
Submitted:
03 December 2025
Posted:
05 December 2025
You are already at the latest version
Abstract
This study evaluated the use of cholesterol-loaded cyclodextrin (CLC) to mitigate cryo-damage, aiming to enhance sperm cryo-survivability and reduce cryo-capacitation. Different CLC levels were tested, and post-thaw motility was higher (p < 0.05) at 1.5-2 mg/mL CLC added to extended semen containing 120 million sperm/mL. Subsequent investigations compared untreated and 2 mg/mL CLC-treated bovine sperm (n=5) and assessed membrane and acrosome integrity, mitochondrial potential, and capacitation status using flow cytometry. Moreover, this study is the first to quantify the post-thaw binding ability of CLC-treated bovine sperm and evaluate their capacitation window using an IVF approach. CLC-treated sperm showed no differences in flow cytometric results. CLC treatment significantly (p < 0.0001) increased the number of viable bound sperm cells to oviduct cells (1.65 × 10⁶ µm²) from 118 ± 15 to 267 ± 15 after three hours, and from 70 ± 15 to 127 ± 15 after seven hours (p < 0.05) of coincubation. When equal numbers of motile sperm were incubated with oocytes, CLC-treated sperm showed penetration and pronuclei formation rates comparable to the control, indicating a similar capacitation window. In conclusion, CLC addition significantly improved post-thaw sperm motility and binding ability, while maintaining sperm capacitation and fertilizing ability.
Keywords:
bovine sperm
; cryopreservation
; cholesterol-loaded cyclodextrin
; capacitation
; cryocapacitation
; flow cytometry
; oviduct culture
; in vitro fertilization
; sperm motility
; sperm membrane
1. Introduction
Frozen sperm cells are exposed to various stressors during processing steps, which include handling, diluting, cooling, cryopreservation, and thawing. These stressors involve alterations in incubation media, ionic imbalance, hyper-osmolarity, dehydration, energy deprivation, membrane phase transition, ice crystal formation, and temperature fluctuations during freezing and thawing [1,2,3,4]. As a result, 40- 50% of the sperm in a semen sample that freezes successfully even under optimum conditions do not survive cryopreservation [5,6]. Additionally, these stressors significantly impact the capacitation status of sperm cells, leading to a wide variety of changes known as capacitation-like changes, also referred to as cryo-capacitation [5,7,8,9]. The plasma membrane and acrosome are the most affected regions by cryopreservation [2,3] because they’re directly exposed to the cryo-environment [10]. During cryopreservation, the sperm membrane undergoes cholesterol and polyunsaturated fatty acids efflux, which leads to changes in membrane permeability to water, ions, and cryoprotectants, besides redistributing phospholipids and reordering lipids within the sperm plasma membrane [1,9,11]. Other cryo-capacitation features include higher cAMP levels, loss of surface proteins and receptors, and an altered sperm proteome [12,13,14]. Furthermore, damage to the mitochondrial membrane may increase reactive oxygen species concentrations, resulting in the generation of free radicals and ultimately causing necrosis in sperm cells [12,15]. These changes, along with membrane damage and acrosome reaction, speed up sperm death if fertilization does not occur [12,16].
Enhancing sperm survivability during cryopreservation could increase the number of insemination doses per ejaculate, especially from bulls whose sperm have a poor ability to survive freezing or lower daily sperm output, such as younger bulls [17,18,19]. Addressing concerns related to sperm quality, artificial insemination protocols, and semen processing are essential [20]. Therefore, innovative cryopreservation strategies targeting enhancing both the quantity and quality of surviving sperm after thawing by reducing cryo-capacitation rates [7] were emphasized [21].
The goal of this study was to improve cryo-survivability and reduce the cryo-capacitation rate of conventional semen by increasing the cholesterol content of sperm plasma membranes. The approach modifies the sperm plasma membrane [22], the most affected part during cryopreservation [2,3]. It enhances membrane protection, helping prevent cold shock and cryoinjury, and might slow down the capacitation process [21], which should normally occur after insemination.
Cholesterol is a hydrophobic molecule and is insoluble in water. Thus, cyclodextrin can be utilized to solubilize cholesterol through the formation of water-soluble inclusion complexes. The incorporation of a methyl or hydroxypropyl group enhances cyclodextrin’s ability to dissolve hydrophobic molecules in water. Cyclodextrin alone can withdraw cholesterol from sperm, leading to cholesterol efflux. However, when loaded with cholesterol, cyclodextrin can deliver cholesterol to sperm cells [21,23]. The addition of cholesterol stabilizes the sperm plasma membrane, enhances thermo-resistance, compensates for the cholesterol efflux during cryopreservation, and can improve post-thaw sperm motility and viability [21,24]. These effects collectively contribute to alleviating cryopreservation damage to the sperm cells.
The impact of cholesterol on capacitation status remains controversial; however, many scientists have reported lower cryo-capacitation levels for cholesterol-treated sperm cells, as highlighted by Mocé et al. [21]. During cryopreservation, both cholesterol-loaded cyclodextrin (CLC) treated and untreated sperm cell undergoes cholesterol loss, a remarkable cryo-capacitation feature [21,22]. Thus, cryo-capacitation may still influence both CLC-treated and untreated cells. For instance, the addition of cholesterol to fresh bull sperm can set back capacitation, while both capacitation and acrosome reaction occur at the same rate for cryopreserved treated and untreated sperm cells [22]. Additionally, treating dog sperm with cholesterol improved post-thaw motility but displayed a similar capacitation rate [25]. Capacitation is a broad and complex process that encompasses numerous biochemical maturational changes at the levels of the sperm head, middle piece, and tail, which are not yet fully understood [12,16]. Thus, the evaluation of capacitation and, consequently, cryo-capacitation varies significantly from laboratory to laboratory due to the numerous approaches used for capacitation induction [22]. Additionally, the various methods to detect capacitation status [8,12] compound the issue and contribute to controversial reports.
Flow cytometry analyses offer a more sensitive, reliable, and accurate approach in evaluating cryo-capacitation and fertile potential when compared to fluorescence microscopy techniques [26,27], because it utilizes large numbers of sperm cells. Flow cytometry analyses in the present study include simultaneous assessment of sperm viability to indicate cryo-survivability using propidium iodide (PI) and acrosome integrity using FITC-PNA [28,29]. Sperm cells with intact membranes are not permeable to PI and thus don’t stain.
Merocyanine 540 (M540) serves as an indicator for assessing membrane fluidity and capacitation at the membrane level. It also helps determine whether cholesterol modification to the sperm plasma membrane has occurred or not. Combining Yo-Pro-1 (YP) with M540 can enable us to simultaneously track membrane changes and viability. Using a combination of M540 and YP, along with Hoechst (to gate out non-sperm events), was found to be a robust approach in evaluating membrane stability and can detect early stages of capacitation in bull sperm [30]. YP infiltrates cells when they undergo membrane destabilization due to increased permeability of pannexin-gated channels in the cell membrane. This occurs before a complete loss of membrane integrity, allowing early detection of cell death when compared to PI [31,32]. Therefore, YP presents a good probe to track membrane changes due to cryoinjuries. During capacitation, sperm cells lose coating glycoproteins, leading to increased membrane fluidity and membrane destabilization (impacting sperm longevity). These are important early events in the capacitation process [33]. These alterations result in intercalating more M540 by the sperm membrane [32,34]. Thus, a combination of YP and M540 can simultaneously identify the early capacitation alterations in lipid packaging and distribution within the sperm plasma membrane.
Other flow cytometric analyses in this study include evaluating post-thaw intracellular calcium levels to track capacitation progress. Capacitated cells have a higher ability to uptake calcium from the surrounding media, accumulating it into their acrosome and cytoplasm [16,35] and undergo acrosome reaction when induced [22,36]. Calcium uptake is essential during the first 2 hours of capacitation induction by heparin [35]. Mitochondrial potential is an important assay that can indicate the motility and activity status of sperm cells. When sperm cells are loaded with JC-1, they exhibit dual orange and green fluorescence; the ratio between both reflects the mitochondrial activity. Active mitochondria aggregate JC-1 and fluoresce orange. While less functional ones accumulate lower concentrations of JC-1, which stay in the monomer state and fluoresce green [27].
The binding of sperm cells to oviduct epithelial cells in the isthmus region of the female reproductive tract is vital for maintaining sperm viability and preventing premature capacitation, as reviewed by Suarez [37]. Cryopreservation may lead to surface alterations that may impact the ability of sperm cells to bind to oviduct cells [38]. Additionally, uncapacitated sperm cells display better ability to interact with oviduct cells [39]. Therefore, binding to oviduct cells can also give us an idea about the capacitation status of sperm cells. The more the sperm cells are capacitated, the less their ability to bind and interact with oviduct cells. Binding to oviductal cells is essential to sperm survival and for completing capacitation at the appropriate time in the cow reproductive tract, which ensures a better chance of sperm cells fertilizing an oocyte. Capacitation is considered complete when sperm penetrates a zona-intact oocyte, fuses with the oolemma, and decondenses in the oocyte cytoplasm [16,40]. Thus, the capacitation window can be tracked based on the penetration time when sperm cells are induced to capacitate in vitro [40,41]. This is important to determine any delays in capacitation due to the CLC treatment.
The objectives of this study were to examine the cryo-survivability traits (motility and viability) and how CLC affects the capacitation status and physiological characteristics of sperm cells. This is the first time for evaluating sperm cells’ ability to bind and interact with oviduct cells and the time of fertilization of CLC-treated sperm cells using an in vitro fertilization (IVF) approach. Additionally, flow cytometric analyses were utilized to accurately assess the progress of capacitation in treated sperm cells.
2. Materials and Methods
Bulls were housed in a commercial facility that collects semen from bulls at ABS (Animal Breeding Services) Global in Deforest, WI, USA. The facility follows the USDA guidelines for animal care, and bulls are under routine veterinary supervision. The Chemicals were procured from Sigma-Aldrich (St. Louis, MO, USA) or ThermoFisher (Pittsburgh, PA, USA) unless otherwise specified.
2.1. Cholesterol-Loaded Cyclodextrin Preparation
The CLC powder was prepared following the Purdy and Graham procedure [22,24,42,43]. Separate solutions of cholesterol and cyclodextrin were prepared by dissolving 0.2 g of cholesterol in 1 mL of chloroform and 1 g of methyl-β-cyclodextrin in 2 mL of methanol, respectively, in two glass tubes. Subsequently, 0.450 mL of the cholesterol solution was added to the 2 mL cyclodextrin solution, and the resulting mixture was vortexed until clear. The mixture was then poured into a glass petri dish and incubated at 37°C for at least 24 hours to remove solvents and yield CLC crystals [23,43]. The crystals were harvested and stored on the shelf at room temperature (around 22°C). The CLC working solution was prepared by adding 0.05 g of CLC crystals to 1 mL of Tris diluent (Triladyl, Minitube USA, 13500/0250, Verona, WI, USA) at 37°C, then vortexing for 1 minute. Although dissolving the crystals in this manner results in a chunky solution, it is considered normal and still effective, as per personal communication with Professor Graham, the founder of this method. To enhance solubilization, the working solution was sonicated for 2 minutes [43]. According to the experimental design, varying amounts of CLC working solution were added to treat sperm cells.
2.2. Semen Collection, Treatment, and Cryopreservation
Semen from four or five different bulls, depending on the experiment (detailed in the Experimental Design below), was obtained from ABS Global, DeForest, WI. Upon arrival, the concentration was determined with a spectrophotometer. The sample was then diluted to 120 × 106 sperm cells per mL before the addition of CLC [22,24]. To prevent interference with CLC, egg yolk was not added until after treating sperm cells with CLC. Therefore, the previous dilution step was carried out using an egg yolk-free Tris extender (Triladyl). The sample was next divided into aliquots, designating one as the control (non-CLC treatment) and others for various levels of CLC depending on the experimental design. Following this, the samples underwent a 15-minute incubation at 22°C as described by Purdy and Graham [24]. Post-incubation, each aliquot was diluted with a 20% egg yolk-Tris extender (Triladyl) to a final concentration of 30 million sperm/mL. The samples were then gradually cooled over 2 hours to 5°C in a cold room. Subsequently, cooled semen underwent a 4-hour equilibration period during which it was packaged into 0.5-mL straws. Post-equilibration, straws were exposed to liquid nitrogen vapor at 4 cm above the liquid nitrogen for 10 minutes. Then straws were plunged into liquid nitrogen for storage at -196°C until thawing.
2.3. Motion Analyses
The motion analyses, including total and progressive motilities of frozen-thawed sperm, were determined using a computer-aided sperm analysis (HTM-IVOS CASA system, Hamilton-Thorn Research, Bedford, MA, USA), equipped with a warming stage at 37°C. The CASA system settings were as described by Mocé and Graham [42], which include analyzing 30 frames per second, minimum contrast = 50, minimum average path velocity = 25 µm/s, straightness = 80%, and nonmotile head size = 5. Semen straws were thawed at 37°C for 30 seconds. A minimum of ten fields with at least 500 sperm per sample were evaluated at time points 0, 30, 60, and maybe 120 minutes based on each experimental design as explained below, then averages were calculated.
2.4. Flow Cytometric Analyses
The flow cytometry analyses were conducted at the UWCCC Flow Cytometry Laboratory, UW-Madison, using a BD LSRII flow cytometer (BD Biosciences, San Jose, CA, USA). Analyses include assessments of sperm viability, membrane stability, acrosome integrity, intracellular calcium, and mitochondrial potential. Semen straws were thawed and diluted to 1 × 106 sperm/mL with Tris diluent (Triladyl) except for calcium analyses, for which Bovine Gamete Media 3, known as BGM3 [16], was used to provide a calcium source. Flow cytometric assays involved evaluating 10,000 Hoechst-positive events, gating out non-sperm events based on H33342 fluorescence, and excluding doublets based on a two-dimensional dot plot of forward scatter area (FSC-A) vs. forward-scatter height (FSC-H). Doublets have a higher signal width or area-to-height ratio than single cells (singlets), and events deviating from the diagonal are considered doublets. All samples were gently mixed using a vortex mixer to separate agglutinated sperm and then filtered through a 40 μm nylon mesh before acquiring the data. The data were acquired using ImageJ software v1.53n (Rasband, W.S., ImageJ, U.S. National Institutes of Health, Bethesda, MA, USA), and subsequent analysis was performed using FlowJo v10.8.2 (Becton, Dickinson, and Company, Ashland, OR, USA, 2022). Two replicates were conducted with the analysis of a different straw from the same freeze batch each time.
2.4.1. Membrane and Acrosome Integrities
The membrane and acrosome integrities were evaluated following the procedure described by Anzar et al., and Inanc et al. [28,29,44]. Briefly, diluted semen samples were incubated at 37°C for 10 minutes with fluorescent dyes: 2 μL H33342 (Biotium, Fremont, California, USA; H3570; stock 5 mg/mL in distilled water), 1 μL FITC-PNA (Sigma-Aldrich, L7381; stock 1 mg/mL in PBS), and 2 μL PI (Molecular Probes, Eugene, Oregon, USA; P1304MP; stock 2.4 mM in water) per mL diluted semen [28,44]. The emission and excitation details of all dyes used in this study are shown in Supplementary Material Table S1.
2.4.2. Membrane Stability and Viability
The membrane stability (capacitation rate on the level of sperm membrane) and viability were evaluated following the procedure of Ortega-Ferrusola et al. [27]. Briefly, diluted semen samples (1 mL) were incubated at 37 °C in the dark for 10 minutes with the addition of 2 μL of H33342 (Biotium, Fremont, California, USA; H3570; stock 5mg/mL in distilled water), 2.6 μL of M540 (Chemodex, St. Gallen, Switzerland; M0033; stock 1 mM in DMSO), and 1 μL of Yo-Pro 1 (Invitrogen, Waltham, MA, USA; Y 3603; stock 25 μM in DMSO) before acquiring the data.
2.4.3. Calcium Level
The intracellular calcium levels were evaluated following the procedure described by Luque et al. [45]. The Fluo-4 AM stock solution was prepared by adding 50 µg Fluo-4 AM (Invitrogen, Waltham, MA, USA; F14201) to 225 µL DMSO for a final concentration of 2 mM. Subsequently, 2.5 μL of the Fluo-4 AM stock solution, 2 μL of PI (2.4 mM water stock), and 2 μL of H33342 (5 mg/mL stock in distilled water) were added per mL of diluted semen and incubated for 20 minutes at 37°C. To induce calcium uptake and capture the calcium spike, 5 μL ionomycin (Calbiochem, Ionomycin, calcium salt, Streptomyces conglobatus; 407952; 1 mM stock in DMSO) was added to the samples, and data were acquired one minute after ionomycin addition. The final ionomycin concentration was 5 µM.
2.4.4. Mitochondrial Activity
The mitochondrial activity was evaluated according to the procedures of Gravance et al., Ortega-Ferrusola et al., and Spizziri et al. [27,36,46]. Briefly, diluted samples were treated with JC-1 (Invitrogen, Waltham, MA, USA; T3168; stock 2mM) by adding 7 μL of the stock solution to 1 mL diluted semen, besides 2 μL of H33342 (5 mg/mL stock in distilled water), with incubating for 20 minutes at 37°C. Mitochondria stained with JC-1 exhibit fluorescence in both orange and green colors, with the proportion depending on the level of mitochondrial activity. Active mitochondria accumulate JC-1 in their interior, forming aggregates that fluoresce orange. In contrast, less functional mitochondria accumulate low concentrations of JC-1, which remain in the monomeric form and fluoresce green. Computer analyses were used to determine the ratio of JC-1 aggregates to monomer fluorescence for each cell [27,36,46]. The control sample was only stained with Hoechst and used to gate out non-Hoechst-stained debris.
2.5. Sperm Binding and Interaction with Oviduct Cells
Bovine oviducts were obtained from a local slaughterhouse, transported in sterile filtered PBS on ice, and washed three times with filtered sterile PBS upon arrival at the laboratory. Oviduct cells were obtained and cultured following the protocol of Medeiros [47]. Briefly, the surrounding connective tissues were trimmed, and epithelial cells were recovered by scraping oviducts with a glass microscope slide into a dish containing PBS (1x) supplemented with 1 µL/mL Gentamicin (stock 50 mg/mL). The epithelial cells were mechanically disrupted by passing them through a 21-gauge syringe needle ten times to eliminate cell aggregates. Afterward, the cells were washed with PBS two times, with a third final wash using culture media. Oviduct cells were washed by allowing the cells to settle and then decanting the supernatant without centrifugation. Oviduct cells (five million per dish) were cultured in 35 mm Petri dishes containing 2 mL of equilibrated TCM199 supplemented with 10% (v/v) fetal calf serum, 50 µg/mL gentamycin, and pyruvate (0.2 mM). The dishes were incubated at 38.5°C under a 5% CO2 in air atmosphere until confluence. Confluence was reached after approximately 6 days of culture. On the day of evaluation, the medium was replaced with equilibrated Bovine Gamete Medium 1 (BGM1) supplemented with 6 mg/mL BSA [16], which is suitable for sperm incubation.
Frozen semen samples were thawed at 37 °C for 30 seconds, stained with Hoechst dye at a final concentration of 65 µM for 10 minutes, washed twice by centrifugation at 3500 × g in BGM1, and resuspended to a concentration of 24 × 106 sperm/mL. Twenty microliters of the semen suspension containing 480,000 sperm cells were added to each dish and incubated for 15 minutes. The dye YOYO-1 (1.6 µL; 0.4 µM stock in DMSO) was then added with swirling for the identification of dead sperm, with an additional 15 minutes of incubation. Excitation was set at 380 nm for Hoechst and 495 nm for YOYO, with emission greater than 510 nm for both dyes. Dishes were examined at four time points post-incubation (0.5, 3, 7, and 20 hours). At each time point, a dish was washed twice with 1 mL PBS to remove unbound sperm and then manually examined using a Nikon Epifluorescent Phase Contrast microscope (Nikon, Minato ku, Japan). Ten fields covering a surface area of 1.65 × 106 µm2 were evaluated per dish. Each field contains an average of 1165 oviduct cells. The evaluations included counting all Hoechst-positive sperm cells in ten random fields, along with counting the dead sperm cells stained with YOYO-1. Oviduct cells do not stain with YOYO-1 due to their viability, and they also do not stain with residual Hoechst. The number of bound and viable sperm was then calculated by multiplying the total number of sperm cells in ten random fields by the percentage viability. Three replicates were conducted with the analysis of a different straw from the same freeze batch each time.
2.6. IVF Experiment
Bovine ovaries were sourced from Applied Reproductive Technologies (Madison, WI, USA). The ovaries were collected, rinsed three times in prewarmed PBS with 10 mL/L of penicillin-streptomycin (100× Pen/Strep stock with 10,000 IU penicillin and 10 mg streptomycin per mL) to remove excess blood. Cumulus-oocyte complexes (COCs) were obtained by slicing follicles < 10 mm in diameter. Oocytes were collected by swirling 12-15 sliced ovaries in 100 mL oocyte collection media, prepared using M199 powder (Gibco, Grand Island, NY, USA) supplemented with bicarbonate (4 mM), Hepes (10 mM), and Pen/Strep (20 mL/L, stock 100× Pen/Strep). Heparin (2.4 μg/mL) to prevent coagulation and fetal bovine serum (20 mL/L) were added on the day of use. Subsequently, the resulting mixture was passed through a 100 μm cell strainer and backwashed into a grid plate containing oocyte collection media. Oocytes displaying homogeneous cytoplasm and surrounded by at least three layers of cumulus cells were selected under a stereomicroscope. After that, the selected oocytes were washed three times using a sterile 20 µL unopette tip attached to a Hamilton syringe.
Oocytes (50 per well) were matured in 4-well plates containing 500 μL of equilibrated maturation media [48] covered with 300 μL of light mineral oil. Maturation occurred over 24 hours at 38.5°C in a humidified atmosphere of 5% CO2 using TCM 199 media (with Earle’s salts), supplemented with 10% fetal calf serum, 50 µg/mL gentamicin, 0.2 mM pyruvate, 2 mM L-glutamine, 5 μg/mL follicle-stimulating hormone, and 1 μg/mL Estradiol [47]. Following maturation, 120 oocytes were transferred to 200 μL low bicarbonate TALP in a 1.5 mL microcentrifuge tube and vortex-mixed for 1 minute to remove cumulus cells following the procedure by Parrish et al. [40]. Oocytes were then washed in HEPES TALP, placed in 4-well Petri dishes with 60 oocytes added per well, containing 425 µL of IVF-TALP and supplemented with heparin (final concentration of 10 µg/mL) and 20 µL of PHE (stock, 0.5 mM penicillamine, 0.25 mM hypotaurine, and 25 µM epinephrine).
Frozen sperm, thawed for 30 seconds in 37°C water, were separated by centrifugation with PureSperm (Irvine Scientific, Santa Ana, CA, USA, 99264) at 3500 × g for 5 minutes in 1.5 mL microcentrifuge tubes. Motile sperm cells only penetrate the PureSperm gradient layers (50%/90%) during centrifugation and accumulate at the bottom. Thus, the bottom 100 μL layer was aspirated and washed in 0.5 mL HEPES TALP media by centrifugation at 900 × g for 3 minutes. The supernatant was discarded, and the sperm concentration was determined using a hemacytometer. The concentration was adjusted to 25 × 106 per mL in IVF TALP media, and 20 μL of the sperm suspension was added to each fertilization well. The final concentration of sperm cells was 1 × 106/mL, following the procedure by Parrish et al. [40].
For fertilization, matured cumulus-free oocytes and sperm were co-incubated at 38.5°C in a humidified atmosphere of 5% CO2 for three different time ranges: 4 hours, 8 hours, and 12 hours. After incubation, oocytes were removed, washed three times in HEPES-TALP, and mounted on slides. Oocytes were then carefully pressed under a coverslip supported with a paraffin wax-Vaseline mixture at its corners [49,50], with two sides of the coverslip sealed with rubber cement. Subsequently, oocytes were fixed in acetic acid:alcohol (1:3) until staining. At the time of evaluation, oocytes were stained with 1% (w/v) orcein in 45% (v/v) acetic acid. Excess stain was cleared with glycerol/acetic acid/water (1:1:3) as described by [51]. The oocytes were then observed under a phase-contrast microscope (400×) for sperm penetration, indicated by a decondensed sperm head with the tail, meiotic stage (anaphase II or beyond), or pronuclei formation [40]. Degenerated oocytes and oocytes with inconclusive observation were excluded. This approach allows for the tracking of fertilizing ability and fertilization progress at each designated time point. A total of 897 oocytes were evaluated across three replicates, each of which utilized a different straw from the same freeze batch.
2.7. Experimental Design
2.7.1. Experiment 1
This experiment aimed to optimize the treatment level and protocol. Semen from five different bulls was provided by ABS for this study. Semen concentration was first evaluated to dilute semen to 120 × 106 sperm cells/ mL, then treated with CLC. Four different CLC levels were tested: 0, 2, 4, and 6 mg per 120 × 106 sperm cells. The semen was then diluted to a final concentration of 30 × 106 sperm cells per mL using a 20% egg yolk Triladyl. Post-thaw samples (at 0-, 30-, and 60-minute time points) were assessed for total and progressive motilities using CASA.
2.7.2. Experiment 2
This experiment focuses on testing CLC levels lower than 2 mg/120 × 106 sperm cells. This experiment was conducted separately, with ABS providing semen from five different bulls than those used in Experiment 1. Semen was first evaluated for concentration and then diluted to 120 × 106 sperm cells/ mL using an egg yolk-free Triladyl. Five different CLC levels were examined: 0, 0.5, 1, 1.5, and 2 mg per 120 × 106 sperm cells. Semen was then diluted to a final concentration of 30 × 106 sperm cells/mL using a 20% egg yolk Triladyl. Post-thaw motion analysis at time points 0, 30, 60, and 120 minutes was evaluated using CASA. Based on the motility data, the 2 mg level of CLC was found to be the most effective and was used in further evaluations that included post-thaw flow cytometric analyses, an oviduct experiment to assess the binding ability of sperm cells, and an IVF experiment to evaluate the fertilizing ability and capacitation window.
2.7.3. Experiment 3
This experiment was designed to compare post-dilution with direct addition of CLC to raw semen, to ensure that CLC treatment can be adapted for industrial protocols. ABS provided semen from four different bulls, and all samples received 2 mg CLC per 120 × 106 sperm cells. For the post-dilution CLC-treated group, semen was prepared exactly as in Experiment 2: it was initially diluted with Tris extender (egg yolk-free) to 120 × 106 sperm/mL for CLC treatment, then further diluted to 30 × 106 sperm/mL using a 20% egg yolk Triladyl. For the raw CLC-treated group, CLC was directly added to raw semen at 2 mg per 120 × 106 sperm cells, then diluted directly to 30 × 106 sperm/mL with the same egg yolk Triladyl extender. Post-thaw total and progressive motilities of both groups were evaluated using CASA at 0, 30, and 60 minutes. Additionally, visual motility was assessed at each time point under a phase-contrast microscope (200×) by examining five fields per sample.
2.8. Statistical Analyses
The statistical analyses utilized analysis of variance using mixed models and/or general linear models. Time and CLC level were considered the main effects, the bull was treated as a random effect, and time was treated as a repeated measure if two or more time points were included. Percentage data were transformed using arcsine before analysis. Significance was determined at the 5% level for all analyses. Statistical analysis was performed using SAS software (SAS Institute Inc., NC, USA, SAS 9.4). All values are presented as means or least square means (LSM) with the corresponding standard error of the mean (SEM).
3. Results
3.1. Experiment 1
The results from the CASA evaluation of sperm (n=5) for Experiment 1 are depicted in Figure 1. There was no interaction between CLC level and post-thaw incubation time, but significant main effects were observed for both time and CLC level (p < 0.01). Specifically, the 2 mg and 4 mg treatments exhibited higher total motility overall (p < 0.05) via CASA evaluation compared to the 0 mg control group (Figure 1A). Progressive motility analysis showed that only the 2 mg level significantly differed from the control (Figure 1B; p < 0.05). The main effect of bull was significant (p < 0.05), but overall, the 2 mg level of CLC was found to be the most effective and was used in further studies.
3.2. Experiment 2
3.2.1. Motion Analyses
In Experiment 2, both total and progressive motilities were significantly affected by time (p < 0.01). The main effect of CLC level was significant for total motility (p < 0.05) but not for progressive motility. There was no interaction between time and CLC level; therefore, the results are presented as main effects of either CLC level or time. For total motility, CLC levels of 1, 1.5, and 2 mg CLC differed significantly from the control (0 mg), whereas 0.5 mg showed no difference. Although progressive motility showed no main effect of CLC level, Dunnett’s test indicated higher values at 1.5 and 2 mg (p < 0.05). Sperm motility declined gradually during incubation, with a significant reduction observed after just 30 minutes (p < 0.01; Table 2). A significant bull effect was detected for total (p < 0.01) and progressive motility (p < 0.05), but no interactions with time or CLC level were observed (p > 0.05). Overall, the 2 mg CLC level was among the most effective and was selected for subsequent flow cytometric, binding, and IVF analyses.

Table 1.
The impact of cholesterol-loaded cyclodextrin (CLC) level on post-thaw bovine sperm motilitya.
Table 1.
The impact of cholesterol-loaded cyclodextrin (CLC) level on post-thaw bovine sperm motilitya.
| CLC-level (mg/120 × 106 sperm cells) | Total Motility via CASA (%) | Progressive Motility via CASA (%) |
|---|---|---|
| 0 | 22.2 ± 2.8 | 5.2 ± 1.1 |
| 0.5 | 29.6 ± 2.8 | 7.8 ± 1.1 |
| 1 | 34.7* ± 2.8 | 8.4 ± 1.1 |
| 1.5 | 35.7* ± 2.8 | 10.4* ± 1.1 |
| 2 | 36.1* ± 2.8 | 9.2* ± 1.1 |
a Motility was assessed at four time points (0, 30, 60, and 120 min.) post-thaw, and then averaged (n=5). All values are expressed as means ± standard error of the mean. CLC had a significant effect on total motility (p < 0.05) but not on progressive motility. An * indicates statistically significant differences (p < 0.05) compared to the 0 CLC level (control) using Dunnett’s test.
Table 2.
Repeated measures analysis of time as a factor affecting sperm motilitya.
| Post-thaw time (min.) | Total Motility via CASA (%) | Progressive Motility via CASA (%) |
|---|---|---|
| 0 | 43 ± 2.1 | 12 ± 0.8 |
| 30 | 37.4** ± 2.5 | 10.2** ± 1.1 |
| 60 | 26.9** ± 2.1 | 6.8** ± 0.8 |
| 120 | 19.5** ± 2.1 | 3.6** ± 0.6 |
a The data include all the CLC-treated groups and the control. Values are expressed as means ± SEM. Time exhibited a significant effect (p < 0.01) for both traits, while the interaction between CLC and time was not significant for either trait. ** within a column indicates p < 0.01 compared to the 0-minute time point using Dunnett’s test.
3.2.2. Flow Cytometric Analyses
Fluorescent data for membrane and acrosome intactness were recorded using photomultiplier detectors and analyzed on a two-dimensional dot plot displaying PI and FITC-PNA intensities. This allowed for the identification of four sperm populations: sperm with intact plasma membrane and intact acrosomes (PI-/FITC-PNA-), sperm with intact plasma membrane and compromised acrosomes (PI-/FITC-PNA+), sperm with compromised plasma membrane and intact acrosomes (PI+/FITC-PNA-), and sperm with compromised plasma membrane and compromised acrosomes (PI+/FITC-PNA+), as described by Anzar et al., and Inanc et al. [28,29,44]. Gating utilized three controls: Hoechst only control (-FITC-PNA, -PI), Hoechst +PI without FITC-PNA, and Hoechst +FITC-PNA without PI. CLC level showed no effect on the percentage of cells with intact membrane and intact acrosome; the control group average was 20.9 ± 2.9 % compared to 30.5 ± 2.9 % for the treated group (Table 3).
For data analysis of membrane stability and viability, a Yo-Pro 1/M540 (YP/M540) dot plot was employed to differentiate viable and stable (uncapacitated with low fluidity) plasma membrane (YP- / M540-), viable and unstable (capacitated with higher fluidity) plasma membrane (YP- / M540+), and dead (YP+) events [27]. Gating was performed using three controls: Hoechst only (-M540, -YP), Hoechst and YP without M540, and Hoechst plus M540 without YP. There was a trend toward a main effect of CLC treatment on the subpopulation of viable sperm with stable plasma membranes (viable uncapacitated), with the control group exhibiting a mean of 20.4 ± 3.7 % and the CLC-treated group 32.8 ± 3.7 % (p = 0.08; Table 3). The mitochondrial membrane potential assay showed no significant difference between groups, with an orange-to-green fluorescence ratio of 0.99 ± 0.04 in the control and 0.91 ± 0.04 in the CLC-treated group (Table 3).
For analyzing calcium data, the Hoechst-positive population was first selected based on the Hoechst-only stained control (-Fluo-4 AM / -PI). Two additional controls with only PI staining (-Fluo-4 AM / +PI) or only Fluo-4 AM staining (+Fluo-4 AM / -PI) were utilized for gating. Subsequently, for each sample, two-dimensional fluorescence dot plots were generated, and the live (negative for PI staining) and dead (positive for PI) sperm populations were gated. These two populations were employed for the analysis of relative [Ca2+]i level, based on the fluorescence intensity of Fluo-4 AM [45]. Relative calcium level within the live sperm population showed no difference between the control and the CLC-treated group (Table 4). Additionally, calcium uptake within the live population was significantly (p < 0.01) shifted by the same magnitude in both control (from a value of 569 ± 129 to 1476 ± 129) and CLC-treated group (from a value of 622 ± 129 to 1491 ± 129) when triggered by ionomycin addition (Table 4).
3.2.3. Oviduct Experiment
The study evaluated the binding ability of bovine frozen sperm cells to oviduct cells in both the control and CLC-treated group at 2 mg CLC/120 × 106 sperm cells. The overall number of viable bound sperm cells to a surface area of 1.65 × 106 µm2 oviduct cells (ten fields) exhibited a highly significant difference between the two groups (p < 0.01). Time also had a highly significant main effect on the number of viable bound sperm cells (p < 0.01), with a linear decrease observed over the incubation period. Furthermore, the interaction between time and CLC level revealed a highly significant difference between the groups (p < 0.01). The CLC treatment significantly (p < 0.05) enhanced the sperm binding capacity, with the effect persisting for up to 7 hours of coincubation (Table 5). However, after 20 h of coincubation, the difference between the two groups was no longer significant (Table 5).
3.2.4. IVF Experiment
The influence of CLC treatment on penetration and pronuclei formation rates was evaluated. The main effect of CLC treatment was not significant for either penetration or pronuclei formation rate. Additionally, the main effect of coincubation time on penetration rate was not significant, suggesting that most oocytes were penetrated within 4 h of coincubation with sperm cells. However, Dunnett’s test revealed a higher fertilization rate at 12 h compared to 4 h (p < 0.05; Table 6). Pronuclei formation rate was significantly (p < 0.0001) increased by the coincubation time. Additionally, the polynomial contrast analysis of the pronuclei formation rate revealed significant linear (p < 0.0001) and quadratic (p < 0.05) effects of coincubation time. This indicates that while the percentage of pronuclei formation continued to increase over time, the rate of increase was gradually declining, suggesting it was beginning to plateau. After 12 h of coincubation, which was the end point of the experiment, approximately 8.3% (26/314) of all oocytes exhibited polyspermy. Oocytes containing a single pronucleus were considered neither penetrated nor having formed pronuclei, as the origin of that pronucleus could not be confirmed as sperm-derived. Oocytes exhibiting polyspermy were considered penetrated. Also, oocytes displaying three or more pronuclei were classified as polyspermy and included in the pronuclei formation population. Overall, CLC-treated sperm cells showed penetration and pronuclei formation rates comparable to the control when equal numbers of motile sperm were used.
3.3. Experiment 3
All motility traits did not differ between post-dilution CLC treatment and direct addition to raw semen when 2 mg of CLC per 120 × 106 sperm cells was used (Figure 2). This indicates that direct CLC addition to raw semen is feasible for industrial use.
4. Discussion
In the present study, motion analysis revealed that the optimal CLC level ranged between 1.5-2 mg CLC/mL extended semen containing 120 × 106 sperm cells. These findings in Experiments 1 and 2 are consistent with previous findings as reviewed by Moce et al. [21]. CLC levels exceeding 4 mg/120 × 106 sperm cells can cause a detrimental effect on sperm motility, as evident in Experiment 1. The optimum CLC treatment aims to elevate sperm cholesterol levels by 2-3 times (exceeding a 3-fold increase may lead to adverse effects) in bull sperm before cryopreservation [24]. This treatment targets raising the cholesterol/phospholipid ratio (Chol/PL) from 0.45 to > 0.8, and this goal can be accomplished by adding 1 to 2 mg CLC per 120 × 106 sperm cells [21]. Cholesterol/Phospholipid ratio is the major determinant of sperm plasma membrane fluidity in animals with lower Chol/PL in sperm plasma membrane, like the bull [21].
Cholesterol constitutes about 40% of mammalian plasma membranes and works as a fluidity buffer, essential for sperm viability and motility after cryopreservation [52,53]. Modification of the sperm plasma membrane with cholesterol affects its fluidity and permeability in a temperature-dependent manner. At temperatures above the phase transition (liquid-ordered phase), the incorporation of cholesterol within the phospholipid tails of sperm plasma membrane pulls the phospholipid tails closer together and decreases their mobility and consequently reducing membrane fluidity. Thus, cholesterol at higher temperatures condenses, stabilizes, and tightens the sperm membrane. Conversely, at colder temperatures, sperm phospholipids tend to pack tightly. Cholesterol supplementation in this scenario would prevent excessive packing while keeping phospholipids apart. This softens the sperm membrane, helping it resists the transition to the solid phase and preventing the lateral separation of phospholipids.
It’s speculated that cholesterol reduces cryodamage (indicated by higher motility and a trend towards higher viable uncapacitated rate in this study) by making membranes less compact (more permeable) at subzero temperatures, allowing more water to exit sperm cells, leading to greater shrinkage and a lower chance of ice crystal formation during cryopreservation. This could also help interpret how higher cholesterol could be detrimental to sperm cells. Higher cholesterol levels may increase membrane fluidity, leading to more water efflux to the extent that sperm cells can’t tolerate the dehydration and volume shrinkage. The other possibility is that holding phospholipids apart by cholesterol, which increases fluidity and permeability while sperm is freezing, may allow the higher concentrated cryoprotectants around the sperm cells to rush inside and cause more damage to sperm. If this is the case, then it’s essential to reduce cryoprotectant agent (CPA) levels and reconsider the entire freezing protocol and composition of the extender when CLC is used.
For example, reducing the glycerol level from 6 % to 3 % when CLC is used at 1.5 mg CLC/120 × 106 sperm cells improved post-thaw sperm viability, motility, capacitation, mitochondrial potential, and fertilizing ability [54]. Glycerol, even at concentrations as low as 4 % and whether added before or after cooling, has been reported by Aly [55] to have detrimental effects on the post-thaw motility, viability, and fertility of rabbit spermatozoa compared with sucrose or a combination of DMSO and sucrose. Aly also reported that sucrose inclusion can mitigate the adverse effects associated with glycerol exposure. Rabbit sperm contain a higher cholesterol-to-phospholipid ratio [56]. Therefore, it’s logical that raising the cholesterol-to-phospholipid ratio in bull sperm might necessitate lowering the glycerol level or changing the CPA to include nonpermeable agents. Another example relates to the interaction between egg yolk and CLC, which is still not well understood. The addition of CLC was found to effectively replace egg yolk, giving similar cleavage and blastocyst rates [44].
One approach that indicates cholesterol incorporation into sperm plasma membranes is by evaluating sperm cryo-survivability when samples are frozen directly from room temperature without prior cooling [57]. In our laboratory, when bovine semen was frozen directly without prior cooling, cholesterol supplementation maintained acceptable post-thaw motility, whereas the control group exhibited nearly zero motility (personal observation, Ahmed Aly). Also, our findings indicate no difference between the CLC-treated groups and the control before freezing. This supports the idea that CLC works at subzero temperatures. Following thawing, sperm cholesterol drastically drops, inducing cryo-capacitation. However, CLC-treated sperm also lose cholesterol during cryopreservation yet retain a higher post-thaw cholesterol content than untreated sperm [11,22,24]. Evaluating post-thaw sperm cholesterol content using phase chromatography appears insufficiently sensitive to detect CLC treatment effects at concentrations below 4 mg per 120 × 106 sperm cells [24]. Therefore, future studies should consider developing and validating flow cytometry–based approaches utilizing Filipin III staining, as demonstrated by Salmon et al. [11] using confocal microscopy, or by tracking cholesterol efflux with BODIPY-cholesterol, as described by Bernecic et al. [58], to enable more accurate assessment in bovine sperm.
Although the present study did not directly measure sperm cholesterol content due to funding constraints, evidence of plasma membrane modification was supported by several observations, including improved post-thaw sperm motility, a trend toward a higher proportion of viable uncapacitated sperm (indicating reduced cryo-capacitation rate), and enhanced binding ability to oviduct epithelial cells. Cryo-capacitated sperm cells lose coating glycoproteins, which increases membrane fluidity and destabilization, allowing more M540 intercalation into the sperm membrane [32,33,34]. In this study, cholesterol addition resulted in a trend towards lower M540 intercalation, which indicates lower post-thaw sperm fluidity.
The addition of CLC did not affect acrosome integrity, consistent with the findings reported by Purdy and Graham [22], but contradicts the results by Batissaco et al., LaVelle et al., Oliveria et al., and Aly et al. [43,59,60,61]. The Purdy and Graham study assessed acrosome reaction using FITC-PNA after inducing calcium uptake with various stimuli, while the present study did not employ any inducers in this experiment. In the calcium experiment, however, ionomycin was used to enhance calcium uptake and was expected to stimulate the acrosome reaction. Both CLC-treated sperm cells and the control exhibited a similar increase in calcium levels, indicating a similar acrosome induction response in both groups, consistent with the findings by Purdy and Graham, and Spizziri et al. [22,36].
Although motility was improved with cholesterol addition in this study, the mitochondrial potential did not show significant improvement. These results align with Spizziri et al., and Aly et al. [36,43]. Additionally, the 6 mg CLC level was found to significantly reduce post-thaw sperm motility, and progressive motility was found to be more sensitive to the 4 mg level compared with total motility (Figure 1A and 1B), indicating that CLC might impact sperm motility, but the mechanism is not clear. This leaves us with two possible speculations: the first one relates to the expected impacts of cholesterol on mitochondrial activity, while the second one relates to the capacitation status of sperm cells. The impact of cholesterol addition on enzyme activity and the metabolic rate of sperm cells remains unknown. However, studies have shown that adding cholesterol to isolated mitochondria in culture increases cholesterol content in both the outer and inner mitochondrial membranes [62,63], decreasing their fluidity and permeability, which can be reversed by adding methyl-β-cyclodextrin [63]. Elevated cholesterol in mitochondrial membranes severely reduces glutathione levels. Glutathione regulates key Krebs cycle enzymes, impacts substrate transport through the cycle, increases reactive oxygen species (ROS) production, affects respiration and ATP production, and may lead to cell death, as reviewed by Martin et al. [64]. Previous studies have indicated that high cholesterol levels are detrimental to sperm motility and viability but did not provide possible mechanisms for this effect. In the present study, the authors suggest that the impact of cholesterol on mitochondria could provide a good explanation for these findings, particularly as to why progressive motility was more sensitive and why mitochondrial potential did not improve with CLC addition. Sperm cells treated with cholesterol appear less capacitated [59,60,61,65]. During capacitation, sperm cells consume more glucose and ATP, displaying increased mitochondrial activity to meet the demands of motility [66,67,68,69]. The more active the mitochondria are, the more ROS is generated, which is a major cryo-capacitation inducer [1,4,70]. Thus, CLC-treated sperm cells likely act as less capacitated cells, consuming less glucose, producing less ROS, and having lower mitochondrial potential.
CLC-treated sperm cells demonstrated a significantly enhanced ability to bind and interact with oviduct cells in the present study. It exhibited a double-binding ability when compared to non-treated sperm cells (calculated by dividing the overall mean of the number of viable bound cells for the treated group by the control). Treating human mesenchymal stem cells with cholesterol increased membrane cholesterol, reduced fluidity, concentrated caveolae, and Caveolin-1, and in turn enhanced adhesion and integrin levels. This effect can be reversed with methyl-β-cyclodextrin addition [71]. Similar improvements are expected in sperm-oviduct cell adhesion and interaction if the same mechanism is applied. Treating porcine sperm cells with CLC was found to improve the interaction with oviduct cells [72]. Following insemination, a functional sperm reservoir (SR) develops at the ampullary–isthmic junction, aiming to sustain a selected population of viable spermatozoa for an extended period, ultimately facilitating coordinated sperm capacitation at the time of ovulation [73,74,75]. For sperm to interact with oviduct cells, they should exhibit a lower cryo-capacitation rate with proper surface receptors [39]. Therefore, successful cryopreserved semen samples are expected to have subpopulations of sperm cells that can be capacitated over a wider range of time and would be expected to increase the chance of sperm to fertilize an egg [76].
The IVF results agree with Purdy and Graham, and Spizziri et al. [22,36]. When an equivalent number of motile sperm is introduced per oocyte, achieving comparable IVF outcomes becomes feasible, suggesting the preserved fertilizing ability of cholesterol-treated sperm [22]. Bovine sperm cells respond to capacitation induction with heparin, with most of the oocytes being penetrated within 4 hours, indicating the completeness of the capacitation process [40]. This experiment, along with the calcium experiment, suggests that CLC-treated sperm cells are physiologically normal and can maintain their fertilizing ability.
This study proposes and discusses three mechanisms by which higher CLC levels (> 4 mg/120 × 106 sperm) adversely affect sperm viability and motility. Firstly, higher CLC levels may impair sperm mitochondrial function. Secondly, they may increase membrane fluidity and permeability at sub-zero temperatures, causing excessive shrinkage or elevated intracellular cryoprotectant accumulation, both of which can damage sperm. Thirdly, increasing CLC levels may impact the sperm plasma membrane domains and lipid rafts [21], which could lead to negative impacts on sperm cells. The impacts of higher cholesterol on sperm lipid domains and receptors were recently supported [77].
The fertility outcomes after sperm are treated with CLC in various species (bull, ram, rooster, boar, and rabbit buck) are controversial, leading to different explanations by previous publications, but the general conclusion is that increasing the cholesterol content of the plasma membrane might delay capacitation [11,24]. Other explanations include possible modifications of the sperm plasma membrane domains and other lipid raft components, including phospholipid redistribution [21]. Capacitation is judged to be complete when sperm can penetrate a zona-intact oocyte, fuse with the oolemma of the oocyte, and decondense in the oocyte cytoplasm [16,40]. The present study evaluated the rates of sperm penetration and pronuclei formation to reflect the capacitation window and help determine the timing of fertilization [40,41], providing insights into how CLC-treated cells respond to capacitation induction in vitro. Both the control and CLC-treated groups responded similarly, exhibiting similar penetration and pronuclei formation rates. Flow cytometry analyses of capacitation progress in a recent study indicated that the addition of cholesterol resulted in a one-hour delay in capacitation compared to the untreated control [60]. Our findings exclude delayed capacitation as a cause of reduced fertility with CLC-treated sperm. However, the potential effects of CLC treatment on lipid rafts, membrane domains, and dynamics, as well as the possible damage to DNA (resulting from increased membrane permeability at sub-zero temperatures and CPA concentrations) or other cellular components, warrant further investigation.
In the IVF work of this study, oocytes were checked every 4 hours, limiting our ability to detect treatment differences in the capacitation time of less than 4 hours. The delay can be detected, but it’s challenging to determine the exact time of change. Another limitation in this study is that the time required to capacitate sperm in vitro is not necessarily that needed to capacitate sperm in vivo. However, we expect the capacitation times to be relative, and if CLC-treated sperm require more time to capacitate in vitro, we would also expect these sperm to require longer capacitation times in vivo. Further investigation, including a fertility trial, is still needed, taking into consideration the possible delay in the in-vivo capacitation window of treated sperm cells [11,18,21]. An additional limitation is that we cultured these embryos for only 12 h, which aligned with our objective at this stage. In future studies, we would consider culturing them to the blastocyst stage or even transferring them to recipient cows.
5. Conclusions
In conclusion, the optimal level of CLC for treating bovine sperm was determined to lie between 1.5 and 2 mg CLC/mL of semen containing 120 × 106 sperm cells, as this range significantly enhanced sperm motility. Although a reduction in cryo-capacitation tendency was observed at the membrane level, downstream indicators, including intracellular calcium concentration and oocyte penetration rate, were comparable to the control, indicating that the overall capacitation window was not altered. Additionally, treated sperm cells exhibited an improved ability to interact with oviduct cells, implying potential improvements in surface proteins and receptors. However, mitochondrial potential remained similar to that of untreated sperm cells. These findings indicate that CLC treatment could increase straw production efficiency, especially from younger bulls with low daily sperm output. The expected increase in the number of straws obtained per ejaculate should correspond to the percentage increase in motility. The CLC treatment shows potential for industrial application; nevertheless, additional validation with commercial semen and fertility trials is required to confirm its practical effectiveness. Future studies should also examine the interactions of CLC with extender components such as egg yolk and glycerol, and explore the inclusion of additional membrane-modifying molecules, such as fatty acids, to achieve a more balanced treatment and improved cryopreservation outcomes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Fluorescent dyes: emissions, excitations, and detection filters used in the present study.
Author Contributions
Conceptualization, methodology, investigation, software, validation, and formal analysis, data curation, A.S.A. and J.J.P.; writing—original draft preparation, A.S.A.; writing—review and editing, J.J.P.; resources, A.S.A., and J.J.P.; supervision: J.J.P. All authors have read and agreed to the published version of the manuscript.
Funding
Ahmed Aly acknowledges and expresses gratitude for the full Ph.D. scholarship (GM-1071) from the Ministry of Higher Education of the Arab Republic of Egypt. The authors acknowledge support from the University of Wisconsin-Madison, Office of the Vice Chancellor for Research and Graduate Education.
Acknowledgments
The authors express gratitude to the UWCCC Flow Cytometry Laboratory at the University of Wisconsin Carbone Cancer Center, supported by P30 CA014520, for providing facilities and services, with special thanks to the laboratory staff for their assistance. Olivia Evans is acknowledged for her help with semen cryopreservation. The authors appreciate Dr. Sofia Ortega for her guidance throughout the IVF experiment. Gratitude to Dr. Adel Talaat for his support. Special thanks to Dr. Kevin J. Rozeboom and ABS for providing the semen and support that made this study possible.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Leahy, T.; Gadella, B.M. Sperm Surface Changes and Physiological Consequences Induced by Sperm Handling and Storage. Reproduction 2011, 142. [Google Scholar] [CrossRef]
- Ozkavukcu, S.; Erdemli, E.; Isik, A.; Oztuna, D.; Karahuseyinoglu, S. Effects of Cryopreservation on Sperm Parameters and Ultrastructural Morphology of Human Spermatozoa. J Assist Reprod Genet 2008, 25. [Google Scholar] [CrossRef] [PubMed]
- Sieme, H.; Oldenhof, H.; Wolkers, W.F. Sperm Membrane Behaviour during Cooling and Cryopreservation. Reproduction in Domestic Animals 2015, 50. [Google Scholar] [CrossRef] [PubMed]
- Ugur, M.R.; Saber Abdelrahman, A.; Evans, H.C.; Gilmore, A.A.; Hitit, M.; Arifiantini, R.I.; Purwantara, B.; Kaya, A.; Memili, E. Advances in Cryopreservation of Bull Sperm. Front Vet Sci 2019, 6. [Google Scholar] [CrossRef]
- Watson, P.F. The Causes of Reduced Fertility with Cryopreserved Semen. In Proceedings of the Animal Reproduction Science; 2000; Vol. 60–61. [CrossRef]
- Watson, P.F. Recent Developments and Concepts in the Cryopreservation of Spermatozoa and the Assessment of Their Post-Thawing Function. Reprod Fertil Dev 1995, 7. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.; Morrier, A.; Cormier, N. Semen Cryopreservation: Successes and Persistent Problems in Farm Species. Can J Anim Sci 2003, 83. [Google Scholar] [CrossRef]
- Bernecic, N.C.; Gadella, B.M.; Leahy, T.; de Graaf, S.P. Novel Methods to Detect Capacitation-Related Changes in Spermatozoa. Theriogenology 2019, 137. [Google Scholar] [CrossRef]
- Thomas, A.D.; Meyers, S.A.; Ball, B.A. Capacitation-like Changes in Equine Spermatozoa Following Cryopreservation. Theriogenology 2006, 65. [Google Scholar] [CrossRef]
- Khan, I.M.; Cao, Z.; Liu, H.; Khan, A.; Rahman, S.U.; Khan, M.Z.; Sathanawongs, A.; Zhang, Y. Impact of Cryopreservation on Spermatozoa Freeze-Thawed Traits and Relevance OMICS to Assess Sperm Cryo-Tolerance in Farm Animals. Front Vet Sci 2021, 8. [Google Scholar] [CrossRef]
- Salmon, V.M.; Leclerc, P.; Bailey, J.L. Cholesterol-Loaded Cyclodextrin Increases the Cholesterol Content of Goat Sperm to Improve Cold and Osmotic Resistance and Maintain Sperm Function after Cryopreservation. Biol Reprod 2016, 94. [Google Scholar] [CrossRef]
- Benko, F.; Ďuračka, M.; Baňas, Š.; Lukáč, N.; Tvrdá, E. Biological Relevance of Free Radicals in the Process of Physiological Capacitation and Cryocapacitation. Oxygen 2022, 2. [Google Scholar] [CrossRef]
- Benko, F.; Fialková, V.; Žiarovská, J.; Ďuračka, M.; Lukáč, N.; Tvrdá, E. In Vitro versus Cryo-Induced Capacitation of Bovine Spermatozoa, Part 2: Changes in the Expression Patterns of Selected Transmembrane Channels and Protein Kinase A. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Westfalewicz, B.; Dietrich, M.A.; Mostek, A.; Partyka, A.; Bielas, W.; Niżański, W.; Ciereszko, A. Analysis of Bull (Bos Taurus) Seminal Vesicle Fluid Proteome in Relation to Seminal Plasma Proteome. J Dairy Sci 2017, 100. [Google Scholar] [CrossRef]
- Benko, F.; Mohammadi-Sangcheshmeh, A.; Ďuračka, M.; Lukáč, N.; Tvrdá, E. In Vitro versus Cryo-Induced Capacitation of Bovine Spermatozoa, Part 1: Structural, Functional, and Oxidative Similarities and Differences. PLoS One 2022, 17. [Google Scholar] [CrossRef]
- Parrish, J.J. Bovine in Vitro Fertilization: In Vitro Oocyte Maturation and Sperm Capacitation with Heparin. Theriogenology 2014, 81. [Google Scholar] [CrossRef]
- Abah, K.O.; Fontbonne, A.; Partyka, A.; Nizanski, W. Effect of Male Age on Semen Quality in Domestic Animals: Potential for Advanced Functional and Translational Research? Vet Res Commun 2023, 47. [Google Scholar] [CrossRef]
- Amann, R.P.; DeJarnette, J.M. Impact of Genomic Selection of AI Dairy Sires on Their Likely Utilization and Methods to Estimate Fertility: A Paradigm Shift. Theriogenology 2012, 77. [Google Scholar] [CrossRef]
- Murphy, E.M.; Kelly, A.K.; O’Meara, C.; Eivers, B.; Lonergan, P.; Fair, S. Influence of Bull Age, Ejaculate Number, and Season of Collection on Semen Production and Sperm Motility Parameters in Holstein Friesian Bulls in a Commercial Artificial Insemination Centre. J Anim Sci 2018, 96. [Google Scholar] [CrossRef]
- DeJarnette, J.M.; Marshall, C.E.; Lenz, R.W.; Monke, D.R.; Ayars, W.H.; Sattler, C.G. Sustaining the Fertility of Artificially Inseminated Dairy Cattle: The Role of the Artificial Insemination Industry. J Dairy Sci 2004, 87. [Google Scholar] [CrossRef]
- Mocé, E.; Blanch, E.; Tomás, C.; Graham, J.K. Use of Cholesterol in Sperm Cryopreservation: Present Moment and Perspectives to Future. Reproduction in Domestic Animals 2010, 45. [Google Scholar] [CrossRef]
- Purdy, P.H.; Graham, J.K. Effect of Adding Cholesterol to Bull Sperm Membranes on Sperm Capacitation, the Acrosome Reaction, and Fertility. Biol Reprod 2004, 71. [Google Scholar] [CrossRef]
- Hartwig, F.P.; Lisboa, F.P.; Hartwig, F.P.; Monteiro, G.A.; Maziero, R.R.D.; Freitas-Dell’Aqua, C.P.; Alvarenga, M.A.; Papa, F.O.; Dell’Aqua, J.A. Use of Cholesterol-Loaded Cyclodextrin: An Alternative for Bad Cooler Stallions. Theriogenology 2014, 81. [Google Scholar] [CrossRef]
- Purdy, P.H.; Graham, J.K. Effect of Cholesterol-Loaded Cyclodextrin on the Cryosurvival of Bull Sperm. Cryobiology 2004, 48. [Google Scholar] [CrossRef]
- Ligocka, Z.; Partyka, A.; Schäfer-Somi, S.; Mucha, A.; Niżański, W. Does Better Post-Thaw Motility of Dog Sperm Frozen with CLC Mean Better Zona Pellucida Binding Ability? Animals 2023, 13. [Google Scholar] [CrossRef]
- de Lima Rosa, J.; de Paula Freitas Dell’Aqua, C.; de Souza, F.F.; Missassi, G.; Kempinas, W.D.G. Multiple Flow Cytometry Analysis for Assessing Human Sperm Functional Characteristics. Reproductive Toxicology 2023, 117. [Google Scholar] [CrossRef]
- Ortega-Ferrusola, C.; Anel-López, L.; Martín-Muñoz, P.; Ortíz-Rodríguez, J.M.; Gil, M.C.; Alvarez, M.; De Paz, P.; Ezquerra, L.J.; Masot, A.J.; Redondo, E.; et al. Computational Flow Cytometry Reveals That Cryopreservation Induces Spermptosis but Subpopulations of Spermatozoa May Experience Capacitation-like Changes. Reproduction 2017, 153. [Google Scholar] [CrossRef]
- Anzar, M.; Kroetsch, T.; Boswall, L. Cryopreservation of Bull Semen Shipped Overnight and Its Effect on Post-Thaw Sperm Motility, Plasma Membrane Integrity, Mitochondrial Membrane Potential and Normal Acrosomes. Anim Reprod Sci 2011, 126. [Google Scholar] [CrossRef]
- Inanc, M.E.; Gungor, S.; Ozturk, C.; Korkmaz, F.; Bastan, I.; Cil, B. Cholesterol-Loaded Cyclodextrin plus Trehalose Improves Quality of Frozen-Thawed Ram Sperm. Vet Med (Praha) 2019, 64, 118–124. [Google Scholar] [CrossRef]
- Hallap, T.; Nagy, S.; Jaakma, Ü.; Johannisson, A.; Rodriguez-Martinez, H. Usefulness of a Triple Fluorochrome Combination Merocyanine 540/Yo-Pro 1/Hoechst 33342 in Assessing Membrane Stability of Viable Frozen-Thawed Spermatozoa from Estonian Holstein AI Bulls. Theriogenology 2006, 65. [Google Scholar] [CrossRef]
- Peña, F.J.; Saravia, F.; Johannisson, A.; Walgren, M.; Rodríguez-Martínez, H. A New and Simple Method to Evaluate Early Membrane Changes in Frozen-Thawed Boar Spermatozoa. Int J Androl 2005, 28. [Google Scholar] [CrossRef]
- Steckler, D.; Stout, T.A.E.; Durandt, C.; Nöthling, J.O. Validation of Merocyanine 540 Staining as a Technique for Assessing Capacitation-Related Membrane Destabilization of Fresh Dog Sperm. Theriogenology 2015, 83. [Google Scholar] [CrossRef]
- Harrison, R.A.P.; Ashworth, P.J.C.; Miller, N.G.A. Bicarbonate/CO2, an Effector of Capacitation, Induces a Rapid and Reversible Change in the Lipid Architecture of Boar Sperm Plasma Membranes. Mol Reprod Dev 1996, 45. [Google Scholar] [CrossRef]
- Guthrie, H.D.; Welch, G.R. Effects of Hypothermic Liquid Storage and Cryopreservation on Basal and Induced Plasma Membrane Phospholipid Disorder and Acrosome Exocytosis in Boar Spermatozoa. Reprod Fertil Dev 2005, 17. [Google Scholar] [CrossRef]
- Parrish, J.J.; Susko-Parrish, J.L.; Graham, J.K. In Vitro Capacitation of Bovine Spermatozoa: Role of Intracellular Calcium. Theriogenology 1999, 51, 461–472. [Google Scholar] [CrossRef]
- Spizziri, B.E.; Fox, M.H.; Bruemmer, J.E.; Squires, E.L.; Graham, J.K. Cholesterol-Loaded-Cyclodextrins and Fertility Potential of Stallions Spermatozoa. Anim Reprod Sci 2010, 118, 255–264. [Google Scholar] [CrossRef]
- Suarez, S.S. Regulation of Sperm Storage and Movement in the Mammalian Oviduct. International Journal of Developmental Biology 2008, 52. [Google Scholar] [CrossRef]
- Ellington, J.E.; Samper, J.C.; Jones, A.E.; Oliver, S.A.; Burnett, K.M.; Wright, R.W. In Vitro Interactions of Cryopreserved Stallion Spermatozoa and Oviduct (Uterine Tube) Epithelial Cells or Their Secretory Products. Anim Reprod Sci 1999, 56. [Google Scholar] [CrossRef]
- Fazeli, A.; Duncan, A.E.; Watson, P.F.; Holt, W. V. Sperm-Oviduct Interaction: Induction of Capacitation and Preferential Binding of Uncapacitated Spermatozoa to Oviductal Epithelial Cells in Porcine Species. Biol Reprod 1999, 60. [Google Scholar] [CrossRef]
- Parrish, J.J.; Susko-Parrish, J.; Winer, M.A.; First, N.L. Capacitation of Bovine Sperm by Heparin. Biol Reprod 1988, 38. [Google Scholar] [CrossRef]
- Koyama, K.; Kang, S.S.; Huang, W.; Yanagawa, Y.; Takahashi, Y.; Nagano, M. Estimation of the Optimal Timing of Fertilization for Embryo Development of in Vitro-Matured Bovine Oocytes Based on the Times of Nuclear Maturation and Sperm Penetration. Journal of Veterinary Medical Science 2014, 76. [Google Scholar] [CrossRef]
- Mocé, E.; Graham, J.K. Cholesterol-Loaded Cyclodextrins Added to Fresh Bull Ejaculates Improve Sperm Cryosurvival. J Anim Sci 2006, 84. [Google Scholar] [CrossRef]
- Aly, A.S.; Rozeboom, K.J.; Parrish, J.J. Using Cholesterol-Loaded Cyclodextrin to Improve Cryo-Survivability and Reduce Capacitation-Like Changes in Gender-Ablated Jersey Semen. Animals 2025, 15. [Google Scholar] [CrossRef]
- Anzar, M.; Rajapaksha, K.; Boswall, L. Egg Yolk-Free Cryopreservation of Bull Semen. PLoS One 2019, 14. [Google Scholar] [CrossRef]
- Luque, G.M.; Dalotto-Moreno, T.; Martín-Hidalgo, D.; Ritagliati, C.; Puga Molina, L.C.; Romarowski, A.; Balestrini, P.A.; Schiavi-Ehrenhaus, L.J.; Gilio, N.; Krapf, D.; et al. Only a Subpopulation of Mouse Sperm Displays a Rapid Increase in Intracellular Calcium during Capacitation. J Cell Physiol 2018, 233. [Google Scholar] [CrossRef]
- Gravance, C.G.; Garner, D.L.; Miller, M.G.; Berger, T. Fluorescent Probes and Flow Cytometry to Assess Rat Sperm Integrity and Mitochondrial Function. Reproductive Toxicology 2000, 15. [Google Scholar] [CrossRef]
- Ortega, M.S.; Moraes, J.G.N.; Patterson, D.J.; Smith, M.F.; Behura, S.K.; Poock, S.; Spencer, T.E. Influences of Sire Conception Rate on Pregnancy Establishment in Dairy Cattle. Biol Reprod 2018, 99. [Google Scholar] [CrossRef]
- De La Torre-Sanchez, J.F.; Preis, K.; Seidel, G.E. Metabolic Regulation of In-Vitro-Produced Bovine Embryos. I. Effects of Metabolic Regulators at Different Glucose Concentrations with Embryos Produced by Semen from Different Bulls. Reprod Fertil Dev 2006, 18. [Google Scholar] [CrossRef]
- Hwang, K.; Lamb, D.J. The Sperm Penetration Assay for the Assessment of Fertilization Capacity. Methods in Molecular Biology 2013, 927. [Google Scholar] [CrossRef]
- Li, C.Y.; Jiang, L.Y.; Chen, W.Y.; Li, K.; Sheng, H.Q.; Ni, Y.; Lu, J.X.; Xu, W.X.; Zhang, S.Y.; Shi, Q.X. CFTR Is Essential for Sperm Fertilizing Capacity and Is Correlated with Sperm Quality in Humans. Human Reproduction 2010, 25. [Google Scholar] [CrossRef]
- Inaba, Y.; Abe, R.; Geshi, M.; Matoba, S.; Nagai, T.; Somfai, T. Sex-Sorting of Spermatozoa Affects Developmental Competence of in Vitro Fertilized Oocytes in a Bull-Dependent Manner. Journal of Reproduction and Development 2016, 62. [Google Scholar] [CrossRef]
- Pinkwart, K.; Schneider, F.; Lukoseviciute, M.; Sauka-Spengler, T.; Lyman, E.; Eggeling, C.; Sezgin, E. Nanoscale Dynamics of Cholesterol in the Cell Membrane. Journal of Biological Chemistry 2019, 294. [Google Scholar] [CrossRef]
- Pöhnl, M.; Trollmann, M.F.W.; Böckmann, R.A. Nonuniversal Impact of Cholesterol on Membranes Mobility, Curvature Sensing and Elasticity. Nat Commun 2023, 14. [Google Scholar] [CrossRef]
- Lv, Y.Q.; Jin, Q.G.; Chen, X.; Wang, S.; Luo, X.T.; Han, Y.; Cheng, M.M.; Qu, X.L.; Zhang, Y.Y.; Jin, Y. Effects of Partially Replacing Glycerol with Cholesterol-Loaded Cyclodextrin on Protamine Deficiency, in Vitro Capacitation and Fertilization Ability of Frozen–Thawed Yanbian Yellow Cattle Sperm. Theriogenology 2022, 179. [Google Scholar] [CrossRef]
- Aly, A.S. Impact of Some Cryoprotectant Agents on Freezing Ability and Fertility of Rabbit Buck Spermatozoa. MSc., Faculty of Agriculture, Ain Shams University: Cairo, 2014.
- Davis, B.K. Timing of Fertilization in Mammals: Sperm Cholesterol/Phospholipid Ratio as a Determinant of the Capacitation Interval. Proc Natl Acad Sci U S A 1981, 78. [Google Scholar] [CrossRef]
- Mocé, E.; Tomás, C.; Blanch, E.; Graham, J.K. Effect of Cholesterol-Loaded Cyclodextrins on Bull and Goat Sperm Processed with Fast or Slow Cryopreservation Protocols. Animal 2014, 8. [Google Scholar] [CrossRef]
- Bernecic, N.C.; Zhang, M.; Gadella, B.M.; Brouwers, J.F.H.M.; Jansen, J.W.A.; Arkesteijn, G.J.A.; de Graaf, S.P.; Leahy, T. BODIPY-Cholesterol Can Be Reliably Used to Monitor Cholesterol Efflux from Capacitating Mammalian Spermatozoa. Sci Rep 2019, 9. [Google Scholar] [CrossRef]
- Batissaco, L.; de Arruda, R.P.; Alves, M.B.R.; Torres, M.A.; Lemes, K.M.; Prado-Filho, R.R.; de Almeida, T.G.; de Andrade, A.F.C.; Celeghini, E.C.C. Cholesterol-Loaded Cyclodextrin Is Efficient in Preserving Sperm Quality of Cryopreserved Ram Semen with Low Freezability. Reprod Biol 2020, 20. [Google Scholar] [CrossRef]
- LaVelle, G.; Cairo, B.; Barfield, J.P. Effect of Cholesterol-Loaded Cyclodextrin Treatment of Bovine Sperm on Capacitation Timing. Reproduction in Domestic Animals 2023, 58. [Google Scholar] [CrossRef]
- Oliveira, C.H.; Vasconcelos, A.B.; Souza, F.A.; Martins-Filho, O.A.; Silva, M.X.; Varago, F.C.; Lagares, M.A. Cholesterol Addition Protects Membrane Intactness during Cryopreservation of Stallion Sperm. Anim Reprod Sci 2010, 118. [Google Scholar] [CrossRef]
- Paradis, S.; Leoni, V.; Caccia, C.; Berdeaux, A.; Morin, D. Cardioprotection by the TSPO Ligand 4′-Chlorodiazepam Is Associated with Inhibition of Mitochondrial Accumulation of Cholesterol at Reperfusion. Cardiovasc Res 2013, 98. [Google Scholar] [CrossRef]
- Yu, W.; Gong, J.S.; Ko, M.; Garver, W.S.; Yanagisawa, K.; Michikawa, M. Altered Cholesterol Metabolism in Niemann-Pick Type C1 Mouse Brains Affects Mitochondrial Function. Journal of Biological Chemistry 2005, 280. [Google Scholar] [CrossRef]
- Martin, L.A.; Kennedy, B.E.; Karten, B. Mitochondrial Cholesterol: Mechanisms of Import and Effects on Mitochondrial Function. J Bioenerg Biomembr 2016, 48. [Google Scholar] [CrossRef]
- Lone, S.A. Possible Mechanisms of Cholesterol-Loaded Cyclodextrin Action on Sperm during Cryopreservation. Anim Reprod Sci 2018, 192. [Google Scholar] [CrossRef]
- Ferreira, J.J.; Cassina, A.; Irigoyen, P.; Ford, M.; Pietroroia, S.; Peramsetty, N.; Radi, R.; Santi, C.M.; Sapiro, R. Increased Mitochondrial Activity upon CatSper Channel Activation Is Required for Mouse Sperm Capacitation. Redox Biol 2021, 48. [Google Scholar] [CrossRef]
- Giaccagli, M.M.; Gómez-Elías, M.D.; Herzfeld, J.D.; Marín-Briggiler, C.I.; Cuasnicú, P.S.; Cohen, D.J.; Da Ros, V.G. Capacitation-Induced Mitochondrial Activity Is Required for Sperm Fertilizing Ability in Mice by Modulating Hyperactivation. Front Cell Dev Biol 2021, 9. [Google Scholar] [CrossRef]
- Hidalgo, D.M.; Romarowski, A.; Gervasi, M.G.; Navarrete, F.; Balbach, M.; Salicioni, A.M.; Levin, L.R.; Buck, J.; Visconti, P.E. Capacitation Increases Glucose Consumption in Murine Sperm. Mol Reprod Dev 2020, 87. [Google Scholar] [CrossRef]
- Sansegundo, E.; Tourmente, M.; Roldan, E.R.S. Energy Metabolism and Hyperactivation of Spermatozoa from Three Mouse Species under Capacitating Conditions. Cells 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Carrageta, D.F.; Guerra-Carvalho, B.; Sousa, M.; Barros, A.; Oliveira, P.F.; Monteiro, M.P.; Alves, M.G. Mitochondrial Activation and Reactive Oxygen-Species Overproduction during Sperm Capacitation Are Independent of Glucose Stimuli. Antioxidants 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Sohn, J.; Lin, H.; Fritch, M.R.; Tuan, R.S. Influence of Cholesterol/Caveolin-1/Caveolae Homeostasis on Membrane Properties and Substrate Adhesion Characteristics of Adult Human Mesenchymal Stem Cells. Stem Cell Res Ther 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Tomás, C.; Blanch, E.; Fazeli, A.; Mocé, E. Effect of a Pre-Freezing Treatment with Cholesterol-Loaded Cyclodextrins on Boar Sperm Longevity, Capacitation Dynamics, Ability to Adhere to Porcine Oviductal Epithelial Cells in Vitro and DNA Fragmentation Dynamics. Reprod Fertil Dev 2013, 25. [Google Scholar] [CrossRef]
- Medeiros, C. Bovine Sperm Interaction with Oviductal Epithelial Cells. Ph.D., University of Wisconsin-Madison, 1999.
- Petrunkina, A.M.; Waberski, D.; Günzel-Apel, A.R.; Töpfer-Petersen, E. Determinants of Sperm Quality and Fertility in Domestic Species. Reproduction 2007, 134. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Martinez, H. Role of the Oviduct in Sperm Capacitation. Theriogenology 2007, 68. [Google Scholar] [CrossRef] [PubMed]
- Vishwanath, R.; Moreno, J.F. Review: Semen Sexing - Current State of the Art with Emphasis on Bovine Species. Animal 2018, 12. [Google Scholar] [CrossRef] [PubMed]
- De Toni, L.; Sabovic, I.; De Filippis, V.; Acquasaliente, L.; Peterle, D.; Guidolin, D.; Sut, S.; Di Nisio, A.; Foresta, C.; Garolla, A. Sperm Cholesterol Content Modifies Sperm Function and Trpv1-mediated Sperm Migration. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
Figure 1.
(Experiment 1). Impact of cholesterol-loaded cyclodextrin (CLC) level on post-thaw bovine sperm motility. Post-thaw motility was assessed at three time points (0, 30, and 60 min.) and then averaged (n=5). All comparisons were made to 0 CLC. Main effects of CLC level and time exhibited a significant effect (p < 0.01) for both traits, while the interaction between CLC and time was not significant for either trait. A) Post-thaw total motility (%). B) Post-thaw progressive motility (%). All values are expressed as means ± standard error of the mean. Asterisks indicate significant differences compared to the 0 CLC, while NS indicates no difference (p > 0.05).
Figure 1.
(Experiment 1). Impact of cholesterol-loaded cyclodextrin (CLC) level on post-thaw bovine sperm motility. Post-thaw motility was assessed at three time points (0, 30, and 60 min.) and then averaged (n=5). All comparisons were made to 0 CLC. Main effects of CLC level and time exhibited a significant effect (p < 0.01) for both traits, while the interaction between CLC and time was not significant for either trait. A) Post-thaw total motility (%). B) Post-thaw progressive motility (%). All values are expressed as means ± standard error of the mean. Asterisks indicate significant differences compared to the 0 CLC, while NS indicates no difference (p > 0.05).
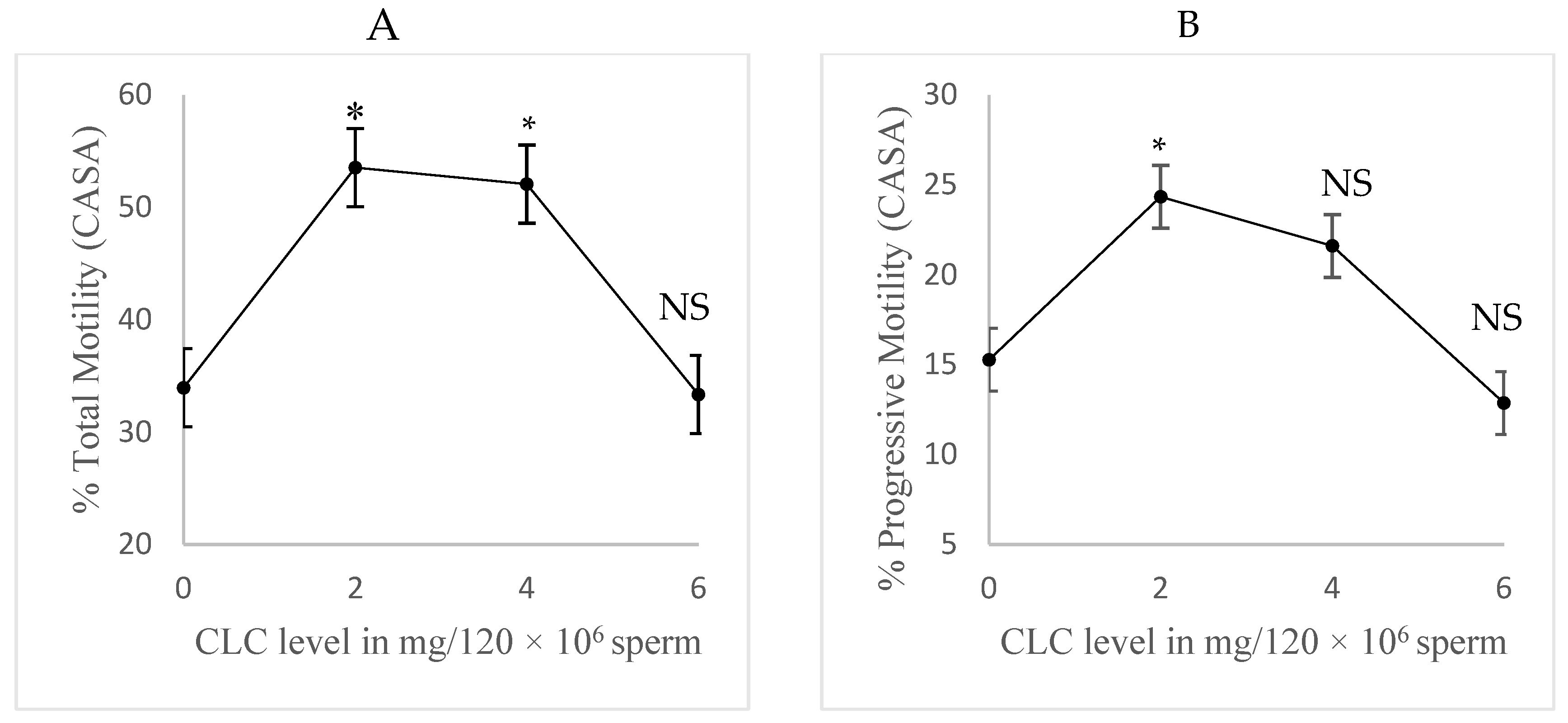
Figure 2.
(Experiment 3). Comparison of cholesterol-loaded cyclodextrin (CLC) addition to raw semen versus post-dilution treatment. Each sample received 2 mg CLC per 120 × 106 sperm cells (n=4). Post-dilution samples were treated after initial dilution to 120 × 106 sperm/mL, then further diluted to 30 x 106 sperm/mL using a commercial Tris buffer with egg yolk. The raw semen group received CLC directly before dilution, then diluted to 30 x 106 sperm/mL with the same diluent. Post-thaw motility was evaluated at 0, 30, and 60 minutes and then averaged. Values are mean ± SEM. No significant difference in sperm motility was detected, indicating CLC can be directly applied to raw semen for industrial use.
Figure 2.
(Experiment 3). Comparison of cholesterol-loaded cyclodextrin (CLC) addition to raw semen versus post-dilution treatment. Each sample received 2 mg CLC per 120 × 106 sperm cells (n=4). Post-dilution samples were treated after initial dilution to 120 × 106 sperm/mL, then further diluted to 30 x 106 sperm/mL using a commercial Tris buffer with egg yolk. The raw semen group received CLC directly before dilution, then diluted to 30 x 106 sperm/mL with the same diluent. Post-thaw motility was evaluated at 0, 30, and 60 minutes and then averaged. Values are mean ± SEM. No significant difference in sperm motility was detected, indicating CLC can be directly applied to raw semen for industrial use.
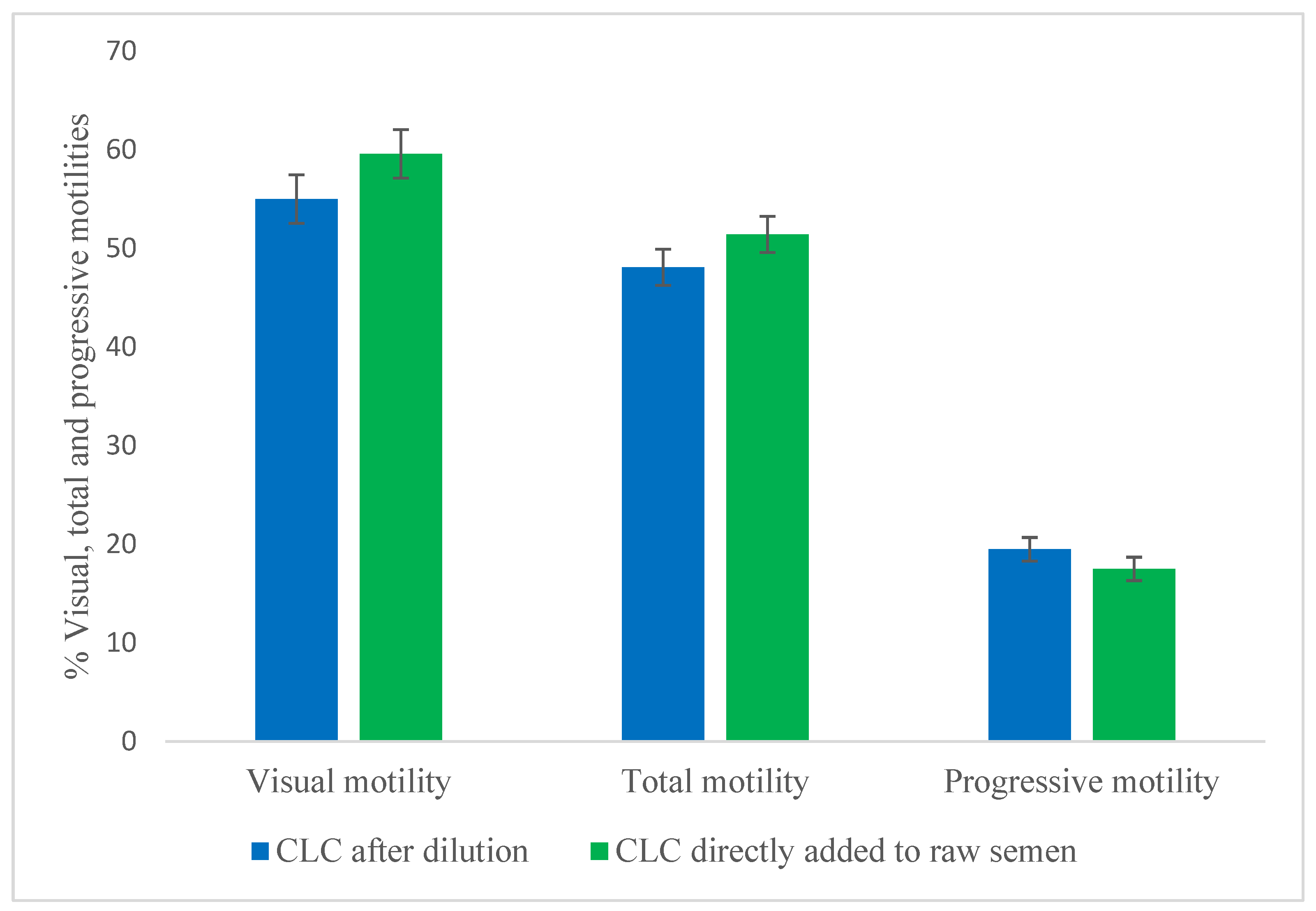
Table 3.
Flow cytometric analyses of frozen bovine semen treated with cholesterol-loaded cyclodextrin (CLC)*.
Table 3.
Flow cytometric analyses of frozen bovine semen treated with cholesterol-loaded cyclodextrin (CLC)*.
| CLC-level (mg/120 × 106 sperm cells) | Subpopulation with intact membranes and acrosomes (%) | Subpopulation with viable un-capacitated sperm cells (%) | Mitochondrial potential (ratio of JC1 orange to green florescence) |
|---|---|---|---|
| 0 | 20.9 ± 2.9 | 20.4 ± 3.7 | 0.99 ± 0.04 |
| 2 | 30.5 ± 2.9 | 32.8 ± 3.7 | 0.91 ± 0.04 |
* The CLC-treated group received 2 mg/120 × 106 sperm cells and was compared to the no CLC control (n=5, with two straws evaluated per sample). Data is presented as mean ± standard error of the mean. The main effect of CLC treatment was not significant for all traits.
Table 4.
Relative intracellular calcium flow cytometric analyses of bovine frozen-thawed semen treated with cholesterol-loaded cyclodextrin (live subpopulation only)a.
Table 4.
Relative intracellular calcium flow cytometric analyses of bovine frozen-thawed semen treated with cholesterol-loaded cyclodextrin (live subpopulation only)a.
| CLC level (mg/67x106 sperm cells) | No Ionomycin | Ionomycin | overall |
|---|---|---|---|
| 0 | 569 ± 129 | 1476** ± 129 | 1022 ± 91 |
| 2 | 622 ± 129 | 1491** ± 129 | 1056 ± 91 |
| Overall | 595 ± 105 | 1484 ± 105 |
a The cholesterol-loaded cyclodextrin (CLC) group received 2 mg per 120 × 106 sperm cells and was compared to the no-CLC control (n=5, with two straws evaluated per sample). Data are presented as mean ± standard error of the mean. The main effect of CLC was not significant, whereas ionomycin addition significantly (p < 0.01) increased calcium uptake. ** indicates p < 0.01 between the no-ionomycin and ionomycin-triggered groups.
Table 5.
Impact of cholesterol-loaded cyclodextrin (CLC) on the binding ability of frozen-thawed bovine sperm cells to oviduct cellsa.
Table 5.
Impact of cholesterol-loaded cyclodextrin (CLC) on the binding ability of frozen-thawed bovine sperm cells to oviduct cellsa.
| Coincubation time (h) | CLC-level (mg/120 × 106 sperm cells) | Number of viable bound sperm cells/ area of 1.65x106 µm2 oviduct cells |
|---|---|---|
| 0.5 | 0 | 209 ± 15 |
| 2 | 424*** ± 15 | |
| 3 | 0 | 118 ± 15 |
| 2 | 267*** ± 15 | |
| 7 | 0 | 70 ± 15 |
| 2 | 127* ± 15 | |
| 20 | 0 | 28 ± 15 |
| 2 | 63 ± 15 |
a The CLC-treated group received 2 mg/120 × 106 in comparison to no CLC (n=5, three replicates). Data is presented as mean ± standard error of the mean. The main effects of both time and CLC were significant (p < 0.01), and the interaction between CLC and time was also significant (p < 0.01). Asterisks within columns indicate statistically significant differences between the CLC-treated group and the control at the same time point, with * indicating p < 0.05 and *** indicating p < 0.0001.
Table 6.
In-vitro fertilizing ability and pronuclei formation rate of frozen-thawed bovine sperm cells treated with Cholesterol-Loaded Cyclodextrin (CLC)a.
Table 6.
In-vitro fertilizing ability and pronuclei formation rate of frozen-thawed bovine sperm cells treated with Cholesterol-Loaded Cyclodextrin (CLC)a.
| Coincubation time (h) | CLC-level (mg/120 × 106 sperm cells) | Penetration (%) | Overall | Pronuclei formation/penetrated oocytes (%) | Overall |
|---|---|---|---|---|---|
| 4 | 0 | 72 ± 4 | 72 ± 3 | 4 ± 2 | 3 ± 2 |
| 2 | 73 ± 4 | 2 ± 2 | |||
| 8 | 0 | 73 ± 3 | 77 ± 2 | 53 ± 7 | 47*** ± 5 |
| 2 | 81 ± 3 | 41 ± 7 | |||
| 12 | 0 | 81 ± 4 | 82* ± 2 | 66 ± 7 | 69*** ± 5 |
| 2 | 83 ± 4 | 72 ± 7 |
a The CLC-treated group received 2 mg/120 × 106 in comparison to no CLC (n=5, three replicates). The total number of examined oocytes was 897. Data is presented as mean ± standard error of the mean. The main effect of CLC was not significant for either trait. Coincubation time had no significant effect on penetration rate but significantly affected pronuclei formation rate (p < 0.0001). The interaction between CLC and coincubation time was not significant for both traits. Asterisks within a column indicate statistically significant differences compared to the 4 h group (*p < 0.05; ***p < 0.0001).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



