpoints would include task-evoked pupil dilation, pupillary light reflex amplitude (expressed as percentage change), and RMSSD as the primary vagal marker. Baseline pupil diameter (as a proxy for tonic LC activity) and task-evoked pupil dilation (as a proxy for phasic LC activity) would be analysed separately.
6.5. Pharmacological Differentiation: N-of-1 Trials
The fifth prediction concerns pharmacological differentiation between substrate-limited and receptor-limited states. The rationale for this approach derives from the model's core claim: if the problem is vesicular depletion rather than receptor dysfunction, then drugs that force vesicular release should produce initial benefit followed by rapid tolerance (as they deplete remaining stores), while drugs that block reuptake without forcing release should produce more modest but sustainable effects.
The proposed design involves double-blind, placebo-controlled N-of-1 crossover trials with a 2-week placebo run-in period. The agents to be compared include atomoxetine (which blocks reuptake without forcing release), modafinil (a DAT/NET blocker), and, if ethically justified, low-dose stimulant (which forces vesicular release). Washout periods between conditions would be a minimum of 5 to 7 half-lives.
The specific prediction is that atomoxetine and modafinil will produce modest, sustainable effects consistent with substrate limitation, while stimulants will produce initial improvement followed by tolerance consistent with emptying of limited vesicular stores.
Given the vulnerable patient population and the potential for adverse effects, rigorous safety monitoring is essential. A Data Safety Monitoring Board would be required, with predefined stopping criteria for cardiovascular events or severe psychiatric symptoms. ECG, blood pressure, sleep quality, and anxiety would be monitored at each visit. All studies would be preregistered on OSF and ClinicalTrials.gov, and careful ethical review would be required before initiation.
Reporting would follow the CONSORT Extension for N-of-1 Trials (CENT 2015), with blinding procedures, washout periods, and stopping rules preregistered before enrolment.
6.6. Transcutaneous Auricular Vagus Nerve Stimulation: A Therapeutic Window to the LC-NE System
The auricular branch of the vagus nerve provides a non-invasive access point to brainstem nuclei implicated in ME/CFS pathophysiology. Anatomical and neuroimaging studies have established that electrical stimulation of the cymba conchae—the ear region predominantly innervated by the auricular branch of the vagus nerve—activates the nucleus tractus solitarius, which projects directly to the locus coeruleus (Frangos et al., 2015; Yakunina et al., 2017). This pathway offers a potential therapeutic target for modulating LC-NE function in ME/CFS. The anatomical basis and proposed mechanisms of taVNS are illustrated in Figure 8.
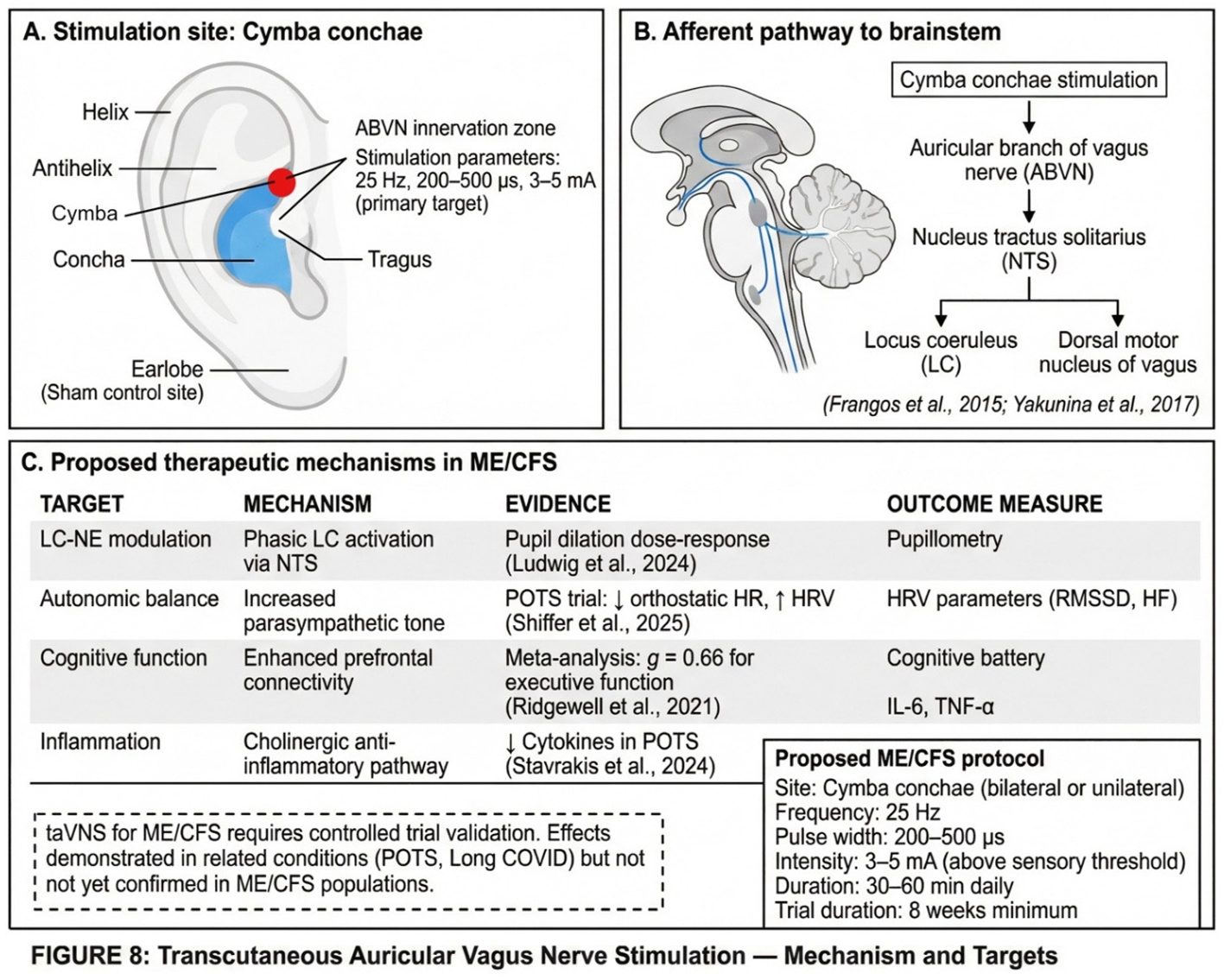
6.6.1. Pupillary Responses as LC Activity Proxy
Pupil diameter is a well-validated non-invasive proxy of LC-NE activity under controlled luminance conditions, though it remains susceptible to confounding by light, cognition, and pharmacological factors (Joshi and Gold, 2020). If taVNS activates the LC as proposed, measurable pupillary changes should occur.
Recent evidence supports this mechanism. Ludwig et al. (2024) demonstrated that phasic taVNS produces significantly greater pupil dilation compared to sham stimulation (0.18 ± 0.03 versus 0.09 ± 0.03, p = 0.001). Higher stimulation intensity (5 mA versus 3 mA) produced larger effects (p < 0.001), suggesting dose-dependent LC activation. Importantly, the influence of intensity on pupil dilation appears stronger than that of frequency, with 25 Hz showing no significant advantage over 10 Hz at matched intensities.
Villani et al. (2022) demonstrated that taVNS modulation of target-related pupil dilation depends on pre-stimulation pupil diameter—an index of baseline tonic LC-NE activity. Active taVNS was associated with differential pupil responses based on baseline state, consistent with the Adaptive Gain Theory prediction that LC-NE effects are state-dependent (Aston-Jones and Cohen, 2005).
A critical caveat is necessary: earlier studies reported null effects on pupillary measures (Keute et al., 2019; Warren et al., 2019). This heterogeneity likely reflects differences in stimulation protocols. Continuous stimulation paradigms show weaker effects than phasic (burst) protocols, and stimulation at perceptual threshold may be insufficient for reliable LC activation. For ME/CFS research applications, phasic protocols with intensity above sensory threshold (3–5 mA) appear most promising.
6.6.2. Orthostatic Intolerance: Direct Evidence from POTS
Orthostatic intolerance affects the majority of ME/CFS patients, with postural orthostatic tachycardia syndrome representing a common comorbidity. Given the shared pathophysiology of autonomic dysregulation, evidence from POTS populations is directly relevant to ME/CFS.
A recent randomised, sham-controlled, double-blind trial (NCT05043051) provides promising preliminary evidence. After two months of daily taVNS at the cymba conchae, the active treatment group demonstrated a reduction in orthostatic tachycardia from 26.4 bpm (SD = 11.3) at baseline to 17.6 bpm (SD = 9.9)—a clinically meaningful improvement of approximately one-third (Shiffer et al., 2025). No change occurred in the sham group. The active treatment group also showed HRV changes reflecting improved autonomic balance, including reduced LF/HF ratio during postural transition.
An earlier open-label study found that four hours of daily transcutaneous vagus nerve stimulation at the cymba conchae for 14 days significantly reduced orthostatic intolerance and gastrointestinal symptoms in POTS patients, with a trend toward reduced standing heart rate. These findings align with mechanistic studies showing that tVNS increases HRV parameters (RMSSD, HF power) and decreases LF/HF ratio in heart failure patients, suggesting enhanced parasympathetic tone and attenuated sympathetic activity (Stavrakis et al., 2020).
The mechanistic plausibility for ME/CFS is considerable. The autonomic phenotype of ME/CFS—characterised by reduced HRV, exaggerated sympathetic responses to orthostasis, and impaired parasympathetic recovery—mirrors the profile responsive to taVNS in POTS. Both conditions show evidence of parasympathetic cardiovagal impairment alongside sympathetic hyperactivation. The reduction in anti-autonomic autoantibodies and inflammatory cytokines observed in the POTS trial suggests that taVNS may address multiple pathophysiological pathways simultaneously.
6.6.3. Cognitive Function and Brain Fog
Cognitive dysfunction—commonly described as 'brain fog'—is a cardinal symptom of ME/CFS, encompassing deficits in attention, working memory, processing speed, and executive function. The LC-NE system is critically involved in all of these cognitive domains (Sara, 2009). If taVNS modulates LC function as proposed, cognitive improvements should follow.
A meta-analysis of 19 taVNS studies in healthy populations found a small but significant overall effect on cognition (Hedges' g = 0.21, p < 0.0001), with the largest domain-specific effect observed for executive function (g = 0.66, 95% CI = 0.14–1.17, p = 0.02) (Ridgewell et al., 2021). Individual studies have reported improvements in cognitive flexibility (d = 0.49), working memory, and attentional processing (Borges et al., 2020; Jacobs et al., 2015; Ventura-Bort et al., 2018).
A Long COVID proof-of-concept trial—relevant given the substantial overlap between Long COVID and ME/CFS—found that four weeks of daily taVNS reduced symptoms in the active group while the sham group showed no change. Following crossover, more than half of participants no longer demonstrated any symptom prevalence (Badran et al., 2022).
Functional connectivity studies have shown that taVNS increases connectivity from hippocampus to prefrontal cortex and cingulate while decreasing connectivity to anterior temporal regions—changes that may reflect normalisation of networks disrupted in ME/CFS (Fang et al., 2017). The medial prefrontal cortex appears particularly responsive, with taVNS-induced connectivity changes correlating with working memory improvements in older adults.
6.6.4. Stimulation Laterality: Unilateral vs. Bilateral
Most taVNS research has employed unilateral left-ear stimulation, based on the historical convention from invasive VNS studies. However, transcutaneous auricular stimulation activates only afferent fibres, which are processed centrally before any efferent cardiac effects—mitigating the safety concerns that drove the left-only convention for invasive stimulation.
A systematic review and meta-analysis of 177 studies (n = 6,322 subjects) found that bilateral taVNS did not significantly increase the likelihood of cardiovascular adverse events compared to unilateral stimulation (Redgrave et al., 2022). Theoretical considerations suggest that bilateral stimulation may enhance effects through increased afferent input to the brainstem.
An fMRI study in 50 healthy controls directly compared left, right, and bilateral taVNS, finding distinct functional connectivity changes depending on stimulation laterality and frequency. Left stimulation preferentially altered connectivity in hippocampal and salience networks, while bilateral stimulation produced the most widespread effects.
6.6.5. Proposed Study Design for ME/CFS
Based on the converging evidence reviewed above, we propose a randomised, sham-controlled, crossover trial design for evaluating taVNS in ME/CFS.
The study population would include at least 40 patients meeting ME-ICC (International Consensus Criteria, 2011) or CCC (Canadian Consensus Criteria, 2003) diagnostic criteria for ME/CFS, stratified by orthostatic intolerance severity. The washout period between conditions would be at least two weeks to exclude carry-over effects.
The active intervention would consist of cymba conchae stimulation at 25 Hz, 200–500 microseconds pulse width, intensity 3–5 mA (above sensory threshold), for 30–60 minutes daily over eight weeks. The sham control would consist of earlobe stimulation with identical parameters.
Primary endpoints would include HRV parameters (RMSSD, HF power, LF/HF ratio), pupillary light reflex and event-related pupil dilation (as LC activity proxies), and orthostatic heart rate response on tilt test.
Secondary endpoints would include fatigue severity (Chalder Fatigue Scale), cognitive function (executive function battery), orthostatic symptom burden, inflammatory markers (IL-6, TNF-α), and post-exertional malaise frequency.
Exploratory analyses would compare unilateral versus bilateral stimulation in a subset and examine whether pupillary responses predict clinical improvement, which could serve as a potential predictive biomarker for treatment selection.
6.7. Prospective ACE-ME/CFS Cohort
The final component of the research agenda addresses the foundational epidemiological question: does ACE exposure prospectively predict ME/CFS development?
The proposed design involves a large general population cohort (n ≥ 10,000) with ACE assessment via the Childhood Trauma Questionnaire at baseline along with biomarkers including HRV and inflammatory markers. Long-term follow-up (10+ years) would document ME/CFS incidence according to IOM/NICE criteria, with careful documentation of infectious triggers.
The primary prediction is a dose-response relationship in which each additional ACE increases ME/CFS risk. Secondary analyses would examine mediation and moderation by HPA axis function and HRV, as well as competing risks.
Observational reporting would follow STROBE guidelines, and analysis plans and amendments would be preregistered on OSF before database lock.
7. Potential Disconfirming Evidence
A hypothesis is only scientifically useful if it can be proven wrong. The LC depletion model should be abandoned or substantially revised if the following observations emerge.
First, the model would be undermined if LC neuromelanin imaging shows no abnormalities in ME/CFS patients compared to controls. This would challenge the core claim that the LC is structurally affected in this condition.
Second, the model would be undermined if cerebrospinal fluid MHPG is normal or elevated rather than low. The model predicts reduced norepinephrine turnover despite high sympathetic tone; normal or elevated MHPG would contradict this prediction.
Third, the model would be undermined if HRV and pupillometry measures show no correlation with symptoms or ACE scores. These physiological markers are proposed as windows on LC function, and the absence of expected correlations would weaken confidence in the model.
Fourth, the model would be undermined if pharmacological probe trials show inconsistent patterns that do not match substrate-limitation predictions. Specifically, if stimulants produce sustained benefit without tolerance, or if reuptake inhibitors fail while stimulants succeed, the substrate-limitation hypothesis would require revision.
Fifth, the model would be undermined if mitochondrial support interventions show no benefit in appropriately powered trials. While such a negative finding could reflect issues with specific interventions or delivery, consistent failure across multiple approaches would challenge the metabolic exhaustion component of the model.
Sixth, and perhaps most fundamentally, the model would be undermined if a prospective cohort study shows no ACE dose-response relationship with ME/CFS incidence. This would call into question the foundational premise that childhood adversity programmes vulnerability to later ME/CFS development.
8. Discussion
8.1. Strengths
The proposed framework has several strengths that distinguish it from previous models of ME/CFS pathophysiology.
First, it integrates disparate observations that have previously been discussed in isolation. ACE epidemiology, clinical phenomenology, autonomic dysfunction, and sleep pathology are brought together within a single mechanistic framework.
Second, the model is mechanistically specific. Rather than invoking vague notions that 'stress causes illness,' it proposes a concrete pathway involving identified neural structures, specific cellular processes, and measurable intermediate variables. This specificity enables targeted investigation.
Third, the model generates testable predictions across multiple levels of analysis—from neuroimaging to cerebrospinal fluid biochemistry to physiological measures to pharmacological responses. This multi-level predictive capacity means that the model can be evaluated through converging evidence rather than relying on any single line of investigation.
Fourth, the model explains heterogeneity through the two-hit framework. Not everyone with ACEs develops ME/CFS, and not everyone with ME/CFS has ACEs. The model accommodates this by proposing that ACEs create vulnerability while a second hit—typically infection—is required to precipitate the full syndrome. This explains why some highly stressed individuals remain healthy while some apparently unstressed individuals become ill.
Fifth, the model has translational potential if validated. Identifying the LC as a critical vulnerable node would enable targeted therapeutic development and patient stratification.
8.2. Weaknesses
Several weaknesses must be acknowledged alongside these strengths.
First, the evidence supporting the model is indirect. No direct measurement of LC vesicular norepinephrine in humans currently exists. The hypothesis is constructed from converging indirect evidence and mechanistic plausibility rather than direct demonstration of the proposed pathology.
Second, ACE ascertainment involves retrospective reporting bias. Adults are asked to recall childhood experiences, and such recall may be systematically affected by current health status, mood, or other factors.
Third, the pharmacological observations regarding stimulant tolerance that helped generate the hypothesis are anecdotal clinical impressions, not findings from controlled studies.
Fourth, the conference data cited as supporting evidence are preliminary and have not undergone peer review.
Fifth, causal direction is unclear and likely bidirectional. While the model proposes that LC dysfunction causes ME/CFS features, it is also possible that ME/CFS features worsen LC dysfunction through positive feedback loops. Distinguishing cause from consequence is methodologically challenging.
Sixth, the model may be incomplete. By focusing on the LC, it may miss other critical nodes in the pathophysiology. ME/CFS is a complex systemic illness, and reducing it to dysfunction of a single brainstem nucleus—however influential—may oversimplify the problem.
8.3. Implications if Hypothesis Is Correct
If the LC depletion hypothesis is validated by future research, several implications would follow.
For understanding ME/CFS, the model would represent a shift from viewing the condition as a 'mysterious post-viral syndrome' to understanding it as a mechanistic process with identifiable vulnerability factors. It would explain why a subset of patients have ACE histories and high achievement orientation phenotypes—these are markers of the chronic stress that programmes LC vulnerability. It would explain the typical clinical course: years of high functioning followed by a trigger event and then failure to recover. And it would explain why the condition is so refractory: the trapped state involves multiple positive feedback loops that resist simple interventions.
For research, the model would provide concrete targets for investigation—the LC itself, its mitochondrial function, its vesicular stores, and its relationship to glymphatic clearance. It would suggest stratification strategies based on ACE history and physiological markers. And it would identify surrogate markers such as HRV and pupillometry that could be used to assess treatment response without waiting for long-term clinical outcomes.
For patients, the model would validate their experiences by demonstrating that symptoms have a neurobiological substrate. This is not 'all in your head' in the dismissive sense—rather, it is specifically in a small but critical part of your brainstem, the chronic dysfunction of which produces the full range of symptoms. Such validation could help reduce the stigma that patients often experience.
However, it is essential to emphasise that no immediate therapeutic implications flow from this hypothesis at present. Research must precede any clinical application. Patients should not change their treatment based on an unvalidated model, and clinicians should not recommend interventions based on theoretical predictions that have not been empirically confirmed.
9. Conclusion
We have proposed a specific, testable hypothesis: ME/CFS in a subset of patients results from chronic norepinephrine depletion of the locus coeruleus, programmed by adverse childhood experiences (or their transgenerational epigenetic transmission) and triggered into clinical manifestation by acute systemic stressors.
The hypothesis integrates ACE epidemiology, LC vulnerability to chronic stress, mitochondrial and vesicular mechanisms, glymphatic dysfunction, clinical phenomenology, and preliminary pharmacological observations into a coherent mechanistic framework. It explains the core features of ME/CFS—including the 'wired-but-tired' paradox, post-exertional malaise, orthostatic intolerance, and unrefreshing sleep—through the single lens of a system that is firing constantly but releasing inadequate neurotransmitter.
The hypothesis generates specific predictions that are testable through neuroimaging, cerebrospinal fluid profiling, physiological surrogates, pharmacological probes, non-invasive neuromodulation, and prospective epidemiology. The predictions are falsifiable: observations inconsistent with the model would require its revision or abandonment.
Most importantly, we emphasise that this is a hypothesis, not established fact. It requires rigorous testing before any conclusions can be drawn. Until empirical evidence either supports or refutes the model, no therapeutic implications should be inferred. We present this framework to stimulate discussion, to guide research, and ultimately to move the field toward a mechanistic understanding of one of medicine's most challenging conditions.
Funding
This work received no external funding.
Acknowledgments
The author thanks colleagues who shared clinical observations and ME/CFS patients who shared their experiences.
Conflicts of Interest
The author has clinical experience with body-based trauma therapies (Somatic Experiencing, Hakomi) and systemic work with ME/CFS patients, which informed the development of this hypothesis. No proprietary methods are promoted. All therapeutic approaches discussed are well-established techniques that would require validation through controlled research before any clinical recommendations could be made. The author has no relationships with the pharmaceutical industry.
Data Availability
Not applicable (conceptual hypothesis paper). Preregistration of proposed studies is intended prior to any data collection.
Ethics
No human participants or data are included in this manuscript. The study designs described are proposals intended for institutional adoption; site-specific OSF preregistration and local IRB approval would precede any data collection.
Abbreviations
ABVN, auricular branch of the vagus nerve; ACE, adverse childhood experience; ATP, adenosine triphosphate; BNST, bed nucleus of the stria terminalis; BPND, non-displaceable binding potential; BRISQ, Biospecimen Reporting for Improved Study Quality; CENT, CONSORT Extension for N-of-1 Trials; COBIDAS, Committee on Best Practices in Data Analysis and Sharing (MRI); CONSORT, Consolidated Standards of Reporting Trials; CNR, contrast-to-noise ratio; CRF, corticotropin-releasing factor; CSF, cerebrospinal fluid; CTQ, Childhood Trauma Questionnaire; DAT, dopamine transporter; DBH, dopamine-β-hydroxylase; DSMB, Data Safety Monitoring Board; EBV, Epstein–Barr virus; ECG, electrocardiogram; GTM, geometric transfer matrix; HF, high-frequency power; HPA, hypothalamic-pituitary-adrenal; HRV, heart rate variability; ICC, intraclass correlation coefficient; IOM, Institute of Medicine; IRB, Institutional Review Board; LC, locus coeruleus; LF, low-frequency power; ME/CFS, myalgic encephalomyelitis/chronic fatigue syndrome; MHPG, 3-methoxy-4-hydroxyphenylglycol; MRI, magnetic resonance imaging; NE, norepinephrine; NET, norepinephrine transporter; NICE, National Institute for Health and Care Excellence; NREM, non-rapid eye movement; NTS, nucleus tractus solitarius; OSF, Open Science Framework; PEM, post-exertional malaise; PET, positron emission tomography; OI, orthostatic intolerance; POTS, postural orthostatic tachycardia syndrome; PVC, partial volume correction; QIBA, Quantitative Imaging Biomarkers Alliance; RMSSD, root mean square of successive differences; ROI, region of interest; ROS, reactive oxygen species; SARS-CoV-2, severe acute respiratory syndrome coronavirus 2; SDNN, standard deviation of normal-to-normal intervals; SFN, small fibre neuropathy; SNMMI, Society of Nuclear Medicine and Molecular Imaging; SNRI, serotonin-norepinephrine reuptake inhibitor; STROBE, Strengthening the Reporting of Observational Studies in Epidemiology; TCA, tricyclic antidepressant; V-ATPase, vacuolar-type ATPase; VMAT, vesicular monoamine transporter.
References
- Amann, R.P. (2008). The cycle of the seminiferous epithelium in humans: A need to revisit? Journal of Andrology, 29(5), 469–487.
- Aston-Jones, G., & Cohen, J.D. (2005). An integrative theory of locus coeruleus-norepinephrine function: Adaptive gain and optimal performance. Annual Review of Neuroscience, 28, 403–450.
- Badran, B.W., Dowdle, L.T., Mithoefer, O.J., LaBate, N.T., Coatsworth, J., Brown, J.C., DeVries, W.H., Austelle, C.W., McTeague, L.M., & George, M.S. (2018). Neurophysiologic effects of transcutaneous auricular vagus nerve stimulation (taVNS) via electrical stimulation of the tragus: A concurrent taVNS/fMRI study and review. Brain Stimulation, 11(3), 492–500.
- Bangasser, D.A., & Valentino, R.J. (2020). The locus coeruleus-norepinephrine system in stress and arousal: Unraveling historical, current, and future perspectives. Frontiers in Psychiatry, 11, 601519.
- Baumeister, D., Akhtar, R., Ciufolini, S., Pariante, C.M., & Mondelli, V. (2016). Childhood trauma and adulthood inflammation: A meta-analysis. Molecular Psychiatry, 21(5), 642–649.
- Betts, M.J., Kirilina, E., Otaduy, M.C.G., et al. (2019). Locus coeruleus imaging as a biomarker for noradrenergic dysfunction in neurodegenerative diseases. Brain, 142(9), 2558–2571.
- Blockmans, D., Persoons, P., Van Houdenhove, B., & Bobbaers, H. (2006). Does methylphenidate reduce the symptoms of chronic fatigue syndrome? The American Journal of Medicine, 119(12), 1047–1053.
- Borges, U., Knops, L., Laborde, S., Klatt, S., & Raab, M. (2020). Transcutaneous vagus nerve stimulation may enhance only specific aspects of the core executive functions: A randomized crossover trial. Frontiers in Neuroscience, 14, 523.
- Bunea, I.M., Szentagotai-Tatar, A., & Miu, A.C. (2017). Early-life adversity and cortisol response to social stress: A meta-analysis. Translational Psychiatry, 7(12), 1274.
- Burkusis, M., Plaks, V., Michael, A., et al. (2015). Environment and the developing embryo. In Principles of Developmental Genetics (2nd ed., pp. 143–154). Academic Press.
- Da Silva, J.P., et al. (2025). Delineating clinical phenotypes and HPA-axis dysfunction in ME/CFS. Presented at IACFS/ME 2025 Conference [conference abstract—not peer-reviewed].
- Dias, B.G., & Ressler, K.J. (2014). Parental olfactory experience influences behavior and neural structure in subsequent generations. Nature Neuroscience, 17(1), 89–96.
- Edwards, R.H. (2007). The neurotransmitter cycle and quantal size. Neuron, 55(6), 835–849.
- Eiden, L.E., & Weihe, E. (2011). VMAT2: A dynamic regulator of brain monoaminergic neuronal function interacting with drugs of abuse. Annals of the New York Academy of Sciences, 1216, 86–98.
- Fang, J., Egorova, N., Rong, P., Liu, J., Hong, Y., Fan, Y., Wang, X., Wang, H., Yu, Y., Ma, Y., Xu, C., Li, S., Zhao, J., Luo, M., Zhu, B., & Kong, J. (2017). Early cortical biomarkers of longitudinal transcutaneous vagus nerve stimulation treatment success in depression. NeuroImage: Clinical, 14, 105–111.
- Felitti, V.J., Anda, R.F., Nordenberg, D., et al. (1998). Relationship of childhood abuse and household dysfunction to many of the leading causes of death in adults. American Journal of Preventive Medicine, 14(4), 245–258.
- Frangos, E., Ellrich, J., & Komisaruk, B.R. (2015). Non-invasive access to the vagus nerve central projections via electrical stimulation of the external ear: fMRI evidence in humans. Brain Stimulation, 8(3), 624–636.
- Goldstein, D.S., & Aregawi, D. (2025). Low brain norepinephrine in ME/CFS. Presented at IACFS/ME 2025 Conference [conference abstract—not peer-reviewed].
- Hauglund, N.L., Andersen, M., Tokarska, K., et al. (2025). Norepinephrine-mediated slow vasomotion drives glymphatic clearance during sleep. Cell, 188(3), 606–622.
- Hendrix, J., et al. (2025). Sympathetic dysfunction in ME/CFS: A meta-analysis. Presented at IACFS/ME 2025 Conference [conference abstract—not peer-reviewed].
- Jacobs, H.I.L., Riphagen, J.M., Razat, C.M., Wiese, S., & Sack, A.T. (2015). Transcutaneous vagus nerve stimulation boosts associative memory in older individuals. Neurobiology of Aging, 36(5), 1860–1867.
- Joshi, S., Li, Y., Kalwani, R.M., & Gold, J.I. (2016). Relationships between pupil diameter and neuronal activity in the locus coeruleus, colliculi, and cingulate cortex. Neuron, 89(1), 221–234.
- Joshi, S., & Gold, J.I. (2020). Pupil size as a window on neural substrates of cognition. Trends in Cognitive Sciences, 24(6), 466–480.
- Kempke, S., Luyten, P., Claes, S., et al. (2013). Self-critical perfectionism and fatigue and pain in chronic fatigue syndrome. Psychological Medicine, 43(5), 995–1002.
- Keute, M., Demirezen, M., Graf, A., Mueller, N.G., & Zaehle, T. (2019). No modulation of pupil size and event-related pupil response by transcutaneous auricular vagus nerve stimulation (taVNS). Scientific Reports, 9, 11452.
- Komaroff, A.L., & Lipkin, W.I. (2021). Insights from myalgic encephalomyelitis/chronic fatigue syndrome may help unravel the pathogenesis of postacute COVID-19 syndrome. Trends in Molecular Medicine, 27(9), 895–906.
- Ludwig, M., Wienke, C., Betts, M.J., Zaehle, T., & Hämmerer, D. (2024). Current and future directions in transcutaneous vagus nerve stimulation: Research, application, and effects on the locus coeruleus-norepinephrine system. Neuroscience & Biobehavioral Reviews, 159, 105607.
- Lumey, L.H., Stein, A.D., & Susser, E. (2011). Prenatal famine and adult health. Annual Review of Public Health, 32, 237–262.
- McGowan, P.O., Sasaki, A., D'Alessio, A.C., et al. (2009). Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nature Neuroscience, 12(3), 342–348.
- Murphy, P.R., O'Connell, R.G., O'Sullivan, M., Robertson, I.H., & Balsters, J.H. (2014). Pupil diameter covaries with BOLD activity in human locus coeruleus. Human Brain Mapping, 35(8), 4140–4154.
- Nakatomi, Y., Mizuno, K., Ishii, A., et al. (2014). Neuroinflammation in patients with chronic fatigue syndrome/myalgic encephalomyelitis: An [11C-(R)]PK11195 PET study. Journal of Nuclear Medicine, 55(6), 945–950.
- Naviaux, R.K., Naviaux, J.C., Li, K., et al. (2016). Metabolic features of chronic fatigue syndrome. Proceedings of the National Academy of Sciences USA, 113(37), E5472–E5480.
- Nedergaard, M. (2013). Garbage truck of the brain. Science, 340(6140), 1529–1530.
- Nelson, M.J., Bahl, J.S., Buckley, J.D., Thomson, R.L., & Davison, K. (2019). Evidence of altered cardiac autonomic regulation in ME/CFS: A systematic review and meta-analysis. Medicine, 98(43), e17600.
- Painter, R.C., Osmond, C., Gluckman, P., et al. (2008). Transgenerational effects of prenatal exposure to the Dutch famine on neonatal adiposity and health in later life. BJOG, 115(10), 1243–1249.
- Perroud, N., Paoloni-Giacobino, A., Prada, P., et al. (2011). Increased methylation of glucocorticoid receptor gene (NR3C1) in adults with a history of childhood maltreatment. Translational Psychiatry, 1(12), e59.
- Pietrzak, R.H., Gallezot, J.D., Ding, Y.S., et al. (2013). Association of PTSD with reduced in vivo norepinephrine transporter availability in the locus coeruleus. JAMA Psychiatry, 70(11), 1199–1205.
- Priovoulos, N., Jacobs, H.I.L., Ivanov, D., Uludag, K., Verhey, F.R.J., & Poser, B.A. (2018). High-resolution in vivo imaging of human locus coeruleus by magnetization transfer MRI at 3T and 7T. NeuroImage, 168, 427–436.
- Redgrave, J.N., Moore, L., Oyekunle, T., Ebrahim, M., Falidas, K., Snowdon, N., Ali, A., & Majid, A. (2022). Transcutaneous auricular vagus nerve stimulation with concurrent upper limb repetitive task practice for poststroke motor recovery: A pilot study. Journal of Stroke and Cerebrovascular Diseases, 27(7), 1998–2005.
- Reik, W., Dean, W., & Walter, J. (2001). Epigenetic reprogramming in mammalian development. Science, 293(5532), 1089–1093.
- Ridgewell, C., Heaton, K.J., Hildebrandt, A., Couse, J., Leber, D., & Neumeier, W.H. (2021). The effects of transcutaneous auricular vagal nerve stimulation on cognition in healthy individuals: A meta-analysis. Neuropsychology, 35(4), 352–365.
- Rodgers, A.B., Morgan, C.P., Bronson, S.L., et al. (2013). Paternal stress exposure alters sperm microRNA content and reprograms offspring HPA stress axis regulation. Journal of Neuroscience, 33(21), 9003–9012.
- Sara, S.J. (2009). The locus coeruleus and noradrenergic modulation of cognition. Nature Reviews Neuroscience, 10(3), 211–223.
- Sasaki, H., & Matsui, Y. (2008). Epigenetic events in mammalian germ-cell development: Reprogramming and beyond. Nature Reviews Genetics, 9(2), 129–140.
- Schwarz, L.A., & Luo, L. (2015). Organization of the locus coeruleus-norepinephrine system. Current Biology, 25(21), R1051–R1056.
- Shiffer, D., Rigo, S., Minonzio, M., Yarsuvat, D.T., Tobaldini, E., Furlan, L., Montano, N., Cairo, B., Porta, A., Zamunér, A.R., Bonovas, S., Urechie, V., Biaggioni, I., Diedrich, A., & Furlan, R. (2025). Short and long term effects of a two-week transcutaneous vagus nerve stimulation in hyperadrenergic postural orthostatic tachycardia syndrome: A proof-of-concept trial. European Journal of Internal Medicine. [CrossRef]
- Shungu, D.C., Weiduschat, N., Murrough, J.W., et al. (2012). Increased ventricular lactate in chronic fatigue syndrome. III. Relationships to cortical glutathione and clinical symptoms. NMR in Biomedicine, 25(9), 1073–1087.
- Stavrakis, S., Stoner, J.A., Humphrey, M.B., Morris, L., Filiberti, A., Reynolds, J.C., Elkholey, K., Javed, I., Twidale, N., Riha, P., Varahan, S., Scherlag, B.J., Jackman, W.M., Dasari, T.W., & Po, S.S. (2020). TREAT AF (Transcutaneous Electrical Vagus Nerve Stimulation to Suppress Atrial Fibrillation): A randomized clinical trial. JACC: Clinical Electrophysiology, 6(3), 282–291.
- Valentino, R.J., & Van Bockstaele, E. (2008). Convergent regulation of locus coeruleus activity as an adaptive response to stress. European Journal of Pharmacology, 583(2–3), 194–203.
- van Campen, C.L.M.C., Rowe, P.C., & Visser, F.C. (2020). Cerebral blood flow is reduced in severe ME/CFS patients during mild orthostatic stress testing. Healthcare, 8(2), 169.
- Van Houdenhove, B., Neerinckx, E., Lysens, R., et al. (2001). Victimization in chronic fatigue syndrome and fibromyalgia in tertiary care. Psychosomatics, 42(1), 21–28.
- Ventura-Bort, C., Garcia de Gurtubay, I., Bermejo, P., et al. (2023). Immediate effects and duration of a short and single application of transcutaneous auricular vagus nerve stimulation on P300 event related potential. Frontiers in Neuroscience, 17, 1096865.
- Villani, V., Tsakiris, M., & Azevedo, R.T. (2022). Transcutaneous vagus nerve stimulation modulates pupil size and interoceptive attention. Psychophysiology, 59(4), e13983.
- Warren, C.M., Tona, K.D., Ber, L., van den Brink, R.L., van Nieuwenhoven, L., de Geus, E.J.C., & Nieuwenhuis, S. (2019). The neuromodulatory and hormonal effects of transcutaneous vagus nerve stimulation as evidenced by salivary alpha amylase, salivary cortisol, pupil diameter, and the P3 event-related potential. Brain Stimulation, 12(3), 635–642.
- Wirth, K., & Scheibenbogen, C. (2020). A unifying hypothesis of the pathophysiology of ME/CFS: Insights from autoantibodies against beta2-adrenergic receptors. Autoimmunity Reviews, 19(6), 102527.
- Xie, L., Kang, H., Xu, Q., et al. (2013). Sleep drives metabolite clearance from the adult brain. Science, 342(6156), 373–377.
- Yakunina, N., Kim, S.S., & Nam, E.C. (2017). Optimization of transcutaneous vagus nerve stimulation using functional MRI. Neuromodulation: Technology at the Neural Interface, 20(3), 290–300.
- Yehuda, R., Daskalakis, N.P., Bierer, L.M., et al. (2016). Holocaust exposure induced intergenerational effects on FKBP5 methylation. Biological Psychiatry, 80(5), 372–380.
- Yehuda, R., & Lehrner, A. (2018). Intergenerational transmission of trauma effects: Putative role of epigenetic mechanisms. World Psychiatry, 17(3), 243–257.
- Young, J.L. (2013). Use of lisdexamfetamine dimesylate in treatment of executive functioning deficits and chronic fatigue syndrome: A double-blind, placebo-controlled study. Psychiatry Research, 207(1–2), 127–133.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



