Submitted:
01 December 2025
Posted:
02 December 2025
You are already at the latest version
Abstract
Background/Objectives: Carbapenem-resistant Klebsiella pneumoniae (CRKP) is a major nosocomial pathogen. Although newer agents have reduced colistin use in high-income countries, this polymyxin remains important in many low- and middle-income settings. Colistin resistance in K. pneumoniae is mainly driven by chromosomal changes in the MgrB–PhoPQ pathway or plasmid-mediated mcr genes. This study aimed to investigate chromosomally mediated colistin resistance in CRKP clinical isolates from a Tunisian ter-tiary hospital. Methods: Between 2010 and 2015, 317 non-duplicate CRKP isolates were collected at Charles Nicolle Hospital, Tunis. Colistin MICs were determined by broth microdilution. Phenotypic tests and PCR characterized carbapenemases, extended-spectrum β-lactamases, AmpC, plasmid-mediated quinolone resistance, mcr and virulence genes. Porins (OmpK35/OmpK36) and the mgrB, phoP and phoQ loci were analyzed by SDS-PAGE and sequencing. Clonal relatedness was assessed by ERIC-PCR and multi-locus sequence typing. Results: Five isolates (1.6%) were colistin-resistant. All were multidrug-resistant, pro-duced OXA-48 and two also carried NDM-1. The isolates belonged to five distinct sequence types, including high-risk clones (ST11, ST101, ST147). No mcr genes were detected. Four isolates carried disruptive mutations in mgrB, and the remaining strain harbored inactivating mutations in both phoP and phoQ with an intact mgrB. Truncating alterations in PhoP/PhoQ and frequent loss or truncation of OmpK35/OmpK36 were observed. Conclusions: In Tunisian CRKP, colistin resistance was mediated by chromosomal alterations, primarily disruption of the MgrB–PhoPQ pathway, in the absence of mcr genes. These mechanisms in both high-risk and emerging sequence types underscore the adaptability of CRKP and the need for surveillance where colistin remains an important therapeutic option.
Keywords:
colistin resistance
; mgrB
; phoP
; phoQ
; carbapenemase
; porins
; multidrug resistance
; virulence factors
1. Introduction
Antimicrobial resistance (AMR) is a global public-health crisis, and carbapenem-resistant Klebsiella pneumoniae (CRKP) epitomizes the problem [1]. Over the past two decades, CRKP has spread throughout hospitals, aided by acquisition of carbapenemase genes and loss or alteration of major porins that impede β-lactam entry. Patients with CRKP infections often have few therapeutic options, so polymyxins such as colistin (polymyxin E) are used as last-resort drugs [2].
Colistin’s bactericidal action involves binding to the negatively charged lipid A component of lipopolysaccharide (LPS), displacing Mg²⁺/Ca²⁺ ions and disrupting the outer membrane [3].
For many years, colistin has been used as a last-resort drug against multidrug-resistant Gram-negative pathogens, including carbapenem-resistant K. pneumoniae (CRKP). More recently, however, several new treatment options have become available for CRKP, such as β-lactam/β-lactamase inhibitor combinations and cefiderocol, which have displaced colistin as a first-line option in many high-income settings [4]. Nevertheless, in many low- and middle-income countries, including North African hospitals, access to these newer agents remains limited and colistin is still widely used in routine practice.
Chromosomal colistin resistance results from mutations that dysregulate lipid A modification pathways. In K. pneumoniae, the PhoPQ and PmrAB two-component systems (TCSs) sense environmental signals and control expression of the pmrCAB and arnBCADTEF (pmrHFIJKLM) operons responsible for adding 4-amino-4-deoxy-L-arabinose (L-Ara4N) or PEtN to lipid A. Disruptive mutations in the mgrB gene (a 47-amino acid negative regulator of PhoPQ) remove feedback inhibition, leading to constitutive activation of PhoPQ and PmrAB and up-regulation of LPS-modifying enzymes. Similarly, activating mutations in phoP/phoQ, pmrA/pmrB,or crrA/crrB can up-regulate these operons; the net effect is lipid A modification and reduced colistin binding. These chromosomal changes are stable and cannot be transferred horizontally, but they can accumulate within clonal lineages [3].
Beyond colistin, CRKP often carries multiple β-lactamases, aminoglycoside-modifying enzymes and plasmid-borne quinolone resistance genes, contributing to multidrug-resistance profiles. The combination of carbapenemase production, porin loss, and colistin-resistance mechanisms produces extensively or pan drug-resistant strains that are difficult to treat.
Tunisia has reported both plasmid-mediated mcr genes [5,6] and chromosomal mgrB mutations [7,8] among clinical K. pneumoniae isolates, highlighting the local public-health importance of this resistance.
This study aimed to investigate the molecular and phenotypic basis of colistin resistance in CRKP clinical isolates recovered from a Tunisian university hospital. We focused on chromosomal mechanisms involving the mgrB–PhoPQ regulatory pathway, while acknowledging that other two-component systems (such as PmrAB and CrrAB) and genome-wide changes may also contribute to colistin resistance but were not explored here due to resource constraints. In addition, because colistin resistance in CRKP usually emerges on a background of broad β-lactam resistance, we also characterized outer membrane porin alterations to better define the multidrug-resistant profile of these isolates.
2. Materials and Methods
2.1. Study Design
Between 2010 and 2015, all clinical CRKP strains were collected from various specimens at the Microbiology Laboratory of Charles Nicolle Hospital in Tunis, Tunisia. Among the collected CRKP isolates, colistin resistance was assessed by determining the minimum inhibitory concentration (MIC) using the broth microdilution method, following the European Committee on Antimicrobial Susceptibility Testing (EUCAST) guidelines [9]. All subsequent phenotypic assays, molecular characterizations, and genetic analyses were performed on the colistin-resistant CRKP strains.
2.2. Species Identification and Antimicrobial Susceptibility Testing
Bacterial species were identified using the API20E system (BioMérieux, Marcy-l’Etoile, France) and confirmed by Matrix-Assisted Laser Desorption Ionization-Time of Flight (MALDI-TOF) (Bruker Daltonics GmbH, Bremen, Germany).
Initially, susceptibility to amoxicillin, amoxicillin–clavulanic acid, cefoxitin, ceftazidime, cefotaxime, cephalotin, cefepime, ertapenem, imipenem, aztreonam, amikacin, tobramycin, netilmicin, nalidixic acid, ofloxacin, ciprofloxacin, fosfomycin, tetracycline, minocycline, and tigecycline was determined by disk diffusion using Mueller-Hinton agar (Bio-Rad), according to EUCAST guidelines [9].
MICs of ertapenem, imipenem, and meropenem were determined using E-test strips (BioMérieux, France), while MICs of amoxicillin–clavulanic acid, cefepime, cefotaxime, ceftazidime, and tigecycline were assessed using the Vitek 2 system (BioMérieux). All results were interpreted according to EUCAST breakpoints and guidelines [9].
2.3. Phenotypic Assays
The modified Hodge test (MHT) was performed according to CLSI guidelines [10] using an ertapenem disk (10 µg). In addition, the Carbapenem Inactivation Method (CIM) was used to detect carbapenemase activity in all isolates, as previously described [11].
Screening for class A carbapenemases was performed using the phenylboronic acid (PBA) method. Inhibition zones obtained with imipenem disks with or without PBA (0.05 M solution) were compared after overnight incubation; an increase of ≥5 mm in the presence of PBA was interpreted as positive for class A carbapenemase production [12].
Screening for class B carbapenemases was performed by comparing inhibition zones around imipenem disks with or without ethylenediaminetetraacetic acid (EDTA, 10 µL of a 0.5 M solution) [13]. An increase in the inhibition zone of >7 mm in the presence of EDTA was considered indicative of metallo-β-lactamase production.
Phenotypic detection of extended-spectrum β-lactamases (ESBLs) was carried out by the double-disk synergy test (DDST) according to CLSI recommendations [10]. AmpC producers were defined as isolates showing a negative ESBL phenotype while being resistant to both cefoxitin and amoxicillin–clavulanic acid.
2.4. Chromosomal Modifications in LPS Biosynthesis
A targeted molecular analysis was conducted to investigate chromosomal alterations in the LPS biosynthesis pathway, focusing on the mgrB, phoP, and phoQ genes. These genes were amplified by polymerase chain reaction (PCR) using gene-specific primers under optimized thermal cycling conditions. The resulting PCR products were purified and subjected to Sanger sequencing.
DNA and deduced amino acid sequences were analyzed using the VECTOR NTI (Invitrogen, California, USA) and compared with the reference genome of the colistin-susceptible K. pneumoniae ATCC 13883 strain (GenBank accession number NZ_JOOW00000000).
2.5. Molecular Detection of Resistance Genes
Multiplex PCR assays were conducted to identify the most commonly encountered carbapenemase genes, including class A (blaKPC and blaGES), class B (blaIMP and blaVIM), and class D (blaOXA-48-like), following previously established protocols [14]. The detection of the blaNDM gene was carried out as described in other studies [15].
In addition, screening was performed for ESBL genes (blaCTX-M, blaTEM, blaSHV, blaVEB, blaGES, and blaPER) and plasmid-mediated AmpC β-lactamase genes (blaCIT, blaMOX, blaFOX, blaEBC, blaDHA, and blaACC) [14]. Furthermore, plasmid-mediated quinolone resistance (PMQR) genes (qnrA, qnrB, qnrC, qnrD, qnrS, qepA, oqxAB, and aac(6’)-Ib-cr) were investigated [16], together with aminoglycoside resistance genes (acc(6’)-Ib, aac(3’)-Ia, aac(3’)-IIa, aac(3’)-IVa, aph(3’)-Ia, aph(3’)-IIa, aph(3’)-VIa, ant(2’’)-Ia) [17].
The presence of plasmid-mediated colistin resistance was assessed through PCR amplification of mcr genes using specific primers for each target [18].
The PCR products were subsequently purified and sequenced, and the resulting DNA sequences were compared with reference nucleotide sequences available in the GenBank database.
2.6. Detection of Virulence-Associated Genes in K. pneumoniae
The virulence profile was analyzed by PCR to check for the presence of nine genes linked to virulence in K. pneumoniae, namely capsular serotype K1 and hypermucoviscosity phenotype (magA), allantoin metabolism (allS), regulator of mucoid phenotype A (rmpA), iron system capture (iroN), capsular serotype K2 and hypermucoviscosity phenotype (cps), adhesion type 3 fimbriae (mrkD), iron transport and phosphotransferase function (kfu), siderophore (entB), and siderophore yersiniabactin (ybtS) [19].
2.7. Molecular Epidemiology and Phylogenetic Analysis
Enterobacterial Repetitive Intergenic Consensus (ERIC) PCR was used as described previously [20] to assess the genetic relatedness of the K. pneumoniae strains. This method targets repetitive DNA sequences within the bacterial genome, providing insight into clonal diversity.
Multilocus sequence typing (MLST) was performed using a previously standardized MLST protocol. The scheme used the following seven housekeeping genes: gapA, infB, mdh, pgi, phoE, rpoB, and tonB [21]. The allelic profile was summarized by assigning a sequence type (ST) via a web database (www.pasteur.fr/recherche/genopole/PF8/mlstKpneumoniae.html).
The phylogenetic relationship among colistin-resistant CRKP isolates was inferred from the concatenated nucleotide sequences of the seven MLST housekeeping genes (gapA, infB, mdh, pgi, phoE, rpoB, and tonB) using the neighbor-joining method implemented in MEGA (version 11.0.13). Prior to tree construction, nucleotide sequences were aligned with the ClustalW algorithm. Evolutionary distances were computed using the Tamura–Nei model, and branch lengths represent the number of base substitutions per site. The tree with the optimal topology was selected automatically, and all positions containing gaps and missing data were eliminated from the final dataset (Figure 1).
2.8. Characterization of Outer Membrane Proteins and Sequencing of ompK35 and ompK36 Genes
Outer membrane protein preparations were obtained by sonication of bacterial cells cultured in Mueller-Hinton broth, followed by selective solubilization of the cytoplasmic components with 2% sodium lauroyl-sarcosynate and ultracentrifugation. The samples were then boiled, loaded onto 11.0% sodium dodecyl sulfate-polyacrylamide gels, and stained with Coomassie blue [22]. The ompK35 and ompK36 genes were amplified, sequenced, and compared with the ompK gene sequences of K. pneumoniae KCTC2242, which produces OmpK35 and lacks OmpK36 (NCBI accession number CP002910), and K. pneumoniae NTUH-K2044, which produces OmpK36 and lacks OmpK35 (NCBI accession number AP006725), using VECTOR NTI (Invitrogen, California, USA) [23].
3. Results
3.1. Prevalence of Colistin Resistance in CRKP Isolates and Antimicrobial Resistance Profile
A total of 317 nonredundant clinical CRKP strains were collected from various wards of Charles Nicolle Hospital between 2010 and 2015. Of these, 5 isolates exhibited resistance to colistin, accounting for 1.6% of the total isolates. The colistin-resistant isolates were primarily recovered from the Intensive Care Unit (ICU) (n=4), with one isolate from the Orthopedics ward. They were recovered from different specimen types: pulmonary (n=2), catheter (n=2), and wound (n=1).
The patients' ages ranged from 24 to 84 years, with 4 of the 5 patients being male. They were admitted either for respiratory distress or polytrauma. Three of these patients were treated during hospitalization with a combination of colistin and imipenem. Besides, combinations of different antibiotic families, namely aminoglycosides (such as amikacin or gentamicin) and fluoroquinolones (mainly ciprofloxacin), in association with imipenem, were used. Despite antibiotic treatment, 1 patient died during hospitalization (Table 1).
All isolates were confirmed as K. pneumoniae by both API20E and MALDI-TOF. Antimicrobial susceptibility testing by disk diffusion revealed resistance to all tested penicillins and cephalosporins, as well as fluoroquinolones and gentamicin; two isolates were non-susceptible to tigecycline based on MICs obtained with the VITEK2 system (Table 2).
E-test results confirmed resistance to ertapenem for all five isolates, with MICs ranging from 2 to >32 mg/L. Two isolates were resistant to imipenem (MICs 2–24 mg/L), and three were resistant to meropenem (MICs 1–24 mg/L). All isolates exhibited colistin resistance, with MICs ranging from 8 to 32 mg/L (Table 2). Furthermore, all isolates tested positive for carbapenemase production using both MHT and CIM. EDTA-based synergy testing was positive for the two blaNDM-1-producing isolates and negative for the remaining strains, whereas no isolate showed a ≥5-mm increase in imipenem inhibition zone in the presence of phenylboronic acid, in keeping with the absence of KPC-type carbapenemases (Table 2).
3.2. Characterization of Antimicrobial Resistance Genes
Molecular fingerprints grouped the 5 isolates into 4 distinct patterns: A (2 strains), B (1 strain), C (1 strain), and D (1 strain). Furthermore, the MLST results revealed the presence of 5 different sequence types (STs): ST11, ST101, ST2502, ST147, and ST4870 (Figure 1).
The phylogenetic analysis of the five colistin-resistant CRKP isolates revealed two main clusters. CRKP1 and CRKP5 grouped closely together, whereas CRKP2 and CRKP4 formed a separate sub-cluster, and CRKP3 appeared more distant from the other isolates (Figure 1).
All strains carried the blaOXA-48 gene, and blaNDM-1 was additionally detected in two strains (CRKP4 and CRKP5). The extended-spectrum β-lactamase (ESBL) blaCTX-M-15 was present in all strains except for CRKP1, which harbored the AmpC β-lactamase blaDHA-1. Other β-lactamase genes identified included blaTEM-1, blaSHV-1, and blaOXA-1, along with plasmid-mediated quinolone resistance genes qnrS1, qnrB1, and aac(6’)-Ib-cr in certain strains. No aminoglycoside resistance genes were detected. No plasmid-mediated colistin resistance genes (mcr) were detected. Three virulence-associated genes, mrkD, entB, and ybtS, were present in all strains, while rmp and kfu were exclusively found in CRKP3 (Figure 1).
3.3. Porin Expression and ompk35/ompk36 Alterations
SDS-PAGE analysis showed that all strains expressed a ~32 kDa protein corresponding to the structural protein OmpA but failed to express a full complement of porins compared to the positive controls K. pneumoniae KCTC2242 (expressing OmpK35 but lacking OmpK36) and K. pneumoniae NTUH-K2044 (expressing OmpK36 but lacking OmpK35) (NCBI accession numbers CP002910 and AP006725, respectively).
Notably, three isolates, CRKP1, CRKP2, and CRKP3 (belonging to pulsotypes B/ST11, C/ST101 and D/ST2502, respectively), lacked the ~39 kDa band corresponding to the major porin OmpK35.
Sequence analysis of ompK35 in CRKP1 revealed the insertion of an ISKpn14 element (780 bp) located 852 nucleotides upstream of the start codon. This IS1-family insertion, positioned within the putative promoter region, is likely responsible for the absence of detectable OmpK35 among outer membrane proteins (Table 3).
In CRKP2 (pulsotype C, ST101) and CRKP3 (pulsotype D, ST2502), sequencing of ompK35 followed by alignment with the reference strain K. pneumoniae KCTC2242 identified multiple alterations. These included point mutations (C156T in CRKP3), deletions [a six-nucleotide deletion (5’-CACCAA-3’) at position 137 in CRKP2 and a single-nucleotide deletion at position 184 in CRKP3], and a single-nucleotide insertion at position 166 in CRKP2. Collectively, these changes introduced premature stop codons, leading to truncated OmpK35 proteins of 314 amino acids in CRKP2 and only 62 amino acids in CRKP3, compared with the full-length 359-amino acid protein in the reference strain (Table 3).
In the CRKP4 strain (pulsotype A, ST147), sequence analysis of the ompK36 gene revealed several point mutations as well as the insertion of a nine-nucleotide sequence [5'-CTGTCTCCT-3'] located at position 550 upstream of the start codon. These genetic alterations result in a protein sequence that diverges from that of the reference strain K. pneumoniae NTUH-K2044. Consistent with these findings, no detectable OmpK36 band was observed in SDS-PAGE profiles(Table 3).
In the CRKP5 strain (pulsotype B, ST4870), two major mutations were identified: the insertion of a guanine at position 65 and a thymine at position 129, both upstream of the start codon. These frameshift events introduce a premature stop codon, leading to the production of a truncated protein of only 22 amino acids instead of the full-length 365-residue OmpK36 (Table 3).
3.4. Chromosome-Mediated Colistin Resistance: Mutations in mgrB, phoP and phoQ
Sequencing of colistin resistance-associated genes (mgrB, phoP, and phoQ) revealed multiple alterations across the five CRKP isolates (Table 4; Figure 2).
Analysis of the mgrB gene revealed distinct alterations among the CRKP isolates. Three isolates (CRKP1, CRKP3, and CRKP5) carried a deletion at nucleotide 132, resulting in frameshift events and altered protein sequences. CRKP4 exhibited multiple point mutations and nucleotide deletions (ΔA42, ΔCC48–49), leading to a premature stop codon at amino acid 19. In contrast, CRKP2 showed no detectable alterations, maintaining an intact mgrB sequence. Phylogenetic analysis supported these observations, with CRKP1, CRKP2, CRKP3, and CRKP5 clustering closely with the reference K. pneumoniae ATCC 13883. CRKP4 was the most divergent isolate, forming a separate branch due to its unique disruptive mutations and premature stop codon.
Comparative analysis of the phoP gene revealed heterogeneous mutational profiles among the five CRKP isolates. CRKP1 carried three nucleotide changes (C29T, C31A, C363T) leading to amino acid substitutions. CRKP2 showed mutations (G57C, T457G, G554C), also resulting in amino acid substitutions without evidence of truncation. CRKP3 harbored three substitutions (A471T, C510A, and C537T) predicted to alter the amino acid sequence. In contrast, CRKP4 exhibited a complex mutation pattern (insertion of nucleotide A at position 7, C32T, C47A, C112A, insertion of nucleotide A at position 143), generating a premature stop codon at amino acid 7, consistent with a truncated non-functional protein. Finally, CRKP5 displayed two substitutions (T17A and C31G) affecting the N-terminal region.
Phylogenetic reconstruction supported these findings: CRKP1 and CRKP5 clustered closely, reflecting their related mutation profiles, while CRKP2 grouped separately but remained relatively close to the reference strain. CRKP3 branched independently, consistent with its distinct substitutions. In contrast, CRKP4 was the most divergent isolate, clearly separated from both the reference and the other strains, in line with its severely truncated protein sequence.
Analysis of the phoQ gene revealed multiple disruptive events across the studied CRKP isolates. CRKP1 carried combined mutations (C219T, InsA486, InsGC498-499), which introduced a premature stop codon at amino acid 175. CRKP2 harbored an extensive set of substitutions and indels (T32G, C35G, A36C, T37A, T38G, ΔC48, InsA58, InsAA65-66, ΔC91) that generated a premature stop codon at amino acid 41, predicting a severely truncated PhoQ protein. CRKP3 exhibited frameshift-inducing mutations (ΔA5, G29C, T130A, ΔC611), leading to an altered protein sequence. The most severe alterations were found in CRKP4, with a cluster of substitutions and deletions (C10T, T11G, G12A, T14C, G15T, G17T, C19T, A20T, ΔT34, ΔGCC105-106-107) that produced a premature stop codon at amino acid 4, consistent with complete loss of function. Finally, CRKP5 carried multiple substitutions and indels (C219T, C512A, C517A, ΔG527, InsT538), resulting in a premature stop codon at amino acid 207.
Phylogenetic reconstruction further highlighted these differences: CRKP1, CRKP2, and CRKP3 formed distinct but relatively close clusters. CRKP5 remained closer to the reference strain but branched separately, consistent with its partial truncation. In contrast, CRKP4 was the most divergent isolate, clearly separated from all other strains and the reference, in agreement with its early stop codon at position 4.
4. Discussion
In this study, five colistin-resistant CRKP isolates were identified among 317 non-duplicate clinical strains collected over a five-year period in a Tunisian hospital.
These colistin-resistant isolates accounted for 1.6% of all CRKP strains and were mostly recovered from intensive care unit (ICU) patients. Although this prevalence is relatively low, it remains clinically concerning. Reported colistin resistance rates vary globally, with higher values in some regions, such as 39.1% in Nigeria, 22.5% in Kenya, and 19.2% in parts of Asia, while many European and North American countries report rates below 5% [24,25,26]. Our findings align more closely with these lower global estimates. However, since our isolates were collected between 2010 and 2015, direct comparison with more recent prevalence data should be interpreted cautiously.
Colistin resistance is primarily driven by the burden of healthcare-associated infections and extensive use of polymyxins, particularly in ICUs [27]. Resistance may also emerge spontaneously through chromosomal mutations or be acquired via horizontal gene transfer, even in the absence of prior colistin exposure [8]. In our study, patients infected with colistin-resistant CRKP ranged in age from 24 to 84 years, were predominantly male, and were admitted for severe conditions such as respiratory distress and polytrauma, clinical scenarios often requiring broad-spectrum antibiotics and invasive procedures, both risk factors for acquiring MDR infections.
Combination therapies incorporating colistin and imipenem, along with aminoglycosides and fluoroquinolones, were commonly used in our cohort, reflecting current clinical practices aimed at enhancing treatment efficacy against CRKP [28]. Although in vitro and observational studies support the synergistic potential of colistin-imipenem combinations [29], therapeutic success remains variable. In our series, only one patient died during hospitalization, suggesting a possible benefit of combination therapy, though clinical outcomes remain unpredictable in critically ill patients.
Colistin-resistant isolates in our study exhibited resistance to multiple other clinically relevant antimicrobials, consistent with many reports documenting extensive multidrug resistance in CRKP worldwide (32, 33). Moreover, literature indicates that colistin MICs tend to be higher in isolates harboring chromosomal mutations affecting regulatory genes compared to isolates with plasmid-mediated resistance mechanisms (mcr genes) [32], highlighting distinct resistance dynamics between genomic and plasmid sources.
All the colistin-resistant CRKP isolates belonged to diverse sequence types (ST11, ST101, ST147, ST2502, and ST4870), reflecting substantial genomic plasticity. All carried the blaOXA-48 carbapenemase gene, while most also harbored ESBL genes such as blaCTX-M-15; additionally, two strains carried blaNDM-1. This resistance gene profile aligns with patterns observed in other studies, where high-risk clones like ST11 and ST147 are key vectors of blaOXA-48 and blaNDM-1, and the regional distribution of these enzymes can vary significantly [33,34,35]. Co-occurrence of OXA-48 with ESBLs, particularly blaCTX-M-15, is also commonly reported [36]. Moreover, the detection of plasmid-mediated quinolone resistance genes (qnrS1, qnrB1, and aac(6′)-Ib-cr) is consistent with previous studies showing their frequent association with ESBL- and carbapenemase-producing K. pneumoniae, especially in NDM- and KPC-positive strains [37]. Importantly, none of our isolates carried plasmid-mediated mcr genes, in line with surveillance data indicating that mcr-driven colistin resistance remains relatively uncommon in K. pneumoniae (39, 40).
Although porin loss primarily contributes to reduced susceptibility to β-lactams rather than to polymyxins, we included OmpK35/OmpK36 characterization to provide a more complete picture of the multidrug-resistant background in which chromosomal colistin resistance arises. We did not investigate other potential contributors to resistance such as efflux pumps or penicillin-binding proteins, which represent an additional limitation of our study.
Carbapenem resistance in K. pneumoniae is frequently associated with loss or functional alteration of the major porins OmpK35 and OmpK36, often caused by insertion sequences or mutations leading to truncated, non-functional proteins, as demonstrated in this study. These findings are consistent with Tunisian reports that highlight porin loss, in combination with carbapenemase production, as a key mechanism of resistance (25, 35). These data underscore the multifactorial nature of CRKP resistance, where both enzymatic degradation and impaired antibiotic influx through porin disruption synergistically reduce treatment efficacy.
The virulence gene profile, dominated by entB, ybtS, and mrkD, with rmpA and kfu detected only in one isolate, suggests that colistin-resistant CRKP typically retain baseline virulence traits but only occasionally acquire hypervirulence markers.
The absence of plasmid-mediated mcr genes in our isolates is consistent with prior studies suggesting that chromosomal mutations are the dominant mechanism of colistin resistance in clinical K. pneumoniae strains [3]. All isolates harbored disruptive mutations in the mgrB-phoPQ regulatory system, including insertions, deletions, or substitutions causing premature stop codons or frameshifts. One isolate (CRKP2) lacked mgrB alterations but presented inactivating mutations in both phoP and phoQ, suggesting that colistin resistance can emerge independently of mgrB disruption. Importantly, although our study did not detect insertion sequences in mgrB, previous studies have reported that IS elements are the most frequent mechanism of mgrB inactivation globally (41, 42). These sequences, such as ISKpn14 or IS5-like elements, often insert into the coding region or promoter of mgrB, thereby silencing its expression and conferring resistance [42]. Our findings support previous reports indicating that while mgrB inactivation is a predominant mechanism, mutations in other components of the PhoPQ system can also confer resistance (43, 44).
In our isolates, the predicted impact of the observed mutations in PhoP and PhoQ ranged from non-synonymous substitutions to very early truncations. For example, several strains carried multiple amino acid changes in PhoP or PhoQ without protein truncation, for which the functional effect on signalling and lipid A modification cannot be inferred from sequence data alone. In contrast, the premature stop codon at position 7 in PhoP in CRKP4 and the extremely early stop codon at position 4 in PhoQ in the same strain almost certainly abolish protein function. Together with the disruptive mutations observed in mgrB, these findings strongly suggest profound dysregulation of the PhoPQ regulatory cascade in some isolates. However, we were not able to perform functional studies such as complementation assays, lipid A profiling or gene expression analysis, so we cannot definitively distinguish between allelic variation with limited impact and mutations that directly drive the colistin-resistant phenotype.
Our study confirms that chromosomal mechanisms remain the primary drivers of colistin resistance in Tunisian CRKP and that these occur in diverse clonal lineages, both globally disseminated and locally emerging. These observations underline the importance of genomic surveillance to monitor the evolution and spread of resistance in critical-care settings.
Limitations
This study has several limitations. First, only five colistin-resistant CRKP isolates were investigated, all recovered from a single tertiary-care hospital, which limits the generalizability of our findings to other Tunisian or regional healthcare settings. Second, the isolates were collected between 2010 and 2015; they therefore reflect an earlier therapeutic context in which colistin was more frequently used against CRKP and may not fully capture the current epidemiology in the era of newer agents such as β-lactam/β-lactamase inhibitor combinations and cefiderocol. Third, our molecular analysis focused on a limited set of chromosomal loci (mgrB, phoP, phoQ), and we did not explore other pathways that may contribute to colistin or carbapenem resistance, such as PmrAB/CrrAB two-component systems, efflux pumps or penicillin-binding proteins. Fourth, we did not have access to whole-genome sequencing for these isolates, which would have provided a more comprehensive view of resistance determinants and mobile genetic elements. Finally, we did not perform functional assays to validate the impact of the identified mutations on lipid A modification or colistin susceptibility; as a result, the causal role of individual amino acid changes in PhoP/PhoQ remains inferred rather than experimentally demonstrated.
5. Conclusions
Our findings show that chromosomally mediated colistin resistance in CRKP isolates from a Tunisian hospital is driven primarily by inactivating mutations in the mgrB–PhoPQ pathway, in the absence of mcr genes. These mutations occur in both globally disseminated and locally emerging sequence types, underscoring the diversity and adaptability of CRKP in clinical environments. Although newer therapeutic options for CRKP have become available in recent years, colistin remains an important component of treatment in many settings, particularly where access to these agents is limited. Continuous surveillance and molecular characterization are therefore essential to detect the emergence of resistance and to inform infection control and treatment strategies.
Author Contributions
Z.H.: Conceptualization, Methodology, Investigation, Formal analysis, Writing—original draft, Writing—review and editing; H.K.: Methodology, Investigation, Writing—original draft, Writing—review and editing; A.O.-S.: Methodology, Investigation, Formal analysis, Writing—review and editing; S.F.: Methodology, Investigation, Writing—original draft, Writing—review and editing; E.M.: Methodology, Investigation, Writing—original draft, Writing—review and editing; L.K.: Writing—review and editing; A.F.: Writing—review and editing; A.R.: Writing—review and editing; L.M.-M.: Funding acquisition, Supervision, Writing—review and editing; I.B.-B.B.: Conceptualization, Supervision, Funding acquisition, Project administration, Writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Higher Education and Scientific Research of Tunisia.
Institutional Review Board Statement
Ethical review and approval were not required for this study because it involved only retrospective analysis of anonymized bacterial isolates, with no identifiable human data, in accordance with institutional and national guidelines.
Informed Consent Statement
Informed consent was not required as the study did not involve identifiable human participants or patient data.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
During the preparation of this study, the author used ChatGPT (OpenAI, accessed November 2025) to assist with English language editing, wording refinement and improvement of clarity in some sections. The authors have reviewed and edited all AI-generated content and take full responsibility for the final version of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AMR | Antimicrobial resistance |
| CIM | Carbapenem Inactivation Method |
| CLSI | Clinical and Laboratory Standards Institute |
| CRKP | Carbapenem-resistant Klebsiella pneumoniae |
| DDST | Double Disk Synergy Test |
| EDTA | Ethylenediaminetetraacetic acid |
| ERIC-PCR | Enterobacterial Repetitive Intergenic Consensus Polymerase Chain Reaction |
| ESBL | Extended-spectrum β-lactamase |
| EUCAST | European Committee on Antimicrobial Susceptibility Testing |
| ICU | Intensive Care Unit |
| LPS | Lipopolysaccharide |
| MALDI-TOF | Matrix-Assisted Laser Desorption/Ionization–Time of Flight |
| MDR | Multidrug-resistant |
| MEGA | Molecular Evolutionary Genetics Analysis |
| MIC | Minimum inhibitory concentration |
| MLST | Multilocus sequence typing |
| NDM | New Delhi Metallo-β-lactamase |
| OMP | Outer membrane protein |
| PBA | Phenylboronic acid |
| PMQR | Plasmid-mediated quinolone resistance |
| SDS-PAGE | Sodium dodecyl sulfate–polyacrylamide gel electrophoresis |
| ST | Sequence type |
| TCS | Two-component system |
References
- Mohammadpour, D.; Memar, M.Y.; Leylabadlo, H.E.; Ghotaslou, A.; Ghotaslou, R. Carbapenem-Resistant Klebsiella Pneumoniae: A Comprehensive Review of Phenotypic and Genotypic Methods for Detection. The Microbe 2025, 6, 100246. [Google Scholar] [CrossRef]
- Li, J.; Shi, Y.; Song, X.; Yin, X.; Liu, H. Mechanisms of Antimicrobial Resistance in Klebsiella: Advances in Detection Methods and Clinical Implications. Infect Drug Resist 2025, 18, 1339–1354. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin Microbiol Rev 2017, 30, 557–596. [Google Scholar] [CrossRef]
- Tayeb, S.; Alharbi, J.; Alattas, B.; Alotaibi, D.; Althibaiti, N.; Alharbi, J.; SafirAldeen, A.; Alqurashi, I.; Wali, S. Promising Future of Novel Beta-Lactam Antibiotics Against Bacterial Resistance. DDDT 2025, Volume 19, 9185–9197. [Google Scholar] [CrossRef]
- Tayh, G.; Nsibi, F.; Abdallah, K.; Abbes, O.; Fliss, I.; Messadi, L. Phenotypic and Molecular Study of Multidrug-Resistant Escherichia Coli Isolates Expressing Diverse Resistance and Virulence Genes from Broilers in Tunisia. Antibiotics 2025, 14, 931. [Google Scholar] [CrossRef]
- Ferjani, S.; Maamar, E.; Ferjani, A.; Meftah, K.; Battikh, H.; Mnif, B.; Hamdoun, M.; Chebbi, Y.; Kanzari, L.; Achour, W.; et al. Tunisian Multicenter Study on the Prevalence of Colistin Resistance in Clinical Isolates of Gram Negative Bacilli: Emergence of Escherichia Coli Harbouring the Mcr-1 Gene. Antibiotics (Basel) 2022, 11, 1390. [Google Scholar] [CrossRef]
- Yengui, M.; Trabelsi, R.; Gdoura, R.; Hamieh, A.; Zerrouki, H.; Rolain, J.-M.; Hadjadj, L. Antibiotic Resistance Profiles of Gram-Negative Bacteria in Southern Tunisia: Focus on ESBL, Carbapenem and Colistin Resistance. Infection, Genetics and Evolution 2025, 133, 105787. [Google Scholar] [CrossRef]
- Jaidane, N.; Bonnin, R.A.; Mansour, W.; Girlich, D.; Creton, E.; Cotellon, G.; Chaouch, C.; Boujaafar, N.; Bouallegue, O.; Naas, T. Genomic Insights into Colistin-Resistant Klebsiella Pneumoniae from a Tunisian Teaching Hospital. Antimicrob Agents Chemother 2018, 62, e01601–17. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). 2014.
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing Twenty-Third Informational Supplement. CLSI Document M100-S23 2013.
- Van Der Zwaluw, K.; De Haan, A.; Pluister, G.N.; Bootsma, H.J.; De Neeling, A.J.; Schouls, L.M. The Carbapenem Inactivation Method (CIM), a Simple and Low-Cost Alternative for the Carba NP Test to Assess Phenotypic Carbapenemase Activity in Gram-Negative Rods. PLoS ONE 2015, 10, e0123690. [Google Scholar] [CrossRef] [PubMed]
- Tamma, P.D.; Simner, P.J. Phenotypic Detection of Carbapenemase-Producing Organisms from Clinical Isolates. J Clin Microbiol 2018, 56, e01140–18. [Google Scholar] [CrossRef]
- Galani, I.; Rekatsina, P.D.; Hatzaki, D.; Plachouras, D.; Souli, M.; Giamarellou, H. Evaluation of Different Laboratory Tests for the Detection of Metallo- -Lactamase Production in Enterobacteriaceae. Journal of Antimicrobial Chemotherapy 2008, 61, 548–553. [Google Scholar] [CrossRef]
- Dallenne, C.; Da Costa, A.; Decré, D.; Favier, C.; Arlet, G. Development of a Set of Multiplex PCR Assays for the Detection of Genes Encoding Important β-Lactamases in Enterobacteriaceae. Journal of Antimicrobial Chemotherapy 2010, 65, 490–495. [Google Scholar] [CrossRef]
- Poirel, L.; Dortet, L.; Bernabeu, S.; Nordmann, P. Genetic Features of BlaNDM-1 -Positive Enterobacteriaceae. Antimicrob Agents Chemother 2011, 55, 5403–5407. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, G.A.; Strahilevitz, J.; Hooper, D.C. Plasmid-Mediated Quinolone Resistance. Microbiol Spectr 2014, 2, 2. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Martínez, M.; Miró, E.; Ortega, A.; Bou, G.; González-López, J.J.; Oliver, A.; Pascual, A.; Cercenado, E.; Oteo, J.; Martínez-Martínez, L.; et al. Molecular Identification of Aminoglycoside-Modifying Enzymes in Clinical Isolates of Escherichia Coli Resistant to Amoxicillin/Clavulanic Acid Isolated in Spain. International Journal of Antimicrobial Agents 2015, 46, 157–163. [Google Scholar] [CrossRef]
- Beyrouthy, R.; Robin, F.; Lessene, A.; Lacombat, I.; Dortet, L.; Naas, T.; Ponties, V.; Bonnet, R. MCR-1 and OXA-48 In Vivo Acquisition in KPC-Producing Escherichia Coli after Colistin Treatment. Antimicrob Agents Chemother 2017, 61, e02540–16. [Google Scholar] [CrossRef]
- Lafeuille, E.; Decré, D.; Mahjoub-Messai, F.; Bidet, P.; Arlet, G.; Bingen, E. OXA-48 Carbapenemase-Producing Klebsiella Pneumoniae Isolated from Libyan Patients. Microbial Drug Resistance 2013, 19, 491–497. [Google Scholar] [CrossRef]
- Rivera, I.G.; Chowdhury, M.A.; Huq, A.; Jacobs, D.; Martins, M.T.; Colwell, R.R. Enterobacterial Repetitive Intergenic Consensus Sequences and the PCR to Generate Fingerprints of Genomic DNAs from Vibrio Cholerae O1, O139, and Non-O1 Strains. Appl Environ Microbiol 1995, 61, 2898–2904. [Google Scholar] [CrossRef]
- Diancourt, L.; Passet, V.; Verhoef, J.; Grimont, P.A.D.; Brisse, S. Multilocus Sequence Typing of Klebsiella Pneumoniae Nosocomial Isolates. J Clin Microbiol 2005, 43, 4178–4182. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Martínez, L.; Conejo, M.C.; Pascual, A.; Hernández-Allés, S.; Ballesta, S.; Ramírez De Arellano-Ramos, E.; Benedí, V.J.; Perea, E.J. Activities of Imipenem and Cephalosporins against Clonally Related Strains of Escherichia Coli Hyperproducing Chromosomal β-Lactamase and Showing Altered Porin Profiles. Antimicrob Agents Chemother 2000, 44, 2534–2536. [Google Scholar] [CrossRef]
- Hamzaoui, Z.; Ocampo-Sosa, A.; Fernandez Martinez, M.; Landolsi, S.; Ferjani, S.; Maamar, E.; Saidani, M.; Slim, A.; Martinez-Martinez, L.; Boutiba-Ben Boubaker, I. Role of Association of OmpK35 and OmpK36 Alteration and blaESBL and/or blaAmpC Genes in Conferring Carbapenem Resistance among Non-Carbapenemase-Producing Klebsiella Pneumoniae. International Journal of Antimicrobial Agents 2018, 52, 898–905. [Google Scholar] [CrossRef] [PubMed]
- Uzairue, L.I.; Rabaan, A.A.; Adewumi, F.A.; Okolie, O.J.; Folorunso, J.B.; Bakhrebah, M.A.; Garout, M.; Alfouzan, W.A.; Halwani, M.A.; Alamri, A.A.; et al. Global Prevalence of Colistin Resistance in Klebsiella Pneumoniae from Bloodstream Infection: A Systematic Review and Meta-Analysis. Pathogens 2022, 11, 1092. [Google Scholar] [CrossRef]
- Mansour, W.; Haenni, M.; Saras, E.; Grami, R.; Mani, Y.; Ben Haj Khalifa, A.; El Atrouss, S.; Kheder, M.; Fekih Hassen, M.; Boujâafar, N.; et al. Outbreak of Colistin-Resistant Carbapenemase-Producing Klebsiella Pneumoniae in Tunisia. Journal of Global Antimicrobial Resistance 2017, 10, 88–94. [Google Scholar] [CrossRef]
- Aris, P.; Robatjazi, S.; Nikkhahi, F.; Amin Marashi, S.M. Molecular Mechanisms and Prevalence of Colistin Resistance of Klebsiella Pneumoniae in the Middle East Region: A Review over the Last 5 Years. Journal of Global Antimicrobial Resistance 2020, 22, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Morand, S.; Rolain, J.-M. Mechanisms of Polymyxin Resistance: Acquired and Intrinsic Resistance in Bacteria. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef]
- Duman, Y.; Tekerekoğlu, M.S.; Kuzucu, Ç.; Yakupoğullari, Y. The Effects of Colistin on Imipenem MICs in OXA-48 Producing Klebsiella Pneumoniae Isolates: An In-Vitro Study. mjima 2021. [CrossRef]
- Katip, W.; Oberdorfer, P.; Kasatpibal, N. Effectiveness and Nephrotoxicity of Loading Dose Colistin–Meropenem versus Loading Dose Colistin–Imipenem in the Treatment of Carbapenem-Resistant Acinetobacter Baumannii Infection. Pharmaceutics 2022, 14, 1266. [Google Scholar] [CrossRef]
- Bogdanovich, T.; Adams-Haduch, J.M.; Tian, G.-B.; Nguyen, M.H.; Kwak, E.J.; Muto, C.A.; Doi, Y. Colistin-Resistant, Klebsiella Pneumoniae Carbapenemase (KPC)-Producing Klebsiella Pneumoniae Belonging to the International Epidemic Clone ST258. Clin Infect Dis 2011, 53, 373–376. [Google Scholar] [CrossRef]
- Jian, Z.; Liu, Y.; Wang, Z.; Liu, P.; Wang, J.; Yan, Q.; Liu, W. Prevalence and Molecular Characteristics of Colistin-Resistant Isolates among Carbapenem-Resistant Klebsiella Pneumoniae in Central South China: A Multicenter Study. Ann Clin Microbiol Antimicrob 2025, 24, 1. [Google Scholar] [CrossRef]
- Riquelme, M.P.; Martinez, R.; Brito, B.; García, P.; Legarraga, P.; Wozniak, A. Chromosome-Mediated Colistin Resistance in Clinical Isolates of Klebsiella Pneumoniae and Escherichia Coli: Mutation Analysis in the Light of Genetic Background. IDR 2023, Volume 16, 6451–6462. [Google Scholar] [CrossRef]
- Dziri, O.; Dziri, R.; Ali El Salabi, A.; Chouchani, C. Carbapenemase Producing Gram-Negative Bacteria in Tunisia: History of Thirteen Years of Challenge. IDR 2020, Volume 13, 4177–4191. [Google Scholar] [CrossRef]
- Hamzaoui, Z.; Ocampo-Sosa, A.; Maamar, E.; Fernandez Martinez, M.; Ferjani, S.; Hammami, S.; Harbaoui, S.; Genel, N.; Arlet, G.; Saidani, M.; et al. An Outbreak of NDM-1-Producing Klebsiella Pneumoniae, Associated with OmpK35 and OmpK36 Porin Loss in Tunisia. Microbial Drug Resistance 2018, 24, 1137–1147. [Google Scholar] [CrossRef]
- Messaoudi, A.; Haenni, M.; Bouallègue, O.; Saras, E.; Chatre, P.; Chaouch, C.; Boujâafar, N.; Mansour, W.; Madec, J.-Y. Dynamics and Molecular Features of OXA-48-like-Producing Klebsiella Pneumoniae Lineages in a Tunisian Hospital. Journal of Global Antimicrobial Resistance 2020, 20, 87–93. [Google Scholar] [CrossRef]
- Carrër, A.; Poirel, L.; Eraksoy, H.; Cagatay, A.A.; Badur, S.; Nordmann, P. Spread of OXA-48-Positive Carbapenem-Resistant Klebsiella Pneumoniae Isolates in Istanbul, Turkey. Antimicrob Agents Chemother 2008, 52, 2950–2954. [Google Scholar] [CrossRef]
- Argente, M.; Miró, E.; Martí, C.; Vilamala, A.; Alonso-Tarrés, C.; Ballester, F.; Calderón, A.; Gallés, C.; Gasós, A.; Mirelis, B.; et al. Molecular Characterization of OXA-48 Carbapenemase-Producing Klebsiella Pneumoniae Strains after a Carbapenem Resistance Increase in Catalonia. Enfermedades Infecciosas y Microbiología Clínica 2019, 37, 82–88. [Google Scholar] [CrossRef]
- Liu, X.; Wu, Y.; Zhu, Y.; Jia, P.; Li, X.; Jia, X.; Yu, W.; Cui, Y.; Yang, R.; Xia, W.; et al. Emergence of Colistin-Resistant Hypervirulent Klebsiella Pneumoniae (CoR-HvKp) in China. Emerging Microbes & Infections 2022, 11, 648–661. [Google Scholar] [CrossRef]
- Zhang, B.; Yu, B.; Zhou, W.; Wang, Y.; Sun, Z.; Wu, X.; Chen, S.; Ni, M.; Hu, Y. Mobile Plasmid Mediated Transition From Colistin-Sensitive to Resistant Phenotype in Klebsiella Pneumoniae. Front. Microbiol. 2021, 12, 619369. [Google Scholar] [CrossRef] [PubMed]
- Cannatelli, A.; Giani, T.; D’Andrea, M.M.; Di Pilato, V.; Arena, F.; Conte, V.; Tryfinopoulou, K.; Vatopoulos, A.; Rossolini, G.M. MgrB Inactivation Is a Common Mechanism of Colistin Resistance in KPC-Producing Klebsiella Pneumoniae of Clinical Origin. Antimicrob Agents Chemother 2014, 58, 5696–5703. [Google Scholar] [CrossRef] [PubMed]
- Fordham, S.M.E.; Mantzouratou, A.; Sheridan, E. Prevalence of Insertion Sequence Elements in Plasmids Relating to mgrB Gene Disruption Causing Colistin Resistance in Klebsiella Pneumoniae. MicrobiologyOpen 2022, 11, e1262. [Google Scholar] [CrossRef]
- Haeili, M.; Javani, A.; Moradi, J.; Jafari, Z.; Feizabadi, M.M.; Babaei, E. MgrB Alterations Mediate Colistin Resistance in Klebsiella Pneumoniae Isolates from Iran. Front. Microbiol. 2017, 8, 2470. [Google Scholar] [CrossRef]
- Gogry, F.A.; Siddiqui, M.T.; Sultan, I.; Haq, Q. Mohd. R. Current Update on Intrinsic and Acquired Colistin Resistance Mechanisms in Bacteria. Front. Med. 2021, 8, 677720. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree of colistin-resistant CRKP isolates constructed using the neighbor-joining method in MEGA (v11.0.13) after multiple sequence alignment with ClustalW. Evolutionary distances were estimated according to the Tamura–Nei model, and branch lengths indicate the number of nucleotide substitutions per site. The dendrogram illustrates the genetic relatedness among the five CRKP isolates (CRKP1–CRKP5) and is annotated with the corresponding ERIC-PCR patterns and sequence types (STs). The heatmap aligned with the tree depicts the distribution of acquired β-lactamase genes (red color), plasmid-mediated quinolone resistance (PMQR) genes (blue color), and virulence-associated genes (green color).
Figure 1.
Phylogenetic tree of colistin-resistant CRKP isolates constructed using the neighbor-joining method in MEGA (v11.0.13) after multiple sequence alignment with ClustalW. Evolutionary distances were estimated according to the Tamura–Nei model, and branch lengths indicate the number of nucleotide substitutions per site. The dendrogram illustrates the genetic relatedness among the five CRKP isolates (CRKP1–CRKP5) and is annotated with the corresponding ERIC-PCR patterns and sequence types (STs). The heatmap aligned with the tree depicts the distribution of acquired β-lactamase genes (red color), plasmid-mediated quinolone resistance (PMQR) genes (blue color), and virulence-associated genes (green color).
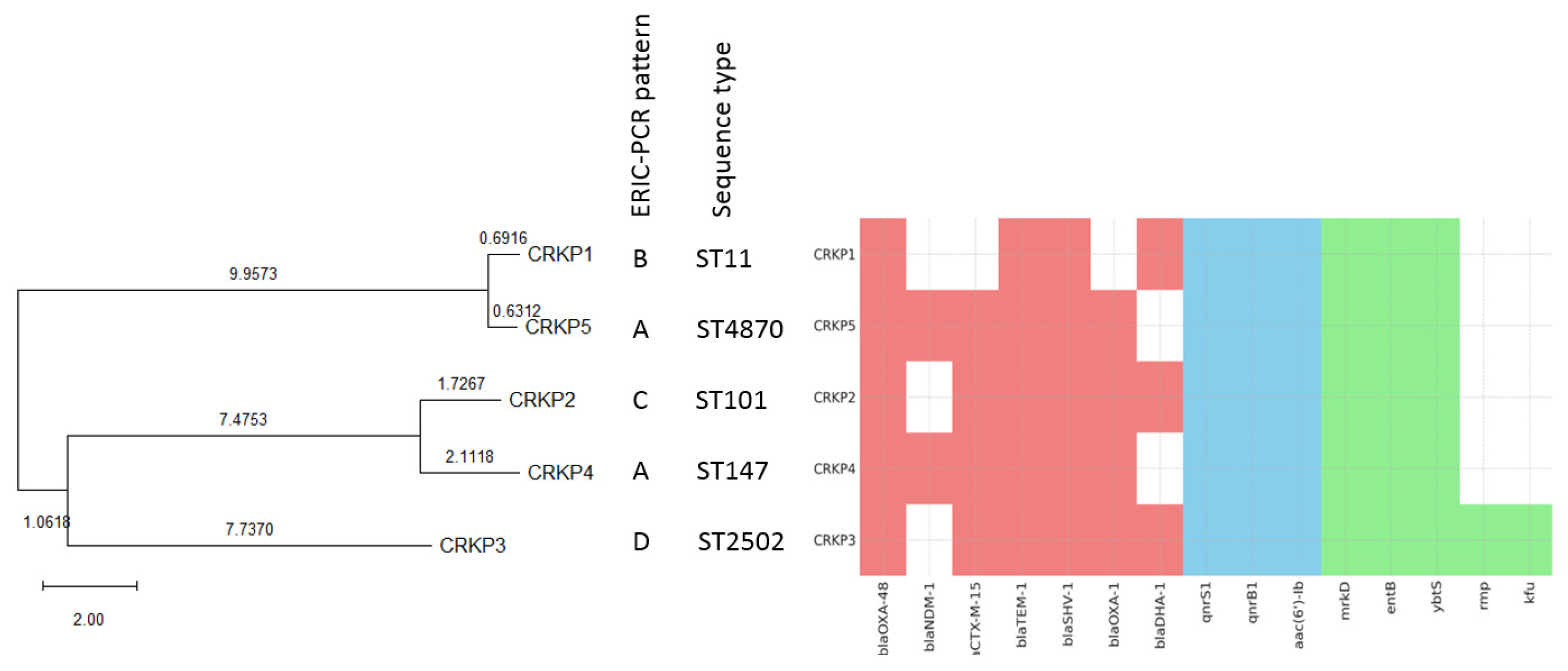
Figure 2.
Multiple sequence alignments of mgrB, phoP, and phoQ genes and their encoded proteins in colistin-resistant CRKP strains. A. Nucleotide sequence alignments of A1 mgrB, A2 phoP, and A3 phoQ genes in colistin-resistant CRKP isolates compared to the K. pneumoniae ATCC 13883 reference strain. B. Corresponding amino acid sequence alignments of B1 MgrB, B2 PhoP, and B3 PhoQ proteins. Conserved regions are indicated by colored boxes, while mismatches, insertions, and deletions highlight genetic variations. Yellow indicates conserved positions across all isolates, blue highlights variable sites (at least one sequence with a difference), and dashes represent insertions/deletions.
Figure 2.
Multiple sequence alignments of mgrB, phoP, and phoQ genes and their encoded proteins in colistin-resistant CRKP strains. A. Nucleotide sequence alignments of A1 mgrB, A2 phoP, and A3 phoQ genes in colistin-resistant CRKP isolates compared to the K. pneumoniae ATCC 13883 reference strain. B. Corresponding amino acid sequence alignments of B1 MgrB, B2 PhoP, and B3 PhoQ proteins. Conserved regions are indicated by colored boxes, while mismatches, insertions, and deletions highlight genetic variations. Yellow indicates conserved positions across all isolates, blue highlights variable sites (at least one sequence with a difference), and dashes represent insertions/deletions.

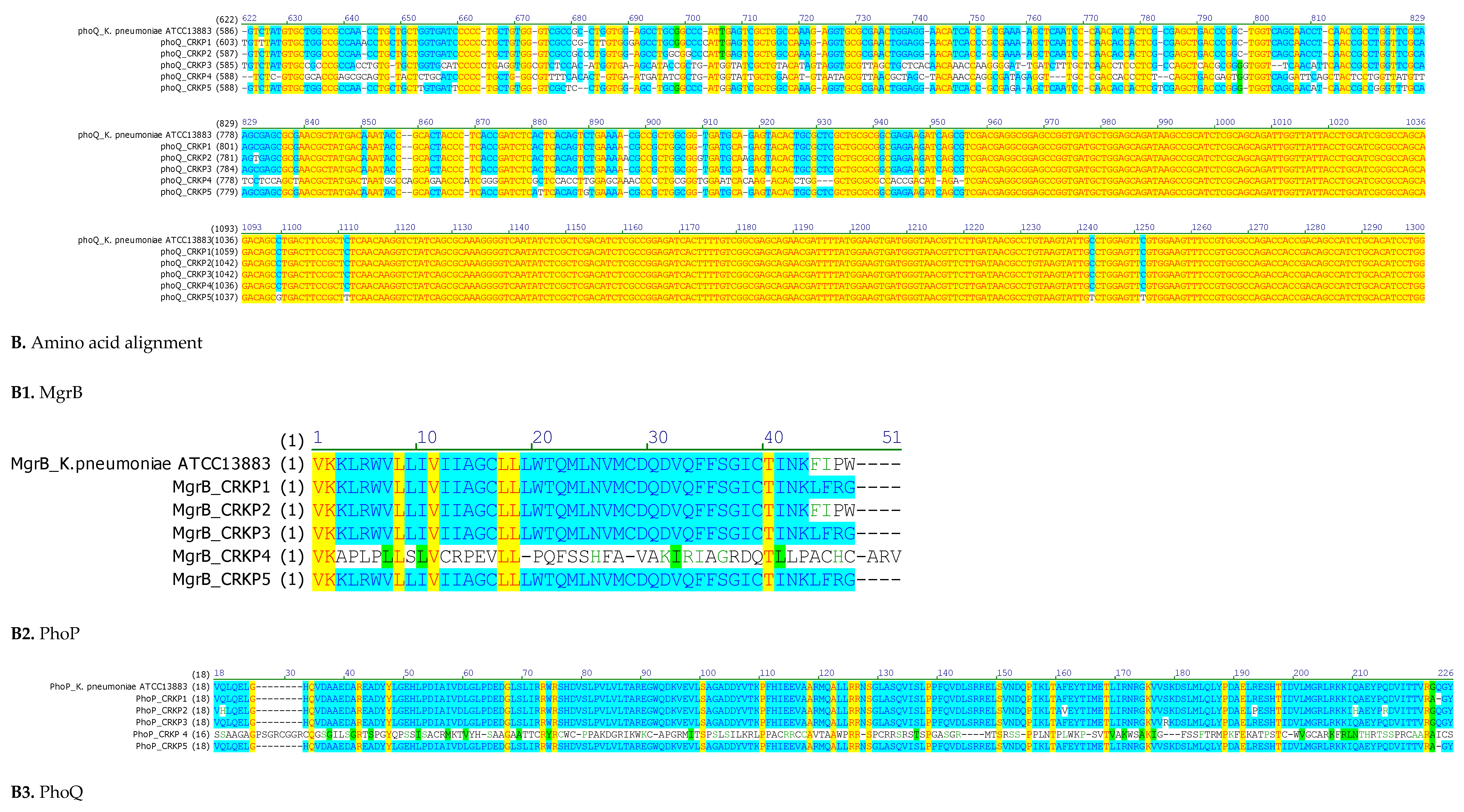
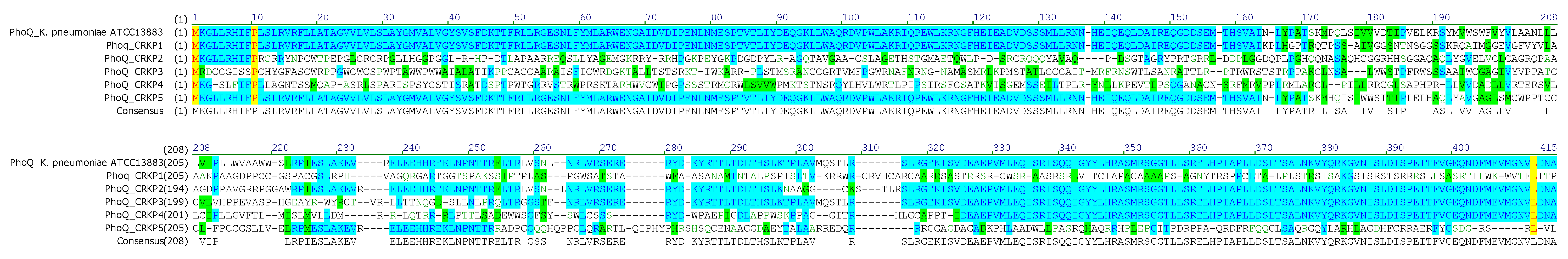

Table 1.
Clinical characteristics, treatment, and outcomes of patients infected with colistin-resistant CRKP.
Table 1.
Clinical characteristics, treatment, and outcomes of patients infected with colistin-resistant CRKP.
| Patient | Strain ID | Ward | Specimen Type | Isolation Date | Age (Years)/Gender | Underlying Disease | Antibiotic Treatment | Outcome | |
| 1 | CRKP 1 | ICU1 | Pulmonary | 2013 | 27/M2 | Respiratory distress | Tigecycline, Gentamicin, Ceftazidime | Improved | |
| 2 | CRKP 2 | Orthopedics | Catheter | 2015 | 84/F3 | Polytraumatism | Amoxicillin+clavulanate | Improved | |
| 3 | CRKP 3 | ICU1 | Catheter | 2015 | 75/M2 | Respiratory distress | Vancomycin, Imipenem, Rifampin, Amikacin, Colistin, Fosfomycin | Died | |
| 4 | CRKP 4 | ICU1 | Pulmonary | 2015 | 32/M2 | Polytraumatism | Gentamicin, Imipenem, Colistin, Vancomycin, Ciprofloxacin | Improved | |
| 5 | CRKP 5 | ICU | Wound | 2015 | 24/M2 | Polytraumatism | Amoxicillin+clavulanate, Gentamicin, Imipenem, Fosfomycin, Colistin, Ciprofloxacin | Improved |
1 ICU, Intensive Care Unit; 2 M, Male; 3 F, Female; A. Nucleotide alignment; A1. mgrB gene.
Table 2.
Antibiotic resistance profiles of colistin-CRKP isolates.
| Strain | Resistance Patterns | Phenotypic Assays | MICs* | ||||||||||||||||||||||||
| MHT1 | CIM2 | EDTA3 | PBA4 | E-Test Strips | VITEK | Broth Microdilution Method | |||||||||||||||||||||
| ETP20 | IMP21 | MEM22 | AMP5 | AMC6 | TZP7 | CXM8 | FOX9 | CTX10 | CAZ11 | FEP13 | AMK14 | 19GM | NAL15 | CIP16 | TGC18 | SXT17 | COL23 | ||||||||||
| CRKP 1 | AMP5, AMC6, TZP7, CXM8, FOX9, CTX, 10 CAZ11, CRO12, FEP13, AMK14, NAL15, 16CIP, SXT1717 | + | + | - | - | >32 | 24 | 24 | ≥32 | ≥32 | ≥128 | ≥64 | ≥64 | ≥64 | ≥64 | ≤1 | 16 | ≤1 | ≥32 | ≥4 | 1 | ≥320 | 32 | ||||
| CRKP 2 | AMP5, AMC6, TZP7, CXM8, FOX9, CTX10, CAZ11, CRO12, FEP13, AMK14, GM19, NAL15, 16CIP, SXT17 | + | + | - | - | >32 | 4 | 12 | ≥32 | ≥32 | ≥128 | ≥64 | ≥64 | ≥64 | ≥64 | ≥64 | 16 | ≥16 | ≥32 | ≥4 | 2 | 40 | 16 | ||||
| CRKP 3 | AMP5, AMC6, TZP7, CXM8, FOX9, CTX10, CAZ11, CRO12, FEP13, AMK14, GM19, NAL15, 16CIP, TGC18, SXT17 | + | + | - | - | >32 | 4 | 6 | ≥32 | ≥32 | ≥128 | ≥64 | ≥64 | ≥64 | 16 | ≥64 | ≥64 | ≥16 | ≥32 | ≥4 | 2 | ≥320 | 8 | ||||
| CRKP 4 | AMP5, AMC6, TZP7, CXM8, FOX9, CTX10, CAZ11, CRO12, FEP13, GM, NAL15, 16CIP, TGC18, SXT17 | + | + | + | - | 32 | 12 | 16 | ≥32 | ≥32 | ≥128 | ≥64 | ≥64 | ≥64 | ≥64 | 16 | 8 | ≥16 | ≥32 | ≥4 | 4 | ≥320 | 8 | ||||
| CRKP 5 | AMP5, AMC6, TZP7, CXM8, FOX9, CTX10, CAZ11, CRO12, FEP13, GM, NAL15, 16CIP, TGC18, SXT17 | + | + | + | - | 2 | 2 | 1 | ≥32 | ≥32 | ≥128 | ≥64 | ≥64 | ≥64 | ≥64 | ≥64 | 8 | ≥16 | ≥32 | ≥4 | 4 | ≥320 | 8 | ||||
1 MHT, Modified Hodge Test ; 2 CIM, Carbapenem Inactivation Method ; 3 EDTA, Ethylene Diamine Tetraacetic Acid ; 4 PBA, Phenyl Boronic Acid ; 5 AMP, Ampicillin ; 6 AMC, Amoxicillin-clavulanic acid ; 7 TZP, piperacillin–tazobactam ; 8 CXM, cefuroxim ; 9 FOX, cefoxitin ; 10 CTX, cefotaxime ; 11 CAZ, ceftazidime ; 12 CRO, ceftriaxone ; 13 FEP, cefepime ; 14 AMK, Amikacin ; 15 NAL, nalidixic acid ; 16 CIP, ciprofloxacin ; 17 SXT, sulfamethoxazole/trimethoprim ; 18 TGC, tigecycline ; 19 GM : Gentamicin ; 20 ETP, Ertapenem ; 21 IMP, Imipenem ; 22 MEM, Meropenem ; 23 COL, Colistin ; *MICs were performed according to EUCAST guidelines (mg/L).
Table 3.
Porin alterations, nucleotide and protein mutations in ompK35 and ompK36 among colistin-resistant CRKP.
Table 3.
Porin alterations, nucleotide and protein mutations in ompK35 and ompK36 among colistin-resistant CRKP.
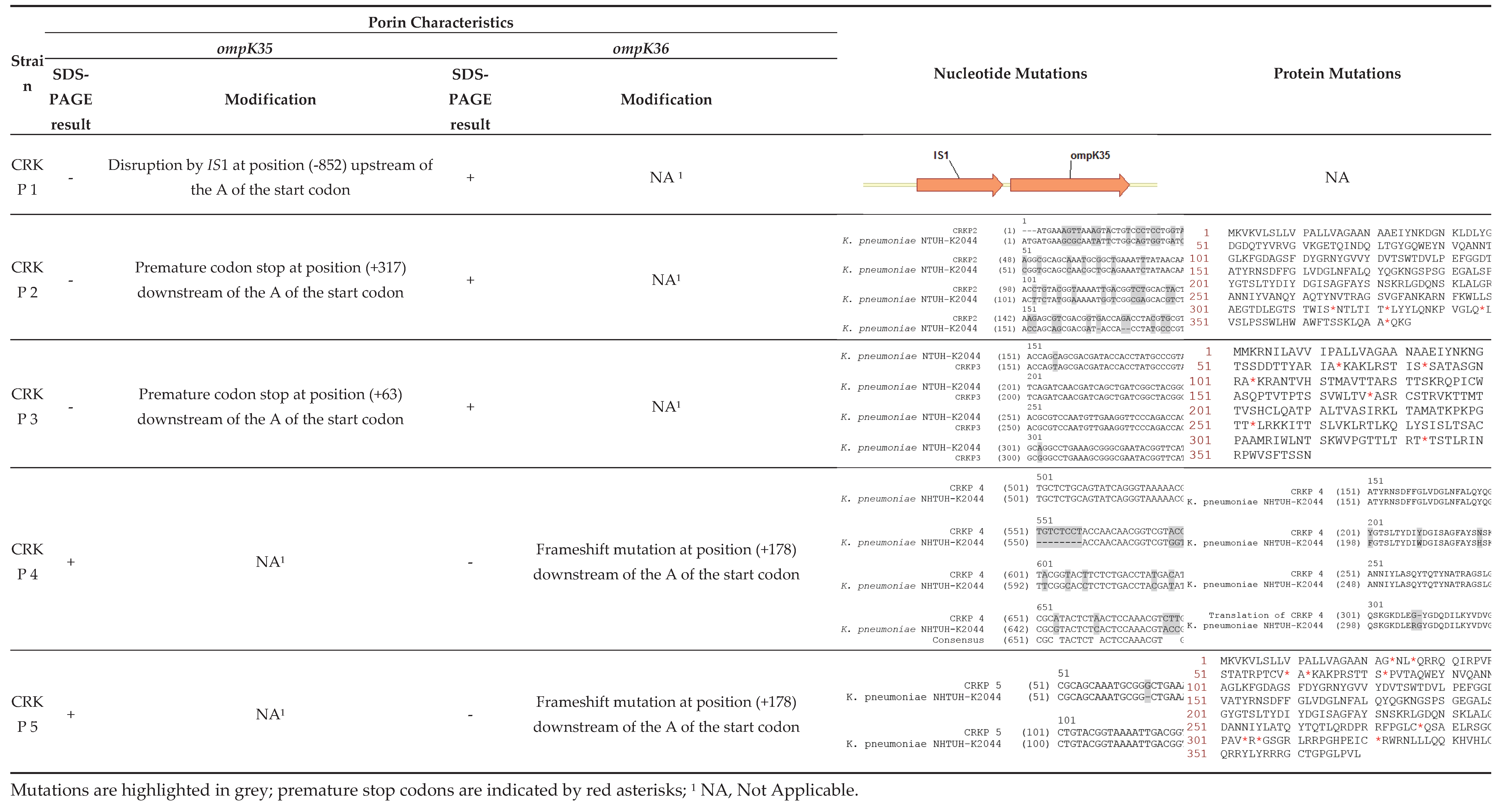
Table 4.
Chromosome-mediated colistin resistance: mutations in mgrB, phoP and phoQ.
| Strain | mgrB Mutations | Predicted Effect | phoP Mutations | Predicted Effect | phoQ Mutations | Predicted Effect |
| CRKP1 | ΔT132 | Frameshift →Altered protein sequence | C29T; C31A; C363T | AA substitutions | C219T; InsA486; InsGC, 498-499 | Premature stop codon at AA 175 |
| CRKP2 | None | Intact MgrB | G57C; T457G; G554C | AA substitutions | T32G; C35G; A36C; T37A; T38G; ΔC48; InsA58; InsAA65-66; ΔC91 |
Premature stop codon at AA 41 |
| CRKP3 | ΔT132 | Frameshift → altered protein sequence | A471T; C510A; C537T | AA substitutions | ΔA5; G29C; T130A; ΔC611 | Frameshift → altered protein sequence |
| CRKP4 | Multiple point mutations; ΔA42, ΔCC48-49 |
Premature stop at AA19 | InsA7; C32T; C47A; C112A; InsA143 | Premature stop codon at AA7 | C10T; T11G; G12A; T14C; G15T; G17T; C19T; A20T ; ΔT34 ; ΔGCC105-106-107 | Premature stop codon at AA 4 |
| CRKP5 | ΔT132 | Frameshift → altered protein sequence | T17A; C31G | AA substitutions | C219T; C512A; C517A; ΔG527; InsT538 | Premature stop codon at AA 207 |
Δ, nucleotide deletion; Ins, nucleotide insertion; AA, amino acid.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



