Submitted:
11 December 2025
Posted:
12 December 2025
You are already at the latest version
Abstract
Existing animal models of post-traumatic stress disorder (PTSD) are often methodologically complex and produce variable outcomes. The aim of this study was to develop a modified PTSD model that accurately recapitulates the clinical progression of the disorder incorporating both behavioral features and objective physiological parameters. We utilized a modified Single Prolonged Stress with Subsequent Stress (SPS&S) protocol, supplemented by a stress reminder phase (without re-exposure to primary stressors) and an evaluation of stress response extinction. Eighty Wistar rats were subjected to the stress protocol, followed by comprehensive behavioral, hematological (leukocytes, hemoglobin, hematocrit), and hormonal (corticosterone, ACTH) assessments 4-5 weeks post-stress. The model produced a PTSD-like phenotype in 25% of animals, characterized by persistent alterations in the investigated biomarkers. The PTSD group exhibited sustained behavioral impairments (increased anxiety), hematological changes (neutrophilic leukocytosis), and endocrine dysregulation (decreased corticosterone, ACTH, and epinephrine). This modified SPS&S model demonstrates validity for studying the long-term consequences of stress, with PTSD markers remaining stable throughout the 28-day observation period.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Animals
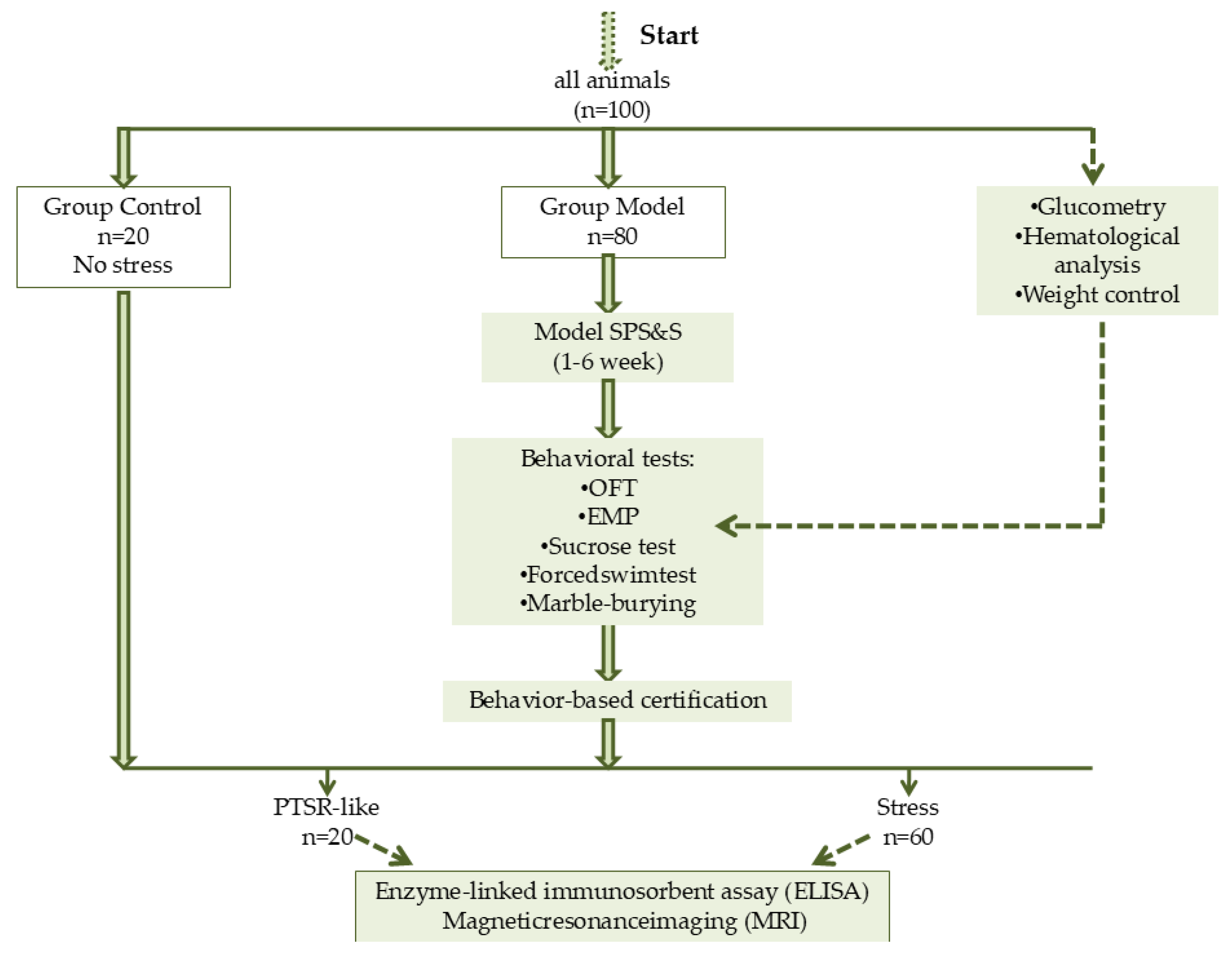
2.2. Experiment Design
2.3. Stress Procedures
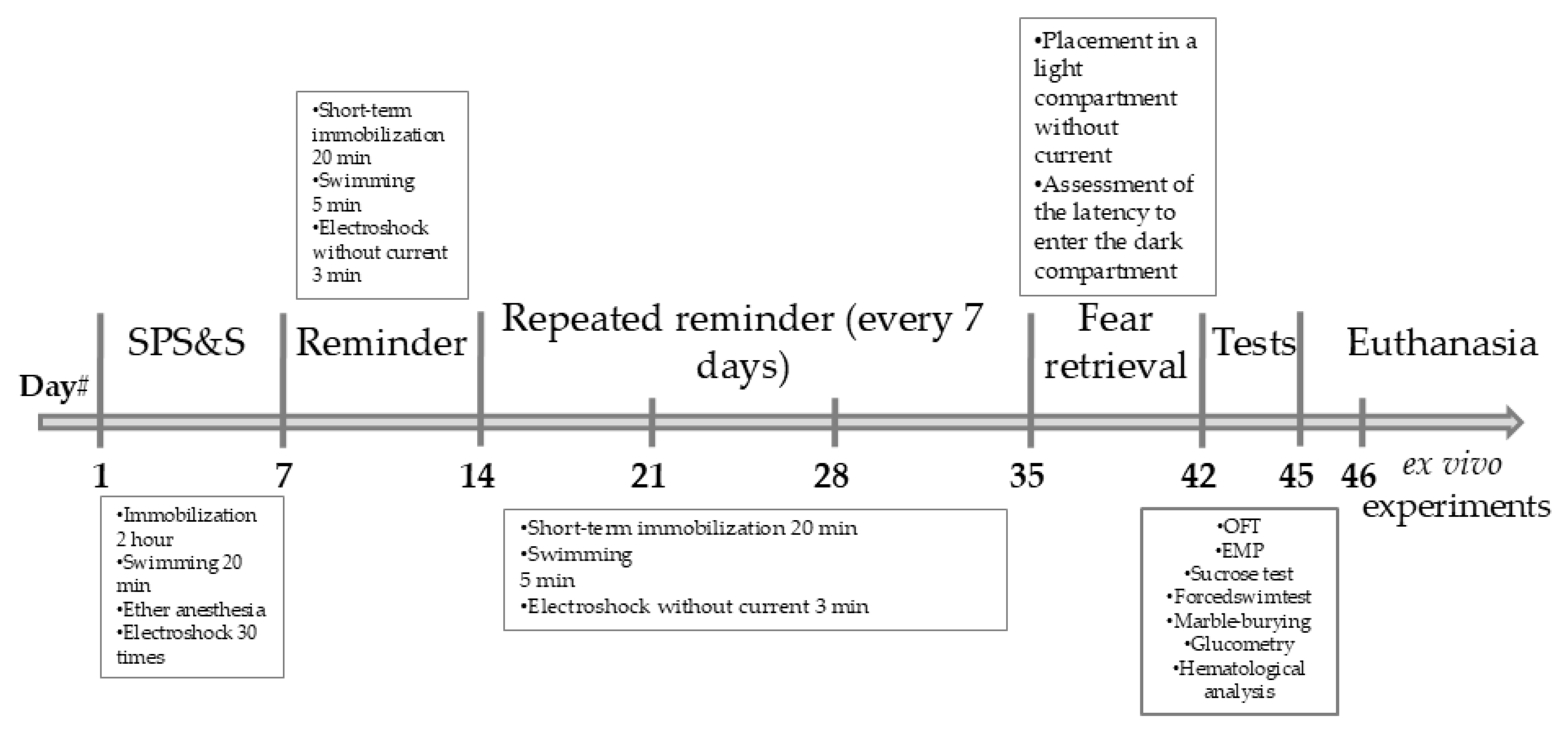
2.3.1. Acute Prolonged Stress Phase (Week 1)
- Immobilization: Rats were placed in restrainers that almost completely immobilized them for 2 hours.
- Forced Swimming: Immediately after immobilization, animals were placed in cylinders (height 65 cm, diameter 35 cm) filled with water (25 ± 1 °C) for 20 minutes.
- Ether Anesthesia: After a 15-minute recovery period, rats were exposed to diethyl ether vapor until the loss of the pain reflex.
- Electric Shock: After recovering from anesthesia, animals were placed in the dark compartment of a conditioned avoidance chamber (580 × 487 × 330 mm, Neurobotics LLC, Russia). Following a 2-minute habituation period, they received 30 electric foot shocks (1.5 mA, 1 s duration, random inter-shock interval of 30-60 s). The animals were returned to their home cages 60 seconds after the final shock.
- For the next 7 consecutive days, the rats were left undisturbed.
2.3.2. Stress Reminder Phase (Weeks 2-5)
- 6.
- Immobilization. Rats were placed in restraints that completely/almost completely immobilized the animals for 20 minutes
- 7.
- Forced swimming. Immediately after immobilization, animals were placed in cylinders (height 65 cm, diameter 30 cm) filled with water at 25±1 °C, in which the animals swam for 5 minutes
- 8.
- Electric shock. After recovery from anesthesia, animals were placed in the dark compartment of a conditioned avoidance chamber (580×487×330 mm, Russia, LLC “Neurobotics”) for 3 minutes without current application.
2.3.3. Extinction Phase (Week 6))
2.4. Physiological and Biochemical Assessments
2.4.1. Body Weight and Sucrose Preference Test
2.4.2. Hematological Parameters and Glucose Level in Peripheral Blood
2.4.3. Enzyme-Linked Immunosorbent Assay (ELISA)
2.4.4. Magnetic Resonance Imaging (MRI)
2.5. Behavioral Tests
2.5.1. Open Field Test
2.5.2. Elevated Plus-Maze Test
2.5.3. Marble-Burying
2.5.4. Forced Swim Test
2.6. Statistical Analysis
3. Results
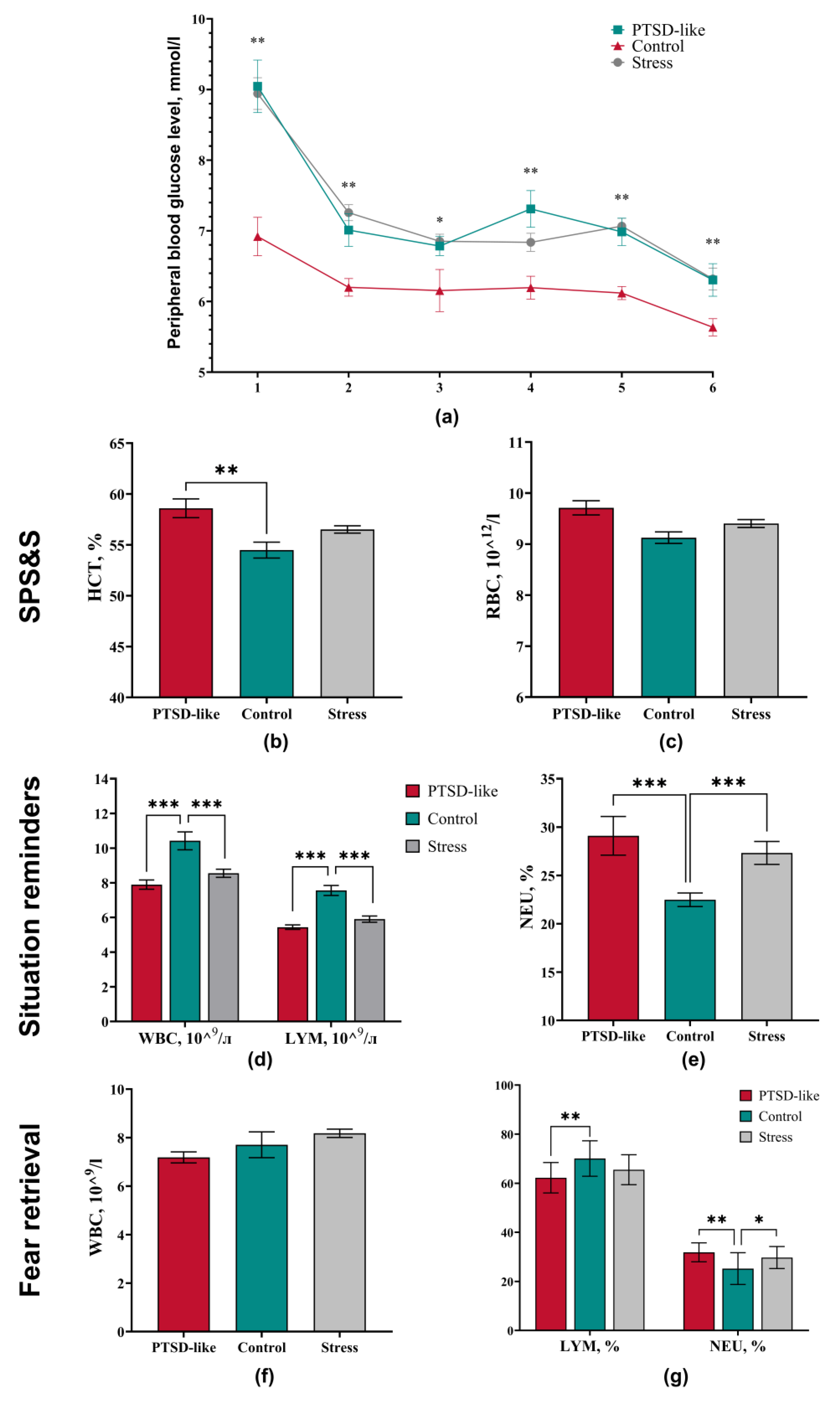
3.1. Animal Stratification into Phenotypic Subgroups
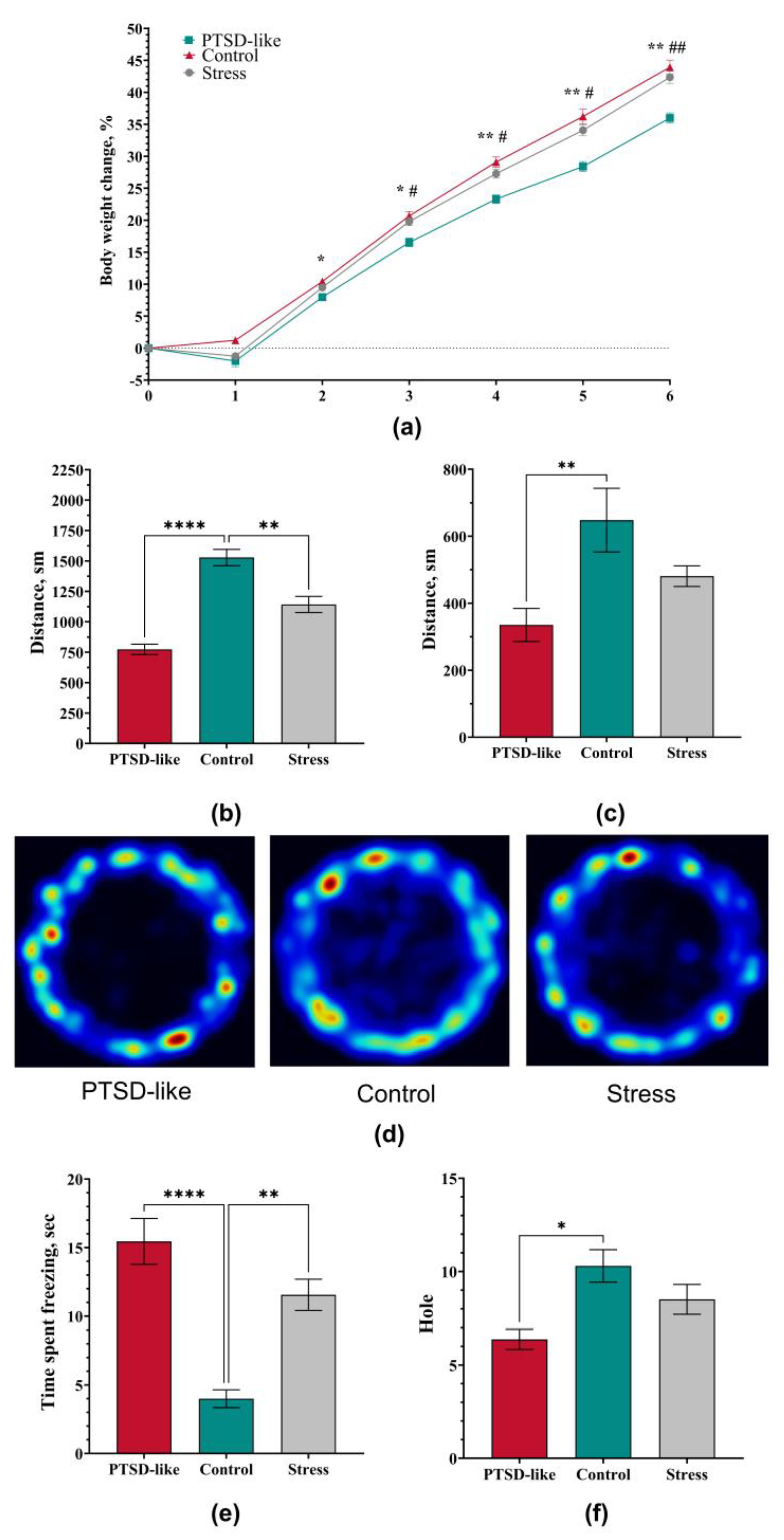
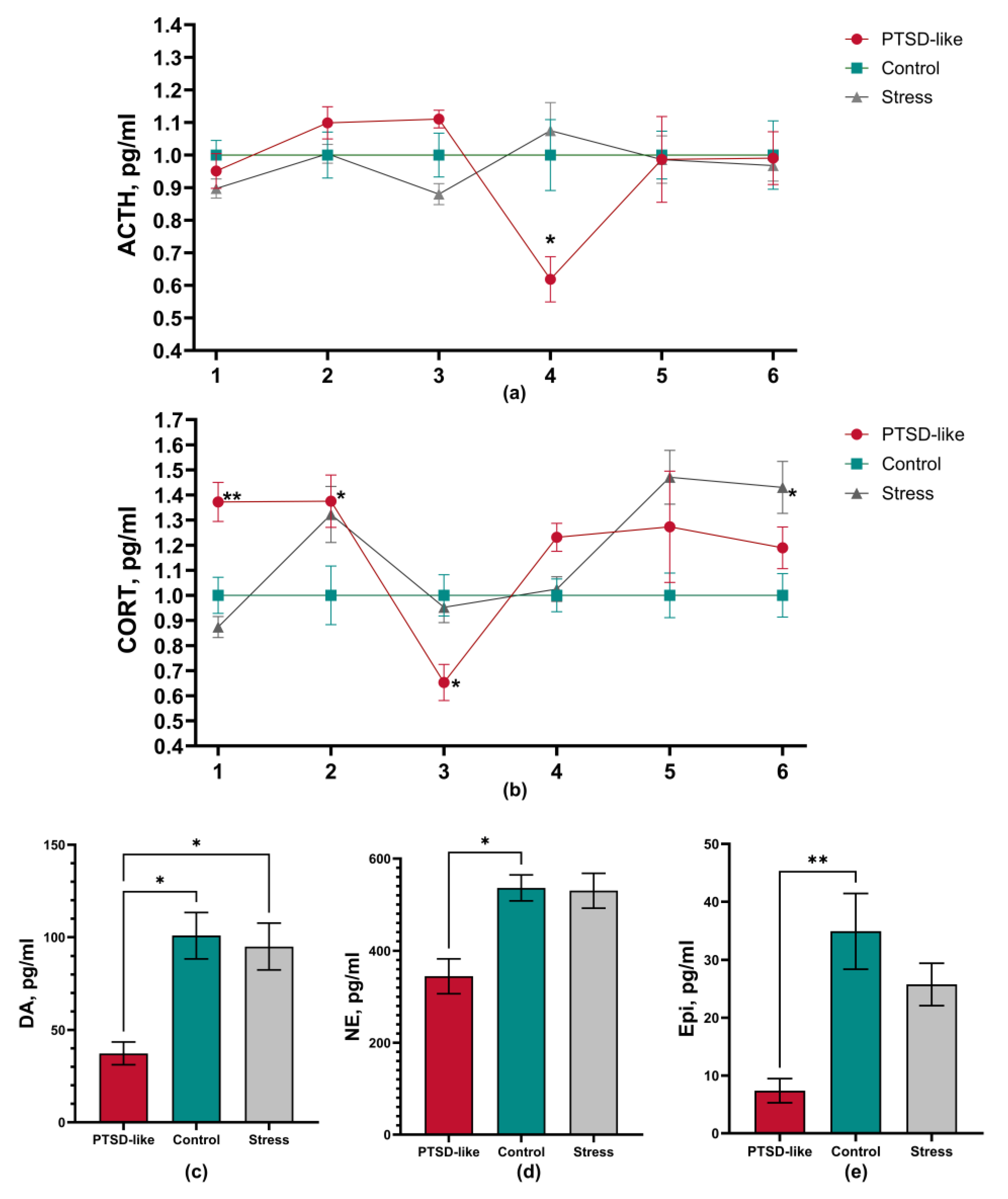
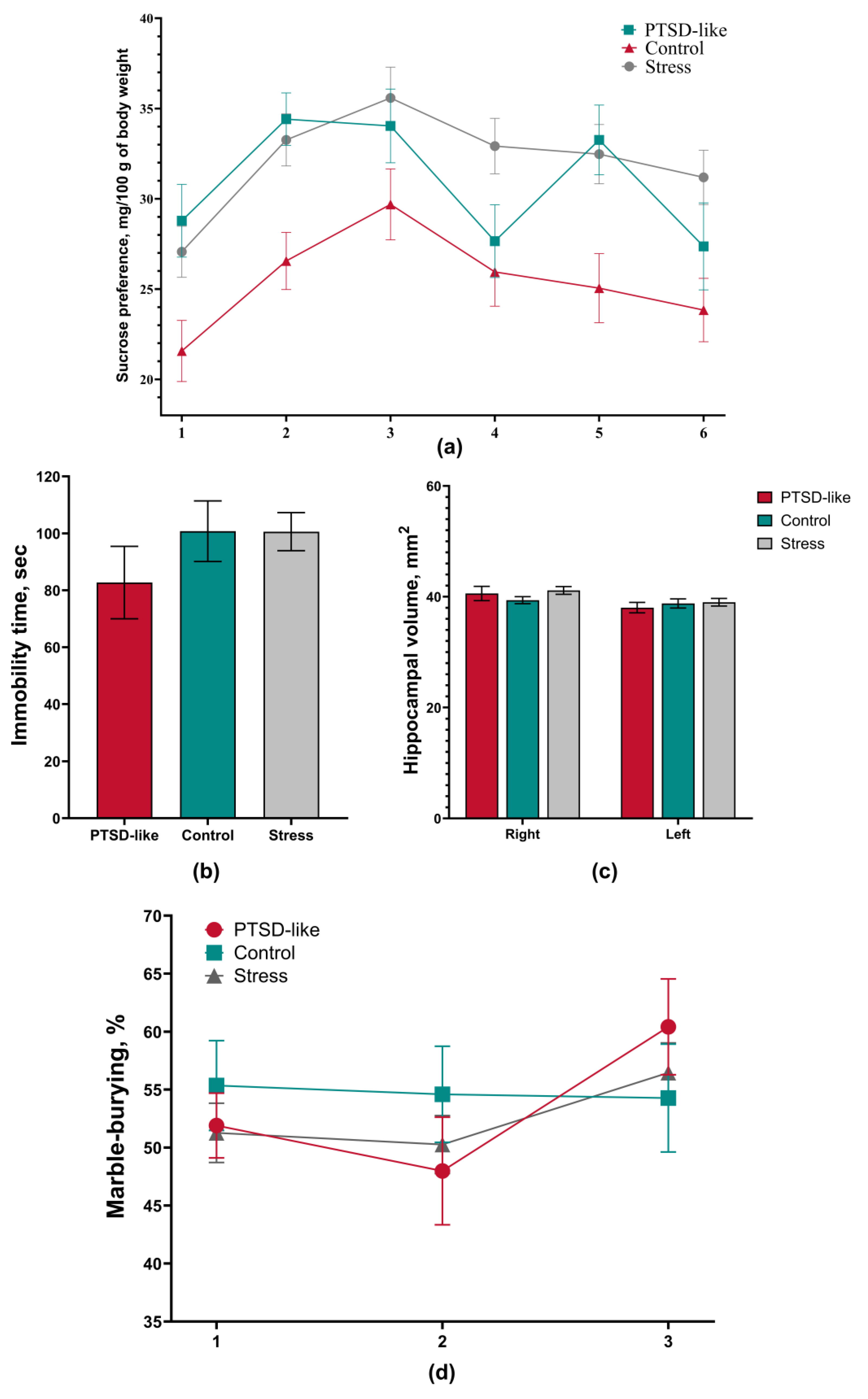
3.2. PTSD-Like Phenotype Is Characterized by Persistent Behavioral and Physiological Changes
3.3. Long-Term Stress Consequences in Physiological and Hematological Markers After Exposure to the Modified SPS&S Mode
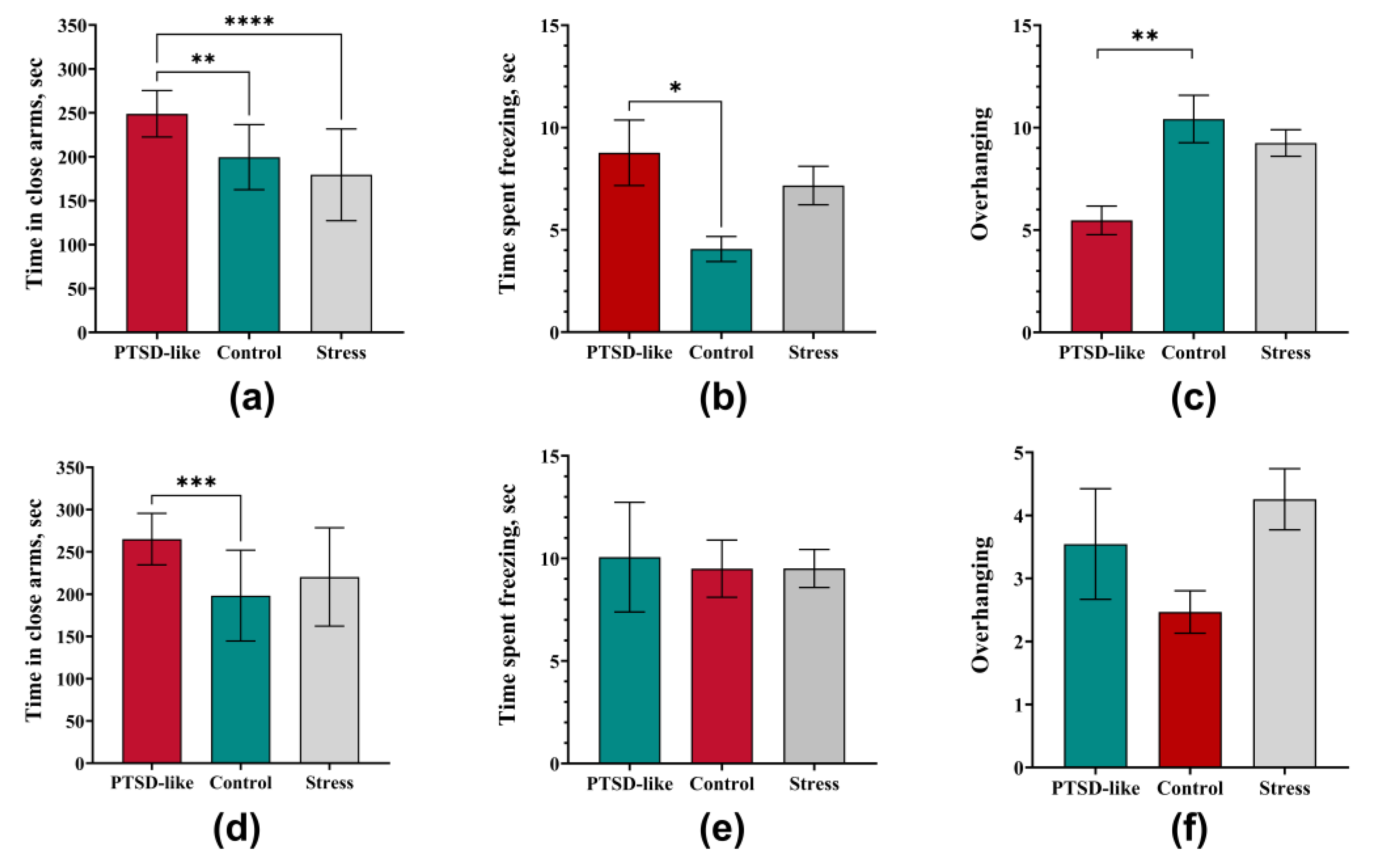
3.4. The Modified SPS&S Model Reproduces an Isolated PTSD-like State Not Accompanied by Comorbid Depressive or Obsessive-Compulsive Manifestations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACTH | Adrenocorticotropic Hormone |
| ADHD | Attention deficit hyperactivity disorder |
| ELISA | Enzyme-linked immunosorbent assay |
| HPA axis | Hypothalamic-pituitary-adrenal axis |
| MRI | Magnetic resonance imaging |
| PTSD | Post-traumatic stress disorder |
| SPS&S | Single Prolonged Stress with Subsequent Stress |
References
- James, K. A.; Stromin, J. I.; Steenkamp, N.; Combrinck, M. I. Understanding the relationships between physiological and psychosocial stress, cortisol and cognition. Front Endocrinol (Lausanne) 2023, 14, 1085950. [CrossRef]
- Maul, S.; Giegling, I.; Fabbri, C.; Corponi, F.; Serretti, A.; Rujescu, D. Genetics of resilience: Implications from genome-wide association studies and candidate genes of the stress response system in posttraumatic stress disorder and depression. Am J Med Genet B Neuropsychiatr Genet 2020, 183 (2), 77-94. [CrossRef]
- Wang, Z.; He, D.; Yang, L.; Wang, P.; Xiao, J.; Zou, Z.; Min, W.; He, Y.; Yuan, C.; Zhu, H.; et al. Similarities and differences between post-traumatic stress disorder and major depressive disorder: Evidence from task-evoked functional magnetic resonance imaging meta-analysis. J Affect Disord 2024, 361, 712-719. [CrossRef]
- Yamamoto, S.; Morinobu, S.; Takei, S.; Fuchikami, M.; Matsuki, A.; Yamawaki, S.; Liberzon, I. Single prolonged stress: toward an animal model of posttraumatic stress disorder. Depress Anxiety 2009, 26 (12), 1110-1117. [CrossRef]
- Flandreau, E. I.; Toth, M. Animal Models of PTSD: A Critical Review. Curr Top Behav Neurosci 2018, 38, 47-68. [CrossRef]
- Lisieski, M. J.; Eagle, A. L.; Conti, A. C.; Liberzon, I.; Perrine, S. A. Single-Prolonged Stress: A Review of Two Decades of Progress in a Rodent Model of Post-traumatic Stress Disorder. Front Psychiatry 2018, 9, 196. [CrossRef]
- Goswami, S.; Rodríguez-Sierra, O.; Cascardi, M.; Paré, D. Animal models of post-traumatic stress disorder: face validity. Front Neurosci 2013, 7, 89. [CrossRef]
- Wang, W.; Liu, Y.; Zheng, H.; Wang, H. N.; Jin, X.; Chen, Y. C.; Zheng, L. N.; Luo, X. X.; Tan, Q. R. A modified single-prolonged stress model for post-traumatic stress disorder. Neuroscience Letters 2008, 441 (2), 237-241. [CrossRef]
- Dykshoorn, K. L. Trauma-related obsessive-compulsive disorder: a review. Health Psychol Behav Med 2014, 2 (1), 517-528. [CrossRef]
- Willner, P.; Towell, A.; Sampson, D.; Sophokleous, S.; Muscat, R. Reduction of sucrose preference by chronic unpredictable mild stress, and its restoration by a tricyclic antidepressant. Psychopharmacology (Berl) 1987, 93 (3), 358-364. [CrossRef]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: a review. Eur J Pharmacol 2003, 463 (1-3), 3-33. [CrossRef]
- Hu, C.; Luo, Y.; Wang, H.; Kuang, S.; Liang, G.; Yang, Y.; Mai, S.; Yang, J. Re-evaluation of the interrelationships among the behavioral tests in rats exposed to chronic unpredictable mild stress. PLoS One 2017, 12 (9), e0185129. [CrossRef]
- Pellow, S.; File, S. E. Anxiolytic and anxiogenic drug effects on exploratory activity in an elevated plus-maze: a novel test of anxiety in the rat. Pharmacol Biochem Behav 1986, 24 (3), 525-529. [CrossRef]
- Walf, A. A.; Frye, C. A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat Protoc 2007, 2 (2), 322-328. [CrossRef]
- de Brouwer, G.; Fick, A.; Harvey, B. H.; Wolmarans, W. A critical inquiry into marble-burying as a preclinical screening paradigm of relevance for anxiety and obsessive-compulsive disorder: Mapping the way forward. Cogn Affect Behav Neurosci 2019, 19 (1), 1-39. [CrossRef]
- Porsolt, R. D.; Le Pichon, M.; Jalfre, M. Depression: a new animal model sensitive to antidepressant treatments. Nature 1977, 266 (5604), 730-732. [CrossRef]
- Yakhkeshi, R.; Roshani, F.; Akhoundzadeh, K.; Shafia, S. Effect of treadmill exercise on serum corticosterone, serum and hippocampal BDNF, hippocampal apoptosis and anxiety behavior in an ovariectomized rat model of post-traumatic stress disorder (PTSD). Physiol Behav 2022, 243, 113629. [CrossRef]
- Mazaheri, M.; Radahmadi, M.; Sharifi, M. R. Effects of chronic social equality and inequality conditions on passive avoidance memory and PTSD-like behaviors in rats under chronic empathic stress. Int J Neurosci 2025, 135 (8), 919-930. [CrossRef]
- Cohen, H.; Kozlovsky, N.; Alona, C.; Matar, M. A.; Joseph, Z. Animal model for PTSD: from clinical concept to translational research. Neuropharmacology 2012, 62 (2), 715-724. [CrossRef]
- Auxéméry, Y. Post-traumatic psychiatric disorders: PTSD is not the only diagnosis. Presse Med 2018, 47 (5), 423-430. [CrossRef]
- Fischer, S.; Schumacher, T.; Knaevelsrud, C.; Ehlert, U.; Schumacher, S. Genes and hormones of the hypothalamic-pituitary-adrenal axis in post-traumatic stress disorder. What is their role in symptom expression and treatment response? J Neural Transm (Vienna) 2021, 128 (9), 1279-1286. [CrossRef]
- Ball, T. M.; Gunaydin, L. A. Measuring maladaptive avoidance: from animal models to clinical anxiety. Neuropsychopharmacology 2022, 47 (5), 978-986. [CrossRef]
- Chu, B.; Marwaha, K.; Sanvictores, T.; Awosika, A. O.; Ayers, D. Physiology, Stress Reaction. In StatPearls, StatPearls Publishing Copyright © 2025, StatPearls Publishing LLC., 2025.
- Austin, A. W.; Patterson, S. M.; von Känel, R. Hemoconcentration and hemostasis during acute stress: interacting and independent effects. Ann Behav Med 2011, 42 (2), 153-173. [CrossRef]
- Dhabhar, F. S.; Miller, A. H.; Stein, M.; McEwen, B. S.; Spencer, R. L. Diurnal and acute stress-induced changes in distribution of peripheral blood leukocyte subpopulations. Brain Behav Immun 1994, 8 (1), 66-79. [CrossRef]
- Dhabhar, F. S.; Miller, A. H.; McEwen, B. S.; Spencer, R. L. Effects of stress on immune cell distribution. Dynamics and hormonal mechanisms. The Journal of Immunology 1995, 154 (10), 5511-5527. (acccessed 11/19/2025). [CrossRef]
- Domínguez-Gerpe, L.; Rey-Méndez, M. Alterations induced by chronic stress in lymphocyte subsets of blood and primary and secondary immune organs of mice. BMC Immunol 2001, 2, 7. [CrossRef]
- Zhang, C.; Liu, B.; Pawluski, J.; Steinbusch, H. W. M.; Kirthana Kunikullaya, U.; Song, C. The effect of chronic stress on behaviors, inflammation and lymphocyte subtypes in male and female rats. Behav Brain Res 2023, 439, 114220. [CrossRef]
- Sanchís-Ollé, M.; Belda, X.; Gagliano, H.; Visa, J.; Nadal, R.; Armario, A. Animal models of PTSD: Comparison of the neuroendocrine and behavioral sequelae of immobilization and a modified single prolonged stress procedure that includes immobilization. J Psychiatr Res 2023, 160, 195-203. [CrossRef]
- Brewin, C. R. Memory and Forgetting. Curr Psychiatry Rep 2018, 20 (10), 87. [CrossRef]
- Arsova, S.; Manusheva, N.; Kopacheva-Barsova, G.; Bajraktarov, S. Enduring Personality Changes after Intense Stressful Event: Case Report. Open Access Maced J Med Sci 2016, 4 (3), 453-454. [CrossRef]
- Sarapultsev, A.; Komelkova, M.; Lookin, O.; Khatsko, S.; Gusev, E.; Trofimov, A.; Tokay, T.; Hu, D. Rat Models in Post-Traumatic Stress Disorder Research: Strengths, Limitations, and Implications for Translational Studies. Pathophysiology 2024, 31 (4), 709-760. [CrossRef]
- Lawrence, S.; Scofield, R. H. Post traumatic stress disorder associated hypothalamic-pituitary-adrenal axis dysregulation and physical illness. Brain Behav Immun Health 2024, 41, 100849. [CrossRef]
- Lowrance, S. A.; Ionadi, A.; McKay, E.; Douglas, X.; Johnson, J. D. Sympathetic nervous system contributes to enhanced corticosterone levels following chronic stress. Psychoneuroendocrinology 2016, 68, 163-170. [CrossRef]
- Xu, J.; Wang, B.; Ao, H. Corticosterone effects induced by stress and immunity and inflammation: mechanisms of communication. Front Endocrinol (Lausanne) 2025, 16, 1448750. [CrossRef]
- Buonacera, A.; Stancanelli, B.; Colaci, M.; Malatino, L. Neutrophil to Lymphocyte Ratio: An Emerging Marker of the Relationships between the Immune System and Diseases. Int J Mol Sci 2022, 23 (7). [CrossRef]
- Cole, S. W.; Mendoza, S. P.; Capitanio, J. P. Social stress desensitizes lymphocytes to regulation by endogenous glucocorticoids: insights from in vivo cell trafficking dynamics in rhesus macaques. Psychosom Med 2009, 71 (6), 591-597. [CrossRef]
- Jia, W. Y.; Zhang, J. J. Effects of glucocorticoids on leukocytes: Genomic and non-genomic mechanisms. World J Clin Cases 2022, 10 (21), 7187-7194. [CrossRef]
- Gasser, P. J.; Lowry, C. A. Organic cation transporter 3: A cellular mechanism underlying rapid, non-genomic glucocorticoid regulation of monoaminergic neurotransmission, physiology, and behavior. Horm Behav 2018, 104, 173-182. [CrossRef]
- Tsetsenis, T.; Broussard, J. I.; Dani, J. A. Dopaminergic regulation of hippocampal plasticity, learning, and memory. Front Behav Neurosci 2022, 16, 1092420. [CrossRef]
- Gulino, R.; Nunziata, D.; de Leo, G.; Kostenko, A.; Emmi, S. A.; Leanza, G. Hippocampal Noradrenaline Is a Positive Regulator of Spatial Working Memory and Neurogenesis in the Rat. Int J Mol Sci 2023, 24 (6). [CrossRef]
- Kitayama, N.; Vaccarino, V.; Kutner, M.; Weiss, P.; Bremner, J. D. Magnetic resonance imaging (MRI) measurement of hippocampal volume in posttraumatic stress disorder: a meta-analysis. J Affect Disord 2005, 88 (1), 79-86. [CrossRef]
- Bremner, J. D.; Randall, P.; Scott, T. M.; Bronen, R. A.; Seibyl, J. P.; Southwick, S. M.; Delaney, R. C.; McCarthy, G.; Charney, D. S.; Innis, R. B. MRI-based measurement of hippocampal volume in patients with combat-related posttraumatic stress disorder. Am J Psychiatry 1995, 152 (7), 973-981. [CrossRef]
- Logue, M. W.; van Rooij, S. J. H.; Dennis, E. L.; Davis, S. L.; Hayes, J. P.; Stevens, J. S.; Densmore, M.; Haswell, C. C.; Ipser, J.; Koch, S. B. J.; et al. Smaller Hippocampal Volume in Posttraumatic Stress Disorder: A Multisite ENIGMA-PGC Study: Subcortical Volumetry Results From Posttraumatic Stress Disorder Consortia. Biol Psychiatry 2018, 83 (3), 244-253. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



