
Submitted:
16 December 2025
Posted:
17 December 2025
You are already at the latest version
Abstract
Oxidative stress and inflammation play a key role in many diseases. This study evaluated the potential of bioactive compounds from Red-belted Bracket and Artist's Bracket mushrooms to mitigate these processes. Multi-step extraction yielded fractions with diversified composition (triterpenoids, polysaccharides) and bioactivities, including antioxidant properties and inhibition of pro-inflammatory enzymes. Both species were rich in triterpenoids: ethanolic extracts from Artist’s Bracket contained mainly ganoderenic and ganoderic acids (≈31 μg/g d.w.), while Red-belted Bracket extracts contained phenolic ac-ids (≈20 μg/g d.w., mainly vanillic and chebulic acids) and triterpenoids (≈73 μg/g d.w., mainly forpinic and formipinic acids). The highest radical-scavenging and reducing activities were observed in alkaline and ethanolic extracts. Lipoxygenase was inhibited only by ethanolic extracts, with IC₅₀ values of 0.93 mg d.w./ml for Artist's Bracket (mixed inhibition) and 0.62 mg d.w./ml for Red-belted Bracket (non-competitive). Artist’s Bracket was also a potent source of xanthine oxidase inhibitors acting competitively (IC₅₀ = 0.71, 1.39 and 2.06 mg d.w./ml for ethanolic, methanolic, and aqueous extracts), whereas Red-belted Bracket was less active (IC₅₀ = 3.84 mg d.w./ml, non-competitive). In conclusion, these mushrooms, particularly their ethanolic extracts, are promising sources of compounds with antioxidant and anti-inflammatory activities, acting as effective inhibitors of lipoxygenase and xanthine oxidase.
Keywords:
medicinal mushrooms
; triterpenoids
; antioxidant activity
; anti-inflammatory activity
; lipoxygenase
; enzymes inhibition
; xanthine oxidase
; multi-step extraction
1. Introduction
Mushrooms and their components play an important role in dietetics and pharmacology due to their unique chemical composition, which determines a range of health-promoting properties [1,2]. Numerous studies indicate that mushrooms exhibit antioxidant, anti-inflammatory, antidiabetic, and anticancer activities. This activity mainly results from the presence of various groups of bioactive compounds in the fruiting bodies, such as terpenoids, nucleotides, polyphenols, lipid derivatives, and functional polysaccharides. Mushroom polysaccharides, mainly β-glucans, modulate the immune system, supporting the body’s response to infectious and cancerous agents [3]. Terpenoids and polyphenols, in turn, show strong antioxidant properties, neutralizing free radicals and protecting cells from oxidative stress. Thanks to this broad spectrum of activity, mushrooms are a valuable component of a functional diet and a potential source of bioactive compounds for the development of new pharmaceutical preparations [4].
The etiology of many civilization-related diseases is directly linked to chronic oxidative stress and the accompanying inflammatory state. Inflammation is the body’s response to tissue damage, infections, or oxidative stress, and its course is driven by key chemical mediators, including pro-inflammatory cytokines (e.g., TNF-α, IL-1β, IL-6) and pro-inflammatory enzymes (e.g. LOX, XO). LOX catalyses the conversion of polyunsaturated fatty acids into eicosanoids, which intensify the inflammatory response, while XO generates reactive oxygen species (ROS) that damage tissues, thereby activating NF-κB and MAPK signalling pathways [5]. Regulating the activity of these enzymes is an important target of pharmacological anti-inflammatory therapy and also a significant area of interest for modern food technology (functional food development)[6,7]. In light of this, studying enzyme inhibition kinetics is a crucial step toward a deeper understanding of the mechanisms regulating enzyme activity. It also facilitates the optimization of enzymatic reactions and the design of effective inhibitors aimed at modulating anti-inflammatory effects in biological systems.
Red-belted bracket (Fomitopsis pinicola (Sw.) P. Karst.) and Artist’s bracket (Ganoderma applanatum (Pers.) Pat.) are rich in bioactive compounds, such as β-glucans (e.g. β-1,3 and β-1,3/1,6), terpenoids (e.g. ganoderic acids, lanostane-type triterpenoids), sterols (e.g. ergosterol, ergosta-7,22-dien-3β-ol) and phenolics (e.g p-hydroxybenzoic acid, gallic acid derivatives) [8,9,10,11,12,13,14]. Due to this unique composition, these mushrooms appear to be promising candidates for targeted therapies of diseases whose etiology and progression are associated with oxidative stress and inflammation. To date, it has been demonstrated that the ethyl acetate and ethanolic extracts of F. pinicola have high antioxidant properties (DPPH, ABTS, nitric oxide and OH tests scavenging activity and inhibition of lipid peroxidation) in vitro, as well as inhibit both acute and chronic inflammation induced with croton oil in a mouse model [15]. In turn, ethanolic extracts (70%) exhibit a potent free radical scavenging (DPPH, ABTS and OH tests) and effectively protect protein against oxidation [8]. Ganoderma triterpenoids attenuate atherosclerotic plaque formation in high-fat diet–fed rabbits by reducing oxidative stress and inflammation, as evidenced by decreased levels of reactive oxygen species (ROS) and malondialdehyde (MDA), resulting from the downregulation of nuclear transcription factor NF-κB p65 and the scavenger receptor LOX-1 [16]. Methanol extract and hot water extracts improve the survival rate of RAW 264.7 macrophages and inhibit the nitric oxide (NO)-mediated expression of inducible nitric oxide synthase (iNOS) protein after lipopolysaccharide treatment [17].
In the case of mushrooms, the active compounds belong to various chemical groups, which necessitates the use of complex extraction systems for their isolation. Typically, organic solvents (e.g. ethanol, methanol, ethyl acetate) are used to isolate terpenoids, sterols, peptides, monosaccharides and polyphenols, while polysaccharides are extracted using hot-water extraction (β-glucans) or alkaline solvents (α-glucans) [18,19,20,21,22]. Of course, the composition of the studied fraction strictly determines the activity, which has been confirmed in many studies [3,23]. Based on the above, in our study, a sequential extraction was used to efficiently recover diverse bioactive compounds, facilitate bioactivity testing, and identify compound groups responsible for specific biological effects.
This study aims to evaluate the antioxidant properties and the effects of extracts obtained through a multi-step extraction method on the activity of pro-inflammatory enzymes (LOX and XO), with a particular focus on the underlying mechanisms and the kinetics of the inhibition process.
2. Materials and Methods
2.1. Materials and Chemicals
2.1.1. Chemicals and Materials
All analytical grade chemicals (if not otherwise stated) were purchased from Merck /Sigma-Aldrich (Poznań, Poland). The mushrooms were collected, manually cleaned, freeze-dried (-42 °C, FreeZone 1 Liter, Labconco, USA), ground (6 °C, 2 × 1 min. MRC SM-450C), and stored in polypropylene boxes at −65 °C. Mushrooms were authenticated by Prof. Michał Świeca (Mushroom Classifier Certificate of Competency 1/RZ/2024) from the Department of Food Chemistry and Biochemistry, University of Life Sciences in Lublin, Poland. The samples are deposited at the Department of Food Chemistry and Biochemistry (voucher specimens: M_UPL_2024_18 for Artist’s bracket (Ganoderma applanatum (Pers.) Pat.) and M_UPL_2024_19 for Red-belted bracket (Fomitopsis pinicola (Sw.) P. Karst.). Additionally, the collected material was determined and identified in the microbiological laboratory of the Department of Biotechnology, Microbiology and Human Nutrition, University of Life Sciences in Lublin (Figure S1).
2.1.2. Preparation of Musroom Extracts
Powdered mushrooms were subjected to multi-step extraction. In the 1st step, 10 g of lyophilised powder was extracted with 0.6 L of 70% ethanol (30 °C for 60 min., constant shaking 60 rpm.), centrifuged (Centrifuge MPW-352R, 3997 × g, 10 min, 20 °C) and stored at - 65 °C (E1). Next, the residues were reextracted with 0.6 L of 50% methanol (30 °C for 60 min., constant shaking 60 rpm.), centrifuged (Centrifuge MPW-352R, 3997 × g, 10 min, 20 °C) and stored at - 65 °C (E2). The resulting residues were reextracted with 0.6 L water (95 °C for 120 min., constant shaking 60 rpm.), centrifuged (Centrifuge MPW-352R, 3997 × g, 10 min, 20 °C) and stored at - 65 °C (E3). Finally, the residues from the 3rd extraction were re-extracted with 2% NaOH (30 °C for 60 min., constant shaking 60 rpm.), centrifuged (Centrifuge MPW-352R, 3997 × g, 10 min, 20 °C), and after the pH adjustment to 7.0 (6 M HCl) stored at - 65 °C (E4).
2.2. Analytical Procedures
2.2.1. Analysis of Bioactive Compounds
2.2.1.1. The Folin-Ciocalteau Reacting Substances (FC-Reacting Substances)
The content of FC-reactive substances, including potential phenolics, was determined according to Singleton et al. [24] and expressed as gallic acid equivalents (GAE) in mg per g of dry weight (d.w.).
2.2.1.2. Total Terpenoids and Sterols
The sum of terpenoids and sterols was determined with the vanillin-glacial acetic acid solution and expressed as ursolic acid equivalents (UAE) in mg per g of dry weight (d.w.) [25].
2.2.1.3. Saccharides
Free saccharides and total polysaccharide contents were determined using the phenol-sulfuric acid method and expressed as glucose equivalents (GluE) in mg per g of dry weight (d.w.) [26].
2.2.1.4. Untargeted Metabolomics
Extracts (E1) were evaporated in a rotary evaporator (R-215 Rotavapor System, Buchi, Switzerland) at 40 ◦C. Concentrated samples were applied to a solid-phase extraction (SPE) a C18 Sep-Pak cartridge (Waters Associates), preconditioned with water. The cartridge was washed first with water to remove sugars. Phenolics and triterpenoids were eluted with MeOH, evaporated, and redissolved in MeOH (1 mL). For UPLC, 150 µL was transferred to another vial, and the solution was made up to a final volume of 1.5 mL with MeOH.
Determination of polyphenolic compounds and triterpenoids was carried out using the ultra-performance liquid chromatography (UPLC) Waters ACQUITY system (Waters, Milford, MA, USA) [27]. The UPLC system was equipped with a binary pump manager, column manager, sample manager, photodiode array (PDA) detector and tandem quadrupole mass spectrometer (TQD) with an electrospray ionisation (ESI) source. Separation of polyphenols was performed using a 1.7 µm, 100 mm × 2.1 mm UPLC BEH RP C18 column (Waters, USA). For the investigation, the mobile phase consisted of 0.1% formic acid in water, v/v (solvent A) and 0.1% formic acid in acetonitrile, v/v (solvent was used). The flow rate was kept constant at 0.35 mL/min for a total run time of 8 min. The system was run with the following gradient program: from 0 min 5% B, from 0 to 8 min linear to 100% B and from 8 to 9.5 min for washing and back to initial conditions. The injection volume of the samples was 5 µL, and the column was maintained at 50 °C. The following TQD parameters were used: cone voltage of 30 V, capillary voltage of 3500 V, source and desolvation temperature of 120 °C and 350 °C, respectively, and desolvation gas flow rate of 800 L/h. Characterisation of the individual polyphenolic compounds and triterpenoids was performed based on retention time, mass-to-charge ratio, fragment ions, and comparison with data obtained from commercial standards and literature findings [8,9,11,13,28,29,30,31,32,33]. The obtained data were processed in Waters MassLynx v.4.1 software (Waters, USA).
2.3.1. Antioxidant Properties
2.3.1.1. Antiradical Properties (ABTS)
The experiments were carried out using the ABTS decolourisation assay [34]. The free radical scavenging ability was expressed as Trolox equivalents in mg per g of dry weight (d.w.).
2.3.1.2. Ferric Reducing Power (RP)
Reducing power was determined according to Oyaizu [35]. It was expressed as Trolox equivalents in mg per g of dry weight (d.w.).
2.4.1. Anti-Inflammatory Properties
2.4.1.1. Ability to Inhibit Xanthine Oxidase Activity (LOXI)
The ability to inhibit lipoxygenase activity was measured using linoleic acid as a substrate using the Lipoxygenase Inhibitor Screening Assay Kit (No. 760700, Cayman Chemicals, Ann Arbour, MI, USA). For the inhibition studies, the enzyme was incubated for 10 min with 10 μL of the studied extract before adding the substrate. One unit (U) will cause an oxidation of 1 μmol of linoleic acid per minute at pH 7.5 at 30 °C. The activity is expressed in IU/g d.w., where IU is defined as an amount of inhibitor decreasing 1 U of enzyme activity [21].
2.4.1.2. Ability to Inhibit Xanthine Oxidase Activity (XOI)
The ability to inhibit xanthine oxidase activity was measured using xanthine as described previously [36]. The reaction mixture contained 120 μL of 1/15 M sodium-phosphate buffer, 20 μL of the enzyme xanthine oxidase (10 μL/mL, X1875 Sigma-Aldrich, Poland) and 20 μL of the substrate (0.015 mmol). The change of absorbance (3 min) was measured at 234 nm. For the inhibition studies, before adding the substrate, the enzyme was incubated for 10 min with 20 μL of the studied extract. One unit of activity converts 1.0 μmol of xanthine to uric acid per minute at pH 7.5 at 25 °C. The activity is expressed in IU/g d.w., where IU is defined as an amount of inhibitor decreasing 1 U of enzyme activity.
2.4.1.3. Determination of IC50 Values and the Mode of Inhibition.
To determine the IC₅₀ value, the relationship between the degree of inhibition of enzyme activity and the concentration of the extract (ranging from 0 to 1 mg d.w./mL) was established. The IC₅₀ values were expressed as mg of mushroom dry weight per 1 ml of the reaction mixture. The inhibition model and kinetic parameters of the process were determined using the inhibitor at the IC₅₀ concentration, according to the Lineweaver–Burk method [37].
2.5. Statistical Analysis
The distribution of the data was estimated using Shapiro–Wilk’s tests. Statistical analysis of the data was performed using Statistica 10 (StatSoft, Tulsa, OK, USA). The analysis of variance (one-way and two-way) and intergroup differences were used with Tukey’s HSD post hoc test with a significance level of P≤ 0.05.
3. Results and Discussion
3.1. Effect of Multi-Step Extraction on the Main Active Constituents and Antioxidant Properties
The mushrooms were subjected to a multistep extraction process aimed at effectively isolating different groups of metabolites potentially exhibiting antioxidant and anti-inflammatory properties (Table 1). In the case of the Artist’s bracket, the highest content of FC-reactive substances was determined after the first extraction step (54% of the total content), while a considerable amount was also detected following the alkaline extraction (23% of the total). Application of the multistep extraction procedure to the Red-belted bracket enabled the recovery of approximately 29 mg of FC-reactive substances from 1 g of the lyophilized sample. Notably, all applied conditions efficiently released these compounds from the mushroom matrix — in successive stages, 28%, 20%, 17%, and 35% of the total content were isolated. Both species proved to be excellent sources of triterpenoids and sterols, which were effectively extracted using 70% ethanol; however, the Red-belted bracket contained nearly three times higher amounts of these compounds than the Artist’s bracket. Both mushrooms contained comparable contents of simple saccharides (mono- and oligosaccharides extractable with organic solvent). Hot-water and alkali extraction steps release polysaccharides from mushrooms; nevertheless, Artist’s bracket was a significantly richer source of these compounds. In its case, the third and fourth extraction steps allowed the isolation of approximately 33% and 65% more of these compounds, respectively, compared to the Red-belted bracket.
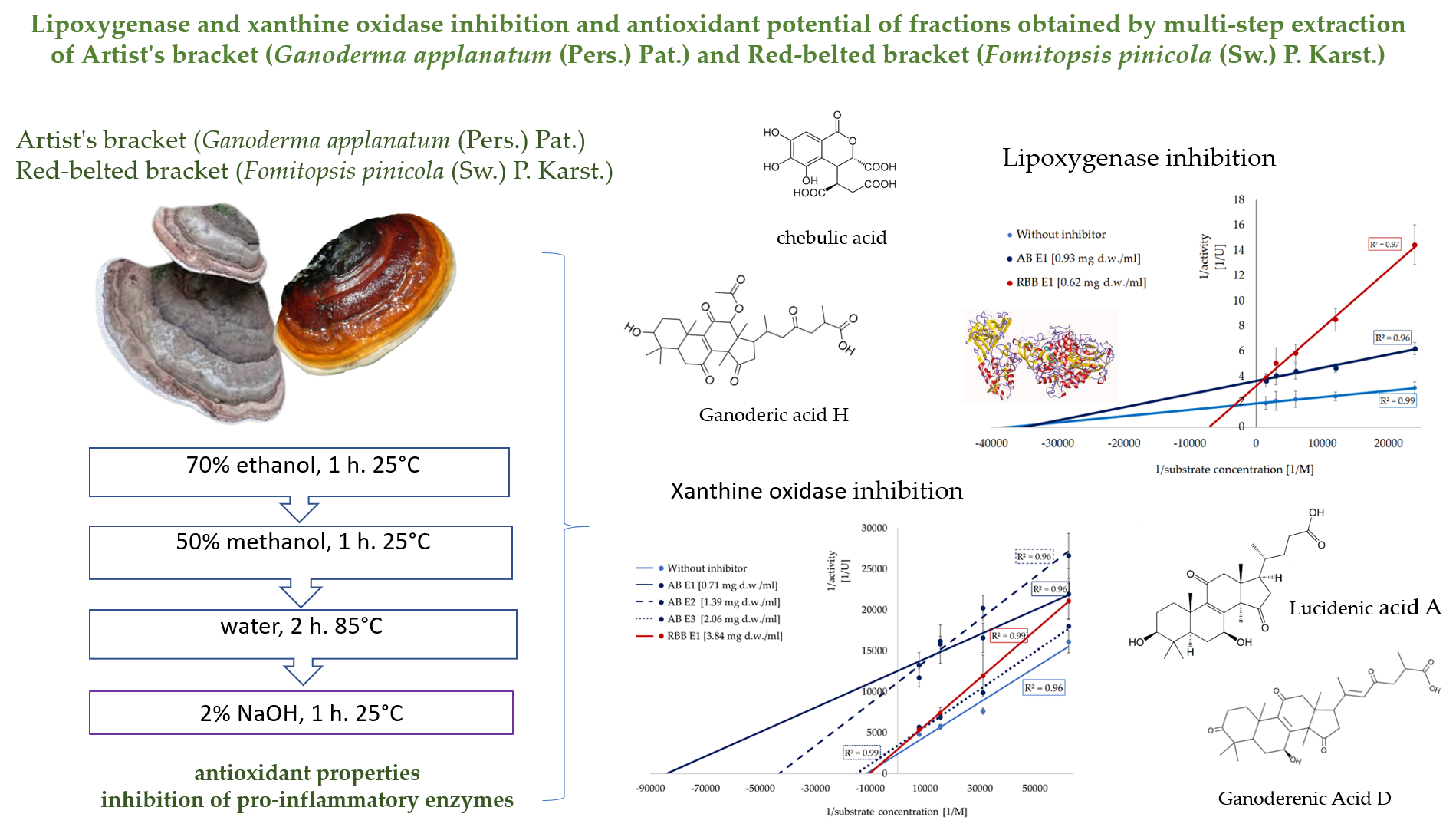
Literature data confirm that polypore is characterised by high antioxidant properties; thus, the obtained extracts were evaluated for their antiradical and reducing properties (Figure 1, Figure S2). The highest antiradical activities were observed in the alkaline and ethanolic fractions. For the Artist’s bracket, these fractions accounted for 35% and 51% of the total activity (sum of all extracts), respectively. Although the ethanolic fraction from the Red-belted bracket showed lower activity compared to that of the Artist’s bracket, the hot-water extract from this mushroom exhibited a considerable ability to quench ABTS radicals (17.2 mg TE/g d.w.) (Figure 1A). A similar trend, was observed in the case of reducing power, where the highest reducing properties were determined in the alkaline and ethanolic extracts from the Artist’s bracket (17.7 and 16.8 mg TE/ g d.w.) and alkaline extracts from the Red-belted bracket (17.5 mg TE/ g d.w.) (Figure 1B).
Both Artist’s bracket and Red-belted bracket are known to be a rich source of active substances, including triterpenoids, phenolics and polysaccharides. In our study, a multi-step extraction enabled us to isolate groups of metabolites with distinct chemical characteristics effectively. So far, for the extraction of phenolics and terpenoids from mushrooms, organic solvents have been used [38]. The application of methanol for the extraction of lyophilized fruiting bodies of Ganoderma applanatum yielded 6.71 mg GAE/g d.w. [12] and 20 mg GAE/g d.w. [39] of FC-reacting substances (potentially phenolics). A high extraction efficiency of terpenoids and phenolics with ethanol as a solvent also confirms the study by Nagadesi and Kannamba [40] , who compared this system with methanolic and cold-water maceration. In the mentioned study, the content of phenolics accounted for 85, 58 and 40 mg GAE/g d.w., respectively. What is important, they proved that both ethanol and 50% methanol used in our study in the 1st and 2nd steps of our extraction process were the most efficient for terpenoids isolation. Considering the Red-belted bracket, our results are consistent with those reported by Sułkowska-Ziaja et al. [14], who obtained 21.88 mg GAE/g d.w. using a Soxhlet apparatus for extraction. A comparable result was also reported by Onar et al. [41], where extraction with 70% ethanol yielded 27.8 mg GAE/g d.w. The use of 70% ethanol in the 1st extraction step in our study yielded 61.5 mg/g d.w. of triterpenes, which is 31% lower than the value reported by Zhang et al. [31]. Instead of differences in the origin of F. pinicola fruiting bodies, this variation may be attributed to the use of sonication to enhance the extractability.
The use of a sequence of polar solvents in the first and second stages of the multistage extraction likely resulted in the extraction of free sugars and oligosaccharides. This step is commonly applied during polysaccharide extraction as a preliminary purification of the research material to remove lipids, phenols, and triterpenes. Hot water extraction effectively isolates β-glucans, while alkaline extraction allows for removing polysaccharides poorly soluble in water (glycogen, alkali-soluble β-glucans, chitoglucans) or structural components of the cell wall that form complexes with proteins or lipids. The amount of sugars released with multistep extraction was c.a. 5-fold higher (102 vs 23 mg/g d.w) than in the study of Iranian Ganoderma applanatum [12]. The conditions applied in our study also resulted in a better yield of polysaccharides (70% higher) than in the study by Kozarsk et al. [42]. Moreover, the obtained extract was characterized by a significantly higher content of Folin-Ciocalteau-reacting substances (potentially polyphenols). Also, in the study conducted by Mohammadifar et al. [12], the total carbohydrate content in the ethanol extract of the Artist’s bracket (23 mg GluE/g d.w.) was more than two times lower than that determined in our study (60 mg GluE/g d.w.). As shown and discussed, the differences in the determined contents of active compounds are influenced not only by the extraction method but also by the environmental conditions and origin of the research material. Nevertheless, the validation of bioactive fractions is essential and plays a crucial role in the conscious application of mushroom extracts in food and medicine, as well as in predicting their health-promoting properties. Previously, Rašeta et al. [43] confirmed the effectiveness of extraction with 70% ethanol and hot water for obtaining fractions from G. applanatum with high antioxidant potential. In the studies, they associated the high ability to neutralize the DPPH radical with a high content of FC-reacting substances and total sugars. Moreover, the antiradical activity (ABTS assay) of the ethanolic and water extracts was approximately 4-fold and 8-fold higher, respectively, than in our study (160 vs 35 mg TE/g d.w. and 60 vs 7 mg TE/g d.w., respectively). Also, the antiradical activity (ABTS test) of G. applanatum extracts (ethanolic and hot-water) was significantly higher than in our study (9- and 8-fold, respectively)[44]. On the other hand, the results of reducing power (17.7 mg TE g d.w.) are nearly the same as those reported by Sułkowska-Ziaja et al. (15 mg TE g d.w.)[45]. The high reducing potential of the polysaccharide extract from Artist’s bracket (EC₅₀ = 0.18 mg/mL) and Red-belted bracket observed in our study is consistent with previous observations [42]. Although a direct comparison of the activity of extracts from the studied fungi is difficult due to methodological differences (extraction, test conditions, and extraction procedures), it should be emphasised that our results are consistent with those obtained by Li et al. [46]for the ABTS and DPPH assays, where F. pinicola was also found to be more effective.
3.2. Effect of Multi-Step Extraction on the Anti-Inflammatory Properties – Inhibition of Lipoxygenase and Xanthine Oxidase
Lipoxygenases (LOX) play a key role in oxidative stress and inflammatory processes so in the next step of the study, the obtained extracts were tested in terms of their ability to inhibit its activity (Figure 2). Only the extract from the 1st step were effective (Figure 2A). Both Artist’s bracket and Red-belted bracket had an inhibitory activity at the comparable level (a decrease of c.a. 65% of initial activity of enzyme); however, a detailed analysis showed that the Red-belted bracket is more effective. The studied extracts worked in a dose-dependent manner, which allowed for the determination of IC50 values (Figure 2B). The parameters, amounting respectively 0.93 and 0.62 mg d.w./ mL for Artist’s bracket and Red-belted bracket, were used in the further analysis of the kinetics of the inhibition process. According to the Lineweaver-Burk plot, the ethanolic extract from Artist’s bracket exhibited a mixed type of inhibition (Vmax was decreased, while Km was significantly increased). On the other hand, Red-belted bracket acted according to a non-competitive mode (Vmax was decreased, while Km was unchanged). It was reflected in a significant reduction of the turnover number (kcat), which was c.a. 50% lower than in the reaction without inhibitor (Table 2).
In turn, xanthine oxidase, which is one of the main sources of reactive oxygen species (ROS) in the body, was inhibited by both the ethanol extracts from the two mushrooms and by compounds from the methanolic extracts and the polysaccharide-rich fraction of Artist’s bracket (Figure 3A). Artist’s bracket turned out to be an excellent source of compounds inhibiting the activity of this enzyme, with IC₅₀ values 0.71, 1.39 and 2.06 mg d.w./ mL for the extracts obtained during the first three stages of isolation, respectively (Figure 3B). Compounds present in this extract act in an uncompetitive mode of action (Vmax and Km were decreased). The strength of inhibition was evidenced by a marked decrease in kcat values; in the presence of the ethanolic and methanolic fractions, it was reduced approximately fivefold compared to the reaction without the inhibitor. Red-belted bracket was less prominent, and only ethanolic extracts exhibited the inhibitory activity (IC50 3.84 mg d.w./ mL). The compounds present in this extract acted in a non-competitive mode of action, did not influence the affinity of the enzyme to the substrate (Km constant) (Figure 3C, Table 3).
The effectiveness of various mushrooms from the order Polyporales in modulating the activity of pro- and antioxidant enzymes has been confirmed in previous studies. LOX activity was inhibited exclusively by methanolic extracts (10 mg/ml) of Trametes hirsuta, Trametes orientalis, Daedaleopsis confragosa, and Roseofomes subflexibilis, which reduced its activity by 31.9%, 33.4%, 42.2%, and 43.4%, respectively. In the mentioned study, F. pinicola and F. nigra had the activity 33% and 43%, respectively [47]. They also proved that inoscavin A, isolated from Phellinus baumii, has a strong inhibitory activity against LOX, with an EC₅₀ value of 6.8 μM. Also, in the study by Nong et al. [48] concerning extracts from Ganoderma lucidum, was confirmed that ganoderic acids effectively inhibit the lipoxygenase activity. They demonstrated that the IC₅₀ values of ganoderic acids A, B, C2, D2, and F were 16.5, 6.9, 8.3, 9.3, and 14.3 μg/mL, respectively.
Similarly, to LOX inhibition, studies on XO activity inhibition are also scarce. In this area, particular emphasis should be placed on a study describing the activity of 47 native, wild Hungarian mushrooms [49]. The strongest XO inhibitory activity included Hypholoma fasciculare, Suillus grevillei, and Tricholoma populinum. The methanolic extract of Hypholoma fasciculare exhibited significant XO inhibition (77.67%, IC₅₀ = 67.76 μg/ml). Reported IC50 values were significantly lower than those determined in our study; however, it should be emphasized that in the cited studies, these values referred to the dried extract, whereas in the present study, they refer to the dry mass of the fungus from which the extract was obtained. The XO inhibitory activity of some species, Fistulina hepatica, Lentinus lepideus, Phellinus linteus, and Pleurotus cornucopiae, among others, has been confirmed [50,51,52]; unfortunately, the results cannot be directly compared due to differences in the methods (extract preparation and expression of activity). Nevertheless, the common denominator, despite the different extraction methods used, is the high activity of the fractions obtained using methanol or ethanol. Therefore, we attempted to determine the quantitative and qualitative composition of the ethanolic extract used in this study (Table 4).
Red-belted bracket contained two main groups of active compounds: phenolic and terpenoids. Among phenolics, vanillic acid and chebulic acids (benzopyran tannin) were dominant. The determined profile of phenolic acids is similar to that recorded previously by Sułkowska-Ziaja et al. [45]; however, the amounts are significantly lower, which may result from the applied extraction conditions – in the cited study, the material was subjected to acid hydrolysis, which made it possible to isolate bound phenolic acids. The presence of phenolic acid, including protocatechuic acid, p-hydroxybenzoic acid and gallic acid in this species was also confirmed [53]. The cited study also reported a significant amount of gallic and ellagic acid (not detected in our samples); however, this may be due to the detection conditions used, as chebulic acids and gallic acids exhibit similar ion-fragmentation patterns. In our extract, a significant amount of terpenoids was also detected, with dominant forpinic acids, which is in line with the previous studies [8,11,31]. In the studied extracts of Artist’s bracket, only ganoderenic acids were detected, which was in line with the previous studies concerning fingerprinting [28] and quality evaluation of Ganoderma Spp.[13]. We have not detected phenolic compounds, which are commonly reported for this genus (also in our study), using the unspecific FC-test [53,54]. This group of functional metabolites has also been identified using more sophisticated techniques (e.g., LC-MS/MS); however, their levels were marginal [53] and seem to be directly related to compounds absorbed from wood rather than synthesised de novo.
The detailed identification of the extract composition allowed for a deeper discussion of the prooxidative enzymes inhibition process. Some previous studies showed that the activity of lipoxidgenase and xanthine oxidase may be effectively inhibited by both phenolics and terpenoids. In the study by Łyko et al. [55] dried terpenoid-rich extracts of Rhododendron luteum obtained after supercritical fluid CO2 (30% ethanol) extraction inhibited LOX and XOI activity according to a competitive mode of action with IC50 values of 0.5 and 2.36 mg/ml, respectively. This activity is slightly higher than that determined in our study for the ethanolic extract, but it should be noted that in this study, IC50 values were expressed per g of dry mass of mushrooms used for the extraction. Also, ferulic, syringic and trans-cinnamic acids [56] and rutin [57] acted as inhibitors of LOX, with an uncompetitive mode of action determined for phenolic acids. Huang et al. [58] showed that terpenoids from Pistacia chinensis leaf inhibit xanthine oxidase according to a mixed type of inhibition. So far, the reports on the modulation of enzyme activity by mushroom components are scarce, especially describing the kinetics of the process, but generally confirm that terpenoid fractions are the most active. Previously, sesquiterpenoids from Fomitopsis pinicola exhibited the most significant inhibition of superoxide anion generation and elastase release with IC50 values of 0.81 and 0.74 mM [59].
The ethyl acetate extracts of Cordyceps militaris possessed the highest XO inhibitory properties, and it was suggested that the activity is responsible for lipid derivatives (pentadecanal, hexadecanoic acid, methyl 2-oxohexadecanoate and N-(2-Hydroxyethyl) octanamide)) and 3`-deoxyadenosine (cordycepin) – after fractionation, the lowest IC50 (68 μg of dried extract/ml) was determined for the 10th fraction containing tetradecanal, pentadecanal and 3`-deoxyadenosine. Amagata et al. [60] suggested that sponge-derived terpenoids, being LOX inhibitors, acted with redox (operate by reducing lipoxygenase to its inactive, ferrous form) and non-redox (enzyme inactivation occurs by competitive or allosteric inhibition) mechanisms. It may be suggested that, in the case of the ethanolic extracts of F. pinicola and G. applanatum, the second mechanism is involved; however, an unambiguous statement is difficult to make because the samples examined are mixtures of multiple compounds. A similar situation is observed in the case of XO inhibition, where uncompetitive inhibition has been identified. In turn, tsugaric acid (lanostanoids) isolated from the fruit bodies of Ganoderma tsugae exhibited significant inhibitory effects on xanthine oxidase (XO) activity with an IC50 value ranging from 90.2 to 182 μM [61]. The purified tsugaric acid D (3α-acetoxy-22-oxo-5α-lanosta-8,24-dien-21-oic acid) inhibited XOI in a dose-dependent manner according to a competitive mode (Ki 0,6 μM). They suggested that, in addition to the chemical skeleton and conformation of the compound, a carbonyl group, including carbonyl in carboxylic acid, acyl group or ester group, may interfere with the interaction between the enzyme and the enzyme-substrate complex. Due to a structural similarity of tsugaric acid D to forpinic acids from red-belted bracket and ganodermic acids from Artist’s bracket, an analogical mechanism of inhibitory action may be postulated. This is also confirmed by the screening study of potential XO inhibitors from G. leucocontextum using the affinity ultrafiltration method [62]. Triterpenoids, including ganoderic acid A, ganoderic acid D, ganodermanontriol, and ganoderal A, demonstrated a significant affinity to native XO, the mechanism was described by molecular docking analysis. They proved that among the studied compounds, ganodermic acids had the highest affinity to the enzyme (intermolecular energy, approximately -9.5 kcal/mol) and interact with the enzyme via hydrophobic interaction, hydrogen bonding, and salt bridges. Ganodermic acid A interacts with a region of the enzyme that does not directly participate in catalysis (I264, R394, K395). In contrast, ganodermic acid D appears to modify the catalytic activity of the molybdenum domain (Mo-cofactor domain) and to interact with amino acids that stabilize the structure and charge of the active site (W336, F337, A338, I358, and D360). The Lineweaver-Burk double reciprocal mapping method showed that, similarly to our studies, ganodermic acid acts as a non-competitive inhibitor with Ki 84 μg/ml (~163 µM).
4. Conclusions
The results confirmed that Red-belted bracket and Artist’s bracket possess significant antioxidant and anti-inflammatory properties, which may support the use of these medicinal mushrooms as part of anti-gout therapy. Importantly, the activity is strongly determined by the extraction method used, which is due to the fact that different groups of active compounds (terpenoids, polysaccharides) require distinct processing conditions. It was also demonstrated that commonly used spectrophotometric methods, although useful for raw material validation, provide incomplete information regarding the composition of active fractions. As shown, the highest activity was observed in the ethanolic fraction, which, in the case of Red-belted bracket, contained both polyphenols and terpenoids, whereas in Artist’s bracket, only terpenoids. The study also provides valuable insights into the kinetics of pro-oxidative enzyme inhibition. The mixture of Artist’s bracket terpenoids acts as a non-competitive inhibitor of LOX and XO. In contrast, the ethanolic extract of Red-belted bracket inhibited LOX via a mixed-type mechanism, while XO was inhibited through an acompetitive mechanism. Given the promising results, particularly regarding the inhibition of pro-oxidant enzymes, in the future, we will attempt to determine whether all compounds contribute equally to the observed activity or if one of them plays a dominant role. On the other hand, the information regarding the activity of the whole mixture, combined with the low costs of obtaining the extracts, opens new possibilities for the application of these mushrooms in food technology, including in the prevention of lifestyle-related diseases.
Supplementary Materials
The following supporting information can be downloaded at: The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1. Genetic identification of mushrooms, Figure S2. Radical scavenging activity-kinetics.
Author Contributions
Conceptualisation, M.Ś. and I.K.; methodology, M.Ś., P.J. and I.K.; software, P.J. and I.K.; validation, M.Ś., and I.K.; formal analysis, M.Ś. and I.K.; investigation, M.Ś., A.M., K.L., M.S., W.M., B.J., P.J., N.Ż. and, I.K.; resources, M.Ś.; data curation, M.Ś., P.J., N.Ż. and, I.K.; writing—original draft preparation, M.Ś., A.M. and, I.K.; writing—review and editing, M.Ś., A.M., K.L., M.S., W.M., B.J., P.J., N.Ż. and, I.K.; visualisation, A.M. and N.Ż.; supervision, M.Ś. and, I.K.; project administration, M.Ś.; funding acquisition, M.Ś. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Centre, Poland (NCN), Grant OPUS No. 2022/45/B/NZ9/01892.
Data Availability Statement
The data that support the findings are available in RepOD.
Acknowledgments
The authors would like to thank Urszula Jakubczak, Weronika Melona and Bartłomiej Janzer for their invaluable help and assistance during laboratory work.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bell, V.; Silva, C.R.P.G.; Fernandes, T.H. Mushrooms as Future Generation Healthy Foods. Front Nutr 2022. [Google Scholar] [CrossRef]
- Łysakowska, P.; Sobota, A.; Wirkijowska, A. Medicinal Mushrooms: Their Bioactive Components, Nutritional Value and Application in Functional Food Production—A Review. Molecules 2023, 28. [Google Scholar] [CrossRef]
- Kumar, K.; Mehra, R.; Guiné, R.P.F.; Lima, M.J.; Kumar, N.; Kaushik, R.; Ahmed, N.; Yadav, A.N.; Kumar, H. Edible Mushrooms: A Comprehensive Review on Bioactive Compounds with Health Benefits and Processing Aspects. Foods 2021, 10, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Michalska, A.; Sierocka, M.; Drzewiecka, B.; Świeca, M. Antioxidant and Anti-Inflammatory Properties of Mushroom-Based Food Additives and Food Fortified with Them—Current Status and Future Perspectives. Antioxidants 2025, 14. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S. Oxidative Stress, Inflammation, and Disease; Elsevier Inc., 2016; ISBN 9780128032701. [Google Scholar]
- Hu, C.; Ma, S. Recent Development of Lipoxygenase Inhibitors as Anti-Inflammatory Agents. Medchemcomm 2018, 9, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, D.; Ghosh, D.; Singh, R.; Dixit, S.; Chopra, S.; Tripathi, S.; Saini, T. Medicinal Mushrooms: Bioactive Components, Pharmacological, Immunological and Toxicological Insights. ChemistrySelect 2025, 10. [Google Scholar] [CrossRef]
- Zhang, Z. fa; Wu, C.; Wang, M.; Chen, J. fei; Lv, G. ying Chemical Fingerprinting and the Biological Properties of Extracts from Fomitopsis Pinicola. Arabian Journal of Chemistry 2023, 16. [Google Scholar] [CrossRef]
- Rosecke, J.; Konig, W.A. Steroids from the Fungus Fomitopsis Pinicola. Phytochemistry 1999, 1621–1627. [Google Scholar] [CrossRef]
- Kuo, P.C.; Tai, S.H.; Hung, C.C.; Hwang, T.L.; Kuo, L.M.; Lam, S.H.; Cheng, K.C.; Kuo, D.H.; Hung, H.Y.; Wu, T.S. Antiinflammatory Triterpenoids from the Fruiting Bodies of Fomitopsis Pinicola. Bioorg Chem 2021, 108. [Google Scholar] [CrossRef]
- Yu, H.; Chen, Q.; Xu, T.C.; Wang, Y.; Xu, W.F.; Li, M.; Zhang, X.; Zhao, C.; Zhang, D.L.; Jin, P.F.; et al. Bioactive Terpenoids and Sterols from the Fruiting Bodies of Fomitopsis Pinicola. Phytochemistry 2025, 236. [Google Scholar] [CrossRef]
- Mohammadifar, S.; Fallahi Gharaghoz, S.; Reza, M.; Shayan, A.; Vaziri, A. Comparison between Antioxidant Activity and Bioactive Compounds of Ganoderma Applanatum (Pers.) Pat. and Ganoderma Lucidum (Curt.) P. Karst from Iran. Vol. 11.
- Liu, L.; Wei, L.; Xu, J.; Xiong, H.; Zhou, S.; Ma, S.; Sun, W.; Tian, L.; Li, Z.; Xu, Z. Molecular Identification and Quality Evaluation of Commercial Ganoderma. Medicinal Plant Biology 2025, 4. [Google Scholar] [CrossRef]
- Sułkowska-Ziaja, K.; Zengin, G.; Gunia-Krzyżak, A.; Popiół, J.; Szewczyk, A.; Jaszek, M.; Rogalski, J.; Muszyńska, B. Bioactivity and Mycochemical Profile of Extracts from Mycelial Cultures of Ganoderma Spp. Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Sankaranarayanan Ravikumar, K.; Ramya, H.; Ajith, T.A.; Janardhanan, K.K.; Shah, A.; Reshi, Z.A.; Andrabi, K.I. ANTIOXIDANT AND ANTI-INFLAMMATORY ACTIVITIES OF BIOACTIVE EXTRACTS OF A POLYPORE MUSHROOM, FOMITOPSIS PINICOLA (SW.:FR.) P. KARST, FROM KASHMIR. Int. J. Adv. Res 6 1144–1156. [CrossRef]
- Li, Y.; Tang, J.; Gao, H.; Xu, Y.; Han, Y.; Shang, H.; Lu, Y.; Qin, C. Ganoderma Lucidum Triterpenoids and Polysaccharides Attenuate Atherosclerotic Plaque in High-Fat Diet Rabbits. Nutrition, Metabolism and Cardiovascular Diseases 2021, 31, 1929–1938. [Google Scholar] [CrossRef]
- Im, K.H.; Choi, J.; Baek, S.A.; Lee, T.S. Journal of Mushrooms 한 국 버 섯 학 회 지 Antioxidant and Inflammation Inhibitory Effects from Fruiting Body Extracts of Ganoderma Applanatum. J. Mushrooms 2021, 261–271. [Google Scholar] [CrossRef]
- Bishop, K.S. Characterisation of Extracts and Anti-Cancer Activities of Fomitopsis Pinicola. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Cheng, J.J.; Lin, C.Y.; Lur, H.S.; Chen, H.P.; Lu, M.K. Properties and Biological Functions of Polysaccharides and Ethanolic Extracts Isolated from Medicinal Fungus, Fomitopsis Pinicola. Process Biochemistry 2008, 43, 829–834. [Google Scholar] [CrossRef]
- Kamiyama, M. Antioxidant/Anti-Inflammatory Activities and Chemical Composition of Extracts from the Mushroom Trametes Versicolor. International Journal of Nutrition and Food Sciences 2013, 2, 85. [Google Scholar] [CrossRef]
- Michalska, A.; Szymanowska, U.; Kapusta, I.; Żurek, N.; Nawrocka, A.; Różyło, R.; Jarocki, P.; Świeca, M. Antioxidant, Anti-Inflammatory, and Antiproliferative Properties of Extracted Turkey Tail (Trametes Versicolor Lloyd) Mushroom Components Microencapsulated with Inulin. Food Chem 2025, 495. [Google Scholar] [CrossRef]
- Karunarathna, S.C.; Patabendige, N.M.; Kumla, J.; Hapuarachchi, K.K.; Suwannarach, N. The Bioactive Compounds, Beneficial Medicinal Properties, and Biotechnological Prospects of Fomitopsis: A Comprehensive Overview. Front Cell Infect Microbiol 2025, 15. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, D.; Ghosh, D.; Singh, R.; Dixit, S.; Chopra, S.; Tripathi, S.; Saini, T. Medicinal Mushrooms: Bioactive Components, Pharmacological, Immunological and Toxicological Insights. ChemistrySelect 2025, 10. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. Methods Enzymol 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Chang, C.L.; Lin, C.S.; Lai, G.H. Phytochemical Characteristics, Free Radical Scavenging Activities, and Neuroprotection of Five Medicinal Plant Extracts. Evidence-based Complementary and Alternative Medicine 2012, Article ID. [Google Scholar] [CrossRef]
- Gao, L.; Wang, Y.; Zhang, F.; Li, S.; Zhao, J.; Zhang, Q.; Ye, J.; Ma, Y.; Wang, Z.; Chen, W. A Standardized Method for the Quantification of Polysaccharides: An Example of Polysaccharides from Tremella Fuciformis. Lwt 2022, 167, 113860. [Google Scholar] [CrossRef]
- Lachowicz-Wiśniewska, S.; Kapusta, I.; Stinco, C.M.; Meléndez-Martínez, A.J.; Bieniek, A.; Ochmian, I.; Gil, Z. Distribution of Polyphenolic and Isoprenoid Compounds and Biological Activity Differences between in the Fruit Skin + Pulp, Seeds, and Leaves of New Biotypes of Elaeagnus Multiflora Thunb. Antioxidants 2021, 10, 1–18. [Google Scholar] [CrossRef]
- Biswal, R.P.; Dandamudi, R.B.; Patnana, D.P.; Pandey, M.; Vutukuri, V.N.R.K. Metabolic Fingerprinting of Ganoderma Spp. Using UHPLC-ESI-QTOF-MS and Its Chemometric Analysis. Phytochemistry 2022, 199. [Google Scholar] [CrossRef]
- Elkhateeb, W.A.; Zaghlol, G.M.; El-Garawani, I.M.; Ahmed, E.F.; Rateb, M.E.; Abdel Moneim, A.E. Ganoderma Applanatum Secondary Metabolites Induced Apoptosis through Different Pathways: In Vivo and in Vitro Anticancer Studies. Biomedicine and Pharmacotherapy 2018, 101, 264–277. [Google Scholar] [CrossRef]
- Ravikumar, K.S.; Ramya, H.; Ajith, T.A.; Shah, M.A.; Janardhanan, K.K. Bioactive Extract of Fomitopsis Pinicola Rich in 11-α- Acetoxykhivorin Mediates Anticancer Activity by Cytotoxicity, Induction of Apoptosis, Inhibition of Tumor Growth, Angiogenesis and Cell Cycle Progression. J Funct Foods 2021, 78. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, B.; Liang, J.; Han, J.; Zhou, L.; Zhao, R.; Liu, H.; Dai, H. Lanostane Triterpenoids with PTP1B Inhibitory and Glucose-Uptake Stimulatory Activities from Mushroom Fomitopsis Pinicola Collected in North America. J Agric Food Chem 2020, 68, 10036–10049. [Google Scholar] [CrossRef]
- Oludemi, T.; Barros, L.; Prieto, M.A.; Heleno, S.A.; Barreiro, M.F.; Ferreira, I.C.F.R. Extraction of Triterpenoids and Phenolic Compounds from: Ganoderma Lucidum: Optimization Study Using the Response Surface Methodology. Proceedings of the Food and Function 2018, Vol. 9, 209–226. [Google Scholar] [CrossRef] [PubMed]
- Kiseleva, I.; Ermoshin, A.; Galishev, B.; Shen, D. Chemical Composition and Antioxidant Activity of Fomitopsis Pinicola Growing on Coniferous and Deciduous Substrates.
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic Biol Med 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Oyaizu, M. Studies on Products of Browning Reaction – Antioxidative Activities of Products of Browning Reaction Prepared from Glucosamine. Japanese Journal of Nutrition 1986, 44, 307–315. [Google Scholar]
- Gawlik-Dziki, U.; Dziki, D.; Świeca, M.; Nowak, R. Mechanism of Action and Interactions between Xanthine Oxidase Inhibitors Derived from Natural Sources of Chlorogenic and Ferulic Acids. Food Chem 2017, 225, 138–145. [Google Scholar] [CrossRef]
- Lineweaver, H.; Burk, D. The Determination of Enzyme Dissociation Constants. J Am Chem Soc 1934, 56, 658–666. [Google Scholar] [CrossRef]
- Agnihotri, C.; Singh, B.P.; Kamal, S.; Ram, H.; Oberoi, H.S.; Mostashari, P.; Mousavi Khaneghah, A. Advancements in Extraction and Encapsulation of Immunomodulatory Mushroom Biomolecules for Enhanced Food Applications. Applied Food Research 2025, 5. [Google Scholar] [CrossRef]
- Zengin, G.; Sarikurkcu, C.; Gunes, E.; Uysal, A.; Ceylan, R.; Uysal, S.; Gungor, H.; Aktumsek, A. Two Ganoderma Species: Profiling of Phenolic Compounds by HPLC-DAD, Antioxidant, Antimicrobial and Inhibitory Activities on Key Enzymes Linked to Diabetes Mellitus, Alzheimer’s Disease and Skin Disorders. Food Funct 2015, 6, 2794–2802. [Google Scholar] [CrossRef]
- Kumar Nagadesi, P.; Kannamba, B. Effect of Extraction and Solvent on Mycochemicals and Proximate Composition of Lignicolous Fungi Ganoderma Lucidum and G. Applanatum: A Comparative Study. SCIREA Journal of Pharmacy 2019, 3, 1–24. [Google Scholar]
- Onar, O.; Akata, I.; Celep, G.S.; Yildirim, O. Antioxidant Activity of Extracts from the Red-Belt Conk Medicinal Mushroom, Fomitopsis Pinicola (Agaricomycetes), and Its Modulatory Effects on Antioxidant Enzymes. Int J Med Mushrooms 2016, 18, 501–508. [Google Scholar] [CrossRef]
- Kozarski, M.; Klaus, A.; Nikšić, M.; Vrvić, M.M.; Todorović, N.; Jakovljević, D.; Van Griensven, L.J.L.D. Antioxidative Activities and Chemical Characterization of Polysaccharide Extracts from the Widely Used Mushrooms Ganoderma Applanatum, Ganoderma Lucidum, Lentinus Edodes and Trametes Versicolor. Journal of Food Composition and Analysis 2012, 26, 144–153. [Google Scholar] [CrossRef]
- Rašeta, M.; Popović, M.; Beara, I.; Šibul, F.; Zengin, G.; Krstić, S.; Karaman, M. Anti-Inflammatory, Antioxidant and Enzyme Inhibition Activities in Correlation with Mycochemical Profile of Selected Indigenous Ganoderma Spp. from Balkan Region (Serbia). Chem Biodivers 2021, 18. [Google Scholar] [CrossRef]
- Rašeta, M.; Karaman, M.; Jakšić, M.; Šibul, F.; Kebert, M.; Novaković, A.; Popović, M. Mineral Composition, Antioxidant and Cytotoxic Biopotentials of Wild-Growing Ganoderma Species (Serbia): G. Lucidum (Curtis) P. Karst vs. G. Applanatum (Pers.) Pat. Int J Food Sci Technol 2016, 51, 2583–2590. [Google Scholar] [CrossRef]
- Sułkowska-Ziaja, K.; Muszyńska, B.; Motyl, P.; Pasko, P.; Ekiert, H. Phenolic Compounds and Antioxidant Activity in Some Species of Polyporoid Mushrooms from Poland. 2012, Vol. 14. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Y.; Li, S.-Y.; Yin, F.; Chen, H.-M.; Yang, D.-F.; Liu, X.-Q.; Jin, Q.-H.; Lv, X.-M.; Mans, D.; Zhang, X.-D.; et al. Antioxidative and Cytoprotective Effects of Ganoderma Applanatum and Fomitopsis Pinicola in PC12 Adrenal Phaeochromocytoma Cells. Int J Med Mushrooms 2022, 24, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Song, J.G.; Hwang, B.S.; Kim, D.W.; Lee, Y.J.; Woo, E.E.; Kim, J.Y.; Lee, I.K.; Yun, B.S. Lipoxygenase Inhibitory Activity of Korean Indigenous Mushroom Extracts and Isolation of an Active Compound from Phellinus Baumii. Mycobiology 2014, 42, 185–188. [Google Scholar] [CrossRef]
- Nong, Y.; Zhou, X.; Li, S.; Liu, Q.; Zhang, Y.; Liang, J.; Zhang, Y.; Liu, C. Efficient and Fast Screening and Separation Based on Computer-aided Screening and Complex Chromatography Methods for Lipoxygenase Inhibitors from. Phytochemical Analysis 2024, 35, 599–616. [Google Scholar] [CrossRef] [PubMed]
- Ványolõs, A.; Orbán-Gyapai, O.; Hohmann, J. Xanthine Oxidase Inhibitory Activity of Hungarian Wild-Growing Mushrooms. Phytotherapy Research 2014, 28, 1204–1210. [Google Scholar] [CrossRef]
- Song, Y.S.; Kim, S.H.; Sa, J.H.; Jin, C.; Lim, C.J.; Park, E.H. Anti-Angiogenic, Antioxidant and Xanthine Oxidase Inhibition Activities of the Mushroom Phellinus Linteus. J Ethnopharmacol 2003, 88, 113–116. [Google Scholar] [CrossRef]
- Ribeiro, B.; Valentão, P.; Baptista, P.; Seabra, R.M.; Andrade, P.B. Phenolic Compounds, Organic Acids Profiles and Antioxidative Properties of Beefsteak Fungus (Fistulina Hepatica). Food and Chemical Toxicology 2007, 45, 1805–1813. [Google Scholar] [CrossRef]
- Alam, N.; Yoon, K.N.; Lee, K.R.; Kim, H.Y.; Shin, P.G.; Cheong, J.C.; Yoo, Y.B.; Shim, M.J.; Lee, M.W.; Lee, T.S. Assessment of Antioxidant and Phenolic Compound Concentrations as Well as Xanthine Oxidase and Tyrosinase Inhibitory Properties of Different Extracts of Pleurotus Citrinopileatus Fruiting Bodies. Mycobiology 2011, 39, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Kozarski, M.; Klaus, A.; Špirović-Trifunović, B.; Miletić, S.; Lazić, V.; Žižak, Ž.; Vunduk, J. Bioprospecting of Selected Species of Polypore Fungi from the Western Balkans. Molecules 2024, 29. [Google Scholar] [CrossRef]
- Kozarski, M.; Klaus, A.; Nikšić, M.; Vrvić, M.M.; Todorović, N.; Jakovljević, D.; Van Griensven, L.J.L.D. Antioxidative Activities and Chemical Characterization of Polysaccharide Extracts from the Widely Used Mushrooms Ganoderma Applanatum, Ganoderma Lucidum, Lentinus Edodes and Trametes Versicolor. Journal of Food Composition and Analysis 2012, 26, 144–153. [Google Scholar] [CrossRef]
- Łyko, L.; Olech, M.; Gawlik, U.; Krajewska, A.; Kalemba, D.; Tyśkiewicz, K.; Piórecki, N.; Prokopiv, A.; Nowak, R. Rhododendron Luteum Sweet Flower Supercritical CO2 Extracts: Terpenes Composition, Pro-Inflammatory Enzymes Inhibition and Antioxidant Activity. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Habza-Kowalska, E.; Kaczor, A.A.; Bartuzi, D.; Piłat, J.; Gawlik-Dziki, U. Some Dietary Phenolic Compounds Can Activate Thyroid Peroxidase and Inhibit Lipoxygenase-Preliminary Study in the Model Systems. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Bouriche, H.; Miles, E.A.; Selloum, L.; Calder, P.C. Effect of Cleome Arabica Leaf Extract, Rutin and Quercetin on Soybean Lipoxygenase Activity and on Generation of Inflammatory Eicosanoids by Human Neutrophils. Prostaglandins Leukot Essent Fatty Acids 2005, 72, 195–201. [Google Scholar] [CrossRef]
- Huang, C.Y.; Chang, Y.Y.; Chang, S.T.; Chang, H.T. Xanthine Oxidase Inhibitory Activity and Chemical Composition of Pistacia Chinensis Leaf Essential Oil. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef]
- Tai, S.H.; Kuo, P.C.; Hung, C.C.; Lin, Y.H.; Hwang, T.L.; Lam, S.H.; Kuo, D.H.; Wu, J. Bin; Hung, H.Y.; Wu, T.S. Bioassay-Guided Purification of Sesquiterpenoids from the Fruiting Bodies of: Fomitopsis Pinicola and Their Anti-Inflammatory Activity. RSC Adv 2019, 9, 34184–34195. [Google Scholar] [CrossRef]
- Amagata, T.; Whitman, S.; Johnson, T.A.; Stessman, C.C.; Loo, C.P.; Lobkovsky, E.; Clardy, J.; Crews, P.; Holman, T.R. Exploring Sponge-Derived Terpenoids for Their Potency and Selectivity against 12-Human, 15-Human, and 15-Soybean Lipoxygenases. J Nat Prod 2003, 66, 230–235. [Google Scholar] [CrossRef]
- Lin, K.W.; Chen, Y.T.; Yang, S.C.; Wei, B.L.; Hung, C.F.; Lin, C.N. Xanthine Oxidase Inhibitory Lanostanoids from Ganoderma Tsugae. Fitoterapia 2013, 89, 231–238. [Google Scholar] [CrossRef]
- Nong, Y.; Liu, Q.; Li, S.; Liang, J.; Zhang, Y.; Zhang, Y.; Hou, W. Comprehensive Screening, Separation, Extraction Optimization, and Bioactivity Evaluation of Xanthine Oxidase Inhibitors from Ganoderma Leucocontextum. Arabian Journal of Chemistry 2025, 18, 132024. [Google Scholar] [CrossRef]
Figure 1.
Antioxidant potential of active fraction from subsequent extraction of mushrooms. Data are mean (n= 9) ± SD. Values with the same letters are not significantly different (p ≤ 0.05). TE- Trolox equivalents; d.w.- dry mass. E1- 70% ethanol extraction, E2- 50% methanol extraction, E3- hot-water extraction, E4- NaOH extraction.
Figure 1.
Antioxidant potential of active fraction from subsequent extraction of mushrooms. Data are mean (n= 9) ± SD. Values with the same letters are not significantly different (p ≤ 0.05). TE- Trolox equivalents; d.w.- dry mass. E1- 70% ethanol extraction, E2- 50% methanol extraction, E3- hot-water extraction, E4- NaOH extraction.
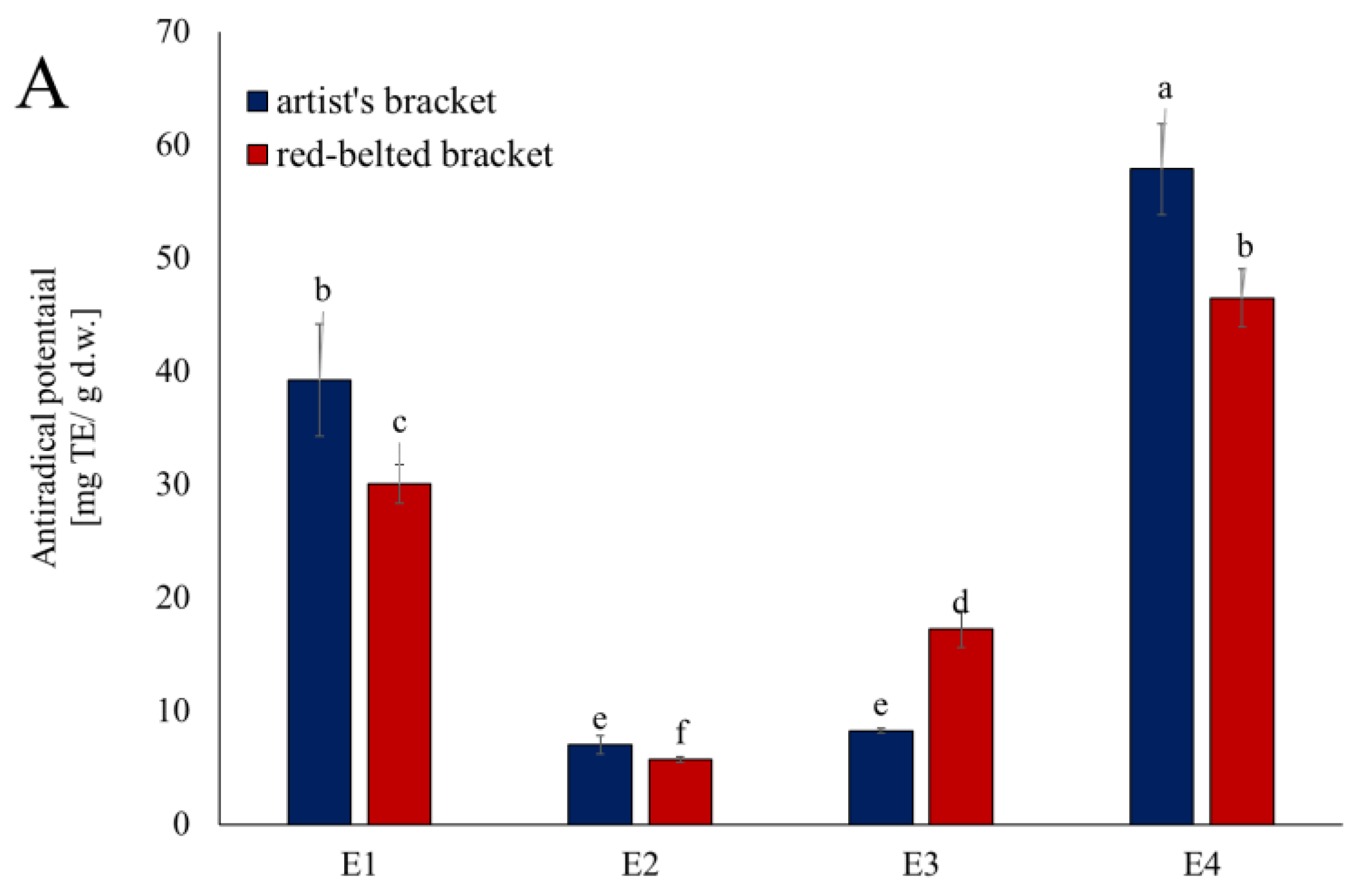
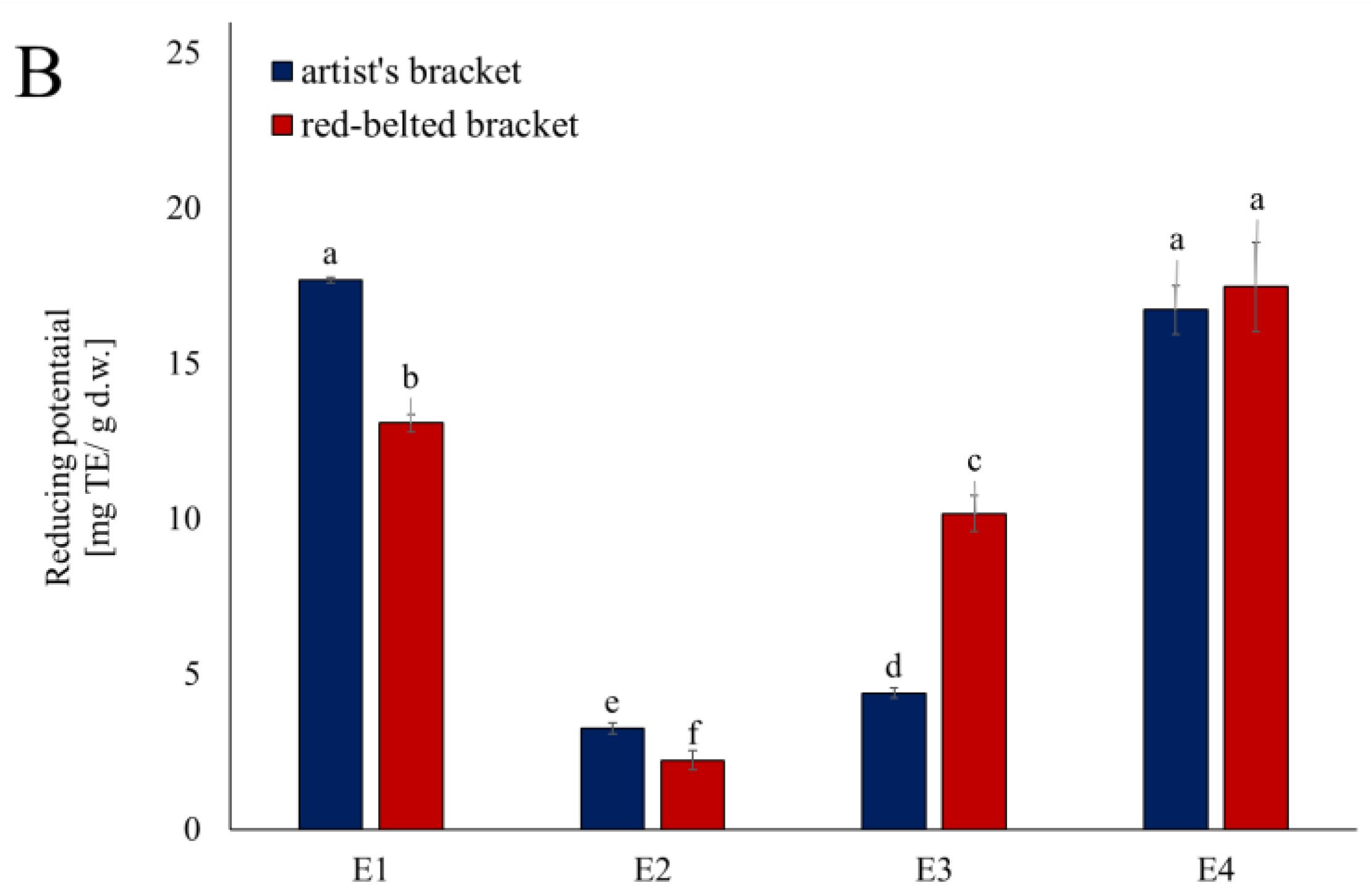
Figure 2.
Inhibition of lipoxygenase activity by fractions from subsequent extraction of mushrooms. Data are mean (n= 9) ± SD. Values with the same letters are not significantly different (p ≤ 0.05). TE- Trolox equivalents; d.w.- dry mass; n.d.- lack of activity. AB- artist’s bracket; RBB- red-belted bracket. E1- 70% ethanol extraction, E2- 50% methanol extraction, E3- hot-water extraction, E4- NaOH extraction.
Figure 2.
Inhibition of lipoxygenase activity by fractions from subsequent extraction of mushrooms. Data are mean (n= 9) ± SD. Values with the same letters are not significantly different (p ≤ 0.05). TE- Trolox equivalents; d.w.- dry mass; n.d.- lack of activity. AB- artist’s bracket; RBB- red-belted bracket. E1- 70% ethanol extraction, E2- 50% methanol extraction, E3- hot-water extraction, E4- NaOH extraction.
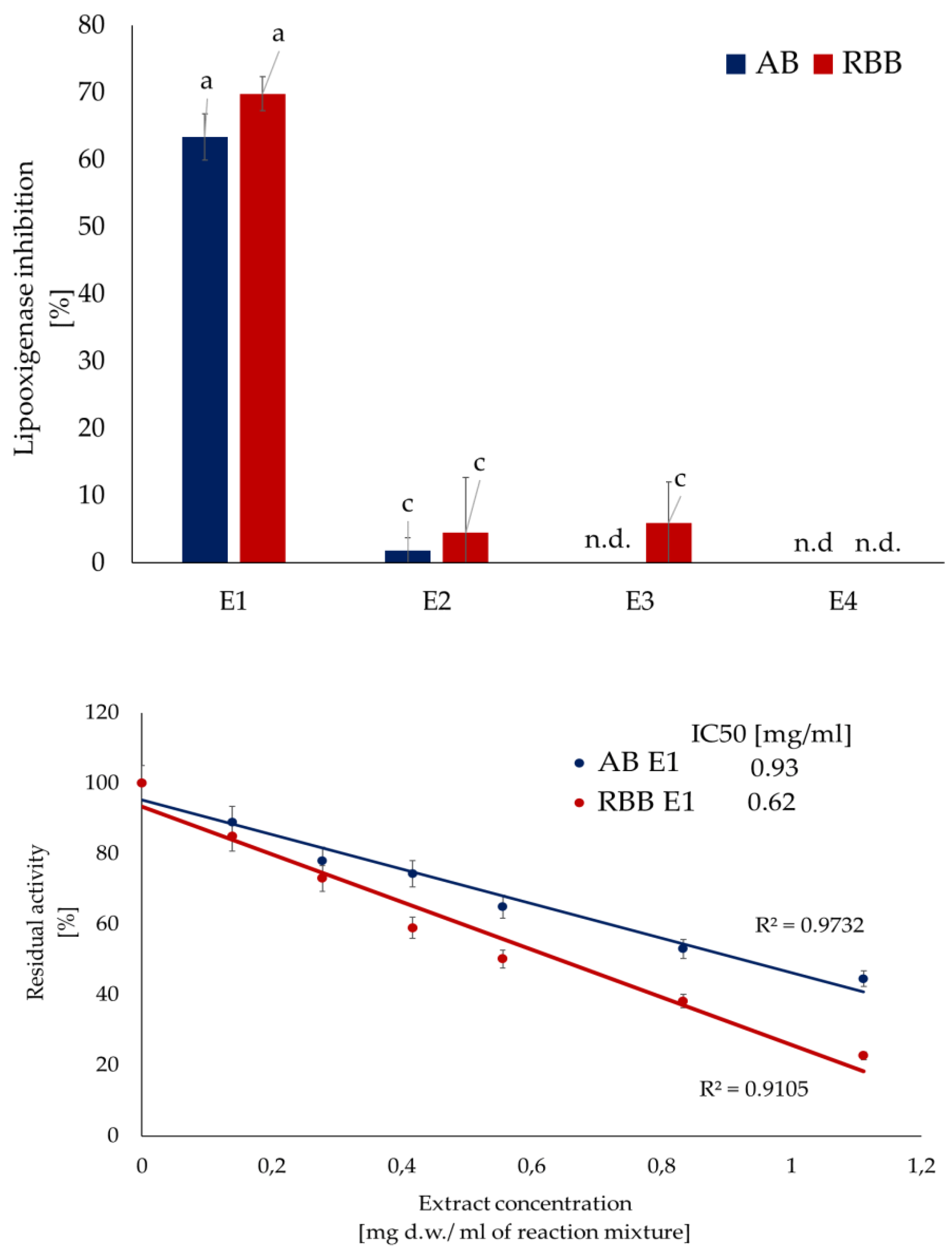
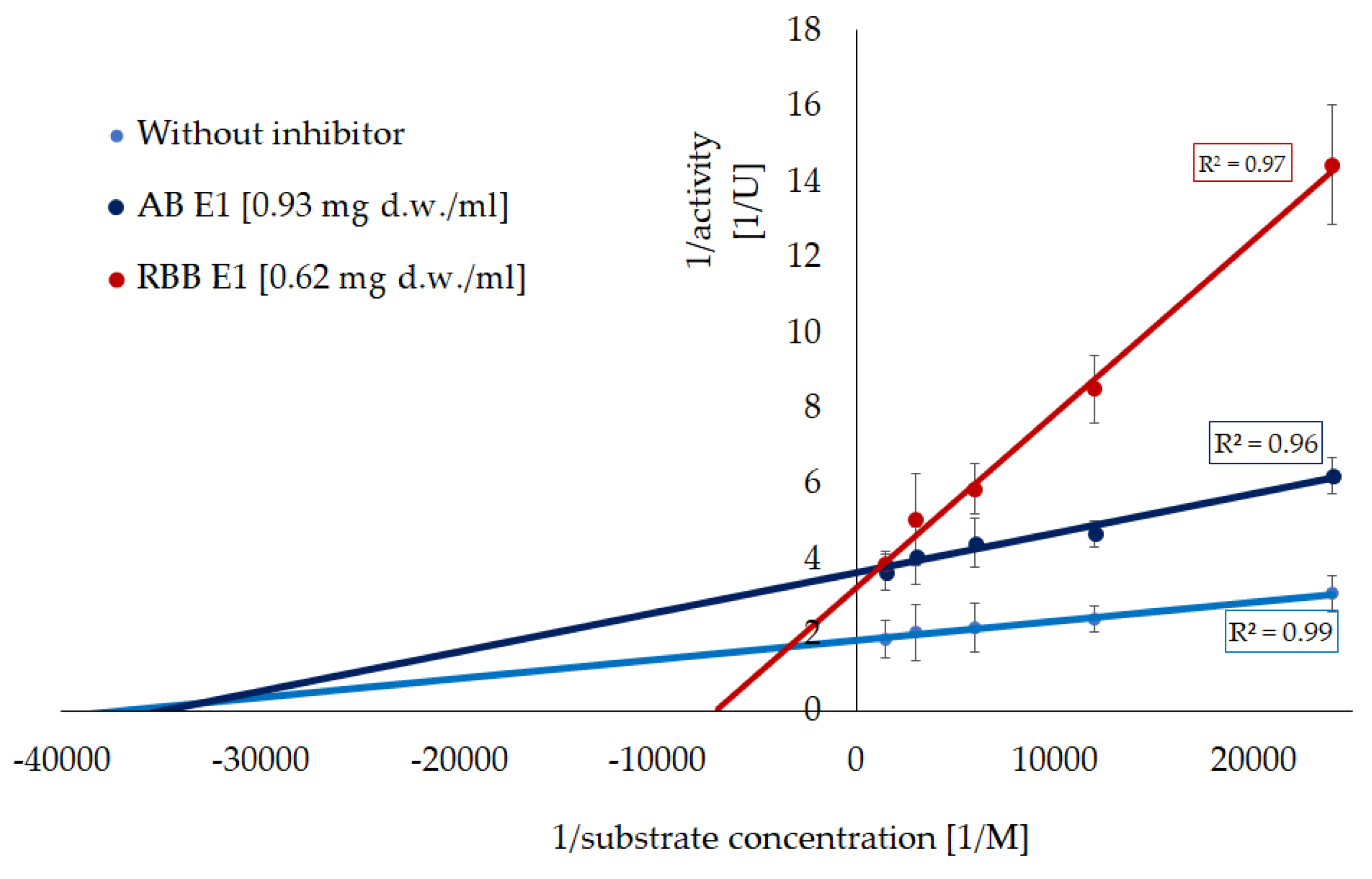
Figure 3.
Inhibition of xanthine oxidase activity by fractions from subsequent extraction of mushrooms. Data are mean (n= 9) ± SD. Values with the same letters are not significantly different (p ≤ 0.05). TE- Trolox equivalents; d.w.- dry mass; n.d.- lack of activity. AB- artist’s bracket; RBB- red-belted bracket. E1- 70% ethanol extraction, E2- 50% methanol extraction, E3- hot-water extraction, E4- NaOH extraction.
Figure 3.
Inhibition of xanthine oxidase activity by fractions from subsequent extraction of mushrooms. Data are mean (n= 9) ± SD. Values with the same letters are not significantly different (p ≤ 0.05). TE- Trolox equivalents; d.w.- dry mass; n.d.- lack of activity. AB- artist’s bracket; RBB- red-belted bracket. E1- 70% ethanol extraction, E2- 50% methanol extraction, E3- hot-water extraction, E4- NaOH extraction.
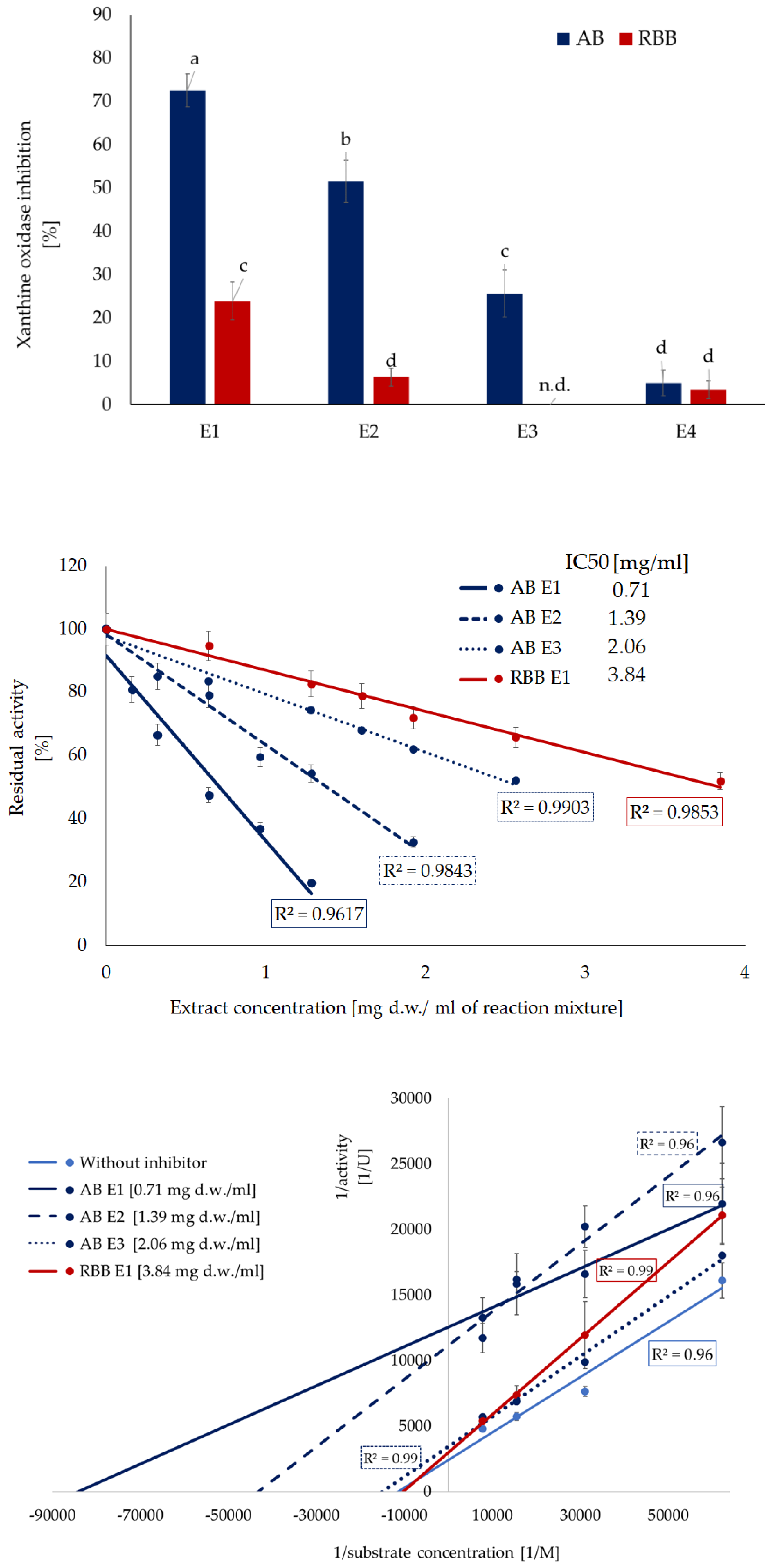

Table 1.
Content of main bioactive components in the fraction from a subsequent extraction of mushrooms.
Table 1.
Content of main bioactive components in the fraction from a subsequent extraction of mushrooms.
| Artist’s bracket | Red-belted bracket | ||
|
FC-reacting substances [mg GAE/ g d.w.] |
E1 | 10.08 ± 0.12 a | 8.11 ± 0.17 b |
| E2 | 1.82 ± 0.04 g | 5.96 ± 0.19 c | |
| E3 | 2.45 ± 0.09 f | 5.07 ± 0.05 d | |
| E4 | 4.28 ± 0.11 e | 10.15 ± 0.11 a | |
| Sum | 18.63 | 29.29 | |
|
Total terpenoids and sterols [mg UAE/ g d.w.] |
E1 | 29.97 ± 1.23 b | 61.51 ± 2.61 a |
| E2 | 2.22 ± 0.22 e | 6.75 ± 0.39 d | |
| E3 | 2.58 ± 0.48 e | 4.98 ± 0.83 d | |
| E4 | 5.09 ± 0.26 d | 14.41 ± 0.34 c | |
| Sum | 39.86 | 87.64 | |
| Mono- and oligosaccharides [mg GluE/ g d.w.] | E1 | 53.28 ± 2.08 b | 50.44 ± 3.03 b |
| E2 | 7.15 ± 1.33 e | 4.75 ± 0.95 e | |
| Sum | 60.42 | 55.24 | |
| Polysaccharides [mg GluE/ g d.w.] |
E3 | 28.73 ± 0.92 c | 19.18 ± 1.86 d |
| E4 | 77.70 ± 0.68 a | 27.18 ± 2.03 c | |
| Sum | 106.4 | 46.4 |
Data are mean (n= 9) ± SD. Values for the features with the same letters are not significantly different (p ≤ 0.05). UAE- ursolic acids equivalents; GAE- gallic acid equivalents; GluE- glucose equivalents; d.w.- dry mass. E1- 70% ethanol extraction, E2- 50% methanol extraction, E3- hot-water extraction, E4- NaOH extraction.
Table 2.
Kinetic parameters of lipoxygenase inhibition by ethanolic extracts (E1) from the mushrooms.
Table 2.
Kinetic parameters of lipoxygenase inhibition by ethanolic extracts (E1) from the mushrooms.
| Without inhibitor | Artist’s bracket | Red-belted bracket | |
| Vmax/Vmaxi [mU] | 536 ± 14.1 a | 274 ± 16.6 b | 272 ± 17.7 b |
| Km/Kmi [μM] | 26.8 ± 0.70 b | 231.7 ± 1.66 a | 27.4 ± 2.94 b |
| Mode of inhibition | - | mixed | noncompetitive |
| kcat [s-1) | 8.40 × 108 a | 4.29 × 108 b | 4.26 × 108 b |
Values with the same letters are not significantly different (p ≤ 0.05). Vmax- maximum reaction velocity, Km- Michaelis constant, kcat- turnover number (catalytic constant).
Table 3.
Kinetic parameters of xanthine oxidase inhibition by extracts from the mushrooms.
| Without inhibitor | Artist’s bracket | Red-belted bracket | |||
| E1 | E2 | E3 | E1 | ||
| Vmax/Vmaxi [U] | 0.415 ± 0.023 a | 0.080 ± 0.010 d | 0.090 ± 0.010 d | 0.292 ± 0.017 c | 0.335 ± 0.026 b |
| Km/Kmi [μM] | 87.7 ± 4.8 | 11.9 ± 1.5 | 23.1 ± 0.0 | 67.0 ± 0.0 | 97.2 ± 7.6 |
| Mode of inhibition | - | acompetitive | acompetitive | acompetitive | noncompetitive |
| kcat [s-1] | 0.072 | 0.014 | 0.016 | 0.051 | 0.058 |
Values with the same letters are not significantly different (p ≤ 0.05). E1- 80% ethanol extraction, E2- 50% methanol extraction, E3- hot-water extraction. Vmax- maximum reaction velocity, Km- Michaelis constant, kcat- turnover number (catalytic constant), kcat/Km- catalytic efficiency.
Table 4.
Individual compounds identified in the ethanolic (E1) extract of the artist’s bracket and red-belted bracket.
Table 4.
Individual compounds identified in the ethanolic (E1) extract of the artist’s bracket and red-belted bracket.
| Red-belted bracket | ||||||
| Compound | Rt | λmax | [M-H] m/z | μg/ g d.w. | ||
| min | nm | MS | MS/MS | |||
| Phenolics | ||||||
| 1. | Vanillic acid | 1.67 | 274 | 167- | 123 | 5.02 ± 0.054 |
| 2. | Protocatechuic acid | 1.76 | 259, 300 | 153- | 109 | 2.26 ± 0.068 |
| 3. | Sinapic acid | 1.92 | 324 | 223- | 164 | 1.81 ± 0.042 |
| 4. | Quercetin 3-O-rutinoside | 2.45 | 255, 355 | 609- | 301 | 1.40 ± 0.013 |
| 5. | Rosmarinic acid glucoside | 2.70 | 324 | 521- | 359 | 1.23 ± 0.013 |
| 6 | Ferulic acid | 2.87 | 327 | 193- | 134 | 3.39 ± 0.074 |
| 7. | Chebulic acid | 2.93 | 320, 402 | 355- | 337, 261 | 5.92 ± 0.104 |
| Terpenes | ||||||
| 8. | 6-α-hydroxy-3,16-dioxolanosta-7(8),9(11), ,24-trien-21-oic acid |
6.43 | - | 481- | 463, 437, 403, 388, 373 | 3.43 ± 0.040 |
| 9. | Dehydrotumulosic acid | 6.55 | - | 483- | 465, 421, 255 | 1.36 ± 0.045 |
| 10. | 16-α-hydroxy-3, oxolanosta-7,9(11),24- -trien-21-oic acid |
6.60 | - | 467- | 423, 407, 389, 373, 311 | 2.20 ± 0.004 |
| 11. | Irpeksolactin E | 6.64 | - | 485- | 467, 423, 407, 353, 337 | 2.90 ± 0.012 |
| 12. | Forpinic acid D | 6.85 | - | 479- | 465, 435, 441 | 1.98 ± 0.014 |
| 13. | Forpinic acid E | 6.98 | - | 495- | 465, 421, 405, 391, 373 | 1.15 ± 0.014 |
| 14. | 16-α-hydroxy-dehydrotraumetenolic acid | 7.06 | - | 469- | 467, 425, 409, 391, 337 | 2.71 ± 0.017 |
| 15. | Unspecified | 7.38 | - | 467- | - | 5.27 ± 0.032 |
| 16. | Formipiniate B | 7.43 | - | 525- | 497, 483, 465, 441 | 2.48 ± 0.037 |
| 17. | Forpinic acid A | 8.52 | - | 599- | 537, 455 | 2.44 ± 0.057 |
| 18. | 20-OH-lucidenic acid A | 8.60 | - | 455- | 149 | 1.56 ± 0.039 |
| 19. | Forpinic acid F | 8.68 | - | 639- | 537, 451, 371, 339 | 1.90 ± 0.007 |
| 20. | Forpinic acid G | 8.74 | - | 583- | 465, 449, 434, 389 | 20.9 ± 0.176 |
| 21. | Forpinic acid C | 8.79 | - | 541- | 451, 371, 339 | 12.0 ± 0.377 |
| 22. | Piptolinic acid D | 8.87 | - | 465- | 435, 421, 405, 369 | 1.97 ± 0.025 |
| 23. | Formipinic acid H | 9.00 | - | 627- | 537, 465, 373 | 3.52 ± 0.038 |
| 24. | Unspecified | 9.10 | - | 495- | - | 2.27 ± 0.068 |
| 25. | Formitopsic acid F | 9.22 | - | 527- | 479, 435, 419, 351 | 2.69 ± 0.062 |
| Artist’s bracket | ||||||
| Terpenes | ||||||
| 1. | Ganoderenic acid A | 4.38 | - | 513- | 495, 451, 249 | 7.78 ± 0.085 |
| 2. | Ganoderenic acid D | 4.75 | - | 511- | 493, 285, 149 | 2.10 ± 0.063 |
| 3. | Ganoderic acid C6 | 4.83 | - | 529- | 481, 467, 437 | 3.57 ± 0.082 |
| 4. | Ganoderic acid A | 4.93 | - | 515- | 497, 303, 2449 | 3.31 ± 0.031 |
| 5. | Ganoderic acid H | 6.93 | - | 571- | 511, 467, 437 | 3.15 ± 0.033 |
| 6. | Ganoderic acid F | 7.16 | - | 569- | 509, 465, 435 | 8.33 ± 0.183 |
Data are mean (n= 9) ± SD. d.w.- dry mass.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



