Submitted:
02 December 2025
Posted:
03 December 2025
Read the latest preprint version here
Abstract
Bioidentical hormone replacement therapy (BHRT) traditionally operates within a triad consisting of sex hormones, thyroid hormones, and adrenal glucocorticoids. Despite widespread adoption, a substantial proportion of patients experience persistent dysglycemia, adrenal instability, fluctuations in symptom control, and inconsistent responses to therapy even when laboratory values appear biochemically normalized. These clinical patterns suggest that an essential regulatory element is missing from the current BHRT conceptual model.This narrative review proposes the Insulin–Cortisol–Vitamin C (ICV) Axis as a previously unrecognized hormonal network central to metabolic and endocrine homeostasis. Insulin profoundly influences sex-hormone binding globulin (SHBG), estradiol and testosterone bioavailability, progesterone responsiveness, thyroid hormone conversion, mitochondrial ATP production, and cortisol reactivity—yet insulin is rarely evaluated in BHRT. Cortisol, in turn, directly modulates insulin sensitivity and metabolic function, while vitamin C is required for cortisol synthesis, adrenal recovery, endothelial nitric oxide signaling, mitochondrial redox regulation, and antioxidant defense. Together, disturbances in these three components can generate characteristic clinical presentations frequently encountered in BHRT practice.In parallel, emerging evidence—including metabolic insights from GLP-1 receptor agonist therapy—indicates that vitamin C status and oxidative stress modulation play broader roles in insulin sensitivity and hormonal signaling than previously recognized. Integrating these findings, the ICV Axis provides a systems-level framework capable of explaining BHRT treatment failures, variable patient responses, and persistent symptomatology despite standard hormone optimization.The purpose of this review is to synthesize biochemical, endocrine, and nutritional evidence supporting this new axis, and to outline a clinically actionable update to BHRT incorporating insulin dynamics and vitamin C sufficiency. Recognition of the ICV Axis represents a conceptual advancement that can improve therapeutic outcomes across metabolic, endocrine, and integrative medical practice.
Keywords:
bioidentical hormone replacement therapy (BHRT)
; cortisol
; endocrine regulation
; insulin
; insulin resistance
; metabolic homeostasis
; oxidative stress
; sex hormones
; thyroid hormones
; vitamin C
1. Introduction
Insulin resistance (IR) affects over 25% of the global adult population and underlies metabolic syndrome, type 2 diabetes, non-alcoholic fatty liver disease (NAFLD), cardiovascular disease (CVD), polycystic ovarian syndrome (PCOS), and accelerated aging. Traditionally, IR is the result of excessive carbohydrate intake, hypercaloric diets, inactivity, and obesity. Dietary interventions—especially low-carbohydrate diets—have proven effective for many individuals. However, clinical experience and heterogeneity of treatment outcomes indicate that carbohydrate intake is only one determinant of insulin homeostasis.
Multiple studies demonstrate that insulin sensitivity is influenced by oxidative stress, inflammation, glucocorticoid activity, magnesium insufficiency, micronutrient status, circadian rhythms, mitochondrial function, and chronic psychosocial stress. For example:
These findings indicate that insulin regulation is embedded in a broader metabolic-endocrine redox network, yet current frameworks often treat insulin in isolation.
1.1. A Missing Link: The Role of Vitamin C in Insulin Sensitivity
Vitamin C is an essential redox molecule and enzymatic cofactor involved in:
- synthesizing carnitine[24]
Vitamin C insufficiency contributes to and is a direct result of oxidative stress—one of the key drivers of insulin resistance[35]. A 2021 meta-analysis of 28 observational studies found significantly lower vitamin C levels in individuals with metabolic syndrome[10]. A randomized trial demonstrated improved glycemic parameters in type 2 diabetes with vitamin C supplementation[12].
1.2. Cortisol, Stress, and Insulin Coupling
Cortisol directly raises blood glucose via gluconeogenesis, reduces insulin-mediated glucose uptake, and promotes visceral fat deposition. Excess cortisol—whether due to chronic stress, sleep disruption, exogenous glucocorticoids, or HPA-axis dysregulation—is strongly associated with insulin resistance[34,37,38,39,40].
Vitamin C is highly concentrated in the adrenal cortex and is required for optimal cortisol synthesis and clearance—a relationship demonstrated in classic adrenal physiology research[34].
1.3. A Unified Model: The Insulin–Cortisol–Vitamin C (ICV) Axis
The cumulative literature suggests a tightly interdependent triad:
Yet no existing metabolic, endocrine, or nutritional framework integrates these three domains into a single explanatory model.
1.4. Purpose of This Review
This narrative review synthesizes evidence to support the conceptual validity of the ICV Axis, provides mechanistic insight, evaluates clinical implications, and identifies research gaps necessary to validate this triad as a clinically relevant regulatory system.
2. Physiology of Insulin Regulation
Insulin is an anabolic hormone essential for maintaining glucose homeostasis, promoting nutrient storage, supporting protein synthesis, and regulating lipid metabolism. While commonly framed as a hormone whose secretion depends primarily on dietary carbohydrate intake, insulin regulation is substantially more complex. It integrates signals from the gastrointestinal tract, autonomic nervous system, inflammatory mediators, oxidative stress, mitochondrial function, and the hypothalamic–pituitary–adrenal (HPA) axis.
Understanding insulin physiology requires examining both baseline secretion mechanisms and contextual modulators that influence insulin sensitivity and metabolic outcomes.
2.1. Glucose-Stimulated Insulin Secretion (GSIS)
Glucose entering pancreatic β-cells via GLUT1/GLUT3 is metabolized through glycolysis and oxidative phosphorylation, increasing intracellular ATP. The resulting rise in ATP/ADP ratio closes ATP-sensitive K+ channels, depolarizing the membrane and opening voltage-gated Ca²⁺ channels. Elevated intracellular Ca²⁺ triggers insulin granule exocytosis.
2.2. Non-Glucose Mediators of Insulin Secretion
- Incretins (GLP-1, GIP)
GLP-1 and GIP enhance insulin secretion in a glucose-dependent manner. GLP-1 also reduces glucagon and lower circulating glucose levels, slows gastric emptying, and decreases appetite[45,46,47].
- Amino acids and dietary proteins
- Free fatty acids (FFAs)
2.3. Insulin Sensitivity and Tissue Uptake
Once secreted, insulin facilitates glucose uptake in[52]:
- skeletal muscle
- adipose tissue
- liver (indirectly via suppression of hepatic glucose output)
The primary mechanism is insulin-stimulated GLUT4 translocation to the cell membrane in muscle and adipose tissue[52].
- physical activity
- mitochondrial function
- oxidative stress
- inflammation
- adipokines (adiponectin, leptin)
- magnesium sufficiency
- stress hormones (especially cortisol)
2.4. Insulin Resistance: A Multifactorial Process
Insulin resistance arises when insulin receptor signaling is impaired. Mechanisms include:
- Oxidative stress
Reactive oxygen species (ROS) interfere with insulin receptor substrate (IRS) signaling[2,61,62,63,64,65,66].
- Inflammation
TNF-α, IL-6, and other cytokines inhibit insulin receptor signaling ADDIN ZOTERO_ITEM CSL_CITATION {"citationID":"PeVL2IDa","properties":{"formattedCitation":"[142\\uc0\\u8211{}144]","plainCitation":"[142,143,144]","noteIndex":0},"citationItems":[{"id":15036,"uris":["http://zotero.org/users/3072536/items/FLSAEQP6"],"itemData":{"id":15036,"type":"article-journal","abstract":"Ascorbic acid (vitamin C) is a cofactor required in catecholamine synthesis for conversion of dopamine to norepinephrine by dopamine β-hydroxylase. Mutant mice lacking the plasma membrane ascorbic acid transporter (SVCT2) have severely reduced tissue levels of ascorbic acid and die after birth. We therefore investigated whether these mice might have impaired synthesis of catecholamines. Levels of catecholamines in brain were unaffected by SVCT2 deficiency. In heart, the only evidence for impaired dopamine β-hydroxylase activity was a twofold increase in tissue dopamine. An influence of the deficiency on tissue catecholamines was most prominent in the adrenals where norepinephrine was decreased by 50% and epinephrine, by 81%. On the ultrastructural level, adrenal chromaffin cells in SVCT2 null mice showed depletion of catecholamine storage vesicles, increased amounts of rough endoplasmic reticulum, signs of apoptosis, and increased glycogen storage. Decreased plasma levels of corticosterone indicated additional effects of the deficiency on adrenal cortical function. These data show that deranged catecholamine system function in SVCT2 null mice is largely restricted to the adrenal medulla and cannot account for the lethality in these animals. The data, however, establish a crucial role for ascorbic acid in adrenal chromaffin cell function.","container-title":"The FASEB Journal","DOI":"10.1096/fj.02-1167fje","ISSN":"1530-6860","issue":"13","language":"en","license":"© 2003 FASEB","note":"_eprint: https://faseb.onlinelibrary.wiley.com/doi/pdf/10.1096/fj.02-1167fje","page":"1-13","source":"Wiley Online Library","title":"Impaired adrenal catecholamine system function in mice with deficiency of the ascorbic acid transporter (SVCT2)","URL":"https://onlinelibrary.wiley.com/doi/abs/10.1096/fj.02-1167fje","volume":"17","author":[{"family":"Bornstein","given":"Stefan R."},{"family":"Yoshida-Hiroi","given":"Mayumi"},{"family":"Sotiriou","given":"Sotiria"},{"family":"Levine","given":"Mark"},{"family":"Hartwig","given":"Hans-Georg"},{"family":"Nussbaum","given":"Robert L."},{"family":"Eisenhofer","given":"Graeme"}],"accessed":{"date-parts":[["2025",11,27]]},"issued":{"date-parts":[["2003"]]}}},{"id":15146,"uris":["http://zotero.org/users/3072536/items/A4FD8G4Z"],"itemData":{"id":15146,"type":"article-journal","abstract":"Ever since Linus Pauling published his studies, the effects of vitamin C have been surrounded by contradictory results. This may be because its effects depend on a number of factors such as the redox state of the body, the dose used, and also on the tissue metabolism. This review deals with vitamin C pharmacokinetics and its participation in neurophysiological processes, as well as its role in the maintenance of redox balance. The distribution and the concentration of vitamin C in the organs depend on the ascorbate requirements of each and on the tissue distribution of sodium-dependent vitamin C transporter 1 and 2 (SVCT1 and SVCT2). This determines the specific distribution pattern of vitamin C in the body. Vitamin C is involved in the physiology of the nervous system, including the support and the structure of the neurons, the processes of differentiation, maturation and neuronal survival; the synthesis of catecholamine, and the modulation of neurotransmission. This antioxidant interacts with self-recycling mechanisms, including its participation in the endogenous antioxidant system. We conclude that the pharmacokinetic properties of ascorbate are related to the redox state and its functions and effects in tissues.","container-title":"Frontiers in Physiology","DOI":"10.3389/fphys.2015.00397","ISSN":"1664-042X","journalAbbreviation":"Front. Physiol.","language":"English","note":"publisher: Frontiers","source":"Frontiers","title":"Vitamin C in Health and Disease: Its Role in the Metabolism of Cells and Redox State in the Brain","title-short":"Vitamin C in Health and Disease","URL":"https://www.frontiersin.org/journals/physiology/articles/10.3389/fphys.2015.00397/full","volume":"6","author":[{"family":"Figueroa-Méndez","given":"Rodrigo"},{"family":"Rivas-Arancibia","given":"Selva"}],"accessed":{"date-parts":[["2025",11,27]]},"issued":{"date-parts":[["2015",12,23]]}}},{"id":15031,"uris":["http://zotero.org/users/3072536/items/N2J2DSQD"],"itemData":{"id":15031,"type":"article-journal","abstract":"Ascorbic acid is well known to acutely stimulate norepinephrine synthesis in neurosecretory cells, but it has also been shown over several days to increase tyrosine hydroxylase mRNA and norepinephrine synthesis in cultured neurons. Since tyrosine hydroxylase is the rate-limiting step in catecholamine synthesis, an effect of ascorbate to increase tyrosine hydroxylase protein could contribute to its ability to increase or sustain catecholamine synthesis. Therefore, we evaluated whether tyrosine hydroxylase protein expression and function is increased in SH-SY5Y neuroblastoma cells by physiologically relevant intracellular ascorbate concentrations. SH-SY5Y neuroblastoma cells did not contain ascorbate and had only very low levels of norepinephrine in culture with L-tyrosine, the substrate for tyrosine hydroxylase. However, treatment of cells with ascorbate for 6 h or more markedly increased norepinephrine synthesis, such that intracellular ascorbate and norepinephrine increased in parallel with half maximal intracellular concentrations of about 1 mM ascorbate and 150 μM norepinephrine. This increase was enhanced by supplementing tetrahydrobiopterin, but was not mimicked by several antioxidants or by catalase or superoxide dismutase. Tyrosine hydroxylase protein expression increased at intracellular ascorbate concentrations above 1.5 mM. This contributed to norepinephrine generation, which was decreased 50-60% by inhibition of protein synthesis with cycloheximide at high intracellular ascorbate. These results suggest that expected physiologic neuronal ascorbate concentrations enhance norepinephrine synthesis both by maintaining tetrahydrobiopterin and increasing tyrosine hydroxylase expression.","container-title":"Biochemical and Biophysical Research Communications","DOI":"10.1016/j.bbrc.2012.08.054","ISSN":"1090-2104","issue":"1","journalAbbreviation":"Biochem Biophys Res Commun","language":"eng","note":"PMID: 22925890\nPMCID: PMC3449284","page":"148-152","source":"PubMed","title":"Mechanisms of ascorbic acid stimulation of norepinephrine synthesis in neuronal cells","volume":"426","author":[{"family":"May","given":"James M."},{"family":"Qu","given":"Zhi-chao"},{"family":"Meredith","given":"M. Elizabeth"}],"issued":{"date-parts":[["2012",9,14]]}}}],"schema":"https://github.com/citation-style-language/schema/raw/master/csl-citation.json"} [142,143,144][67,68,69,70,71,72].
- Mitochondrial dysfunction
- Lipotoxicity / Ectopic fat
Accumulation of diacylglycerol and ceramides in liver or muscle blocks insulin signaling [71,72,73].
- Circadian disruption
- Chronic stress & cortisol elevation
2.5. Insulin Variability and Fluctuations Matter
Emerging evidence suggests that insulin variability, not just absolute levels, contributes to metabolic dysfunction.
2.6. Vitamin C as a Modulator of Insulin Physiology
Although traditionally excluded from mainstream models of insulin regulation, vitamin C participates in multiple biochemical pathways that directly influence insulin sensitivity and metabolic homeostasis. A growing body of evidence demonstrates that vitamin C status affects key regulatory systems involved in glucose uptake, mitochondrial function, endothelial signaling, and inflammatory control. These mechanisms provide a scientifically grounded basis for understanding vitamin C as a physiologically important modulator of insulin action.
Key mechanisms include:
- Support of GLUT4 activation and glucose uptake, partly through antioxidant protection of skeletal muscle and pancreatic β-cells, and through improved mitochondrial redox status[9].
- Augmentation of endothelial nitric oxide activity, mediated by preservation of tetrahydrobiopterin and reduction of NO oxidative degradation, which improves insulin-mediated vasodilation and tissue glucose delivery[30].
Human studies support the physiological relevance of these mechanisms:
- Supplementation lowers fasting glucose, triglycerides, LDL cholesterol, HbA1c, and serum insulin in type 2 diabetes, suggesting improved glycemic control and insulin sensitivity[96].
Collectively, these findings establish vitamin C as a significant modulator of insulin physiology, acting through convergent pathways involving oxidative stress reduction, GLUT4 activation, mitochondrial support, nitric oxide–mediated vascular function, and inflammatory control. These mechanisms form the foundation for Section 3, which examines direct evidence linking vitamin C status to insulin sensitivity and metabolic disease outcomes.
3. Evidence Linking Vitamin C Status and Insulin Sensitivity
A growing body of human, clinical, and mechanistic research demonstrates a significant association between vitamin C status and insulin sensitivity. While vitamin C is traditionally framed as an antioxidant, the evidence outlined below shows that ascorbate participates directly in biological processes central to insulin signaling, glucose uptake, endothelial function, and inflammatory regulation. The totality of this literature supports the hypothesis that vitamin C is an important modulator of insulin homeostasis.
3.1. Observational Evidence: Vitamin C Levels Inversely Correlate with Insulin Resistance and Metabolic Syndrome
Multiple population-based studies report that low circulating vitamin C levels are strongly associated with features of metabolic syndrome—including elevated fasting glucose, central adiposity, hypertension, dyslipidemia, and insulin resistance. A recent NHANES-based analysis confirmed a robust inverse relationship between plasma ascorbate and cardiometabolic risk markers, with individuals in the lowest quartile of vitamin C demonstrating the highest HOMA-IR and metabolic syndrome prevalence [92]. These findings are consistent with prior epidemiologic studies showing that inadequate dietary vitamin C intake predicts higher fasting glucose and impaired insulin sensitivity [93].
3.2. Interventional Trials: Vitamin C Supplementation Improves Glycemic Control and Insulin Biomarkers
Controlled supplementation trials further support a causal role for vitamin C in improving insulin dynamics. Daily oral vitamin C (typically 500–1,000 mg twice per day) has been shown to significantly reduce fasting glucose, HbA1c, triglycerides, LDL cholesterol, and serum insulin levels in individuals with type 2 diabetes, indicating improved insulin sensitivity and glycemic handling [94]. Meta-analytic data reinforce these findings: pooled results from randomized trials reveal that vitamin C supplementation lowers oxidative stress markers, improves glycemic control, and reduces HOMA-IR in patients with metabolic syndrome and type 2 diabetes [95].
Although study heterogeneity exists—particularly regarding dose, duration, and population characteristics—the consistency of these results across multiple cohorts provides evidence that vitamin C favorably influences insulin physiology under metabolic stress conditions.
3.3. Mechanistic Studies Demonstrating Improvements in Insulin Signaling
Vitamin C exerts several mechanistic effects directly relevant to insulin action:
- Reduction of Oxidative Stress: Vitamin C decreases reactive oxygen species that otherwise impair insulin receptor substrate (IRS) phosphorylation and downstream signaling, thereby maintaining insulin responsiveness[93].
These mechanistic pathways align with clinical observations showing improved insulin sensitivity following vitamin C replenishment.
3.4. Endothelial Function: Vitamin C Enhances NO Bioavailability and Insulin-Mediated Vasodilation
Insulin’s ability to stimulate glucose uptake in skeletal muscle partly depends on endothelial nitric oxide (NO) availability, which regulates the vasodilation required for glucose and insulin delivery to peripheral tissues. Vitamin C sustains endothelial NO synthase (eNOS) activity by maintaining tetrahydrobiopterin (BH4) and reducing oxidative inactivation of NO [99]. Interventional trials in humans show that vitamin C supplementation improves endothelial-dependent vasodilation, a process strongly linked to improved insulin-mediated glucose disposal and overall metabolic health[96].
3.5. Anti-Inflammatory Effects: Vitamin C Reduces Cytokines That Impair Insulin Signaling
Chronic low-grade inflammation is a major driver of insulin resistance. Vitamin C supplementation has been shown to reduce proinflammatory cytokines—such as TNF-α, IL-1β, and CRP—that impair insulin receptor kinase activity and GLUT4 signaling pathways[10]. By lowering inflammatory burden, vitamin C indirectly improves insulin sensitivity and helps restore metabolic homeostasis, particularly in individuals with metabolic syndrome, obesity, or chronic stress.
3.6. Summary of Evidence
Consistent observational, interventional, and mechanistic findings support the conclusion that vitamin C is a significant modulator of insulin physiology. The effects are mediated through:
- reduction of oxidative stress
- enhancement of GLUT4 activation and glucose uptake
- mitochondrial support and metabolic flexibility
- improvement of endothelial NO-dependent vasodilation
- suppression of inflammatory pathways that impair insulin receptor activity
Taken together, these findings establish vitamin C as a biologically plausible and clinically relevant component of insulin regulation and provide the scientific rationale for examining vitamin C as a central element within the proposed Insulin–Cortisol–Vitamin C (ICV) Axis.
4. The Insulin–Cortisol–Vitamin C (ICV) Axis: A Unified Regulatory Framework
Insulin homeostasis is commonly assessed in isolation—primarily as a function of dietary carbohydrate intake, pancreatic β-cell output, and peripheral insulin receptor sensitivity. However, evidence reviewed in Section 2 and Section 3 indicates that insulin dynamics cannot be fully understood without integrating (1) cortisol regulation and (2) vitamin C status. Both are key determinants of oxidative stress, inflammation, glucose disposal, mitochondrial function, and vascular signaling—each of which directly modulates insulin action.
The ICV Axis proposes that insulin, cortisol, and vitamin C form a tightly interdependent regulatory triad. Disturbances in any one component propagate through the other two, creating self-reinforcing cycles of metabolic dysfunction. This section outlines the physiological basis for this coupling, describes known interactions between these pathways, and highlights their combined relevance to metabolic and endocrine homeostasis.
4.1. Bidirectional Coupling Between Insulin and Cortisol
Cortisol is a potent counter-regulatory hormone whose primary role is to increase glucose availability during stress and upregulate vitamin C uptake into the cells[15,100,101]. Cortisol directly antagonizes insulin by:
Elevated or dysregulated cortisol patterns—whether due to chronic psychosocial stress, sleep disruption, systemic inflammation, or HPA-axis dysfunction—consistently correlate with impaired insulin sensitivity and higher fasting insulin levels[37,103,104,116,119,120]. Numerous observational and clinical studies demonstrate:
Conversely, hyperinsulinemia may affect the HPA axis through central feedback mechanisms, promoting additional cortisol release. Thus, cortisol and insulin form a bidirectional loop, where abnormalities in one reinforce abnormalities in the other.
4.2. The Central Role of Vitamin C in Adrenal Physiology and Cortisol Regulation
Vitamin C concentrates in the adrenal cortex at the highest levels found in any human organ, reflecting its essential role in adrenal steroidogenesis. Ascorbate is required for:
4.3. Vitamin C as a Regulator of Insulin Sensitivity
Section 3 establishes that vitamin C influences multiple pathways essential for insulin action, including:
- oxidative stress reduction
- preservation of insulin receptor function
- enhancement of GLUT4-mediated glucose uptake
- mitochondrial redox support
- maintenance of endothelial nitric oxide bioavailability
- suppression of pro-inflammatory cytokines
Together, these effects position vitamin C as a direct modulator of insulin sensitivity, and not merely a general antioxidant. Vitamin C deficiency or suboptimal status—common in individuals with chronic illness, metabolic syndrome, or elevated oxidative stress—can therefore contribute directly to impaired insulin signaling[10,35,36,140,141,142].
4.4. Integration of Insulin, Cortisol, and Vitamin C: A Systems Physiology Model
The combined evidence supports a unified model wherein insulin, cortisol, and vitamin C act as a coupled metabolic–endocrine axis:
4.4.1. Core Relationships
- Cortisol → Insulin: cortisol elevation increases glucose output and insulin secretion.
- Insulin → Cortisol: hyperinsulinemia amplifies central HPA-axis activation and promotes visceral adiposity, further elevating cortisol.
- Cortisol → Vitamin C: cortisol directly increases intracellular vitamin C levels by stimulating the synthesis of vitamin C transporters and resulting in an anti-inflammatory impact in cells and tissues.
- Vitamin C → Cortisol: vitamin C regulates cortisol synthesis and shutdown; deficiency prolongs cortisol elevation.
- Vitamin C → Insulin: vitamin C improves insulin sensitivity via oxidative stress reduction, GLUT4 facilitation, mitochondrial support, and endothelial function.
- Insulin → Vitamin C: hyperglycemia and oxidative load increase vitamin C turnover, depleting tissue reserves.
Importantly, recent commentary on modern metabolic therapies aligns with the ICV-axis concept. As illustrated in GLP-1 Receptor Agonists and Vitamin C: A Powerful Anti-Aging Combination, chronic use of GLP-1RAs may exert many of their beneficial metabolic and anti-aging effects via enhanced intracellular vitamin C uptake, redox normalization, improved mitochondrial function, and optimized insulin sensitivity — effectively functioning as proof-of-concept that vitamin C status can modulate the same pathways described in the ICV model[143].
4.4.2. The Dysregulation Cycle
A disturbance in any one component—cortisol excess, vitamin C deficiency, or insulin resistance—propagates through the other two, forming a self-amplifying cycle:
- 1.
- Stress → cortisol ↑
- 2.
- Cortisol ↑ → glucose ↑ → insulin ↑
- 3.
- Insulin ↑ → oxidative stress ↑ → vitamin C depletion
- 4.
- Vitamin C ↓ → impaired cortisol regulation + reduced insulin sensitivity
- 5.
- Cycle repeats, leading to entrenched metabolic dysfunction.
This cycle helps explain:
- persistently high insulin despite low-carbohydrate diets
- stress-related metabolic deterioration
- mitochondrial dysfunction
- endothelial dysfunction
- BHRT instability (via SHBG alterations, thyroid conversion issues, and altered adrenal output)
- heterogeneity in response to GLP-1 receptor agonists
- metabolic syndrome and fatigue syndromes refractory to standard treatment
4.5. Clinical Relevance of the ICV Axis
The ICV Axis provides a mechanistic explanation for numerous clinical observations:
- Low-carb nonresponders
Individuals who do not improve insulin sensitivity despite dietary carbohydrate restriction often show:
- elevated cortisol
- low vitamin C status
- increased oxidative stress
- inflammation or sleep disruptionThe ICV model explains persistent insulin resistance in this subgroup.
- BHRT instability
Insulin sensitivity influences:
- SHBG levels
- free estrogen/testosterone
- progesterone sensitivity
- thyroid conversion
Cortisol and vitamin C both modulate these pathways, suggesting an integrated framework for hormone therapy optimization.
- Metabolic syndrome & cardiometabolic risk
ICV dysregulation aligns with hallmark features of metabolic syndrome, including:
- hyperinsulinemia
- central adiposity
- endothelial dysfunction
- hypertension
- dyslipidemia
- GLP-1 agonist variability
Variations in vitamin C status and HPA-axis function may partially explain heterogeneous responses to GLP-1 therapies.
4.6. Implications for Research and Clinical Practice
The ICV Axis suggests new directions for:
- biomarker development (vitamin C status, cortisol rhythms, oxidative stress indices)
- clinical trials (vitamin C repletion + stress modulation + metabolic therapy)
- personalized medicine approaches integrating diet, micronutrition, and endocrine regulation
Despite strong mechanistic and associative evidence, causal validation of the ICV Axis requires prospective interventional studies.
4.7. Summary of Section 4
The Insulin–Cortisol–Vitamin C Axis offers a new conceptual framework for understanding metabolic and hormonal homeostasis. Evidence from endocrinology, nutrition science, stress physiology, and redox biology supports a tightly interdependent triad, where disturbances in one element propagate through the others and contribute to chronic metabolic disease. Integrating this model into clinical practice may improve treatment outcomes for insulin resistance, metabolic syndrome, BHRT, and cardiometabolic disease.
5. Integrating the ICV Axis Into Existing BHRT Frameworks
Bioidentical hormone replacement therapy (BHRT) is widely used to address age-related declines in sex hormones, mitigate menopausal and andropausal symptoms, support metabolic health, and improve quality of life. Traditional BHRT models focus on the gonadal axis (estrogen, progesterone, testosterone), thyroid optimization, and—less consistently—the adrenal pathway. However, insulin is not typically conceptualized as a formal hormonal axis within BHRT, despite its central regulatory role in metabolism, inflammation, vascular function, body composition, and endocrine crosstalk.
The Insulin–Cortisol–Vitamin C (ICV) Axis offers a new integrative framework that bridges metabolic endocrinology with micronutrient biochemistry and stress physiology, providing a more comprehensive foundation for BHRT clinical decision-making. Incorporating the ICV Axis into BHRT represents an advancement in root-cause physiology and may improve therapeutic consistency, resilience, and long-term outcomes.
5.1. Why Insulin Should Be Considered a Hormonal Axis in BHRT
Insulin is an anabolic hormone with broad effects on receptor expression, steroidogenesis, hepatic sex-hormone binding globulin (SHBG) synthesis, thyroid conversion, adrenal signaling, and mitochondrial function[144,145,146]. Dysregulated insulin—whether chronically elevated, suppressed, or highly variable—can significantly alter the clinical response to BHRT[147,148].
Key reasons insulin belongs in BHRT frameworks:
Yet BHRT practice rarely measures fasting insulin, postprandial insulin, or insulin variability, and almost never addresses insulin as a therapeutic target.
5.2. How Cortisol Links Insulin and Sex Hormone Physiology
The adrenal cortex produces cortisol in response to physiologic and psychologic stress. Cortisol directly elevates glucose and induces insulin secretion. Conversely, insulin fluctuations affect circadian cortisol patterns and HPA-axis reactivity[160,161,162].
In BHRT:
This triad becomes especially relevant in peri- and post-menopausal physiology, where adrenal compensation is already taxed.
5.3. A Critical Missing Component: Vitamin C in BHRT Physiology
Vitamin C has not historically been included in BHRT, yet it is essential for:
Vitamin C insufficiency amplifies BHRT side effects such as emotional lability, sleep disruption, hot flashes, fatigue, poor stress tolerance, and suboptimal metabolic response[175,176].
By introducing vitamin C into the BHRT conceptual model, the ICV Axis provides the first micronutrient-integrated BHRT physiology framework.
5.4. Why Existing BHRT Models Fail Without Insulin–Cortisol–Vitamin C Integration
- inconsistent symptom improvement
- persistent fatigue
- continued weight gain
- elevated inflammatory markers
- fluctuating hot flashes or night sweats
- unstable mood
- reduced libido
- plateaued metabolic progress
Many of these clinical patterns correlate more closely with insulin, cortisol, redox status, and vitamin C levels than with sex hormone dosing.
Without addressing the ICV Axis, clinicians may mistakenly escalate estrogen, progesterone, or testosterone dosing when the root cause is metabolic-endocrine imbalance—not sex hormone deficiency.
5.5. Practical Integration: Updating BHRT to the ICV Model
Incorporating the ICV Axis into BHRT practice includes (Figure 1):
Figure 1.
The ICV Axis as a regulatory hub in BHRT.
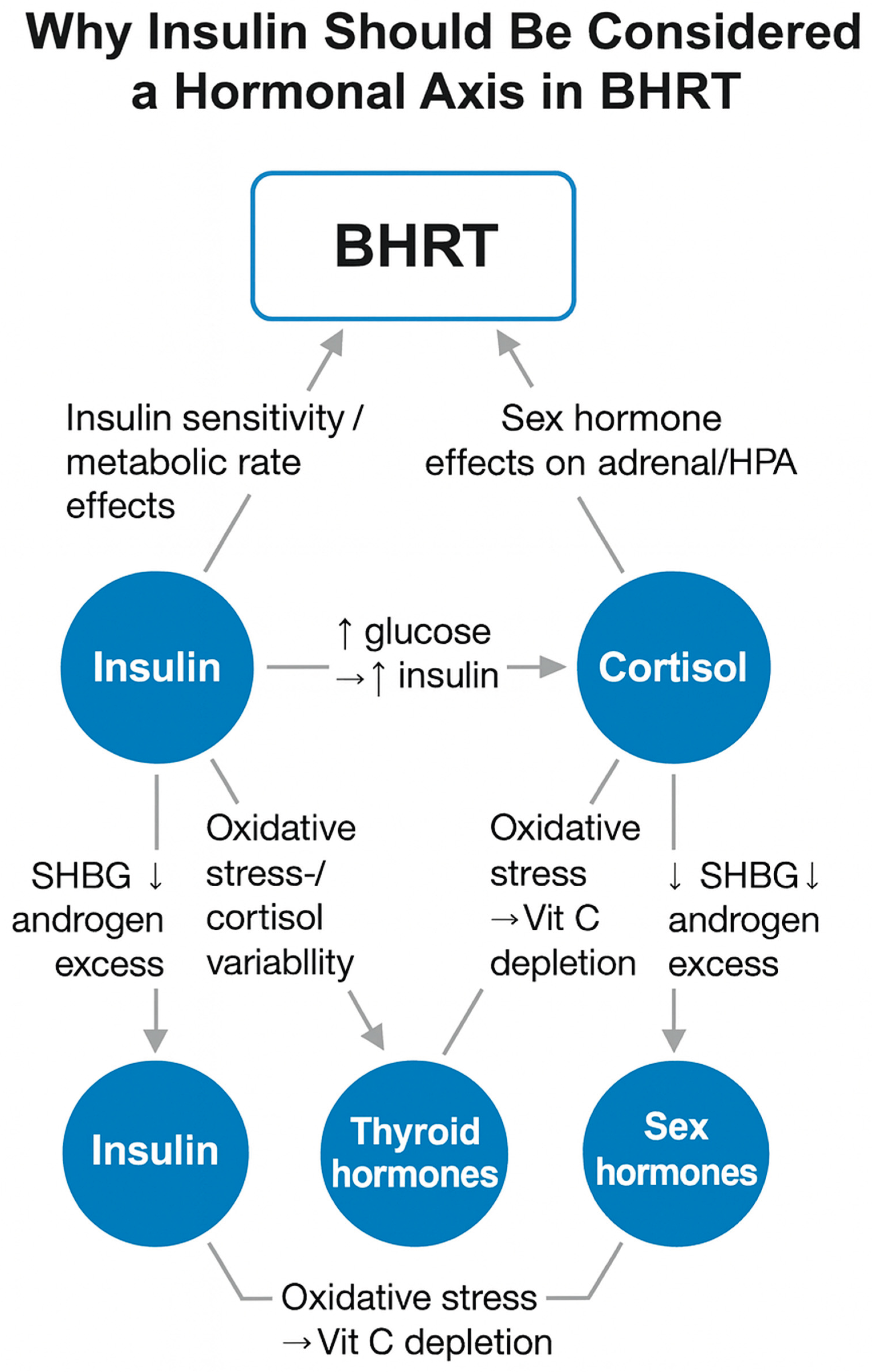
This integrated approach is more physiologic, more personalized, and more aligned with systems biology.
- Legend:
- Blue circles = key hormonal/metabolic nodes
- Blue rounded rectangle = BHRT regulatory inputs
- Gray arrows = primary directional influences
- Dashed arrows (if present) = feedback effects
- Labels = dominant mechanistic pathways (e.g., ↑ glucose → ↑ insulin; oxidative stress → vitamin C depletion; SHBG ↓ → androgen excess)
The ICV Axis (insulin–cortisol–vitamin C) interacts directly with thyroid and sex hormone physiology, influencing SHBG, androgen balance, adrenal load, oxidative stress, thyroid conversion, and metabolic rate. These relationships help explain variable or unstable BHRT outcomes when insulin and cortisol dynamics or vitamin C status are not addressed.
5.6. Summary: Why This Is a Landmark Advancement in BHRT
Introducing insulin as a hormonal axis within BHRT—and connecting it mechanistically to cortisol and vitamin C—represents a paradigm shift. BHRT has long operated within a tripartite model (sex hormones, thyroid, adrenal), but the ICV Axis reveals an overlooked regulatory dimension essential for metabolic stability and hormone responsiveness.
This framework positions BHRT within a broader systems-medicine context and lays the groundwork for:
- more effective interventions
- fewer treatment failures
- better metabolic outcomes
- improved patient resilience
- a unified model connecting nutrition, hormones, stress physiology, and redox biology
The ICV Axis therefore provides a novel, scientifically grounded, and clinically actionable expansion to the existing BHRT paradigm.
6. Implications for Research and Clinical Practice
The proposed Insulin–Cortisol–Vitamin C (ICV) Axis expands existing models of metabolic regulation by integrating endocrine physiology, redox biology, and micronutrient status into a unified framework. While traditional approaches to insulin resistance and metabolic syndrome emphasize carbohydrate reduction, pharmacologic therapies, or isolated hormonal modulation, the ICV model underscores the importance of systems-level interactions often overlooked in clinical practice.
6.1. Implications for Endocrinology and BHRT Practice
Current BHRT paradigms focus primarily on sex hormones (estradiol, progesterone, testosterone), thyroid hormones, and occasionally DHEA. Insulin is rarely conceptualized as a hormone requiring balanced homeostasis within the same framework, despite its broad downstream effects on SHBG, free hormone fractions, adipokines, and metabolic signaling. Incorporating insulin dynamics and micronutrient status—particularly vitamin C sufficiency—may improve BHRT outcomes, reduce dose variability, and help explain clinical scenarios where hormone replacement appears biochemically adequate but symptomatically insufficient.
6.2. Implications for Metabolic and Nutritional Medicine
The ICV Axis suggests that insulin resistance cannot be fully corrected by diet alone. Individuals adhering to low-carbohydrate or ketogenic interventions may exhibit persistent dysglycemia due to unaddressed oxidative stress, chronic cortisol elevation, micronutrient depletion, or endothelial dysfunction. Integrating vitamin C assessment and adrenal–stress evaluation may help identify resistant phenotypes and guide more comprehensive treatment strategies.
6.3. Implications for Cardiometabolic and Chronic Disease Care
Because insulin resistance, oxidative stress, and cortisol dysregulation contribute to cardiovascular disease, NAFLD, and accelerated biological aging, the ICV Axis may provide a framework for identifying individuals at high risk even when traditional parameters appear normal. Addressing vitamin C insufficiency and cortisol disruption may improve metabolic flexibility, endothelial function, and mitochondrial resilience across diverse patient populations.
6.4. Implications for Lifestyle, Stress, and Circadian Medicine
The ICV model reinforces the need for stress-modulating interventions—sleep optimization, circadian alignment, physical activity, and psychosocial resilience—alongside nutritional support. This broader approach aligns with real-world clinical experience that metabolic therapies often fail when chronic stress or micronutrient depletion remains untreated.
6.5. A New Conceptual Lens for Integrative Orthomolecular Medicine (IOM)
The ICV Axis is consistent with IOM principles emphasizing biochemical individuality, redox balance, mitochondrial function, and structural nutrient sufficiency. By framing insulin dysregulation within a broader network of micronutrients and hormonal physiology, the ICV model provides a conceptual bridge between orthomolecular medicine, metabolic therapy, and modern endocrinology.
7. Conclusions
Insulin homeostasis is traditionally viewed as a function of dietary carbohydrate load, adiposity, and lifestyle behavior. However, substantial evidence indicates that insulin dynamics are deeply intertwined with cortisol physiology, oxidative stress, mitochondrial function, endothelial nitric oxide signaling, and micronutrient availability—particularly vitamin C.
This narrative review synthesizes these domains and proposes the Insulin–Cortisol–Vitamin C (ICV) Axis, a novel conceptual framework that unifies metabolic, endocrine, and redox physiology. The model suggests that disturbances in any component of the axis may propagate through the system, contributing to insulin resistance, metabolic syndrome, BHRT instability, dysglycemia, and impaired responses to lifestyle or pharmacologic interventions.
Although the ICV Axis remains a theoretical construct, it is grounded in established endocrine physiology, vitamin C biochemistry, oxidative stress research, and clinical observations in metabolic and chronic disease practice. Future mechanistic studies, interventional trials, and clinical evaluations are needed to validate the ICV model, assess its predictive utility, and explore therapeutic strategies targeting this triad.
The ICV Axis represents a paradigm shift in the understanding and treatment of insulin resistance and related chronic diseases—expanding the scope of metabolic and hormonal medicine to include redox biology and micronutrient sufficiency as central determinants of metabolic health.
Additional detail establishing the important impact of this ICV model is available[143].
Author Contributions
Conceptualization: T.E.L., R.H., R.Z.C. Writing – original draft: R.Z.C. Writing – review & editing: All authors. Supervision: R.Z.C.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable.
Acknowledgments
The authors gratefully acknowledge Dr. Thomas E. Levy for originating the conceptual foundation of the Insulin–Cortisol–Vitamin C (ICV) Axis. His early insights into the interplay between redox biology, vitamin C physiology, and metabolic–endocrine regulation directly inspired the development of this integrated model.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rains, J.L.; Jain, S.K. Oxidative Stress, Insulin Signaling, and Diabetes. Free Radic Biol Med 2011, 50, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.L.; Goldfine, I.D.; Maddux, B.A.; Grodsky, G.M. Are Oxidative Stress−Activated Signaling Pathways Mediators of Insulin Resistance and β-Cell Dysfunction? Diabetes 2003, 52, 1–8. [Google Scholar] [CrossRef]
- Karin, O.; Raz, M.; Tendler, A.; Bar, A.; Korem Kohanim, Y.; Milo, T.; Alon, U. A New Model for the HPA Axis Explains Dysregulation of Stress Hormones on the Timescale of Weeks. Mol Syst Biol 2020, 16, e9510. [Google Scholar] [CrossRef]
- James, K.A.; Stromin, J.I.; Steenkamp, N.; Combrinck, M.I. Understanding the Relationships between Physiological and Psychosocial Stress, Cortisol and Cognition. Front Endocrinol (Lausanne) 2023, 14, 1085950. [Google Scholar] [CrossRef]
- Young, E.S.; Doom, J.R.; Farrell, A.K.; Carlson, E.A.; Englund, M.M.; Miller, G.E.; Gunnar, M.R.; Roisman, G.I.; Simpson, J.A. Life Stress and Cortisol Reactivity: An Exploratory Analysis of the Effects of Stress Exposure across Life on HPA-Axis Functioning. Dev Psychopathol 2021, 33, 301–312. [Google Scholar] [CrossRef]
- Ortiz, R.; Kluwe, B.; Lazarus, S.; Teruel, M.N.; Joseph, J.J. Cortisol and Cardiometabolic Disease: A Target for Advancing Health Equity. Trends Endocrinol Metab 2022, 33, 786–797. [Google Scholar] [CrossRef] [PubMed]
- Hackett, R.A.; Kivimäki, M.; Kumari, M.; Steptoe, A. Diurnal Cortisol Patterns, Future Diabetes, and Impaired Glucose Metabolism in the Whitehall II Cohort Study. J Clin Endocrinol Metab 2016, 101, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Maury, E.; Ramsey, K.M.; Bass, J. Circadian Rhythms and Metabolic Syndrome: From Experimental Genetics to Human Disease. Circ Res 2010, 106, 447–462. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.-Y.; Ima-Nirwana, S. Vitamin C: A Review on Its Role in the Management of Metabolic Syndrome. Int J Med Sci 2020, 17, 1625–1638. [Google Scholar] [CrossRef]
- Guo, H.; Ding, J.; Liu, Q.; Li, Y.; Liang, J.; Zhang, Y. Vitamin C and Metabolic Syndrome: A Meta-Analysis of Observational Studies. Front. Nutr. 2021, 8. [Google Scholar] [CrossRef]
- Sabri, M.; Ghaffari, G.; Hashemipour, M.; Mostofizadeh, N.; Koushki, A.M. Effect of Long-Term Vitamin C Intake on Vascular Endothelial Function in Diabetic Children and Adolescents: A Pilot Study. J Res Med Sci 2016, 21, 119. [Google Scholar] [CrossRef] [PubMed]
- Mason, S.A.; Keske, M.A.; Wadley, G.D. Effects of Vitamin C Supplementation on Glycemic Control and Cardiovascular Risk Factors in People With Type 2 Diabetes: A GRADE-Assessed Systematic Review and Meta-Analysis of Randomized Controlled Trials. Diabetes Care 2021, 44, 618–630. [Google Scholar] [CrossRef]
- May, J.M.; Harrison, F.E. Role of Vitamin C in the Function of the Vascular Endothelium. Antioxid Redox Signal 2013, 19, 2068–2083. [Google Scholar] [CrossRef]
- Beglaryan, N.; Hakobyan, G.; Nazaretyan, E. Vitamin C Supplementation Alleviates Hypercortisolemia Caused by Chronic Stress. Stress Health 2024, 40, e3347. [Google Scholar] [CrossRef]
- Marik, P.E. Vitamin C: An Essential “Stress Hormone” during Sepsis. J Thorac Dis 2020, 12 (Suppl 1), S84–S88. [Google Scholar] [CrossRef] [PubMed]
- Nooraei, N.; Fathi, M.; Edalat, L.; Behnaz, F.; Mohajerani, S.A.; Dabbagh, A. Effect of Vitamin C on Serum Cortisol after Etomidate Induction of Anesthesia.
- Panahi, J.R.; Paknezhad, S.P.; Vahedi, A.; Shahsavarinia, K.; Laleh, M.R.; Soleimanpour, H. Effect of Vitamin C on Adrenal Suppression Following Etomidate for Rapid Sequence Induction in Trauma Patients: A Randomized Clinical Trial. BMC Anesthesiol 2023, 23, 104. [Google Scholar] [CrossRef]
- Patak, P.; Willenberg, H.S.; Bornstein, S.R. Vitamin C Is an Important Cofactor for Both Adrenal Cortex and Adrenal Medulla. Endocr Res 2004, 30, 871–875. [Google Scholar] [CrossRef]
- Redmann, A.; Möbius, K.; Hiller, H.H.; Oelkers, W.; Bähr, V. Ascorbate Depletion Prevents Aldosterone Stimulation by Sodium Deficiency in the Guinea Pig. Eur J Endocrinol 1995, 133, 499–506. [Google Scholar] [CrossRef]
- Kumar, S.; Saxena, J.; Srivastava, V.K.; Kaushik, S.; Singh, H.; Abo-El-Sooud, K.; Abdel-Daim, M.M.; Jyoti, A.; Saluja, R. The Interplay of Oxidative Stress and ROS Scavenging: Antioxidants as a Therapeutic Potential in Sepsis. Vaccines (Basel) 2022, 10, 1575. [Google Scholar] [CrossRef]
- Zheng, H.; Xu, Y.; Liehn, E.A.; Rusu, M. Vitamin C as Scavenger of Reactive Oxygen Species during Healing after Myocardial Infarction. Int J Mol Sci 2024, 25, 3114. [Google Scholar] [CrossRef]
- Foyer, C.H.; Kunert, K. The Ascorbate-Glutathione Cycle Coming of Age. J Exp Bot 2024, 75, 2682–2699. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of Ascorbate-Glutathione Pathway in Mitigating Oxidative Damage in Plants under Abiotic Stress. Antioxidants (Basel) 2019, 8, 384. [Google Scholar] [CrossRef]
- Rebouche, C.J. Ascorbic Acid and Carnitine Biosynthesis. Am J Clin Nutr 1991, 54 (6 Suppl), 1147S–1152S. [Google Scholar] [CrossRef]
- Aumailley, L.; Bourassa, S.; Gotti, C.; Droit, A.; Lebel, M. Vitamin C Modulates the Levels of Several Proteins of the Mitochondrial Complex III and Its Activity in the Mouse Liver. Redox Biol 2022, 57, 102491. [Google Scholar] [CrossRef]
- Hong, R.; Min, S.; Huang, J.; Zou, M.; Zhou, D.; Liang, Y. High-Dose Vitamin C Promotes Mitochondrial Biogenesis in HCT116 Colorectal Cancer Cells by Regulating the AMPK/PGC-1α Signaling Pathway. J Cancer Res Clin Oncol 2025, 151, 167. [Google Scholar] [CrossRef] [PubMed]
- Sasidharan Nair, V.; Huehn, J. Impact of Vitamin C on the Development, Differentiation and Functional Properties of T Cells. Eur J Microbiol Immunol (Bp) 2024, 14, 67–74. [Google Scholar] [CrossRef]
- Cerullo, G.; Negro, M.; Parimbelli, M.; Pecoraro, M.; Perna, S.; Liguori, G.; Rondanelli, M.; Cena, H.; D’Antona, G. The Long History of Vitamin C: From Prevention of the Common Cold to Potential Aid in the Treatment of COVID-19. Front Immunol 2020, 11, 574029. [Google Scholar] [CrossRef]
- Fesahat, F.; Norouzi, E.; Seifati, S.M.; Hamidian, S.; Hosseini, A.; Zare, F. Impact of Vitamin C on Gene Expression Profile of Inflammatory and Anti-Inflammatory Cytokines in the Male Partners of Couples with Recurrent Pregnancy Loss. Int J Inflam 2022, 2022, 1222533. [Google Scholar] [CrossRef] [PubMed]
- Uscio, L.V. d’; Milstien, S.; Richardson, D.; Smith, L.; Katusic, Z.S. Long-Term Vitamin C Treatment Increases Vascular Tetrahydrobiopterin Levels and Nitric Oxide Synthase Activity. Circ Res 2003, 92, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Taddei, S.; Virdis, A.; Ghiadoni, L.; Magagna, A.; Salvetti, A. Vitamin C Improves Endothelium-Dependent Vasodilation by Restoring Nitric Oxide Activity in Essential Hypertension. Circulation 1998, 97, 2222–2229. [Google Scholar] [CrossRef]
- Williams, M.J.A.; Sutherland, W.H.F.; McCormick, M.P.; Jong, S.A. de; McDonald, J.R.; Walker, R.J. Vitamin C Improves Endothelial Dysfunction in Renal Allograft Recipients. Nephrol Dial Transplant 2001, 16, 1251–1255. [Google Scholar] [CrossRef]
- Mitani, F.; Ogishima, T.; Mukai, K.; Suematsu, M. Ascorbate Stimulates Monooxygenase-Dependent Steroidogenesis in Adrenal Zona Glomerulosa. Biochemical and Biophysical Research Communications 2005, 338, 483–490. [Google Scholar] [CrossRef]
- Wu, X.; Iguchi, T.; Itoh, N.; Okamoto, K.; Takagi, T.; Tanaka, K.; Nakanishi, T. Ascorbic Acid Transported by Sodium-Dependent Vitamin C Transporter 2 Stimulates Steroidogenesis in Human Choriocarcinoma Cells. Endocrinology 2008, 149, 73–83. [Google Scholar] [CrossRef]
- Liu, M.; Park, S. A Causal Relationship between Vitamin C Intake with Hyperglycemia and Metabolic Syndrome Risk: A Two-Sample Mendelian Randomization Study. Antioxidants (Basel) 2022, 11, 857. [Google Scholar] [CrossRef] [PubMed]
- Nosratabadi, S.; Ashtary-Larky, D.; Hosseini, F.; Namkhah, Z.; Mohammadi, S.; Salamat, S.; Nadery, M.; Yarmand, S.; Zamani, M.; Wong, A.; Asbaghi, O. The Effects of Vitamin C Supplementation on Glycemic Control in Patients with Type 2 Diabetes: A Systematic Review and Meta-Analysis. Diabetes & Metabolic Syndrome: Clinical Research & Reviews 2023, 17, 102824. [Google Scholar] [CrossRef]
- Schernthaner-Reiter, M.H.; Wolf, P.; Vila, G.; Luger, A. The Interaction of Insulin and Pituitary Hormone Syndromes. Front. Endocrinol. 2021, 12. [Google Scholar] [CrossRef]
- Adam, T.C.; Hasson, R.E.; Ventura, E.E.; Toledo-Corral, C.; Le, K.-A.; Mahurkar, S.; Lane, C.J.; Weigensberg, M.J.; Goran, M.I. Cortisol Is Negatively Associated with Insulin Sensitivity in Overweight Latino Youth. J Clin Endocrinol Metab 2010, 95, 4729–4735. [Google Scholar] [CrossRef]
- Geer, E.B.; Islam, J.; Buettner, C. Mechanisms of Glucocorticoid-Induced Insulin Resistance: Focus on Adipose Tissue Function and Lipid Metabolism. Endocrinol Metab Clin North Am 2014, 43, 75–102. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Maleki, M.; Butler, A.E.; Jamialahmadi, T.; Sahebkar, A. Molecular Mechanisms Linking Stress and Insulin Resistance. EXCLI J 2022, 21, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, M.; Takei, M.; Ishii, H.; Sato, Y. Glucose-Stimulated Insulin Secretion: A Newer Perspective. J Diabetes Investig 2013, 4, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Plecitá-Hlavatá, L.; Jabůrek, M.; Holendová, B.; Tauber, J.; Pavluch, V.; Berková, Z.; Cahová, M.; Schröder, K.; Brandes, R.P.; Siemen, D.; Ježek, P. Glucose-Stimulated Insulin Secretion Fundamentally Requires H2O2 Signaling by NADPH Oxidase 4. Diabetes 2020, 69, 1341–1354. [Google Scholar] [CrossRef]
- Deepa Maheshvare, M.; Raha, S.; König, M.; Pal, D. A Pathway Model of Glucose-Stimulated Insulin Secretion in the Pancreatic β-Cell. Front Endocrinol (Lausanne) 2023, 14, 1185656. [Google Scholar] [CrossRef]
- Peng, X.; Wang, K.; Chen, L. Biphasic Glucose-Stimulated Insulin Secretion over Decades: A Journey from Measurements and Modeling to Mechanistic Insights. Life Metab 2025, 4, loae038. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.K. Mechanisms of Action and Therapeutic Applications of GLP-1 and Dual GIP/GLP-1 Receptor Agonists. Front Endocrinol (Lausanne) 2024, 15, 1431292. [Google Scholar] [CrossRef] [PubMed]
- Reimann, F.; Diakogiannaki, E.; Hodge, D.; Gribble, F.M. Cellular Mechanisms Governing Glucose-Dependent Insulinotropic Polypeptide Secretion. Peptides 2020, 125, 170206. [Google Scholar] [CrossRef]
- Holst, J.J.; Rosenkilde, M.M. GIP as a Therapeutic Target in Diabetes and Obesity: Insight From Incretin Co-Agonists. J Clin Endocrinol Metab 2020, 105, e2710–2716. [Google Scholar] [CrossRef]
- Newsholme, P.; Krause, M. Nutritional Regulation of Insulin Secretion: Implications for Diabetes. Clin Biochem Rev 2012, 33, 35–47. [Google Scholar]
- Zhang, T.; Li, C. Mechanisms of Amino Acid-Stimulated Insulin Secretion in Congenital Hyperinsulinism. Acta Biochim Biophys Sin (Shanghai) 2013, 45, 36–43. [Google Scholar] [CrossRef]
- Itoh, Y.; Kawamata, Y.; Harada, M.; Kobayashi, M.; Fujii, R.; Fukusumi, S.; Ogi, K.; Hosoya, M.; Tanaka, Y.; Uejima, H.; Tanaka, H.; Maruyama, M.; Satoh, R.; Okubo, S.; Kizawa, H.; Komatsu, H.; Matsumura, F.; Noguchi, Y.; Shinohara, T.; Hinuma, S.; Fujisawa, Y.; Fujino, M. Free Fatty Acids Regulate Insulin Secretion from Pancreatic Beta Cells through GPR40. Nature 2003, 422, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-F. Free Fatty Acid Receptors in the Endocrine Regulation of Glucose Metabolism: Insight from Gastrointestinal-Pancreatic-Adipose Interactions. Front Endocrinol (Lausanne) 2022, 13, 956277. [Google Scholar] [CrossRef]
- Honka, M.-J.; Latva-Rasku, A.; Bucci, M.; Virtanen, K.A.; Hannukainen, J.C.; Kalliokoski, K.K.; Nuutila, P. Insulin-Stimulated Glucose Uptake in Skeletal Muscle, Adipose Tissue and Liver: A Positron Emission Tomography Study. Eur J Endocrinol 2018, 178, 523–531. [Google Scholar] [CrossRef]
- Chadt, A.; Al-Hasani, H. Glucose Transporters in Adipose Tissue, Liver, and Skeletal Muscle in Metabolic Health and Disease. Pflugers Arch 2020, 472, 1273–1298. [Google Scholar] [CrossRef] [PubMed]
- Chutia, H.; Lynrah, K.G. Association of Serum Magnesium Deficiency with Insulin Resistance in Type 2 Diabetes Mellitus. J Lab Physicians 2015, 7, 75–78. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Tripathy, D. Skeletal Muscle Insulin Resistance Is the Primary Defect in Type 2 Diabetes. Diabetes Care 2009, 32 (suppl_2), S157–S163. [Google Scholar] [CrossRef] [PubMed]
- Humphries, S.; Kushner, H.; Falkner, B. Low Dietary Magnesium Is Associated with Insulin Resistance in a Sample of Young, Nondiabetic Black Americans*. Am J Hypertens 1999, 12, 747–756. [Google Scholar] [CrossRef]
- Mooren, F.C.; Krüger, K.; Völker, K.; Golf, S.W.; Wadepuhl, M.; Kraus, A. Oral Magnesium Supplementation Reduces Insulin Resistance in Non-Diabetic Subjects - a Double-Blind, Placebo-Controlled, Randomized Trial. Diabetes Obes Metab 2011, 13, 281–284. [Google Scholar] [CrossRef]
- Simental-Mendía, L.E.; Sahebkar, A.; Rodríguez-Morán, M.; Guerrero-Romero, F. A Systematic Review and Meta-Analysis of Randomized Controlled Trials on the Effects of Magnesium Supplementation on Insulin Sensitivity and Glucose Control. Pharmacol Res 2016, 111, 272–282. [Google Scholar] [CrossRef]
- Ferrannini, E.; Iozzo, P.; Virtanen, K.A.; Honka, M.-J.; Bucci, M.; Nuutila, P. Adipose Tissue and Skeletal Muscle Insulin-Mediated Glucose Uptake in Insulin Resistance: Role of Blood Flow and Diabetes. The American Journal of Clinical Nutrition 2018, 108, 749–758. [Google Scholar] [CrossRef]
- BOERSMA, G.J.; HEURLING, K.; PEREIRA, M.J.; JOHANSSON, E.; LUBBERINK, M.; KATSOGIANNOS, P.; SKRTIC, S.; KULLBERG, J.; AHLSTRÖM, H.; ERIKSSON, J.W. Glucose Uptake in Muscle, Visceral Adipose Tissue, and Brain Strongly Predict Whole-Body Insulin Resistance in the Development of Type 2 Diabetes. Diabetes 2018, 67 (Supplement_1), 1790-P. [Google Scholar] [CrossRef]
- Henriksen, E.J.; Diamond-Stanic, M.K.; Marchionne, E.M. Oxidative Stress and the Etiology of Insulin Resistance and Type 2 Diabetes. Free Radic Biol Med 2011, 51, 993–999. [Google Scholar] [CrossRef]
- Hurrle, S.; Hsu, W.H. The Etiology of Oxidative Stress in Insulin Resistance. Biomed J 2017, 40, 257–262. [Google Scholar] [CrossRef]
- Tangvarasittichai, S. Oxidative Stress, Insulin Resistance, Dyslipidemia and Type 2 Diabetes Mellitus. World J Diabetes 2015, 6, 456–480. [Google Scholar] [CrossRef]
- Andreadi, A.; Bellia, A.; Di Daniele, N.; Meloni, M.; Lauro, R.; Della-Morte, D.; Lauro, D. The Molecular Link between Oxidative Stress, Insulin Resistance, and Type 2 Diabetes: A Target for New Therapies against Cardiovascular Diseases. Current Opinion in Pharmacology 2022, 62, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Fiorenza, M.; Onslev, J.; Henríquez-Olguín, C.; Persson, K.W.; Hesselager, S.A.; Jensen, T.E.; Wojtaszewski, J.F.P.; Hostrup, M.; Bangsbo, J. Reducing the Mitochondrial Oxidative Burden Alleviates Lipid-Induced Muscle Insulin Resistance in Humans. Science Advances 2024, 10, eadq4461. [Google Scholar] [CrossRef]
- Yesupatham, A.; Saraswathy, R. Role of Oxidative Stress in Prediabetes Development. Biochemistry and Biophysics Reports 2025, 43, 102069. [Google Scholar] [CrossRef] [PubMed]
- Sangwung, P.; Petersen, K.F.; Shulman, G.I.; Knowles, J.W. Mitochondrial Dysfunction, Insulin Resistance, and Potential Genetic Implications: Potential Role of Alterations in Mitochondrial Function in the Pathogenesis of Insulin Resistance and Type 2 Diabetes. Endocrinology 2020, 161, bqaa017. [Google Scholar] [CrossRef] [PubMed]
- Sergi, D.; Naumovski, N.; Heilbronn, L.K.; Abeywardena, M.; O’Callaghan, N.; Lionetti, L.; Luscombe-Marsh, N. Mitochondrial (Dys)Function and Insulin Resistance: From Pathophysiological Molecular Mechanisms to the Impact of Diet. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef]
- Montgomery, M.K.; Turner, N. Mitochondrial Dysfunction and Insulin Resistance: An Update. Endocr Connect 2015, 4, R1–R15. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, X.; Liu, Y.; Li, Y.; Li, D.; Mei, Z.; Deng, Y. Mitochondrial Quality Control in Diabetes Mellitus and Complications: Molecular Mechanisms and Therapeutic Strategies. Cell Death Dis 2025, 16, 652. [Google Scholar] [CrossRef]
- Petersen, M.C.; Shulman, G.I. Roles of Diacylglycerols and Ceramides in Hepatic Insulin Resistance. Trends Pharmacol Sci 2017, 38, 649–665. [Google Scholar] [CrossRef]
- Sokolowska, E.; Blachnio-Zabielska, A. The Role of Ceramides in Insulin Resistance. Front. Endocrinol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Szendroedi, J.; Yoshimura, T.; Phielix, E.; Koliaki, C.; Marcucci, M.; Zhang, D.; Jelenik, T.; Müller, J.; Herder, C.; Nowotny, P.; Shulman, G.I.; Roden, M. Role of Diacylglycerol Activation of PKCθ in Lipid-Induced Muscle Insulin Resistance in Humans. Proceedings of the National Academy of Sciences 2014, 111, 9597–9602. [Google Scholar] [CrossRef]
- Leproult, R.; Holmbäck, U.; Van Cauter, E. Circadian Misalignment Augments Markers of Insulin Resistance and Inflammation, Independently of Sleep Loss. Diabetes 2014, 63, 1860–1869. [Google Scholar] [CrossRef]
- Catalano, F.; De Vito, F.; Cassano, V.; Fiorentino, T.V.; Sciacqua, A.; Hribal, M.L. Circadian Clock Desynchronization and Insulin Resistance. Int J Environ Res Public Health 2022, 20, 29. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.T.; Kondo, T.; Ashry, A.; Fu, Y.; Okawa, H.; Sawangmake, C.; Egusa, H. Effect of Circadian Clock Disruption on Type 2 Diabetes. Front. Physiol. 2024, 15. [Google Scholar] [CrossRef]
- Morris, C.J.; Yang, J.N.; Garcia, J.I.; Myers, S.; Bozzi, I.; Wang, W.; Buxton, O.M.; Shea, S.A.; Scheer, F.A.J.L. Endogenous Circadian System and Circadian Misalignment Impact Glucose Tolerance via Separate Mechanisms in Humans. Proceedings of the National Academy of Sciences 2015, 112, E2225–E2234. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Chauhan, R.; Devi, S. Biological Connection of Circadian Rhythm and Insulin Resistance: A Review. Biological Rhythm Research 2025, 56, 524–540. [Google Scholar] [CrossRef]
- Beaupere, C.; Liboz, A.; Fève, B.; Blondeau, B.; Guillemain, G. Molecular Mechanisms of Glucocorticoid-Induced Insulin Resistance. Int J Mol Sci 2021, 22, 623. [Google Scholar] [CrossRef]
- Guo, K.; Zhang, L.; Ye, J.; Niu, X.; Jiang, H.; Gan, S.; Zhou, J.; Yang, L.; Zhou, Z. Metabolic Syndrome Associated with Higher Glycemic Variability in Type 1 Diabetes: A Multicenter Cross-Sectional Study in China. Front. Endocrinol. 2022, 13. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Yang, Y.; Wang, Q.; Sun, Y.; Shen, R.; Zhang, M. Glycemic Variability in Type 2 Diabetic Patients with Metabolic Dysfunction-Associated Steatotic Liver Disease: A Case-Control Study. Ann Med 2025, 57, 2548976. [Google Scholar] [CrossRef]
- Galgani, J.E.; Moro, C.; Ravussin, E. Metabolic Flexibility and Insulin Resistance. Am J Physiol Endocrinol Metab 2008, 295, E1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Buscemi, S.; Verga, S.; Cottone, S.; Azzolina, V.; Buscemi, B.; Gioia, D.; Cerasola, G. Glycaemic Variability and Inflammation in Subjects with Metabolic Syndrome. Acta Diabetol 2009, 46, 55–61. [Google Scholar] [CrossRef]
- Metwally, A.A.; Perelman, D.; Park, H.; Wu, Y.; Jha, A.; Sharp, S.; Celli, A.; Ayhan, E.; Abbasi, F.; Gloyn, A.L.; McLaughlin, T.; Snyder, M.P. Prediction of Metabolic Subphenotypes of Type 2 Diabetes via Continuous Glucose Monitoring and Machine Learning. Nat. Biomed. Eng 2025, 9, 1222–1239. [Google Scholar] [CrossRef]
- Ogiso, K.; Shayo, S.C.; Kawade, S.; Hashiguchi, H.; Deguchi, T.; Nishio, Y. Repeated Glucose Spikes and Insulin Resistance Synergistically Deteriorate Endothelial Function and Bardoxolone Methyl Ameliorates Endothelial Dysfunction. PLOS ONE 2022, 17, e0263080. [Google Scholar] [CrossRef]
- Node, K.; Inoue, T. Postprandial Hyperglycemia as an Etiological Factor in Vascular Failure. Cardiovasc Diabetol 2009, 8, 23. [Google Scholar] [CrossRef]
- Koska, J.; Schwartz, E.A.; Mullin, M.P.; Schwenke, D.C.; Reaven, P.D. Improvement of Postprandial Endothelial Function After a Single Dose of Exenatide in Individuals With Impaired Glucose Tolerance and Recent-Onset Type 2 Diabetes. Diabetes Care 2010, 33, 1028–1030. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Tsai, M.-F.; Thorat, R.S.; Xiao, D.; Zhang, X.; Sandhu, A.K.; Edirisinghe, I.; Burton-Freeman, B.M. Endothelial Function and Postprandial Glucose Control in Response to Test-Meals Containing Herbs and Spices in Adults With Overweight/Obesity. Front Nutr 2022, 9, 811433. [Google Scholar] [CrossRef]
- Mah, E.; Bruno, R.S. Postprandial Hyperglycemia on Vascular Endothelial Function: Mechanisms and Consequences. Nutr Res 2012, 32, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Kc, S.; Cárcamo, J.M.; Golde, D.W. Vitamin C Enters Mitochondria via Facilitative Glucose Transporter 1 (Glut1) and Confers Mitochondrial Protection against Oxidative Injury. FASEB J 2005, 19, 1657–1667. [Google Scholar] [CrossRef]
- Besse-Patin, A.; Estall, J.L. An Intimate Relationship between ROS and Insulin Signalling: Implications for Antioxidant Treatment of Fatty Liver Disease. Int J Cell Biol 2014, 2014, 519153. [Google Scholar] [CrossRef]
- Picklo, M.J.; Thyfault, J.P. Vitamin E and Vitamin C Do Not Reduce Insulin Sensitivity but Inhibit Mitochondrial Protein Expression in Exercising Obese Rats. Appl Physiol Nutr Metab 2015, 40, 343–352. [Google Scholar] [CrossRef]
- Carr, A.C.; Frampton, C.; Lunt, H. Metabolic Syndrome Is Associated with Increased Vitamin C Requirements in the US National Health and Nutrition Examination Survey. Nutrition Research 2025, 141, 1–9. [Google Scholar] [CrossRef]
- Pleiner, J.; Schaller, G.; Mittermayer, F.; Bayerle-Eder, M.; Roden, M.; Wolzt, M. FFA-Induced Endothelial Dysfunction Can Be Corrected by Vitamin C. J Clin Endocrinol Metab 2002, 87, 2913–2917. [Google Scholar] [CrossRef]
- Ashor, A.W.; Lara, J.; Mathers, J.C.; Siervo, M. Effect of Vitamin C on Endothelial Function in Health and Disease: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Atherosclerosis 2014, 235, 9–20. [Google Scholar] [CrossRef]
- Afkhami-Ardekani, M.; Shojaoddiny-Ardekani, A. Effect of Vitamin C on Blood Glucose, Serum Lipids & Serum Insulin in Type 2 Diabetes Patients. Indian J Med Res 2007, 126, 471–474. [Google Scholar]
- Wang, T.; Wang, J.; Hu, X.; Huang, X.-J.; Chen, G.-X. Current Understanding of Glucose Transporter 4 Expression and Functional Mechanisms. World J Biol Chem 2020, 11, 76–98. [Google Scholar] [CrossRef] [PubMed]
- Albaik, M.; Sheikh Saleh, D.; Kauther, D.; Mohammed, H.; Alfarra, S.; Alghamdi, A.; Ghaboura, N.; Sindi, I.A. Bridging the Gap: Glucose Transporters, Alzheimer’s, and Future Therapeutic Prospects. Front. Cell Dev. Biol. 2024, 12. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.Y.; Otsuka, M.; Arakawa, N. Ascorbate Indirectly Stimulates Fatty Acid Utilization in Primary Cultured Guinea Pig Hepatocytes by Enhancing Carnitine Synthesis123. The Journal of Nutrition 1994, 124, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Eigler, N.; Saccà, L.; Sherwin, R.S. Synergistic Interactions of Physiologic Increments of Glucagon, Epinephrine, and Cortisol in the Dog: A MODEL FOR STRESS-INDUCED HYPERGLYCEMIA. J Clin Invest 1979, 63, 114–123. [Google Scholar] [CrossRef]
- Gelfand, R.A.; Matthews, D.E.; Bier, D.M.; Sherwin, R.S. Role of Counterregulatory Hormones in the Catabolic Response to Stress. J Clin Invest 1984, 74, 2238–2248. [Google Scholar] [CrossRef]
- Joseph, J.J.; Golden, S.H. Cortisol Dysregulation: The Bidirectional Link between Stress, Depression, and Type 2 Diabetes Mellitus. Ann N Y Acad Sci 2017, 1391, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Kamba, A.; Daimon, M.; Murakami, H.; Otaka, H.; Matsuki, K.; Sato, E.; Tanabe, J.; Takayasu, S.; Matsuhashi, Y.; Yanagimachi, M.; Terui, K.; Kageyama, K.; Tokuda, I.; Takahashi, I.; Nakaji, S. Association between Higher Serum Cortisol Levels and Decreased Insulin Secretion in a General Population. PLOS ONE 2016, 11, e0166077. [Google Scholar] [CrossRef]
- Ohira, M.; Kawagoe, N.; Kameyama, C.; Kondou, Y.; Igarashi, M.; Ueshiba, H. Association of Serum Cortisol with Insulin Secretion and Plasma Aldosterone with Insulin Resistance in Untreated Type 2 Diabetes: A Cross-Sectional Study. Diabetol Metab Syndr 2025, 17, 144. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, R.; Kluwe, B.; Odei, J.B.; Echouffo Tcheugui, J.B.; Sims, M.; Kalyani, R.R.; Bertoni, A.G.; Golden, S.H.; Joseph, J.J. The Association of Morning Serum Cortisol with Glucose Metabolism and Diabetes: The Jackson Heart Study. Psychoneuroendocrinology 2019, 103, 25–32. [Google Scholar] [CrossRef]
- Sharma, A.; Vella, A. Glucose Metabolism in Cushing’s Syndrome. Curr Opin Endocrinol Diabetes Obes 2020, 27, 140–145. [Google Scholar] [CrossRef]
- DeFronzo, R.A.; Auchus, R.J. Cushing Syndrome, Hypercortisolism, and Glucose Homeostasis: A Review. Diabetes 2025, 74, 2168–2178. [Google Scholar] [CrossRef]
- Li, J.-X.; Cummins, C.L. Fresh Insights into Glucocorticoid-Induced Diabetes Mellitus and New Therapeutic Directions. Nat Rev Endocrinol 2022, 18, 540–557. [Google Scholar] [CrossRef]
- Lundgren, M.; Burén, J.; Ruge, T.; Myrnäs, T.; Eriksson, J.W. Glucocorticoids Down-Regulate Glucose Uptake Capacity and Insulin-Signaling Proteins in Omental But Not Subcutaneous Human Adipocytes. J Clin Endocrinol Metab 2004, 89, 2989–2997. [Google Scholar] [CrossRef]
- Varlamov, E.V.; Purnell, J.Q.; Fleseriu, M. Glucocorticoid Receptor Antagonism as a New “Remedy” for Insulin Resistance-Not There Yet! J Clin Endocrinol Metab 2021, 106, e2447–e2449. [Google Scholar] [CrossRef]
- Chapela, S.P.; Simancas-Racines, D.; Montalvan, M.; Frias-Toral, E.; Simancas-Racines, A.; Muscogiuri, G.; Barrea, L.; Sarno, G.; Martínez, P.I.; Reberendo, M.J.; Llobera, N.D.; Stella, C.A. Signals for Muscular Protein Turnover and Insulin Resistance in Critically Ill Patients: A Narrative Review. Nutrients 2023, 15, 1071. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Wang, H.; Lee, I.H.; Du, J.; Mitch, W.E. Endogenous Glucocorticoids and Impaired Insulin Signaling Are Both Required to Stimulate Muscle Wasting under Pathophysiological Conditions in Mice. J Clin Invest 2009, 119, 3059–3069. [Google Scholar] [CrossRef]
- Louard, R.J.; Bhushan, R.; Gelfand, R.A.; Barrett, E.J.; Sherwin, R.S. Glucocorticoids Antagonize Insulin’s Antiproteolytic Action on Skeletal Muscle in Humans. J Clin Endocrinol Metab 1994, 79, 278–284. [Google Scholar] [CrossRef]
- Simmons, P.S.; Miles, J.M.; Gerich, J.E.; Haymond, M.W. Increased Proteolysis. An Effect of Increases in Plasma Cortisol within the Physiologic Range. J Clin Invest 1984, 73, 412–420. [Google Scholar] [CrossRef]
- Wang, X.; Hu, Z.; Hu, J.; Du, J.; Mitch, W.E. Insulin Resistance Accelerates Muscle Protein Degradation: Activation of the Ubiquitin-Proteasome Pathway by Defects in Muscle Cell Signaling. Endocrinology 2006, 147, 4160–4168. [Google Scholar] [CrossRef]
- Akalestou, E.; Genser, L.; Rutter, G.A. Glucocorticoid Metabolism in Obesity and Following Weight Loss. Front. Endocrinol. 2020, 11. [Google Scholar] [CrossRef]
- Boscaro, M.; Giacchetti, G.; Ronconi, V. Visceral Adipose Tissue: Emerging Role of Gluco- and Mineralocorticoid Hormones in the Setting of Cardiometabolic Alterations. Ann N Y Acad Sci 2012, 1264, 87–102. [Google Scholar] [CrossRef]
- Misra, M.; Bredella, M.A.; Tsai, P.; Mendes, N.; Miller, K.K.; Klibanski, A. Lower Growth Hormone and Higher Cortisol Are Associated with Greater Visceral Adiposity, Intramyocellular Lipids, and Insulin Resistance in Overweight Girls. American Journal of Physiology-Endocrinology and Metabolism 2008, 295, E385–E392. [Google Scholar] [CrossRef]
- Ould Bessi, N.; Touahria Miliani, Y.; Damou, R.; EL Mehdaoui, M.A.; Kemache, A.; Ait Abdelkader, B. Association between 8 a.m. Cortisol Levels and Insulin Resistance in Healthy Individuals from Algiers. Obesity Medicine 2025, 58, 100648. [Google Scholar] [CrossRef]
- Yan, Y.-X.; Xiao, H.-B.; Wang, S.-S.; Zhao, J.; He, Y.; Wang, W.; Dong, J. Investigation of the Relationship Between Chronic Stress and Insulin Resistance in a Chinese Population. J Epidemiol 2016, 26, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Lazzarino, A.I.; Hamer, M.; Gaze, D.; Collinson, P.; Steptoe, A. The Association between Cortisol Response to Mental Stress and High-Sensitivity Cardiac Troponin T Plasma Concentration in Healthy Adults. J Am Coll Cardiol 2013, 62, 1694–1701. [Google Scholar] [CrossRef] [PubMed]
- Vage, A.; Gormley, G.; Hamilton, P.K. The Effects of Controlled Acute Psychological Stress on Serum Cortisol and Plasma Metanephrine Concentrations in Healthy Subjects. Ann Clin Biochem 2025, 62, 165–173. [Google Scholar] [CrossRef]
- Zhu, X.; Zhu, Y.; Huang, J.; Zhou, Y.; Tong, J.; Zhang, P.; Luo, X.; Chen, S.; Tian, B.; Tan, S.; Wang, Z.; Han, X.; Tian, L.; Li, C.-S.R.; Hong, L.E.; Tan, Y. Abnormal Cortisol Profile during Psychosocial Stress among Patients with Schizophrenia in a Chinese Population. Sci Rep 2022, 12, 18591. [Google Scholar] [CrossRef]
- Andreadi, A.; Andreadi, S.; Todaro, F.; Ippoliti, L.; Bellia, A.; Magrini, A.; Chrousos, G.P.; Lauro, D. Modified Cortisol Circadian Rhythm: The Hidden Toll of Night-Shift Work. Int J Mol Sci 2025, 26, 2090. [Google Scholar] [CrossRef]
- Marhefkova, N.; Sládek, M.; Sumová, A.; Dubsky, M. Circadian Dysfunction and Cardio-Metabolic Disorders in Humans. Front. Endocrinol. 2024, 15. [Google Scholar] [CrossRef]
- Schrader, L.A.; Ronnekleiv-Kelly, S.M.; Hogenesch, J.B.; Bradfield, C.A.; Malecki, K.M.C. Circadian Disruption, Clock Genes, and Metabolic Health. J Clin Invest 2024, 134. [Google Scholar] [CrossRef]
- Beaudry, J.L.; D’souza, A.M.; Teich, T.; Tsushima, R.; Riddell, M.C. Exogenous Glucocorticoids and a High-Fat Diet Cause Severe Hyperglycemia and Hyperinsulinemia and Limit Islet Glucose Responsiveness in Young Male Sprague-Dawley Rats. Endocrinology 2013, 154, 3197–3208. [Google Scholar] [CrossRef]
- Pofi, R.; Othonos, N.; Marjot, T.; Bonaventura, I.; Barrett, A.; White, S.; Miller, H.; Potter, T.; Bailey, M.; Eastell, R.; Gossiel, F.; Woods, C.; Hazlehurst, J.M.; Hodson, L.; Tomlinson, J.W. Dose-Dependent and Tissue-Specific Adverse Effects of Exogenous Glucocorticoids: Insights for Optimizing Clinical Practice. J Endocrinol Invest 2025, 48, 2067–2076. [Google Scholar] [CrossRef]
- Qi, D.; Rodrigues, B. Glucocorticoids Produce Whole Body Insulin Resistance with Changes in Cardiac Metabolism. American Journal of Physiology-Endocrinology and Metabolism 2007, 292, E654–E667. [Google Scholar] [CrossRef]
- Das, D.; Sen, C.; Goswami, A. Effect of Vitamin C on Adrenal Suppression by Etomidate Induction in Patients Undergoing Cardiac Surgery: A Randomized Controlled Trial. Ann Card Anaesth 2016, 19, 410–417. [Google Scholar] [CrossRef]
- Hornsby, P.J.; Harris, S.E.; Aldern, K.A. The Role of Ascorbic Acid in the Function of the Adrenal Cortex: Studies in Adrenocortical Cells in Culture. Endocrinology 1985, 117, 1264–1271. [Google Scholar] [CrossRef]
- Patani, A.; Balram, D.; Yadav, V.K.; Lian, K.-Y.; Patel, A.; Sahoo, D.K. Harnessing the Power of Nutritional Antioxidants against Adrenal Hormone Imbalance-Associated Oxidative Stress. Front Endocrinol (Lausanne) 2023, 14, 1271521. [Google Scholar] [CrossRef]
- Campión, J.; Milagro, F.I.; Fernández, D.; Martínez, J.A. Vitamin C Supplementation Influences Body Fat Mass and Steroidogenesis-Related Genes When Fed a High-Fat Diet. Int J Vitam Nutr Res 2008, 78, 87–95. [Google Scholar] [CrossRef]
- Idahor, C.O.; Ogunfuwa, O.; Ogbonna, N.; Adigwe, A.; Ogbeide, O.A. Beyond Fluid Therapy: The Role of Vitamin C, Steroids, and Thiamine in Sepsis Management. Cureus 2025, 17, e84666. [Google Scholar] [CrossRef]
- Meščić Macan, A.; Gazivoda Kraljević, T.; Raić-Malić, S. Therapeutic Perspective of Vitamin C and Its Derivatives. Antioxidants (Basel) 2019, 8, 247. [Google Scholar] [CrossRef]
- Bornstein, S.R.; Yoshida-Hiroi, M.; Sotiriou, S.; Levine, M.; Hartwig, H.-G.; Nussbaum, R.L.; Eisenhofer, G. Impaired Adrenal Catecholamine System Function in Mice with Deficiency of the Ascorbic Acid Transporter (SVCT2). The FASEB Journal 2003, 17, 1–13. [Google Scholar] [CrossRef]
- Figueroa-Méndez, R.; Rivas-Arancibia, S. Vitamin C in Health and Disease: Its Role in the Metabolism of Cells and Redox State in the Brain. Front. Physiol. 2015, 6. [Google Scholar] [CrossRef]
- May, J.M.; Qu, Z.; Meredith, M.E. Mechanisms of Ascorbic Acid Stimulation of Norepinephrine Synthesis in Neuronal Cells. Biochem Biophys Res Commun 2012, 426, 148–152. [Google Scholar] [CrossRef]
- Moritz, B.; Schmitz, A.E.; Rodrigues, A.L.S.; Dafre, A.L.; Cunha, M.P. The Role of Vitamin C in Stress-Related Disorders. The Journal of Nutritional Biochemistry 2020, 85, 108459. [Google Scholar] [CrossRef]
- Senmaru, T.; Yamazaki, M.; Okada, H.; Asano, M.; Fukui, M.; Nakamura, N.; Obayashi, H.; Kondo, Y.; Maruyama, N.; Ishigami, A.; Hasegawa, G. Pancreatic Insulin Release in Vitamin C-Deficient Senescence Marker Protein-30/Gluconolactonase Knockout Mice. J Clin Biochem Nutr 2012, 50, 114–118. [Google Scholar] [CrossRef]
- Shu, Y.; Zou, C.; Cai, Y.; He, Q.; Wu, X.; Zhu, H.; Qv, M.; Chao, Y.; Xu, C.; Tang, L.; Wu, X. Vitamin C Deficiency Induces Hypoglycemia and Cognitive Disorder through S-Nitrosylation-Mediated Activation of Glycogen Synthase Kinase 3β. Redox Biol 2022, 56, 102420. [Google Scholar] [CrossRef]
- Yahaya, T.O.; Yusuf, A.B.; Danjuma, J.K.; Usman, B.M.; Ishiaku, Y.M. Mechanistic Links between Vitamin Deficiencies and Diabetes Mellitus: A Review. Egyptian Journal of Basic and Applied Sciences 2021, 8, 189–202. [Google Scholar] [CrossRef]
- Levy, T.E. GLP-1 Receptor Agonists and Vitamin C: A Powerful Anti-Aging Combination. Orthomolecular Medicine News Service 2025, 21. Available online: https://orthomolecular.org/resources/omns/v21n66.shtml.
- Norton, L.; Shannon, C.; Gastaldelli, A.; DeFronzo, R.A. Insulin: The Master Regulator of Glucose Metabolism. Metabolism 2022, 129, 155142. [Google Scholar] [CrossRef]
- Tessari, P. Stepwise Discovery of Insulin Effects on Amino Acid and Protein Metabolism. Nutrients 2023, 16, 119. [Google Scholar] [CrossRef]
- Vargas, E.; Joy, N.V.; Carrillo Sepulveda, M.A. Biochemistry, Insulin Metabolic Effects. In StatPearlsStatPearls Publishing, Treasure Island (FL), 2025. Available online: http://www.ncbi.nlm.nih.gov/books/NBK525983/.
- Bitoska, I.; Krstevska, B.; Milenkovic, T.; Subeska-Stratrova, S.; Petrovski, G.; Mishevska, S.J.; Ahmeti, I.; Todorova, B. Effects of Hormone Replacement Therapy on Insulin Resistance in Postmenopausal Diabetic Women. Open Access Maced J Med Sci 2016, 4, 83–88. [Google Scholar] [CrossRef]
- Members, M. New Meta-Analysis Shows That Hormone Therapy Can Significantly Reduce Insulin Resistance. Available online: https://menopause.org/press-releases/new-meta-analysis-shows-that-hormone-therapy-can-significantly-reduce-insulin-resistance (accessed on 27 November 2025).
- Aroda, V.R.; Christophi, C.A.; Edelstein, S.L.; Perreault, L.; Kim, C.; Golden, S.H.; Horton, E.; Mather, K.J. Circulating Sex Hormone Binding Globulin Levels Are Modified with Intensive Lifestyle Intervention, but Their Changes Did Not Independently Predict Diabetes Risk in the Diabetes Prevention Program. BMJ Open Diab Res Care 2020, 8. [Google Scholar] [CrossRef]
- Le, T.N.; Nestler, J.E.; Strauss, J.F.; Wickham, E.P. Sex Hormone-Binding Globulin and Type 2 Diabetes Mellitus. Trends Endocrinol Metab 2012, 23, 32–40. [Google Scholar] [CrossRef]
- Wallace, I.R.; McKinley, M.C.; Bell, P.M.; Hunter, S.J. Sex Hormone Binding Globulin and Insulin Resistance. Clin Endocrinol (Oxf) 2013, 78, 321–329. [Google Scholar] [CrossRef]
- Ding, H.; Zhang, J.; Zhang, F.; Zhang, S.; Chen, X.; Liang, W.; Xie, Q. Resistance to the Insulin and Elevated Level of Androgen: A Major Cause of Polycystic Ovary Syndrome. Front. Endocrinol. 2021, 12. [Google Scholar] [CrossRef]
- Pateguana, N.B.; Janes, A. The Contribution of Hyperinsulinemia to the Hyperandrogenism of Polycystic Ovary Syndrome. Journal of Metabolic Health 2019, 4, 3. [Google Scholar] [CrossRef]
- Unluhizarci, K.; Karaca, Z.; Kelestimur, F. Role of Insulin and Insulin Resistance in Androgen Excess Disorders. World J Diabetes 2021, 12, 616–629. [Google Scholar] [CrossRef]
- El-Eshmawy, M.M.; Hegazy, A.; El-Baiomy, A.A. Relationship Between IGF-1 and Cortisol/ DHEA-S Ratio in Adult Men With Diabetic Metabolic Syndrome Versus Non-Diabetic Metabolic Syndrome. Journal of Endocrinology and Metabolism 2011, 1, 188–195. [Google Scholar] [CrossRef]
- Xie, M.; Zhong, Y.; Xue, Q.; Wu, M.; Deng, X.; O Santos, H.; Tan, S.C.; Kord-Varkaneh, H.; Jiao, P. Impact of Dehydroepianrosterone (DHEA) Supplementation on Serum Levels of Insulin-like Growth Factor 1 (IGF-1): A Dose-Response Meta-Analysis of Randomized Controlled Trials. Exp Gerontol 2020, 136, 110949. [Google Scholar] [CrossRef]
- Bos, M.M.; Smit, R.A.J.; Trompet, S.; Heemst, D. van; Noordam, R. Thyroid Signaling, Insulin Resistance, and 2 Diabetes Mellitus: A Mendelian Randomization Study. J Clin Endocrinol Metab 2017, 102, 1960–1970. [Google Scholar] [CrossRef]
- Mendez, D.A.; Ortiz, R.M. Thyroid Hormones and the Potential for Regulating Glucose Metabolism in Cardiomyocytes during Insulin Resistance and T2DM. Physiol Rep 2021, 9, e14858. [Google Scholar] [CrossRef]
- Wei, Y.; Li, X.; Cui, R.; Liu, J.; Wang, G. Associations between Sensitivity to Thyroid Hormones and Insulin Resistance in Euthyroid Adults with Obesity. Front. Endocrinol. 2024, 15. [Google Scholar] [CrossRef]
- Begemann, K.; Rawashdeh, O.; Olejniczak, I.; Pilorz, V.; Assis, L.V.M. de; Osorio-Mendoza, J.; Oster, H. Endocrine Regulation of Circadian Rhythms. npj Biol Timing Sleep 2025, 2, 10. [Google Scholar] [CrossRef]
- O’Byrne, N.A.; Yuen, F.; Butt, W.Z.; Liu, P.Y. Sleep and Circadian Regulation of Cortisol: A Short Review. Curr Opin Endocr Metab Res 2021, 18, 178–186. [Google Scholar] [CrossRef]
- Van Cauter, E.; Blackman, J.D.; Roland, D.; Spire, J.P.; Refetoff, S.; Polonsky, K.S. Modulation of Glucose Regulation and Insulin Secretion by Circadian Rhythmicity and Sleep. J Clin Invest 1991, 88, 934–942. [Google Scholar] [CrossRef]
- Anjum, A.; Anwar, H.; Sohail, M.U.; Ali Shah, S.M.; Hussain, G.; Rasul, A.; Ijaz, M.U.; Nisar, J.; Munir, N.; Shahzad, A. The Association between Serum Cortisol, Thyroid Profile, Paraoxonase Activity, Arylesterase Activity and Anthropometric Parameters of Undergraduate Students under Examination Stress. Eur J Inflamm 2021, 19, 20587392211000884. [Google Scholar] [CrossRef]
- Petrowski, K.; Kahaly, G.J. Stress and Thyroid Function—From Bench to Bedside. Endocr Rev 2025, 46, 709–735. [Google Scholar] [CrossRef]
- Sinha, S.R.; Prakash, P.; Keshari, J.R.; Kumari, R.; Prakash, V. Assessment of Serum Cortisol Levels in Hypothyroidism Patients: A Cross-Sectional Study. Cureus 2023, 15, e50199. [Google Scholar] [CrossRef]
- Chattu, V.K.; Chattu, S.K.; Burman, D.; Spence, D.W.; Pandi-Perumal, S.R. The Interlinked Rising Epidemic of Insufficient Sleep and Diabetes Mellitus. Healthcare (Basel) 2019, 7, 37. [Google Scholar] [CrossRef]
- Wei, Z.; Chen, Y.; Upender, R.P. Sleep Disturbance and Metabolic Dysfunction: The Roles of Adipokines. Int J Mol Sci 2022, 23, 1706. [Google Scholar] [CrossRef]
- May, J.M.; Qu, Z.-C.; Nazarewicz, R.; Dikalov, S. Ascorbic Acid Efficiently Enhances Neuronal Synthesis of Norepinephrine from Dopamine. Brain Res Bull 2013, 90, 35–42. [Google Scholar] [CrossRef]
- Biswas, P.; Dellanoce, C.; Vezzoli, A.; Mrakic-Sposta, S.; Malnati, M.; Beretta, A.; Accinni, R. Antioxidant Activity with Increased Endogenous Levels of Vitamin C, E and A Following Dietary Supplementation with a Combination of Glutathione and Resveratrol Precursors. Nutrients 2020, 12, 3224. [Google Scholar] [CrossRef]
- Shang, F.; Lu, M.; Dudek, E.; Reddan, J.; Taylor, A. Vitamin C and Vitamin E Restore the Resistance of GSH-Depleted Lens Cells to H2O2. Free Radical Biology and Medicine 2003, 34, 521–530. [Google Scholar] [CrossRef]
- Tram, N.K.; McLean, R.M.; Swindle-Reilly, K.E. Glutathione Improves the Antioxidant Activity of Vitamin C in Human Lens and Retinal Epithelial Cells: Implications for Vitreous Substitutes. Current Eye Research 2021, 46, 470–481. [Google Scholar] [CrossRef]
- Hoorsan, H.; Simbar, M.; Tehrani, F.R.; Fathi, F.; Mosaffa, N.; Riazi, H.; Akradi, L.; Nasseri, S.; Bazrafkan, S. The Effectiveness of Antioxidant Therapy (Vitamin C) in an Experimentally Induced Mouse Model of Ovarian Endometriosis. Womens Health (Lond Engl) 2022, 18, 17455057221096218. [Google Scholar] [CrossRef]
- Jing, Y.; Lu, H.; Li, J.; He, Z.; Zhao, L.; Zhang, C.; Huang, Z.; Liu, L.; Sun, S.; Ma, S.; Rodriguez Esteban, C.; Fu, X.; Zhao, G.; Izpisua Belmonte, J.C.; Zhang, W.; Qu, J.; Wang, S.; Liu, G.-H. Vitamin C Conveys Geroprotection on Primate Ovaries. Cell Stem Cell 2025, 32, 1723–1740.e9. [Google Scholar] [CrossRef]
- Tatone, C.; Di Emidio, G.; Battaglia, R.; Di Pietro, C. Building a Human Ovarian Antioxidant ceRNA Network “OvAnOx”: A Bioinformatic Perspective for Research on Redox-Related Ovarian Functions and Dysfunctions. Antioxidants (Basel) 2024, 13, 1101. [Google Scholar] [CrossRef]
- Kuo, S.-M.; Stout, A.; Wactawski-Wende, J.; Leppert, P.C. Ascorbic Acid Status in Postmenopausal Women with Hormone Replacement Therapy. Maturitas 2002, 41, 45–50. [Google Scholar] [CrossRef]
- Office of Dietary Supplements - Vitamin C. Available online: https://ods.od.nih.gov/factsheets/VitaminC-HealthProfessional/ (accessed on 27 November 2025).
- Compounded Bioidentical Menopausal Hormone Therapy. Available online: https://www.acog.org/clinical/clinical-guidance/clinical-consensus/articles/2023/11/compounded-bioidentical-menopausal-hormone-therapy (accessed on 27 November 2025).
- Greendale, G.A.; Sternfeld, B.; Huang, M.; Han, W.; Karvonen-Gutierrez, C.; Ruppert, K.; Cauley, J.A.; Finkelstein, J.S.; Jiang, S.-F.; Karlamangla, A.S. Changes in Body Composition and Weight during the Menopause Transition. JCI Insight 2019, 4. [Google Scholar] [CrossRef]
- McBane, S.E.; Borgelt, L.M.; Barnes, K.N.; Westberg, S.M.; Lodise, N.M.; Stassinos, M. Use of Compounded Bioidentical Hormone Therapy in Menopausal Women: An Opinion Statement of the Women’s Health Practice and Research Network of the American College of Clinical Pharmacy. Pharmacotherapy 2014, 34, 410–423. [Google Scholar] [CrossRef]
- National Academies of Sciences, E.; Division, H. and M.; Policy, B. on H.S.; Therapy, C. on the C.U. of T.P. with C.B.H.R.; Jackson, L.M.; Parker, R.M.; Mattison, D.R. The Safety and Effectiveness of Compounded Bioidentical Hormone Therapy. In The Clinical Utility of Compounded Bioidentical Hormone Therapy: A Review of Safety, Effectiveness, and UseNational Academies Press (US), 2020. Available online: https://www.ncbi.nlm.nih.gov/books/NBK562865/.
- Patel, P.; Patil, S.; Kaur, N. Estrogen and Metabolism: Navigating Hormonal Transitions from Perimenopause to Postmenopause. J Midlife Health 2025, 16, 247–256. [Google Scholar] [CrossRef]
- Ruiz, A.D.; Daniels, K.R.; Barner, J.C.; Carson, J.J.; Frei, C.R. Effectiveness of Compounded Bioidentical Hormone Replacement Therapy: An Observational Cohort Study. BMC Womens Health 2011, 11, 27. [Google Scholar] [CrossRef]
- Sood, R.; Shuster, L.; Smith, R.; Vincent, A.; Jatoi, A. Counseling Postmenopausal Women about Bioidentical Hormones: Ten Discussion Points for Practicing Physicians. J Am Board Fam Med 2011, 24, 202–210. [Google Scholar] [CrossRef]
- Stephenson, K.; Neuenschwander, P.F.; Kurdowska, A.K. The Effects of Compounded Bioidentical Transdermal Hormone Therapy on Hemostatic, Inflammatory, Immune Factors; Cardiovascular Biomarkers; Quality-of-Life Measures; and Health Outcomes in Perimenopausal and Postmenopausal Women. Int J Pharm Compd 2013, 17, 74–85. [Google Scholar]
- admin How Bioidentical HRT Impacts Diabetes Risk. Available online: https://womenshormonenetwork.org/2024/02/22/how-bioidentical-hrt-impacts-diabetes-risk/ (accessed on 27 November 2025).
- Cooper, B.C.; Burger, N.Z.; Toth, M.J.; Cushman, M.; Sites, C.K. Insulin Resistance with Hormone Replacement Therapy: Associations with Markers of Inflammation and Adiposity. Am J Obstet Gynecol 2007, 196, 123.e1–7. [Google Scholar] [CrossRef]
- Bornstein, S.R.; Allolio, B.; Arlt, W.; Barthel, A.; Don-Wauchope, A.; Hammer, G.D.; Husebye, E.S.; Merke, D.P.; Murad, M.H.; Stratakis, C.A.; Torpy, D.J. Diagnosis and Treatment of Primary Adrenal Insufficiency: An Endocrine Society Clinical Practice Guideline. J Clin Endocrinol Metab 2016, 101, 364–389. [Google Scholar] [CrossRef]
- Choi, M.H. Clinical and Technical Aspects in Free Cortisol Measurement. Endocrinol Metab (Seoul) 2022, 37, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, T.; Elahi, A.; Wijns, W.; Shahzad, A. Cortisol Detection Methods for Stress Monitoring in Connected Health. Health Sciences Review 2023, 6, 100079. [Google Scholar] [CrossRef]
- Donin, A.S.; Dent, J.E.; Nightingale, C.M.; Sattar, N.; Owen, C.G.; Rudnicka, A.R.; Perkin, M.R.; Stephen, A.M.; Jebb, S.A.; Cook, D.G.; Whincup, P.H. Fruit, Vegetable and Vitamin C Intakes and Plasma Vitamin C: Cross-Sectional Associations with Insulin Resistance and Glycaemia in 9-10 Year-Old Children. Diabet Med 2016, 33, 307–315. [Google Scholar] [CrossRef]
- VA.Gov | Veterans Affairs. Available online: https://www.va.gov/WHOLEHEALTHLIBRARY/tools/hormone-replacement-therapy.asp (accessed on 27 November 2025).
- Weinstock, D. The Menopause Society: Hormone Therapy Statement. Available online: https://www.obgproject.com/2022/11/21/north-american-menopause-society-releases-2017-hormone-therapy-statement/ (accessed on 27 November 2025).
- Committee on the Clinical Utility of Treating Patients with Compounded Bioidentical Hormone Replacement Therapy; Board on Health Sciences Policy; Health and Medicine Division; National Academies of Sciences, Engineering, and Medicine The Clinical Utility of Compounded Bioidentical Hormone Therapy: A Review of Safety, Effectiveness, and Use; National Academies Press, Washington, D.C., 2020. https://doi.org/10.17226/25791. Available online: https://www.nap.edu/catalog/25791.
- Kleis-Olsen, A.S.; Farlov, J.E.; Petersen, E.A.; Schmücker, M.; Flensted-Jensen, M.; Blom, I.; Ingersen, A.; Hansen, M.; Helge, J.W.; Dela, F.; Larsen, S. Metabolic Flexibility in Postmenopausal Women: Hormone Replacement Therapy Is Associated with Higher Mitochondrial Content, Respiratory Capacity, and Lower Total Fat Mass. Acta Physiol (Oxf) 2024, 240, e14117. [Google Scholar] [CrossRef]
- Unlu, Y.; Vinales, K.L.; Hollstein, T.; Chang, D.; Cabeza de Baca, T.; Walter, M.; Krakoff, J.; Piaggi, P. The Association between Gut Hormones and Diet-Induced Metabolic Flexibility in Metabolically Healthy Adults. Obesity (Silver Spring) 2023, 31, 139–149. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



