
Submitted:
13 December 2025
Posted:
16 December 2025
You are already at the latest version
Abstract
In this work, we analyzed the ability to incorporate 8-oxo-dATP by several human DNA polymerases: replicative Pol ε (exo-), BER enzymes Pol β and Pol λ, and trans-lesion Pol η, Pol ι, and Pol κ. We demonstrated that human DNA polymerases differ in their abilities to discriminate against 8-oxo-dATP. Among tested DNA polymerases, Pol λ demonstrated the worst ability to discriminate against 8-oxo-dATP opposite template T on both singles-stranded DNA and double-stranded DNA substrates with a 1 nt gap. In contrast, Pol β was quite accurate on singe-stranded DNA substrate but incorporated 8-oxo-dATP opposite template T in a 1 nt gap. Unexpectedly, the catalytic subunit of high-fidelity Pol ε (exo-) incorporated 8-oxodATP opposite templates T and G with weak but higher efficiency compare to error-prone polymerases of Family Y. While structures of human polymerases with incoming 8oxo-dATP are not available, we speculate possible mechanism of 8-oxo-dATP discriminations.
Keywords:
8-oxo-dATP
; DNA polymerase
; mutagenesis
; ROS
1. Introduction
Guanine is the most susceptible DNA base to oxidation due to its low redox potential. It readily forms 7,8-dihydro-8-oxoguanine (8-oxoG) lesion when attacked by reactive oxygen species (ROS) in DNA [1,2]. Moreover, guanine is susceptible to oxidation in the nucleotide pool and its mutagenic potential is realized also through the incorporation of 2ʹ-deoxy-7,8-dihydro-8-oxo-guanosine-5ʹ-triphosphate (8-oxo-dGTP) into DNA [3,4]. The active site of some DNA polymerases can be less demanding to the structure of DNA and/or incoming nucleotides enabling incorporation of modified nucleotides directly from the nucleotide pool. To enhance the selection of complementary canonical nucleotides during replication and DNA repair, enzyme Human MutT Homolog 1 (MTH1) recognizes and hydrolyzes 8-oxo-dGTP to the corresponding monophosphate, preventing the promutagenic events [5].
7,8-dihydro-8-oxoadenine (8-oxoA) is another abundant oxidative lesion with dual miscoding properties. The number of 8-oxoA lesions ranged from 10 to 50% of 8-oxoG in DNA [6], but in some cells it was detected at the level of 0.7 lesions per 106 nucleotides, which corresponds to ~2200 lesions per human genome and is comparable with 8-oxoG levels in mammalian cells [7]. In some cancer cells the ratio of 8-oxoA to 8-oxoG reaches 1:1 [8].
Oxidation of adenine, like guanine, can also occurs at comparable levels in the nucleotide pool, for example after treatment of HepG2 and LO2 cells with acetamiprid [9]. MTH1 exhibits catalytic activity towards 2’-deoxy-7,8-dihydro-8-oxoadenosine-5’-triphosphate (8-oxo-dATP) with efficiency comparable to 8-oxo-dGTP [10]. However, the incorporation of 8-oxo-dATP into DNA has not been studied. Only a few DNA polymerases have been biochemically tested with 8-oxo-dATP. In this work, we carry out systemic analysis of incorporation of 8-oxo-dATP into DNA by six human DNA polymerases belonging to several families. Our data demonstrate that human DNA polymerases differ in their abilities to discriminate against 8-oxo-dATP.
2. Materials and Methods
2.1. Equipment and Reagents for Chemical Synthesis
All reagents were purchased from Sigma-Aldrich (USA). Solvents were purchased from CHIMMED (Russia). 1H and 31P NMR spectra were recorded on a Bruker Avance III 600 spectrometer (Germany) at 600 and 243 MHz, respectively. The multiplicity of signals in the spectra is shown using the following abbreviations: s (singlet), d (doublet), t (triplet), quartet (q) and m (multiplet). The spin–spin coupling constants (J) are given in Hz. Ion-exchange chromatography was performed on an Akta Explorer 100 instrument (Cytiva, Sweden). ESI HR mass spectrum was acquired on a LTQ FT Ultra (Germany) mass spectrometer in a positive ion mode.
2.2. 8-oxo-dATP Synthesis
8-oxo-dATP substrate was prepared from 2’-deoxy-7,8-dihydro-8-oxo-adenosine (8-oxo-dA) as was previously described for the synthesis of 8-oxo-εATP [11] with minor modification (Figure 1). Tributylamine (Bu3N, 1.43 mL) and freshly distilled trimethyl phosphate ((CH3O)3PO, 12.0 mL) were added to 8-oxo-dA [12] (0.80 g, 3.00 mmol) in a Schlenk flask (100 mL) under inert gas atmosphere. The mixture was vigorously stirred at room temperature for 30 min and then cooled to −10 °C. Under inert gas atmosphere, to the reaction mixture phosphorus oxychloride (POCl3, 0.50 mL, 5.40 mmol) was added and the mixture was stirred at −10 °C for 1 h. Then, to the reaction mixture a phosphorylating agent prepared by vigorously stirring acetonitrile CH3CN (30 ml), Bu3N (4.2 ml, 17.7 mmol) and bis(tributylammonium)pyrophosphate ((NHBu3)2H2P2O7, 1.8 g, 3.30 mmol) under inert atmosphere for 20 min at −20 °C was added dropwise. After stirring the mixture at −10 °C for 1 h, cold water (65 ml) was added. The mixture was stirred at 0 °C for 1 h, then transferred to a separatory funnel, and washed with methylene chloride (25 ml, 5 times). The aqueous layer was collected and aqueous ammonia solution was added to adjust the pH to 7.0. The resulting 8-oxo-dATP solution was stored in a refrigerator until purification by ion-exchange chromatography on a 50×250 mm column packed with HEMA-BIO 1000 DEAE 70 μm sorbent (Germany) with a gradient of 50-600 mM triethylammonium bicarbonate (pH 7.6). Fractions containing the target product were evaporated. The residue was re-dissolved in water and evaporated to remove residual buffer to afford the bis-triethylammonium salt of 8-oxo-dATP (0.86 g, 1.21 mmol, 40%) as a white flaky powder. 1H NMR (600 MHz, D2O): δ 8.21 (s, 1H), 6.37 (d, J = 7.1 Hz, 1H), 4.83 – 4.78 (m, 1H), 4.32 – 4.27 (m, 1H), 4.26 – 4.22 (m, 1H), 4.20 – 4.15 (m, 1H), 3.34 – 3.29 (m, 1H), 3.25 (q, J = 7.3 Hz, 12H), 2.41 – 2.36 (m, 1H), 1.33 (t, J = 7.3 Hz, 18H). 31P NMR (243 MHz, D2O): δ -10.9 (d, J = 19.8 Hz, 1P), -11.2 (d, J = 20.0 Hz, 1P), -23.3 (dd, J = 19.8 Hz, J = 20.0 Hz, 1P). HRMS (ESI) m/z: calculated for C10H17N5O13P3+ [M - 2(C2H5)3N + H]+: 508.0030; found 508.0037. NMR spectra are shown in Supplementary.
2.3. DNA Templates and Enzymes
Pol η, Pol ι, Pol β, Pol λ were purified from S. cerevisiae and E. coli as described in [13,14,15]. Human Pol κ was kindly provided by L.V. Gening (Institute of Molecular Genetics, Russia). Plasmid encoding the POLE gene of the catalytic subunit of human Pol ε lacking the 3`-5`-exonuclease activity was kindly provided by Prof. T. Tahirov (UNMC, Omaha). POLE was fused with the N-terminal 6хHIS-SUMO tag. Protein expression was carried out in C3013 E. coli strain at 15°С for 12 h following induction with 0.15 µg/ml tetracycline. After metal-affinity chromatography with Ni2+-NTA agarose SUMO-tag was removed by Ulp protease and enzyme was purified on heparin-sepharose column.
DNA oligonucleotides used in this study (Table 1) were synthetized as described previously [16]. To prepare DNA substrates, the 5′-Cy5-labelled primer Cy5-Pr16 was annealed to the corresponding unlabeled template oligonucleotides at a molar ratio of 1:1.1 in 100 mM NaCl by heating to 97 °C and slowly cooling to 4 °C.
2.4. DNA Polymerase Reactions for the Primer Extension Assay
Standard primer extension reactions were performed in 20 µl containing 100 nM DNA substrate, 50 μM dNTP (including 8-oxo-dATP), 30 mM HEPES рН 7.4, 10 mM MgCl2 (for Pol ε, Pol κ, Pol η, Pol β) or 1 mM MgCl2 (for Pol ι and Pol λ), 100 µg/ml BSA, 1 mM DTT, 8% glycerol and 2 – 60 nM of polymerase as indicated on the figure legends. Reactions were incubated at 37 °C for 10 min or as indicated on figure legends and placed on ice. Reactions were stopped by the addition of an equal volume of 2х loading buffer (20 mM EDTA, 0.001% bromophenol blue, 96% formamide) and heated for 5 min at 95°С. The reaction products were resolved on 21% polyacrylamide gels with 8 M urea, visualized on Typhoon 9400 (GE Healthcare, USA) and analyzed with ImageQuant software. All experiments were repeated three times. The percent of the extended primer (PrExt) was calculated for each reaction and the mean values of PrExt with the standard errors are shown in figures.
3. Results
In this work, we analyzed the ability of a number of human DNA polymerases involved in DNA translesion synthesis, DNA repair and replication to incorporate 8-oxo-dATP into DNA. We tested the incorporation of dATP and 8-oxo-dATP in primer extension reactions opposite G and T at the +1 position of template DNA with a protruding single-stranded 5’-end. Additionally, the activity of BER enzymes Pol β and Pol λ from family X was analyzed on DNA templates containing a 1-nt gap.
3.1. Negligible Incorporation of 8-oxo-dATP by Translesion DNA Polymerases of Family Y
Human DNA polymerases Pol η, Pol κ and Pol ι play a key role as an inserter polymerase during DNA translesion synthesis. Their ability to incorporate 8-oxo-dATP have been analyzed for the first time. The activities of Pol κ and Pol η were comparable: both enzymes incorporated the control dATP opposite template T with high efficiency, but successfully discriminated against the incorporation of 8-oxo-dATP regardless of the template nucleotide (Figure 2). Pol κ incorporated 8-oxodATP opposite template T slightly more efficient than Pol η. Pol ι demonstrated extremely low activity opposite template T. It is known that Pol ι prefers to incorporate dGTP over dATP and abrogate DNA synthesis opposite template T; this phenomenon have been called the “T-stop” rule. Minor incorporation of 8-oxo-dATP by Pol ι was detected opposite template G (Figure 2).
3.2. Incorporation of 8-oxo-dATP by Pol β and Pol λ of Family X
Pol β and Pol λ are major DNA polymerases involved in DNA synthesis during BER. Their activity was tested on DNA with a protruding single-stranded 5’-end and double-stranded DNA substrates containing a 1 nt gap. On a DNA substrate with a protruding single-stranded 5’-end, Pol β possessed highly discriminatory behavior: Pol β efficiently incorporated control dATP opposite template T, but in other cases enzyme successfully discriminated against all non-cognate incoming nucleoside triphosphates (Figure 3A). Pol λ, in turn, was capable of incorporating both intact dATP and modified 8-oxo-dATP opposite template T with comparable efficiency (Figure 3A). At the same time, Pol λ incorporated nucleoside triphosphates opposite template G with very low efficiency (Figure 3A). On DNA substrate with a 1 nt gap both enzymes efficiently incorporated 8-oxo-dATP opposite template T (Figure 3B) demonstrating low ability to discriminate an oxidized form of dATP.
2.3. Incorporation of 8-oxo-dATP by Replicative B-family Pol ε
High-fidelity human Pol ε replicates the majority of the leading strand of genomic DNA [17]. Surprisingly, the Pol ε catalytic subunit variant lacking the 3`-5`-exonuclease activity possessed relatively low ability to discriminate against 8-oxo-dATP when incorporating both opposite templates T and G (Figure 4A). However, this synthesis was highly distributive (Figure 4B). Unlike incorporation of dATP opposite template T, Pol ε (exo-) was not able to continue replication beyond the non-canonical nucleotide substrate even under prolonged incubation time.
4. Discussion
The concentrations of 8-oxo-dATP under physiological conditions and under oxidative stress can reach levels comparable with 8-oxo-dGTP [9]. However, the mutagenic effect of 8-oxo-dATP incorporation in DNA in mammalian cells have not been estimated yet. A single study indicates that 8-oxo-dATP can be fairly well discriminated by DNA polymerases in vitro. Among a few tested enzymes, human Pol β demonstrated the most efficient incorporation of 8-oxo-dATP, especially opposite the complementary template T. Low incorporation efficiency was observed in reactions with E. coli KF and mammalian Pol α, which incorporated the oxidized nucleotide substrate preferentially opposite non-complementary G and A, while human Pol λ did not incorporate 8-oxo-dATP onto any of the DNA substrates [18].
Our data are in agreement with quite efficient incorporation of 8-oxo-dATP opposite template T by Pol β. However, we demonstrated that incorporation of non-canonical nucleotide substrate is more efficient on double-stranded DNA with a 1 nt gap than on single-stranded DNA substrate. In contrast, our data indicated very efficient incorporation of 8-oxo-dATP on both DNA substrates by human Pol λ. Pol ε (exo-), Pol κ, Pol η and Pol ι were tested with 8-oxo-dATP in this work for the first time. Very limited incorporation of 8-oxo-dATP was observed by translesion error-prone DNA polymerases of Family Y but small amounts of 8-oxo-dATP were incorporated by the catalytic subunit of high-fidelity Pol ε (exo-).
Ability to discriminate against 8-oxo-dATP during DNA synthesis depends on the unique architectures of polymerases` active sites. However, human DNA polymerase structures with incoming 8-oxo-dATP are not available yet. We speculate that 8-oxodATP incorporation opposite template T should occur in the anti conformation and may cause the steric hindrance between the 8-oxo group and the triphosphate group or Me2+ ions in Pol β [19,20], Pol η [21], Pol ι [22] and Pol kappa [23] resulting in shift of the triphosphate of incoming nucleotide and phosphate backbone of templating nucleotide and less efficient pairing with T (Figure S3). In Pol ε, the 8-oxo group may cause the steric hindrance with deoxyribose ring possibly changing ribose conformation [24] (Figure S3). Therefore, efficiency of 8-oxo-dATP incorporation by a polymerase is likely depends on the ability of its active site to rotate base to accommodate the modification. Remarkably, Pol λ very efficiently incorporated 8-oxo-dATP opposite template T. Unlike Pol β which contains negatively charged Asp276 close to the incoming dATP opposite a templating T, Pol λ active site at similar position contains neutral Ala510 which will not interfere with the 8-oxo group [25] (Figure S3). Such efficient 8-oxo group accommodation will not interfere with 8-oxodATP discrimination.
Pol ι, Pol λ and Pol ε (exo-) also demonstrated minor incorporation of 8-oxo-dATP opposite template G. Such activity likely requires the rotation of oxidized adenine to the syn conformation and formation of Hoogsteen hydrogen bonds with the templating G. Similar accommodation of oxidized dGTP was demonstrated for incoming 8-oxo-dGTP opposite template A in the active sites of Pol λ and Pol β [19,25]. Moreover, residues Asp276, Lys280 in Pol β and Tyr112, Tyr138 in Pol κ might also clash with shifting oxidized base contributing to 8-oxo-dATP discrimination [19,23].
5. Conclusions
Our data show that human DNA polymerases significantly differ in their ability to discriminate against 8-oxodATP during DNA synthesis. While some enzymes such as Pol β, Pol λ and Pol ε possibly might contribute to the 8-oxodATP-induced mutagenesis, the exact deleterious effect of dATP oxidation in human cells is still unclear and is a subject of future studies.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org.
Author Contributions
Methodology A.V.M, and A.V.A.; investigation, A.A.K., P.N.K., M.S.B., Y.G.B., E.O.B.; resources A.V.M. and A.V.A.; data curation A.A.K. and P.N.K.; writing—original draft preparation, A.V.M.; writing—review and editing, A.V.A.; visualization, A.V.M.; supervision, A.V.M. and A.V.A.; funding acquisition, A.V.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Russian Scientific Foundation, grant number 23-14-00209.
Data Availability Statement
All data are available upon request.
Acknowledgments
We thank Tair Tahirov for providing POLE plasmid, Andrey Baranovskiy for useful discussion and Timofei Zatsepin for conducting oligonucleotide synthesis.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Helbock, H.J.; Beckman, K.B.; Shigenaga, M.K.; Walter, P.B.; Woodall, A.A.; Yeo, H.C.; Ames, B.N. DNA Oxidation Matters: The HPLC–Electrochemical Detection Assay of 8-Oxo-Deoxyguanosine and 8-Oxo-Guanine. Proc. Natl. Acad. Sci. U.S.A. 1998, 95, 288–293. [Google Scholar] [CrossRef]
- Møller, P.; Cooke, M.S.; Collins, A.; Olinski, R.; Rozalski, R.; Loft, S. Harmonising Measurements of 8-Oxo-7,8-Dihydro-2′-Deoxyguanosine in Cellular DNA and Urine. Free Radical Research 2012, 46, 541–553. [Google Scholar] [CrossRef]
- Hayakawa, H.; Taketomi, A.; Sakumi, K.; Kuwano, M.; Sekiguchi, M. Generation and Elimination of 8-Oxo-7,8-Dihydro-2’-Deoxyguanosine 5’-Triphosphate, a Mutagenic Substrate for DNA Synthesis, in Human Cells. Biochemistry 1995, 34, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, Y.I.; Minnick, D.T.; Izuta, S.; Kunkel, T.A. DNA Replication Fidelity with 8-Oxodeoxyguanosine Triphosphate. Biochemistry 1994, 33, 4695–4701. [Google Scholar] [CrossRef] [PubMed]
- Nakabeppu, Y.; Ohta, E.; Abolhassani, N. MTH1 as a Nucleotide Pool Sanitizing Enzyme: Friend or Foe? Free Radical Biology and Medicine 2017, 107, 151–158. [Google Scholar] [CrossRef]
- Fuciarelli, A.F.; Wegher, B.J.; Gajewski, E.; Dizdaroglu, M.; Blakely, W.F. Quantitative Measurement of Radiation-Induced Base Products in DNA Using Gas Chromatography-Mass Spectrometry. Radiation Research 1989, 119, 219. [Google Scholar] [CrossRef]
- Tuo, J.; Jaruga, P.; Rodriguez, H.; Dizdaroglu, M.; Bohr, V.A. The Cockayne Syndrome Group B Gene Product Is Involved in Cellular Repair of 8-Hydroxyadenine in DNA. Journal of Biological Chemistry 2002, 277, 30832–30837. [Google Scholar] [CrossRef] [PubMed]
- Jałoszyński, P.; Jaruga, P.; Oliński, R.; Biczysko, W.; Szyfter, W.; Nagy, E.; Möller, L.; Szyfter, K. Oxidative DNA Base Modifications and Polycyclic Aromatic Hydrocarbon DNA Adducts in Squamous Cell Carcinoma of Larynx. Free Radical Research 2003, 37, 231–240. [Google Scholar] [CrossRef]
- Zhang, H.-X.; Yu, D.; Sun, J.-F.; Zeng, L.; Wang, C.-Y.; Bai, L.-P.; Zhu, G.-Y.; Jiang, Z.-H.; Zhang, W. An Integrated Approach to Evaluate Acetamiprid-Induced Oxidative Damage to tRNA in Human Cells Based on Oxidized Nucleotide and tRNA Profiling. Environment International 2023, 178, 108038. [Google Scholar] [CrossRef]
- Fujikawa, K.; Kamiya, H.; Yakushiji, H.; Fujii, Y.; Nakabeppu, Y.; Kasai, H. The Oxidized Forms of dATP Are Substrates for the Human MutT Homologue, the hMTH1 Protein. Journal of Biological Chemistry 1999, 274, 18201–18205. [Google Scholar] [CrossRef]
- Petushkov, I.V.; Aralov, A.V.; Ivanov, I.A.; Baranov, M.S.; Zatsepin, T.S.; Kulbachinskiy, A.V. Effect of 8-Oxo-1,N6-Ethenoadenine Derivatives on the Activity of RNA Polymerases from SARS-CoV-2 and Escherichia Coli. Biochemistry Moscow 2024, 89, 2263–2273. [Google Scholar] [CrossRef] [PubMed]
- Chatgilialoglu, C.; Navacchia, M.L.; Postigo, A. A Facile One-Pot Synthesis of 8-Oxo-7,8-Dihydro-(2′-Deoxy)Adenosine in Water. Tetrahedron Letters 2006, 47, 711–714. [Google Scholar] [CrossRef]
- Kazachenko, K.Y.; Miropolskaya, N.A.; Gening, L.V.; Tarantul, V.Z.; Makarova, A.V. Alternative Splicing at Exon 2 Results in the Loss of the Catalytic Activity of Mouse DNA Polymerase Iota in Vitro. DNA Repair 2017, 50, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Shilkin, E.S.; Petrova, D.V.; Kruchinin, A.A.; Zharkov, D.O.; Makarova, A.V. The Effect of Methylation and Hydroxymethylation of Cytosine on Activity and Fidelity of Pol λ and Pol β. DNA Repair 2025, 148, 103815. [Google Scholar] [CrossRef] [PubMed]
- Boldinova, E.O.; Yudkina, A.V.; Shilkin, E.S.; Gagarinskaya, D.I.; Baranovskiy, A.G.; Tahirov, T.H.; Zharkov, D.O.; Makarova, A.V. Translesion Activity of PrimPol on DNA with Cisplatin and DNA–Protein Cross-Links. Sci Rep 2021, 11, 17588. [Google Scholar] [CrossRef]
- Boldinova, E.O.; Kruchinin, A.A.; Kamzeeva, P.N.; Aralov, A.V.; Makarova, A.V. Accurate DNA Synthesis Across 8-Oxoadenine by Human PrimPol. IJMS 2025, 26, 6796. [Google Scholar] [CrossRef]
- Bedaiwi, S.; Usmani, A.; Carty, M.P. Canonical and Non-Canonical Roles of Human DNA Polymerase η. Genes 2024, 15, 1271. [Google Scholar] [CrossRef]
- Grin, I.R.; Vasilyeva, S.V.; Dovgerd, A.P.; Silnikov, V.N.; Zharkov, D.O. Human and Bacterial DNA Polymerases Discriminate against 8-Oxo-2’-Deoxyadenosine- 5’-Triphosphate. Biopolym. Cell 2012, 28, 306–309. [Google Scholar] [CrossRef]
- Batra, V.K.; Beard, W.A.; Shock, D.D.; Pedersen, L.C.; Wilson, S.H. Structures of DNA Polymerase β with Active-Site Mismatches Suggest a Transient Abasic Site Intermediate during Misincorporation. Molecular Cell 2008, 30, 315–324. [Google Scholar] [CrossRef]
- Freudenthal, B.D.; Beard, W.A.; Perera, L.; Shock, D.D.; Kim, T.; Schlick, T.; Wilson, S.H. Uncovering the Polymerase-Induced Cytotoxicity of an Oxidized Nucleotide. Nature 2015, 517, 635–639. [Google Scholar] [CrossRef]
- Zhao, Y.; Gregory, M.T.; Biertümpfel, C.; Hua, Y.-J.; Hanaoka, F.; Yang, W. Mechanism of Somatic Hypermutation at the WA Motif by Human DNA Polymerase η. Proc. Natl. Acad. Sci. U.S.A. 2013, 110, 8146–8151. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Nair, D.T.; Johnson, R.E.; Prakash, L.; Prakash, S.; Aggarwal, A.K. Replication across Template T/U by Human DNA Polymerase-ι. Structure 2009, 17, 974–980. [Google Scholar] [CrossRef]
- Yockey, O.P.; Jha, V.; Ghodke, P.P.; Xu, T.; Xu, W.; Ling, H.; Pradeepkumar, P.I.; Zhao, L. Mechanism of Error-Free DNA Replication Past Lucidin-Derived DNA Damage by Human DNA Polymerase κ. Chem. Res. Toxicol. 2017, 30, 2023–2032. [Google Scholar] [CrossRef]
- Roske, J.J.; Yeeles, J.T.P. Structural Basis for Processive Daughter-Strand Synthesis and Proofreading by the Human Leading-Strand DNA Polymerase Pol ε. Nat Struct Mol Biol 2024, 31, 1921–1931. [Google Scholar] [CrossRef] [PubMed]
- Gosavi, R.A.; Moon, A.F.; Kunkel, T.A.; Pedersen, L.C.; Bebenek, K. The Catalytic Cycle for Ribonucleotide Incorporation by Human DNA Pol λ. Nucleic Acids Research 2012, 40, 7518–7527. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The synthesis of the bis-triethylammonium salt of 8-oxo-dATP.
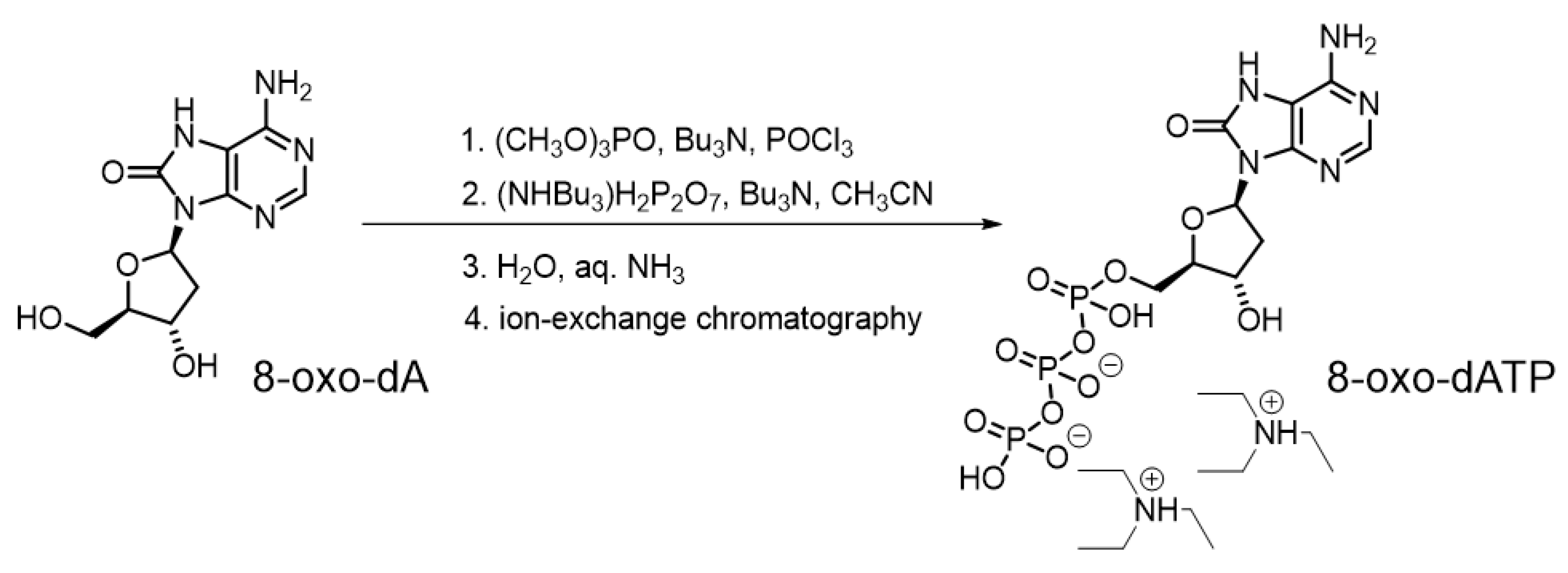
Figure 2.
Incorporation of 8-oxo-dATP opposite templates T and G by human DNA polymerases of Family Y. The mean values of primer extension and standard errors are indicated on the diagrams.
Figure 2.
Incorporation of 8-oxo-dATP opposite templates T and G by human DNA polymerases of Family Y. The mean values of primer extension and standard errors are indicated on the diagrams.
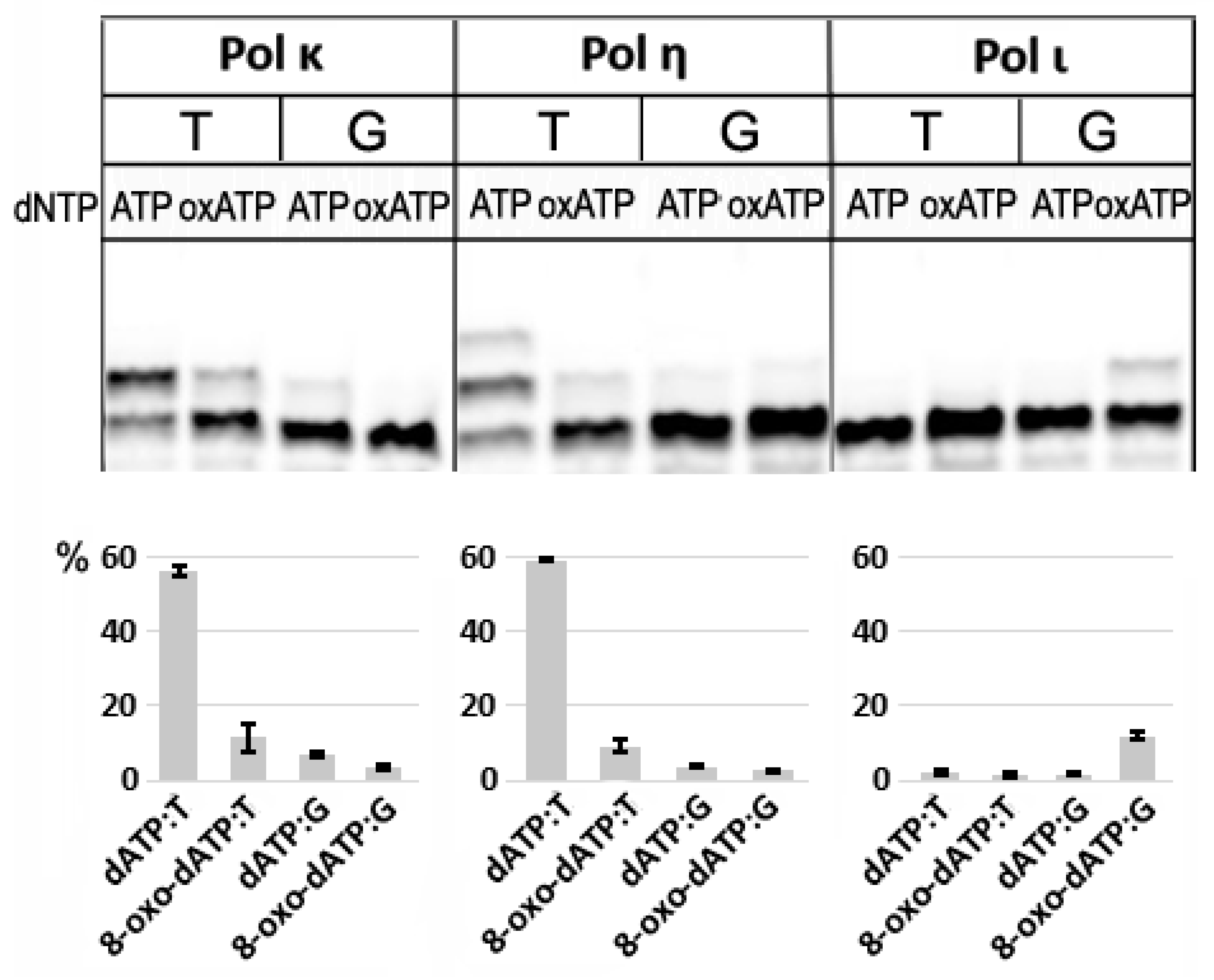
Figure 3.
Incorporation of 8-oxo-dATP opposite templates T and G by human DNA polymerases of Family X. (A) Incorporation of 8-oxo-dATP by Pol β and Pol λ on a DNA substrate with a protruding single-stranded 5’-end. (B) Incorporation of 8-oxo-dATP by Pol β and Pol λ on DNA templates containing a 1-nt gap. The mean values of primer extension and standard errors are indicated on the diagrams.
Figure 3.
Incorporation of 8-oxo-dATP opposite templates T and G by human DNA polymerases of Family X. (A) Incorporation of 8-oxo-dATP by Pol β and Pol λ on a DNA substrate with a protruding single-stranded 5’-end. (B) Incorporation of 8-oxo-dATP by Pol β and Pol λ on DNA templates containing a 1-nt gap. The mean values of primer extension and standard errors are indicated on the diagrams.
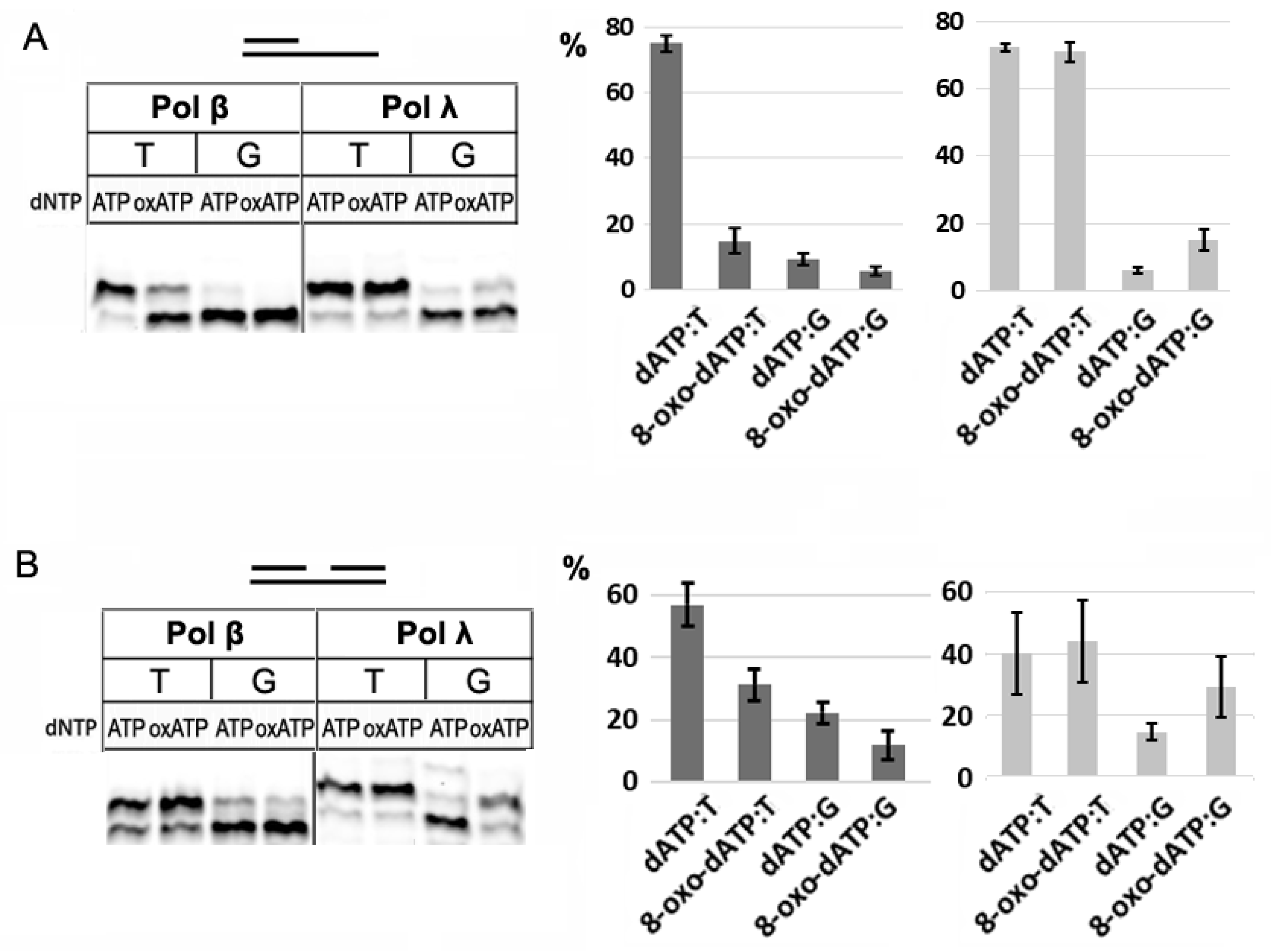
Figure 4.
Incorporation of 8-oxo-dATP by human catalytic subunit Pol ε (exo-). (A) Incorporation of 8-oxo-dATP opposite templates T and G. The mean values of primer extension and standard errors are indicated on the diagram. (B) The distributive character of 8-oxo-dATP opposite template T by Pol ε (exo-). Reactions were incubated 10 min with 15 nM Pol ε for dATP and 20 min with 20 nM Pol ε for 8-oxo-dATP.
Figure 4.
Incorporation of 8-oxo-dATP by human catalytic subunit Pol ε (exo-). (A) Incorporation of 8-oxo-dATP opposite templates T and G. The mean values of primer extension and standard errors are indicated on the diagram. (B) The distributive character of 8-oxo-dATP opposite template T by Pol ε (exo-). Reactions were incubated 10 min with 15 nM Pol ε for dATP and 20 min with 20 nM Pol ε for 8-oxo-dATP.

Table 1.
Oligonucleotides used in this study.
| Oligonucleotide | Sequence 5′-3′ |
| Сy5-Pr16 | Cy5-GTCACAGAGATACTAC |
| TemplateTA | GAGCAGTCGCACATGTAGTATCTCTGTGAC |
| TemplateGA | GAGCAGTCGCACAGGTAGTATCTCTGTGAC |
| Closing_oxodATP | /P/-TGTGCGACTGCTC |
P = 5`-phosphate.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



