Submitted:
09 December 2025
Posted:
10 December 2025
You are already at the latest version
Abstract
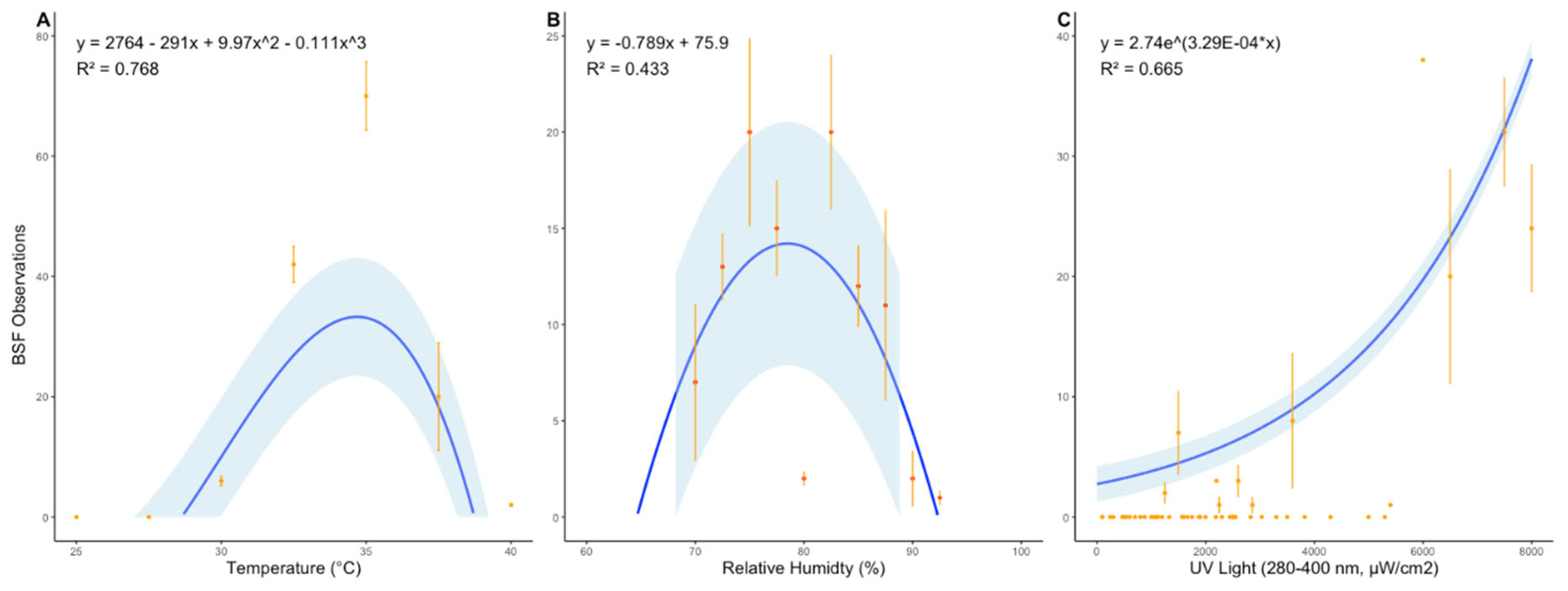
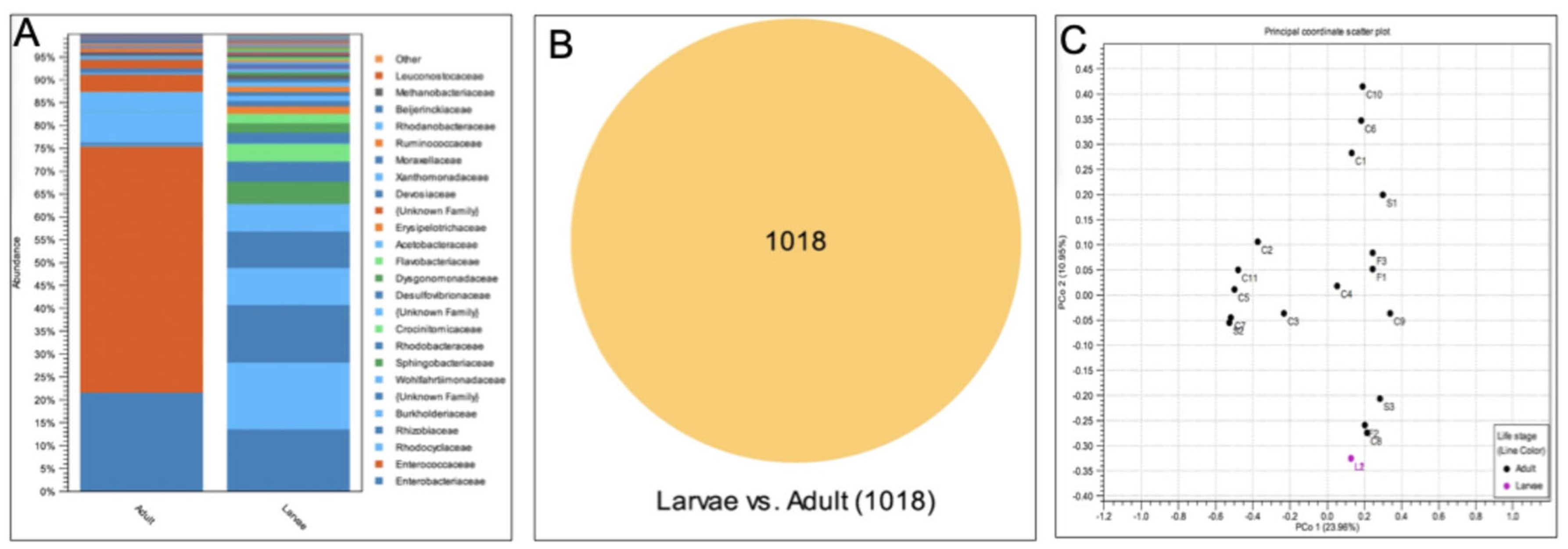
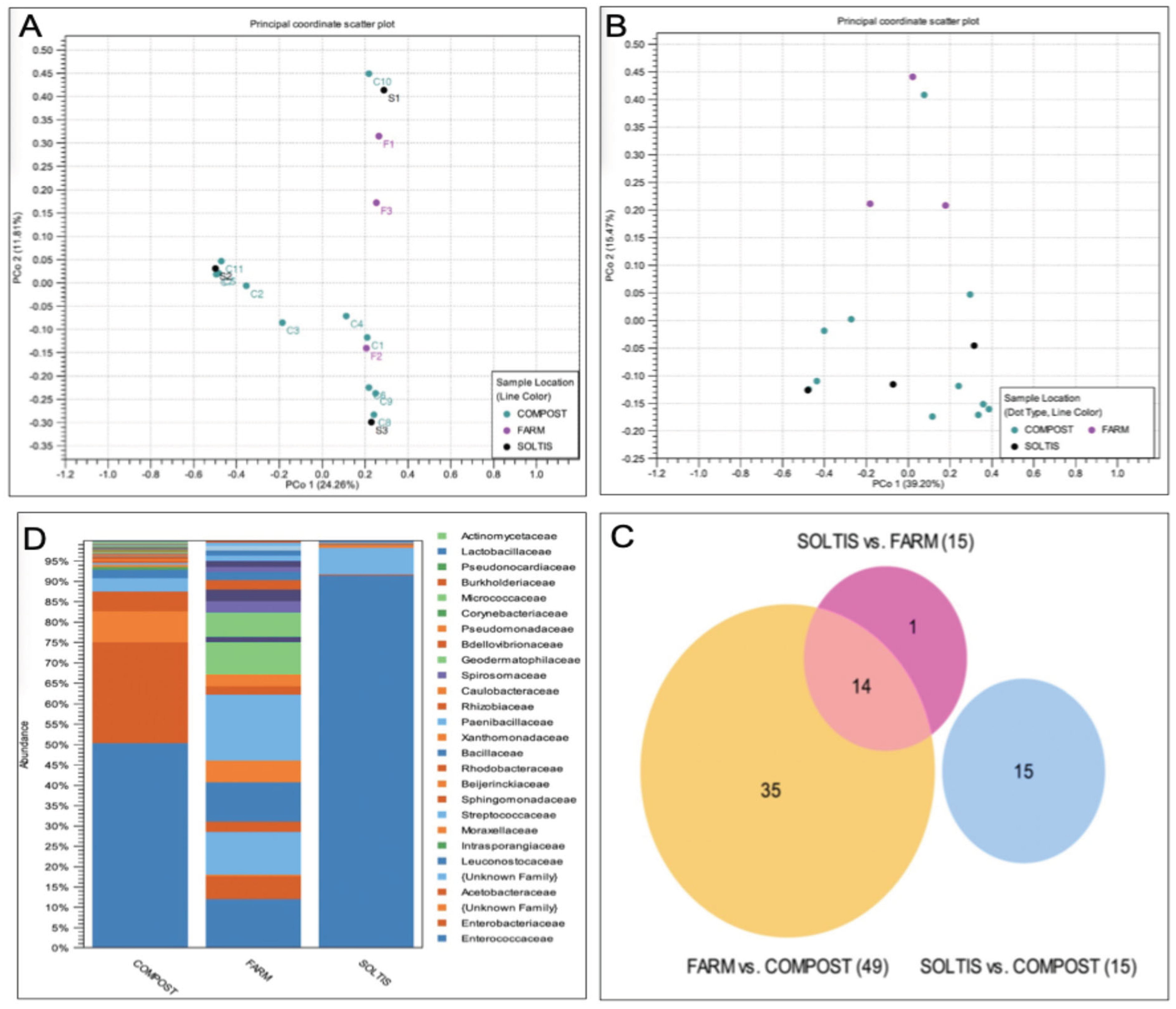
The black soldier fly (Hermetia illucens) is a globally important species, yet little is known about the behavior of adults in nature. Here, we investigated (a) whether wild populations exhibit lekking behaviors, (b) whether wild adults feed, and (c) the microbial diversity of the adult gut. Fieldwork was conducted in Alajuela Province, San Ramon, Costa Rica at three sites (a field station, a local farm, and an ecolodge compost facility). At all three sites, a highly female-skewed sex ratio (F:M=0.98) was observed. Despite many sampling attempts within 30-m of the oviposition site, no mating behaviors were directly observed within this radius, mostly due to the lack of males. Modeling confirmed female abundance declined sharply with increasing distance, and indeed no individuals were observed beyond a 7.5-m radius from the oviposition site, supporting the hypothesis that males and females may occupy distinct habitats in the wild, consistent with lekking. Modeling also revealed BSF were mostly observed between 9:00-14:30, with a peak at 12:00-noon. BSF sightings peaked at 34.7 °C, 78.5% RH, and 8000 µW/cm2 of UV-AB light, while none were observed past 40°C. The presence of vertebrate DNA in the guts of adults revealed that 100% of individuals previously fed as adults, possibly to supplement nutritional reserves during long-distance flights. Lastly, microbial community analysis revealed a core community of microbes present within the gut of wild-caught BSF. By considering spatial molecular ecology at three field sites, this study highlights the strong association of the black soldier fly with human-modified environments, suggesting environmental structure and resource availability may play a large role in shaping wild fly behavior. This work presents the first detailed field report of H. illucens in the Neotropics and offers some critical insights for future fieldwork as well as baseline traits for industrial applications.
Keywords:
Introduction
Methods
Results
Discussion
Conclusions
Supplementary Materials
Funding
Acknowledgments
References
- Alcock, J. A large male competitive advantage in a lekking fly, Hermetia comstocki Williston (Diptera: Stratiomyidae). Psyche: A Journal of Entomology 1990, 97, 267–279. [Google Scholar] [CrossRef]
- Alcock, J. Possible causes of variation in territory tenure in a lekking pompilid wasp (Hemipepsis ustulata) (Hymenoptera). Journal of Insect Behavior 2000, 13, 439–453. [Google Scholar] [CrossRef]
- Alcock, J.; Gwynne, D.T.; Dadour, I.R. Acoustic signaling, territoriality, and mating in whistling moths,Hecatesia thyridion (Agaristidae). Journal of Insect Behavior 1989, 2, 27–37. [Google Scholar] [CrossRef]
- Ancona, S.; Dénes, F.V.; Krüger, O.; Székely, T.; Beissinger, S.R. Estimating adult sex ratios in nature. Philosophical Transactions of the Royal Society B: Biological Sciences 2017, 372, 20160313. [Google Scholar] [CrossRef]
- Andrade, D.; Pasini, F.; Scarano, F.R. Syntropy and innovation in agriculture. Current Opinion in Environmental Sustainability 2020, 45, 20–24. [Google Scholar] [CrossRef]
- Auger, L.; Tegtmeier, D.; Caccia, S.; Klammsteiner, T.; De Smet, J. BugBook: How to explore and exploit the insect-associated microbiome 2025. [CrossRef]
- Barbosa, F. An integrative view of postcopulatory sexual selection in a soldier fly: Interplay between cryptic male choice and sperm competition. In Cryptic Female Choice in Arthropods: Patterns, Mechanisms and Prospects; Peretti, A. V., Aisenberg, A., Eds.; Springer International Publishing, 2015; pp. 385–401. [Google Scholar] [CrossRef]
- Barbosa, T.M.; Jales, J.T.; Medeiros, J.R.; Gama, R.A. Sarcosaprophagous dipterans associated with differentially-decomposed substrates in Atlantic Forest environments. Acta Brasiliensis 2023, 7, 14–21. [Google Scholar] [CrossRef]
- Barrett, M.; Chia, S.Y.; Fischer, B.; Tomberlin, J.K. Welfare considerations for farming black soldier flies, Hermetia illucens (Diptera: Stratiomyidae): a model for the insects as food and feed industry. Journal of Insects as Food and Feed 2022, 1–30. [Google Scholar] [CrossRef]
- Barrett, M.; Patel, N.; McCarry, B.; Shellenberger, G.; Schwartz, E.; Fiocca, K.; Waddell, E. Dietary preferences and impacts of feeding on behavior, longevity, and reproduction in adult black soldier flies (Diptera: Stratiomyidae; Hermetia illucens). Journal of Insects as Food and Feed 2025, Online, 1–12. [Google Scholar] [CrossRef]
- Beehler, B.M.; Foster, M.S. Hotshots, hotspots, and female preference in the organization of lek mating systems. The American Naturalist 1988, 131, 203–219. [Google Scholar] [CrossRef]
- Beenakkers, A.M. Th.; Van der Horst, D.J.; Van Marrewijk, W.J.A. Insect flight muscle metabolism. Insect Biochemistry 1984, 14, 243–260. [Google Scholar] [CrossRef]
- Booth, D.C.; Sheppard, C. Oviposition of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae): Eggs, masses, timing, and site characteristics. Environmental Entomology 1984, 13, 421–423. [Google Scholar] [CrossRef]
- Bradbury, J.; Gibson, R.; Tsai, I.M. Hotspots and the dispersion of leks. Animal Behaviour 1986, 34, 1694–1709. [Google Scholar] [CrossRef]
- Bradbury, J.W.; Gibson, R.M. Leks and mate choice. In Mate Choice; Bateson, P., Ed.; Cambridge University Press, 1983; pp. 109–140. [Google Scholar]
- Breviglieri, C.P.B.; Romero, G.Q.; Mega, A.C.G.; da Silva, F.R. Are Cecropia trees ecosystem engineers? The effect of decomposing Cecropia leaves on arthropod communities. Biotropica 2019, 51, 562–571. [Google Scholar] [CrossRef]
- Brown, J.S.; Laundré, JW; Gurung, M. The ecology of fear: Optimal foraging, game theory, and trophic interactions. Journal of Mammalogy 1999, 80, 385–399. [Google Scholar] [CrossRef]
- Bruno, D.; Bonelli, M.; Cadamuro, A.G.; Reguzzoni, M.; Grimaldi, A.; Casartelli, M.; Tettamanti, G. The digestive system of the adult Hermetia illucens (Diptera: Stratiomyidae): Morphological features and functional properties. Cell and Tissue Research 2019, 378, 221–238. [Google Scholar] [CrossRef]
- Buzatto, B.A.; Machado, G. Resource defense polygyny shifts to female defense polygyny over the course of the reproductive season of a Neotropical harvestman. Behavioral Ecology and Sociobiology 2008, 63, 85–94. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinformatics 2009, 10, 421. [Google Scholar] [CrossRef]
- Chiabotto, C.; Grosso, F.; Doretto, A.; Meneguz, M. Observation of mating behavior using marked flies of black soldier fly (Hermetia illucens) under sunlight condition. Journal of Insects as Food and Feed 2024, 10, 2017–2029. [Google Scholar] [CrossRef]
- Copello, A. Biologia de Hemetia illuscens Latr. (La mosca de nuetras colmenas). Revista de La Sociedata Entomologica Argentina 1926, 1, 1–5. [Google Scholar]
- Cornaby, B.W. Carrion reduction by animals in contrasting tropical habitats. Biotropica 1974, 6, 51–63. [Google Scholar] [CrossRef]
- da Silva, G.D.P.; Hesselberg, T. A review of the use of black soldier fly larvae, Hermetia illucens (Diptera: Stratiomyidae), to compost organic waste in tropical regions. Neotropical Entomology 2020, 49, 151–162. [Google Scholar] [CrossRef]
- Diener, S.; Studt Solano, N.M.; Roa Gutiérrez, F.; Zurbrügg, C.; Tockner, K. Biological treatment of municipal organic waste using black soldier fly larvae. Waste and Biomass Valorization 2011, 2, 357–363. [Google Scholar] [CrossRef]
- Dodson, G. Lek mating system and large male aggressive advantage in a gall-forming tephritid fly (Diptera: Tephritidae). Ethology 1986, 72, 99–108. [Google Scholar] [CrossRef]
- Emlen, S.T.; Oring, L.W. Ecology, sexual selection, and the evolution of mating systems. Science 1977, 197, 215–223. [Google Scholar] [CrossRef]
- Fachin, D.A.; González, C.R.; Elgueta, M.; Hauser, M. A catalog of Stratiomyidae (Diptera: Brachycera) from Chile, with a new synonym and notes on the species. Zootaxa 2021, 5004, 1–57. [Google Scholar] [CrossRef] [PubMed]
- Fachin, D.A.; Hauser, M. Large flies overlooked: The genus Hermetia Latreille, 1804 (Diptera: Stratiomyidae) in the Neotropics, with 11 synonyms and a new species to Brazil. Neotropical Entomology 2022, 51, 660–690. [Google Scholar] [CrossRef]
- Fu, J.; Wang, J.; Huang, X.; Guan, B.; Feng, Q.; Deng, H. Composition and diversity of gut microbiota across developmental stages of Spodoptera frugiperda and its effect on the reproduction. Frontiers in Microbiology 2023, 14. [Google Scholar] [CrossRef]
- Furman, D.P.; Young, R.D.; Catts Paul, E. Hermetia illucens (Linnaeus) as a Factor in the Natural Control of Musca domestica Linnaeus. Journal of Economic Entomology 1959, 52, 917–921. [Google Scholar] [CrossRef]
- Gasnier, T.R.; Magnusson, W.E.; Lima, A.P. Foraging activity and diet of four sympatric lizard species in a tropical rainforest. Journal of Herpetology 1994, 28, 187–192. [Google Scholar] [CrossRef]
- Giunti, G.; Campolo, O.; Laudani, F.; Palmeri, V. Male courtship behaviour and potential for female mate choice in the black soldier fly Hermetia illucens L. (Diptera: Stratiomyidae). Entomologia Generalis 2018, 38, 29–46. [Google Scholar] [CrossRef]
- Harjoko, D.N.; Hua, Q.Q.H.; Toh, E.M.C.; Goh, C.Y.J.; Puniamoorthy, N. A window into fly sex: Mating increases female but reduces male longevity in black soldier flies. Animal Behaviour 2023, 200, 25–36. [Google Scholar] [CrossRef]
- Hastings, J.M.; Dodson, G.N.; Heckman, J.L. Male perch selection and the mating system of the robber fly,Promachus albifacies (Diptera: Asilidae). Journal of Insect Behavior 1994, 7, 829–841. [Google Scholar] [CrossRef]
- Herberstein, M.E.; Painting, C.J.; Holwell, G.I. Scramble competition polygyny in terrestrial arthropods. In Advances in the study of behavior; Naguib, M., Podos, J., Simmons, L.W., Barrett, L., Healy, S.D., Zuk, M., Eds.; Elsevier, 2017; Vol. 49, pp. 237–295. [Google Scholar] [CrossRef]
- Hoc, B.; Noël, G.; Carpentier, J.; Francis, F.; Megido, R.C. Optimization of black soldier fly (Hermetia illucens) artificial reproduction. PLOS ONE 2019, 14, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, L.; Hull, K.L.; Bierman, A.; Badenhorst, R.; Bester-van der Merwe, A.E.; Rhode, C. Patterns of genetic diversity and mating systems in a mass-reared black soldier fly colony. Insects 2021, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Höglund, J.; Alatalo, R.V. Leks; 1995. [Google Scholar]
- James, M.T. The genus Hermetia in the United States (Diptera: Stratiomyidae). Bulletin of the Brooklyn Entomological Society 1935, 30, 165–170. Available online: https://biostor.org/reference/169315.
- Jensen, K.; Thormose, S.F.; Noer, N.K.; Schou, T.M.; Kargo, M.; Gligorescu, A.; Nørgaard, J.V.; Hansen, L.S.; Zaalberg, R.M.; Nielsen, H.M.; Kristensen, T.N. Controlled and polygynous mating in the black soldier fly: Advancing breeding programs utilizing quantitative genetic designs 2025. [CrossRef]
- Kaufmann, C.; Reim, C.; Blanckenhorn, W.U. Size-dependent insect flight energetics at different sugar supplies. Biological Journal of the Linnean Society 2013, 108, 565–578. [Google Scholar] [CrossRef]
- Kaya, C.; Generalovic, T.N.; Ståhls, G.; Hauser, M.; Samayoa, A.C.; Nunes-Silva, C.G.; Roxburgh, H.; Wohlfahrt, J.; Ewusie, E.A.; Kenis, M.; Hanboonsong, Y.; Orozco, J.; Carrejo, N.; Nakamura, S.; Gasco, L.; Rojo, S.; Tanga, C.M.; Meier, R.; Rhode, C.; Sandrock, C. Global population genetic structure and demographic trajectories of the black soldier fly, Hermetia illucens. BMC Biology 2021, 19, 94. [Google Scholar] [CrossRef]
- Khamis, F.M.; Ombura, F.L.O.; Akutse, K.S.; Subramanian, S.; Mohamed, S.A.; Fiaboe, K.K.M.; Saijuntha, W.; Van Loon, J.J.A.; Dicke, M.; Dubois, T.; Ekesi, S.; Tanga, C.M. Insights in the global genetics and gut microbiome of black soldier fly, Hermetia illucens: Implications for animal feed safety control. Frontiers in Microbiology 2020, 11. Available online: https://www.frontiersin.org/articles/10.3389/fmicb.2020.01538. [CrossRef]
- Kimsey, L.S. The behaviour of male orchid bees (Apidae, Hymenoptera, Insecta) and the question of leks. Animal Behaviour 1980, 28, 996–1004. [Google Scholar] [CrossRef]
- Klüber, P.; Arous, E.; Zorn, H.; Rühl, M. Protein- and carbohydrate-rich supplements in feeding adult black soldier flies (Hermetia illucens) affect life history traits and egg productivity. Life 2023, 13, Article 2. [Google Scholar] [CrossRef]
- Kotzé, Z; Tomberlin, J.K. Influence of substrate age and interspecific colonization on oviposition behavior of a generalist feeder, black soldier fly (Diptera: Straiomyidae), on carrion. Journal of Medical Entomology 2020, 57, 987–993. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-Index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Applied and Environmental Microbiology 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed]
- Laudani, F.; Campolo, O.; Latella, I.; Modafferi, A.; Palmeri, V.; Giunti, G. Does Hermetia illucens recognize sibling mates to avoid inbreeding depression? Entomologia Generalis 2024, 44, 1225–1232. [Google Scholar] [CrossRef]
- Lemke, N.B.; Dickerson, A.J.; Tomberlin, J.K. No neonates without adults. BioEssays 2023, 45, 2200162. [Google Scholar] [CrossRef]
- Lemke, N.B.; Li, C.; De Smet, J.; Tomberlin, J.K. Temporal trends: Phase-shifted time-series analysis reveals highly correlated reproductive behaviors in the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae) (p. 2025.08.26.672371). bioRxiv 2025. [Google Scholar] [CrossRef]
- Lemke, N.B.; Li, C.; Dickerson, A.J.; Salazar, D.A.; Rollinson, L.N.; Mendoza, J.E.; Miranda, C.D.; Crawford, S.; Tomberlin, J.K. Heterogeny in cages: Age-structure and attractant availability impacts fertile egg production in the black soldier fly, Hermetia illucens. Journal of Insects as Food and Feed. Online. 2025. [CrossRef]
- Lemke, N.B.; Rollison, L.N.; Tomberlin, J.K. Sex-specific perching: Monitoring of artificial plants reveals dynamic female-biased perching behavior in the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Insects 2024, 15, 770. [Google Scholar] [CrossRef]
- Lemke, N.; Puniamoorthy, N. Beyond the Black Box: Reproductive Strategies of the Black Soldier Fly as a Model for Bridging Evolutionary Biology and Applied Entomology. Biology and Life Sciences 2025. [Google Scholar] [CrossRef]
- Lima, S.L. Predators and the breeding bird: Behavioral and reproductive flexibility under the risk of predation. Biological Reviews 2009, 84, 485–513. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, L. The game birds and wild fowl of Sweden and Norway: With an account of the seals and salt-water fishes of those countries; F. Warne and Company, 1867. [Google Scholar]
- Manas, F.; Labrousse, C.; Bressac, C. Plastic responses in sperm expenditure to sperm competition risk in black soldier fly (Hermetia illucens, Diptera) males. Journal of Insect Physiology 2025, 161, 104751. [Google Scholar] [CrossRef] [PubMed]
- Manas, F.; Piterois, H.; Labrousse, C.; Beaugeard, L.; Uzbekov, R.; Bressac, C. Gone but not forgotten: Dynamics of sperm storage and potential ejaculate digestion in the black soldier fly Hermetia illucens. Royal Society Open Science 2024, 11, 241205. [Google Scholar] [CrossRef] [PubMed]
- Manas, F.; Venon, P.; Yang, L.; Labrousse, C.; Bressac, C. Multiple mating is not driven by size and sperm management in black soldier fly (Hermetia illucens). Entomologia Experimentalis et Applicata 2025, 173, 815–825. [Google Scholar] [CrossRef]
- Munsch-Masset, P.; Labrousse, C.; Beaugeard, L.; Bressac, C. The reproductive tract of the black soldier fly (Hermetia illucens) is highly differentiated and suggests adaptations to sexual selection. Entomologia Experimentalis et Applicata 2023, n/a. [Google Scholar] [CrossRef]
- Muraro, T.; Lalanne, L.; Pelozuelo, L.; Calas-List, D. Mating and oviposition of a breeding strain of black soldier fly Hermetia illucens (Diptera: Stratiomyidae): polygynandry and multiple egg-laying. Journal of Insects as Food and Feed 2024, 1(aop), 1–13. [Google Scholar] [CrossRef]
- Norris, K.R. The bionomics of blow flies. Annual Review of Entomology 1965, 10, 47–68. [Google Scholar] [CrossRef]
- Nyakeri, E.M.; Ogola, H.J.O.; Ayieko, M.A.; Amimo, F.A. Valorisation of organic waste material: Growth performance of wild black soldier fly larvae (Hermetia illucens) reared on different organic wastes. Journal of Insects as Food and Feed 2017, 3, 193–202. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; Volk, N.; Diehl, J.J.E.; van Loon, J.J.A.; Belušič. G Photoreceptor spectral sensitivity of the compound eyes of black soldier fly (Hermetia illucens) informing the design of LED-based illumination to enhance indoor reproduction. Journal of Insect Physiology 2016, 95, 133–139. [Google Scholar] [CrossRef]
- Owings, C.G. Developmental plasticity of Cochliomyia macellaria Fabricius (Diptera: Calliphoridae) from three distinct ecoregions in Texas. PhD Thesis. 2012. Available online: https://oaktrust.library.tamu.edu/items/093ad7a9-ca1d-49d5-b3d1-8cf965af9a20.
- Owings, C.G.; Banerjee, A.; Asher, T.M.D.; Gilhooly, W.P.; Tuceryan, A.; Huffine, M.; Skaggs, C.L.; Adebowale, I.M.; Manicke, N.E.; Picard, C.J. Female blow flies as vertebrate resource indicators. Scientific Reports 2019, 9, 10594. [Google Scholar] [CrossRef]
- Price, E.O. Animal domestication and behavior; CABI Pub, 2002. [Google Scholar]
- Rathore, A.; Isvaran, K.; Guttal, V. Lekking as collective behaviour. Philosophical Transactions of the Royal Society B: Biological Sciences 2023, 378, 20220066. [Google Scholar] [CrossRef] [PubMed]
- Rhode, C.; Badenhorst, R.; Hull, K.L.; Greenwood, M.P.; der Merwe, A.E.B.; Andere, A.A.; Picard, C.J.; Richards, C. Genetic and phenotypic consequences of early domestication in black soldier flies (Hermetia illucens). Animal Genetics 2020, 51, 752–762. [Google Scholar] [CrossRef]
- Salari, S.P.; De Goede, M.L. (2024, June 20). Study into Hermetia illucens behaviour in large artificial mating chambers and possible optimizations. 5th Insects to Feed the World, Singapore Expo, Singapore.
- Sandrock, C.; Kaya, C.; Kapun, M.; Wohlfahrt, J.; Blanckenhorn, W.U. (2021). Deciphering the evolutionary history of the black soldier fly, Hermetia illucens, on a global scale. Book of Abstracts of the 72nd Annual Meeting of the European Federation of Animal Sciences. Davos, Switzerland. 30 August - 3 September 2021, 295. 295. Available online: https://orgprints.org/id/eprint/43302/.
- Shelly, T.E. Sexual selection on leks: A fruit fly primer. Journal of Insect Science 2018, 18. [Google Scholar] [CrossRef]
- Shelly, T.E.; Whittier, T.S. Lek distribution in the Mediterranean fruit fly (Diptera: Tephritidae): Influence of tree size, foliage density, and neighborhood. Proceedings of the Hawaiian Entomological Society 1993, 32, 113–121. [Google Scholar]
- Sheppard, C.; Tomberlin, J.; Joyce, J.; Kiser, B.; Sumner, S. Rearing methods for the black soldier fly (Diptera: Stratiomyidae). Journal of Medical Entomology 2002, 39, 695–698. [Google Scholar] [CrossRef]
- The evolution of insect mating systems; Shuker, D.M., Simmons, L.W., Eds.; Oxford University Press, 2014. [Google Scholar]
- Silva, J.P.; Moreira, F.; Palmeirim, J.M. Spatial and temporal dynamics of lekking behaviour revealed by high-resolution GPS tracking. Animal Behaviour 2017, 129, 197–204. [Google Scholar] [CrossRef]
- Spielman, A. Bionomics of autogenous mosquitoes. Annual Review of Entomology 1971, 16, 231–248. [Google Scholar] [CrossRef]
- Stephens, P.A.; Boyd, I.L.; McNamara, J.M.; Houston, A.I. Capital breeding and income breeding: Their meaning, measurement, and worth. Ecology 2009, 90, 2057–2067. [Google Scholar] [CrossRef]
- Studt-Solano, N.M.; Flores-Mora, D. Uso de larvas de mosca soldado negro (Hermetia illucens) para el manejo de residuos municipales orgánicos en el campus de la Universidad Earth, Costa Rica. 2010. Available online: https://agris.fao.org/search/en/providers/124451/records/66470d98e62948cba00ad208.
- Sugiura, S. Predators as drivers of insect defenses. Entomological Science 2020, 23, 316–337. [Google Scholar] [CrossRef]
- Tettamanti, G.; Van Campenhout, L.; Casartelli, M. A hungry need for knowledge on the black soldier fly digestive system. Journal of Insects as Food and Feed 2022, 8, 217–222. [Google Scholar] [CrossRef]
- Thinn, A.A.; Kainoh, Y. Effect of diet on the longevity and oviposition performance of black soldier flies, Hermetia illucens (Diptera: Stratiomyidae). Japan Agricultural Research Quarterly: JARQ 2022, 56, 211–217. [Google Scholar] [CrossRef]
- Thornhill, R.; Alcock, J. The evolution of insect mating systems; Harvard University Press, 1983. [Google Scholar]
- Tingle, F.C.; Mitchell, E.R.; Copeland, W.W. The soldier fly Hermetia Illucens in poultry houses in North Central Florida. Journal of the Georgia Entomological Society 1975. [Google Scholar]
- Tomberlin, J.K.; Sheppard, D.C. Lekking behavior of the black soldier fly (Diptera: Stratiomyidae). Florida Entomologist 2001, 84, 729–730. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; Sheppard, D.C.; Joyce, J.A. Selected life-history traits of black soldier flies (Diptera: Stratiomyidae) reared on three artificial diets. Annals of the Entomological Society of America 2002, 95, 379–386. [Google Scholar] [CrossRef]
- Tomberlin, J.; Sheppard, D. Factors influencing mating and oviposition of black soldier flies (Diptera: Straiomyidae) in a colony. Journal of Entomological Science 2002, 37, 345–352. [Google Scholar] [CrossRef]
- Van Huis, A. Welfare of farmed insects. Journal of Insects as Food and Feed 2021, 7, 573–584. [Google Scholar] [CrossRef]
- Van Looveren, N.; IJdema, F.; van der Heijden, N.; Van Der Borght, M.; Vandeweyer, D. Microbial dynamics and vertical transmission of Escherichia coli across consecutive life stages of the black soldier fly (Hermetia illucens). Animal Microbiome 2024, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- van Rijn, P.C.J.; Kooijman, J.; WäckersF, L. The contribution of floral resources and honeydew to the performance of predatory hoverflies (Diptera: Syrphidae). Biological Control 2013, 67, 32–38. [Google Scholar] [CrossRef]
- Wise, D.H. Spiders in ecological webs; Cambridge University Press, 1995. [Google Scholar]
- Xiao, K.; Shen, K.; Zhong, J.-F.; Li, G.-Q. Effects of dietary sodium on performance, flight and compensation strategies in the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Frontiers in Zoology 2010, 7, 11. [Google Scholar] [CrossRef]
- Yuval, B.; Kaspi, R.; Shloush, S.; Warburg, M.S. Nutritional reserves regulate male participation in Mediterranean fruit fly leks. Ecological Entomology 1998, 23, 211–215. [Google Scholar] [CrossRef]
- Zahavi, A.; Zahavi, A. The handicap principle: A missing piece of Darwin’s puzzle; Oxford University Press, 1999. [Google Scholar]
- Zhang, J.; Huang, L.; He, J.; Tomberlin, J.K.; Li, J.; Lei, C.; Sun, M.; Liu, Z.; Yu, Z. An artificial light source influences mating and oviposition of black soldier flies, Hermetia illucens. Journal of Insect Science 2010, 10, 1–7. [Google Scholar] [CrossRef]
- Zheng, L.; Crippen, T.L.; Singh, B.; Tarone, A.M.; Dowd, S.; Yu, Z.; Wood, T.K.; Tomberlin, J.K. A survey of bacterial diversity from successive life stages of black soldier fly (Diptera: Stratiomyidae) by Using 16S rDNA pyrosequencing. Journal of Medical Entomology 2013, 50, 647–658. [Google Scholar] [CrossRef] [PubMed]
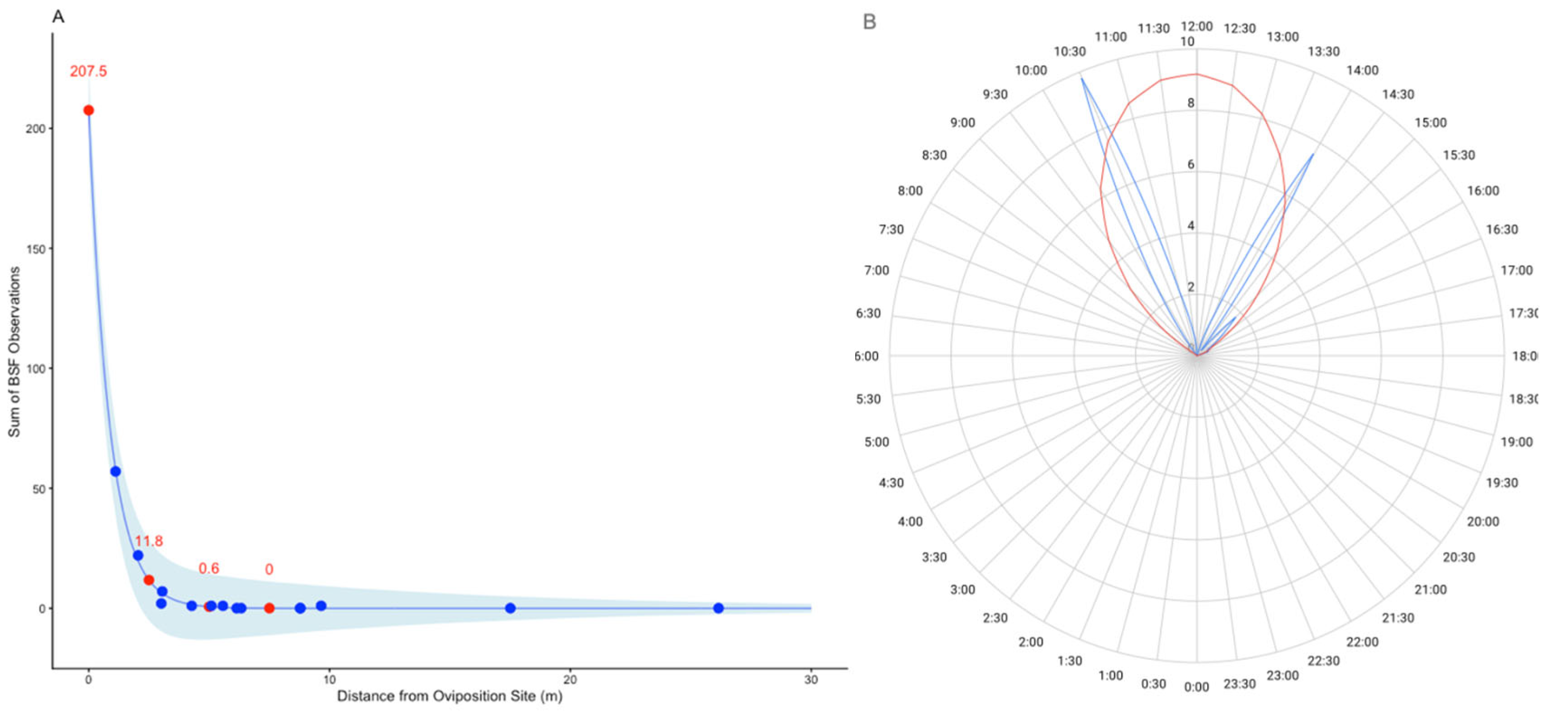
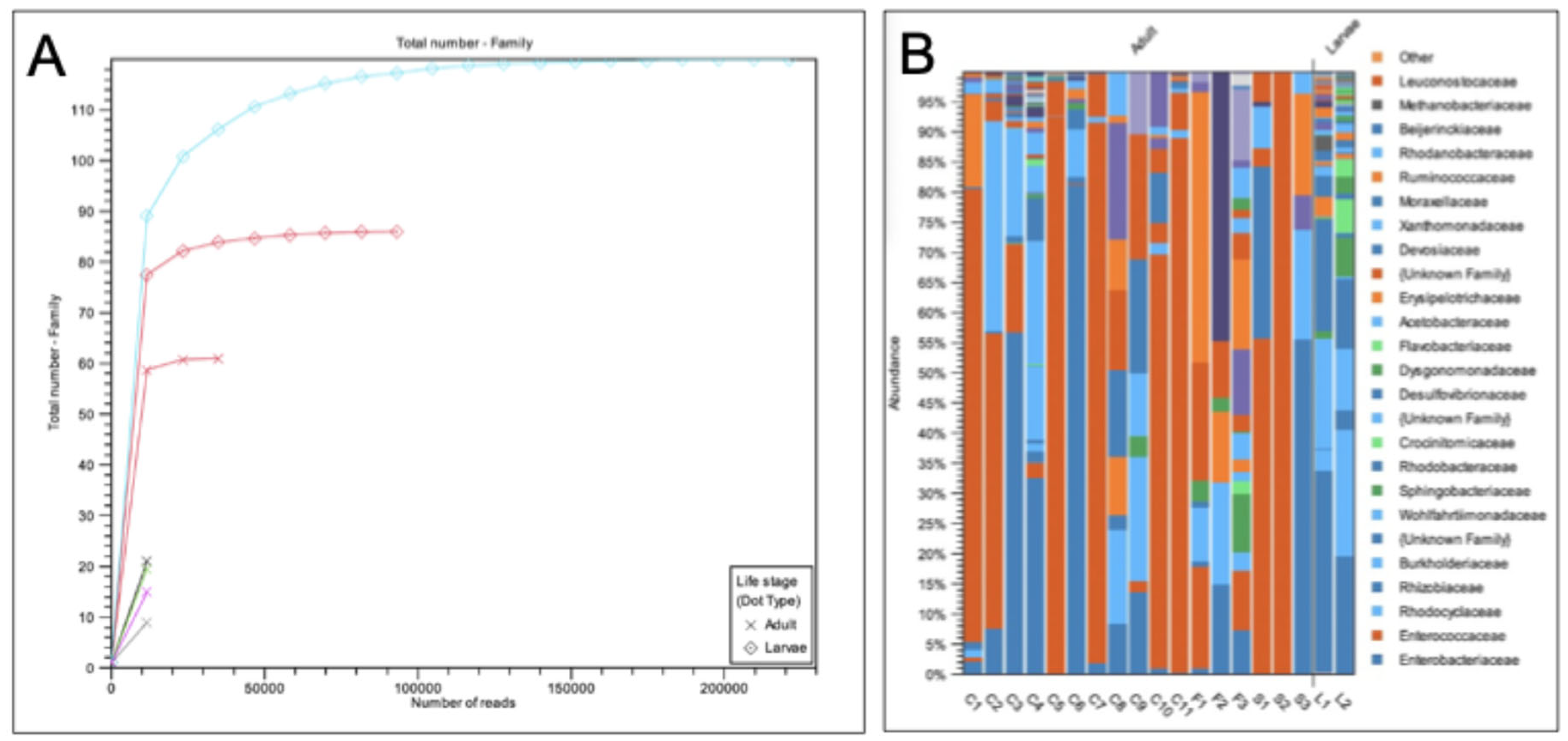

| Site | Female | Male | Total | Sex Ratio (F:M) |
|---|---|---|---|---|
| Soltis Center | 7 | 0 | 7 | 1.000 |
| Farm in San Juan | 6 | 0 | 6 | 1.000 |
| Finca Luna Nueva | 38 | 1 | 39 | 0.974 |
| Total | 51 | 1 | 52 | 0.980 |
| Temp (C) | % RH | UV | Abs Humidity | Dew Point | |
|---|---|---|---|---|---|
| Absolute Min | 22.7 | 66.3 | 100 | 11.4 | 16 |
| Absolute Max | 38.8 | 91.6 | 8000 | 41.5 | 37 |
| Range of weekly averages | 31.4 - 32.4 | 76.5 - 82.3 | 2700 - 3200 | 22.4 - 25.6 | 27 - 29 |
| Counts of BSF | Temp | Humidity | UV | |
| Counts of BSF | 1 | |||
| Temp | 0.28 | 1 | ||
| Humidity | -0.16 | -0.44 | 1 | |
| UV | 0.29 | 0.559 | 0.01 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



