Submitted:
08 December 2025
Posted:
09 December 2025
You are already at the latest version
Abstract
Humans and other terrestrial vertebrates contain two estrogen receptors (ERs), ERa and ERb. Among cartilaginous fish (sharks, rays, skates), which are jawless vertebrates that evolved about 525 million years ago, only activation by steroids of ERb orthologs has been characterized. To remedy this gap in understanding estrogen signaling in jawless vertebrates, we studied estrogen activation of orthologs of human ERa and ERb from elephant shark (Callorhynchus milii). Unexpectedly, we found that C. milii contained three estrogen-responsive ERa genes: ERa1 (596 amino acids), ERa2 (600 amino acids), and ERa3 (599 amino acids) with strong sequence similarity to each other. We also found an estrogen-unresponsive gene, ERa4 (561 amino acids), with a 39 amino acid deletion in the DNA-binding domain. An estrogen-responsive ERb ortholog (580 amino acids) also was present in C. milii. The three active C. milii ERas have a similar length to human ERa (595 amino acids); however, C. milii ERb is longer than human ERb (530 amino acids). We determined the half-maximal response (EC50) and fold-activation to estradiol (E2), estrone (E1), and estriol (E3) of C. milii ERa1, ERa2, ERa3, and ERb. Among these estrogens, E2 had the lowest EC50 for all four ERs. Fold-activation by E2 and E3 was similar for ERa1, ERa2, ERa3, and ERb. Overall, estrogen activation of C. milii ERa and ERb was similar to that for human ERa and ERb, indicating substantial conservation of the vertebrate ER in the 525 million years since the divergence of cartilaginous fish and humans from a common ancestor.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Chemical Reagents
2.3. Cloning of Estrogen Receptors
2.4. Database and Sequence Analysis
2.5. Reporter Gene Assay
3. Results
3.1. Cloning and Sequence Analysis of cDNAs for four Elephant Shark ERa Genes and one ERb Gene
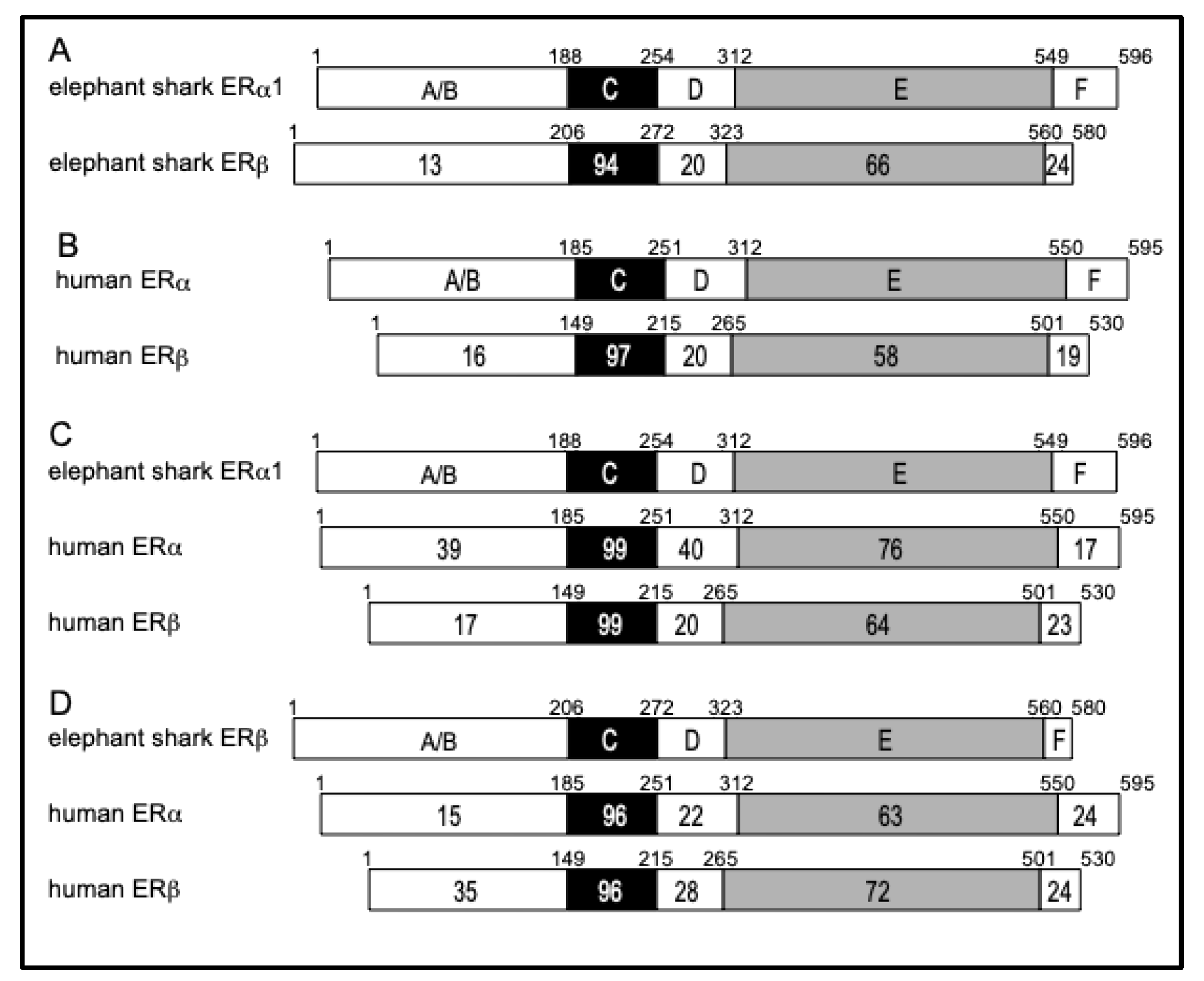
3.2. Comparison of Functional Domains on Human ERa and ERb with Functional Domains in Elephant Shark ERa and ERb
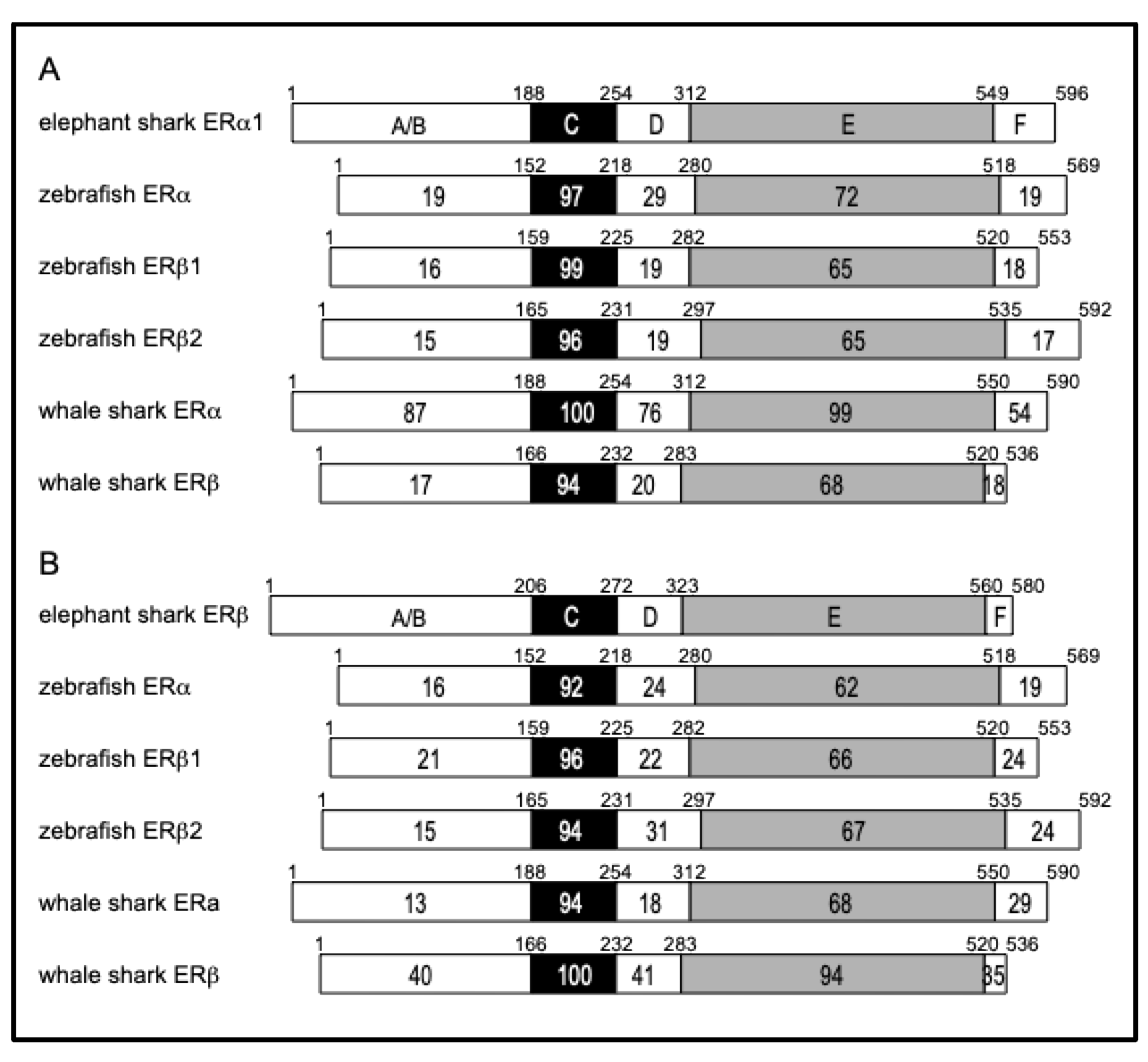
3.3. Comparison of Functional Domains on Elephant Shark ERa and ERb with Functional Domains in Zebrafish, and Whale Shark
3.4. Transcriptional Activities of Elephant Shark ERa and ERb.
4. Discussion
Author Contribution
Funding
Declaration of competing interest
Data availability
Acknowledgment
References
- Bondesson, M.; Hao, R.; Lin, C.-Y.; Williams, C.; Gustafsson, J. Estrogen receptor signaling during vertebrate development. Biochim. et Biophys. Acta (BBA) - Gene Regul. Mech. 2015, 1849, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Dahlman-Wright, K. The gene regulatory networks controlled by estrogens. Mol. Cell. Endocrinol. 2011, 334, 83–90. [Google Scholar] [CrossRef]
- DeRoo, B.J.; Korach, K.S. Estrogen receptors and human disease. J. Clin. Investig. 2006, 116, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, K.J.; Hewitt, S.C.; Arao, Y.; Korach, K.S. Estrogen Hormone Biology. Curr. Top. Dev. Biol. 2017, 125, 109–146. [Google Scholar]
- Paterni, I.; Granchi, C.; Katzenellenbogen, J.A.; Minutolo, F. Estrogen receptors alpha (ERalpha) and beta (ERbeta): subtype-selective ligands and clinical potential. Steroids 2014, 90, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Heldring, N.; Pawson, T.; McDonnell, D.; Treuter, E.; Gustafsson, J.; Pike, A.C. Structural Insights into Corepressor Recognition by Antagonist-bound Estrogen Receptors. J. Biol. Chem. 2007, 282, 10449–10455. [Google Scholar] [CrossRef]
- Dahlman-Wright, K.; Cavailles, V.; Fuqua, S.A.; Jordan, V.C.; Katzenellenbogen, J.A.; Korach, K.S.; Maggi, A.; Muramatsu, M.; Parker, M.G.; Gustafsson, J. International Union of Pharmacology. LXIV. Estrogen Receptors. Pharmacol. Rev. 2006, 58, 773–781. [Google Scholar] [CrossRef]
- Markov, G.V.; Tavares, R.; Dauphin-Villemant, C.; Demeneix, B.A.; Baker, M.E.; Laudet, V. Independent elaboration of steroid hormone signaling pathways in metazoans. Proc. Natl. Acad. Sci. 2009, 106, 11913–11918. [Google Scholar] [CrossRef]
- Baker, M.E. Origin and diversification of steroids: Co-evolution of enzymes and nuclear receptors. Mol. Cell. Endocrinol. 2011, 334, 14–20. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Enmark, E.; Pelto-Huikko, M.; Nilsson, S.; Gustafsson, J.A. Cloning of a novel receptor expressed in rat prostate and ovary. Proc. Natl. Acad. Sci. USA 1996, 93, 5925–5930. [Google Scholar] [CrossRef]
- Bridgham, J.T.; Brown, J.E.; Rodríguez-Marí, A.; Catchen, J.M.; Thornton, J.W. Evolution of a New Function by Degenerative Mutation in Cephalochordate Steroid Receptors. PLOS Genet. 2008, 4, e1000191. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.E.; Nelson, D.R.; Studer, R.A. Origin of the response to adrenal and sex steroids: Roles of promiscuity and co-evolution of enzymes and steroid receptors. J. Steroid Biochem. Mol. Biol. 2015, 151, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Escande, A.; Servant, N.; Rabenoelina, F.; Auzou, G.; Kloosterboer, H.; Cavaillès, V.; Balaguer, P.; Maudelonde, T. Regulation of activities of steroid hormone receptors by tibolone and its primary metabolites. J. Steroid Biochem. Mol. Biol. 2009, 116, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, B.; Lee, A.P.; Ravi, V.; Maurya, A.K.; Lian, M.M.; Swann, J.B.; Ohta, Y.; Flajnik, M.F.; Sutoh, Y.; Kasahara, M.; et al. Elephant shark genome provides unique insights into gnathostome evolution. Nature 2014, 505, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Filowitz, G.L.; Rajakumar, R.; O'Shaughnessy, K.L.; Cohn, M.J. Cartilaginous Fishes Provide Insights into the Origin, Diversification, and Sexually Dimorphic Expression of Vertebrate Estrogen Receptor Genes. Mol. Biol. Evol. 2018, 35, 2695–2701. [Google Scholar] [CrossRef]
- Katsu, Y.; Kohno, S.; Narita, H.; Urushitani, H.; Yamane, K.; Hara, A.; Clauss, T.M.; Walsh, M.T.; Miyagawa, S.; Guillette, L.J.; et al. Cloning and functional characterization of Chondrichthyes, cloudy catshark, Scyliorhinus torazame and whale shark, Rhincodon typus estrogen receptors. Gen. Comp. Endocrinol. 2010, 168, 496–504. [Google Scholar] [CrossRef]
- Inoue, J.G.; Miya, M.; Lam, K.; Tay, B.-H.; Danks, J.A.; Bell, J.; I Walker, T.I.; Venkatesh, B. Evolutionary Origin and Phylogeny of the Modern Holocephalans (Chondrichthyes: Chimaeriformes): A Mitogenomic Perspective. Mol. Biol. Evol. 2010, 27, 2576–2586. [Google Scholar] [CrossRef]
- Yu, W.-P.; Rajasegaran, V.; Yew, K.; Loh, W.-L.; Tay, B.-H.; Amemiya, C.T.; Brenner, S.; Venkatesh, B. Elephant shark sequence reveals unique insights into the evolutionary history of vertebrate genes: A comparative analysis of the protocadherin cluster. Proc. Natl. Acad. Sci. 2008, 105, 3819–3824. [Google Scholar] [CrossRef]
- Thornton, J.W. Evolution of vertebrate steroid receptors from an ancestral estrogen receptor by ligand exploitation and serial genome expansions. Proc. Natl. Acad. Sci. 2001, 98, 5671–5676. [Google Scholar] [CrossRef]
- Baker, M.E. Steroid receptors and vertebrate evolution. Mol. Cell. Endocrinol. 2019, 496, 110526. [Google Scholar] [CrossRef]
- Kakumura, K.; Watanabe, S.; Bell, J.D.; Donald, J.A.; Toop, T.; Kaneko, T.; Hyodo, S. Multiple urea transporter proteins in the kidney of holocephalan elephant fish (Callorhinchus milii). Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol. 2009, 154, 239–247. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Cotnoir-White, D.; Laperrière, D.; Mader, S. Evolution of the repertoire of nuclear receptor binding sites in genomes. Mol. Cell. Endocrinol. 2011, 334, 76–82. [Google Scholar] [CrossRef]
- Paris, M.; Pettersson, K.; Schubert, M.; Bertrand, S.; Pongratz, I.; Escriva, H.; Laudet, V. An amphioxus orthologue of the estrogen receptor that does not bind estradiol: Insights into estrogen receptor evolution. BMC Evol. Biol. 2008, 8, 219–219. [Google Scholar] [CrossRef] [PubMed]
- Katsu, Y.; Kubokawa, K.; Urushitani, H.; Iguchi, T. Estrogen-Dependent Transactivation of Amphioxus Steroid Hormone Receptor via Both Estrogen and Androgen Response Elements. Endocrinology 2010, 151, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Awruch, C. Reproductive endocrinology in chondrichthyans: The present and the future. Gen. Comp. Endocrinol. 2013, 192, 60–70. [Google Scholar] [CrossRef]
- Green, S.; Walter, P.; Kumar, V.; Krust, A.; Bornert, J.-M.; Argos, P.; Chambon, P. Human oestrogen receptor cDNA: sequence, expression and homology to v-erb-A. Nature 1986, 320, 134–139. [Google Scholar] [CrossRef]
- Matthews, J.; Gustafsson, J.A. Estrogen Signaling: A Subtle Balance Between ER alpha and ER beta. Mol. Interv. 2003, 3, 281–292. [Google Scholar] [CrossRef]
- Katzenellenbogen, B.S.; A Katzenellenbogen, J. Estrogen receptor transcription and transactivation Estrogen receptor alpha and estrogen receptor beta: regulation by selective estrogen receptor modulators and importance in breast cancer. Breast Cancer Res. 2000, 2, 335–44. [Google Scholar] [CrossRef]
- Katsu, Y.; Lange, A.; Urushitani, H.; Ichikawa, R.; Paull, G.C.; Cahill, L.L.; Jobling, S.; Tyler, C.R.; Iguchi, T. Functional Associations between Two Estrogen Receptors, Environmental Estrogens, and Sexual Disruption in the Roach (Rutilus rutilus). Environ. Sci. Technol. 2007, 41, 3368–3374. [Google Scholar] [CrossRef]
- Katsu, Y.; Kohno, S.; Hyodo, S.; Ijiri, S.; Adachi, S.; Hara, A.; Guillette, L.J.; Iguchi, T. Molecular Cloning, Characterization, and Evolutionary Analysis of Estrogen Receptors from Phylogenetically Ancient Fish. Endocrinology 2008, 149, 6300–6310. [Google Scholar] [CrossRef]
- Katsu, Y.; Taniguchi, E.; Urushitani, H.; Miyagawa, S.; Takase, M.; Kubokawa, K.; Tooi, O.; Oka, T.; Santo, N.; Myburgh, J.; et al. Molecular cloning and characterization of ligand- and species-specificity of amphibian estrogen receptors. Gen. Comp. Endocrinol. 2010, 168, 220–230. [Google Scholar] [CrossRef]
- Lange, A.; Katsu, Y.; Miyagawa, S.; Ogino, Y.; Urushitani, H.; Kobayashi, T.; Hirai, T.; Shears, J.A.; Nagae, M.; Yamamoto, J.; et al. Comparative responsiveness to natural and synthetic estrogens of fish species commonly used in the laboratory and field monitoring. Aquat. Toxicol. 2012, 109, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Yatsu, R.; Katsu, Y.; Kohno, S.; Mizutani, T.; Ogino, Y.; Ohta, Y.; Myburgh, J.; van Wyk, J.H.; Guillette, L.J.; Miyagawa, S.; et al. Characterization of evolutionary trend in squamate estrogen receptor sensitivity. Gen. Comp. Endocrinol. 2016, 238, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, H. Pharmacology of estrogens and progestogens: influence of different routes of administration. Climacteric 2005, 8, 3–63. [Google Scholar] [CrossRef] [PubMed]
- Stanczyk, F.Z. Metabolism of endogenous and exogenous estrogens in women. J. Steroid Biochem. Mol. Biol. 2024, 242, 106539. [Google Scholar] [CrossRef]

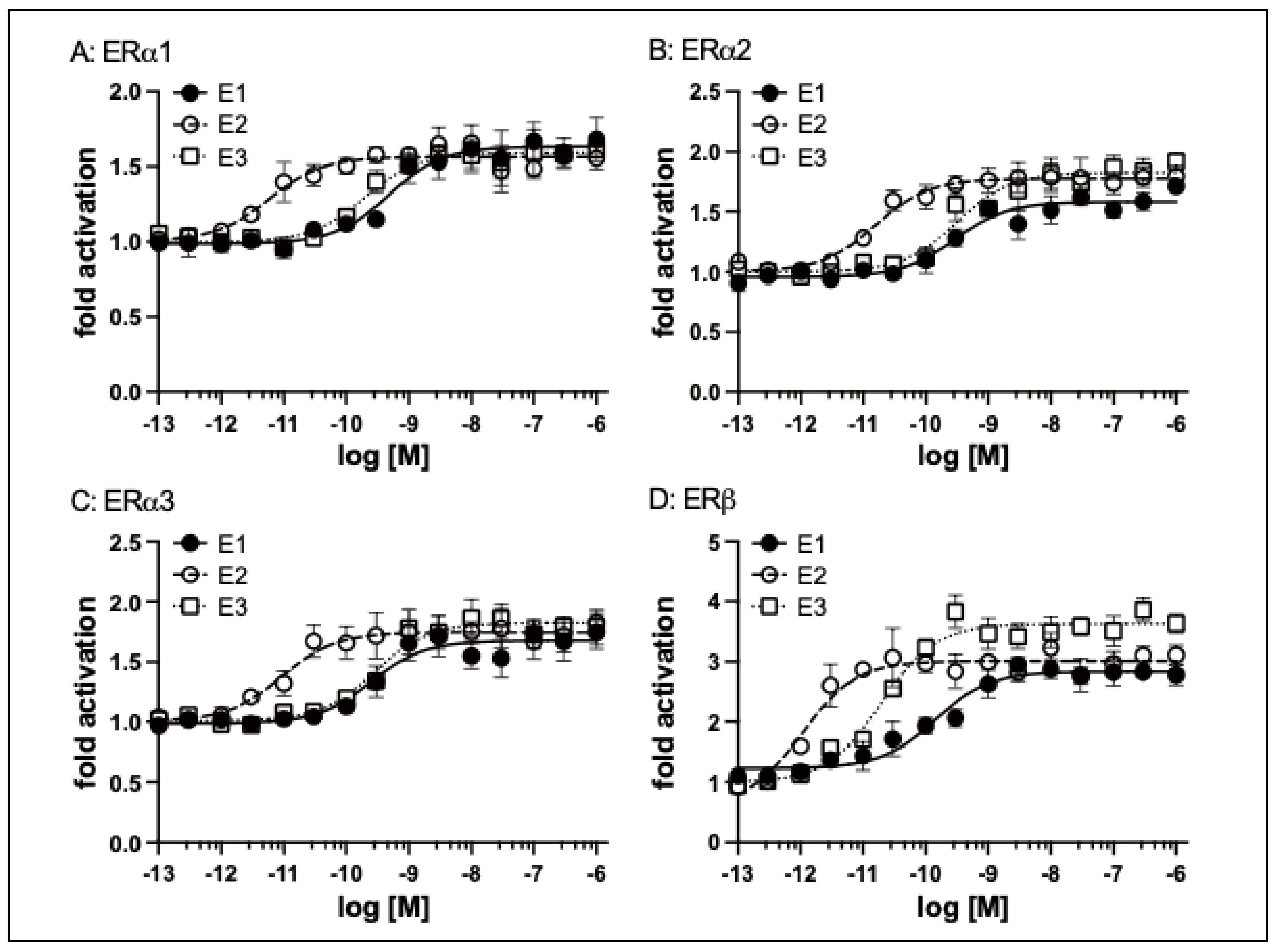

| E1 | E2 | E3 | ||
| ERa1 | EC50 (nM) | 0.49 | 0.0059 | 0.2 |
| Fold-Activation (± SEM) * |
1.62 (± 0.16) |
1.66 (± 0.04) |
1.57 (± 0.09) |
|
| Ratio** | 0.98 | 1.00 | 0.95 | |
| ERa2 | EC50 (nM) | 0.29 | 0.016 | 0.31 |
| Fold-Activation (± SEM) * |
1.51 (± 0.12) |
1.79 (± 0.12) |
1.82 (± 0.13) |
|
| Ratio** | 0.84 | 1.00 | 1.02 | |
| ERa3 | EC50 (nM) | 0.25 | 0.0093 | 0.29 |
| Fold-Activation (± SEM) * |
1.55 (± 0.11) |
1.75 (± 0.08) |
1.87 (± 0.15) |
|
| Ratio** | 0.89 | 1.00 | 1.07 | |
| ERb | EC50 (nM) | 0.14 | 0.001 | 0.02 |
| Fold-Activation (± SEM) * |
2.87 (± 0.16) |
3.23 (± 0.26) |
3.49 (± 0.26) |
|
| Ratio** | 0.89 | 1.00 | 1.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



