
Submitted:
05 December 2025
Posted:
08 December 2025
You are already at the latest version
Abstract
From clinical and experimental studies, it appears that supplementation with specific amino acids may be relevant to correct for amino acid deficiency in case of insufficient supply from dietary proteins in regards with the amounts needed for optimal metabolism and physiological functions. Clinical and experimental arguments suggest that amino acid supplementation may be indicated in specific situations under a specific nutritional context. However, it is essential not to overdose with excessive quantities of amino acids in supplements thus beyond the upper levels of safe intake (ULSI). In this narrative review, we recapitulate the protein and amino acid requirements for the general population and for subgroups of population including vulnerable subgroups, and these requirements are compared to the usual consumption. Typical examples of clinical trials showing benefits from amino acid supplementation in different physiological and pathophysiological contexts are presented together with results obtained from experimental studies in that field. Finally, parameters such as the no-observed-adverse-effect-level (NOAEL) values used to determine the ULSI for amino acid supplementation are defined, and values determined in clinical trials are given and discussed.
Keywords:
amino acid supplementation
; human nutrition
; tolerable upper intake
; no -observed -adverse-effect level
; amino acid requirement
; amino acid usual consumption
1. Introduction
As recapitulated recently [1], the 20 common amino acids (and the uncommon amino acid selenocysteine) are well known to represent the building blocks for protein synthesis in the different tissues and organs in the human body. In addition, specific amino acids such as glycine, aspartate, and glutamine are precursors for the synthesis of the purine and pyrimidine rings of RNAs and DNA in cells, as well as precursors for the synthesis of the free energy donor adenosine triphosphate (ATP).
Amino acids are precursors for the synthesis of numerous bioactive peptides [2] as well as for the synthesis of metabolic regulators. N-acetylglutamate for instance is synthesized from the amino acid glutamate, and this compound is acting as an allosteric cofactor for carbamylphosphate synthetase I, an enzyme involved in the liver urea cycle [3]. Several neurotransmitters (such as serotonin, dopamine, adrenaline, noradrenaline, and gamma aminobutyrate); hormones like melatonin and pigment like melanin require specific amino acids as precursors for their synthesis. Some amino acids, mainly alanine and glutamine, are precursors for the synthesis of glucose [4,5] and serine/methionine are involved in the synthesis of phospholipids. Specific amino acids can be used as energy substrate in different cell phenotypes within tissues and organs.
Amino acids are also used as precursors for numerous bioactive compounds with central metabolic and physiological functions. Several among the 20 common amino acids are involved in cell signaling in target tissues. The branched-chain amino acids leucine, isoleucine, and valine for instance are involved through specific signaling pathways in the regulation of protein, glucose, and lipid metabolism [6]. Some amino acids are precursors of compounds involved in central physiological functions. Arginine for instance is the precursor of nitric oxide (NO) which is involved in vascular muscle relaxation [7]. Some amino acids are required for nitrogenous waste disposal. Regarding this last point, the functioning of the urea cycle for example involves amino acids such as arginine and aspartate. Finally, an amino acid such as methionine is involved, through its conversion into S-adenosylmethionine, in DNA and histone methylation [8]. These methylation processes are an important component for the epigenetic regulation of gene expression in human cells [9].
The percentages of amino acids involved as precursors for the synthesis of the different amino acid-derived compounds are evidently much different according to the pathways of use considered. As we will see in the next paragraph, protein renewal in the body requires large amounts of amino acids. In contrast, as a role of thumb, the utilization of amino acids for compounds such as metabolic regulators, gaseous mediators (like nitric oxide), neurotransmitters, and oligopeptides (like glutathione) represents only a minor part of their total utilization in the body. However, as observed in animal models, in specific situations such as infection and intestinal inflammation, glutathione synthesis from the three amino acid precursors cysteine, glutamate, and glycine is markedly increased. In such situation, cysteine for the synthesis of glutathione may represent a significant part of the whole-body utilization of this amino acid [10,11]. The main metabolic and physiological functions of amino acids in humans are briefly recapitulated in Figure 1.
Adequate amounts of amino acids, mostly from dietary proteins and to a minor extent from amino acids in the diet in their free form, must be regularly provided. Also, amino acids are provided from endogenous synthesis and protein recycling in the body. Amino acid availability from these different sources must meet the metabolic and physiological needs of the different tissues. Amino acid utilization is also required for excretion of nitrogenous compounds within the biological fluids (urine, feces etc.) (Figure 2). A large part of the utilization of amino acids is devoted to the renewal of the body proteins. Indeed, in healthy adults weighing 70 kg, approximately 300-400 g body proteins are renewed every day among the 10-12 kg mass of body proteins [12].
Among the 20 amino acids present in dietary proteins, 9 of them (isoleucine, leucine, valine, methionine, tryptophan, threonine, phenylalanine, histidine, and lysine) must be provided by the diet because the body is not able to synthesize them (or not in sufficient quantities) to meet the body’s requirement [13]. These amino acids are then considered as indispensable. However, they are not all equivalent since for some of them, like lysine and threonine, no metabolic capacities for their synthesis have been identified [14], while for some other indispensable amino acids, minor capacities of synthesis can be detected in some tissues. For instance, homocysteine, which is found in low amounts in some dietary products [15], can represent a precursor for the synthesis of methionine through vitamin B12-dependent pathways [16]. Different criteria are used to estimate the quality of dietary proteins originating either from animal and plant sources, or from microorganisms, and these criteria include the digestibility of the alimentary proteins as well as their content in indispensable amino acids [17].
The other 11 amino acids (thus alanine, arginine, asparagine, aspartate, cysteine, glutamate, glutamine, glycine, proline, serine, and tyrosine), by difference, are considered as not indispensable (or dispensable), and for these ones, anabolic pathways from other amino acids or non-amino acid precursors have been characterized and found to allow significant endogenous production of these amino acids [1].
Among these 11 amino acids, another subclassification has been made gathering 6 amino acids considered as conditionally indispensable. These amino acids, although being synthesized in significant amounts within the body, are not sufficiently produced in different physiological and pathophysiological situations to cover the metabolic and physiological needs [18]. These conditionally indispensable amino acids are arginine, cysteine, glutamine, glycine, proline and tyrosine. Regarding arginine, this amino acid becomes for instance indispensable in severely burned patients [19], while cysteine supplementation appears clinically relevant in subgroups of critically ill patients. In these patients, such supplementation can increase the endogenous synthesis of the cysteine-derived tripeptide glutathione [20]. Glutathione is well known to limit the severity of chronic inflammation [20] in different clinical contexts [21,22]. Regarding glutamine, this amino acid has been shown to become indispensable in patients undergoing severe catabolic states like the one encountered in sepsis [23] while glycine may become indispensable in the late stages of pregnancy [24].
In order to evaluate the relevance of supplementation with one or several amino acids in a given physiological/pathophysiological situation, it is important to cross various parameters including protein and amino acid requirement, usual consumption, clinical endpoints determined in clinical intervention studies, together with results of experimental studies which can provide a better understanding of the mechanisms at the basis of the observations made.
In this narrative review, we present different situations where supplementation with one specific or with mixtures of different amino acids showed beneficial effects in clinical trials and/or in experimental studies. We then present what is known from clinical studies about the upper limits for safe utilization of amino acids in supplements.
2. Protein and Amino Acid Requirement Along the Life Cycle and Comparison with Usual Consumption in Different Geographical Areas
The dietary protein requirement in adult humans can be defined as the amounts of dietary proteins that will maintain the body protein mass in individuals with moderate level of physical activity. In children and pregnant or lactating women, the amounts of dietary proteins that will ensure the maintenance of body protein mass must consider the additional amounts of proteins that will ensure adequate tissue deposition in children and pregnant women, or secretion of milk with adequate composition in lactating women, respectively. From the notion of protein requirement, the recommended protein nutritional intake corresponds to the value which cover the requirements of most of the population, on a statistical basis of 97.5% of the individuals among the populations studied. This value is close to the average requirements to which two standard deviations are added [25].
2.1. Protein Requirement in Different Situations
The median average requirement for proteins in the general population of healthy adults has been estimated to be 0.65 g proteins per kg body weight per day, while the recommended dietary intake is 0.83 g proteins per kg body weight per day, thus representing 58.1 g of good quality dietary proteins for an individual weighing 70 kg [26]. These values can be compared with the usual consumption in different geographical areas. In Europe and USA, the average dietary protein consumption is approximately 85 g per day in adults [27,28], thus largely above the recommended intake. However, this average value determined in Western countries must not mask subgroups of population for who the usual consumption can be much lower. To take an example, in vegans, the total protein intake is lower when compared to intake in non-vegan groups [29] and approximately one third of vegans are found below the adequate protein intake [30].
Some other countries are below the average protein consumption found in the Western world. In Thailand for instance, the average consumption of proteins is 63.4 g per day [31], thus close to the recommended intake. In some countries, the usual protein consumption is largely below the recommended intake. For instance, in Southern Ethiopia, the prevalence of inadequate dietary protein intake is found in 94% of the female population [32].
In infants between birth and 3 years old, the estimation of the protein requirement is within 0.76-1.80 g proteins/kg/day, with a recommended intake between 0.94 and 2.60 g proteins/kg/day [33]. In children between 4 and 10 years, the protein requirement is 0.70 g proteins/kg/day, and the recommended intake is 0.88 g proteins/kg/day. In adolescents (11-18 years), the protein requirement and the recommended intake is between 0.63 and 0.72 g proteins/kg/day, and between 0.78 and 0.90 g proteins/kg/day, respectively. These estimations have been also made in pregnant women, averaging between 0.65 and 0.73 g proteins/kg/day and between 0.82 and 1.00 g proteins/kg/day for the requirement and the recommended intake, respectively [33]. However, it is important to mention here that protein requirements in these human subgroups are originating from only a limited number of studies [34].
The requirement and recommendation for the elderly are generally higher than for young adults, and in several recent publications, the recommended intake has been evaluated to range between 1.0 and 1.6 g proteins/kg/day, thus representing between 70 and 105 g of good quality proteins [35,36,37,38]. Despite this recommendation to increase the intake of dietary proteins in the elderly population, protein-energy malnutrition is detected in approximately 1% of this elderly population in Northern Europe, 11% in Southern Europe, 15% in Central and West Africa, 18% in Australia/New Zealand while protein-energy malnutrition represents as much as 25% of the elderly population in South-East Asia [39].
In specific situations such as athletes practising endurance or strength sports, the recommended intake of dietary proteins is higher than the one in individuals with moderate physical activity, being situated between 1.3-1.8 g proteins/kg/day (91-126 g proteins per day for individuals weighing 70 kg) [40].
2.2. Indispensable Amino Acid Requirement in Adults and Comparison with Usual Consumption in Western Countries
The requirement and usual consumption (UC) of indispensable amino acids have been reported in several reports [33,41,42] and in one recent review [43]. Consumption data for indispensable amino acids in food and supplements are expressed in mg per kg body weight in Table 1 to allow comparison with the requirements. As indicated in this table, the requirements for indispensable amino acids in adults are somewhat different for some amino acids when comparing the values given in the three reports. However, it is worth noting that for methionine and phenylalanine, the presented requirement values do not relate exactly to the same parameters.
The FNB/IOM data for the usual consumption (UC) referred to the USA and Canadian population for all life stages and gender groups [41]. In the same table are presented the UC of indispensable amino acids by European adults from the EPIC cohort (NMCD values) [44,45]. These mean values of UC are similar when comparing the two populations, being higher than the requirement for all the 9 indispensable amino acids.
3. Amino Acid Supplementation in Specific Situations: Lessons from Clinical Trials and Experimental Studies
Beneficial effects of amino acid supplementation have been tested in physiological and pathophysiological situations of presumed insufficient dietary protein intake in regards with requirements. This may be for instance the case in the elderly population, which is recognized at risk for sarcopenia, and thus which may experiment progressive loss of muscle mass and strength [46].
3.1. Leucine Supplementation in the Sarcopenic Elderly
The rationale for testing supplementation with specific amino acids together with protein of good quality in the sarcopenic elderly people is rather robust for several reasons. Firstly, in a context of increased requirement for proteins in the elderly, muscle protein synthesis is reduced in aging adults [47], and such reduction is paralleled by reduction of protein synthesis within human muscle mitochondria [48]. Such reduction occurs in conjunction with reduced mitochondrial capacity for ATP production [49]. Secondly, protein synthesis in muscles of elderly people is less responsive to amino acids than in young subjects, a situation defined as part of anabolic resistance [50]. Positive effects of higher versus lower levels of proteins in foods on the preservation of lean mass have been observed in older adults [51,52].
The branched-chain amino acids (BCAAs) leucine, isoleucine and valine have raised considerable interest because of their different metabolic and physiological functions. BCAAs are used for protein synthesis in mammalian cells, and transaminated in the presence of α-ketoglutarate, allowing the production of glutamate and the corresponding α-ketoacids [53,54]. Each α-ketoacids can then undergo several steps of conversion resulting in the synthesis of acetyl-CoA and succinyl-CoA. These compounds can then enter the tricarboxylic acid cycle allowing the synthesis of reduced equivalents. These equivalents are finally used in the mitochondrial respiratory chain allowing ATP synthesis. BCAAs are precursors not only for the dispensable amino acid glutamate, but also for alanine and glutamine. BCAAs are involved in the regulation of metabolic pathways engaged in glucose and lipid metabolism, and in protein synthesis in skeletal muscles. Regarding more specifically leucine, this BCAA has been shown to act on muscle protein synthesis through the stimulation of mTORC1/P70S6K signaling pathway, which represents the major myocellular signaling pathway [55].
Leucine (52 mg/kg body weight) has been found able to increase protein synthesis in skeletal muscles when measured over the 5-hour period of feeding [56]. As expected, leucine supplementation is paralleled by increased circulating concentration of leucine. Generally, large increases in peripheral indispensable amino acid concentrations (notably leucine) are necessary to drive significant increase in muscle and whole-body protein synthesis [57]. Then, leucine has been tested for its effects on muscle mass and strength in the elderly in controlled clinical trials with and without physical exercise. The results of these trials are rather heterogeneous due to different volunteer characteristics and experimental design (doses of leucine given orally and duration of supplementation, quantity and quality of proteins given together with the amino acid supplements, presence of other amino acids and vitamins in the supplements, chosen endpoints, etc.).
In the double blind, randomized controlled clinical study by Rondanelli and collaborators, supplements containing 10.9 g indispensable amino acids including 4 g of leucine together with 22 g of whey protein and vitamin D, when associated with physical activity for 12 weeks, increased skeletal muscle mass and handgrip strength in sarcopenic elderly subjects [58]. In other randomized controlled studies, supplement containing 2.8 g leucine, 20 g whey protein, and vitamin D given for 4-8 weeks increased muscle mass and improved physical performance in older adults [59], while 3g leucine, 20 g whey proteins, and vitamin D in supplements given for 13 weeks to volunteers increased muscle mass in older adults [60].
From these studies, several comments can be made. Firstly, considering that 1 g whey protein bring 81 mg leucine [61], the total amount of leucine brought by whey protein and supplements in these three studies are between 4 and 6 g. We can compare these values with the approximative amount of leucine that is contained within 91 g of proteins, thus the mean recommended intake for old adults weighing 70 kg. By considering that the usual consumption of leucine in North America is 5.8 g/day from a diet containing 85 g proteins, 91 g proteins provide 6.2 g leucine, a value close to the upper level of leucine provided in the three clinical trials mentioned above. Secondly, it must be kept in mind that optimal protein synthesis in muscles required sufficient supply of all amino acids, the ones that are provided by whey proteins and amino acids in their free form in supplements. Thirdly, the requirement for leucine appears higher in older than young adults [62]. This suggests that in seniors, leucine could represent a limiting amino acid for protein synthesis, for utilization in ATP production, and even maybe for signal functions in skeletal muscles when the supply of protein is largely below the recommendation for older adults.
It is of major importance to consider both the age-related decline in appetite observed in subpopulations among the elderly and the satiating properties of proteins [63,64,65]. Studies in older adults suggest that the supplements with indispensable amino acids are non-satiating, or at least not as much satiating as high protein diet, in both acute and medium-term experiments [66,67,68,69]. Then, supplements with indispensable amino acids with an abundant leucine content represent an interesting strategy to enhance the anabolic properties of a meal containing a suboptimal supply of proteins in older adults with little appetite.
The exhaustive mention of all the randomized controlled studies dealing with leucine supplementation in the elderly in different experimental contexts is outside the scope of this review, and readers are referred to excellent reviews on that topic [70,71,72,73,74,75]. From these reviews, several important conclusions can be drawn. Leucine supplementation exerts beneficial effects on muscle protein fractional synthetic rate, muscle mass, and lean body mass in older adults prone to sarcopenia or with diagnosed sarcopenia. Regular physical exercise together with supplements with indispensable amino acids (with high leucine content) and/or whey proteins together with vitamin D improve the maintenance or gains of skeletal muscle mass and total lean mass in sarcopenic older adults. Importantly, the effects of physical exercise and dietary supplements appear synergistic. Since concentrations of some amino acids, including not only BCAAs but also dispensable amino acids such as arginine, glutamine, glycine, and serine, are diminished in the plasma of older adults, further clinical trials are required to test supplements with combination of individual amino acids (and some of their metabolites) for their effects in limiting sarcopenia in older adults prone to sarcopenia.
3.2. Amino Acid Supplementation in Athletes
Branched-chain amino acids (BCAAs) are among the most popular supplements, marketed under the promise that they enhance muscular adaptations to physical exercise. Despite their prevalent consumption among athletes and the general sportsmen and sportswomen, large controversies remain in this sports nutrition field. Although it appears clearly that, in the context of sufficient protein-energy intake, supplementation with BCAAs has little or no efficacy for enhancing gains of muscle mass and strength-based physical performance during resistance training, there is some clinical evidence that muscle loss may be minimized by BCAAs during protein-energy restriction [76,77].
Among BCAAs, leucine given by intravenous infusion in healthy volunteers has been shown to decrease protein degradation [78]. BCAA supplementation (100 mg BCAAs/kg body weight) increases protein synthesis in muscles during the recovery phase (2 hours) after 1 hour of endurance sports [79]. In volunteers, BCAA infusion increases the phosphorylation of proteins involved in the translation of mRNA corresponding to proteins involved in the synthesis of muscles proteins [80]. In a clinical trial, oral supplementation with arginine and the nine indispensable amino acids (including leucine which represent 20% in mass of the amino acid mixture (40 g)) after heavy resistance training, results in the transition from net muscle protein degradation to net protein synthesis when compared to placebo [81].
Experimental studies in animal model have revealed mechanisms involved in the biological effects of BCAAs on skeletal muscles. Notably, in the piglet model, leucine has been shown to upregulate the mammalian target of rapamycin (mTOR) complex (mTORC)1, an element of the signaling cascade involved in muscle protein synthesis [55,82]. In the rat model, it has been demonstrated that physical exercise promoted BCAA oxidation in skeletal muscles [83]. The mechanisms at the basis of such effects involve the activation of the branched-chain α-ketoacid dehydrogenase (BCKDH) complex which catalyses the second step reaction in the BCAA catabolic pathway [84].
The role of creatine for the metabolism and physiology of skeletal muscles in physical exercise is out of the scope of the present review because this compound is not belonging to the 21 amino acids used for protein synthesis, but is synthesized from arginine, glycine and methionine [85]. The interested readers are referred to recent reviews on that topic [86,87].
3.3. Amino Acid Supplementation for Intestinal Mucosa Healing
Inflammatory bowel diseases (IBD) are characterized by mucosal lesions in the intestine which are associated with exacerbated immune functions of still unclear etiology [88]. IBD, mainly Crohn’s disease (CD) and ulcerative colitis (UC), are identified by chronic inflammation of the mucosa with alternating relapse and remission periods. In CD, remission episode offers the possibility of mucosal healing, allowing in some cases the total disappearance of all mucosal ulcerations [89]. However, in clinical practice, this endpoint is not easy to achieve. For patients with UC, mucosal healing is described as the absence of friability, blood, erosions, and ulcers in all segments of the gut mucosa [90,91]. An advanced mucosal healing is associated with sustained remission, improved clinical outcomes, and reduced rates of hospitalization, thus explaining why mucosal healing represents a therapeutical goal for gastroenterologists in charge of CD and UC patients [92,93,94,95].
Some clinical and experimental studies report on the requirement of dietary proteins in the process of intestinal mucosa healing and on the implication of specific amino acids used in supplements for accelerating this process [96]. Mucosal healing is a combination of different protein-energy dependent processes which allow to restore the continuity of the intestinal epithelium [94,97]. Indeed, poor nutritional status may characterize patients with IBD in active phase because of anorexia, intestinal malabsorption, increased intestinal losses, and increased catabolism [98]. Poor nutritional status is likely to affect the efficiency of mucosal healing, and protein-energy malnutrition together with deficiencies in minerals and vitamins have been reported in patients with IBD [99,101]. Overall, elemental formulas with amino acids do not appear to be more efficient for clinical remission in adults and children than polymeric formulas containing whole proteins [102,103,104,105].
Although the protein requirements for intestinal mucosal healing in CD and UC patients in remission have not be determined, there are some reasons, mostly from experimental studies, to presume that the daily protein requirements in IBD patients in remission may differ from those in healthy adults. The European Society for Clinical Nutrition and Metabolism recommends increasing the protein intake to 1.2-1.5 g/kg body weight/day in adult patients with active IBD, and to maintain an intake of 1g proteins/kg body weight/day during remission [106].
In the mouse model, mucosal healing after colitis induction with dextran sodium sulfate is paralleled by increased fractional protein synthesis rate in colon when compared with the situation observed in the animals before colitis induction [107]. Some experimental studies point to potentially beneficial effects of moderately increased dietary protein ingestion and of supplementation with specific amino acids for mucosal healing after an inflammatory episode. In a mouse model of chemically-induced colitis, a moderately high-protein diet (30% of energy supplied by casein and whey proteins) given to animals at different times after the time of maximal colitis intensity showed beneficial effects during the epithelial repair process within the colonic mucosa when compared with animals receiving a normoproteic diet (14% of energy as proteins) [108]. In sharp contrast, a frankly high-protein diet (53% of energy supplied by proteins) resulted in the worsening of the inflammation both in intensity and duration. Although the reasons that would explain the deleterious effects of high protein diet on the colonic mucosa of mice in situation of experimental colitis remains unknown, high protein diet consumption has been shown to be associated with changes of the amounts of bacterial metabolites within the colonic luminal fluid in a way that is globally deleterious for the colonic epithelium [109]. These data suggest that moderate increase of dietary protein intake facilitates mucosal healing in colon, but that excessive intake of dietary protein is deleterious. This indicates a tricky choice to set the correct level of proteins to bring in this experimental model.
Supplementation with threonine, serine, proline, and cysteine (15, 10, 15, and 7.2 g/kg diet, respectively) have been shown to increase mucin synthesis when given 8 days before experimental colitis induction in the rat model and 28 days thereafter [110]. Since an intact mucus layer is necessary for providing an “initial seal” after intestinal mucosa injury [111,112,113], these data are of major interest. Although the respective roles of each amino acids used in the mixture cannot be determined in such studies, it is worth noting that the indispensable amino acid threonine is abundant in mucins and optimal synthesis of mucins require a relatively high supply of threonine [114]. Supplementation with a mixture of glutamate, methionine, and threonine (0.57, 0.30, and 0.50 g/day, respectively) given in the mucosal healing phase after chemical induction of colitis in rats improves the colonic mucosal regeneration/reepithelialization after supplementation for 10 days [115]. Here again, the respective roles of each of the three amino acids in the mixture could not be delineated, but glutamate has been identified as a major energy substrate in colonocytes and as a precursor for the synthesis of the dispensable amino acids aspartate and alanine, as well as for the synthesis of the tripeptide glutathione [116]. This latter compound is involved in the maintenance of the intracellular redox status and in the control of the intracellular concentration of both the oxygen- and nitrogen reactive species [117,118]. These reactive species are produced at abnormal levels in inflammatory bowel diseases [119]. Regarding methionine, this indispensable amino acid is largely metabolized in the intestine and is a precursor of cysteine, another precursor for glutathione synthesis [120]. In a mice model with experimentally induced colitis, an elemental diet enriched with a mixture of amino acids (192 g amino acids per kg food) was capable to slow down colonic mucus degradation [121].
3.4. Amino Acid Supplementation During Weight Loss Programs
In a context of worldwide increased prevalence of obesity in children, adolescents, and adults [122,123], different strategies such as changes in the lifestyle [124], pharmacological treatments [125], and surgical procedures have been implemented [126,127]. Whatever the strategy used for significant weight loss, the loss of fat mass is paralleled by significant decrease in lean mass, especially regarding skeletal muscles, thus increasing the risk of sarcopenia [128,129,130,131]. The most efficient preservation of lean mass during weight loss periods requires notably the presence of adequate anabolic stimuli [132].
Regular physical exercise, especially resistance-type exercise training, and higher protein intake that the one recommended for the general adult population are both recommended for obese individuals who undertake weight-loss programs. In fact, high protein diets, which, as introduced above, are more satiating than carbohydrates and fats on a basis of an equal energy content, are effective in attenuating the loss of lean mass which parallel weight loss [133,134]. Physical exercise appears also effective for reducing the loss of lean mass in case of body weight reduction [135,136,137] and thus can act in conjunction with high protein diet.
Regarding supplementation with amino acids, effects of such supplementation on the maintenance of the lean body have been documented in the dietary context of lower protein intake when associated with physical exercise [138]. In obese adults recruited for a four-week randomized controlled trial, meal replacement containing 6 g proteins and enriched with 16g of a mixture of indispensable amino acids showed beneficial effects on skeletal muscle cross-sectional area when compared to isoenergetic standard meal replacement containing 16 g protein but no supplement [139]. Amino acid supplementation has been tested in a randomized controlled study with adult men characterized by severe obesity who undergo a 4-week intervention program. This program associated low-calorie balanced diet with physical activity program. In such situation, supplementation with a mixture of indispensable amino acids together with citrate, succinate, and malate was found able to increase muscle mass when compared with the control group receiving no supplements [140]. However, the respective role of the amino acids and of the tricarboxylic acid intermediates could not be delineated in this study.
3.5. Amino Acid Supplementation in Metabolic Syndrome
Metabolic syndrome refers to the co-occurrence of several heterogenous parameters including biochemical anomalies, physiological dysfunctions, and anthropometric characteristics. Although the list of these parameters may differ between scientists, they usually gather criteria such as dyslipidemia (hypertriglyceridemia, reduced high-density lipoprotein (HDL) cholesterolemia), hyperglycemia, insulin resistance, endothelial dysfunction, elevated arterial blood pressure, and abdominal obesity [141,142]. Other parameters such as microalbumineria and non-alcoholic fatty liver may be included as parts of parameters which define metabolic syndrome [143,144]. The occurrence of several among these parameters in each individual correlates with increased risks of cardiovascular adverse events and of type 2 diabetes [145,146].
From some clinical trials and experimental studies, there are some indications that supplementation with mixture of indispensable amino acids or individual amino acids such as arginine, leucine, glycine, histidine, and isoleucine can alleviate one or several criteria usually associated with metabolic syndrome.
In a randomized controlled clinical trial, arginine supplementation (8.3 g/day) added to hypocaloric diet for 21 days, when combined with exercise training in obese and insulin resistant type 2 diabetic patients, exerts additive effect on glucose metabolism and insulin sensitivity when compared to dietary restriction and exercise training alone [147]. In a double blind, randomized controlled study with type 2 diabetic patients, arginine in supplements (9 g/day) given for one month significantly improves peripheral and hepatic insulin sensitivity [148]. Furthermore, in a randomized controlled study, a single dose of arginine (8g) given to hypertensive patients was able to induce post-exercise lower systolic blood pressure [149]. In a double blind, randomized controlled clinical trial, long-term (18 months) oral arginine supplementation (6.4 g/day) improved insulin sensitivity in volunteers with impaired glucose tolerance [150]. Lastly, intravenous infusion of arginine in obese individuals improved insulin sensitivity and restored the insulin-mediated vasodilatation [151]. Some of the effects of arginine may be partly related to the role of this amino acid as precursor for nitric oxide (NO) synthesis by nitric oxide synthase activities, but this hypothesis must be confronted to new experiments. The rationale at the basis of this hypothesis is because among the numerous physiological functions of NO, generation of NO from arginine by the vascular endothelium appears essential for the regulation of blood flow and blood pressure [152,153].
Experiments with animal models reinforce the view that supplementation with arginine can improve some of the anomalies which are associated with metabolic syndrome. Indeed, in rats with experimentally induced metabolic syndrome, arginine supplementation given for 8 weeks further increases the beneficial effects of physical training on hypertension, adipose tissue mass, and hepatic steatosis [154]. Also, in a rodent model with metabolic syndrome, arginine supplementation (0.50% in drinking water) given for 4 weeks alleviates hypertension [155]. In addition, chronic arginine supplementation (5% in diet) given for 8 weeks in a rodent model of diet-induced metabolic syndrome improves glucose tolerance, decreased blood pressure, and decrease abdominal fat pads [156]. In a rodent model of insulin resistance and hypertriglyceridemia, arginine supplementation given for 4 weeks counteracts hypertension and hypertriglyceridemia [157]. Of note, in a model of type 2 diabetes in mice, 12 week-treatment with arginine in supplement (1 mg arginine/kg/day) allows increased mitochondrial respiration and biogenesis in cardiomyocytes isolated from animals when compared to the situation in cardiomyocytes isolated from control animals without supplement [158].
Regarding BCAAs, in a paper published more than 5 decades ago, circulating concentrations of these amino acids were found elevated in obese humans [159]. The same situation prevails in genetically obese rodents [160,161,162], but the relationship between BCAA concentrations and parameters defining metabolic syndrome in animal models remain obscure. A unifying hypothesis proposes that obesity associated insulin resistance affects BCAA metabolism (and then the circulating concentrations of BCAAs) [163], but it remains also possible that increased circulating BCAA concentrations play a role in insulin resistance/glucose tolerance which are associated with metabolic syndrome. In animal model, BCAA supplementation does not worsen insulin resistance or glucose tolerance [164], thus suggesting that increased BCAA concentrations is likely more a consequence than a cause of insulin resistance. However, it is not possible to totally exclude that ingestion of massive dose of BCAAs may promote detrimental effects in specific situations. For instance, in a mice model of atherosclerosis, supplementation with BCAAs (BCAAs in drinking water at 3 millimolar concentration) accelerates atherosclerosis progression [165].
BCAA have been tested in clinical trials and animal models for their effect on different parameters associated with metabolic syndrome. In a double blind, randomized controlled trial, leucine supplementation (3g/day) given for 8 weeks during energy restriction results in improved fat free mass and lean tissue mass compared to placebo in adults with metabolic syndrome [166]. In mice, leucine supplementation (6%, which corresponds to the amount of leucine in a high protein diet) given for 20 weeks shows beneficial effects on liver triglyceride content and insulin sensitivity [167]. The other BCAA isoleucine (2.5%), when given as supplement during 4 weeks to dietary-induced obese mice prevents partly the accumulation of triglycerides in liver and lowers the epididymal white adipose tissue mass [168].
The effects of glycine supplementation (15 g/day) vs. placebo given for 3 months have been tested in volunteers with characteristics of metabolic syndrome and this amino acid decreases the systolic blood pressure [169]. Glycine supplementation (1%) improves insulin sensitivity in a rodent model of sucrose-induced insulin resistance [170], and interestingly, this improvement is paralleled by increased concentration of glutathione in liver.
In a randomized controlled trial with obese individuals with characteristics of metabolic syndrome, histidine supplementation (4 g/day) when given for 12 weeks improves insulin resistance [171].
Lastly, when phenylalanine is given to rats in drinking water (2 g/L) for 28 weeks, it results in an increased hepatic lipid deposition [172]. Such phenylalanine supplementation represents 55 mg phenylalanine per day. This amount can be compared to the usual consumption of phenylalanine from rat regular diet, that is 136 mg phenylalanine per day [43].
3.6. Amino Acid Supplementation in Miscellaneous Situations
There are some data from preclinical and clinical studies which suggest that amino acid supplementation could be of value in different other situations, notably in case of inadequate protein intake. Although this narrative review does not claim to be exhaustive, several typical situations are presented here. For instance, regarding osteoporosis prevention, in a randomized study increased protein intake (from 58 to 69 g per day together with increased calcium intake) in the elderly receiving an adequate vitamin D supply is associated with significant reduction of all fractures, hip fractures, and falls [173]. Because glutamate has been implicated in osteoblast and osteoclast differentiation [174,175,176], because this amino acid displays a mitogenic effect on osteoblasts [177], and because glutamate prevents decreased bone mineral density in femur and tibia after systemic administration [178], glutamate supplementation has been tested for its effect on bone quality in situation when such quality is compromised. Monosodium glutamate (from 5 to 20 g/kg diet) given for 6 to 12 weeks partly preserves bone quality in mice under moderate protein restriction [179]. However, relatively high doses of glutamate in supplements were used to observe significant effects in this latter study, and this can be explained by the fact that virtually all glutamate in a regular diet is metabolized by the gut during absorption [180,181]. Then, it would be of interest to test the effects of glutamate in supplements in randomized controlled clinical trial with older volunteers at risk of osteoporosis. The study of the effects of proline supplementation on bone quality would be also of interest because this amino acid can be hydroxylated in the collagen proteins, giving rise to hydroxyproline [182]. Proline and hydroxyproline are abundant in collagens [183].
Supplementation with amino acids has been tested in hemodialysis patients and in cirrhotic patients. Interestingly, oral administration of a mixture containing the 9 indispensable amino acids together with 9 dispensable amino acids (5.4 g amino acids) administered on interdialytic days to hemodialysis patients was able to counteract the amino acid loss in these patients [184]. In a randomized controlled clinical trial with compensated cirrhotic patients, supplementation with BCAAs (4.06, 3.52, and 3.26 g/day of leucine, isoleucine, and valine, respectively) for 16 weeks improves liver frailty index and muscle mass [185].
Although an observational study found that substituting dietary low-quality carbohydrates with isocaloric animal protein is significantly associated with lower prevalence of cognitive decline in older people [186], and although higher protein intake is associated with lower odds of subjective cognitive decline in two recent studies [187,188], little is known on the effects of supplementation with one or several specific amino acids on such processes. One study in mice shows that dietary supplementation with indispensable amino acids together with cysteine and arginine increased the mitochondrial biogenesis and endogenous antioxidant response in the hippocampus, to an extent comparable to those elicited by exercise [189]. These results suggest beneficial effects of exercise and indispensable amino acids on this cerebral structure involved notably in memory and learning [190,191].
4. Safe Utilization of Amino Acids in Supplements
The Upper Levels of Safe Intake (ULSI) for amino acid must refer to the highest level of daily amino acid intake (in the form of proteins and as free amino acids) from all sources that is likely to pose no risk of adverse outcomes in healthy individuals. Such ULSI can be determined in reference to the No-Observed-Adverse-Effect level (NOAEL) values and to the Lowest-Observed-Adverse-Effect level (LOAEL) values [45]. The Food and Nutrition Board (FNB) and the Institute of Medicine (IOM) in their 2005 report suggest that the tolerable upper intake levels should preferentially referred to NOAEL values when available [41]. The NOAEL values for all individual amino acids can be determined from clinical trials. NOAEL and LOAEL values have been determined in preclinical studies using mostly the rat model. These values cannot be extrapolated to humans because such extrapolation is hazardous, notably because the usual amino consumption from all sources in rats is one order of magnitude higher than the consumption of amino acids in humans when expressed as mg amino acids per kg body weight per day. Then, NOAEL and LOAEL values for amino acids determined in rats are more informative when considered in relation with the animal usual consumption (UC) and the NOAEL/UC ratios can provide a rough estimation of the relative tolerance of the individual amino acids in this experimental model [43] as detailed below.
The duration of the dietary intervention with amino acid supplements in clinical trials is rather variable ranging usually from three to 13 weeks, and the parameters used as well as the dietary context, age, and sex of individuals often differ between studies. The NOAEL values are based on parameters which include anthropomorphic analysis (body weight, body composition, body mass index), dietary and energy intake, blood and urine biochemical analyses, maximal capacity to oxidize amino acids, circulating concentrations of amino acids and their metabolites, gastrointestinal signs (nausea, diarrhea, etc.), questionnaires on the perceived quality of life, adverse events often classified as mild (which require no treatment), moderate (which require brief pharmacological treatment), and severe (which require hospitalization) and clinical parameters such as blood pressure, renal, cardiovascular, and hepatic functions.
The NOAEL values for amino acids have been determined in clinical trials for histidine, leucine, lysine, methionine, phenylalanine, threonine, tryptophan, arginine, glycine, and serine. Regarding the indispensable amino acid histidine, this amino acid was given as supplements for 4 weeks to men and women, and different parameters including anthropometric analysis, determination of body composition, sleep patterns, dietary intake, blood biochemistry including measurement of histamine (produced from histidine by decarboxylation), and urine analysis were performed. According to the results obtained, a NOAEL value for histidine in supplement of 8 g/day was proposed [192], a value significantly higher than UC for adults in Western countries (2.0 g/day).
The amino acid leucine was given to adult men as supplements for 8 hours and based on parameters such as the upper capacity to oxidize leucine, as well as glycemia, insulinemia, alanine aminotransferase and ammonemia, it was found that the safe upper limit of leucine intake is 35-38 g/day [193,194]. In elderly men, the upper limit for leucine intake is like the value determined in young men [194]. This value is much higher when compared to the UC (5.8 g/day) for adults in Western countries, but it must be underlined here that this upper safe limit has been determined in very short-term trials so that no NOAEL value can be proposed for this amino acid.
Supplementation with lysine added to an ordinary diet performed for different periods of time (up to 36 months) can cause symptoms related to the gastrointestinal tract such as nausea and diarrhea. These results led to propose in a literature review a provisional NOAEL for lysine in food and supplements equal to 6 g/day [195]. This later value is close to the UC for adults (5.1 g/day), thus suggesting that relatively small amounts of lysine in supplements may lead to mild adverse events.
When methionine is given as supplement for 4 weeks to older adults (men and women), based on plasma homocysteine concentration used as the primary determinant (homocysteine is produced from methionine in three steps), but also on numerous other biochemical blood variables, the NOAEL value for methionine in supplements was found to be equal to 3.2 g/day [196]. This value is above the UC for adults (1.7 g/day).
Regarding phenylalanine, when given as supplement to male volunteers for 4 weeks, based on parameters including blood biochemistry and absence of treatment-related adverse events, the NOAEL value for phenylalanine in supplement was found to be 12 g/day [197], a value which can be compared to the UC of this amino acid for adults (3.3 g/day). The difference between the NOAEL value and UC suggest that phenylalanine in supplements is well tolerated in humans.
The NOAEL value was determined for threonine in supplements in a double blind randomized controlled trials performed for 4 weeks in men. Based on anthropometric parameters, blood chemistry (including amino acid concentrations), dietary intake, and the detection of adverse events, a NOAEL value of 12 g/day was proposed [198]. Interestingly, when measured after an overnight fast in plasma obtained from volunteers at the end of the 4 week-trial, a marked specific increase of threonine concentration was measured after consumption of threonine in supplements at doses between 6 and 12 g/day when compared to placebo value. Since the amino acid plasma concentrations can be considered in the first place as the net result of intestinal absorption and uptake/release from tissues, and since absorption of amino acids resulting from dietary protein digestion is minimal after an overnight fast [199,200], these results suggest possible modifications of threonine metabolism after 4 week-supplementation with this amino acid. When compared with the UC for threonine in adults (2.9 g/day), the NOAEL value is about four times higher.
For tryptophan, the NOAEL value for supplementation was determined in a double blind randomized controlled clinical study performed with female volunteers using values from blood and urine biochemistry (notably the tryptophan metabolites nicotinamide, kynurenin, and kynurenic acid in urine) and profile of mood states category measurement. The tryptophan NOAEL value was found to be 5g/day [201]. This value, when compared to the UC in adults (0.8 g/day), suggests that tryptophan is well tolerated when given in supplements.
Regarding the dispensable amino acids arginine and serine, NOAEL values have been determined in volunteers. Regarding arginine, in a randomized controlled clinical trial with overweight or obese but otherwise healthy volunteers (men and women), the NOAEL value was determined from blood biochemistry measurement, as well as measurement of cardiovascular, renal, and hepatic parameters. The results obtained indicate that supplementation with arginine at dose of 30 g/day for three months can be considered as safe [202]. In their review of literature, Shao and Hathcock, after considering the observed safe level of arginine in supplements, propose to not go beyond 20 g arginine per day [203]. By reviewing the randomized controlled trials performed with arginine supplementation in volunteers, Kuramochi and collaborators have proposed a lower NOAEL value (7.5 g/day) for this amino acid in supplements because of light gastrointestinal symptoms recorded in some participants at higher doses [204]. If we consider the lower value of NOAEL proposed for arginine (thus 7.5 g/day) and if we compare this value with the UC (4.2 g/day), this amino acid appears to be relatively well tolerated since the NOAEL/UC ratio is equal to 1.8.
For serine, the NOAEL value has been determined in male volunteers based on the circulating concentrations of biochemical analytes (including amino acids) and found to be 12.0 g/day in supplement [197]. When compared with the UC for serine (3.9 g/day), the NOAEL value for this amino acid is approximately 3 times higher.
Determination of NOAEL values has been also made for the non-proteinogenic amino acids ornithine and citrulline which are used as dietary supplements in different situations [205,206,207]. The NOAEL values for ornithine and citrulline in supplements were determined in a 4-week clinical trial performed in men based on the measurement of circulating biochemical analytes (including amino acid) concentrations, the occurrence of adverse events, and mental self-assessment [208]. According to this study, the NOAEL values for ornithine and citrulline were 12 g/day and 24 g/day, respectfully. Literature review performed on clinical studies dealing with oral intake with ornithine up to 22-week consumption has confirmed a NOAEL value equal to 12 g/day for this amino acid in supplements [209]. The NOAEL values for the amino acids for which it has been determined are summarized in Table 2.
For the other indispensable amino acids (isoleucine and valine), and dispensable amino acids (alanine, asparagine, aspartate, cysteine, glutamate, glutamine, glycine, proline, and tyrosine), no NOAEL or LOAEL values have been determined in clinical trials. For glutamine, an observed safe level of 14 g/day has been proposed for human consumption [203]. Then, only data from animal studies are available. Although as previously said, it is not possible to extrapolate from animal models to humans, some studies in rats can bring useful information [43]. These rat studies suggest that the NOAEL values can be much different for some amino acids according to the sex of animals, raising the possibility of differential tolerance to supplements between males and females in other mammals including humans. Indeed, the NOAEL values for valine and tyrosine are higher in male than female rats [210,211], while for tryptophan, the NOAEL value is higher in female than male rats [212].
By using the NOAEL and UC values determined in rats, it is possible to calculate the NOAEL/UC ratios for some amino acids. Such calculations give a rough estimation of the tolerance of amino acids in supplements for rats [43]. By doing so, alanine appears well tolerated in rats (NOAEL/UC ratio is 6.9), followed by serine (4.8), leucine (3.7), isoleucine (3.1), proline (2.3), and aspartate (1.0), this latter amino acid appearing thus to be the least tolerated amino acid among this list.
5. Conclusions and Prospects
The NOAEL values for amino acid in supplements in humans have been determined for only 9 amino acids among the 20 common amino acids found in proteins. In addition to these 9 proteinogenic amino acids, NOAEL values have been ascertained for the non-proteinogenic amino acids ornithine and citrulline. Thus, for the other 11 proteinogenic amino acids, additional clinical trials are needed to estimate the upper levels of safe intake of these amino acids in supplements.
What could be the mechanistic reasons, beyond the clinical signs, that would explain deleterious effects of excessive amounts of amino acids in supplements? Although we still have a limited vision on the mechanisms involved, some studies suggest that excessive amounts of amino acids could disturb the normal overall metabolism of these compounds. For instance, it is well known that the amino acid transporters situated in the luminal brush border- and in the baso-lateral membranes of the enterocytes can transport several amino acids with different affinity [213]. Furthermore, a particular amino acid may modulate the transport and metabolism of other amino acids along the small intestine [214]. Then, a supply of an excessive amount of one given amino acid may have an impact on the transport of other amino acids and may then affect their metabolism firstly in enterocytes and then in peripheral tissues after absorption [215]. Such modifications may affect amino acid-dependent physiological processes. From this point of view, capacity of metabolic adaptation after medium term amino acid supplementation should be considered to evaluate the possible impact of such supplementation in humans.
In specific situations of insufficient protein intake, supplementation with individual amino acids with relevant quantities may be recommended to allow optimal physiological and metabolic functions as documented in this review. This is the case for instance for subpopulations among the elderly with protein-energy deficiency status. According to clinical trials performed in elderly with sarcopenia or at risk for sarcopenia, such amino acid mixtures should contain a relatively high content in leucine among the other amino acids. Such dietary supplementation appears more effective when combined with regular physical exercise.
In athletes, branched-chain amino acids appear able to increase muscle protein synthesis during the recovery phase. In experiments with healthy volunteers receiving leucine by intravenous infusion, results suggest that this BCAA decreases the overall protein degradation in the body. Possible supplementation with BCAAs in athletes should be reasoned in relation to the recommended intake of proteins in this subpopulation. As indicated above, the recommended intake of protein in athletes is 1.6 to 2.2 times higher than in the general adult population.
The question of the supply of amino acids above the usual requirements for obtaining optimized metabolic and physiological outcomes in some specific situations remains to be answered in an unequivocal manner. For instance, as presented above, after an episode of experimental intestinal inflammatory flare in rodents, the mucosal healing phase seems to require a higher uptake of alimentary proteins than in healthy animals, and some specific amino acids in supplements appear efficient for such a process. These results encourage further innovative experimental and clinical studies on the protein and amino acid requirement during the healing phase after inflammatory episode.
Importantly, the multifunction roles of amino acids both as precursors for the synthesis of macromolecules (proteins and the purine and pyrimidine rings in RNAs and DNA) and as precursors for the synthesis of bioactive compounds should be considered when evaluating the relevance of supplementation with amino acid(s) in each situation. In fact, these numerous amino acid-derived compounds exert central metabolic and physiological roles, and the chemical identity and roles of these compounds are different according to the amino acid considered [1]. In addition, specific amino acids are precursors for ATP synthesis and are involved as energy substrates in numerous cell phenotypes.
During weight loss programs in obese volunteers, supplementation with mixtures of indispensable amino acids, in a context of lower protein-energy intake, has shown some positive effects on the maintenance of the lean mass. In clinical trials with volunteers characterized by biological parameters associated with metabolic syndrome, the indispensable amino acids leucine and histidine, as well as the conditionally indispensable amino acids arginine and glycine, have shown some beneficial effects on the underlying criteria which define this syndrome.
Overall, the challenge for the coming years is to identify more precisely the different physiological and pathophysiological situations, in different subpopulations, for which supplementation with amino acids will prove to be useful in a preventive or curative perspective (Figure 3).
The determination of the amino acid requirement in these different situations is an important part of this research. Finally, the identification of the doses of amino acids, either in individual or mixture form, together with the determination of the usage duration which are efficient and safe, are needed. Regarding the safety of amino acids in supplement, the NOAEL values for all the amino acids for which we have no values need to be determined in future clinical trials to get robust reference values for users.
Funding
The writing of this review received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analysed in this review. Data sharing is not applicable to this article.
Conflicts of Interest
The author declare no conflicts of interest.
References
- Blachier, F. Amino acid metabolism for human physiology. In The evolutionary journey of amino acids. From the origin of life to human metabolism; Springer Nature: Cham, Switzerland, 2025. [Google Scholar]
- Bechaux, J.; Gatellier, P.; Le Page, J.F.; Drillet, Y.; Sante-Lhoutellier, V. A comprehensive review of bioactive peptides obtained from animal byproducts and their applications. Food Funct. 2019, 10, 6244–6266. [Google Scholar] [CrossRef] [PubMed]
- Caldovic, L.; Tuchman, M. N-acetylglutamate and its changing role through evolution. Biochem. J. 2003, 372, 279–290. [Google Scholar] [CrossRef]
- Newsholme, P.; Procopio, J.; Lima, M.M.; Pithon-Curi, T.C.; Curi, R. Glutamine and glutamate--their central role in cell metabolism and function. Cell Biochem. Funct. 2003, 21, 1–9. [Google Scholar] [CrossRef]
- Wahren, J.; Ekberg, K. Splanchnic regulation of glucose production. Annu. Rev. Nutr. 2007, 27, 329–45. [Google Scholar] [CrossRef]
- Neinast, M.; Murashige, D.; Arany, Z. Branched Chain Amino Acids. Annu. Rev. Physiol. 2019, 81, 139–164. [Google Scholar] [CrossRef]
- Ignarro, L.J. Endothelium-derived nitric oxide: actions and properties. FASEB J. 1989, 3, 31–36. [Google Scholar] [CrossRef]
- Brosnan, J. T.; Brosnan, M.E. The sulfur-containing amino acids: an overview. J. Nutr. 2006, 136, 1636S–1640S. [Google Scholar] [CrossRef] [PubMed]
- Etchegaray, J.P.; Mostoslavsky, R. Interplay between metabolism and epigenetics: a nuclear adaptation to environmental changes. Mol. Cell. 2016, 62, 695–711. [Google Scholar] [CrossRef] [PubMed]
- Malmezat, T.; Breuillé, D.; Capitan, P.; Mirand, P.P.; Obled, C. Glutathione turnover is increased during the acute phase of sepsis in rats. J. Nutr. 2000, 130, 1239–1246. [Google Scholar] [CrossRef]
- Rémond, D.; Buffière, C.; Pouyet, C.; Papet, I.; Dardevet, D.; Savary-Auzeloux, I.; Williamson, G.; Faure, M.; Breuillé, D. Cysteine fluxes across the portal-drained viscera of enterally fed minipigs: effect of an acute intestinal inflammation. Amino Acids 2011, 40, 543–552. [Google Scholar] [CrossRef]
- Schutz, Y. Protein turnover, ureagenesis and gluconeogenesis. Int. J. Vitam. Nutr. Res. 2011, 81, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Gietzen, D.W.; Rogers, QR. Nutritional homeostasis and indispensable amino acid sensing: a new solution to an old puzzle. Trends Neurosci. 2006, 29, 91–99. [Google Scholar] [CrossRef]
- Reeds, P.J. Dispensable and indispensable amino acids for humans. J. Nutr. 2000, 130, 1835S–1840S. [Google Scholar] [CrossRef]
- Sakamoto, A.; Nishimura, Y.; Ono, H.; Sakura, N. Betaine and homocysteine concentrations in foods. Pediatr. Int. 2002, 44, 409–413. [Google Scholar] [CrossRef]
- Blachier, F.; Andriamihaja, M.; Blais, A. Sulfur-containing amino acids and lipid metabolism. J. Nutr. 2020, 150, 2524S–2531S. [Google Scholar] [CrossRef] [PubMed]
- Manary, M.J.; Wegner, D.R.; Maleta, K. Protein quality malnutrition. Front. Nutr. 2024, 11, 1428810. [Google Scholar] [CrossRef]
- Fürst, P.; Stehle, P. What are the essential elements needed for the determination of amino acid requirements in humans? J. Nutr. 2004, 134, 1558S–1565S. [Google Scholar] [CrossRef]
- Yu, Y.M.; Ryan, C.M.; Castillo, L.; Lu, X.M.; Beaumier, L.; Tompkins, R.G.; Young, V.R. Arginine and ornithine kinetics in severely burned patients: increased rate of arginine disposal. Am. J. Physiol. 2001, 280, E509–E517. [Google Scholar] [CrossRef]
- McPherson, R.A.; Hardy, G. Clinical and nutritional benefits of cysteine-enriched protein supplements. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 562–568. [Google Scholar] [CrossRef]
- Muro, P.; Zhang, L.; Li, S.; Zhao, Z.; Jin, T.; Mao, F.; Mao, Z. The emerging role of oxidative stress in inflammatory bowel disease. Front. Endocrinol. (Lausanne) 2024, 15, 1390351. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; Biswas, S.K.; Jimenez, L.A.; Torres, M.; Forman, H.J. Glutathione, stress responses, and redox signaling in lung inflammation. Antioxid. Redox. Signal. 2005, 7, 42–59. [Google Scholar] [CrossRef] [PubMed]
- Lacey, J.M.; Wilmore, D.W. Is glutamine a conditionally essential amino acid? Nutr. Rev. 1990, 48, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, B.F.; Ennis, M.A.; Dyer, R.A.; Lim, K.; Elango, R. Glycine, a dispensable amino acid, is conditionally indispensable in late stages of human pregnancy. J. Nutr. 2021, 151, 361–369. [Google Scholar] [CrossRef]
- Wu, G. Dietary protein intake and human health. Food Funct. 2016, 7, 1251–1265. [Google Scholar] [CrossRef]
- Rand, W.M.; Pellett, P.L.; Young, V.R. Meta-analysis of nitrogen balance studies for estimating protein requirements in healthy adults. Am. J. Clin. Nutr. 2003, 77, 109–127. [Google Scholar] [CrossRef]
- Dubuisson, C.; Lioret, S.; Touvier, M.; Dufour, A.; Calamassi-Tran, G.; Volatier, J.L.; Lafay, L. Trends in food and nutritional intakes of French adults from 1999 to 2007: results from the INCA surveys. Br. J. Nutr. 2010, 103, 1035–1048. [Google Scholar] [CrossRef] [PubMed]
- Pasiakos, S.M.; Agarwal, S.; Lieberman, H.R.; Fulgoni, V.L, 3rd. Sources and amounts of animal, dairy, and plant protein intake of US adults in 2007-2010. Nutrients 2015, 7, 7058–7069. [Google Scholar] [CrossRef]
- Bakaloudi, D.R.; Halloran, A.; Rippin, H.L.; Oikonomidou, A.C.; Dardavesis, T.I.; Williams, J.; Wickramasinghe, K.; Breda, J.; Chourdakis, M. Intake and adequacy of the vegan diet. A systematic review of the evidence. Clin. Nutr. 2021, 40, 3503–3521. [Google Scholar] [CrossRef]
- Allès, B.; Baudry, J.; Méjean, C.; Touvier, M.; Péneau, S.; Hercberg, S.; Kesse-Guyot, E. Comparison of sociodemographic and nutritional characteristics between self-reported vegetarians, vegans, and meat-eaters from the NutriNet-Santé study. Nutrients 2017, 9, 1023. [Google Scholar] [CrossRef]
- Al Hasan, S.M.; Saulam, J.; Mikami, F.; Kanda, K.; Ngatu, N.R.; Yokoi, H.; Hirao, T. Trends in per capita food and protein availability at the national level of the Southeast Asian countries: An Analysis of the FAO’s Food Balance Sheet Data from 1961 to 2018. Nutrients 2022, 14, 603. [Google Scholar] [CrossRef]
- Asayehu, T.T.; Lachat, C.; Henauw, S.; Gebreyesus, S.H. Dietary behaviour, food and nutrient intake of women do not change during pregnancy in Southern Ethiopia. Matern. Child. Nutr. 2017, 13, e12343. [Google Scholar] [CrossRef]
- Agence Française de Sécurité Sanitaire des Aliments/ French Agency for Food Safety. Protein intake: dietary intake, quality, requirements and recommendations. 2007.
- Burstad, K.M.; Lamina, T.; Erickson, A.; Gholizadeh, E.; Namigga, H.; Claussen, A.M.; Slavin, J.L.; Teigen, L.; Hill Gallant, K.M.; Stang, J.; Steffen, L.M.; Harindhanavudhi, T.; Kouri, A.; Duval, S.; Butler, M. Evaluation of dietary protein and amino acid requirements: a systematic review. Am. J. Clin. Nutr. 2025, 122, 285–305. [Google Scholar] [CrossRef]
- Rogeri, P.S.; Zanella, R., Jr.; Martins, G.L.; Garcia, M.D.A.; Leite, G.; Lugaresi, R.; Gasparini, S.O.; Sperandio, G.A.; Ferreira, L.H.B.; Souza-Junior, T.P.; Lancha, A.H., Jr. Strategies to prevent sarcopenia in the aging process: role of protein intake and exercise. Nutrients 2021, 14, 52. [Google Scholar] [CrossRef]
- Wolfe, R.R.; Miller, S.L.; Miller, K.B. Optimal protein intake in the elderly. Clin. Nutr. 2008, 27, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Baerlocher, K.; Bauer, J.M.; Elmadfa, I.; Heseker, H.; Leschik-Bonnet, E.; Stangl, G.; Volkert, D.; Stehle, P.; on behalf of the German Nutrition Society (DGE). Revised Reference Values for the Intake of Protein. Ann. Nutr. Metab. 2019, 74, 242–250. [Google Scholar] [CrossRef]
- Lancha, A.H., Jr.; Zanella, R., Jr.; Tanabe, S.G.; Andriamihaja, M.; Blachier, F. Dietary protein supplementation in the elderly for limiting muscle mass loss. Amino Acids 2017, 49, 33–47. [Google Scholar] [CrossRef]
- Crichton, M.; Craven, D.; Mackay, H.; Marx, W.; de van der Schueren, M.; Marshall, S. A systematic review, meta-analysis and meta-regression of the prevalence of protein-energy malnutrition: associations with geographical region and sex. Age Ageing 2019, 48, 38–48. [Google Scholar] [CrossRef]
- Phillips, S.M.; Van Loon, L.J. Dietary protein for athletes: from requirements to optimum adaptation. J. Sports Sci. 2011, 29, S29–38. [Google Scholar] [CrossRef] [PubMed]
- Food and Nutrition Board (FNB); Institute of Medicine (IOM). Dietary reference intake for energy, carbohydrate, fiber, fat, fatty acids, cholesterol, protein, and amino acids; The National Academies Press: Washington DC, 2005. [Google Scholar]
- World Health Organization (WHO). Protein and amino acid requirements in human nutrition. Report of a Joints WHO/FAO/UNU Expert Consultation; WHO Press: Geneva, Switzerland, 2007. [Google Scholar]
- Blachier, F.; Blais, A.; Elango, R.; Saito, K.; Shimomura, Y.; Kadowaki, M.; Matsumoto, H. Tolerable amounts of amino acids for human supplementation: summary and lessons from published peer-reviewed studies. Amino Acids 2021, 53, 1313–1328. [Google Scholar] [CrossRef]
- Iguacel, I.; Perez-Cornago, A.; Schmidt, J.A.; Van Puyvelde, H.; Travis, R.; Casagrande, C.; Nicolas, G.; Riboli, E.; Weiderpass, E.; Ardanaz, E.; et al. Evaluation of protein and amino acid intake estimates from the EPIC dietary questionnaires and 24-h dietary recalls using different food composition databases. Nutr. Metab. Cardiovasc. Dis. 2022, 32, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Elango, R. Tolerable upper intake for individual amino acids in humans: a narrative review of recent clinical studies. Adv. Nutr. 2023, 14, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyère, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; Schneider, S.M.; Sieber, C.C.; Topinkova, E.; Vandewoude, M.; Visser, M.; Zamboni, M. Writing Group for the European Working Group on Sarcopenia in Older People 2 (EWGSOP2), and the Extended Group for EWGSOP2.Sarcopenia: revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 16–31. [Google Scholar] [CrossRef]
- Balagopal, P.; Rooyackers, O.E.; Adey, D.B.; Ades, P.A.; Nair, K.S. Effects of aging on in vivo synthesis of skeletal muscle myosin heavy-chain and sarcoplasmic protein in humans. Am. J. Physiol. 1997, 273, E790–E800. [Google Scholar] [CrossRef]
- Rooyackers, O.E.; Adey, D.B.; Ades, P.A.; Nair, K.S. Effect of age on in vivo rates of mitochondrial protein synthesis in human skeletal muscle. Proc. Natl. Acad. Sci. U S A 1996, 93, 15364–15369. [Google Scholar] [CrossRef]
- Short, K.R.; Bigelow, M.L.; Kahl, J.; Singh, R.; Coenen-Schimke, J.; Raghavakaimal, S.; Nair, K.S. Decline in skeletal muscle mitochondrial function with aging in humans. Proc. Natl. Acad. Sci. U S A 2005, 102, 5618–5623. [Google Scholar] [CrossRef]
- Tezze, C; Sandri, M; Tessari, P. Anabolic resistance in the pathogenesis of sarcopenia in the elderly: role of nutrition and exercise in young and old people. Nutrients 2023, 15, 4073. [Google Scholar] [CrossRef]
- Houston, D.; K. Nicklas, B.J.; Ding, J.; Harris, T.B.; Tylavsky, F.A.; Newman, A.B.; Lee, J.S.; Sahyoun, N.R.; Visser, M.; Kritchevsky, S.B. Health ABC Study. Dietary protein intake is associated with lean mass change in older, community-dwelling adults: the Health, Aging, and Body Composition (Health ABC) Study. Am. J. Clin. Nutr. 2008, 87, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Paddon-Jones, D.; Rasmussen, B.B. Dietary protein recommendations and the prevention of sarcopenia. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Li, F.; Li, Y.; Tang, Y.; Kong, X.; Feng, Z.; Anthony, T.G.; Watford, M.; Hou, Y.; Wu, G; Yin, Y. The role of leucine and its metabolites in protein and energy metabolism. Amino Acids 2016, 48, 41–51. [Google Scholar] [CrossRef]
- Nie, C.; He, T.; Zhang, W.; Zhang, G.; Ma, X. Branched chain amino acids: beyond nutrition metabolism. Int. J. Mol. Sci. 2018, 19, 954. [Google Scholar] [CrossRef]
- Wilkinson, K.; Koscien, C.P.; Monteyne, A.J.; Wall, B.T.; Stephens, F.B. Association of postprandial postexercise muscle protein synthesis rates with dietary leucine: A systematic review. Physiol. Rep. 2023, 11, e15775. [Google Scholar] [CrossRef] [PubMed]
- Rieu, I.; Balage, M.; Sornet, C.; Giraudet, C.; Pujos, E.; Grizard, J.; Mosoni, L.; Dardevet, D. Leucine supplementation improves muscle protein synthesis in elderly men independently of hyperaminoacidaemia. J. Physiol. 2006, 575, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Church, D.D.; Hirsch, K.R.; Park, S.; Kim, I.Y.; Gwin, J.A.; Pasiakos, S.M.; Wolfe, R.R.; Ferrando, A.A. Essential amino acids and protein synthesis: insights into maximizing the muscle and whole-body response to feeding. Nutrients 2020, 12, 3717. [Google Scholar] [CrossRef]
- Rondanelli, M.; Klersy, C.; Terracol, G.; Talluri, J.; Maugeri, R.; Guido, D.; Faliva, M.A.; Solerte, B.S.; Fioravanti, M.; Lukaski, H.; Perna, S. Whey protein, amino acids, and vitamin D supplementation with physical activity increases fat-free mass and strength, functionality, and quality of life and decreases inflammation in sarcopenic elderly. Am. J. Clin. Nutr. 2016, 103, 830–840. [Google Scholar] [CrossRef]
- Rondanelli, M.; Cereda, E.; Klersy, C.; Faliva, M.A.; Peroni, G.; Nichetti, M.; Gasparri, C.; Iannello, G.; Spadaccini, D.; Infantino, V.; Caccialanza, R.; Perna, S. Improving rehabilitation in sarcopenia: a randomized-controlled trial utilizing a muscle-targeted food for special medical purposes. J. Cachexia Sarcopenia Muscle 2020, 11, 1535–1547. [Google Scholar] [CrossRef]
- Bauer, J.M.; Verlaan, S.; Bautmans, I.; Brandt, K.; Donini, L.M.; Maggio, M.; McMurdo, M.E.; Mets, T.; Seal, C.; Wijers, S.L.; Ceda, G.P.; De Vito, G.; Donders, G.; Drey, M.; Greig, C.; Holmbäck, U.; Narici, M.; McPhee, J.; Poggiogalle, E.; Power, D.; Scafoglieri, A.; Schultz, R.; Sieber, C.C.; Cederholm, T. Effects of a vitamin D and leucine-enriched whey protein nutritional supplement on measures of sarcopenia in older adults, the PROVIDE study: a randomized, double-blind, placebo-controlled trial. J. Am. Med. Dir. Assoc. 2015, 16, 740–747. [Google Scholar] [CrossRef]
- Rafiq, S.; Huma, N.; Pasha, I.; Sameen, A.; Mukhtar, O.; Khan, MI. Chemical composition, nitrogen fractions and amino acids profile of milk from different animal species. Asian-Australas J. Anim. Sci. 2016, 29, 1022–1028. [Google Scholar] [CrossRef]
- Szwiega, S.; Pencharz, P.B.; Rafii, M.; Lebarron, M.; Chang, J.; Ball, R.O.; Kong, D.; Xu, L.; Elango, R.; Courtney-Martin, G. Dietary leucine requirement of older men and women is higher than current recommendations. Am. J. Clin. Nutr. 2021, 113, 410–419. [Google Scholar] [CrossRef]
- Ispoglou, T.; Witard, O.C.; Duckworth; L, C.; Lees, M.J. The efficacy of essential amino acid supplementation for augmenting dietary protein intake in older adults: implications for skeletal muscle mass, strength and function. Proc. Nutr. Soc. 2021, 80, 230–242. [Google Scholar] [CrossRef]
- Dent, E.; Hoogendijk, E.O.; Visvanathan, R.; Wright, O.R.L. Malnutrition screening and assessment in hospitalised older people: a review. J. Nutr. Health Aging 2019, 23, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Fielding, R.A.; Landi, F.; Smoyer, K.E.; Tarasenko, L.; Groarke, J. Association of anorexia/appetite loss with malnutrition and mortality in older populations: A systematic literature review. J. Cachexia Sarcopenia Muscle 2023, 14, 706–729. [Google Scholar] [PubMed]
- Butterworth, M.; Lees, M.; Harlow, P.; Hind, K.; Duckworth, L.; Ispoglou, T. Acute effects of essential amino acid gel-based and whey protein supplements on appetite and energy intake in older women. Appl. Physiol. Nutr. Metab. 2019, 44, 1141–1149. [Google Scholar]
- Ispoglou, T.; Deighton, K.; King, R.F.; White, H.; Lees, M. Novel essential amino acid supplements enriched with L-leucine facilitate increased protein and energy intakes in older women: a randomised controlled trial. Nutr. J. 2017, 16, 75. [Google Scholar] [CrossRef]
- Rondanelli, M.; Opizzi, A.; Antoniello, N.; Boschi, F.; Iadarola, P.; Pasini, E.; Aquilani, R.; Dioguardi, F.S. Effect of essential amino acid supplementation on quality of life, amino acid profile and strength in institutionalized elderly patients. Clin. Nutr. 2011, 30, 571–577. [Google Scholar] [CrossRef]
- Ispoglou, T.; White, H.; Preston, T.; McElhone, S.; McKenna, J.; Hind, K. Double-blind, placebo-controlled pilot trial of L-Leucine-enriched amino-acid mixtures on body composition and physical performance in men and women aged 65-75 years. Eur. J. Clin. Nutr. 2016, 70, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Komar, B.; Schwingshackl, L.; Hoffmann, G. Effects of leucine-rich protein supplements on anthropometric parameter and muscle strength in the elderly: a systematic review and meta-analysis. J. Nutr. Health Aging 2015, 19, 437–446. [Google Scholar] [CrossRef]
- Gielen, E.; Beckwée, D.; Delaere, A.; De Breucker, S.; Vandewoude, M.; Bautmans, I.; Sarcopenia Guidelines Development Group of the Belgian Society of Gerontology and Geriatrics (BSGG). Nutritional interventions to improve muscle mass, muscle strength, and physical performance in older people: an umbrella review of systematic reviews and meta-analyses. Nutr. Rev. 2021, 79, 121–147. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Lepe, M.A.; Miranda-Gil, M.I.; Valbuena-Gregorio, E.; Olivas-Aguirre, F.J. Exercise programs combined with diet supplementation improve body composition and physical function in older adults with sarcopenia: A systematic review. Nutrients 2023, 15, 1998. [Google Scholar] [CrossRef]
- Giacosa, A.; Barrile, G.C.; Mansueto, F.; Rondanelli, M. The nutritional support to prevent sarcopenia in the elderly. Front. Nutr. 2024, 11, 1379814. [Google Scholar] [CrossRef]
- Conde Maldonado, E.; Marqués-Jiménez, D.; Casas-Agustench, P.; Bach-Faig, A. Effect of supplementation with leucine alone, with other nutrients or with physical exercise in older people with sarcopenia: a systematic review. Endocrinol. Diabetes Nutr. 2022, 69, 601–613. [Google Scholar] [CrossRef]
- Xu, Z.R.; Tan, Z.J.; Zhang, Q.; Gui, Q.F.; Yang, Y.M. The effectiveness of leucine on muscle protein synthesis, lean body mass and leg lean mass accretion in older people: a systematic review and meta-analysis. Br. J. Nutr. 2015, 113, 25–34. [Google Scholar] [CrossRef]
- Plotkin, D.L.; Delcastillo, K.; Van Every, D.W.; Tipton, K.D.; Aragon, A.A.; Schoenfeld, B.J. Isolated leucine and branched-chain amino acid supplementation for enhancing muscular strength and hypertrophy: a narrative review. Int. J. Sport Nutr. Exerc. Metab. 2021, 31, 292–301. [Google Scholar] [CrossRef]
- van Hall, G.; Raaymakers, J.S.; Saris, W.H.; Wagenmakers, A.J. Ingestion of branched-chain amino acids and tryptophan during sustained exercise in man: failure to affect performance. J. Physiol. 1995, 486, 789–794. [Google Scholar] [CrossRef]
- Nair, K.S.; Schwartz, R.G.; Welle, S. Leucine as a regulator of whole body and skeletal muscle protein metabolism in humans. Am. J. Physiol. 1992, 263, E928–E934. [Google Scholar] [CrossRef]
- Blomstrand, E.; Saltin, B. BCAA intake affects protein metabolism in muscle after but not during exercise in humans. Am. J. Physiol. 2001, 281, E365–E374. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Jahn, L.A.; Long, W.; Fryburg, D.A.; Wei, L.; Barrett, E.J. Branched chain amino acids activate messenger ribonucleic acid translation regulatory proteins in human skeletal muscle, and glucocorticoids blunt this action. J. Clin. Endocrinol. Metab. 2001, 86, 2136–2143. [Google Scholar] [CrossRef] [PubMed]
- Tipton, K.D.; Ferrando, A.A.; Phillips, S.M.; Doyle, D., Jr.; Wolfe, R.R. Postexercise net protein synthesis in human muscle from orally administered amino acids. Am. J. Physiol. 1999, 276, E628–E634. [Google Scholar] [CrossRef] [PubMed]
- West, D.W.; Burd, N.A.; Coffey, V.G.; Baker, S.K.; Burke, L.M.; Hawley, J.A; Moore, D.R.; Stellingwerff, T.; Phillips, S.M. Rapid aminoacidemia enhances myofibrillar protein synthesis and anabolic intramuscular signaling responses after resistance exercise. Am. J. Clin. Nutr. 2011, 94, 795–803. [Google Scholar] [CrossRef]
- Shimomura, Y.; Murakami, T.; Nakai, N.; Nagasaki, M.; Harris, R.A. Exercise promotes BCAA catabolism: effects of BCAA supplementation on skeletal muscle during exercise. J. Nutr. 2004, 134, 1583S–1587S. [Google Scholar] [CrossRef]
- Shimomura, Y.; Fujii, H.; Suzuki, M.; Murakami, T.; Fujitsuka, N.; Nakai, N. Branched-chain alpha-keto acid dehydrogenase complex in rat skeletal muscle: regulation of the activity and gene expression by nutrition and physical exercise. J. Nutr. 1995, 125, 1762S–1765S. [Google Scholar] [CrossRef]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef] [PubMed]
- Antonio, J.; Candow, D.G.; Forbes, S.C.; Gualano, B.; Jagim, A.R.; Kreider, R.B.; Rawson, E.S.; Smith-Ryan, A.E.; VanDusseldorp, T.A.; Willoughby, D.S.; et al. Common questions and misconceptions about creatine supplementation: what does the scientific evidence really show? J. Int. Soc. Sports Nutr. 2021, 18, 13. [Google Scholar] [CrossRef] [PubMed]
- Burke, R.; Piñero, A.; Coleman, M.; Mohan, A.; Sapuppo, M.; Augustin, F.; Aragon, A.A.; Candow, D.G.; Forbes, S.C.; Swinton, P.; et al. The effects of creatine supplementation combined with resistance training on regional measures of muscle hypertrophy: a systematic review with meta-analysis. Nutrients 2023, 15, 2116. [Google Scholar] [CrossRef]
- Jarmakiewicz-Czaja, S.; Zielińska, M.; Sokal, A.; Filip, R. Genetic and epigenetic etiology of inflammatory bowel disease: an update. Genes (Basel) 2022, 13, 2388. [Google Scholar] [CrossRef]
- Sandborn, W.J.; Feagan, B.G.; Hanauer, S.B.; Lochs, H.; Löfberg, R.; Modigliani, R.; Present, D.H.; Rutgeerts, P.; Schölmerich, J.; Stange, E.F.; Sutherland, L.R. A review of activity indices and efficacy endpoints for clinical trials of medical therapy in adults with Crohn’s disease. Gastroenterology 2002, 122, 512–530. [Google Scholar] [CrossRef]
- D’Haens, G.; Sandborn, W.J.; Feagan, B.G.; Geboes, K.; Hanauer, S.B.; Irvine, E.J.; Lémann, M.; Marteau, P.; Rutgeerts, P.; Schölmerich, J.; et al. A review of activity indices and efficacy end points for clinical trials of medical therapy in adults with ulcerative colitis. Gastroenterology 2007, 132, 763–786. [Google Scholar] [CrossRef] [PubMed]
- Lichtenstein, G.R.; Rutgeerts, P. Importance of mucosal healing in ulcerative colitis. Inflamm. Bowel Dis. 2010, 16, 338–346. [Google Scholar] [CrossRef]
- Pineton de Chambrun, G.; Peyrin-Biroulet, L.; Lémann, M.; Colombel, J.F. Clinical implications of mucosal healing for the management of IBD. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 15–29. [Google Scholar] [CrossRef]
- Dave, M.; Loftus, E.V., Jr. Mucosal healing in inflammatory bowel disease-a true paradigm of success? Gastroenterol. Hepatol. (N Y) 2012, 8, 29–38. [Google Scholar]
- Neurath, M.F.; Travis, S.P. Mucosal healing in inflammatory bowel diseases: a systematic review. Gut 2012, 61, 1619–1635. [Google Scholar] [CrossRef]
- Colombel, J.F.; Rutgeerts, P.; Reinisch, W.; Esser, D.; Wang, Y.; Lang, Y.; Marano, C.W.; Strauss, R.; Oddens, B.J.; Feagan, B.G.; et al. Early mucosal healing with infliximab is associated with improved long-term clinical outcomes in ulcerative colitis. Gastroenterology 2011, 141, 1194–201. [Google Scholar] [CrossRef]
- Lan, A.; Blachier, F.; Benamouzig, R.; Beaumont, M.; Barrat, C.; Coelho, D.; Lancha, A., Jr.; Kong, X.; Yin, Y.; Marie, J.C.; et al. Mucosal healing in inflammatory bowel diseases: is there a place for nutritional supplementation? Inflamm. Bowel Dis. 2015, 21, 198–207. [Google Scholar] [CrossRef]
- Sturm, A.; Dignass, A.U. Epithelial restitution and wound healing in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 348–353. [Google Scholar] [CrossRef]
- Alastair, F.; Emma, G.; Emma, P. Nutrition in inflammatory bowel disease. JPEN J. Parenter. Enteral Nutr. 2011, 35, 571–580. [Google Scholar] [CrossRef]
- Kulnigg, S.; Gasche, C. Systematic review: managing anaemia in Crohn’s disease. Aliment Pharmacol. Ther. 2006, 24, 1507–1523. [Google Scholar] [CrossRef]
- Nguyen, G.C.; Munsell, M.; Harris, M.L. Nationwide prevalence and prognostic significance of clinically diagnosable protein-calorie malnutrition in hospitalized inflammatory bowel disease patients. Inflamm. Bowel Dis. 2008, 14, 1105–1111. [Google Scholar] [CrossRef]
- Hartman, C.; Eliakim, R.; Shamir, R. Nutritional status and nutritional therapy in inflammatory bowel diseases. World J. Gastroenterol. 2009, 15, 2570–2578. [Google Scholar] [CrossRef]
- González-Huix, F.; de León, R.; Fernández-Bañares, F.; Esteve, M.; Cabré, E.; Acero, D.; Abad-Lacruz, A.; Figa, M.; Guilera, M.; Planas, R.; et al. Polymeric enteral diets as primary treatment of active Crohn’s disease: a prospective steroid controlled trial. Gut 1993, 34, 778–782. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Banares, F.; Cabré, E.; Esteve-Comas, M.; Gassull, M.A. How effective is enteral nutrition in inducing clinical remission in active Crohn’s disease ? A meta-analysis of the randomized clinical trials. JPEN J. Parenter. Enteral Nutr. 1995, 19, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Messori, A.; Trallori, G.; D’Albasio, G.; Milla, M.; Vannozzi, G.; Pacini, F. Defined-formula diets versus steroids in the treatment of active Crohn’s disease: a meta-analysis. Scand. J. Gastroenterol. 1996, 31, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, A.M.; Ohlsson, A.; Sherman, P.M.; Sutherland, L.R. Meta-analysis of enteral nutrition as a primary treatment of active Crohn’s disease. Gastroenterology 1995, 108, 1056–1067. [Google Scholar] [CrossRef] [PubMed]
- Forbes, A.; Escher, J.; Hébuterne, X.; Kłęk, S.; Krznaric, Z.; Schneider, S.; Shamir, R.; Stardelova, K.; Wierdsma, N.; Wiskin, A.E.; et al. ESPEN guideline: Clinical nutrition in inflammatory bowel disease. Clin. Nutr. 2017, 36, 321–347. [Google Scholar] [CrossRef]
- Vidal-Lletjós, S.; Khodorova, N.V.; Piscuc, M.; Gaudichon, C.; Blachier, F.; Lan, A. Tissue-specific effect of colitis on protein synthesis in mice: impact of the dietary protein content. Eur. J. Nutr. 2021, 60, 1669–1677. [Google Scholar] [CrossRef]
- Vidal-Lletjós, S.; Andriamihaja, M.; Blais, A.; Grauso, M.; Lepage, P.; Davila, A.M.; Viel, R.; Gaudichon, C.; Leclerc, M.; Blachier, F.; et al. Dietary Protein Intake Level Modulates Mucosal Healing and Mucosa-Adherent Microbiota in Mouse Model of Colitis. Nutrients 2019, 11, 514. [Google Scholar] [CrossRef]
- Blachier, F.; Beaumont, M.; Andriamihaja, M.; Davila, A.M.; Lan, A.; Grauso, M.; Armand, L.; Benamouzig, R.; Tomé, D. Changes in the luminal environment of the colonic epithelial cells and physiopathological consequences. Am. J. Pathol. 2017, 187, 476–486. [Google Scholar] [CrossRef]
- Faure, M; Mettraux, C; Moennoz, D; Godin, JP; Vuichoud, J; Rochat, F; Breuillé, D; Obled, C; Corthésy-Theulaz, I. Specific amino acids increase mucin synthesis and microbiota in dextran sulfate sodium-treated rats. J. Nutr. 2006, 136, 1558–1564. [Google Scholar] [CrossRef] [PubMed]
- Rieder, F.; Karrasch, T.; Ben-Horin, S.; Schirbel, A.; Ehehalt, R.; Wehkamp, J.; de Haar, C.; Velin, D.; Latella, G.; Scaldaferri, F.; et al. Results of the 2nd scientific workshop of the ECCO (III): basic mechanisms of intestinal healing. J. Crohns Colitis 2012, 6, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Kiesslich, R.; Goetz, M.; Angus, E.M.; Hu, Q.; Guan, Y.; Potten, C.; Allen, T.; Neurath, M.F.; Shroyer, N.F.; Montrose, M.H.; et al. Identification of epithelial gaps in human small and large intestine by confocal endomicroscopy. Gastroenterology 2007, 133, 1769–1778. [Google Scholar] [CrossRef]
- Watson, A.J.; Chu, S.; Sieck, L.; Gerasimenko, O.; Bullen, T.; Campbell, F.; McKenna, M.; Rose, T.; Montrose, M.H. Epithelial barrier function in vivo is sustained despite gaps in epithelial layers. Gastroenterology 2005, 129, 902–912. [Google Scholar] [CrossRef]
- Faure, M.; Moënnoz, D.; Montigon, F.; Mettraux, C.; Breuillé, D.; Ballèvre, O. Dietary threonine restriction specifically reduces intestinal mucin synthesis in rats. J. Nutr. 2005, 135, 486–491. [Google Scholar] [CrossRef]
- Liu, X.; Beaumont, M.; Walker, F.; Chaumontet, C.; Andriamihaja, M.; Matsumoto, H.; Khodorova, N.; Lan, A.; Gaudichon, C.; Benamouzig, R.; et al. Beneficial effects of an amino acid mixture on colonic mucosal healing in rats. Inflamm. Bowel Dis. 2013, 19, 2895–2905. [Google Scholar] [CrossRef]
- Blachier, F.; Boutry, C.; Bos, C.; Tomé, D. Metabolism and functions of L-glutamate in the epithelial cells of the small and large intestines. Am. J. Clin. Nutr. 2009, 90, 814S–821S. [Google Scholar] [CrossRef]
- Chakravarthi, S.; Jessop, C.E.; Bulleid, N.J. The role of glutathione in disulphide bond formation and endoplasmic-reticulum-generated oxidative stress. EMBO Rep. 2006, 7, 271–275. [Google Scholar] [CrossRef]
- Kemp, M.; Go, Y.M.; Jones, D.P. Nonequilibrium thermodynamics of thiol/disulfide redox systems: a perspective on redox systems biology. Free Radic. Biol. Med. 2008, 44, 921–937. [Google Scholar] [CrossRef]
- Muro, P.; Zhang, L.; Li, S.; Zhao, Z.; Jin, T.; Mao, F.; Mao, Z. The emerging role of oxidative stress in inflammatory bowel disease. Front. Endocrinol. (Lausanne) 2024, 15, 1390351. [Google Scholar] [CrossRef]
- Riedijk, M.A.; Stoll, B.; Chacko, S.; Schierbeek, H.; Sunehag, A.L.; van Goudoever, J.B.; Burrin, D.G. Methionine transmethylation and transsulfuration in the piglet gastrointestinal tract. Proc. Natl. Acad. Sci. U S A 2007, 104, 3408–3413. [Google Scholar] [CrossRef]
- Zhang, B.; Zhao, C.; Zhang, X.; Li, X.; Zhang, Y.; Liu, X.; Yin, J.; Li, X.; Wang, J.; Wang, S. An elemental diet enriched in amino acids alters the gut microbial community and prevents colonic mucus degradation in mice with colitis. mSystems 2022, 7, e0088322. [Google Scholar] [CrossRef] [PubMed]
- Jebeile, H.; Kelly, A.S.; O’Malley, G.; Baur, L.A. Obesity in children and adolescents: epidemiology, causes, assessment, and management. Lancet Diabetes Endocrinol. 2022, 10, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M. Obesity: global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef]
- Lee, K.X.; Quek, K.F.; Ramadas, A. Dietary and lifestyle risk factors of obesity among young adults: a scoping review of observational studies. Curr. Nutr. Rep. 2023, 12, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.M. On the causes of obesity and its treatment: The end of the beginning. Cell Metab. 2025, 37, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Yan, Y.; Qiu, X.; Lin, S.; Wen, J. Endoscopic bariatric surgery for adults with overweight and obesity: a systematic review and network meta-analysis. Int. J. Obes. (Lond) 2025, 49, 237–245. [Google Scholar] [CrossRef]
- Idris, I.; Anyiam, O. The latest evidence and guidance in lifestyle and surgical interventions to achieve weight loss in people with overweight or obesity. Diabetes Obes. Metab. 2025, 27, 20–34. [Google Scholar] [CrossRef]
- Cava, E.; Yeat, N.C.; Mittendorfer, B. Preserving healthy muscle during weight loss. Adv. Nutr. 2017, 8, 511–519. [Google Scholar]
- McCarthy, D.; Berg, A. Weight loss strategies and the risk of skeletal muscle mass loss. Nutrients 2021, 13, 2473. [Google Scholar] [CrossRef]
- Nuijten, M.A.H.; Eijsvogels, T.M.H.; Monpellier, V.M.; Janssen, I.M.C.; Hazebroek, E.J.; Hopman, M.T.E. The magnitude and progress of lean body mass, fat-free mass, and skeletal muscle mass loss following bariatric surgery: A systematic review and meta-analysis. Obes. Rev. 2022, 23, e13370. [Google Scholar] [PubMed]
- Sargeant, J.A.; Henson, J.; King, J.A.; Yates, T.; Khunti, K.; Davies, M.J. A review of the effects of glucagon-like peptide-1 receptor agonists and sodium-glucose cotransporter 2 inhibitors on lean body mass in humans. Endocrinol. Metab. (Seoul) 2019, 34, 247–262. [Google Scholar] [CrossRef] [PubMed]
- James, E.; Kelsey, R.J.; Sargeant, J.A.; Henson, J.; Yates, T.; Wilkinson, T.J. Nutritional supplementation to preserve healthy lean mass and function during periods of pharmacological and nonpharmacological-induced weight loss: protocol for a systematic review and meta-analysis. Health Sci. Rep. 2024, 7, e70219. [Google Scholar]
- Wycherley, T.P.; Moran, L.J.; Clifton, P.M.; Noakes, M.; Brinkworth, G.D. Effects of energy-restricted high-protein, low-fat compared with standard-protein, low-fat diets: a meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2012, 96, 1281–1298. [Google Scholar] [CrossRef]
- Leidy, H.J.; Clifton, P.M.; Astrup, A.; Wycherley, T.P.; >Westerterp-Plantenga, M.S.; Luscombe-Marsh, N.D.; Woods, S.C.; Mattes, R.D. The role of protein in weight loss and maintenance. Am. J. Clin. Nutr. 2015, 101, 1320S–1329S. [Google Scholar] [CrossRef]
- Villareal, D.T.; Chode, S.; Parimi, N.; Sinacore, D.R.; Hilton, T.; Armamento-Villareal, R.; Napoli, N.; Qualls, C.; Shah, K. Weight loss, exercise, or both and physical function in obese older adults. N. Engl. J. Med. 2011, 364, 1218–1229. [Google Scholar] [CrossRef]
- Frimel, T,N.; Sinacore, D.R.; Villareal, D.T. Exercise attenuates the weight-loss-induced reduction in muscle mass in frail obese older adults. Med. Sci. Sports Exerc. 2008, 40, 1213–1219. [Google Scholar] [CrossRef]
- Yoshimura, E.; Kumahara, H.; Tobina, T.; Matsuda, T.; Watabe, K.; Matono, S.; Ayabe, M.; Kiyonaga, A.; Anzai, K.; Higaki, Y.; et al. Aerobic exercise attenuates the loss of skeletal muscle during energy restriction in adults with visceral adiposity. Obes. Facts 2014, 7, 26–35. [Google Scholar] [CrossRef]
- Cannavaro, D.; Leva, F.; Caturano, A.; Berra, C.C.; Bonfrate, L.; Conte, C. Optimizing body composition during weight loss: the role of amino acid supplementation. Nutrients 2025, 17, 2000. [Google Scholar] [CrossRef]
- Coker, M.S.; Barati, Z.; Murphy, C.J.; Bateman, T.; Newcomer, B.R.; Wolfe, R.R.; Coker, R.H. Essential amino acid enriched meal replacement improves body composition and physical function in obese older adults: A randomized controlled trial. Clin. Nutr. ESPEN 2022, 51, 104–111. [Google Scholar] [CrossRef]
- Brunani, A.; Cancello, R.; Gobbi, M.; Lucchetti, E.; Di Guglielmo, G.; Maestrini, S.; Cattaldo, S.; Piterà, P.; Ruocco, C.; Milesi, A.; et al. Comparison of protein- or amino acid-based supplements in the rehabilitation of men with severe obesity: a randomized controlled pilot study. J. Clin. Med. 2023, 12, 4257. [Google Scholar] [CrossRef]
- Alkhulaifi, F.; Darkoh, C. Meal timing, meal frequency and metabolic syndrome. Nutrients 2022, 14, 1719. [Google Scholar] [CrossRef]
- Després, J.P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.L. A comprehensive definition for metabolic syndrome. Dis. Model Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Kassi, E.; Pervanidou, P.; Kaltsas, G.; Chrousos, G. Metabolic syndrome: definitions and controversies. BMC Med. 2011, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Peterseim, C.M.; Jabbour, K.; Kamath Mulki, A. Metabolic syndrome: an updated review on diagnosis and treatment for primary care clinicians. J. Prim. Care Community Health 2024, 15, 21501319241309168. [Google Scholar] [CrossRef]
- Miglani, N.; Bains, K. Interplay between proteins and metabolic syndrome-A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 2483–2496. [Google Scholar] [CrossRef] [PubMed]
- Lucotti, P.; Setola, E.; Monti, L.D.; Galluccio, E.; Costa, S.; Sandoli, E.P.; Fermo, I.; Rabaiotti, G.; Gatti, R.; Piatti, P. Beneficial effects of a long-term oral L-arginine treatment added to a hypocaloric diet and exercise training program in obese, insulin-resistant type 2 diabetic patients. Am. J. Physiol. 2006, 291, E906–E912. [Google Scholar] [CrossRef] [PubMed]
- Piatti, P.M.; Monti, L.D.; Valsecchi, G.; Magni, F.; Setola, E.; Marchesi, F.; Galli-Kienle, M.; Pozza, G.; Alberti, K.G. Long-term oral L-arginine administration improves peripheral and hepatic insulin sensitivity in type 2 diabetic patients. Diabetes Care 2001, 24, 875–880. [Google Scholar] [CrossRef]
- Casonatto, J.; Cavalari, J.V. A single dosage of l-arginine oral supplementation induced post-aerobic exercise hypotension in hypertensive patients. J. Diet. Suppl. 2023, 20, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Monti, L.D.; Setola, E.; Lucotti, P.C.; Marrocco-Trischitta, M.M.; Comola, M.; Galluccio, E.; Poggi, A.; Mammì, S.; Catapano, A.L.; Comi, G.; et al. Effect of a long-term oral l-arginine supplementation on glucose metabolism: a randomized, double-blind, placebo-controlled trial. Diabetes Obes. Metab. 2012, 14, 893–900. [Google Scholar] [CrossRef]
- Wascher, T.C.; Graier, W.F.; Dittrich, P.; Hussain, M.A.; Bahadori, B.; Wallner, S.; Toplak, H. Effects of low-dose L-arginine on insulin-mediated vasodilatation and insulin sensitivity. Eur. J. Clin. Invest. 1997, 27, 690–695. [Google Scholar] [CrossRef]
- Lyons, D. Impairment and restoration of nitric oxide-dependent vasodilation in cardiovascular disease. Int. J. Cardiol. 1997, 62, S101–S109. [Google Scholar] [CrossRef]
- Giles, T.D. Aspects of nitric oxide in health and disease: a focus on hypertension and cardiovascular disease. J. Clin. Hypertens. (Greenwich) 2006, 8, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Reginato, G.S.; de Jager, L.; Martins, A.B.; Lucchetti, B.F.C.; de Campos, B.H.; Lopes, F.N.C.; Araujo, E.J.A.; Zaia, C.T.B.V.; Pinge-Filho, P.; Martins-Pinge, M.C. Differential benefits of physical training associated or not with l-arginine supplementation in rats with metabolic syndrome: Evaluation of cardiovascular, autonomic and metabolic parameters. Physiol. Behav. 2023, 268, 114251. [Google Scholar] [CrossRef] [PubMed]
- Tagami, K.; Okuzawa, T.; Yoshida, K.; Mishima, R.; Obara, N.; Kunimatsu, A.; Koide, M.; Teranishi, T.; Itakura, K.; Ikeda, K.; et al. L-arginine ameliorates hypertension and cardiac mitochondrial abnormalities but not cardiac injury in male metabolic syndrome rats. Physiol. Rep. 2025, 13, e70183. [Google Scholar] [CrossRef]
- Alam, M.A.; Kauter, K.; Withers, K.; Sernia, C.; Brown, L. Chronic l-arginine treatment improves metabolic, cardiovascular and liver complications in diet-induced obesity in rats. Food Funct. 2013, 4, 83–91. [Google Scholar] [CrossRef]
- Kawano, T.; Nomura, M.; Nisikado, A.; Nakaya, Y.; Ito, S. Supplementation of L-arginine improves hypertension and lipid metabolism but not insulin resistance in diabetic rats. Life Sci. 2003, 73, 3017–3026. [Google Scholar] [CrossRef] [PubMed]
- Fiordelisi, A.; Cerasuolo, F.A.; Avvisato, R.; Buonaiuto, A.; Maisto, M.; Bianco, A.; D’Argenio, V.; Mone, P.; Perrino, C.; D’Apice, S.; et al. L-arginine supplementation as mitochondrial therapy in diabetic cardiomyopathy. Cardiovasc. Diabetol. 2024, 23, 450. [Google Scholar] [CrossRef]
- Felig, P.; Marliss, E.; Cahill, G.F., Jr. Plasma amino acid levels and insulin secretion in obesity. N. Engl. J. Med. 1969, 281, 811–816. [Google Scholar] [CrossRef] [PubMed]
- She, P.; Van Horn, C.; Reid, T.; Hutson, S.M.; Cooney, R.N.; Lynch, C.J. Obesity-related elevations in plasma leucine are associated with alterations in enzymes involved in branched-chain amino acid metabolism. Am. J. Physiol. 2007, 293, E1552–E1563. [Google Scholar] [CrossRef]
- Zhou, M.; Shao, J.; Wu, C.Y.; Shu, L.; Dong, W.; Liu, Y.; Chen, M.; Wynn, R.M.; Wang, J.; Wang, J.; et al. Targeting BCAA catabolism to treat obesity-associated insulin resistance. Diabetes 2019, 68, 1730–1746. [Google Scholar] [CrossRef]
- Giesbertz, P.; Padberg, I.; Rein, D.; Ecker, J.; Höfle, A.S.; Spanier, B.; Daniel, H. Metabolite profiling in plasma and tissues of ob/ob and db/db mice identifies novel markers of obesity and type 2 diabetes. Diabetologia 2015, 58, 2133–2143. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Holmes, M.V.; Davey Smith, G.; Ala-Korpela, M. Genetic support for a causal role of insulin resistance on circulating branched-chain amino acids and inflammation. Diabetes Care 2017, 40, 1779–1786. [Google Scholar] [CrossRef]
- Lee, J.; Vijayakumar, A.; White, P.J.; Xu, Y.; Ilkayeva, O.; Lynch, C.J.; Newgard, C.B.; Kahn, B.B. BCAA supplementation in mice with diet-induced obesity alters the metabolome without impairing glucose homeostasis. Endocrinology 2021, 162, bqab062. [Google Scholar] [CrossRef]
- Zhao, S.; Zhou, L.; Wang, Q.; Cao, J.H.; Chen, Y.; Wang, W.; Zhu, B.D.; Wei, Z.H.; Li, R.; Li, C.Y.; et al. Elevated branched-chain amino acid promotes atherosclerosis progression by enhancing mitochondrial-to-nuclear H(2)O(2)-disulfide HMGB1 in macrophages. Redox Biol. 2023, 62, 102696. [Google Scholar] [CrossRef]
- Pathak, K.; Zhao, Y.; Calton, E.K.; James, A.P.; Newsholme, P.; Sherriff, J.; Soares, M.J. The impact of leucine supplementation on body composition and glucose tolerance following energy restriction: an 8-week RCT in adults at risk of the metabolic syndrome. Eur. J. Clin. Nutr. 2024, 78, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Freudenberg, A.; Petzke, K.J.; Klaus, S. Comparison of high-protein diets and leucine supplementation in the prevention of metabolic syndrome and related disorders in mice. J. Nutr. Biochem. 2012, 23, 1524–1530. [Google Scholar] [CrossRef]
- Nishimura, J.; Masaki, T.; Arakawa, M.; Seike, M.; Yoshimatsu, H. Isoleucine prevents the accumulation of tissue triglycerides and upregulates the expression of PPARalpha and uncoupling protein in diet-induced obese mice. J. Nutr. 2010, 140, 496–500. [Google Scholar] [CrossRef]
- Díaz-Flores, M.; Cruz, M.; Duran-Reyes, G.; Munguia-Miranda, C.; Loza-Rodríguez, H.; Pulido-Casas, E.; Torres-Ramírez, N.; Gaja-Rodriguez, O.; Kumate, J.; Baiza-Gutman, L.A.; et al. Oral supplementation with glycine reduces oxidative stress in patients with metabolic syndrome, improving their systolic blood pressure. Can. J. Physiol. Pharmacol. 2013, 91, 855–860. [Google Scholar] [CrossRef] [PubMed]
- El-Hafidi, M.; Franco, M.; Ramírez, A.R.; Sosa, J.S.; Flores, J.A.P.; Acosta, O.L.; Salgado, M.C.; Cardoso-Saldaña, G. Glycine increases insulin sensitivity and glutathione biosynthesis and protects against oxidative stress in a model of sucrose-induced insulin resistance. Oxid. Med. Cell. Longev. 2018, 2018, 2101562. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.N.; Niu, Y.C.; Sun, X.W.; Li, Q.; Zhao, C.; Wang, C.; Guo, F.C.; Sun, C.H.; Li, Y. Histidine supplementation improves insulin resistance through suppressed inflammation in obese women with the metabolic syndrome: a randomised controlled trial. Diabetologia 2013, 56, 985–994. [Google Scholar] [CrossRef]
- Sun, Y.; Cai, L.; Yu, B.; Zhang, H.; Zhang, Z.; Xu, X.; Yu, Y.; Li, J.; Chen, C.; Xia, F. L.; et al. Phenylalanine promotes liver steatosis by inhibiting BNIP3-mediated mitophagy. Mol. Med. 2025, 31, 250. [Google Scholar] [CrossRef]
- Iuliano, S.; Poon, S.; Robbins, J.; Bui, M.; Wang, X.; De Groot, L.; Van Loan, M.; Zadeh, A.G.; Nguyen, T.; Seeman, E. Effect of dietary sources of calcium and protein on hip fractures and falls in older adults in residential care: cluster randomised controlled trial. B.M.J 2021, 375, n2364. [Google Scholar] [CrossRef]
- Cowan, R.W.; Seidlitz, E.P.; Singh, G. Glutamate signaling in healthy and diseased bone. Front. Endocrinol. (Lausanne) 2012, 3, 89. [Google Scholar] [CrossRef]
- Lin, T.H.; Yang, R.S.; Tang, C.H.; Wu, M.Y.; Fu, W.M. Regulation of the maturation of osteoblasts and osteoclastogenesis by glutamate. Eur. J. Pharmacol. 2008, 589, 37–44. [Google Scholar] [CrossRef]
- Xie, W.; Dolder, S.; Siegrist, M.; Wetterwald, A.; Hofstetter, W. Glutamate receptor agonists and glutamate transporter antagonists regulate differentiation of osteoblast lineage cells. Calcif. Tissue Int. 2016, 99, 142–54. [Google Scholar] [CrossRef]
- Fatokun, A.A.; Stone, T.W.; Smith, R.A. Hydrogen peroxide-induced oxidative stress in MC3T3-E1 cells: The effects of glutamate and protection by purines. Bone 2006, 39, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Hinoi, E.; Takarada, T.; Uno, K.; Inoue, M.; Murafuji, Y.; Yoneda, Y. Glutamate suppresses osteoclastogenesis through the cystine/glutamate antiporter. Am. J. Pathol. 2007, 170, 1277–1290. [Google Scholar] [CrossRef] [PubMed]
- Blais, A.; Rochefort, G.Y.; Moreau, M.; Calvez, J.; Wu, X.; Matsumoto, H.; Blachier, F. Monosodium glutamate supplementation improves bone status in mice under moderate protein restriction. J.B.M.R. Plus 2019, 3, e10224. [Google Scholar] [CrossRef]
- Reeds, P.J.; Burrin, D.G.; Jahoor, F.; Wykes, L.; Henry, J.; Frazer, E.M. Enteral glutamate is almost completely metabolized in first pass by the gastrointestinal tract of infant pigs. Am. J. Physiol. 1996, 270, E413–E418. [Google Scholar] [CrossRef] [PubMed]
- Janeczko, M.J.; Stoll, B.; Chang, X.; Guan, X.; Burrin, D.G. Extensive gut metabolism limits the intestinal absorption of excessive supplemental dietary glutamate loads in infant pigs. J. Nutr. 2007, 137, 2384–2390. [Google Scholar] [CrossRef]
- Gorres, K.L.; Raines, R.T. Prolyl 4-hydroxylase. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 106–124. [Google Scholar] [CrossRef]
- Albaugh, V.L.; Mukherjee, K.; Barbul, A. Proline precursors and collagen synthesis: biochemical challenges of nutrient supplementation and wound healing. J. Nutr. 2017, 147, 2011–2017. [Google Scholar] [CrossRef]
- Murtas, S.; Reggiardo, G.; Contu, R.; Cadeddu, M.; Secci, R.; Putzu, P.; Mocco, C.; Leoni, M.; Gigante, Maria V.; Marras, C.; et al. Replacement of the massive amino acid losses induced by hemodialysis: A new treatment option proposal for a largely underestimated issue. Clin. Nutr. ESPEN 2024, 63, 354–363. [Google Scholar] [CrossRef]
- Siramolpiwat, S.; Limthanetkul, N.; Pornthisarn, B.; Vilaichone, R.K.; Chonprasertsuk, S.; Bhanthumkomol, P.; Nunanan, P.; Issariyakulkarn, N. Branched-chain amino acids supplementation improves liver frailty index in frail compensated cirrhotic patients: a randomized controlled trial. BMC Gastroenterol. 2023, 23, 154. [Google Scholar] [CrossRef]
- Liu, C.; Meng, Q.; Zu, C.; Wei, Y.; Su, X.; Zhang, Y.; He, P.; Zhou, C.; Liu, M.; Ye, Z. Dietary low- and high-quality carbohydrate intake and cognitive decline: A prospective cohort study in older adults. Clin. Nutr. 2023, 42, 1322–1329. [Google Scholar] [CrossRef]
- Yeh, T.S.; Yuan, C.; Ascherio, A.; Rosner, B.A.; Blacker, D.; Willett, W.C. Long-term dietary protein intake and subjective cognitive decline in US men and women. Am. J. Clin. Nutr. 2022, 115, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Hill, E.; Li, Y.; He, M. Energy and macronutrient intakes at breakfast and cognitive declines in community-dwelling older adults: a 9-year follow-up cohort study. Am. J. Clin. Nutr. 2021, 113, 1093–1103. [Google Scholar] [CrossRef]
- Ragni, M.; Fenaroli, F.; Ruocco, C.; Segala, A.; D’Antona, G.; Nisoli, E.; Valerio, A. A balanced formula of essential amino acids promotes brain mitochondrial biogenesis and protects neurons from ischemic insult. Front. Neurosci. 2023, 17, 1197208. [Google Scholar] [CrossRef] [PubMed]
- Jaroudi, W.; Garami, J.; Garrido, S.; Hornberger, M.; Keri, S.; Moustafa, AA. Factors underlying cognitive decline in old age and Alzheimer’s disease: the role of the hippocampus. Rev. Neurosci. 2017, 28, 705–714. [Google Scholar] [CrossRef]
- Pronier, É.; Morici, J.F.; Girardeau, G. The role of the hippocampus in the consolidation of emotional memories during sleep. Trends Neurosci. 2023, 46, 912–925. [Google Scholar] [CrossRef]
- Gheller, M.E.; Vermeylen, F.; Handzlik, M.K.; Gheller, B.J.; Bender, E.; Metallo, C.; Aydemir, T.B.; Smriga, M.; Thalacker-Mercer, AE. Tolerance to graded dosages of histidine supplementation in healthy human adults. Am. J. Clin. Nutr. 2020, 112, 1358–1367. [Google Scholar] [CrossRef]
- Pencharz, P.B.; Elango, R.; Ball, R.O. Determination of the tolerable upper intake level of leucine in adult men. J. Nutr. 2012, 142, 2220S–2224S. [Google Scholar] [CrossRef]
- Elango, R.; Rasmussen, B.; Madden, K. Safety and tolerability of leucine supplementation in elderly men. J Nutr. 2016, 146, 2630S–2634S. [Google Scholar] [CrossRef] [PubMed]
- Hayamizu, K.; Oshima, I.; Fukuda, Z.; Kuramochi, Y.; Nagai, Y.; Izumo, N.; Nakano, M. Safety assessment of L-lysine oral intake: a systematic review. Amino Acids 2019, 51, 647–659. [Google Scholar] [CrossRef]
- Deutz, N.E.; Simbo, S.Y.; Ligthart-Melis, G.C.; Cynober, L.; Smriga, M.; Engelen, MP. Tolerance to increased supplemented dietary intakes of methionine in healthy older adults. Am. J. Clin. Nutr. 2017, 106, 675–683. [Google Scholar] [CrossRef]
- Miura, N.; Matsumoto, H.; Cynober, L.; Stover, P.J.; Elango, R.; Kadowaki, M.; Bier, D.M.; Smriga, M. Subchronic tolerance trials of graded oral supplementation with phenylalanine or serine in healthy adults. Nutrients 2021, 13, 1976. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Miura, N.; Naito, M.; Elango, R. Evaluation of safe utilization of L-threonine for supplementation in healthy adults: a randomized double blind controlled trial. Amino Acids 2025, 57, 30. [Google Scholar] [CrossRef]
- Boirie, Y.; Dangin, M.; Gachon, P.; Vasson, M.P.; Maubois, J.L.; Beaufrère, B. Slow and fast dietary proteins differently modulate postprandial protein accretion. Proc. Natl. Acad. Sci. U S A 1997, 94, 14930–14935. [Google Scholar] [CrossRef]
- Luttikhold, J.; van Norren, K.; Buijs, N.; Ankersmit, M.; Heijboer, A.C.; Gootjes, J.; Rijna, H.; van Leeuwen, P.A.; van Loon, L.J. Jejunal casein feeding is followed by more rapid protein digestion and amino acid absorption when compared with gastric feeding in healthy young men. J. Nutr. 2015, 145, 2033–2038. [Google Scholar] [CrossRef]
- Hiratsuka, C.; Fukuwatari, T.; Sano, M.; Saito, K.; Sasaki, S.; Shibata, K. Supplementing healthy women with up to 5.0 g/d of L-tryptophan has no adverse effects. J. Nutr. 2013, 143, 859–866. [Google Scholar] [CrossRef] [PubMed]
- McNeal, C.J.; Meininger, C.J.; Wilborn, C.D.; Tekwe, C.D.; Wu, G. Safety of dietary supplementation with arginine in adult humans. Amino Acids 2018, 50, 1215–1229. [Google Scholar] [CrossRef]
- Shao, A.; Hathcock, J.N. Risk assessment for the amino acids taurine, L-glutamine and L-arginine. Regul. Toxicol. Pharmacol. 2008, 50, 376–399. [Google Scholar] [CrossRef] [PubMed]
- Kuramochi, Y.; Murata, M.; Sumino, A.; Sone, H.; Hayamizu, K. Safety assessment of L-Arg oral intake in healthy subjects: a systematic review of randomized control trials. Amino Acids 2023, 55, 1949–1964. [Google Scholar] [CrossRef]
- Bucci, L.R.; Hickson, J.F., Jr.; Wolinsky, I.; Pivarnik, J.M. Ornithine supplementation and insulin release in bodybuilders. Int. J. Sport Nutr. 1992, 2, 287–91. [Google Scholar] [CrossRef]
- Papadia, C.; Osowska, S.; Cynober, L.; Forbes, A. Citrulline in health and disease. Review on human studies. Clin. Nutr. 2018, 37, 1823–1828. [Google Scholar] [CrossRef] [PubMed]
- Dillon, K.N.; Kang, Y.; Maharaj, A.; Martinez, M.A.; Fischer, S.M.; Figueroa, A. L-Citrulline supplementation attenuates aortic pressure and pressure waves during metaboreflex activation in postmenopausal women. Br. J. Nutr. 2024, 131, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Miura, N.; Morishita, K.; Yasuda, T.; Akiduki, S.; Matsumoto, H. Subchronic tolerance trials of graded oral supplementation with ornithine hydrochloride or citrulline in healthy adults. Amino Acids 2023, 55, 299–311. [Google Scholar] [CrossRef]
- Yang, H.; Kuramochi, Y.; Sato, S.; Sakai, R.; Hayamizu, K. Safety assessment of L-ornithine oral intake in healthy subjects: a systematic review. Amino Acids 2025, 57, 23. [Google Scholar] [CrossRef]
- Tsubuku, S.; Hatayama, K.; Katsumata, T.; Nishimura, N.; Mawatari, K.; Smriga, M.; Kimura, T. Thirteen-week oral toxicity study of branched-chain amino acids in rats. Int. J. Toxicol. 2004, 23, 119–126. [Google Scholar] [CrossRef]
- Shibui, Y.; Manabe, Y.; Kodama, T.; Gonsho, A. 13-week repeated dose toxicity study of l-tyrosine in rats by daily oral administration. Food Chem. Toxicol. 2016, 87, 55–64. [Google Scholar] [CrossRef]
- Shibui, Y.; Matsumoto, H.; Masuzawa, Y.; Ohishi, T.; Fukuwatari, T.; Shibata, K.; Sakai, R. Thirteen week toxicity study of dietary l-tryptophan in rats with a recovery period of 5 weeks. J. Appl. Toxicol. 2018, 38, 552–563. [Google Scholar] [CrossRef]
- Mailliard, M.E.; Stevens, B.R.; Mann, G.E. Amino acid transport by small intestinal, hepatic, and pancreatic epithelia. Gastroenterology 1995, 108, 888–910. [Google Scholar] [CrossRef] [PubMed]
- Munck, L.K.; Munck, B.G. The rabbit jejunal ‘imino carrier’ and the ileal ‘imino acid carrier’ describe the same epithelial function. Biochim. Biophys. Acta 1992, 1116, 91–96. [Google Scholar] [CrossRef]
- Soultoukis, G.A.; Partridge, L. Dietary protein, metabolism, and aging. Annu. Rev. Biochem. 2016, 85, 5–34. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of the implication of amino acids as precursors for numerous compounds, as energy substrates, and as modulators of cell signaling. The compounds produced from amino acids include macromolecules and smaller molecules such as bioactive peptides, neurotransmitters, bioactive metabolites, ATP, glucose, and phospholipids. Specific amino acids are used as energy substrates in different cells within tissues. Some amino acids are involved in different physiological functions through cell signaling, epigenetic modifications, and utilization in metabolic pathways involved in nitrogenous waste disposal.
Figure 1.
Schematic representation of the implication of amino acids as precursors for numerous compounds, as energy substrates, and as modulators of cell signaling. The compounds produced from amino acids include macromolecules and smaller molecules such as bioactive peptides, neurotransmitters, bioactive metabolites, ATP, glucose, and phospholipids. Specific amino acids are used as energy substrates in different cells within tissues. Some amino acids are involved in different physiological functions through cell signaling, epigenetic modifications, and utilization in metabolic pathways involved in nitrogenous waste disposal.
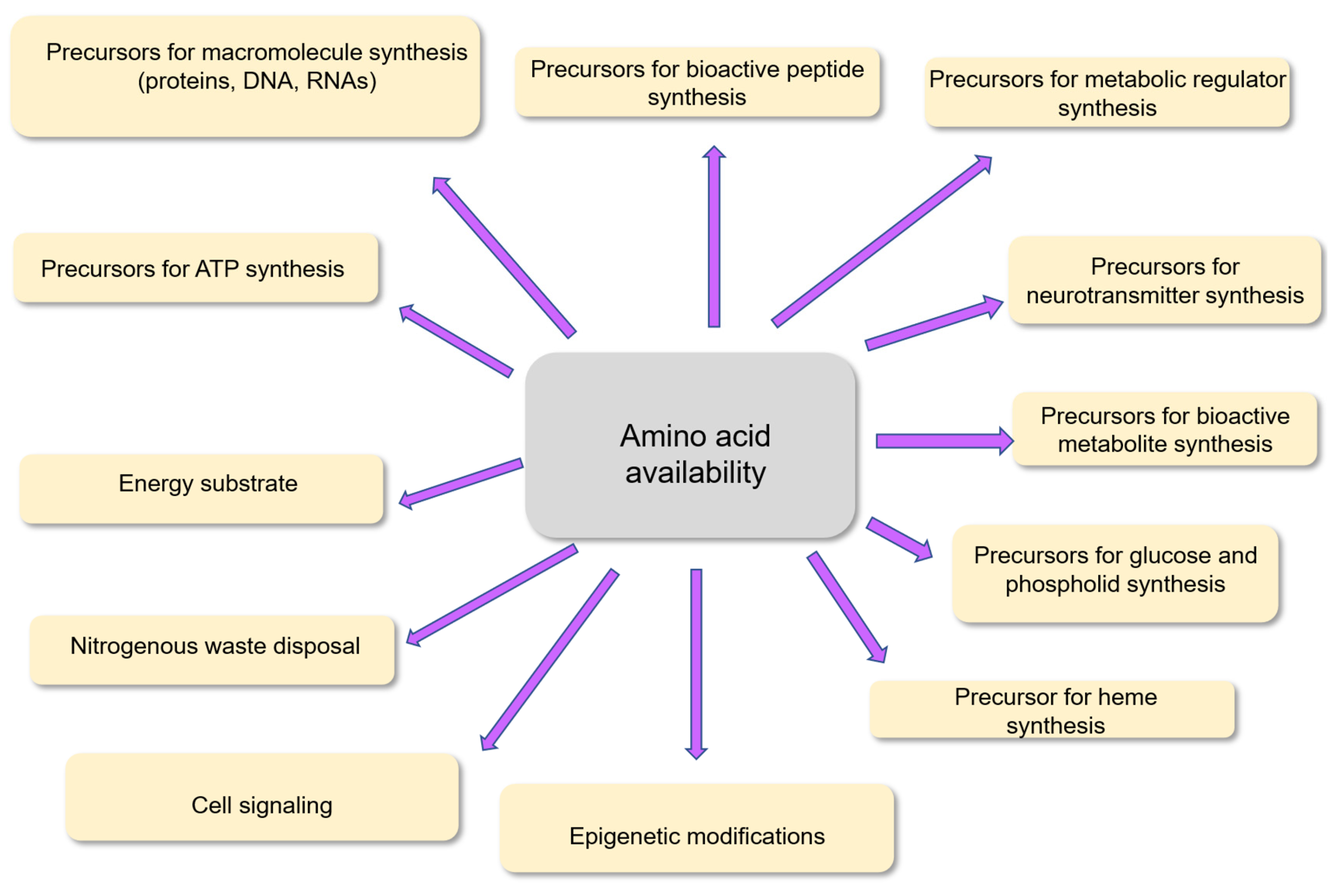
Figure 2.
Schematic representation of the supply and utilization of amino acids in the human body.
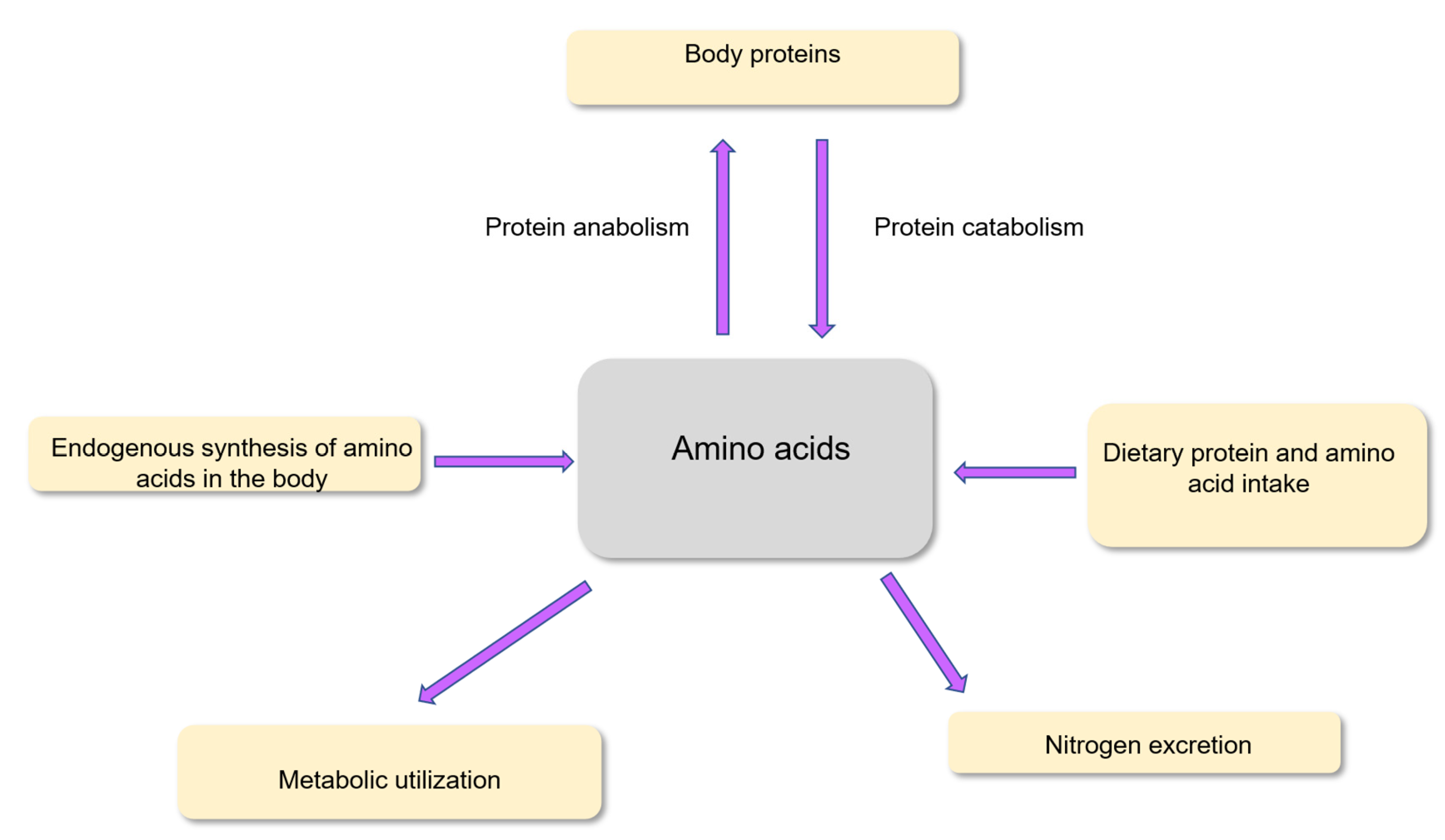
Figure 3.
Paradigm for amino acid supplementation. The amino acid requirements in different physiological, pre-pathological, and pathophysiological situations are used to determine the nutritional recommendations for the intake of the different amino acids. When the usual consumption of each amino acid is lower than the recommendation, amino acid supplementation can be envisaged in the limits of the No-Observed-Adverse-Effects Levels (NOAELs). The doses of amino acids in supplements, as well as the NOAEL values are determined in clinical trials. Experimental studies are useful for the identification of the mechanisms underlying the effects of amino acid supplementation. Amino acid supplementation can be made with individual amino acids or with mixtures of amino acids. Usual consumption of amino acids is determined from intake from dietary proteins and from intake of dietary amino acids in their free form.
Figure 3.
Paradigm for amino acid supplementation. The amino acid requirements in different physiological, pre-pathological, and pathophysiological situations are used to determine the nutritional recommendations for the intake of the different amino acids. When the usual consumption of each amino acid is lower than the recommendation, amino acid supplementation can be envisaged in the limits of the No-Observed-Adverse-Effects Levels (NOAELs). The doses of amino acids in supplements, as well as the NOAEL values are determined in clinical trials. Experimental studies are useful for the identification of the mechanisms underlying the effects of amino acid supplementation. Amino acid supplementation can be made with individual amino acids or with mixtures of amino acids. Usual consumption of amino acids is determined from intake from dietary proteins and from intake of dietary amino acids in their free form.
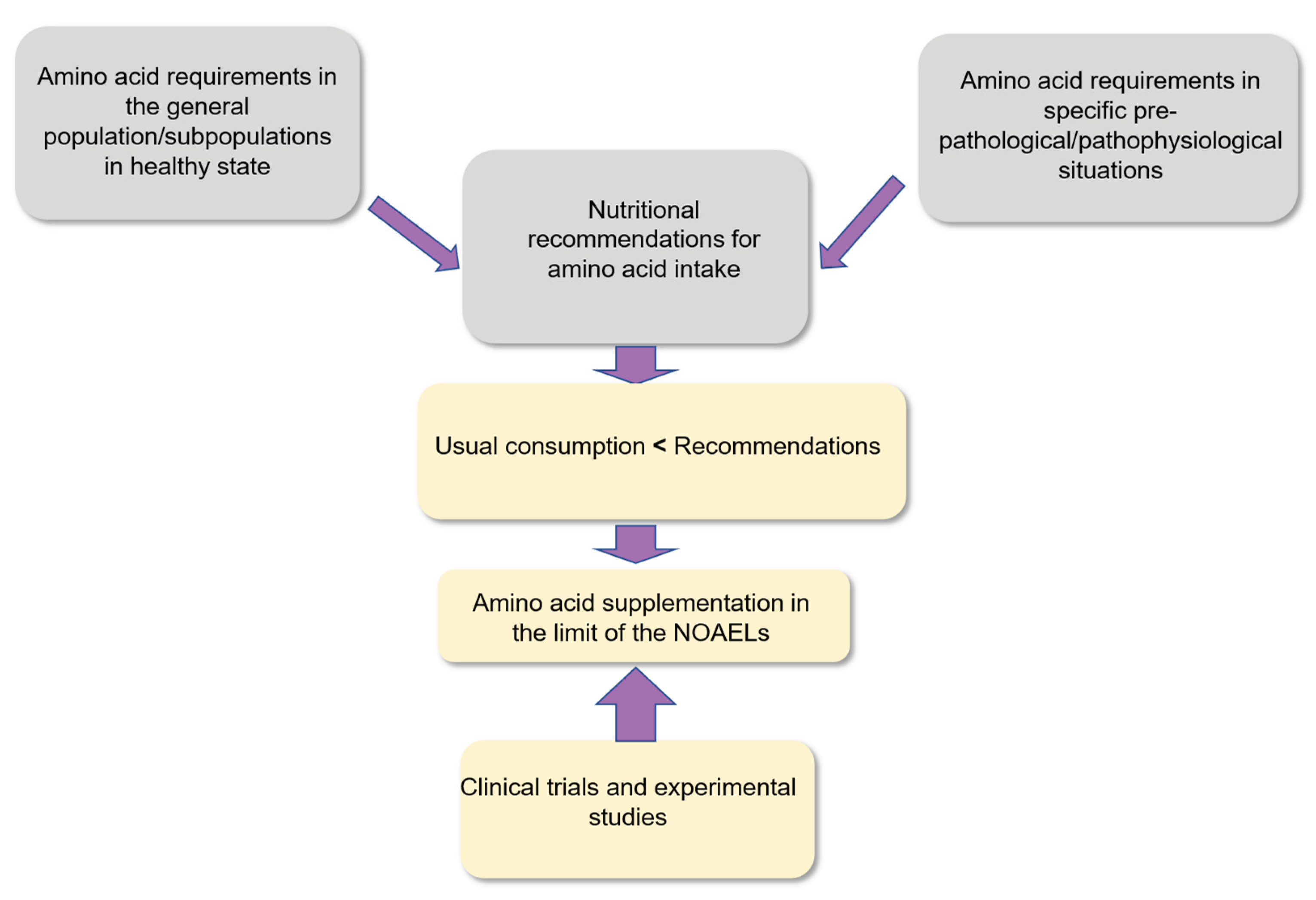

Table 1.
Requirement of indispensable amino acids in adults and usual consumption in North America and Europe.
Table 1.
Requirement of indispensable amino acids in adults and usual consumption in North America and Europe.
| Indispensable AA Requirements Usual Consumption |
| (mg/kg body weight/day) |
| FNB/IOM1 WHO2 AFSSA3 FNB/IOM4 NMCD5 |
| Histidine 11 10 11 31 28 |
| Isoleucine 15 20 18 51 45 |
| Leucine 34 39 39 87 80 |
| Lysine 31 30 30 75 71 |
| Methionine 10a 10 15b 25 23 |
| Phenylalanine 27c 25c 27d 49 46 |
| Threonine 16 15 16 43 40 |
| Tryptophan 4 4 4 13 11 |
| Valine 19 26 18 57 54 |
Note. Requirements and usual consumption are given in mg amino acid per kg body weight per day. 1 Values from the Food and Nutrition Board/ Institute of Medicine (FNB/IOM) 2005 report [41]; 2 Values from the World Health Organization (WHO) 2007 report [42]; 3 Values from the French Agency for Food Safety (AFSSA) 2007 report [33];4 Usual consumption (UC) from food and supplements in North America for all life stage groups 5 UC values for European adults from Iguacel et al. Nutr. Metab. Cardiovasc. Dis. (NMCD) 2022 [44]. a Methionine requirement in the presence of an excess of dietary cysteine; b Sulfur-containing amino acids; c Phenylalanine and tyrosine, d Aromatic amino acids. AA are amino acids.
Table 2.
No-observed-adverse-effect-level (NOAEL) values for amino acids in supplements in humans.
| Amino acids NOAEL values (g/day) References |
| Histidine 8.0 Gheller et al. Am. J. Clin. Nutr. 2020 |
| Lysine 6.0 Hayamizu et al. Amino Acids 2019 |
| Methionine 3.2 Deutz et al. Am. J. Clin. Nutr. 2017 |
| Phenylalanine 12.0 Miura et al. Nutrients 2021 |
| Threonine 12.0 Matsumoto et al. Amino Acids 2025 |
| Tryptophan 5.0 Hiratsuka et al. J. Nutr. 2013 |
| Arginine 7.5-20 Kuramochi et al. Amino Acids 2023, Shao et al. Regul. Toxicol. Pharmacol. 2008 |
| Serine 12.0 Miura et al. Nutrients 2021 |
| Ornithine 12.0 Miura et al. Amino Acids 2023, Yang et al. Amino Acids 2025 |
| Citrulline 24.0 Miura et al. Amino Acids 2023 |
Note: The NOAEL values for amino acids in supplements are given for the 8 proteinogenic amino acids and for the two non-proteinogenic amino acids (ornithine and citrulline) for which values are available. The NOAEL value 7.5 g/day for arginine in supplements originate from the recording of light gastrointestinal symptoms recorded in some volunteers.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



