
Submitted:
03 December 2025
Posted:
04 December 2025
You are already at the latest version
Abstract
Background: The three Synuclein family members (α-, β-, and γ-synuclein) are presynaptic proteins that regulate synaptic vesicle trafficking and thereby influence neurotransmitter release. Synucleins belong to a class of intrinsically disordered proteins and are prone to aggregation into pathological deposits, which may impair their physiological synaptic functions. Knockout (KO) mouse lines, commonly used to model synuclein depletion in the nervous system, reveal a range of phenotypes with different motor and behavioral deficits. However, given the high sequence homology and functional interplay among the three synucleins, the specific contribution of each family member to these phenotypes remains poorly understood. Objective: In this study, we conducted a comparative phenotypic analysis of γ-synuclein KO, α- and β-synuclein KO, and αβγ-synuclein KO mice. Methods: Mice were subjected to a battery of behavioral tests assessing motor activity and coordination, anxiety-like behavior, and spatial learning and memory. Synaptic vesicle proteins were analyzed in brain tissues using Western blotting. Results: We observed that knocking out γ-synuclein but not α- and β-synucleins reduces mouse lifespan and leads to sustained reduction in muscle strength implicating that γ-synuclein is essential for neuromuscular function. Another consequence of γ-synuclein deficiency is altered anxiety-like behavior manifested as a diminished aversive response, while exploratory behavior and memory remain intact. The triple KO mice mirror γ-synuclein KO mice in some behavioral changes, including shortened lifespan, reduced muscle strength, and decreased anxiety-like behavior. However, the triple KO mice additionally exhibit hyperactivity, which is not present in the other groups. No changes in synaptic vesicle marker levels were detected, indicating that the observed motor and behavioral abnormalities are not attributable to impaired synaptic connectivity. Conclusions: Taken together, these findings demonstrate nonredundant functions of individual synuclein family members and highlight a distinct role of γ-synuclein in regulating motor performance and behavioral responses.
Keywords:
γ-synuclein
; knockout (KO) mice
; motor performance
; behavioral phenotyping
; aging
1. Introduction
Synucleins are highly conserved cytosolic proteins that play an important role in regulating synaptic transmission in the central nervous system [1,2,3]. The synuclein family comprises three structurally similar proteins, α-, β-, and γ-synucleins, predominantly expressed in neural cells, and the patterns of their localization in different brain regions partially overlap [4,5,6]. Synucleins share structural similarities, but the functional specialization of individual synucleins remains poorly understood.
α-Synuclein is the most extensively studied member of the family because it is involved in the pathogenesis of several neurodegenerative diseases. It is the major component of Lewy bodies in Parkinson’s disease and dementia with Lewy bodies. α-Synuclein is also found in senile plaques in Alzheimer’s disease [7,8,9]. Similarly, γ-synuclein has the capacity to aggregate forming pathological deposits identified in amyotrophic lateral sclerosis and dementia with Lewy bodies [10,11,12,13]. Moreover, γ-synuclein is implicated in oncogenesis, because its expression is increased in various types of tumors [14,15,16,17]. In contrast, β-synuclein is generally considered non-amyloidogenic due to the absence of the NAC domain in its sequence [18,19,20]. Furthermore, β-synuclein can inhibit α-synuclein aggregation [21,22,23].
Transgenic mice lacking one, two, or all three synuclein family proteins are widely used to study biological functions of synucleins [24,25,26,27]. Some studies report that deletion of one or several synuclein genes does not result in significant impairments: animals remain fertile, exhibit no expressed morphological or behavioral deviations, and their lifespan is comparable to the lifespan of wild-type (WT) mice [28]. However, other studies report shortened lifespan and presence of a neurological phenotype with sensory dysfunction, such as age-related retinal degeneration and pronounced neurological deficits in triple KO mice [25,29].
The absence of overt phenotypic changes following synuclein inactivation suggests an auxiliary role of these proteins in nervous system physiology, with deficits becoming apparent only under specific conditions, such as during aging [29]. Indeed, aged synuclein-deficient mice exhibit statistically significant, although physiologically subthreshold alterations in the nigrostriatal system, including reduced dopamine levels [24]. Given that aging is accompanied by progressive functional decline in the central nervous system, the loss of one or more synucleins may exert a cumulative effect that becomes evident only at advanced age [25,29]. Supporting this hypothesis, more pronounced impairments have been reported in αβ-synuclein double KO mice compared to α-synuclein KO mice. Also, αβγ-synuclein triple KO mice demonstrated a more severe phenotype than αβ-synuclein double KOs [24,28].
Although structural similarity of synucleins implies potential functional redundancy, existing data indicate at least partial specialization within the synuclein family. β-Synuclein appears to play a primary role in regulating motor function: its absence leads to substantial coordination impairments, while additional loss of α- or γ-synuclein does not further worsen this phenotype [28]. Deletion of the α-synuclein gene is more strongly associated with behavioral alterations, including increased activation thresholds, reduced impulsivity [30], and impaired learning and memory [26,31]. Notably, α-synuclein KO mice do not differ from WT controls in anxiety-related behaviors [32]. In contrast, γ-synuclein KOs display reduced anxiety, enhanced exploratory activity, and accelerated adaptation to novel environments, suggesting that γ-synuclein may exert an inhibitory influence on cognitive processes [33]. These animals also exhibit enhanced working memory compared to WT mice, while long-term spatial memory performance in the Morris water maze remains unchanged [33]. Taken together, these findings indicate that α-synuclein KOs and γ-synuclein KOs exhibit oppositely directed alterations in memory and certain cognitive functions.
To elucidate the specific role of γ-synuclein in the development of motor and behavioral changes observed in synuclein-deficient mice, we performed a comparative analysis of motor and cognitive functions in γ-synuclein KO mice, α- and β-synuclein double KO mice, and αβγ-synuclein triple KO mice at 6, 13, and 24 months of age. Additionally, we quantified levels of some synaptic protein markers in brain tissues from these animals. The results may help clarify the role of γ-synuclein in neurodegenerative and psychiatric disorders characterized by pathological protein aggregation and consequent depletion of the functional protein pool in the cell, such as Parkinson’s disease and dementia with Lewy bodies. Furthermore, these results may inform therapeutic approaches aimed at preserving or compensating γ-synuclein function [16,34,35].
2. Materials and Methods
2.1. Animals
This study used mouse lines with constitutive knockout of synuclein genes: JAXMice 028559 B6(Cg)-Sncatm1.2Vlb/J (SncaΔflox, Snca Δflox KO) with Snca deletion [36], JAXMice 006390 B6.129-Sncbtm1.1Sud/J with Sncb deletion [24], and JAXMice 008843 B6.129P2-Sncgtm1Vlb/J with Sncg deletion [37]. All KO lines were maintained on a C57BL/6J background. α- and β-synuclein double KO mice, and αβγ-synuclein triple KO mice were generated by intercrossing the corresponding single KO lines. C57BL/6J mice served as a WT control group. Animal colonies were maintained under standard vivarium conditions at a temperature of 22 °C on a 12/12 h light/dark cycle. Each experimental group included at least 20 males. Exact sample sizes are indicated in the figure legends. All procedures adhered to the “Guidelines for accommodation and care of animals. Species-specific provisions for laboratory rodents and rabbits” (GOST 33216-2014) and complied with Directive 2010/63/EU on the protection of animals used for scientific purposes, and were approved by the local Institute Ethics Review Committee of the IPAC RAS (protocol No. 48, 15 January 2021). All reasonable measures were taken to minimize animal numbers and suffering. All mice were genotyped by PCR using DNA obtained from ear biopsies, as described previously [27].
2.2. Behavioral Testing
Animals from all four experimental groups (γ-synuclein KO, αβ-synuclein KO, αβγ-synuclein KO, and WT) underwent a standardized battery of motor tests at 6, 13, and 24 months of age, including the grip-strength test, inverted-grid hanging test, and accelerating rotarod. At 13 months, exploratory behavior and psychoemotional state were additionally assessed using the open field, Y-maze, elevated plus maze, and Morris water maze tests. Animals were placed in the testing room 30 minutes before each experiment for habituation. Motor and cognitive tests were conducted under lighting conditions comparable to standard housing (~20 lx). The open field, Y-maze, and elevated plus maze tests were video-recorded and analyzed using EthoVision XT 11.5 software (Noldus, Wageningen, Netherlands). The Morris water maze test was analyzed using ANY-maze software (ANY-maze, Dublin, Ireland).
2.2.1. Grip Strength Test
Mice were held by the tail and allowed to grasp a metal grid (7×12.1 cm, 6×6 mm spacing) attached to a grip strength meter (ALMEMO, Holzkirchen, Germany). The mean force across six trials was used for analysis [38].
2.2.2. Inverted Grid Test
Mice were placed on an inverted metal grid (33.6×34.3 cm, 1.3×1.3 cm spacing), held 50-60 cm above a padded surface. The latency to fall was recorded, with a maximum test duration of 60 s. Mice that fell earlier received two additional trials at 15-minute intervals. The best result of the three trials was used for analysis [39].
2.2.3. Accelerating Rotarod Test
Mice were trained one day before testing at a constant speed (4 rpm) for 10 min. On the test day, mice were placed on a rod accelerating from 4 to 40 rpm over 4 min followed by 1 min at maximum speed (Ugo Basile 7650, Gemonio, Italy). Each mouse completed four trials with 45-min intervals. The latency to fall was recorded [40].
2.2.4. Open Field Test
Mice explored a 40 × 40 cm arena with 40 cm high walls (OpenScience, Moscow, Russia) under diffuse illumination (15 lx center, 10 lx corners). Activity was recorded for 5 minutes (Basler acA1300-60 camera) and analyzed using EthoVision XT 11.5 software (Noldus, Wageningen, Netherlands). The following parameters were evaluated: total distance traveled, time spent in the center zone (20 × 20 cm), and number of center entries [41,42,43,44].
2.2.5. Y-Maze
The maze consisted of three arms (40 × 6 × 20 cm) arranged at 120° angles (OpenScience, Moscow, Russia). The test included two 5-min sessions separated by 30 min. In session 1, two arms were open; in session 2, all three were open. Preference for the novel arm was calculated as the percentage of entries into the new arm during the first 2 minutes of session 2. Total distance, and time spent in each arm were also recorded [45].
2.2.6. Elevated Plus Maze
The apparatus (OpenScience, Moscow, Russia) consisted of two open and two closed arms arranged at 90° around a central platform. Mice were placed in the center facing an open arm and allowed to explore for 5 minutes. Illumination at the center was 380-400 lx. Time and number of entries into open arms and distance and speed were recorded [46].
2.2.7. Morris Water Maze
A circular pool, 150 cm in diameter, (OpenScience, Moscow, Russia) was filled with water to a height 60 cm above the base; water temperature was 22 °C. A hidden platform (10 cm diameter) was submerged 1 cm below the surface. Mice were trained over 4 days (4 trials/day; max trial length 60 s). On day 5, a 90-s probe trial was performed without the platform. The time to locate the platform and the number of platform-zone crossings were analyzed. Swim speed and total distance served as measures of physical activity [47].
2.3. Survival Analysis
After behavioral testing, animals were maintained under standard vivarium conditions until natural death. Lifespan (in days) was used to generate Kaplan-Meier survival curves and perform survival analysis.
2.4. Analysis of Synaptic Markers
Dorsal striatum and cerebral cortex samples were homogenized in 2× Laemmli buffer and heat-denatured at 95 °C for 5 minutes. Proteins were separated on 14% SDS-PAGE gels and transferred to PVDF membranes (Hybond-P, Amersham, Sheffield, UK) by semi-dry transfer. Membranes were blocked with 4% non-fat dry milk in TBST (Tris-buffered saline with 0.1% Tween 20) and incubated with primary antibodies against synaptic proteins: synaptophysin (1:5000, S31620), synapsin IIa (1:10000, 610666), and amphiphysin (1:10000, A59420; all BD Biosciences; Transduction Laboratories, San Jose, CA, USA). HRP-conjugated secondary antibodies (1:4000, cat. 5178-2504, Bio-Rad, Hercules, CA, USA) were used for detection by enhanced chemiluminescence kit (ECL Plus, Thermo Fisher Scientific, Waltham, MA, USA) and X-ray film (Thermo Scientific, Waltham, MA, USA). Membranes were subsequently reprobed with α-tubulin antibodies (1:10000, Proteintech, Rosemont, IL, USA) as a loading control.
2.5. Statistical Analysis
Statistical analysis was performed using GraphPad Prism 6 (GraphPad Software, San Diego, CA, USA). Data are presented as mean ± SEM unless otherwise indicated. The numbers inside the bars represent the number of animals per group. Normality was assessed using the D’Agostino-Pearson test. For normally distributed data, one-way ANOVA was applied; otherwise, the Kruskal-Wallis test was used. The statistical test and p-value threshold for each analysis are specified in the figure legends. Data analysis was performed in a blinded manner: the experimenters were unaware of group assignments until completion of the analysis.
3. Results
3.1. γ-Synuclein KO Mice Exhibit Reduced Lifespan
To assess the contribution of γ-synuclein to organismal viability, we conducted a comparative survival analysis of Sncg knockout (γ-KO) mice relative to WT controls. Two additional groups were included for comparison: mice with a double knockout of Snca and Sncb (αβ-KO), which retain an intact Sncg gene, and mice lacking all three synuclein genes (αβγ-KO). Analysis of the Kaplan-Meier curves demonstrated a statistically significant reduction in lifespan in γ-KO mice and in synuclein null mice (αβγ-KO), whereas αβ-KOs did not differ from WT controls (Figure 1). Median survival (age at which 50% of animals remained alive) was 763 days in the γ-KO group, 859 days in the αβγ-KO group, 910 days in the αβ-KO group, and 913 days in the WT group. These findings indicate that loss of γ-synuclein and the combined loss of all synucleins negatively affect longevity, implicating the importance of synucleins in maintaining organismal viability.
3.2. γ-Synuclein KO Mice Exhibit Decreased Muscle Strength
Motor functions and selected behavioral responses were evaluated in all KO groups compared to WT using the grip strength test, inverted grid test, and accelerating rotarod.
At 6 months of age, which corresponds to young adult mice, γ-synuclein KO and αβγ-synuclein KO mice showed reduced forelimb grip strength compared with WT controls (Figure 2A). In contrast, αβ-synuclein KO mice, which express γ-synuclein as the only remaining synuclein, displayed normal grip strength. Overall, mice lacking γ-synuclein (γ-synuclein KO and αβγ-synuclein KO) exhibited an approximately 30% reduction in strength relative to WT animals. Thus, γ-synuclein plays a key role in maintaining muscle strength, whereas α- and β-synucleins do not appear to compensate for its loss. In the inverted grid test, no statistically significant differences were observed among groups, although γ-synuclein KOs showed a trend toward shorter hang times (Figure 2B). These results suggest that short-term muscle endurance under dynamic load distribution is largely preserved in KO young adult mice. In the accelerating rotarod test, αβ-synuclein KOs exhibited a 28% reduction in fall latency compared with WT (Figure 2C) indicating reduced motor coordination and/or endurance. However, γ-synuclein KOs performed similarly to WT controls, which suggests that α- and β-synuclein, but not γ-synuclein, contribute more prominently to rotarod performance. Interestingly, αβγ-synuclein null mice showed increased fall latency relative to WT, possibly reflecting compensatory mechanisms resulting from the complete loss of all synucleins or increased activity in these animals (see below).
At 13 months (middle-aged mice), γ-synuclein KO and αβγ-synuclein KO mice again demonstrated reduced grip strength compared to WT mice, which is consistent with the 6-month findings (Figure 3A). αβ-synuclein KO mice were comparable to WT in grip strength. Also at 13 months, γ-synuclein KO mice exhibited a statistically significant reduction in hang time in the inverted grid test, whereas αβ-synuclein KOs and αβγ-synuclein KOs did not differ from WT controls (Figure. 3B). This finding indicates progressive impairment of motor endurance in γ-synuclein KO mice with age. In the accelerating rotarod test, only αβ-synuclein KO mice showed a trend toward reduced fall latency, and in contrast to the 6-month time point, αβγ-synuclein KO mice did not show increased performance (Figure 3C).
By 24 months (old age in mice), γ-synuclein KO mice no longer differed from WT animals in grip strength, although αβγ-synuclein KO mice still showed significantly reduced performance (Figure 4A). Notably, WT animals demonstrated age-related declines in grip strength (p = 0.0040, mixed-effects analysis with Tukey’s correction for 13 vs. 24 months), consistent with normal aging. These findings suggest that γ-synuclein loss reduces muscle strength at younger ages but does not exacerbate age-related decline, whereas complete synuclein deficiency has more severe long-term effects. In the inverted grid test, all KO groups demonstrated progressively reduced performance compared with WT (Figure 4B). In the accelerating rotarod test, the triple KO mice again showed increased fall latency, similar to the 6-month results, whereas γ-synuclein KO and αβ-synuclein KO mice did not differ from WT (Figure 4C). As in the grip strength test, WT mice demonstrated lower performance at 24 months (p = 0.0044, mixed-effects analysis with Tukey’s correction for 13 vs. 24 months).
3.3. γ-Synuclein KO Mice Display Reduced Anxiety-Like behavior and no Memory Impairments
To determine which behavioral responses beyond motor activity might be influenced by the loss of γ-synuclein we performed a series of tests assessing the general state of the nervous system and some higher neurological functions in middle-aged mice (13 months old). We first conducted the open field test. Overall locomotor and exploratory activity were not altered in γ-synuclein KO mice. Parameters such as the total distance traveled and number of center entries also did not differ from control values (Figure 5A, C). In contrast, both the αβ-synuclein KO and triple KO mice exhibited significantly higher values for these parameters compared to WT. Interestingly, the hyperactivity observed in the triple KO mice is consistent with their motor test results. In contrast, the double KO mice manifested hyperactivity only in the open field test. It is possible that the exploratory component of activity is more pronounced than pure locomotion in αβ-synuclein KO mice. This is indirectly supported by a trend toward increased time in the arena center observed in the αβ-synuclein KO group compared to other groups (Figure 5B).
We performed the Y-maze test to assess short-term memory. No group differences were found for novel-arm exploration, and all animals showed a high preference (>40%) for the previously unexplored arm (Figure 6A) indicating intact short-term memory across groups. A hyperactive phenotype was again evident in the triple and double KO mice, reflected by an increased total distance traveled during the test (Figure 6B), which was consistent with the open field results.
Long-term memory was examined using the Morris water maze. All groups demonstrated a progressive decrease in latency to find the platform during training, which is indicative of normal learning (Figure 7A). Nonetheless, KO mice appeared to learn more slowly during the early days of training. Notably, γ-synuclein KO and αβγ-synuclein KO mice showed significantly lower swimming speeds (Figure 7C), which may indicate reduced muscle strength and endurance or diminished aversive response to water.
Regarding long-term memory, KO mice did not differ from WT controls. During the probe trial on day 5, all groups located the former platform area with similar speed and made a comparable number of crossings (Figure 7B, D). Thus, synucleins are likely not required for short- or long-term memory formation. Performance differences in the Morris water maze are more plausibly attributed to alterations in motor function or anxiety levels.
To assess anxiety-like behavior more specifically, we performed the elevated plus maze (EPM) test. γ-Synuclein KO and triple KO mice made more entries and spent significantly more time in the open and more stress-inducing arm of the maze compared to WT controls (Figure 8A, B), which indicates reduced anxiety-like behavior. As in the previous tests, triple KO mice exhibited a hyperactive phenotype—the total distance traveled was markedly greater compared to other groups (Figure 8C).
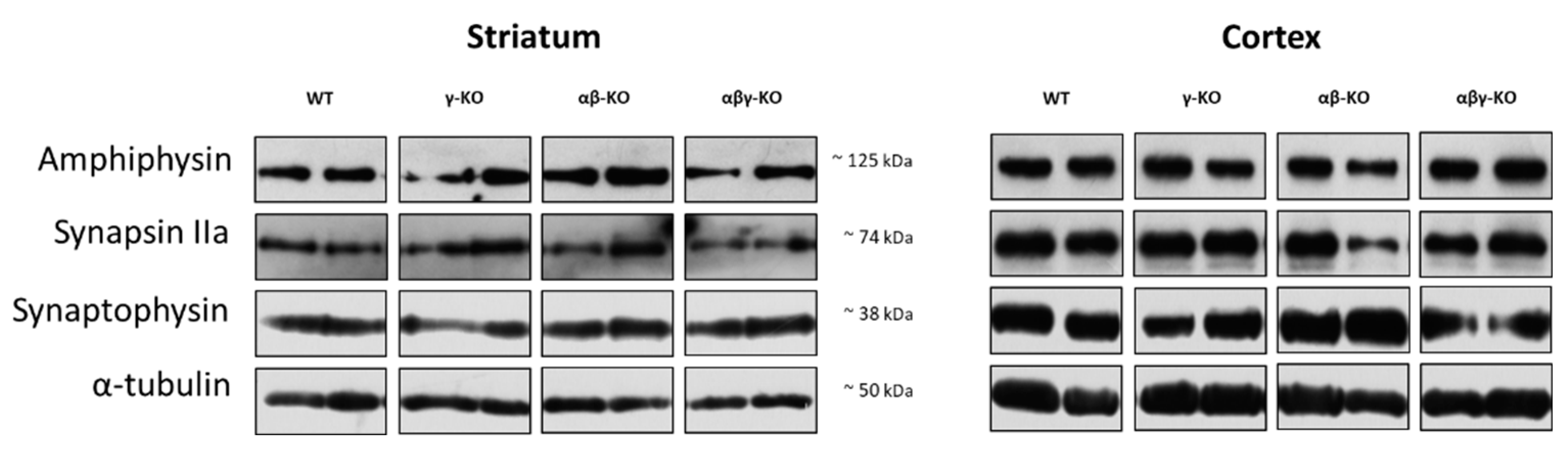
3.4. Absence of Synucleins Does not Alter Synaptic Marker Levels in the Striatum and Cortex
Synuclein functions are closely linked to synaptic transmission, and its disruption could potentially underlie the observed phenotypes. Therefore, we analyzed several synaptic proteins involved in neurotransmission. We examined the dorsal striatum and cerebral cortex—key structures responsible for controlling motor and cognitive functions. Tissue samples from 13-month-old animals were analyzed by Western blotting.
Synaptophysin, a synaptic vesicle marker involved in exocytosis [48,49,50], is widely used as an indicator of synaptic density. Synaptophysin levels in both striatum and cortex did not differ significantly between KO and WT mice (Figure 9, Figure S1).
Synapsin II is a neuron-specific phosphoprotein that maintains the reserve pool of neurotransmitter vesicles and mobilizes them when necessary, especially during high-frequency stimulation [51,52,53]. The IIa isoform of this protein is the principal regulator of vesicle dynamics and has a higher affinity for neurotransmitter vesicles. The quantity of synapsin IIa showed no genotype-related differences between KO and WT. (Figure 9, Figure S1).
Levels of Amphiphysin, which plays a major role in synaptic vesicle endocytosis [54,55,56], also did not differ among groups (Figure 9, Figure S1). Taken together, these findings indicate that the absence of synucleins does not significantly affect synapse quantity, vesicle turnover, and synaptic plasticity in the examined brain regions implicating preserved synaptic structure.
4. Discussion
The results obtained in this study reveal complex and multidirectional patterns of behavioral changes in γ-synuclein KO mice. Despite high homology and overlapping expression patterns among α-, β-, and γ-synucleins, our findings indicate that these proteins possess specialized, nonredundant functions that manifest under specific physiological conditions.
One of the most striking observations was lifespan reduction in γ-synuclein-deficient mice (γ-synuclein KO and αβγ-synuclein KO), whereas double αβ-synuclein KO mice displayed normal survival (Figure 1). Earlier work conducted on C57BL/6-background knockouts did not detect lifespan differences [28], but that study was limited to a 2-year observation window (~750 days). Extending the observation period to 3 years (>1000 days) in the present study revealed a late-onset phenotype, suggesting that γ-synuclein may uniquely influence long-term survival and that this effect becomes apparent only in advanced age.
A prominent characteristic of γ-synuclein deficiency was reduced muscle strength consistently observed in young (6-month-old) and middle-aged (13-month-old) mice (Figure 2-3). However, this deficit was no longer distinguishable by 24 months from the age-related deficit in WT mice. The early reduction in muscle strength may reflect the importance of γ-synuclein in synaptic transmission in central and neuromuscular synapses [37]. The disappearance of differences in older animals could indicate physiological compensatory mechanisms developing with age, possibly mediated by other synuclein family members [24,57]. Supporting this, mice lacking all three synucleins displayed reduced grip strength across all ages, including 24 months, suggesting a failure of compensatory mechanisms in the complete absence of synucleins (Figure 2-4A).
In the inverted grid test, evaluating endurance and motor coordination along with muscle strength, KO mice displayed a progressive, age-dependent decline (Figure 2-4B). At 6 months, no deviations were observed in any of the KO groups compared to control. At 13 months, a decrease in grid retention time was observed in the γ-synuclein KO group, and at 24 months—in all three groups of KO mice, particularly in the γ-synuclein KO group. These findings differ partially from the findings reported by Connor-Robson et al. [28] who found no reduction in 24-month-old γ-synuclein KO mice, but they are consistent with their observations for αβ-synuclein KO and αβγ-synuclein KO animals. Potential explanations for this discrepancy include subtle variations in diet, environmental conditions, or genetic drift within the C57BL/6 mouse line.
In contrast, γ-synuclein deficiency did not affect performance on the accelerating rotarod—a task that is more reliant on coordination and motor learning than muscle strength. In all age periods, γ-synuclein KOs did not differ from controls. A decrease in latency to fall was noted for the αβ-synuclein KO group at 6 months compared to WT, but these differences converged as the animals aged. (Figure 2-4C). The triple KOs in our study performed significantly better than WTs at 24 months, in contrast with the decreased performance reported earlier [28]. We observed this enhanced performance in αβγ-synuclein KO mice across all age groups and attribute it to the general hyperactivity of these animals, previously reported and evident in other behavioral tests [27]. A key methodological distinction of this triple KO line is a different α-KO allele used in its generation [36] that lacks the unintended disruption of the neighboring Mmrn1 gene present in the previous line [58]. Direct comparisons using both α-KO lines are required to clarify these differences.
Exploratory and cognitive functions were assessed for middle-aged mice at 13 months. At this age, the motor phenotype was already well developed, but age-related deficits had not yet fully emerged. The most prominent behavioral alterations included reduced anxiety-like behavior in γ-synuclein KO mice and increased exploratory activity in αβ- and αβγ-synuclein KO mice in the open-field test (Figure 5, 8). γ-synuclein KOs consistently spent more time in the open arms of the elevated plus maze indicating diminished aversion to stress-associated stimuli. A similar reduction in anxiety-like behavior was observed in the synuclein null group. Nevertheless, this reduction was accompanied by a pronounced hyperactivity phenotype manifested by increased locomotor activity (Figure 8C). Across multiple behavioral assays, augmented locomotor activity was characteristic of αβ- and αβγ-synuclein KO mice, whereas reduced anxiety-like behavior was specifically associated with γ-synuclein deficiency. These findings suggest that γ-synuclein uniquely contributes to modulating anxiety-related behaviors, while locomotor hyperactivity emerges only when two or all three synuclein family members are absent.
Assessment of short-term and long-term memory in the Y-maze and Morris water maze tests revealed no significant cognitive impairments in any KO group (Figure 6, 7). Although KO mice exhibited slower learning during early water maze training, this likely reflects reduced motivation, altered aversion to water, and changes in the emotional state rather than memory dysfunction. This interpretation is consistent with prior studies showing that synuclein deletion influences stress reactivity, dopaminergic signaling, and initiation thresholds for behavioral responses [33,59,60,61]. Reduced anxiety in γ-synuclein KO mice previously documented in younger animals [33,62] appears to be a stable phenotypic manifestation of γ-synuclein loss.
A particularly notable finding is the preserved expression of synaptic markers (synaptophysin, synapsin IIa, and amphiphysin) in both the striatum and cerebral cortex across all KO groups (Figure 9). These results indicate that synaptic density and vesicular neurotransmitter turnover remain intact despite the absence of synucleins. Previously published data reported decreased dopamine and amphiphysin levels in aged α-synuclein KO mice but not in γ-synuclein KO mice [63]. In the present study, we confirmed that 13-month-old γ-synuclein KO mice show no changes in synaptic marker levels. Moreover, no differences were detected in both the striatum and cerebral cortex of same age αβ-synuclein KO or αβγ-synuclein KO mice. Thus, the behavioral and motor deviations observed in the KO lines are more likely to result not from structural synaptic abnormalities but from more subtle physiological alterations in neurotransmission dynamics.
In summary, our phenotypic analysis of γ-synuclein KO mice compared to mice that retain γ-synuclein but lack the other two family members (α- and β-synuclein) and to mice completely lacking all three synucleins indicates that γ-synuclein is crucial for neuromuscular system functions. Moreover, loss of γ-synuclein leads to a sustained reduction in muscle strength. Another phenotypic consequence of γ-synuclein knockout in mice is change in anxiety-like behavior leading to a diminished response to aversive stimuli. This may suggest inability to adequately evaluate danger in the environment. At the same time, no other impairments of higher nervous functions, such as altered exploratory activity or memory deficits, were observed. In the absence of all three synucleins, some behavioral changes replicate changes in γ-synuclein KO mice, such as decreased muscle strength and reduced anxiety. However, triple KOs also exhibit additional abnormalities, such as hyperactivity, which is not present in other KO groups.
In the inverted grid test, no differences were observed in young mice, but significant impairments emerged with age. This may suggest gradual accumulation of subtle deficits in motor regulation detectable with age progression. However, differences between groups diminished with age in other assays, such as grip strength. These opposing results suggest that disruption of synuclein function does not produce a straightforward cumulative phenotype but instead triggers complex, multidirectional compensatory processes within the nervous system. Notably, even the complete absence of all three synucleins does not lead to overt physiological failure, yet it results in dysregulation of distinct, not directly linked behavioral aspects, such as motor performance and anxiety. These findings support the existence of specialized, non-overlapping functions of different synucleins and highlight a distinct role of γ-synuclein in regulating both motor performance and behavioral responses [33,64,65].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Original, unmodified images of X-ray films for Western blots.
Author Contributions
Conceptualization, M.S.K., and N.N.N.; methodology, R.K.O. and K.D.Ch.; validation, O.A.L., and M.S.K.; formal analysis, M.S.K.; investigation, I.S.S., O.A.M., V.N.Z., N.M.Yu., and A.A.L.; resources, M.S.K. and K.D.Ch.; data curation, M.S.K.; writing—original draft preparation, M.S.K. and I.S.S.; writing—review and editing, N.N.N.; visualization, I.S.S.; supervision, M.S.K. and N.N.N.; project administration, M.S.K. and N.N.N.; funding acquisition, M.S.K. and K.D.Ch. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the IPAC RAS State Target (№ FFSG-2024-0023 to M.S.K. and FFSG-2024-0020 to K.D.Ch.). The analysis of synaptic markers was supported by the Ministry of Science and Higher Education of the Russian Federation (agreement № 075-15-2025-558).
Institutional Review Board Statement
The animal study protocol was approved by the local Institute Ethics Review Committee of the IPAC RAS (protocol No. 48, 15 January 2021). All animal work was carried out in accordance with the “Guidelines for accommodation and care of animals. Species-specific provisions for laborato-ry rodents and rabbits” (GOST 33216-2014) in compliance with the principles enunciated in the Directive 2010/63/EU on the protection of animals used for scientific purposes.
Data Availability Statement
Data are contained within the current article and its supplementary material.
Acknowledgments
Animals were provided and supported by Bioresource Collection and Centre for Collective Use of IPAC RAS.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ANOVA | Analysis of variance |
| ECL | Enhanced chemiluminescence |
| EPM | Elevated plus maze |
| KO | Knockout |
| PVDF | Polyvinylidene fluoride |
| rpm | Revolutions per minute |
| SDS-PAGE | Sodium dodecyl sulfate–polyacrylamide gel electrophoresis |
| SEM | Standard error of the mean |
| TBST | Tris-buffered saline with Tween-20 |
| WT | Wild type |
| αβ-KO | double knockoutfor α- and β-synucleins (Snca−/−; Sncb−/−) |
| αβγ-KO | triple knockout for α-, β- and γ-synucleins (Snca−/−; Sncb−/−; Sncg−/−) |
| γ-KO | γ-synuclein knockout mice (Sncg−/−) |
References
- Carnazza, K.E.; Komer, L.E.; Xie, Y.X.; Pineda, A.; Briano, J.A.; Gao, V.; Na, Y.; Ramlall, T.; Buchman, V.L.; Eliezer, D.; et al. Synaptic Vesicle Binding of α-Synuclein Is Modulated by β- and γ-Synucleins. Cell Rep. 2022, 39, 110675. [Google Scholar] [CrossRef] [PubMed]
- Mechanisms of Alpha-Synuclein Toxicity: An Update and Outlook. In Progress in Brain Research; Elsevier, 2020; Vol. 252, pp. 91–129. ISBN 978-0-444-64260-8.
- Intracellular Dynamics of Synucleins. In International Review of Cell and Molecular Biology; Elsevier, 2015; Vol. 320, pp. 103–169. ISBN 978-0-12-802277-1.
- Baltic, S.; Perovic, M.; Mladenovic, A.; Raicevic, N.; Ruzdijic, S.; Rakic, L.; Kanazir, S. α-Synuclein Is Expressed in Different Tissues During Human Fetal Development. J. Mol. Neurosci. 2004, 22, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Burré, J. The Synaptic Function of α-Synuclein. J. Park. Dis. 2015, 5, 699–713. [Google Scholar] [CrossRef] [PubMed]
- Malatynska, E.; Pinhasov, A.; Crooke, J.; Horowitz, D.; Brenneman, D.E.; Ilyin, S.E. Levels of mRNA Coding for α-, β-, and γ-Synuclein in the Brains of Newborn, Juvenile, and Adult Rats. J. Mol. Neurosci. 2006, 29, 269–278. [Google Scholar] [CrossRef]
- Burré, J.; Sharma, M.; Südhof, T.C. Cell Biology and Pathophysiology of α-Synuclein. Cold Spring Harb. Perspect. Med. 2018, 8, a024091. [Google Scholar] [CrossRef]
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.-Y.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. α-Synuclein in Lewy Bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef]
- Uéda, K.; Fukushima, H.; Masliah, E.; Xia, Y.; Iwai, A.; Yoshimoto, M.; Otero, D.A.; Kondo, J.; Ihara, Y.; Saitoh, T. Molecular Cloning of cDNA Encoding an Unrecognized Component of Amyloid in Alzheimer Disease. Proc. Natl. Acad. Sci. 1993, 90, 11282–11286. [Google Scholar] [CrossRef]
- Ninkina, N.; Peters, O.; Millership, S.; Salem, H.; Van Der Putten, H.; Buchman, V.L. γ-Synucleinopathy: Neurodegeneration Associated with Overexpression of the Mouse Protein. Hum. Mol. Genet. 2009, 18, 1779–1794. [Google Scholar] [CrossRef]
- Peters, O.M.; Millership, S.; Shelkovnikova, T.A.; Soto, I.; Keeling, L.; Hann, A.; Marsh-Armstrong, N.; Buchman, V.L.; Ninkina, N. Selective Pattern of Motor System Damage in Gamma-Synuclein Transgenic Mice Mirrors the Respective Pathology in Amyotrophic Lateral Sclerosis. Neurobiol. Dis. 2012, 48, 124–131. [Google Scholar] [CrossRef]
- Peters, O.M.; Shelkovnikova, T.; Highley, J.R.; Cooper-Knock, J.; Hortobágyi, T.; Troakes, C.; Ninkina, N.; Buchman, V.L. Gamma-synuclein Pathology in Amyotrophic Lateral Sclerosis. Ann. Clin. Transl. Neurol. 2015, 2, 29–37. [Google Scholar] [CrossRef]
- Surgucheva, I.; Newell, K.L.; Burns, J.; Surguchov, A. New α- and γ-Synuclein Immunopathological Lesions in Human Brain. Acta Neuropathol. Commun. 2014, 2, 132. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, W.; Wu, Y.; Zhou, Y.; Xue, R.; Luo, C.; Wang, L.; Zhao, W.; Jiang, J.-D.; Liu, J. Loss of Epigenetic Control of Synuclein-γ Gene as a Molecular Indicator of Metastasis in a Wide Range of Human Cancers. Cancer Res. 2005, 65, 7635–7643. [Google Scholar] [CrossRef] [PubMed]
- Ninkina, N. Organization, Expression and Polymorphism of the Human Persyn Gene. Hum. Mol. Genet. 1998, 7, 1417–1424. [Google Scholar] [CrossRef] [PubMed]
- Surguchov, A. γ-Synuclein as a Cancer Biomarker: Viewpoint and New Approaches. Oncomedicine 2016, 1, 1–3. [Google Scholar] [CrossRef]
- Zanotti, L.C.; Malizia, F.; Cesatti Laluce, N.; Avila, A.; Mamberto, M.; Anselmino, L.E.; Menacho-Márquez, M. Synuclein Proteins in Cancer Development and Progression. Biomolecules 2023, 13, 980. [Google Scholar] [CrossRef]
- Biere, A.L.; Wood, S.J.; Wypych, J.; Steavenson, S.; Jiang, Y.; Anafi, D.; Jacobsen, F.W.; Jarosinski, M.A.; Wu, G.-M.; Louis, J.-C.; et al. Parkinson’s Disease-Associated α-Synuclein Is More Fibrillogenic than β- and γ-Synuclein and Cannot Cross-Seed Its Homologs. J. Biol. Chem. 2000, 275, 34574–34579. [Google Scholar] [CrossRef]
- Bonaccorsi Di Patti, M.C.; Meoni, M.; Toni, M. Comparative Analysis of Aggregation of β- and γ-Synucleins in Vertebrates. Biomolecules 2025, 15, 1231. [Google Scholar] [CrossRef]
- Giasson, B.I.; Murray, I.V.J.; Trojanowski, J.Q.; Lee, V.M.-Y. A Hydrophobic Stretch of 12 Amino Acid Residues in the Middle of α-Synuclein Is Essential for Filament Assembly. J. Biol. Chem. 2001, 276, 2380–2386. [Google Scholar] [CrossRef]
- Brown, J.W.P.; Buell, A.K.; Michaels, T.C.T.; Meisl, G.; Carozza, J.; Flagmeier, P.; Vendruscolo, M.; Knowles, T.P.J.; Dobson, C.M.; Galvagnion, C. β-Synuclein Suppresses Both the Initiation and Amplification Steps of α-Synuclein Aggregation via Competitive Binding to Surfaces. Sci. Rep. 2016, 6, 36010. [Google Scholar] [CrossRef]
- Li, X.; Yu, L.; Liu, X.; Shi, T.; Zhang, Y.; Xiao, Y.; Wang, C.; Song, L.; Li, N.; Liu, X.; et al. β-Synuclein Regulates the Phase Transitions and Amyloid Conversion of α-Synuclein. Nat. Commun. 2024, 15, 8748. [Google Scholar] [CrossRef]
- Park, J.-Y.; Lansbury, P.T. β-Synuclein Inhibits Formation of α-Synuclein Protofibrils: A Possible Therapeutic Strategy against Parkinson’s Disease. Biochemistry 2003, 42, 3696–3700. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Fornai, F.; Kwon, H.-B.; Yazdani, U.; Atasoy, D.; Liu, X.; Hammer, R.E.; Battaglia, G.; German, D.C.; Castillo, P.E.; et al. Double-Knockout Mice for α- and β-Synucleins: Effect on Synaptic Functions. Proc. Natl. Acad. Sci. 2004, 101, 14966–14971. [Google Scholar] [CrossRef] [PubMed]
- Greten-Harrison, B.; Polydoro, M.; Morimoto-Tomita, M.; Diao, L.; Williams, A.M.; Nie, E.H.; Makani, S.; Tian, N.; Castillo, P.E.; Buchman, V.L.; et al. Aβγ-Synuclein Triple Knockout Mice Reveal Age-Dependent Neuronal Dysfunction. Proc. Natl. Acad. Sci. 2010, 107, 19573–19578. [Google Scholar] [CrossRef] [PubMed]
- Kokhan, V.S.; Afanasyeva, M.A.; Van’kin, G.I. α-Synuclein Knockout Mice Have Cognitive Impairments. Behav. Brain Res. 2012, 231, 226–230. [Google Scholar] [CrossRef]
- Anwar, S.; Peters, O.; Millership, S.; Ninkina, N.; Doig, N.; Connor-Robson, N.; Threlfell, S.; Kooner, G.; Deacon, R.M.; Bannerman, D.M.; et al. Functional Alterations to the Nigrostriatal System in Mice Lacking All Three Members of the Synuclein Family. J. Neurosci. 2011, 31, 7264–7274. [Google Scholar] [CrossRef]
- Connor-Robson, N.; Peters, O.M.; Millership, S.; Ninkina, N.; Buchman, V.L. Combinational Losses of Synucleins Reveal Their Differential Requirements for Compensating Age-Dependent Alterations in Motor Behavior and Dopamine Metabolism. Neurobiol. Aging 2016, 46, 107–112. [Google Scholar] [CrossRef]
- Burré, J.; Sharma, M.; Tsetsenis, T.; Buchman, V.; Etherton, M.R.; Südhof, T.C. α-Synuclein Promotes SNARE-Complex Assembly in Vivo and in Vitro. Science 2010, 329, 1663–1667. [Google Scholar] [CrossRef]
- Peña-Oliver, Y.; Buchman, V.L.; Dalley, J.W.; Robbins, T.W.; Schumann, G.; Ripley, T.L.; King, S.L.; Stephens, D.N. Deletion of Alpha-synuclein Decreases Impulsivity in Mice. Genes Brain Behav. 2012, 11, 137–146. [Google Scholar] [CrossRef]
- Brown, J.L.; Hart, D.W.; Boyle, G.E.; Brown, T.G.; LaCroix, M.; Baraibar, A.M.; Pelzel, R.; Kim, M.; Sherman, M.A.; Boes, S.; et al. SNCA Genetic Lowering Reveals Differential Cognitive Function of Alpha-Synuclein Dependent on Sex. Acta Neuropathol. Commun. 2022, 10, 180. [Google Scholar] [CrossRef]
- Peña-Oliver, Y.; Buchman, V.L.; Stephens, D.N. Lack of Involvement of Alpha-Synuclein in Unconditioned Anxiety in Mice. Behav. Brain Res. 2010, 209, 234–240. [Google Scholar] [CrossRef]
- Kokhan, V.S.; Van’kin, G.I.; Bachurin, S.O.; Shamakina, I.Y. Differential Involvement of the Gamma-Synuclein in Cognitive Abilities on the Model of Knockout Mice. BMC Neurosci. 2013, 14, 53. [Google Scholar] [CrossRef] [PubMed]
- Oeckl, P.; Metzger, F.; Nagl, M.; Von Arnim, C.A.F.; Halbgebauer, S.; Steinacker, P.; Ludolph, A.C.; Otto, M. Alpha-, Beta-, and Gamma-Synuclein Quantification in Cerebrospinal Fluid by Multiple Reaction Monitoring Reveals Increased Concentrations in Alzheimer′s and Creutzfeldt-Jakob Disease but No Alteration in Synucleinopathies. Mol. Cell. Proteomics 2016, 15, 3126–3138. [Google Scholar] [CrossRef] [PubMed]
- Mukaetova-Ladinska, E.B.; Milne, J.; Andras, A.; Abdel-All, Z.; Cerejeira, J.; Greally, E.; Robson, J.; Jaros, E.; Perry, R.; McKeith, I.G.; et al. Alpha- and Gamma-Synuclein Proteins Are Present in Cerebrospinal Fluid and Are Increased in Aged Subjects with Neurodegenerative and Vascular Changes. Dement. Geriatr. Cogn. Disord. 2008, 26, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Ninkina, N.; Connor-Robson, N.; Ustyugov, A.A.; Tarasova, T.V.; Shelkovnikova, T.A.; Buchman, V.L. A Novel Resource for Studying Function and Dysfunction of α-Synuclein: Mouse Lines for Modulation of Endogenous Snca Gene Expression. Sci. Rep. 2015, 5, 16615. [Google Scholar] [CrossRef]
- Ninkina, N.; Papachroni, K.; Robertson, D.C.; Schmidt, O.; Delaney, L.; O’Neill, F.; Court, F.; Rosenthal, A.; Fleetwood-Walker, S.M.; Davies, A.M.; et al. Neurons Expressing the Highest Levels of γ-Synuclein Are Unaffected by Targeted Inactivation of the Gene. Mol. Cell. Biol. 2003, 23, 8233–8245. [Google Scholar] [CrossRef]
- Arudkumar, J.; Chinn Joshua Chey, Y.; Piltz, S.; Quinton Thomas, P.; Adikusuma, F. Forelimb Grip Strength Testing V1 2024.
- Chaprov, K.D.; Teterina, E.V.; Roman, A.Yu.; Ivanova, T.A.; Goloborshcheva, V.V.; Kucheryanu, V.G.; Morozov, S.G.; Lysikova, E.A.; Lytkina, O.A.; Koroleva, I.V.; et al. Comparative Analysis of MPTP Neurotoxicity in Mice with a Constitutive Knockout of the α-Synuclein Gene. Mol. Biol. 2021, 55, 133–142. [Google Scholar] [CrossRef]
- Jakkamsetti, V.; Scudder, W.; Kathote, G.; Ma, Q.; Angulo, G.; Dobariya, A.; Rosenberg, R.N.; Beutler, B.; Pascual, J.M. Quantification of Early Learning and Movement Sub-Structure Predictive of Motor Performance. Sci. Rep. 2021, 11, 14405. [Google Scholar] [CrossRef]
- Gellért, L.; Varga, D. Locomotion Activity Measurement in an Open Field for Mice. BIO-Protoc 2016, 6. [Google Scholar] [CrossRef]
- Sestakova, N.; Puzserova, A.; Kluknavsky, M.; Bernatova, I. Determination of Motor Activity and Anxiety-Related Behaviour in Rodents: Methodological Aspects and Role of Nitric Oxide. Interdiscip. Toxicol. 2013, 6, 126–135. [Google Scholar] [CrossRef]
- Tatem, K.S.; Quinn, J.L.; Phadke, A.; Yu, Q.; Gordish-Dressman, H.; Nagaraju, K. Behavioral and Locomotor Measurements Using an Open Field Activity Monitoring System for Skeletal Muscle Diseases. J. Vis. Exp. 2014, 51785. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.-H.; Jang, E.-H.; Choi, J.-H.; Lee, H.-R.; Bakes, J.; Kong, Y.-Y.; Kaang, B.-K. Basal Anxiety during an Open Field Test Is Correlated with Individual Differences in Contextually Conditioned Fear in Mice. Anim. Cells Syst. 2013, 17, 154–159. [Google Scholar] [CrossRef]
- Dellu, F.; Contarino, A.; Simon, H.; Koob, G.F.; Gold, L.H. Genetic Differences in Response to Novelty and Spatial Memory Using a Two-Trial Recognition Task in Mice. Neurobiol. Learn. Mem. 2000, 73, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Walf, A.A.; Frye, C.A. The Use of the Elevated plus Maze as an Assay of Anxiety-Related Behavior in Rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef]
- Kapadia, M.; Xu, J.; Sakic, B. The Water Maze Paradigm in Experimental Studies of Chronic Cognitive Disorders: Theory, Protocols, Analysis, and Inference. Neurosci. Biobehav. Rev. 2016, 68, 195–217. [Google Scholar] [CrossRef]
- Tarsa, L.; Goda, Y. Synaptophysin Regulates Activity-Dependent Synapse Formation in Cultured Hippocampal Neurons. Proc. Natl. Acad. Sci. 2002, 99, 1012–1016. [Google Scholar] [CrossRef]
- Wiedenmann, B.; Franke, W.W. Identification and Localization of Synaptophysin, an Integral Membrane Glycoprotein of Mr 38,000 Characteristic of Presynaptic Vesicles. Cell 1985, 41, 1017–1028. [Google Scholar] [CrossRef]
- Fujise, K.; Mishra, J.; Rosenfeld, M.S.; Rafiq, N.M. Synaptic Vesicle Characterization of iPSC-Derived Dopaminergic Neurons Provides Insight into Distinct Secretory Vesicle Pools. Npj Park. Dis. 2025, 11, 16. [Google Scholar] [CrossRef]
- Gitler, D.; Augustine, G.J. Synapsins and Regulation of the Reserve Pool. In Encyclopedia of Neuroscience; Elsevier, 2009; pp. 709–717. ISBN 978-0-08-045046-9. [Google Scholar]
- Stavsky, A.; Parra-Rivas, L.A.; Tal, S.; Riba, J.; Madhivanan, K.; Roy, S.; Gitler, D. Synapsin E-Domain Is Essential for α-Synuclein Function. eLife 2024, 12, RP89687. [Google Scholar] [CrossRef]
- Ferreira, A.; Chin, L.-S.; Li, L.; Lanier, L.M.; Kosik, K.S.; Greengard, P. Distinct Roles of Synapsin I and Synapsin II during Neuronal Development. Mol. Med. 1998, 4, 22–28. [Google Scholar] [CrossRef]
- Bauerfeind, R.; Takei, K.; De Camilli, P. Amphiphysin I Is Associated with Coated Endocytic Intermediates and Undergoes Stimulation-Dependent Dephosphorylation in Nerve Terminals. J. Biol. Chem. 1997, 272, 30984–30992. [Google Scholar] [CrossRef] [PubMed]
- David, C.; McPherson, P.S.; Mundigl, O.; De Camilli, P. A Role of Amphiphysin in Synaptic Vesicle Endocytosis Suggested by Its Binding to Dynamin in Nerve Terminals. Proc. Natl. Acad. Sci. 1996, 93, 331–335. [Google Scholar] [CrossRef]
- Kontaxi, C.; Kim, N.; Cousin, M.A. The Phospho-regulated Amphiphysin/Endophilin Interaction Is Required for Synaptic Vesicle Endocytosis. J. Neurochem. 2023, 166, 248–264. [Google Scholar] [CrossRef] [PubMed]
- Gao, V.; Briano, J.A.; Komer, L.E.; Burré, J. Functional and Pathological Effects of α-Synuclein on Synaptic SNARE Complexes. J. Mol. Biol. 2023, 435, 167714. [Google Scholar] [CrossRef] [PubMed]
- Abeliovich, A.; Schmitz, Y.; Fariñas, I.; Choi-Lundberg, D.; Ho, W.-H.; Castillo, P.E.; Shinsky, N.; Verdugo, J.M.G.; Armanini, M.; Ryan, A.; et al. Mice Lacking α-Synuclein Display Functional Deficits in the Nigrostriatal Dopamine System. Neuron 2000, 25, 239–252. [Google Scholar] [CrossRef]
- Somayaji, M.; Cataldi, S.; Choi, S.J.; Edwards, R.H.; Mosharov, E.V.; Sulzer, D. A Dual Role for α-Synuclein in Facilitation and Depression of Dopamine Release from Substantia Nigra Neurons in Vivo. Proc. Natl. Acad. Sci. 2020, 117, 32701–32710. [Google Scholar] [CrossRef]
- Ninkina, N.; Millership, S.J.; Peters, O.M.; Connor-Robson, N.; Chaprov, K.; Kopylov, A.T.; Montoya, A.; Kramer, H.; Withers, D.J.; Buchman, V.L. β-Synuclein Potentiates Synaptic Vesicle Dopamine Uptake and Rescues Dopaminergic Neurons from MPTP-Induced Death in the Absence of Other Synucleins. J. Biol. Chem. 2021, 297, 101375. [Google Scholar] [CrossRef]
- Xia, D.; Xiong, M.; Yang, Y.; Wang, X.; Chen, Q.; Li, S.; Meng, L.; Zhang, Z. Chronic Stress Induces Depression-like Behaviors and Parkinsonism via Upregulating α-Synuclein. Npj Park. Dis. 2025, 11, 139. [Google Scholar] [CrossRef]
- Kokhan, V.S.; Bolkunov, A.V.; Ustiugov, A.A.; Van’kin, G.I.; Shelkovnikova, T.A.; Redkozubova, O.M.; Strekalova, T.V.; Buchman, V.L.; Ninkina, N.N.; Bachurin, S.O. Targeted Inactivation of the Gene Encoding Gamma-Synuclein Affects Anxiety Levels and Investigative Activity in Mice. Neurosci. Behav. Physiol. 2012, 42, 575–581. [Google Scholar] [CrossRef]
- Al-Wandi, A.; Ninkina, N.; Millership, S.; Williamson, S.J.M.; Jones, P.A.; Buchman, V.L. Absence of α-Synuclein Affects Dopamine Metabolism and Synaptic Markers in the Striatum of Aging Mice. Neurobiol. Aging 2010, 31, 796–804. [Google Scholar] [CrossRef]
- Kim, B.; Yang, M.; Lee, J.; Kim, J.-S.; Hyun, S.-H.; Moon, C. Upregulation of γ-Synuclein in the Prefrontal Cortex and Hippocampus Following Dopamine Depletion: A Study Using the Striatal 6-Hydroxydopamine Hemiparkinsonian Rat Model. Neurosci. Lett. 2024, 839, 137936. [Google Scholar] [CrossRef]
- Boyer, F.; Dreyer, J. The Role of Gamma-synuclein in Cocaine-induced Behaviour in Rats. Eur. J. Neurosci. 2008, 27, 2938–2951. [Google Scholar] [CrossRef]
Figure 1.
Survival curves of synuclein knockout and WT mice. (A) γ-Synuclein KO mice (γ-KO). (B) Double α- and β-synuclein KO mice (αβ-KO). (C) Triple synuclein KO mice (αβγ-KO). Statistical analysis: Mantel-Cox (log-rank) test. * p < 0.05; *** p < 0.001.
Figure 1.
Survival curves of synuclein knockout and WT mice. (A) γ-Synuclein KO mice (γ-KO). (B) Double α- and β-synuclein KO mice (αβ-KO). (C) Triple synuclein KO mice (αβγ-KO). Statistical analysis: Mantel-Cox (log-rank) test. * p < 0.05; *** p < 0.001.
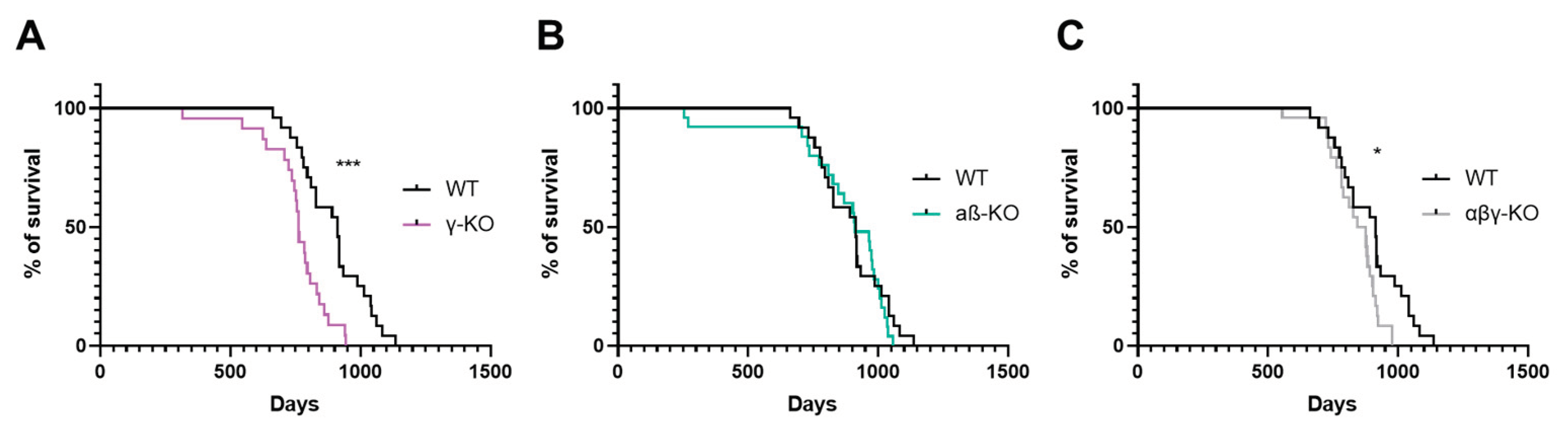
Figure 2.
Motor performance of 6-month-old synuclein KOs (γ-KO, αβ-KO and αβγ-KO) and WT mice. (A) Grip strength test. (B) Inverted grid test. (C) Accelerating rotarod. Data are presented as mean ± SEM. Statistical analysis: Kruskal-Wallis test with Dunn’s multiple comparisons test. *p < 0.05; **p < 0.01; ***p < 0.001. Sample sizes shown within the bars.
Figure 2.
Motor performance of 6-month-old synuclein KOs (γ-KO, αβ-KO and αβγ-KO) and WT mice. (A) Grip strength test. (B) Inverted grid test. (C) Accelerating rotarod. Data are presented as mean ± SEM. Statistical analysis: Kruskal-Wallis test with Dunn’s multiple comparisons test. *p < 0.05; **p < 0.01; ***p < 0.001. Sample sizes shown within the bars.
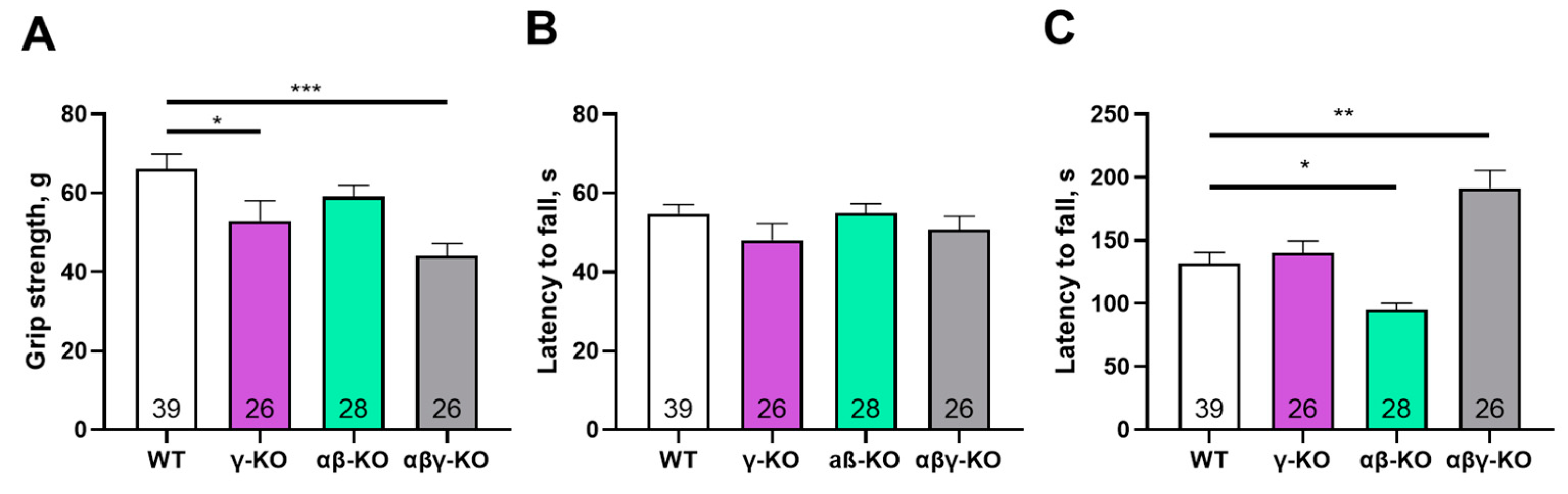
Figure 3.
Motor performance of 13-month-old synuclein knockout (γ-synuclein KO, αβ-KO and αβγ-KO) and WT mice. (A) Grip strength. (B) Inverted grid. (C) Accelerating rotarod. Data are presented as mean ± SEM. Statistical analysis: Kruskal-Wallis test with Dunn’s multiple comparisons test. ***p < 0.001 or the exact p values are shown. Sample sizes shown within the bars.
Figure 3.
Motor performance of 13-month-old synuclein knockout (γ-synuclein KO, αβ-KO and αβγ-KO) and WT mice. (A) Grip strength. (B) Inverted grid. (C) Accelerating rotarod. Data are presented as mean ± SEM. Statistical analysis: Kruskal-Wallis test with Dunn’s multiple comparisons test. ***p < 0.001 or the exact p values are shown. Sample sizes shown within the bars.
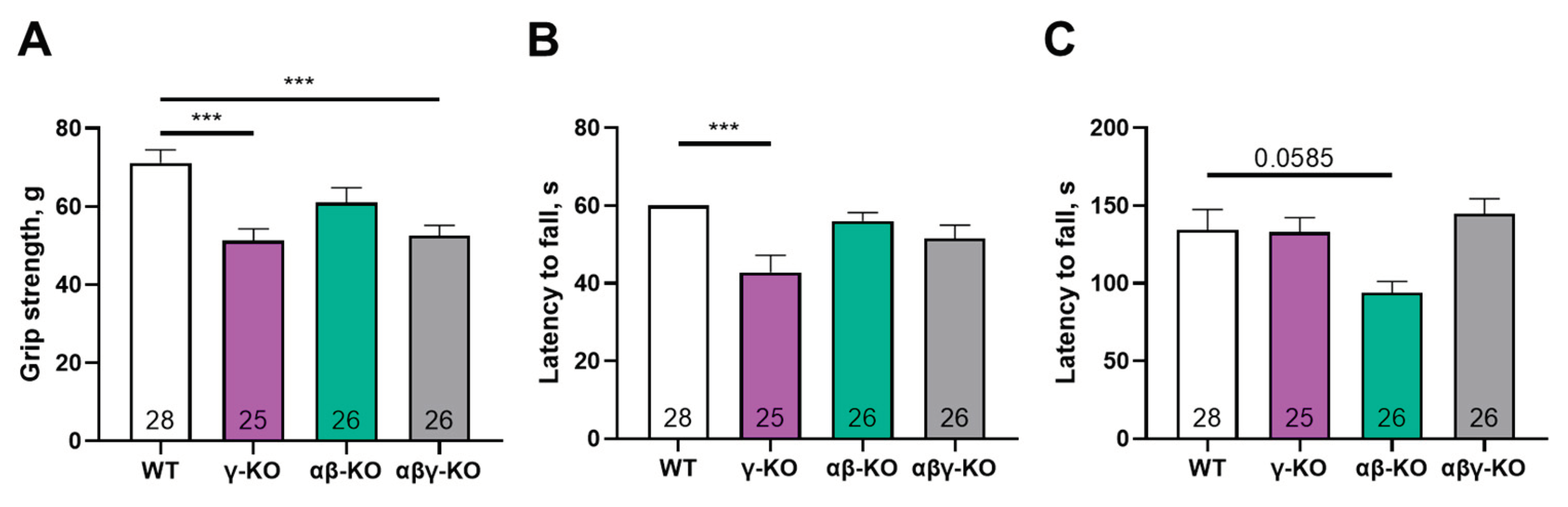
Figure 4.
Motor performance of 24-month-old synuclein KO (γ-KO, αβ-KO and αβγ-KO) and WT mice. (A) Grip strength. (B) Inverted grid. (C) Accelerating rotarod. Data are presented as mean ± SEM. Statistical analysis: Kruskal-Wallis test with Dunn’s multiple comparisons test (B); one-way ANOVA with Dunnett’s correction (A, C). *p < 0.05; **p < 0.01; ****p < 0.0001. Sample sizes shown within the bars.
Figure 4.
Motor performance of 24-month-old synuclein KO (γ-KO, αβ-KO and αβγ-KO) and WT mice. (A) Grip strength. (B) Inverted grid. (C) Accelerating rotarod. Data are presented as mean ± SEM. Statistical analysis: Kruskal-Wallis test with Dunn’s multiple comparisons test (B); one-way ANOVA with Dunnett’s correction (A, C). *p < 0.05; **p < 0.01; ****p < 0.0001. Sample sizes shown within the bars.
Figure 5.
Open field test for 13-month-old synuclein KO (γ-KO, αβ-KO and αβγ-KO) and WT mice. (A) Average distance traveled. (B) Time spent in the center. (C) Number of center entries. Data are presented as mean ± SEM. Statistical analysis: Kruskal-Wallis test with Dunn’s multiple comparisons test (B, C) and one-way ANOVA with Dunnett’s test (A). p<0.05; p<0.01; p<0.001; p<0.0001. Sample sizes shown within the bars.
Figure 5.
Open field test for 13-month-old synuclein KO (γ-KO, αβ-KO and αβγ-KO) and WT mice. (A) Average distance traveled. (B) Time spent in the center. (C) Number of center entries. Data are presented as mean ± SEM. Statistical analysis: Kruskal-Wallis test with Dunn’s multiple comparisons test (B, C) and one-way ANOVA with Dunnett’s test (A). p<0.05; p<0.01; p<0.001; p<0.0001. Sample sizes shown within the bars.
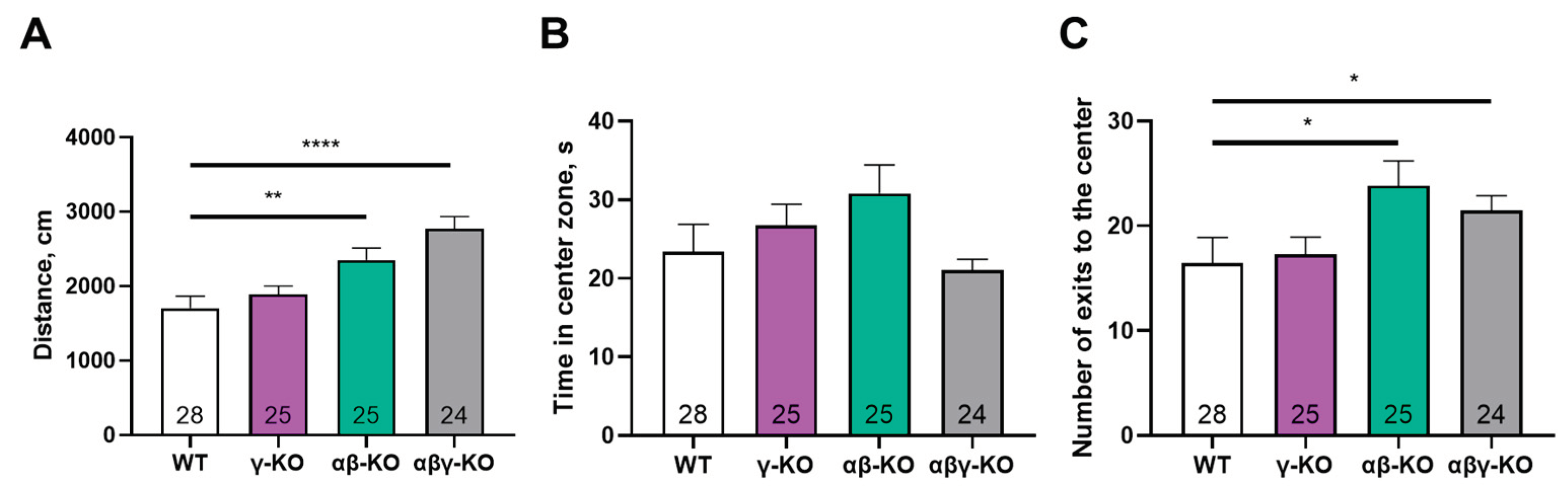
Figure 6.
Y-maze test for 13-month-old synuclein knockout (γ-synuclein KO, αβ-synuclein KO and αβγ-synuclein KO) and WT mice. (A) Total distance traveled. (B) Fraction of novel-arm entries. Data are presented as mean ± SEM. Statistical analysis: Kruskal-Wallis test with Dunn’s multiple comparisons test (B) and one-way ANOVA with Dunnett’s test (A). Symbols: * p<0.05; **** p<0.0001. Sample sizes are indicated by the numbers within the bars.
Figure 6.
Y-maze test for 13-month-old synuclein knockout (γ-synuclein KO, αβ-synuclein KO and αβγ-synuclein KO) and WT mice. (A) Total distance traveled. (B) Fraction of novel-arm entries. Data are presented as mean ± SEM. Statistical analysis: Kruskal-Wallis test with Dunn’s multiple comparisons test (B) and one-way ANOVA with Dunnett’s test (A). Symbols: * p<0.05; **** p<0.0001. Sample sizes are indicated by the numbers within the bars.
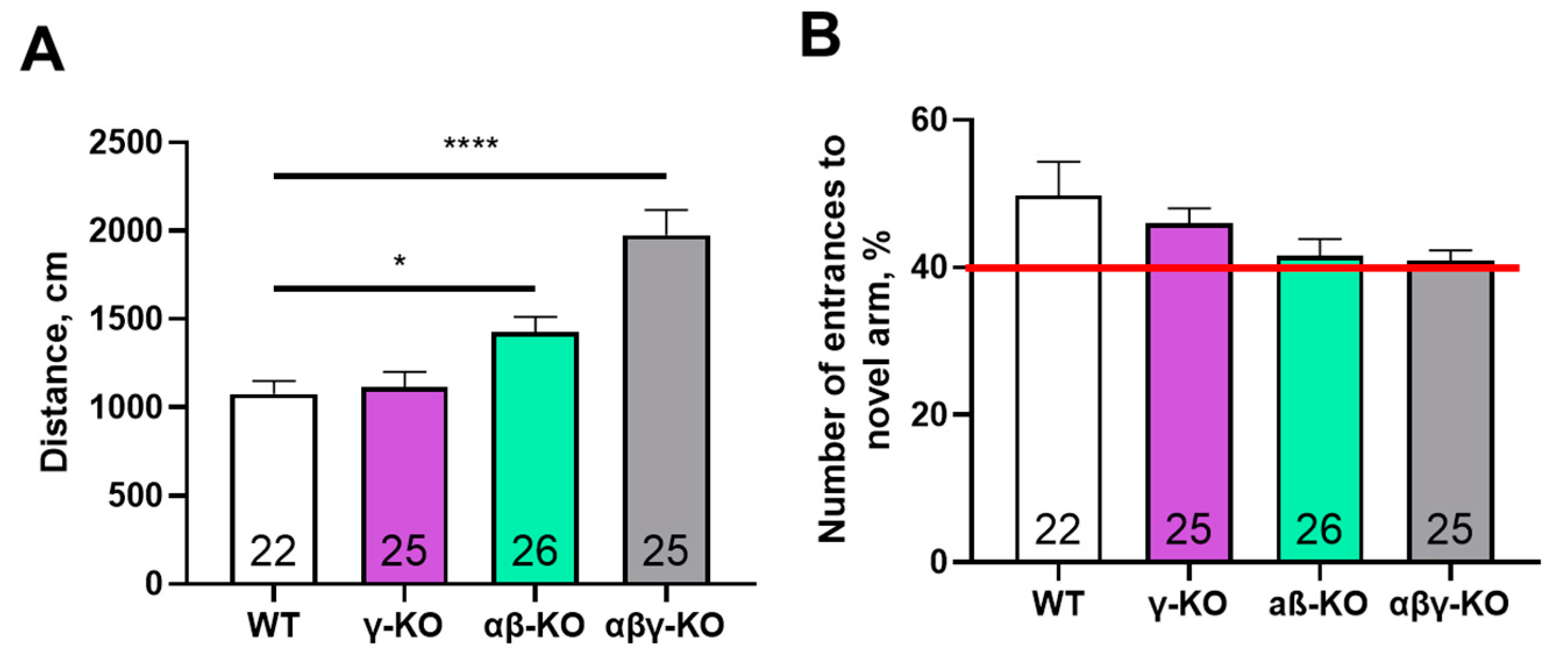
Figure 7.
Morris water maze test for 13-month-old synuclein KO (γ-KO, αβ-KO and αβγ-KO) and WT mice. (A) Latency to find the platform across 4 training days. (B) Number of platform-zone crossings during the day-5 probe trial. (C) Average swimming speed across 4 training days. (D) Latency to first platform-zone entry at the day-5 probe trial. Data are presented as mean ± SEM. Statistical analysis: Kruskal-Wallis test with Dunn’s multiple comparisons test (B, D) and two-way ANOVA with Dunnett’s test (A, C). Symbols: * p<0.05, comparison between WT and γ-KO at all time points; **** p<0.0001, comparison between WT and αβγ-KO at all time points. Sample sizes are indicated by the numbers within the bars.
Figure 7.
Morris water maze test for 13-month-old synuclein KO (γ-KO, αβ-KO and αβγ-KO) and WT mice. (A) Latency to find the platform across 4 training days. (B) Number of platform-zone crossings during the day-5 probe trial. (C) Average swimming speed across 4 training days. (D) Latency to first platform-zone entry at the day-5 probe trial. Data are presented as mean ± SEM. Statistical analysis: Kruskal-Wallis test with Dunn’s multiple comparisons test (B, D) and two-way ANOVA with Dunnett’s test (A, C). Symbols: * p<0.05, comparison between WT and γ-KO at all time points; **** p<0.0001, comparison between WT and αβγ-KO at all time points. Sample sizes are indicated by the numbers within the bars.
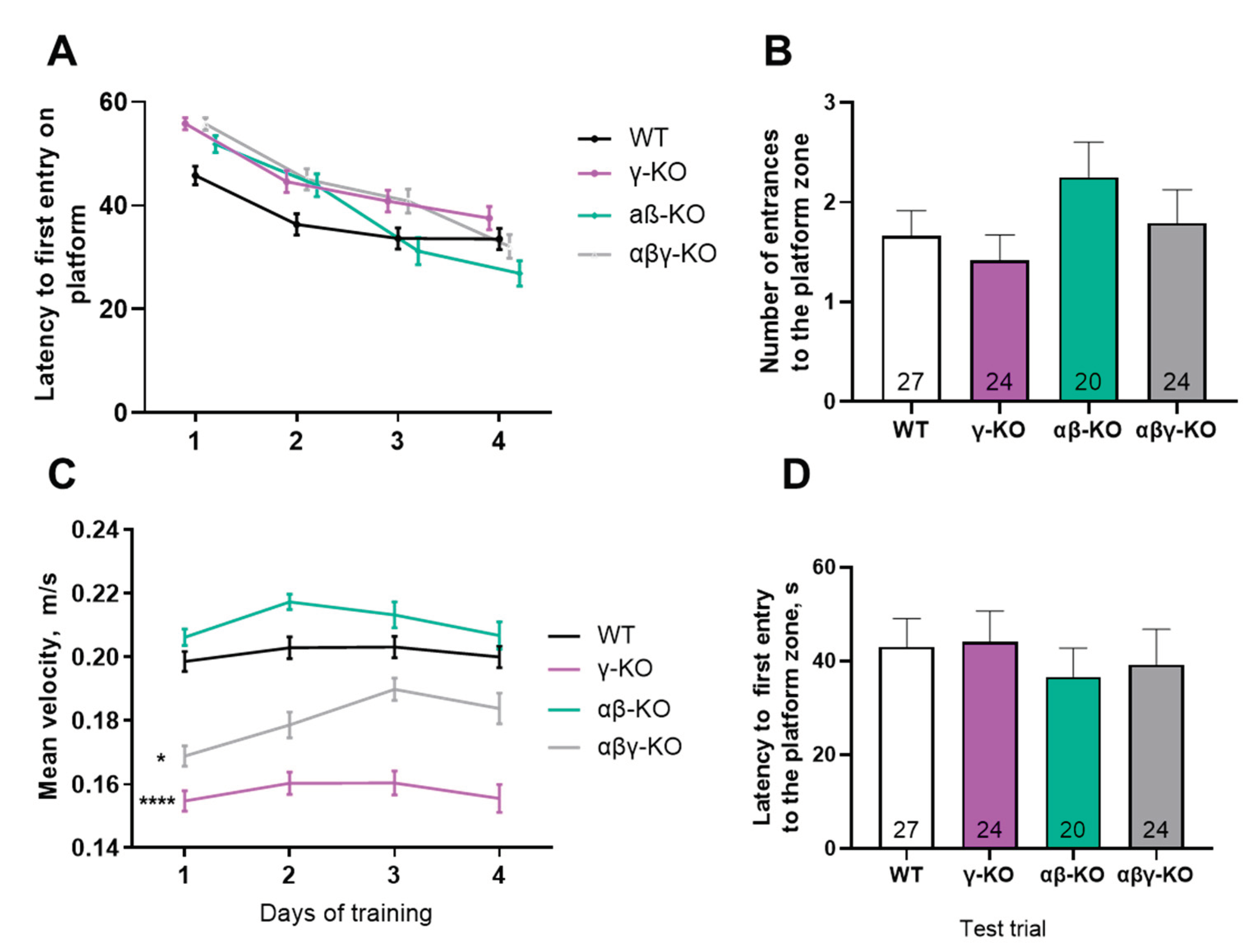
Figure 8.
Elevated plus maze test for 13-month-old synuclein KO (γ-synuclein KO, αβ-synuclein KO and αβγ-synuclein KO) and WT mice. (A) Number of entries into open arms. (B) Time spent in open arms. (C) Total distance traveled. Data are presented as mean ± SEM. Statistical analysis: Kruskal-Wallis test with Dunn’s multiple comparisons test (A, B) and one-way ANOVA with Dunnett’s test (C). Symbols: * p<0.05; *** p<0.001; **** p<0.0001. Sample sizes are indicated by the numbers within the bars.
Figure 8.
Elevated plus maze test for 13-month-old synuclein KO (γ-synuclein KO, αβ-synuclein KO and αβγ-synuclein KO) and WT mice. (A) Number of entries into open arms. (B) Time spent in open arms. (C) Total distance traveled. Data are presented as mean ± SEM. Statistical analysis: Kruskal-Wallis test with Dunn’s multiple comparisons test (A, B) and one-way ANOVA with Dunnett’s test (C). Symbols: * p<0.05; *** p<0.001; **** p<0.0001. Sample sizes are indicated by the numbers within the bars.
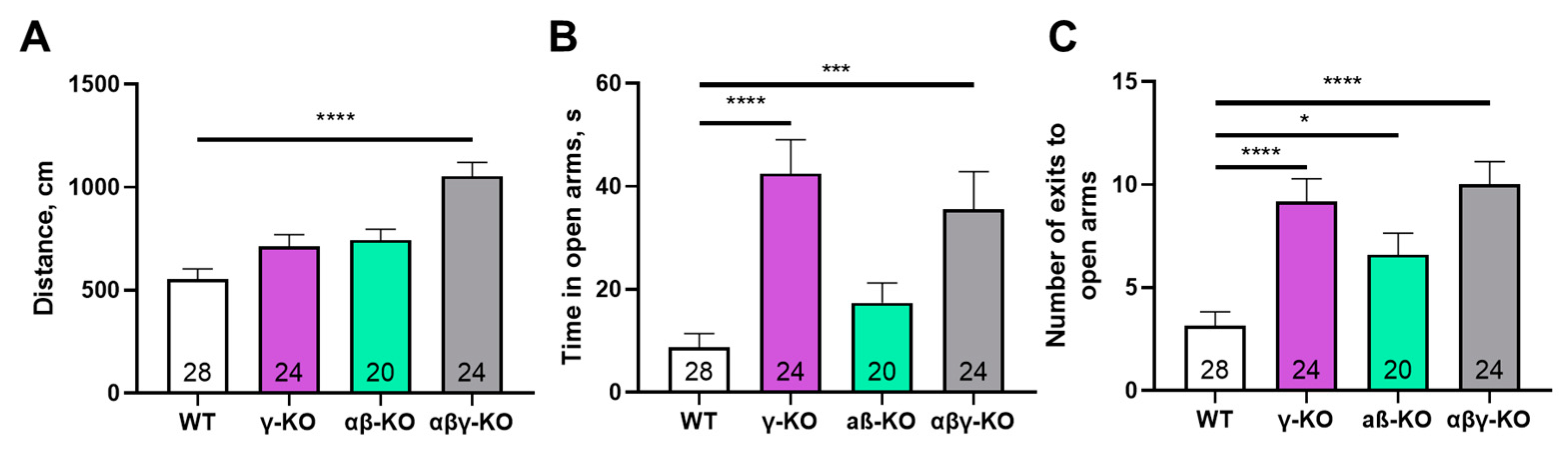
Figure 9.
Synaptic proteins in the brain of 13-month-old synuclein KO (γ-KO, αβ-KO and αβγ-KO) and WT mice, representative western-blots.
Figure 9.
Synaptic proteins in the brain of 13-month-old synuclein KO (γ-KO, αβ-KO and αβγ-KO) and WT mice, representative western-blots.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



