Submitted:
27 November 2025
Posted:
27 November 2025
You are already at the latest version
Abstract
Nutritional immunity is a major facet of host defense, wherein the host immune system strategically limits pathogen access to critical nutrients, including iron, zinc, vitamins, lipids, and amino acids, to repress microbial proliferation and virulence. This review provides a comprehensive synthesis of the molecular mechanisms that power nutrient immunity, including metal homeostasis, transporter modulation, hormonal regulation, and direct antimicrobial actions. We examine nutrient-specific strategies employed by the host, from iron withholding mechanisms to vitamin deprivation and copper-mediated toxicity. We also explore how diverse pathogens, including extracellular, intracellular, and eukaryotic, adapt to these hostile nutritional landscapes through siderophore diversification, regulatory integration, and metabolic rewiring. Comparative genomic analyses reveal convergent evolution in nutrient acquisition systems, illuminating the dynamic arms race between host restriction and microbial evasion. Further, we discuss the translational potential of nutritional immunity, cutting across nutrient-based therapies, host-directed interventions, and emerging diagnostic biomarkers. Finally, we suggest future directions that synergize nutritional immunity with microbiome ecology, global malnutrition, and personalized medicine. By elucidating the interconnection between metabolism and immunity, this review highlights the therapeutic promise of starving the pathogen to save the host.
Keywords:
nutritional immunity
; host–pathogen interactions
; metal sequestration
; immunometabolism
; pathogen adaptation
; host-directed therapeutics
; antimicrobial resistance
1. Introduction
Infectious diseases constitute a major public health concern across the globe [1]. The situation is further aggravated by the increasing emergence of antimicrobial resistance and inadequate development of new antibiotics [2,3]. At the heart of every infection lies a fierce competition for nutrients between the host and an invading pathogen [4]. Pathogens, including bacteria, fungi, and protozoa, need access to essential elements such as zinc, iron, manganese, amino acids, and vitamins to uphold structural integrity, power replication, and initiate their virulence programs [5,6]. In response, the host deploys a repertoire of defense mechanisms collectively known as nutritional immunity, which are strategies that limit microbial access to these resources while maintaining host metabolic homeostasis [7]. Interestingly, this nutrient tug-of-war is not just a passive consequence of infection but rather a dynamic and evolutionarily conserved interface between metabolism and immunity. For example, iron sequestration via transferrin, lactoferrin, and hepcidin is tightly controlled to restrict microbial proliferation while minimizing oxidative stress [8]. Similarly, calprotectin-mediated zinc and manganese chelation hampers bacterial enzymatic function and growth [9]. These host strategies are opposed by microbial adaptations, including siderophore production, metal transporter upregulation, and metabolic rewiring, underlining the complexity of host-pathogen nutrient interactions [10]. Furthermore, nutritional immunity is increasingly recognized as a key pillar of innate and adaptive defense. It borders cytokine signaling, hormonal regulation, and cellular immunity, determining the outcome of infections across different anatomical niches [5,11]. For instance, macrophages regulate phagosomal nutrient availability through NRAMP1-mediated metal efflux and autophagy-linked amino acid deprivation [12,13]. In parallel, systemic responses such as hypoferremia and altered vitamin metabolism reveal the host’s attempt to starve pathogens while maintaining immune function [14]. Notably, the concept of nutritional immunity is not only true within the context of bacterial infections. Eukaryotic pathogens such as Plasmodium falciparum, Leishmania spp., and Candida albicans also exhibit complex nutrient acquisition systems that allow immune evasion and persistence [6,15]. Importantly, the interplay between nutrient availability and pathogen virulence is further modulated by host nutritional status, microbiome composition, and environmental factors, making nutritional immunity a nexus of immunology, metabolism, and ecology [16,17]. This review aims to provide a comprehensive synthesis of the mechanisms, pathogen-specific strategies, and therapeutic implications of nutritional immunity. We begin by detailing the molecular and cellular processes through which the host limits pathogen access to critical nutrients. We then explore nutrient-specific strategies, such as iron sequestration, copper toxicity, and amino acid deprivation, and examine how diverse pathogens adapt to these pressures. We offer comparative genomic insights that reveal evolutionary patterns in nutrient acquisition and translational perspectives that highlight emerging therapies and diagnostics rooted in nutritional immunity. We conclude by outlining future directions that would interlink nutritional immunity with microbiome science, personalized medicine, and global health.
2. Mechanisms of Nutritional Immunity
The concept of nutritional immunity encompasses a suite of host strategies designed to restrict microbial access to essential nutrients within the host milieu, thereby impairing pathogen proliferation and virulence. These mechanisms, illustrated in Figure 1 and described extensively in Table 1, are multifaceted and involve metal sequestration, metal toxicity, nutrient competition, metabolic reprogramming, hormonal regulation, siderophore interference, and direct antimicrobial actions. Together, they constitute a dynamic and context-dependent defense system that integrates innate and adaptive immune responses. This section presents the various strategies employed by the host to exert nutritional immunity.
2.1. Metal Homeostasis and Disruption
Metal homeostasis is a major cornerstone in nutritional immunity, particularly in the regulation of iron, zinc, manganese, and copper. A diagrammatic snapshot of various mechanisms employed by the host to sequester critical metal nutrients from pathogens, as well as pathogens’ counterstrategies, is presented in Figure 2. Iron, an essential cofactor for DNA synthesis and microbial respiration, is tightly controlled by host proteins such as transferrin, ferritin, lactoferrin, hemopexin, and haptoglobin [7]. During infection, the liver amplifies hepcidin, a peptide hormone that degrades ferroportin, the sole iron exporter, thereby lowering serum iron levels and restricting microbial access [11]. Similarly, zinc and manganese are sequestered by calprotectin, a neutrophil-derived protein that chelates these metals at sites of inflammation [33]. This sequestration disrupts microbial enzymatic functions, especially those involved in DNA replication and oxidative stress resistance. Pathogens respond by upregulating high-affinity transporters such as ZnuABC and MntH, highlighting the evolutionary arms race between host sequestration and microbial acquisition [17]. On the other hand, the host weaponizes transition metals like copper and zinc as toxic antimicrobial agents. For example, macrophages accumulate copper and zinc in phagosomes via ATP7A transporters and zinc transporters (ZIP8, ZnT family), respectively [34,35]. These toxic metal levels overwhelm microbial detoxification systems and interfere with critical metabolic processes. For instance, the correct metalation of bacterial enzyme cofactors is disrupted when excess copper and zinc replace iron and manganese. Also, excess metals in phagosomes catalyze the generation of reactive oxygen species and impair microbial iron-sulfur clusters [36]. Pathogens negate this by expressing copper efflux pumps and chaperones, such as CopA and CueO, which attenuate toxicity and preserve intracellular homeostasis [21]. The implementation of metal homeostasis during infection portrays a finely tuned balance between host defense and systemic physiology. Dysregulation can lead to immunopathology, as observed in anemia of inflammation or metal overload syndromes [37,38]. Thus, an understanding of the molecular mechanisms of metal sequestration and toxicity is critical for the development of targeted therapies that promote nutritional immunity without impairing host health.
2.2. Metabolic and Hormonal Regulation
Metabolic and hormonal regulation plays a key role in nutritional immunity by regulating nutrient availability and immune cell function [4,39]. For example, immune cells undergo metabolic reprogramming during infection by shifting from oxidative phosphorylation to glycolysis to support rapid proliferation and cytokine production [40]. This shift elevates host immune cells’ consumption of glucose and glutamine, depriving pathogens of major carbon sources and creating a competitive metabolic environment. Hormonal signals further control nutrient flux. For instance, hepcidin, induced by interleukin-6 (IL-6) during inflammation, diminishes the export of iron and facilitates intracellular sequestration [41]. Similarly, indoleamine 2,3-dioxygenase (IDO), which is switched on by interferon-γ, catabolizes tryptophan into kynurenine, lowering its availability to intracellular pathogens such as Toxoplasma gondii and Chlamydia trachomatis [42]. These hormonal pathways interconnect immune signaling with nutrient control, boosting host defense. Metabolic regulation also influences the differentiation and function of immune cells. For example, the depletion of arginine by arginase activity represses T cell proliferation and facilitates the development of regulatory T cells, thereby controlling the immune response to chronic infections [43]. Contrarily, amino acid sufficiency promotes cytokine production and effector functions, which underlines the dual role of nutrients as both substrates and stimuli [44]. The interplay between metabolism, hormones, and immunity highlights the complexity of nutritional immunity. The therapeutic modulation of these pathways, through metabolic inhibitors, hormone analogs, or dietary interventions, could present promising avenues for boosting host defense. However, such strategies must be carefully calibrated to avert inadvertent immunosuppression or metabolic dysregulation.
2.3. Resource Competition
Resource competition is a fundamental mechanism of nutritional immunity, wherein host cells alongside commensal microbes compete with pathogens for critical nutrients. This competition occurs at multiple levels, including direct substrate limitation, niche exclusion, and metabolic interference. For instance, activated macrophages consume large amounts of glucose and amino acids, reducing their availability to intracellular pathogens [26]. This metabolic competition is further elevated by the Warburg effect, which amplifies glycolytic flux in immune cells [45]. Additionally, commensal microbiota play a pivotal role in resource competition. For example, beneficial microbes in the gut withhold iron and synthesize siderophores, which inhibit pathogen growth, effectively increasing the host’s nutritional immunity [16]. These microbes also compete for amino acids and vitamins, which lowers the nutrient pool available to invading pathogens. Notably, dysbiosis or microbial imbalance can impair this competitive landscape and elevate susceptibility to infection [46]. Pathogens retaliate to resource competition by developing flexible metabolic pathways and high-affinity transport systems. For instance, Listeria monocytogenes can turn to host-derived glycerol and acetate during intracellular growth, bypassing glucose restriction [47]. Likewise, Salmonella enterica expresses multiple carbon and amino acid transporters to adapt to nutrient-limited vacuoles [48]. These adaptations portray the selective pressure imposed by host and microbial competitors. Understanding resource competition dynamics is necessary for designing interventions that fortify host and microbiome defenses. Dietary modulation, probiotic therapies, and prebiotic supplementation can improve competitive exclusion and decrease pathogen colonization [49,50]. Moreover, targeting pathogen-specific nutrient acquisition pathways may tip the balance in favor of host immunity, presenting a novel approach to infection control.
2.4. Host Regulation of Nutrient Transporters and Storage
Nutrient transporters and storage proteins are highly regulated by the host to control systemic and local nutrient availability during infection. For instance, iron transport is controlled by ferroportin, the only known iron exporter, and its regulation by hepcidin ensures that iron is kept within hepatocytes and macrophages during inflammation [11]. Also, transferrin and lactoferrin bind extracellular iron, while ferritin stores intracellular iron, collectively diminishing the levels of free iron and restricting microbial access [7]. Similarly, the Zrt-/Irt-like Proteins (ZIP) and Zinc Transporters (ZnT) families of transporters regulate zinc and manganese transport, thereby controlling their cellular influx and efflux [51]. During infection, immune cells upregulate metallothioneins to neutralize excess zinc and avert toxicity, while downregulating transporters to limit pathogen access [52]. These changes are directed by metal-responsive transcription factors like MTF-1 and NRF2, which integrate oxidative stress and nutrient signals [53]. Furthermore, amino acid transporters are also modulated during infection. For example, SLC1A5 and SLC7A5, which mediate leucine and glutamine uptake, are amplified in activated T cells to support proliferation and cytokine production [54]. By contrast, downregulation of tryptophan transporters and activation of IDO restricts pathogen access and modulates immune responses [55]. These changes illustrate the bipartite role of transporters in acquiring nutrients and regulating immunity. Storage proteins such as albumin, ferritin, and transferrin serve as reservoirs and buffers, maintaining homeostasis while limiting pathogen access [56,57,58]. Their expression and localization are dynamically controlled during infection, influenced by cytokines, hormones, and cellular stress. Therapeutic modulation of transporter and storage protein activity, through small molecules, biologics, or gene editing, offers a promising strategy for strengthening nutritional immunity and infection control.
2.5. Direct Antimicrobial Actions
Nutritional immunity also involves direct antimicrobial actions mediated by nutrient-binding proteins and metabolic byproducts. For example, calprotectin not only chelates zinc and manganese but also punctures microbial membranes and blocks enzymatic activity, exerting bacteriostatic effects [59]. Also, lipocalin-2 binds bacterial siderophores, hampering iron uptake and triggering oxidative stress in pathogens [29]. These proteins act as both nutrient scavengers and antimicrobial effectors. Oxidative radicals, such as reactive oxygen species (ROS) and nitrogen oxygen species (RNS), generated during immune activation, further facilitate nutritional immunity. For instance, these molecules oxidize and degrade microbial cofactors, such as iron-sulfur clusters and thiol groups, compromising metabolic function and replication [60]. Also, copper and zinc, concentrated in phagosomes, catalyze ROS production, which impairs microbial redox balance, enhancing killing efficiency [21]. Metabolic byproducts also play a role in enhancing nutritional immunity. As an example, kynurenine, produced from tryptophan catabolism via IDO, has immunomodulatory and antimicrobial properties. It suppresses T cell proliferation and triggers apoptosis in infected cells, facilitating pathogen clearance [42]. Similarly, lactate and succinate, produced during glycolysis and the TCA cycle, modulate immune responses and suppress pathogen growth in specific contexts [61]. These direct antimicrobial actions complement other nutritional immunity strategies, creating an unwelcome environment for pathogens. Essentially, we see the integration of metabolic, immunological, and biochemical defenses in nutritional immunity. Future research should explore the therapeutic potential of these antimicrobial molecules, including their use as adjuvants, antimicrobial agents, or immunomodulators in treating infectious diseases.
3. Nutrient-Specific Strategies in Nutritional Immunity
Nutritional immunity is orchestrated through the targeted restriction of specific nutrients that are critical for microbial survival and virulence. Each nutrient plays a unique role in microbial physiology, and the host has developed specialized mechanisms to restrict their availability during infection. This section examines the molecular and immunological strategies engaged by the host to restrict iron, zinc, manganese, copper, magnesium, sulfur, nitrogen, vitamins, carbon sources, and amino acids, and how pathogens adapt to these pressures.
3.1. Iron Limitation: A Key Mechanism of Nutritional Immunity
Iron is indispensable for almost all living organisms owing to its role in enzymatic reactions, oxygen transport, DNA synthesis, and cellular respiration [62]. During infection, the host withholds iron using a pool of proteins, including ferritin, lactoferrin, transferrin, hemopexin, and haptoglobin, which bind iron and iron-containing complexes in extracellular and intracellular compartments [7]. The liver-derived peptide hormone, hepcidin, plays a pivotal role by degrading ferroportin, the only known iron exporter, thereby lowering serum iron levels and trapping iron within macrophages [11]. This strategy of iron sequestration is particularly effective against extracellular pathogens that depend on free iron for replication [63]. However, many bacteria have developed countermeasures, including the production of siderophores, which are small, high-affinity iron-chelating molecules [64]. Pathogens like Escherichia coli and Salmonella enterica produce covert siderophores like salmochelin [65,66]. This salmochelin evades host proteins like lipocalin-2, which normally neutralize traditional siderophores (Smith, 2007). These adaptations illustrate the evolutionary arms race between host iron sequestration and microbial iron acquisition. Meanwhile, iron limitation also influences host immunity beyond direct antimicrobial effects. For instance, iron deprivation can modulate macrophage polarization, increase oxidative burst, and regulate cytokine production [68]. Moreover, iron status affects the outcome of infections such as tuberculosis and malaria, where both iron deficiency and overload can aggravate disease [69]. Iron regulation, therefore, is a double-edged sword that must be well balanced to optimize host defense. Notably, iron chelation has been therapeutically explored as an adjunct to antimicrobial therapy [70,71,72]. For example, agents like deferiprone and deferoxamine can diminish iron availability to pathogens, although they must be used with care to avert the impairment of host hematopoiesis and immune function [73]. A complete understanding of the nuances of iron metabolism during infection is necessary for developing safe and effective interventions that leverage nutritional immunity.
3.2. Zinc and Manganese: Essential Metals in Host-Pathogen Interactions
Zinc and manganese are critical cofactors for several microbial enzymes, including those involved in metabolic regulation, oxidative stress resistance, and DNA replication [9]. The host limits pathogen access to these metals through calprotectin, a neutrophil-derived protein that chelates zinc and manganese at inflammation sites [33]. This sequestration disrupts microbial enzymatic function and facilitates pathogen clearance [9]. In response, pathogens counteract this strategy by upregulating high-affinity metal transporters. For example, the ZnuABC system in Gram-negative bacteria and the MntABC system in Gram-positive bacteria promote zinc and manganese uptake under limiting conditions [17]. These transporters are often controlled by metal-responsive transcription factors such as Zur and MntR, which direct gene expression based on intracellular metal concentrations [74]. Furthermore, zinc limitation also affects host immunity. For instance, zinc is critical for cytokine production, T cell development, and barrier integrity [75]. During infection, zinc redistribution promotes the activation of immune cells while restricting microbial access [52]. However, excessive zinc sequestration can compromise host function [76], which underlines the need for careful regulation of zinc homeostasis. Manganese plays a special role in microbial resistance to oxidative stress by serving as a cofactor for superoxide dismutase and other antioxidant enzymes [77]. By reducing manganese availability, the host leaves the pathogens prone to reactive oxygen species, facilitating microbial killing [68]. This twofold role of manganese in microbial metabolism and immune defense underpins its relevance in nutritional immunity.
3.3. Copper Toxicity and Homeostasis
Contrary to zinc and iron, the host employs copper as a toxic antimicrobial agent [78]. For instance, under hypoxic conditions, which commonly occur during infections, macrophages have been shown to ramp up copper concentrations in phagosomes via ATP7A transporters, where copper catalyzes the production of reactive oxygen species and disrupts microbial iron-sulfur clusters [36]. This targeted copper delivery depicts a direct antimicrobial strategy rooted within nutritional immunity. Copper toxicity impairs multiple microbial processes, including DNA repair, respiration, and redox balance [79]. Pathogens respond by expressing chaperones, copper efflux pumps like CopA, and detoxifying enzymes that alleviate copper-induced damage [21]. Mycobacterium tuberculosis, for example, activates the RicR regulon to handle copper stress during intracellular infection [80]. Host copper homeostasis is highly regulated to avert collateral damage. Excess copper can be toxic to host cells, which necessitates the use of metallothioneins and copper-binding proteins to buffer intracellular levels [81]. Striking a balance between antimicrobial efficacy and host toxicity is an important consideration in copper-based immunity. Therapeutically, copper-enhancing agents and copper-mimetic compounds are being explored as adjuncts to antimicrobial therapy [82,83]. These strategies aim to exploit pathogen vulnerabilities to copper stress while maintaining host homeostasis. Further research into copper trafficking and regulation during infection may produce novel approaches to utilizing this potent antimicrobial metal.
3.4. Magnesium Limitation and Membrane Integrity
Magnesium is necessary for stabilizing cellular membranes, nucleic acids, and ribosomes in both host and microbes [84]. During infection, the host restricts magnesium availability within phagosomes, disrupting bacterial membrane integrity and signaling pathways [85]. This strategy is precisely potent against intracellular pathogens that depend on magnesium for survival in acidic vacuolar environments. Salmonella enterica senses magnesium limitation through the PhoP/PhoQ two-component system, which activates genes that reinforce resistance to acid stress and antimicrobial peptides [86]. This system also controls virulence factors, connecting magnesium sensing to pathogenicity. S. enterica mutants deficient in PhoP/PhoQ signaling exhibit reduced survival in macrophages, highlighting the significance of magnesium adaptation [87]. Magnesium starvation from pathogens makes them vulnerable to host defense mechanisms [4]. It impairs ATP synthesis, disrupts membrane potential, and heightens susceptibility to antibiotics and immune effectors. The host takes advantage of these vulnerabilities by modulating magnesium transport and buffering systems during infection [4]. Despite its significance, magnesium restriction is not as well-characterized as iron or zinc limitation. Future research should probe the molecular mechanisms of magnesium sequestration, its impact on immune cell function, and its potential as a therapeutic target. A deeper understanding of magnesium dynamics may present new strategies for controlling intracellular infections.
3.5. Sulfur and Nitrogen Metabolism
Sulfur and nitrogen are vital elements for microbial survival, which serve as building blocks for nucleotides, amino acids, and cofactors [88,89]. The host can limit pathogen access to these nutrients through enzymatic degradation and metabolic rerouting. For nitrogen, a major host strategy is the depletion of arginine[90] and tryptophan [55]. Arginine is consumed by host arginase, which diminishes its availability for the synthesis of nitric oxide and microbial growth, while tryptophan is broken down by indoleamine 2,3-dioxygenase (IDO), generating immunomodulatory metabolites like kynurenine [55]. Interestingly, pathogens have also developed mechanisms to bypass these restrictions. For example, Chlamydia trachomatis encodes tryptophan synthase, with which it synthesizes tryptophan de novo when host levels drop [91]. Similarly, Mycobacterium tuberculosis upregulates nitrogen absorption pathways under nutrient stress, promoting the bacteria’s survival within macrophages [48]. These adaptations point to the metabolic flexibility of intracellular pathogens in response to nitrogen limitations orchestrated by the host. Sulfur metabolism is not much characterized in nutritional immunity, but it is gaining attention. Meanwhile, amino acids that contain sulfur, such as methionine and cysteine, are necessary for microbial antioxidant defenses and protein synthesis [92]. The host may decrease sulfur availability by downregulating the expression of transporters and enzymes involved in sulfur intake (Kies & Hammer, 2022). Additionally, oxidative stress caused by immune cells can oxidize sulfur compounds, making them inaccessible to the invading pathogens [94]. An in-depth understanding of how the host restricts sulfur and nitrogen will unveil new avenues for antimicrobial development. For example, targeting microbial sulfur assimilation or exploiting nitrogen starvation responses could facilitate pathogen clearance. Moreover, modulating host enzymes like arginase and IDO offers potential for immunotherapy, particularly in chronic pathogenic infections and cancer, where nutrient metabolism is intertwined with immune regulation.
3.6. Vitamin Sequestration
Vitamins are critically required as cofactors in metabolic pathways and enzymatic reactions (Hanna et al., 2022; Tardy et al., 2020). Following bacterial infection, the host can restrict access to vitamins such as folate, biotin, and vitamin B12, which creates an impediment for microbial metabolism [97]. This strategy is particularly potent against microbes that depend on host-derived vitamins due to incomplete biosynthetic pathways. Lipocalin-2, classically known for binding bacterial siderophores, has also been implicated in withholding vitamins. It can bind vitamin-like molecules and interfere with microbial uptake, extending its role beyond iron restriction [98]. Additionally, host cells may downregulate vitamin transporters or alter vitamin metabolism during infection, further limiting pathogen access. For example, human cytomegalovirus (CVM) infection has been shown to downregulate the vitamin D receptor (VDR) in host cells, impacting the vitamin D system and influencing immunity [99]. Some pathogens overcome this roadblock by synthesizing vitamins de novo. For example, Salmonella typhimurium possesses a full vitamin B12 biosynthesis pathway, which permits it to flourish in the nutrient-limited gastric mucosa [100]. Also, Mycobacterium tuberculosis expresses high-affinity transporters for biotin and folate, which are critical for fatty acid synthesis and nucleotide metabolism, respectively [101]. Fungal pathogens like Candida albicans and Cryptococcus neoformans also possess redundant vitamin acquisition systems, enabling survival in nutrient-limited niches [102,103]. Therapeutically, vitamin restriction presents a novel angle for antimicrobial intervention as inhibitors of microbial vitamin biosynthesis or transport could selectively hamper pathogen metabolism without affecting host cells. Overall, vitamin sequestration is a budding edge in nutritional immunity, and understanding how pathogens obtain and utilize these micronutrients could uncover new weak points for therapeutic targeting.
3.7. Carbon Source Restriction
Carbon sources such as glucose, amino acids, and fatty acids are fundamental to microbial biosynthetic processes and energy production (Bhagwat et al., 2025; Passalacqua et al., 2016). The host restricts carbon availability for the invading pathogens by modifying transporter expression, nutrient compartmentalization, and metabolic flux [106,107]. Upon activation, immune cells utilize large amounts of glucose and glutamine, denying pathogens of these substrates and creating a competitive metabolic environment [40]. Pathogens adapt by resorting to alternative carbon sources or scavenging metabolites generated by the host. Listeria monocytogenes, for example, circumvents glucose limitation during intracellular growth by utilizing host glycerol and acetate [47]. Also, Salmonella enterica expresses several carbon transporters and metabolic enzymes that help it to thrive in nutrient-poor vacuoles [48]. These adaptations demonstrate the metabolic versatility required for intracellular survival. Carbon source restriction also shapes immune responses. For example, metabolites such as lactate and succinate, generated during glycolysis and the TCA cycle, modulate cytokine production and immune cell differentiation [61]. Therefore, nutrient competition between host and pathogen not only affects microbial survival but also influences the immune landscape. Consequently, targeting microbial carbon metabolism offers therapeutic potential. For example, inhibitors of key glycolytic enzymes, glycerol uptake, or acetate metabolism could hinder pathogen replication. Moreover, modulating host metabolism to increase nutrient competition or immune activation may offer synergistic benefits in infection control.
3.8. Amino Acid Deprivation and Metabolic Reprogramming
Amino acids are necessary for protein synthesis, metabolic regulation, and immune signaling [44,108]. The host limits amino acid availability to pathogens through enzymatic degradation and transporter modulation. For instance, tryptophan is broken down by IDO, while arginine is used up by arginase, reducing their availability to pathogens and modulating immune responses [42,43]. These mechanisms are particularly efficacious against intracellular pathogens that depend on host amino acids [25]. Pathogens respond by amplifying biosynthetic pathways or scavenging peptides from the host. For example, Chlamydia trachomatis generates tryptophan de novo, while Salmonella enterica expresses amino acid transporters that enhance uptake under limiting conditions [48,91]. These adaptations are often controlled by nutrient-sensing systems that coordinate metabolic and virulence gene expression. Further, amino acid deprivation also reprograms host immunity by activating autophagy, stress responses, and metabolic checkpoints that facilitate pathogen clearance. For instance, glutamine and leucine availability influence mTOR signaling, which regulates T cell activation and differentiation [54]. Thus, amino acid metabolism is a junction of nutritional immunity and immune regulation. Manipulating amino acid availability presents therapeutic opportunities for infection control and immunomodulation. Arginase modulators, IDO inhibitors, and amino acid analogs are being researched in infectious disease and cancer. Understanding the context-specific effects of amino acid deprivation will be key to safely and effectively exploiting these strategies.
4. Nutritional Immunity in the Context of Specific Pathogens
Nutritional immunity is not a monolithic defense strategy. Rather, it is dynamic, pathogen-specific, and shaped by the ecological niche, virulence mechanisms, and metabolic dependencies of invading microbes. This section examines how different classes of pathogens, including extracellular, intracellular, and eukaryotic, interact with host nutrient sequestration systems, and how comparative genomics illustrate evolutionary adaptations in nutrient acquisition.
4.1. Extracellular Pathogens: Confronting Nutrient Sequestration Head-On
Extracellular pathogens, which dwell outside host cells, are directly exposed to the host’s nutrient-sequestering proteins at mucosal surfaces and the bloodstream (Figure 2) [109]. For example, Neisseria meningitidis must compete with iron-binding proteins like lactoferrin and transferrin in the cerebrospinal fluid and blood [110]. To overcome this, N. meningitidis expresses transferrin-binding proteins like TbpA and TbpB, which directly extract iron from host transferrin [111,112]. This direct contest for iron is a major attribute of extracellular pathogens, which often develop surface receptors that mimic host ligands. A different strategy is employed by Streptococcus pneumoniae, which expresses pneumococcal surface protein A (PspA) that suppresses complement deposition and indirectly protects against nutritional immunity by inhibiting immune activation [113,114]. S. pneumoniae also uses ABC transporters to obtain zinc and manganese, opposing sequestrations mediated by calprotectin [115]. These metal acquisition systems are highly regulated and often connected with virulence gene expression, highlighting the twofold role of nutrient sensing in pathogenesis. Staphylococcus aureus and Pseudomonas aeruginosa are particularly proficient at surviving in nutrient-constrained settings. For example, S. aureus expresses iron-regulated surface determinant (Isd) proteins, which extract heme from hemoglobin and deliver it across the bacterial envelope [116]. Similarly, P. aeruginosa synthesizes multiple siderophores, including pyochelin and pyoverdine, which scavenge iron from host proteins and are regulated by quorum sensing [117,118]. These systems are critical for survival in iron-deprived niches such as abscesses. Another extracellular pathogen, Escherichia coli, especially uropathogenic strains (UPEC), deploys covert siderophores like salmochelin to elude lipocalin-2, a host protein that binds and neutralizes bacterial siderophores [65,119]. Multiple iron acquisition systems are also expressed by UPEC, allowing it to adjust to different iron sources in the urinary tract [120]. Altogether, extracellular pathogens express a diverse array of nutrient acquisition strategies that demonstrate their exposure to host defenses and their need for rapid adaptation.
4.2. Intracellular Pathogens: Navigating the Nutrient Desert Within
Intracellular pathogens contend with a different challenge of surviving within host cells, where nutrients are highly regulated and often withheld in organelles [121]. Interestingly, this group of pathogens has also evolved various strategies for overcoming these nutrient limitations. Mycobacterium tuberculosis, for example, resides within macrophage phagosomes [122], where iron and zinc are restricted [123]. To overcome this limitation, the pathogen expresses siderophores called mycobactins and amplifies the expression of iron acquisition genes in low-iron situations [124]. Additionally, M. tuberculosis modulates host lipid metabolism to acquire fatty acids as carbon sources, revealing its metabolic flexibility [125]. Salmonella enterica survives within Salmonella-containing vacuoles (SCVs) in macrophages and epithelial cells [126]. It recognizes magnesium limitation through the PhoP/PhoQ system and activates genes that increase survival in magnesium-limited environments [127]. It also expresses ZnuABC and SitABCD transporters to acquire zinc and manganese, respectively, countering calprotectin-mediated sequestration [128,129]. These adaptations are necessary for the pathogen’s intracellular replication and systemic dissemination. Toxoplasma gondii, an obligate intracellular parasite, lives in a parasitophorous vacuole that is largely detached from host endocytic pathways [130]. It scavenges host amino acids and cholesterol through specialized transporters and manipulates host autophagy to access nutrients [131,132]. Nutrient limitation triggers stage conversion from tachyzoite to bradyzoite, a dormant form that enhances persistence[133]. This developmental plasticity is a trademark of intracellular eukaryotic pathogens. Coxiella burnetii, Chlamydia trachomatis, and Rickettsia prowazekii each occupy distinct intracellular niches[25]. For example, C. burnetii thrives in acidic lysosome-like vacuoles and expresses transporters for amino acids and metals [134]. C. trachomatis resides in an inclusion body and encodes tryptophan synthase to compensate for host IDO-mediated tryptophan depletion [91]. R. prowazekii, which replicates in the cytosol, scavenges host amino acids and ATP, reflecting its reductive genome and reliance on host metabolism [25]. Overall, these pathogens embody the diverse strategies used to navigate intracellular nutrient deserts.
4.3. Eukaryotic Pathogens: Complexity, Redundancy, and Immune Evasion
Eukaryotic pathogens, including fungi, parasites, and protozoa, have complex life cycles and redundant metabolic pathways that help them evade nutritional immunity [4,6]. For example, Plasmodium falciparum, the protozoan causative agent of malaria, invades red blood cells and breaks down hemoglobin to access amino acids and heme [135,136]. It detoxifies heme into hemozoin and expresses transporters for iron and other metals [136]. Host iron status determines malaria severity, and iron supplementation can worsen infection, underscoring the delicate balance of iron homeostasis [137]. Leishmania species reside in macrophage phagolysosomes and express ZIP family transporters for iron and zinc [138]. They also synthesize surface glycoconjugates like lipophosphoglycan (LPG) that manipulate host immune responses, enhancing nutrient acquisition [139]. The ability of Leishmania to survive in acidic, nutrient-limited compartments highlights its evolutionary adaptation to unfriendly intracellular environments. Fungal pathogens such as Candida albicans, Cryptococcus neoformans, and Aspergillus fumigatus possess robust nutrient acquisition systems. For instance, C. albicans expresses siderophore transporters and ferric reductases to acquire iron, and its metabolic flexibility permits it to flourish in diverse host niches [140]. C. neoformans synthesizes melanin and capsule components that defend the pathogen against oxidative stress and facilitate iron uptake [141]. A. fumigatus expresses high-affinity iron transporters and secretes siderophores, which are critical for virulence in immunocompromised hosts [142]. These eukaryotic pathogens also dodge nutritional immunity through immune modulation, antigenic variation, and metabolic redundancy [143,144]. The fact that their genomes encode multiple isoforms of nutrient transporters and enzymes allows them to adapt to varying host environments. Furthermore, their ability to modulate host cell signaling and immune responses adds a layer of complexity to their nutrient acquisition strategies, making them formidable adversaries in the context of nutritional immunity.
4.4. Comparative Genomics: Mapping the Evolutionary Landscape of Nutrient Acquisition
Comparative genomics has elucidated the evolutionary trajectories of nutrient acquisition systems across diverse pathogens. By evaluating genome content, gene expression, and regulatory networks, researchers have spotted conserved and lineage-specific adaptations that depict ecological niches and host interactions. For instance, siderophore biosynthesis genes are common among Gram-negative bacteria but differ in structure and regulation, indicating convergent evolution under iron-constrained settings [145]. Intracellular pathogens often exhibit genome reduction, shedding redundant metabolic pathways while preserving or expanding nutrient transporters [25]. Chlamydia and Rickettsia species, for example, have streamlined genomes but encode specialized systems for extracting host metabolites, which illustrates their obligate intracellular lifestyle [146,147]. Contrastingly, facultative intracellular pathogens like Salmonella maintain versatile metabolic networks, allowing survival in both intracellular and extracellular environments[148]. Eukaryotic pathogens show extensive gene duplication and diversification in nutrient acquisition genes [149,150]. For example, Plasmodium species have several copies of enzymes and transporters involved in amino acid and metal metabolism, demonstrating their complex life cycle and host transitions [151]. Also, fungal pathogens possess expanded gene families for oxidative stress resistance and iron uptake, which corresponds with their capacity to colonize diverse tissues [152]. Comparative genomics also reveals horizontal gene transfer events that facilitate nutrient acquisition [153]. Pathogens may acquire metal transporters, or vitamin synthesis genes, or siderophore biosynthesis clusters from other microbes, expanding their metabolic repertoire. These genomic insights underline the evolutionary arms race between host nutritional immunity and microbial adaptation, presenting targets for therapeutic intervention and vaccine development.
5. Pathogen Adaptation and Evasion
Pathogens have evolved sophisticated mechanisms to counteract nutrient restrictions imposed by the host. These adaptations not only ensure survival in hostile environments but also boost virulence and persistence. This section examines the molecular strategies pathogens employ to overcome nutritional immunity and the evolutionary dynamics that shape nutrient acquisition systems.
5.1. Pathogen Strategies to Overcome Nutritional Immunity
5.1.1. Siderophore Diversification and Stealth
Siderophores are high-affinity iron-chelating molecules produced by bacteria to scavenge iron from host proteins [154]. Pathogens have diversified their siderophore structures to evade host defenses such as lipocalin-2, which binds and neutralizes siderophores [64]. Escherichia coli, for instance, synthesizes salmochelin, a glycosylated derivative of enterobactin that is not recognized by lipocalin-2 [155]. This structural modification permits pathogens to sustain iron acquisition under immune pressure. Diversification also includes the production of multiple siderophores with unique chemical properties. For example, Pseudomonas aeruginosa produces both pyochelin and pyoverdine, which differ in regulatory control and affinity, allowing the pathogen to adapt to varying iron availability [117]. Similarly, Yersinia pestis synthesizes yersiniabactin, which not only chelates iron but also binds copper, playing a twofold role in metal acquisition and detoxification[156]. Stealth siderophores are often encoded within pathogenicity islands and controlled by iron-responsive transcription factors such as Fur (Katumba et al., 2022; Leon-Sicairos et al., 2015). These systems are highly connected with virulence, as iron acquisition is critical for replication and immune evasion. The ability to overcome host sequestration while sustaining iron uptake underlines the evolutionary pressure driving siderophore innovation.
5.1.2. Metal Transporter Upregulation
In response to metal limitation imposed by the host, pathogens upregulate high-affinity transporters for zinc, manganese, and magnesium. For instance, the ZnuABC and MntH systems are widely conserved among Gram-negative bacteria and are critical for survival under calprotectin-mediated sequestration [33]. These transporters are often regulated by metal-responsive repressors such as Zur [159,160] and MntR [161,162], which fine-tune expression based on intracellular metal levels. Staphylococcus aureus exemplifies this strategy by expressing multiple metal transporters and metalloregulatory proteins [163]. It uses the MntABC system for manganese uptake and adapts its metalloproteome to replace scarce metals with available alternatives [164]. This flexibility permits pathogens to sustain enzymatic function and combat oxidative stress even under metal deprivation. Transporter upregulation is usually integrated with virulence gene expression. For example, Salmonella enterica activates SitABCD [165] and ZnuABC [166] during intracellular infection, enhancing survival in macrophage phagosomes. These systems facilitate nutrient acquisition and also contribute to immune evasion and persistence, underscoring their dual role in pathogenesis.
5.1.3. Metabolic Rewiring and Carbon Source Flexibility
Pathogens often reprogram their metabolism to utilize alternative carbon sources when preferred substrates are limited [167]. This metabolic adaptability is important for survival in nutrient-restricted environments such as inflamed tissues or intracellular compartments. For instance, Listeria monocytogenes transitions from glucose to host-derived acetate and glycerol during intracellular growth [168,169]. This switch is governed by PrfA, a master virulence regulator that integrates metabolic and pathogenic signals [168]. Similarly, Mycobacterium tuberculosis displays extreme metabolic plasticity by utilizing host lipids as carbon sources during intracellular infection. For example, it upregulates genes involved in β-oxidation and the glyoxylate shunt, allowing it to persist in macrophages where glucose is scarce [125]. This adaptation not only supports energy production but also manipulates immune responses by reshaping host lipid metabolism. Carbon source flexibility often supports virulence and immune evasion, in that pathogens that can exploit host metabolites are better prepared to survive nutrient limitation and immune activation. Therefore, metabolic rewiring represents a strategic adaptation that improves microbial fitness and pathogenicity in diverse host environments.
5.1.4. Amino Acid Scavenging and Biosynthesis
Amino acid restriction is a common host strategy to repress pathogen growth [170,171]. In response, pathogens either upregulate biosynthetic pathways or scavenge amino acids from the host. For example, Chlamydia trachomatis encodes tryptophan synthase to counteract host IDO-mediated tryptophan depletion, enabling sustained replication in epithelial cells [91]. This enzyme is controlled by nutrient availability and immune signals, highlighting its role in adaptation. Similarly, Mycobacterium tuberculosis [172] and Salmonella enterica [173] also express biosynthetic enzymes and amino acid transporters that are upregulated during intracellular infection. These systems permit the pathogens to thrive in vacuoles where amino acid levels are tightly controlled [48]. Amino acid scavenging is often coupled with stress responses and the expression of virulence genes, which altogether improve survival under immune pressure. Further, some pathogens manipulate host autophagy to access amino acids. Toxoplasma gondii [174] and Coxiella burnetii [175], for instance, both induce host autophagy to release nutrients into the parasitophorous vacuole. This strategy not only supplies amino acids to the pathogens but also modulates host immunity, illustrating the multidimensional role of amino acid acquisition in pathogenesis.
5.1.5. Host Manipulation and Immune Evasion
Aside from acquiring nutrients, pathogens actively modulate host processes to evade nutritional immunity. For example, Leishmania spp alter phagosome maturation and acidification, creating a niche for optimal nutrient acquisition and minimized immune detection [176]. These modifications are facilitated by surface glycoconjugates and secreted effectors that interfere with host signaling. Similarly, Plasmodium falciparum reprograms red blood cells to enhance nutrient uptake and waste disposal [177]. It sends out proteins that form new permeability pathways, creating access to amino acids and glucose [178]. These alterations also decrease immune recognition, improving parasite survival and replication. Immune evasion strategies often involve modulation of host cytokine responses and antigen presentation [179]. Mycobacterium tuberculosis produces ESAT-6 and other effectors that block phagosome-lysosome fusion and suppress pro-inflammatory signaling [180]. These tactics not only keep the pathogen from immune clearance but also preserve access to intracellular nutrients, reinforcing the connection between immune evasion and nutritional adaptation.
5.2. Evolution of Nutrient Acquisition Mechanisms in Pathogens
5.2.1. Gene Expansion and Operon Architecture
Pathogens often expand gene families involved in acquiring nutrients to increase redundancy and adaptability [181]. Genes involved in siderophore biosynthesis and transport are frequently duplicated and organized into operons, enabling coordinated expression [182,183,184]. For example, Pseudomonas aeruginosa has several operons for pyoverdine synthesis, each controlled by the availability of iron and environmental cues [185]. Operon architecture promotes rapid response to nutrient limitation, since genes encoding transporters, regulators, and biosynthetic enzymes are co-transcribed, ensuring efficient resource allocation. This organization is particularly visible in metal acquisition systems such as ZnuABC and SitABCD, which are tightly regulated by metal-responsive repressors [186]. The expansion of genes also supports functional diversification. The duplicated genes may develop new substrate specificities or regulatory mechanisms, elevating pathogen fitness in diverse environments [187]. This evolutionary strategy illustrates the selective pressure imposed by nutritional immunity and the need for pathogens’ metabolic versatility.
5.2.2. Horizontal Gene Transfer and Convergent Evolution
Horizontal gene transfer (HGT) plays a critical role in the dissemination of nutrient acquisition and catabolism systems [153]. Pathogenicity islands commonly contain siderophore biosynthesis genes, metal transporters, and vitamin synthesis pathways obtained from other microbes [188]. Yersinia pestis, for example, acquired the yersiniabactin cluster via HGT, improving its iron acquisition capabilities [189]. Convergent evolution also determines nutrient acquisition strategies. Thus, unrelated pathogens may develop similar mechanisms to conquer host defenses, such as stealth siderophores or high-affinity transporters. This convergence reflects general selective pressures and underscores the functional importance of these adaptations [145]. HGT and convergence contribute to the speedy evolution of virulence characteristics. By acquiring and optimizing nutrient acquisition genes, pathogens can adapt to new hosts and niches, expanding their ecological range and pathogenic potential. These evolutionary dynamics reflect the fluidity of microbial genomes and the pivotal role of nutritional immunity in determining pathogen evolution.
5.2.3. Reductive Evolution in Obligate Intracellular Pathogens
Obligate intracellular pathogens commonly undergo reductive evolution, losing redundant metabolic pathways while preserving essential nutrient acquisition systems. For instance, Rickettsia and Chlamydia species have streamlined genomes but encode specialized transporters for amino acids, nucleotides, and ATP [147,190]. This dependency highlights their adaptation to nutrient-rich intracellular environments. Reductive evolution is fueled by genome decay and host dependency. Genes that are no longer needed for extracellular survival are lost, while those involved in host interaction and nutrient uptake are preserved or expanded. While reductive evolution increases efficiency by transferring biosynthetic functions to the host, it simultaneously creates nutritional fragility for the pathogen. For example, the reliance of Chlamydia on host tryptophan makes it highly susceptible to IDO-mediated starvation [91]. Likewise, Coxiella’s requirement for host-derived lipids and amino acids restricts its replication to the specialized parasitophorous vacuole [191]. These vulnerabilities highlight how nutritional immunity may not only limit pathogen growth but also influence long-term genome evolution.
6. Therapeutic Implications
Nutritional immunity not only serves as a major pillar for host defense but also presents a fertile landscape for therapeutic innovation. Through the comprehension and manipulation of nutrient availability, researchers and clinicians can develop targeted interventions that either starve pathogens or strengthen host defenses. Therapeutic strategies inspired by nutritional immunity, including their mechanisms of action, examples, clinical trials, challenges, and translational outlook, are presented in Table 2. Overall, this section evaluates the translational potential of nutritional immunity across antimicrobial strategies, host-directed therapies, and diagnostic applications.
6.1. Targeting Nutrient Availability for Infection Control
Manipulating the availability of nutrients is a promising approach for controlling infections, particularly in an era of rising antimicrobial resistance. Iron chelation, for example, has been considered as a strategy to restrict microbial growth by depriving pathogens of this critical cofactor. Agents such as deferoxamine and deferiprone bind free iron, lowering its bioavailability and thereby hindering the proliferation of iron-dependent pathogens like Staphylococcus aureus and Escherichia coli [242]. However, therapeutic iron chelation must be delicately balanced to avoid exacerbating anemia or impairing host immunity [7]. Beyond iron, zinc and manganese sequestration have emerged as viable antimicrobial strategies. For instance, calprotectin, a host protein that chelates these metals, has inspired the development of synthetic mimetics that mimic calprotectin’s metal-binding properties[243]. These agents can be deployed locally at infection sites to increase metal starvation without systemic toxicity [33]. Moreover, targeting microbial metal transporters with small-molecule inhibitors offers a pathogen-specific approach that circumvents traditional antibiotic mechanisms [244,245]. Carbon source restriction is another avenue for controlling infection. By manipulating host metabolism or blocking microbial access to key substrates, such as glucose or fatty acids, researchers can create unfavorable environments for pathogens. For instance, inhibitors of bacterial glycerol uptake have demonstrated efficacy against Listeria monocytogenes, which depends on host-derived glycerol during intracellular growth [47]. These strategies underscore the potential of metabolic interference as a non-antibiotic antimicrobial approach. Nevertheless, targeting nutrient availability must factor in host-pathogen specificity and tissue context. This is because nutrient manipulation may have inadvertent implications on host cells, particularly immune cells that depend on similar substrates for their activation and function. Thus, precision targeting, guided by pathogen biology and host physiology, is critical for the safe and effective deployment of nutrient-based therapies.
6.2. Novel Therapeutic Strategies Based on Nutritional Immunity
Recent advances in molecular biology and immunology have enabled the design of novel therapeutics that exploit or mimic nutritional immunity. One example of such a strategy involves the development of siderophore-antibiotic conjugates, also known as “Trojan horse” antibiotics. These compounds utilize bacterial iron uptake systems to deliver antibiotics directly into the pathogen. For instance, cefiderocol, a siderophore-cephalosporin conjugate, has shown potent activity against multidrug-resistant Gram-negative bacteria by seizing iron transport pathways [246]. Another innovative approach involves the use of engineered probiotics that battle with pathogens for nutrients. These beneficial microbes can be designed to sequester iron, zinc, or amino acids in the gut, thereby impeding pathogen colonization. For example, Lactobacillus strains engineered to express siderophore receptors have demonstrated the potential to reduce Salmonella burden in animal models [16]. Such strategies present a microbiome-friendly alternative to conventional antimicrobials. Also, standard antibiotics combined with nutrient iron chelators have shown amplified antimicrobial activity. For instance, the impregnation of catheters with both antimicrobial agents and iron chelators gave increased antimicrobial and anti-biofilm activity than just antimicrobial agents alone [72]. Also, thiostrepton, a Gram-positive thiopeptide antibiotic imported via pyoverdine receptors, synergized with the iron chelator deferasirox to yield higher growth suppression for P. aeruginosa and Acinetobacter baumannii in clinical isolates than thiostrepton alone [247]. Further, nanotechnology finds application in nutritional immunity-based therapeutics. For example, nanoparticles functionalized with metal-binding ligands can be employed to sequester iron or zinc at infection sites, creating localized nutrient deserts [248]. These platforms can be further adjusted to deliver immunomodulatory agents or antibiotics, enhancing their therapeutic efficacy [249,250]. The modularity of nanomedicine permits tailored interventions based on infection site and pathogen type. Essentially, these novel strategies must be evaluated for their impact on host nutrient homeostasis and immune function. While mimicking nutritional immunity can improve pathogen clearance, excessive nutrient deprivation may impair tissue repair or compromise immune activation. Thus, therapeutic design must integrate insights from host-pathogen interactions, nutrient biology, and immunometabolism to ensure the best outcomes.
6.3. Nutrient-Based Therapies for Specific Infections
Nutrient-based therapies have demonstrated promise, particularly in treating infections where nutrient acquisition is closely linked to pathogenesis. Malaria, caused by Plasmodium falciparum, is a prime example where the parasite digests hemoglobin to access amino acids and iron [135,177], making iron availability a pivotal determinant of disease severity [251]. Consequently, iron chelation therapy using agents like deferoxamine has been explored to control parasite growth, although its clinical utility remains controversial owing to potential host toxicity [252,253,254]. In tuberculosis, iron and lipid metabolism are pivotal to Mycobacterium tuberculosis survival [255]. Host-directed therapies that modulate iron availability or lipid metabolism have also been proposed to improve bacterial clearance. Similarly, statins, which alter host lipid metabolism, may hinder M. tuberculosis access to carbon sources and improve immune responses [256]. Fungal infections also offer opportunities for nutrient-based interventions. Candida albicans and Cryptococcus neoformans depend on iron acquisition for virulence [141,192]. Iron chelation has been shown to lower fungal burden in animal models, and iron-binding compounds are being researched as adjuncts to antifungal therapy [257]. These approaches are especially relevant in immunocompromised patients, where conventional antifungals may be inadequate [258]. However, nutrient-based therapies must be essentially customized for the pathogen’s metabolic profile and the host’s nutritional status. Iron chelation, for example, may be unadvised in anemic patients or those with chronic infections. Personalized medicine approaches that integrate nutritional biomarkers, pathogen genomics, and host physiology are necessary for the safe and effective use of these therapies.
6.4. Host-Directed Therapies to Enhance Nutritional Immunity
Host-directed therapies (HDTs) aim to strengthen the host’s natural defenses, including nutritional immunity, to fight infection. One strategy involves increasing the expression or activity of nutrient-sequestering proteins such as calprotectin, hepcidin, and lipocalin-2. For example, synthetic hepcidin analogs have been developed to lower serum iron levels and repress bacterial growth in sepsis models [11]. These agents mimic the host’s iron-chelating response and may serve as supplements to antibiotic therapy. Immunomodulatory agents can also be used to switch on pathways that limit nutrient availability. Interferon-γ, for instance, triggers indoleamine 2,3-dioxygenase (IDO), which depletes tryptophan and restrains intracellular pathogens like Toxoplasma gondii and Chlamydia trachomatis [42]. Enhancing IDO activity or mimicking its effects may present therapeutic benefits in chronic infections where immune evasion is common. Further, gene therapy and RNA-based approaches are emerging tools for controlling host nutrient responses. CRISPR-based editing of genes involved in iron metabolism or metal transport could also be used to improve resistance to specific pathogens. Similarly, siRNA targeting of host nutrient transporters may lower pathogen access to critical substrates without impairing host function [259]. These technologies provide precision control over host-pathogen interactions. Despite their potential, HDTs must be carefully examined for off-target effects and long-term safety. Modulating nutrient availability can affect immune cell function, tissue repair, and systemic metabolism [260]. Therefore, HDTs should be integrated into a broader therapeutic framework that considers nutritional status, pathogen biology, and host immunity.
6.5. Diagnostic Biomarkers of Nutritional Immunity
The active interplay between host nutrient sequestration and pathogen adaptation produces quantifiable changes in biomarker profiles that can be explored for diagnostics. The levels of ferritin, transferrin, and hepcidin saturation in the serum are commonly used to determine iron status and inflammation. For example, elevated ferritin and hepcidin levels are suggestive of iron sequestration during infection and can help differentiate between iron-deficiency anemia and anemia of inflammation [11]. Calprotectin, a manganese- and zinc-binding protein, is a well-established biomarker of neutrophil activation and mucosal inflammation [261,262]. Fecal calprotectin is commonly used to monitor inflammatory bowel disease, and its elevation in systemic infections may indicate the activation of nutritional immunity [263]. Similarly, the serum levels of lipocalin-2 may correlate with bacterial infection and may serve as a marker of iron-chelating responses [264]. Metabolomic profiling presents a high-resolution view of nutrient flux during infection. Changes in amino acid levels, such as tryptophan depletion and kynurenine accumulation, portray IDO activity and immune modulation. These metabolites can be measured using mass spectrometry and integrated into diagnostic algorithms for infections like HIV and tuberculosis [42]. Such approaches allow real-time monitoring of host-pathogen interactions. Emerging technologies such as biosensors and wearable devices may further improve the utility of nutritional immunity biomarkers. By continuously tracking nutrient-related parameters, clinicians can personalize therapy and assess the efficacy of treatment. Ultimately, merging biomarker data with genomic and immunologic profiles will facilitate precision diagnostics and targeted therapeutic interventions.
7. Future Directions
As the field of nutritional immunity evolves, it is increasingly clear that nutrient sequestration is not merely a passive defense but a dynamic and integrative component of host-pathogen interactions. Future efforts and research must address the mechanistic complexity, translational potential, and global significance of nutritional immunity, particularly in the context of emerging infectious diseases, malnutrition, and microbiome science. Essentially, this section offers a window into the future of a complete understanding of the components and applications of the concept of nutritional immunity.
7.1. Emerging Research Areas in Nutritional Immunity
Recent advances in systems biology and high-throughput technologies offer new avenues for exploring nutritional immunity at unprecedented resolution. For example, single-cell transcriptomics and spatial proteomics now permit researchers to map nutrient sequestration responses at the cellular and tissue level during infection [265]. These platforms are unveiling heterogeneity in immune cell responses, including differential expression of metal transporters, amino acid catabolic enzymes, and nutrient-sensing receptors across immune subsets. Another emerging area involves the exploration of non-classical nutrients and their role in immunity. For instance, while iron, manganese, and zinc have been extensively studied, elements such as copper, selenium, and molybdenum are receiving attention for their roles in redox regulation and enzymatic activity during infection [266]. A good understanding of how these trace elements are regulated and exploited by pathogens could reveal novel therapeutic targets and biomarkers. The interconnection between nutritional immunity and immunometabolism is also a rising field. For example, we now know that immune cells undergo metabolic reprogramming during activation, consuming large quantities of glucose, glutamine, and fatty acids [267]. This competition for nutrients between host and pathogen creates a metabolic tug-of-war that shapes the outcome of infection [40]. Investigating how nutrient availability affects immune cell fate decisions, cytokine production, and pathogen clearance will be relevant for creating new metabolic interventions. Finally, not much is known yet about the role of nutritional immunity in viral infections. While most research has focused on bacterial and fungal pathogens, viruses also modulate host nutrient pathways in a way that supports replication. For instance, HIV [69] and SARS-CoV-2 [268] alter iron metabolism and amino acid availability, which impacts immune function and disease progression. Expanding nutritional immunity studies to include viral pathogens will provide a more comprehensive understanding of host defense.
7.2. Translating Nutritional Immunity Into Clinical Applications
Translating the principles of nutritional immunity into clinical practice needs a multidisciplinary approach that synergizes microbiology, immunology, pharmacology, and nutrition science. One promising direction is the development of host-directed therapies that amplify nutrient sequestration. For example, synthetic hepcidin analogs are being explored to treat iron overload disorders and limit iron availability during bacterial infections [269,270,271]. These agents could be repurposed to boost nutritional immunity in sepsis or chronic infections. Another translational route involves the use of nutritional biomarkers to inform clinical decision-making. For instance, serum levels of ferritin, hepcidin, and calprotectin can tell the inflammatory status and predict infection severity. Incorporating these biomarkers into diagnostic panels may enhance early detection and stratification of infectious diseases [272]. Further, metabolomic profiling of amino acid and metal fluxes could offer real-time insights into host-pathogen dynamics and therapeutic efficacy. Personalized medicine strategies are also gaining traction. This is because genetic polymorphisms in immune regulators and nutrient transporters shape individual susceptibility to infection and response to therapy. For instance, mutations in the SLC11A1 gene, which encodes the NRAMP1 transporter, affect macrophage metal handling and resistance to intracellular pathogens [273]. Genotyping patients for such variants could guide tailored interventions that improve nutritional immunity. Clinical translation must also factor in safety and off-target effects. The manipulation of nutrient availability can influence host metabolism, immune function, and tissue repair. Thus, therapeutic strategies must be precisely targeted and context-specific. Robust clinical trials, pharmacokinetic modeling, and long-term safety assessments will be necessary to ensure that nutritional immunity-based therapies are both safe and effective.
7.3. Nutritional Immunity in the Context of Global Health and Malnutrition
Nutritional immunity has immense implications for global health, especially in regions burdened by infectious diseases and malnutrition. For example, iron deficiency is the most common nutritional disorder globally and can impair immune function while paradoxically protecting against certain infections like malaria [137]. A major challenge in public health nutrition remains how to balance iron supplementation with the risk of infection. Malnutrition alters the host’s ability to mount effective nutritional immunity. Protein-energy malnutrition and micronutrient deficiencies hamper the synthesis of key immune proteins such as hepcidin, calprotectin, and lipocalin-2, weakening the host’s ability to sequester nutrients from pathogens [274]. This creates a vicious cycle where infection aggravates malnutrition, and malnutrition impairs immune defense, particularly in children and immunocompromised populations. Global health efforts must integrate nutritional immunity into their frameworks for disease prevention and treatment. Programs that offer micronutrient supplementation should review infection status and tailor interventions accordingly. Zinc supplementation, for instance, has been shown to lower the duration and severity of diarrheal diseases in children, partly by improving innate immune responses [275]. Increasing our understanding of the immunological context of nutrient interventions will help in enhancing their efficacy and safety. Notably, urbanization, climate change, and food insecurity are modifying the landscape of infectious disease and nutrition [276,277]. Consequently, emerging pathogens may take advantage of novel nutrient niches, while shifts in dietary patterns could affect host immunity. Future global health strategies must embrace a systems-level perspective that considers the interplay between nutrition, immunity, and pathogen ecology to build strong health systems.
7.4. Nutritional Immunity and the Microbiome
The human microbiome plays a critical role in shaping nutritional immunity. Commensal microbes compete with pathogens for nutrients, generate antimicrobial metabolites, and influence host nutrient metabolism [23,278]. For example, gut bacteria can sequester iron and produce siderophores that inhibit pathogen growth, effectively acting as an extension of the host’s nutritional defense [279,280]. This microbial competition is a major determinant of colonization resistance. The composition of the microbiome also influences host nutrient sensing and immune responses. For example, short-chain fatty acids (SCFAs) synthesized by microbial fermentation of dietary fiber modulate immune cell function and elevate barrier integrity [281,282,283]. These metabolites affect the expression of nutrient transporters and immune regulators, influencing the host’s ability to respond to infection [284]. Dysbiosis, or microbial imbalance, can disrupt these processes and compromise nutritional immunity [285,286]. Probiotic and prebiotic interventions offer a promising strategy to improve nutritional immunity [49]. By promoting beneficial microbes that compete with pathogens and support host nutrient metabolism, these therapies can reinforce immune defenses. For instance, Lactobacillus and Bifidobacterium strains have been shown to amplify host antimicrobial peptides and modulate iron metabolism, lowering susceptibility to enteric infections [287,288,289]. Such strategies are especially relevant during antibiotic treatment and in vulnerable populations. Future research must illuminate the molecular mechanisms by which the microbiome interacts with nutritional immunity. Integrating microbiome profiling with immunometabolic analysis will essentially provide a holistic view of the interactions between host, pathogen, and microbe. Ultimately, harnessing the microbiome to support nutritional immunity is a frontier in precision medicine and infectious disease prevention.
8. Conclusion
Nutritional immunity is a chief cornerstone of host defense, which is intricately intertwined with the molecular and cellular fabric of immune responses. Its role in influencing pathogen behavior, determining disease outcomes, and guiding therapeutic innovation has become increasingly evident across disciplines. Notably, the concept of nutritional immunity has evolved from a narrow focus on iron sequestration to a multifaceted framework encompassing a wide array of nutrients, including metals, vitamins, amino acids, and carbon sources. Host organisms deploy a sophisticated arsenal of nutrient-binding proteins, transport regulators, and metabolic reprogramming strategies to deprive pathogens of critical resources. These mechanisms are not independent but are deeply interwoven with immune signaling pathways, cellular stress responses, and tissue-specific defense programs. In response, pathogens have equally developed complex countermeasures, including the synthesis of stealth siderophores, amplification of high-affinity transporters, metabolic rewiring, and modulation of host nutrient pathways. The diversity of these adaptations illustrates the ecological niches and evolutionary pressures faced by different microbial taxa. Therapeutically, nutritional immunity presents a rich landscape for innovation. For instance, strategies that mimic or bolster nutrient sequestration, such as siderophore-antibiotic conjugates, metal chelators, and host-directed therapies, have shown promise in preclinical and clinical settings. Additionally, diagnostic biomarkers derived from nutrient fluxes and immune responses are also emerging as critical tools for infection monitoring and personalized medicine. Future research must essentially prioritize the integration of nutritional immunity into clinical frameworks. This includes the creation of diagnostic platforms that monitor nutrient-related biomarkers, the design of therapeutics that modulate host nutrient responses, and the refinement of nutritional interventions based on infection status. To optimize these strategies, personalized medicine approaches that take into account genetic variation in nutrient handling, immune function, and microbiome composition will be relevant. Importantly, global health initiatives must also factor in the principles of nutritional immunity, such as understanding the interplay between nutrient status and immune competence in regions affected by malnutrition and infectious disease. Accordingly, programs that provide micronutrient supplementation should be tailored to local pathogen burdens and host nutritional profiles. Moreover, emerging infectious diseases, facilitated by urbanization, ecological disruption, and climate change, may take advantage of novel nutrient niches, necessitating adaptive public health responses. Finally, the microbiome represents a frontier for nutritional immunity research. Commensal microbes affect host nutrient metabolism, compete with pathogens, and influence immune responses. Harnessing the microbiome to support nutritional immunity, through probiotics, prebiotics, and microbiome-targeted therapies, offers a promising approach to prevent and control infections. As studies continue to unravel the molecular complexities of nutritional immunity, its translation into clinical and public health practice could redefine the landscape of infectious disease control.

Box 1.
Nutritional immunity with lipids: nutrients and niche modulators.
|
Significance. Beyond metals and small metabolites, host lipids, including fatty acids, cholesterol, phospholipids, and lipid droplets (LDs), are both critical nutrient sources and structural building blocks for many pathogens, and they actively shape intracellular niches [290]. Host control over lipid trafficking and storage therefore functions as a form of nutritional immunity that can limit pathogen growth or, paradoxically, be subverted to enhance pathogen persistence [291,292]. Mechanisms and host responses. Infected cells reprogram lipid metabolism. For example, macrophages and other innate cells ramp up the uptake and esterification of cholesterol, synthesize neutral lipids, form LDs, and alter fatty-acid flux. LDs act as metabolic hubs and may be “weaponized” by the host, for example, by concentrating antimicrobial lipids or lipid-derived mediators, or, if hijacked, provide a nutrient depot for invaders [292,293,294]. Pathogen strategies. Intracellular bacteria and parasites exploit host lipids in multiple ways. These include import and catabolism of host cholesterol as seen in Mycobacterium tuberculosis [125], redirection of host fatty-acid metabolism [293], and interception of LDs and secretory trafficking to obtain membranes and energy as observed in Chlamydia, Salmonella, and Toxoplasma [290]. Dedicated bacterial systems such as M. tuberculosis cholesterol uptake and degradation pathways, coordinated by proteins like LucA enable utilization of host sterols for persistence [125,290,295]. Consequences and translational outlook. Targeting lipid access or utilization with statins, inhibitors of microbial cholesterol catabolism, modulation of LD biology, or host-directed metabolic reprogramming, offers promising adjunctive strategies to traditional antimicrobials especially in the wake of rising antibiotic resistance. However, given that lipids are central to host physiology, therapeutic windows are narrow and require precision [291,293]. |
Box 2.
Nutritional immunity in the context of viral infections.
| Nutritional immunity has been predominantly studied in bacterial and fungal pathogenesis. Yet, accumulating evidence illuminates its relevance during viral infections. Unlike bacteria, viruses do not directly acquire metals for replication but rely heavily on host cellular metabolic processes that are metal-dependent, including enzymatic activity, DNA/RNA synthesis, and immune signaling [296]. Therefore, the host–virus interaction with nutritional immunity is largely indirect, mediated through regulation of metal homeostasis, nutrients redistribution, and modulation of host immune responses. Iron metabolism is strongly implicated in viral infections. Many viruses, including Human Immunodeficiency Virus (HIV), hepatitis C virus (HCV), and cytomegalovirus (CMV), exploit host iron-dependent pathways for replication [297]. High iron levels can enhance viral replication, while iron limitation via hepcidin upregulation or therapeutic chelation can suppress viral spread [298]. Conversely, iron overload in conditions such as hemochromatosis predisposes patients to more severe viral infections [299]. Zinc plays a twofold role in antiviral defense and viral pathogenesis. For instance, intracellular zinc bolsters antiviral immunity by promoting interferon signaling and stabilizing antiviral proteins, while zinc-finger antiviral protein (ZAP) directly restricts viral RNA [300]. Supplementation of zinc has been shown to lower the replication of coronaviruses, influenza virus, and HIV in vitro [300,301]. However, pathogens may oppose host zinc redistribution strategies. For instance, HIV can alter zinc transporter expression to favor its persistence in macrophages [300]. Copper and manganese also intersect with viral infections. For example, copper possesses intrinsic antiviral activity, disrupting viral proteins and nucleic acids, a property exploited in both innate immunity and copper-based surface coatings for infection control [302]. Meanwhile, manganese, is critical for the cGAS–STING pathway, a pivotal antiviral sensing mechanism, and its depletion impairs interferon-mediated viral clearance [303]. Taken together, nutritional immunity in viral infections represents a subtler and more host-centric phenomenon than in bacterial infections, where the pathogen directly competes for metals. Viruses rewire host nutrient availability to their benefit, while the host leverages nutrient redistribution to limit viral replication and enhance immune defense. This emerging field opens new therapeutic avenues, including modulation of iron homeostasis, zinc supplementation, and copper-based antivirals which are already under investigation in both preclinical and clinical settings [304,305,306,307,308]. |
Box 3.
Clinical relevance of nutritional immunity.
|
A common clinical axis. Nutritional immunity is fundamental to several clinical syndromes and therapeutic dilemmas, including anemia of inflammation, infection risk in iron-overloaded hosts, and both the promise and perils of altering metal metabolism in patients. Anemia of inflammation. Proinflammatory cytokines (notably IL-6) trigger hepatic hepcidin, which downregulates ferroportin and hides iron within macrophages and hepatocytes [309]. This adaptive response restricts extracellular iron available to pathogens but also causes hypoferremia and anemia that can worsen patient outcomes if prolonged [7,308]. Managing anemia in infection therefore requires balancing limiting pathogen access to iron with host oxygen-carrying needs. Iron overload and infection susceptibility. Patients with hereditary hemochromatosis or transfusional iron overload have an elevated risk for siderophilic infections because excess labile iron overwhelms host sequestering mechanisms [310,311]. Similarly, iron chelation can be protective in some infections but may exacerbate others if it becomes bioavailable to microbes via their strategies of recapturing withheld iron [7,56,312]. Therapeutic manipulation: promise and caveats. Clinical interventions that modulate metal availability, including localized metal delivery, systemic chelators, or hepcidin-modulating agents, are evolving. However, risks include inducing iatrogenic anemia, impairing immune cell function, or unintentionally supplying accessible iron to pathogens [313,314]. There is need, therefore, for translational strategies such as localized delivery and pathogen-directed siderophore conjugates to be precisely targeted and tested in context-specific clinical trials. |
Box 4.
Future perspectives: nutritional immunity meets drug discovery.
 |
|
Convergence of fields. Nutritional immunity presents mechanistic targets for new therapeutics. However, the realization of that potential will be facilitated through the integration of computational design, nanomedicine, and immunometabolic insight. Recent advances in artificial intelligence (AI)-driven molecule discovery, precision nanodelivery, and systems immunometabolism offers a pipeline for agent development that is faster and more targeted than conventional antibiotic screens. AI and discovery. Machine-learning and deep-learning platforms have already identified novel antimicrobial scaffolds and optimized siderophore-antibiotic conjugates in silico and in vitro, expediting hit discovery and de-risking medicinal chemistry [315,316]. AI can prioritize compounds that selectively impair pathogen metal acquisition or exploit species-specific uptake systems. Nanomedicine and targeted modulation. Nanocarriers allow for precise spatial control in that they can deliver chelators, metal-mimetic agents (e.g., gallium), or zinc/copper payloads to infected tissues or intracellular vacuoles while limiting systemic toxicity [317,318,319,320]. Functionalization with targeting ligands such as antibodies and peptides can improve pathogen-cell specificity. Immunometabolic therapeutics. Host-directed modulation of immune cell metabolism such as altering macrophage metabolism to support nutrient withholding or enhanced antimicrobial effector functions, complements direct antimicrobial strategies and can lower resistance pressure [55,321]. Roadmap. A translational program should: (1) pair mechanistic animal models with multiomics readouts, especially metabolomics and metallomics (2) use AI to triage candidate chemistries that engage pathogen uptake systems, (3) validate targeted nanodelivery in safety models, and (4) design early-phase trials with robust biomarkers, including hepcidin, labile plasma iron, and tissue metallome, to ascertain on-target effects before large efficacy trials [315,317]. This integrated approach promises host-informed, pathogen-targeted therapeutics that exploit nutritional immunity while lowering collateral host injury. |
Authors contributions
CEU and OCA: Conceptualization. CEU and SS: Visualization. CEU, OCA, TFA, FAA, SS, VAA, MJO, II, KOO, JKE, AAO, CBU, MIA, CCU, LCU: Writing – original draft; Writing – review and editing.
Declaration of Interest Statement
The authors declare that the research was conducted in the absence of any competing interests.
References
- Zhang XX, Jin YZ, Lu YH, Huang LL, Wu CX, Lv S, et al. Infectious disease control: from health security strengthening to health systems improvement at global level. Global Health Research and Policy. 2023 Sept 5;8(1):38.
- Salam MdA, Al-Amin MdY, Salam MT, Pawar JS, Akhter N, Rabaan AA, et al. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare (Basel). 2023 July 5;11(13):1946.
- Muteeb G, Rehman MT, Shahwan M, Aatif M. Origin of Antibiotics and Antibiotic Resistance, and Their Impacts on Drug Development: A Narrative Review. Pharmaceuticals (Basel). 2023 Nov 15;16(11):1615.
- Kreimendahl S, Pernas L. Metabolic immunity against microbes. Trends in Cell Biology. 2024 June 1;34(6):496–508.
- Murdoch CC, Skaar EP. Nutritional immunity: the battle for nutrient metals at the host–pathogen interface. Nat Rev Microbiol. 2022 Nov;20(11):657–70.
- Wildeman AS, Culotta VC. Nutritional Immunity and Fungal Pathogens: A New Role for Manganese. Curr Clin Micro Rpt. 2024 June 1;11(2):70–8.
- Cassat JE, Skaar EP. Iron in Infection and Immunity. Cell Host & Microbe. 2013 May 15;13(5):509–19.
- Haley KP, Skaar EP. A battle for iron: host sequestration and Staphylococcus aureus acquisition. Microbes and Infection. 2012 Mar 1;14(3):217–27.
- Kehl-Fie TE, Skaar EP. Nutritional immunity beyond iron: a role for manganese and zinc. Curr Opin Chem Biol. 2010 Apr;14(2):218–24.
- Núñez G, Sakamoto K, Soares MP. Innate Nutritional Immunity. J Immunol. 2018 July 1;201(1):11–8.
- Ganz T, Nemeth E. Iron homeostasis in host defence and inflammation. Nat Rev Immunol. 2015 Aug;15(8):500–10.
- Cellier MF, Courville P, Campion C. Nramp1 phagocyte intracellular metal withdrawal defense. Microbes and Infection. 2007 Nov 1;9(14):1662–70.
- Kolliniati O, Ieronymaki E, Vergadi E, Tsatsanis C. Metabolic Regulation of Macrophage Activation. J Innate Immun. 2021 July 9;14(1):51–68.
- Drakesmith H, Prentice AM. Hepcidin and the Iron-Infection Axis. Science. 2012 Nov 9;338(6108):768–72.
- Malavia D, Crawford A, Wilson D. Chapter Three - Nutritional Immunity and Fungal Pathogenesis: The Struggle for Micronutrients at the Host–Pathogen Interface. In: Poole RK, editor. Advances in Microbial Physiology [Internet]. Academic Press; 2017 [cited 2025 Aug 23]. p. 85–103. (Microbiology of Metal Ions; vol. 70). Available from: https://www.sciencedirect.com/science/article/pii/S0065291117300061.
- Deriu E, Liu JZ, Pezeshki M, Edwards RA, Ochoa RJ, Contreras H, et al. Probiotic Bacteria Reduce Salmonella Typhimurium Intestinal Colonization by Competing for Iron. Cell Host & Microbe. 2013 July 17;14(1):26–37.
- Hood MI, Skaar EP. Nutritional immunity: transition metals at the pathogen-host interface. Nat Rev Microbiol. 2012 July 16;10(8). [CrossRef]
- Monteith AJ, Skaar EP. The impact of metal availability on immune function during infection. Trends in Endocrinology & Metabolism. 2021 Nov 1;32(11):916–28.
- Brophy MB, Nolan EM. Manganese and Microbial Pathogenesis: Sequestration by the Mammalian Immune System and Utilization by Microorganisms. ACS Chem Biol. 2015 Mar 20;10(3):641–51.
- Damo SM, Kehl-Fie TE, Sugitani N, Holt ME, Rathi S, Murphy WJ, et al. Molecular basis for manganese sequestration by calprotectin and roles in the innate immune response to invading bacterial pathogens. Proceedings of the National Academy of Sciences. 2013 Mar 5;110(10):3841–6.
- Djoko KY, Ong C lynn Y, Walker MJ, McEwan AG. The Role of Copper and Zinc Toxicity in Innate Immune Defense against Bacterial Pathogens*. Journal of Biological Chemistry. 2015 July 31;290(31):18954–61.
- Barwinska-Sendra A, Waldron KJ. Chapter Eight - The Role of Intermetal Competition and Mis-Metalation in Metal Toxicity. In: Poole RK, editor. Advances in Microbial Physiology [Internet]. Academic Press; 2017 [cited 2025 Sept 7]. p. 315–79. (Microbiology of Metal Ions; vol. 70). Available from: https://www.sciencedirect.com/science/article/pii/S0065291117300036.
- Abt MC, Pamer EG. Commensal bacteria mediated defenses against pathogens. Current Opinion in Immunology. 2014 Aug 1;29:16–22.
- Horrocks V, King OG, Yip AYG, Marques IM, McDonald JAK. Role of the gut microbiota in nutrient competition and protection against intestinal pathogen colonization. Microbiology (Reading). 2023 Aug 4;169(8):001377.
- Mandel CG, Sanchez SE, Monahan CC, Phuklia W, Omsland A. Metabolism and physiology of pathogenic bacterial obligate intracellular parasites. Front Cell Infect Microbiol. 2024 Mar 22;14:1284701.
- Kelly B, O’Neill LA. Metabolic reprogramming in macrophages and dendritic cells in innate immunity. Cell Res. 2015 July;25(7):771–84.
- Nakashige TG, Zhang B, Krebs C, Nolan EM. Human Calprotectin Is an Iron-Sequestering Host-Defense Protein. Nat Chem Biol. 2015 Oct;11(10):765–71.
- Zackular JP, Chazin WJ, Skaar EP. Nutritional Immunity: S100 Proteins at the Host-Pathogen Interface. J Biol Chem. 2015 July 31;290(31):18991–8.
- Flo TH, Smith KD, Sato S, Rodriguez DJ, Holmes MA, Strong RK, et al. Lipocalin 2 mediates an innate immune response to bacterial infection by sequestrating iron. Nature. 2004 Dec;432(7019):917–21.
- Guo BX, Wang QQ, Li JH, Gan ZS, Zhang XF, Wang YZ, et al. Lipocalin 2 regulates intestine bacterial survival by interplaying with siderophore in a weaned piglet model of Escherichia coli infection. Oncotarget. 2017 June 16;8(39):65386–96.
- Bachman MA, Miller VL, Weiser JN. Mucosal Lipocalin 2 Has Pro-Inflammatory and Iron-Sequestering Effects in Response to Bacterial Enterobactin. PLoS Pathog. 2009 Oct 16;5(10):e1000622.
- Valdebenito M, Müller SI, Hantke K. Special conditions allow binding of the siderophore salmochelin to siderocalin (NGAL-lipocalin). FEMS Microbiol Lett. 2007 Dec 1;277(2):182–7.
- Kehl-Fie TE, Chitayat S, Hood MI, Damo S, Restrepo N, Garcia C, et al. Nutrient Metal Sequestration by Calprotectin Inhibits Bacterial Superoxide Defense, Enhancing Neutrophil Killing of Staphylococcus aureus. Cell Host & Microbe. 2011 Aug 18;10(2):158–64.
- White C, Lee J, Kambe T, Fritsche K, Petris MJ. A Role for the ATP7A Copper-transporting ATPase in Macrophage Bactericidal Activity. J Biol Chem. 2009 Dec 4;284(49):33949–56.
- Stafford SL, Bokil NJ, Achard MES, Kapetanovic R, Schembri MA, McEwan AG, et al. Metal ions in macrophage antimicrobial pathways: emerging roles for zinc and copper. Biosci Rep. 2013 July 16;33(4):e00049.
- White C, Kambe T, Fulcher YG, Sachdev SW, Bush AI, Fritsche K, et al. Copper transport into the secretory pathway is regulated by oxygen in macrophages. J Cell Sci. 2009 May 1;122(9):1315–21.
- Chen L, Shen Q, Liu Y, Zhang Y, Sun L, Ma X, et al. Homeostasis and metabolism of iron and other metal ions in neurodegenerative diseases. Sig Transduct Target Ther. 2025 Feb 3;10(1):31.
- Rosenblum SL. Inflammation, dysregulated iron metabolism, and cardiovascular disease. Front Aging. 2023 Feb 3;4:1124178.
- Pernas L. Cellular metabolism in the defense against microbes. J Cell Sci. 2021 Feb 8;134(5):jcs252023.
- O’Neill LAJ, Kishton RJ, Rathmell J. A guide to immunometabolism for immunologists. Nat Rev Immunol. 2016 Sept;16(9):553–65.
- Nemeth E, Tuttle MS, Powelson J, Vaughn MB, Donovan A, Ward DM, et al. Hepcidin Regulates Cellular Iron Efflux by Binding to Ferroportin and Inducing Its Internalization. Science. 2004 Dec 17;306(5704):2090–3.
- Munn DH, Mellor AL. Indoleamine 2,3-dioxygenase and tumor-induced tolerance. J Clin Invest. 2007 May 1;117(5):1147–54.
- Thomas AC, Mattila JT. “Of Mice and Men”: Arginine Metabolism in Macrophages. Front Immunol [Internet]. 2014 Oct 7 [cited 2025 Aug 24];5. Available from: https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2014.00479/full.
- Kelly B, Pearce EL. Amino Assets: How Amino Acids Support Immunity. Cell Metabolism. 2020 Aug 4;32(2):154–75.
- Kornberg MD. The immunologic Warburg effect: Evidence and therapeutic opportunities in autoimmunity. Wiley Interdiscip Rev Syst Biol Med. 2020;12(5):e1486.
- Kamel M, Aleya S, Alsubih M, Aleya L. Microbiome Dynamics: A Paradigm Shift in Combatting Infectious Diseases. J Pers Med. 2024 Feb 18;14(2):217.
- Eylert E, Schär J, Mertins S, Stoll R, Bacher A, Goebel W, et al. Carbon metabolism of Listeria monocytogenes growing inside macrophages. Mol Microbiol. 2008 Aug;69(4):1008–17.
- Rohmer L, Hocquet D, Miller SI. Are pathogenic bacteria just looking for food? Metabolism and microbial pathogenesis. Trends in Microbiology. 2011 July 1;19(7):341–8.
- Klaenhammer TR, Kleerebezem M, Kopp MV, Rescigno M. The impact of probiotics and prebiotics on the immune system. Nat Rev Immunol. 2012 Oct;12(10):728–34.
- Ji J, Jin W, Liu S, Jiao Z, Li X. Probiotics, prebiotics, and postbiotics in health and disease. MedComm (2020). 2023 Nov 4;4(6):e420.
- Bin BH, Seo J, Kim ST. Function, Structure, and Transport Aspects of ZIP and ZnT Zinc Transporters in Immune Cells. J Immunol Res. 2018 Oct 2;2018:9365747.
- Lichten LA, Cousins RJ. Mammalian zinc transporters: nutritional and physiologic regulation. Annu Rev Nutr. 2009;29:153–76.
- He F, Ru X, Wen T. NRF2, a Transcription Factor for Stress Response and Beyond. Int J Mol Sci. 2020 July 6;21(13):4777.
- Sinclair LV, Rolf J, Emslie E, Shi YB, Taylor PM, Cantrell DA. Control of amino-acid transport by antigen receptors coordinates the metabolic reprogramming essential for T cell differentiation. Nat Immunol. 2013 May;14(5):500–8.
- Munn DH, Mellor AL. Indoleamine 2,3 dioxygenase and metabolic control of immune responses. Trends Immunol. 2013 Mar;34(3):137–43.
- Ullah I, Lang M. Key players in the regulation of iron homeostasis at the host-pathogen interface. Front Immunol [Internet]. 2023 Oct 24 [cited 2025 Sept 13];14. Available from: https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2023.1279826/full.
- Mackenzie EL, Iwasaki K, Tsuji Y. Intracellular Iron Transport and Storage: From Molecular Mechanisms to Health Implications. Antioxid Redox Signal. 2008 June;10(6):997–1030.
- Belinskaia DA, Voronina PA, Goncharov NV. Integrative Role of Albumin: Evolutionary, Biochemical and Pathophysiological Aspects. J Evol Biochem Physiol. 2021;57(6):1419–48.
- Besold AN, Culbertson EM, Nam L, Hobbs RP, Boyko A, Maxwell CN, et al. Antimicrobial action of calprotectin that does not involve metal withholding. Metallomics. 2018 Dec 12;10(12):1728–42.
- Nathan C, Ding A. Nonresolving Inflammation. Cell. 2010 Mar 19;140(6):871–82.
- Tannahill GM, Curtis AM, Adamik J, Palsson-McDermott EM, McGettrick AF, Goel G, et al. Succinate is an inflammatory signal that induces IL-1β through HIF-1α. Nature. 2013 Apr;496(7444):238–42.
- Charlebois E, Pantopoulos K. Nutritional Aspects of Iron in Health and Disease. Nutrients. 2023 Jan;15(11):2441.
- Parrow NL, Fleming RE, Minnick MF. Sequestration and Scavenging of Iron in Infection. Infect Immun. 2013 Oct;81(10):3503–14.
- Schalk IJ. Bacterial siderophores: diversity, uptake pathways and applications. Nat Rev Microbiol. 2025 Jan;23(1):24–40.
- Wiles TJ, Kulesus RR, Mulvey MA. Origins and virulence mechanisms of uropathogenic Escherichia coli. Experimental and Molecular Pathology. 2008 Aug 1;85(1):11–9.
- Hantke K, Nicholson G, Rabsch W, Winkelmann G. Salmochelins, siderophores of Salmonella enterica and uropathogenic Escherichia coli strains, are recognized by the outer membrane receptor IroN. Proceedings of the National Academy of Sciences. 2003 Apr;100(7):3677–82.
- Smith KD. Iron metabolism at the host pathogen interface: lipocalin 2 and the pathogen-associated iroA gene cluster. Int J Biochem Cell Biol. 2007;39(10):1776–80.
- Nairz M, Schroll A, Sonnweber T, Weiss G. The struggle for iron - a metal at the host-pathogen interface. Cell Microbiol. 2010 Dec;12(12):1691–702.
- Drakesmith H, Prentice A. Viral infection and iron metabolism. Nature Reviews Microbiology. 2008 July;6(7):541–52.
- Reid DW, O’May C, Kirov SM, Roddam L, Lamont IL, Sanderson K. Iron chelation directed against biofilms as an adjunct to conventional antibiotics. American Journal of Physiology-Lung Cellular and Molecular Physiology [Internet]. 2009 May 1 [cited 2025 Sept 29]; Available from: https://journals.physiology.org/doi/10.1152/ajplung.00058.2009.
- Weinberg GA. Iron chelators as therapeutic agents against Pneumocystis carinii. Antimicrobial Agents and Chemotherapy [Internet]. 1994 May [cited 2025 Sept 29]; Available from: https://journals.asm.org/doi/10.1128/aac.38.5.997.
- Itoh K, Tsutani H, Mitsuke Y, Iwasaki H. Potential additional effects of iron chelators on antimicrobial- impregnated central venous catheters. Front Microbiol [Internet]. 2023 Aug 7 [cited 2025 Aug 3];14. Available from: https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2023.1210747/full.
- Raymond KN, Allred BE, Sia AK. Coordination Chemistry of Microbial Iron Transport. Acc Chem Res. 2015 Sept 15;48(9):2496–505.
- Baksh KA, Zamble DB. Allosteric control of metal-responsive transcriptional regulators in bacteria. Journal of Biological Chemistry. 2020 Feb 7;295(6):1673–84.
- Wessels I, Maywald M, Rink L. Zinc as a Gatekeeper of Immune Function. Nutrients. 2017 Nov 25;9(12):1286.
- Makthal N, Kumaraswami M. Zinc’ing it out: Zinc homeostasis mechanisms and their impact on the pathogenesis of human pathogen group A streptococcus. Metallomics. 2017 Dec 1;9(12):1693–702.
- Liu M, Sun X, Chen B, Dai R, Xi Z, Xu H. Insights into Manganese Superoxide Dismutase and Human Diseases. Int J Mol Sci. 2022 Dec 14;23(24):15893.
- Salah I, Parkin IP, Allan E. Copper as an antimicrobial agent: recent advances. RSC Adv. 2021 May 19;11(30):18179–86.
- Samanovic MI, Ding C, Thiele DJ, Darwin KH. Copper in Microbial Pathogenesis: Meddling with the Metal. Cell Host & Microbe. 2012 Feb 16;11(2):106–15.
- Rowland JL, Niederweis M. Resistance mechanisms of Mycobacterium tuberculosis against phagosomal copper overload. Tuberculosis (Edinb). 2012 May;92(3):202–10.
- Calvo J, Jung H, Meloni G. Copper metallothioneins. IUBMB Life. 2017 Apr;69(4):236–45.
- Arendsen LP, Thakar R, Sultan AH. The Use of Copper as an Antimicrobial Agent in Health Care, Including Obstetrics and Gynecology. Clin Microbiol Rev. 2019 Aug 14;32(4):e00125-18.
- Lemire JA, Harrison JJ, Turner RJ. Antimicrobial activity of metals: mechanisms, molecular targets and applications. Nat Rev Microbiol. 2013 June;11(6):371–84.
- Fiorentini D, Cappadone C, Farruggia G, Prata C. Magnesium: Biochemistry, Nutrition, Detection, and Social Impact of Diseases Linked to Its Deficiency. Nutrients. 2021 Mar 30;13(4):1136.
- Papp-Wallace KM, Maguire ME. Magnesium Transport and Magnesium Homeostasis. EcoSal Plus. 2008 Sept 25;3(1):10.1128/ecosalplus.5.4.4.2.
- Groisman EA. The Pleiotropic Two-Component Regulatory System PhoP-PhoQ. Journal of Bacteriology. 2001 Mar 15;183(6):1835–42.
- Gunn JS, Ernst RK, McCoy AJ, Miller SI. Constitutive Mutations of the Salmonella entericaSerovar Typhimurium Transcriptional Virulence RegulatorphoP. Infection and Immunity. 2000 June;68(6):3758–62.
- Lensmire JM, Hammer ND. Nutrient sulfur acquisition strategies employed by bacterial pathogens. Current Opinion in Microbiology. 2019 Feb 1;47:52–8.
- Amon J, Titgemeyer F, Burkovski A. Common patterns - unique features: nitrogen metabolism and regulation in Gram-positive bacteria. FEMS Microbiol Rev. 2010 July;34(4):588–605.
- Ryan BE, Mike LA. Arginine at the host-pathogen interface. Infect Immun. 93(8):e00612-24.
- Wang L, Hou Y, Yuan H, Chen H. The role of tryptophan in Chlamydia trachomatis persistence. Front Cell Infect Microbiol. 2022 Aug 2;12:931653.
- Kim G, Weiss SJ, Levine RL. Methionine oxidation and reduction in proteins. Biochimica et Biophysica Acta (BBA) - General Subjects. 2014 Feb 1;1840(2):901–5.
- Kies PJ, Hammer ND. A Resourceful Race: Bacterial Scavenging of Host Sulfur Metabolism during Colonization. Infect Immun. 90(5):e00579-21.
- Kharwar S, Bhattacharjee S, Chakraborty S, Mishra AK. Regulation of sulfur metabolism, homeostasis and adaptive responses to sulfur limitation in cyanobacteria. Biologia. 2021 Oct 1;76(10):2811–35.
- Tardy AL, Pouteau E, Marquez D, Yilmaz C, Scholey A. Vitamins and Minerals for Energy, Fatigue and Cognition: A Narrative Review of the Biochemical and Clinical Evidence. Nutrients. 2020 Jan 16;12(1):228.
- Hanna M, Jaqua E, Nguyen V, Clay J. B Vitamins: Functions and Uses in Medicine. Perm J. 26(2):89–97.
- Pham VT, Dold S, Rehman A, Bird JK, Steinert RE. Vitamins, the gut microbiome and gastrointestinal health in humans. Nutrition Research. 2021 Nov 1;95:35–53.
- Schröder SK, Gasterich N, Weiskirchen S, Weiskirchen R. Lipocalin 2 receptors: facts, fictions, and myths. Front Immunol [Internet]. 2023 Aug 11 [cited 2025 Aug 27];14. Available from: https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2023.1229885/full.
- Rieder FJ, Gröschel C, Kastner MT, Kosulin K, Laengle J, Zadnikar R, et al. Human Cytomegalovirus Infection Downregulates Vitamin-D Receptor in Mammalian Cells. J Steroid Biochem Mol Biol. 2017 Jan;165(Pt B):356–62.
- Roth JR, Lawrence JG, Rubenfield M, Kieffer-Higgins S, Church GM. Characterization of the cobalamin (vitamin B12) biosynthetic genes of Salmonella typhimurium. J Bacteriol. 1993 June;175(11):3303–16.
- Minato Y, Thiede JM, Kordus SL, McKlveen EJ, Turman BJ, Baughn AD. Mycobacterium tuberculosis Folate Metabolism and the Mechanistic Basis for para-Aminosalicylic Acid Susceptibility and Resistance. Antimicrobial Agents and Chemotherapy [Internet]. 2015 June 1 [cited 2025 Aug 1]; Available from: https://journals.asm.org/doi/10.1128/aac.00647-15.
- Mayer FL, Wilson D, Hube B. Candida albicans pathogenicity mechanisms. Virulence. 2013 Feb 15;4(2):119–28.
- Perfect JR, Kronstad JW. Cryptococcal nutrient acquisition and pathogenesis: dining on the host. Microbiology and Molecular Biology Reviews. 2025 Feb 10;89(1):e00015-23.
- Passalacqua KD, Charbonneau ME, O’Riordan MXD. Bacterial Metabolism Shapes the Host–Pathogen Interface. Microbiology Spectrum. 2016 May 13;4(3):10.1128/microbiolspec.vmbf-0027–2015.
- Bhagwat A, Haldar T, Kanojiya P, Saroj SD. Bacterial metabolism in the host and its association with virulence. Virulence. 16(1):2459336.
- Eisenreich W, Heesemann J, Rudel T, Goebel W. Metabolic host responses to infection by intracellular bacterial pathogens. Front Cell Infect Microbiol [Internet]. 2013 July 9 [cited 2025 Aug 27];3. Available from: https://www.frontiersin.org/journals/cellular-and-infection-microbiology/articles/10.3389/fcimb.2013.00024/full.
- Tang J, Wang X, Chen S, Chang T, Gu Y, Zhang F, et al. Disruption of glucose homeostasis by bacterial infection orchestrates host innate immunity through NAD+/NADH balance. Cell Reports. 2024 Sept 24;43(9):114648.
- Ling ZN, Jiang YF, Ru JN, Lu JH, Ding B, Wu J. Amino acid metabolism in health and disease. Sig Transduct Target Ther. 2023 Sept 13;8(1):345.
- Prentice AM, Ghattas H, Cox SE. Host-Pathogen Interactions: Can Micronutrients Tip the Balance?123. The Journal of Nutrition. 2007 May 1;137(5):1334–7.
- Perkins-Balding D, Ratliff-Griffin M, Stojiljkovic I. Iron Transport Systems in Neisseria meningitidis. Microbiol Mol Biol Rev. 2004 Mar;68(1):154–71.
- Cornelissen CN, Sparling PF. Iron piracy: acquisition of transferrin-bound iron by bacterial pathogens. Mol Microbiol. 1994 Dec;14(5):843–50.
- Litt DJ, Palmer HM, Borriello SP. Neisseria meningitidis Expressing Transferrin Binding Proteins of Actinobacillus pleuropneumoniae Can Utilize Porcine Transferrin for Growth. Infect Immun. 2000 Feb;68(2):550–7.
- Andre GO, Converso TR, Politano WR, Ferraz LFC, Ribeiro ML, Leite LCC, et al. Role of Streptococcus pneumoniae Proteins in Evasion of Complement-Mediated Immunity. Front Microbiol [Internet]. 2017 Feb 20 [cited 2025 Aug 29];8. Available from: https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2017.00224/full.
- Tu AHT, Fulgham RL, McCrory MA, Briles DE, Szalai AJ. Pneumococcal Surface Protein A Inhibits Complement Activation by Streptococcus pneumoniae. Infection and Immunity. 1999 Sept;67(9):4720–4.
- Brown JS, Gilliland SM, Holden DW. A Streptococcus pneumoniae pathogenicity island encoding an ABC transporter involved in iron uptake and virulence. Molecular Microbiology. 2001;40(3):572–85.
- Tiedemann MT, Muryoi N, Heinrichs DE, Stillman MJ. Iron acquisition by the haem-binding Isd proteins in Staphylococcus aureus: studies of the mechanism using magnetic circular dichroism. Biochem Soc Trans. 2008 Nov 19;36(6):1138–43.
- Ghssein G, Ezzeddine Z. A Review of Pseudomonas aeruginosa Metallophores: Pyoverdine, Pyochelin and Pseudopaline. Biology (Basel). 2022 Nov 25;11(12):1711.
- Cornelis P, Tahrioui A, Lesouhaitier O, Bouffartigues E, Feuilloley M, Baysse C, et al. High affinity iron uptake by pyoverdine in Pseudomonas aeruginosa involves multiple regulators besides Fur, PvdS, and FpvI. Biometals. 2023 Apr 1;36(2):255–61.
- Robinson AE, Heffernan JR, Henderson JP. The iron hand of uropathogenic Escherichia coli: the role of transition metal control in virulence. Future Microbiol. 2018 June;13(7):745–56.
- Garcia EC, Brumbaugh AR, Mobley HLT. Redundancy and specificity of Escherichia coli iron acquisition systems during urinary tract infection. Infect Immun. 2011 Mar;79(3):1225–35.
- Thakur A, Mikkelsen H, Jungersen G. Intracellular Pathogens: Host Immunity and Microbial Persistence Strategies. J Immunol Res. 2019 Apr 14;2019:1356540.
- Clemens DL, Lee BY, Horwitz MA. The Mycobacterium tuberculosis Phagosome in Human Macrophages Is Isolated from the Host Cell Cytoplasm. Infect Immun. 2002 Oct;70(10):5800–7.
- Sheldon JR, Skaar EP. Metals as phagocyte antimicrobial effectors. Curr Opin Immunol. 2019 Oct;60:1–9.
- Zhang L, Kent JE, Whitaker M, Young DC, Herrmann D, Aleshin AE, et al. A periplasmic cinched protein is required for siderophore secretion and virulence of Mycobacterium tuberculosis. Nat Commun. 2022 Apr 26;13:2255.
- Pandey AK, Sassetti CM. Mycobacterial persistence requires the utilization of host cholesterol. Proceedings of the National Academy of Sciences. 2008 Mar 18;105(11):4376–80.
- Zhao H, Zhang X, Zhang N, Zhu L, Lian H. The interplay between Salmonella and host: Mechanisms and strategies for bacterial survival. Cell Insight. 2025 Apr 1;4(2):100237.
- Chamnongpol S, Cromie M, Groisman EA. Mg2+ sensing by the Mg2+ sensor PhoQ of Salmonella enterica. J Mol Biol. 2003 Jan 24;325(4):795–807.
- Ikeda JS, Janakiraman A, Kehres DG, Maguire ME, Slauch JM. Transcriptional Regulation of sitABCD of Salmonella enterica Serovar Typhimurium by MntR and Fur. J Bacteriol. 2005 Feb;187(3):912–22.
- Ammendola S, Pasquali P, Pistoia C, Petrucci P, Petrarca P, Rotilio G, et al. High-Affinity Zn2+ Uptake System ZnuABC Is Required for Bacterial Zinc Homeostasis in Intracellular Environments and Contributes to the Virulence of Salmonella enterica. Infection and Immunity. 2007 Dec;75(12):5867–76.
- Mordue DG, Håkansson S, Niesman I, David Sibley L. Toxoplasma gondii Resides in a Vacuole That Avoids Fusion with Host Cell Endocytic and Exocytic Vesicular Trafficking Pathways. Experimental Parasitology. 1999 June 1;92(2):87–99.
- Blader IJ, Koshy AA. Toxoplasma gondii Development of Its Replicative Niche: in Its Host Cell and Beyond. Eukaryotic Cell. 2014 July 30;13(8):965–76.
- Fan YM, Zhang QQ, Pan M, Hou ZF, Fu L, Xu X, et al. Toxoplasma gondii sustains survival by regulating cholesterol biosynthesis and uptake via SREBP2 activation. J Lipid Res. 2024 Oct 28;65(12):100684.
- Skariah S, McIntyre MK, Mordue DG. Toxoplasma gondii: determinants of tachyzoite to bradyzoite conversion. Parasitology Research. 2010 June 1;107(2):253.
- Sandoz KM, Beare PA, Cockrell DC, Heinzen RA. Complementation of Arginine Auxotrophy for Genetic Transformation of Coxiella burnetii by Use of a Defined Axenic Medium. Applied and Environmental Microbiology. 2016 May 15;82(10):3042–51.
- Liu J, Istvan ES, Gluzman IY, Gross J, Goldberg DE. Plasmodium falciparum ensures its amino acid supply with multiple acquisition pathways and redundant proteolytic enzyme systems. Proceedings of the National Academy of Sciences. 2006 June 6;103(23):8840–5.
- Counihan NA, Modak JK, de Koning-Ward TF. How Malaria Parasites Acquire Nutrients From Their Host. Front Cell Dev Biol. 2021 Mar 25;9:649184.
- Prentice AM, Ghattas H, Doherty C, Cox SE. Iron metabolism and malaria. Food Nutr Bull. 2007 Dec;28(4 Suppl):S524-539.
- Huynh C, Sacks DL, Andrews NW. A Leishmania amazonensis ZIP family iron transporter is essential for parasite replication within macrophage phagolysosomes. J Exp Med. 2006 Sept 25;203(10):2363–75.
- Chandra S, Ruhela D, Deb A, Vishwakarma RA. Glycobiology of the Leishmania parasite and emerging targets for antileishmanial drug discovery. Expert Opinion on Therapeutic Targets. 2010 July 1;14(7):739–57.
- Almeida RS, Brunke S, Albrecht A, Thewes S, Laue M, Jr JEE, et al. The Hyphal-Associated Adhesin and Invasin Als3 of Candida albicans Mediates Iron Acquisition from Host Ferritin. PLOS Pathogens. 2008 Nov 21;4(11):e1000217.
- Jung WH, Kronstad JW. Iron and fungal pathogenesis: a case study with Cryptococcus neoformans. Cell Microbiol. 2008 Feb;10(2):277–84.
- Hissen AHT, Wan ANC, Warwas ML, Pinto LJ, Moore MM. The Aspergillus fumigatus Siderophore Biosynthetic Gene sidA, Encoding l-Ornithine N5-Oxygenase, Is Required for Virulence. Infection and Immunity. 2005 Sept;73(9):5493–503.
- Deitsch KW, Lukehart SA, Stringer JR. Common strategies for antigenic variation by bacterial, fungal and protozoan pathogens. Nat Rev Microbiol. 2009 July;7(7):493–503.
- Maizels RM. Parasite immunomodulation and polymorphisms of the immune system. J Biol. 2009;8(7):62.
- Miethke M, Marahiel MA. Siderophore-Based Iron Acquisition and Pathogen Control. Microbiol Mol Biol Rev. 2007 Sept;71(3):413–51.
- Andersson SGE, Zomorodipour A, Andersson JO, Sicheritz-Pontén T, Alsmark UCM, Podowski RM, et al. The genome sequence of Rickettsia prowazekii and the origin of mitochondria. Nature. 1998 Nov;396(6707):133–40.
- Stephens RS, Kalman S, Lammel C, Fan J, Marathe R, Aravind L, et al. Genome Sequence of an Obligate Intracellular Pathogen of Humans: Chlamydia trachomatis. Science. 1998 Oct 23;282(5389):754–9.
- Taylor SJ, Winter SE. Salmonella finds a way: Metabolic versatility of Salmonella enterica serovar Typhimurium in diverse host environments. PLOS Pathogens. 2020 June 11;16(6):e1008540.
- Templeton TJ. The varieties of gene amplification, diversification and hypervariability in the human malaria parasite, Plasmodium falciparum. Molecular and Biochemical Parasitology. 2009 Aug 1;166(2):109–16.
- Dunn MJ, Kinney GM, Washington PM, Berman J, Anderson MZ. Functional diversification accompanies gene family expansion of MED2 homologs in Candida albicans. PLOS Genetics. 2018 Apr 9;14(4):e1007326.
- Martin RE, Henry RI, Abbey JL, Clements JD, Kirk K. The “permeome” of the malaria parasite: an overview of the membrane transport proteins of Plasmodium falciparum. Genome Biology. 2005 Mar 2;6(3):R26.
- Haas H. Fungal siderophore metabolism with a focus on Aspergillus fumigatus. Nat Prod Rep. 2014 Sept 11;31(10):1266–76.
- Goyal A. Horizontal gene transfer drives the evolution of dependencies in bacteria. iScience. 2022 Apr 27;25(5):104312.
- Kramer J, Özkaya Ö, Kümmerli R. Bacterial siderophores in community and host interactions. Nat Rev Microbiol. 2020 Mar;18(3):152–63.
- Lin H, Fischbach MA, Liu DR, Walsh CT. In Vitro Characterization of Salmochelin and Enterobactin Trilactone Hydrolases IroD, IroE, and Fes. J Am Chem Soc. 2005 Aug 1;127(31):11075–84.
- Price SL, Vadyvaloo V, DeMarco JK, Brady A, Gray PA, Kehl-Fie TE, et al. Yersiniabactin contributes to overcoming zinc restriction during Yersinia pestis infection of mammalian and insect hosts. Proceedings of the National Academy of Sciences. 2021 Nov 2;118(44):e2104073118.
- Katumba GL, Tran H, Henderson JP. The Yersinia High-Pathogenicity Island Encodes a Siderophore-Dependent Copper Response System in Uropathogenic Escherichia coli. mBio. 13(1):e02391-21.
- Leon-Sicairos N, Reyes-Cortes R, Guadrón-Llanos AM, Madueña-Molina J, Leon-Sicairos C, Canizalez-Román A. Strategies of Intracellular Pathogens for Obtaining Iron from the Environment. BioMed Research International. 2015;2015(1):476534.
- Gu Y, Liu Y, Mao W, Peng Y, Han X, Jin H, et al. Functional versatility of Zur in metal homeostasis, motility, biofilm formation, and stress resistance in Yersinia pseudotuberculosis. Microbiol Spectr. 2024 May 2;12(5):e0375623.
- Cai R, Gao F, Pan J, Hao X, Yu Z, Qu Y, et al. The transcriptional regulator Zur regulates the expression of ZnuABC and T6SS4 in response to stresses in Yersinia pseudotuberculosis. Microbiological Research. 2021 Aug 1;249:126787.
- Bosma EF, Rau MH, van Gijtenbeek LA, Siedler S. Regulation and distinct physiological roles of manganese in bacteria. FEMS Microbiol Rev. 2021 Nov 1;45(6):fuab028.
- Waters LS, Sandoval M, Storz G. The Escherichia coli MntR Miniregulon Includes Genes Encoding a Small Protein and an Efflux Pump Required for Manganese Homeostasis ▿. J Bacteriol. 2011 Nov;193(21):5887–97.
- Cassat JE, Skaar EP. Metal ion acquisition in Staphylococcus aureus: overcoming nutritional immunity. Semin Immunopathol. 2012 Mar 1;34(2):215–35.
- Kehl-Fie TE, Zhang Y, Moore JL, Farrand AJ, Hood MI, Rathi S, et al. MntABC and MntH Contribute to Systemic Staphylococcus aureus Infection by Competing with Calprotectin for Nutrient Manganese. Infection and Immunity. 2013 Aug 13;81(9):3395–405.
- Zaharik ML, Cullen VL, Fung AM, Libby SJ, Kujat Choy SL, Coburn B, et al. The Salmonella enterica Serovar Typhimurium Divalent Cation Transport Systems MntH and SitABCD Are Essential for Virulence in an Nramp1G169 Murine Typhoid Model. Infection and Immunity. 2004 Sept;72(9):5522–5.
- Campoy S, Jara M, Busquets N, Pérez de Rozas AM, Badiola I, Barbé J. Role of the High-Affinity Zinc Uptake znuABC System in Salmonella enterica Serovar Typhimurium Virulence. Infection and Immunity. 2002 Aug;70(8):4721–5.
- Nair A, Sarma SJ. The impact of carbon and nitrogen catabolite repression in microorganisms. Microbiological Research. 2021 Oct 1;251:126831.
- Joseph B, Mertins S, Stoll R, Schär J, Umesha KR, Luo Q, et al. Glycerol Metabolism and PrfA Activity in Listeria monocytogenes. Journal of Bacteriology. 2008 Aug;190(15):5412–30.
- Freeman MJ, Eral NJ, Sauer JD. Listeria monocytogenes requires phosphotransferase systems to facilitate intracellular growth and virulence. PLOS Pathogens. 2025 Apr 15;21(4):e1012492.
- Zhang YJ, Rubin EJ. Feast or famine: the host–pathogen battle over amino acids. Cell Microbiol. 2013 July;15(7):1079–87.
- Ren W, Rajendran R, Zhao Y, Tan B, Wu G, Bazer FW, et al. Amino Acids As Mediators of Metabolic Cross Talk between Host and Pathogen. Front Immunol [Internet]. 2018 Feb 27 [cited 2025 Aug 30];9. Available from: https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2018.00319/full.
- Borah K, Beyß M, Theorell A, Wu H, Basu P, Mendum TA, et al. Intracellular Mycobacterium tuberculosis Exploits Multiple Host Nitrogen Sources during Growth in Human Macrophages. Cell Rep. 2019 Dec 10;29(11):3580-3591.e4.
- Liu Y, Zhang Q, Hu M, Yu K, Fu J, Zhou F, et al. Proteomic Analyses of Intracellular Salmonella enterica Serovar Typhimurium Reveal Extensive Bacterial Adaptations to Infected Host Epithelial Cells. Infection and Immunity [Internet]. 2015 May 4 [cited 2025 Sept 16]; Available from: https://journals.asm.org/doi/10.1128/iai.02882-14.
- Coppens I, Dunn JD, Romano JD, Pypaert M, Zhang H, Boothroyd JC, et al. Toxoplasma gondii Sequesters Lysosomes from Mammalian Hosts in the Vacuolar Space. Cell. 2006 Apr 21;125(2):261–74.
- Wang T, Wang C, Li C, Song L. The intricate dance: host autophagy and Coxiella burnetii infection. Front Microbiol. 2023 Sept 22;14:1281303.
- McConville MJ, Handman E. The molecular basis of Leishmania pathogenesis. International Journal for Parasitology. 2007 Aug 1;37(10):1047–51.
- Counihan NA, Chisholm SA, Bullen HE, Srivastava A, Sanders PR, Jonsdottir TK, et al. Plasmodium falciparum parasites deploy RhopH2 into the host erythrocyte to obtain nutrients, grow and replicate. Soldati-Favre D, editor. eLife. 2017 Mar 2;6:e23217.
- Martin RE. The transportome of the malaria parasite. Biol Rev Camb Philos Soc. 2020 Apr;95(2):305–32.
- Sorci G, Cornet S, Faivre B. Immune Evasion, Immunopathology and the Regulation of the Immune System. Pathogens. 2013 Mar;2(1):71–91.
- Stanley SA, Raghavan S, Hwang WW, Cox JS. Acute infection and macrophage subversion by Mycobacterium tuberculosis require a specialized secretion system. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):13001–6.
- Lažetić V, Troemel ER. Conservation lost: host-pathogen battles drive diversification and expansion of gene families. FEBS J. 2021 Sept;288(18):5289–99.
- Moon YH, Tanabe T, Funahashi T, Shiuchi K ichi, Nakao H, Yamamoto S. Identification and characterization of two contiguous operons required for aerobactin transport and biosynthesis in Vibrio mimicus. Microbiol Immunol. 2004;48(5):389–98.
- Stork M, Di Lorenzo M, Welch TJ, Crosa JH. Transcription termination within the iron transport-biosynthesis operon of Vibrio anguillarum requires an antisense RNA. J Bacteriol. 2007 May;189(9):3479–88.
- Lynch D, O’Brien J, Welch T, Clarke P, Cuív PO, Crosa JH, et al. Genetic organization of the region encoding regulation, biosynthesis, and transport of rhizobactin 1021, a siderophore produced by Sinorhizobium meliloti. J Bacteriol. 2001 Apr;183(8):2576–85.
- Stintzi A, Johnson Z, Stonehouse M, Ochsner U, Meyer JM, Vasil ML, et al. The pvc Gene Cluster of Pseudomonas aeruginosa: Role in Synthesis of the Pyoverdine Chromophore and Regulation by PtxR and PvdS. J Bacteriol. 1999 July;181(13):4118–24.
- Patzer SI, Hantke K. The ZnuABC high-affinity zinc uptake system and its regulator Zur in Escherichia coli. Mol Microbiol. 1998 June;28(6):1199–210.
- Kondrashov FA. Gene duplication as a mechanism of genomic adaptation to a changing environment. Proc Biol Sci. 2012 Dec 22;279(1749):5048–57.
- Schmidt H, Hensel M. Pathogenicity Islands in Bacterial Pathogenesis. Clin Microbiol Rev. 2004 Jan;17(1):14–56.
- Carniel E. The Yersinia high-pathogenicity island: an iron-uptake island. Microbes and Infection. 2001 June 1;3(7):561–9.
- Zomorodipour A, Andersson SGE. Obligate intracellular parasites: Rickettsia prowazekii and Chlamydia trachomatis. FEBS Letters. 1999 June 4;452(1):11–5.
- Ghigo E, Colombo MI, Heinzen RA. The Coxiella burnetii Parasitophorous Vacuole. In: Toman R, Heinzen RA, Samuel JE, Mege JL, editors. Coxiella burnetii: Recent Advances and New Perspectives in Research of the Q Fever Bacterium [Internet]. Dordrecht: Springer Netherlands; 2012 [cited 2025 Sept 16]. p. 141–69. Available from: https://doi.org/10.1007/978-94-007-4315-1_8. [CrossRef]
- Puri S, Kumar R, Rojas IG, Salvatori O, Edgerton M. Iron Chelator Deferasirox Reduces Candida albicans Invasion of Oral Epithelial Cells and Infection Levels in Murine Oropharyngeal Candidiasis. Antimicrob Agents Chemother. 2019 Apr;63(4):e02152-18.
- Ni T, Chi X, Wu H, Xie F, Bao J, Wang J, et al. Design, synthesis and evaluation of novel deferasirox derivatives with high antifungal potency in vitro and in vivo. European Journal of Medicinal Chemistry. 2024 Jan 15;264:116026.
- Feizi S, Awad M, Ramezanpour M, Cooksley C, Murphy W, Prestidge CA, et al. Promoting the Efficacy of Deferiprone-Gallium-Protoporphyrin (IX) against Mycobacterium abscessus Intracellular Infection with Lipid Liquid Crystalline Nanoparticles. ACS Appl Mater Interfaces. 2024 Dec 25;16(51):70274–83.
- Feizi S, Awad M, Nepal R, Cooksley CM, Psaltis AJ, Wormald PJ, et al. Deferiprone-gallium-protoporphyrin (IX): A promising treatment modality against Mycobacterium abscessus. Tuberculosis. 2023 Sept 1;142:102390.
- Tarnow-Mordi WO, Abdel-Latif ME, Martin A, Pammi M, Robledo K, Manzoni P, et al. The effect of lactoferrin supplementation on death or major morbidity in very low birthweight infants (LIFT): a multicentre, double-blind, randomised controlled trial. The Lancet Child & Adolescent Health. 2020 June 1;4(6):444–54.
- Pammi M, Suresh G. Enteral lactoferrin supplementation for prevention of sepsis and necrotizing enterocolitis in preterm infants. Cochrane Database Syst Rev. 2017 June 28;2017(6):CD007137.
- Kadiyala U, Turali-Emre ES, Bahng JH, Kotov NA, VanEpps JS. Unexpected insights into antibacterial activity of zinc oxide nanoparticles against methicillin resistant Staphylococcus aureus (MRSA). Nanoscale. 2018 Mar 8;10(10):4927–39.
- Caron AJ, Ali IJ, Delgado MJ, Johnson D, Reeks JM, Strzhemechny YM, et al. Zinc oxide nanoparticles mediate bacterial toxicity in Mueller-Hinton Broth via Zn2+. Front Microbiol [Internet]. 2024 Apr 22 [cited 2025 Sept 11];15. Available from: https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2024.1394078/full.
- Mendes CR, Dilarri G, Forsan CF, Sapata V de MR, Lopes PRM, de Moraes PB, et al. Antibacterial action and target mechanisms of zinc oxide nanoparticles against bacterial pathogens. Sci Rep. 2022 Feb 16;12(1):2658.
- O’Brien H, Davoodian T, Johnson MDL. The promise of copper ionophores as antimicrobials. Current Opinion in Microbiology. 2023 Oct 1;75:102355.
- Dalecki AG, Haeili M, Shah S, Speer A, Niederweis M, Kutsch O, et al. Disulfiram and Copper Ions Kill Mycobacterium tuberculosis in a Synergistic Manner. Antimicrob Agents Chemother. 2015 Aug;59(8):4835–44.
- Ngwane AH, Petersen RD, Baker B, Wiid I, Wong HN, Haynes RK. The evaluation of the anti-cancer drug elesclomol that forms a redox-active copper chelate as a potential anti-tubercular drug. IUBMB Life. 2019 May;71(5):532–8.
- Goss CH, Kaneko Y, Khuu L, Anderson GD, Ravishankar S, Aitken ML, et al. Gallium disrupts bacterial iron metabolism and has therapeutic effects in mice and humans with lung infections. Science Translational Medicine. 2018 Sept 26;10(460):eaat7520.
- Chan YR, Liu JS, Pociask DA, Zheng M, Mietzner TA, Berger T, et al. Lipocalin 2 Is Required for Pulmonary Host Defense against Klebsiella Infection. J Immunol. 2009 Apr 15;182(8):4947–56.
- Wang Q, Li S, Tang X, Liang L, Wang F, Du H. Lipocalin 2 Protects Against Escherichia coli Infection by Modulating Neutrophil and Macrophage Function. Front Immunol [Internet]. 2019 Nov 8 [cited 2025 Sept 11];10. Available from: https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2019.02594/full.
- Lin YM, Ghosh M, Miller PA, Möllmann U, Miller MJ. Synthetic sideromycins (skepticism and optimism): selective generation of either broad or narrow spectrum Gram-negative antibiotics. Biometals. 2019 June;32(3):425–51.
- Page MGP, Dantier C, Desarbre E. In vitro properties of BAL30072, a novel siderophore sulfactam with activity against multiresistant gram-negative bacilli. Antimicrob Agents Chemother. 2010 June;54(6):2291–302.
- Straubinger M, Blenk H, Naber KG, Wagenlehner FME. Urinary Concentrations and Antibacterial Activity of BAL30072, a Novel Siderophore Monosulfactam, against Uropathogens after Intravenous Administration in Healthy Subjects. Antimicrobial Agents and Chemotherapy. 2016 May 23;60(6):3309–15.
- Selvaraj P, Harishankar M, Afsal K. Vitamin D: Immuno-modulation and tuberculosis treatment. Can J Physiol Pharmacol. 2015 May;93(5):377–84.
- Coussens AK, Wilkinson RJ, Hanifa Y, Nikolayevskyy V, Elkington PT, Islam K, et al. Vitamin D accelerates resolution of inflammatory responses during tuberculosis treatment. Proceedings of the National Academy of Sciences. 2012 Sept 18;109(38):15449–54.
- Liu Q, Yang Z, Miao Y, Liu X, Peng J, Wei H. Effects of methionine restriction and methionine hydroxy analogs on intestinal inflammation and physical barrier function in mice. Journal of Future Foods. 2025 Jan 1;5(1):68–78.
- Schnadower D, Tarr PI, Casper TC, Gorelick MH, Dean JM, O’Connell KJ, et al. Lactobacillus rhamnosus GG versus Placebo for Acute Gastroenteritis in Children. New England Journal of Medicine. 2018 Nov 22;379(21):2002–14.
- PiLeJe. Effect and Tolerability of Lactobacillus Rhamnosus GG LA801 for the Preventive Nutritional Care of Nosocomial Diarrhea in Children [Internet]. clinicaltrials.gov; 2021 Apr [cited 2025 Sept 12]. Report No.: NCT04628819. Available from: https://clinicaltrials.gov/study/NCT04628819.
- Vazquez-Gutierrez P, de Wouters T, Werder J, Chassard C, Lacroix C. High Iron-Sequestrating Bifidobacteria Inhibit Enteropathogen Growth and Adhesion to Intestinal Epithelial Cells In vitro. Front Microbiol. 2016 Sept 22;7:1480.
- Sonnenborn U, Schulze J. The non-pathogenic Escherichia coli strain Nissle 1917 – features of a versatile probiotic. Microbial Ecology in Health and Disease. 2009 Jan 1;21(3–4):122–58.
- Helmy YA, Closs G, Jung K, Kathayat D, Vlasova A, Rajashekara G. Effect of Probiotic E. coli Nissle 1917 Supplementation on the Growth Performance, Immune Responses, Intestinal Morphology, and Gut Microbes of Campylobacter jejuni Infected Chickens. Infection and Immunity. 2022 Sept 22;90(10):e00337-22.
- Vaughn BP, Fischer M, Kelly CR, Allegretti JR, Graiziger C, Thomas J, et al. Effectiveness and Safety of Colonic and Capsule Fecal Microbiota Transplantation for Recurrent Clostridioides difficile Infection. Clinical Gastroenterology and Hepatology. 2023 May 1;21(5):1330-1337.e2.
- Tariq R, Pardi DS, Khanna S. Resolution rates in clinical trials for microbiota restoration for recurrent Clostridioides difficile infection: an updated systematic review and meta-analysis. Therap Adv Gastroenterol. 2023 May 30;16:17562848231174293.
- Wilcox MH, McGovern BH, Hecht GA. The Efficacy and Safety of Fecal Microbiota Transplant for Recurrent Clostridiumdifficile Infection: Current Understanding and Gap Analysis. Open Forum Infect Dis [Internet]. 2020 May 1 [cited 2025 Sept 12];7(5). Available from: https://dx.doi.org/10.1093/ofid/ofaa114. [CrossRef]
- Kelly CR, Yen EF, Grinspan AM, Kahn SA, Atreja A, Lewis JD, et al. Fecal Microbiota Transplantation Is Highly Effective in Real-World Practice: Initial Results From the FMT National Registry. Gastroenterology. 2021 Jan;160(1):183-192.e3.
- Shin J, Lee JH, Park SH, Cha B, Kwon KS, Kim H, et al. Efficacy and Safety of Fecal Microbiota Transplantation for Clearance of Multidrug-Resistant Organisms under Multiple Comorbidities: A Prospective Comparative Trial. Biomedicines. 2022 Sept 26;10(10):2404.
- Woodworth MH, Babiker A, Prakash-Asrani R, Mehta CC, Steed DB, Ashley A, et al. Microbiota Transplantation Among Patients Receiving Long-Term Care: The Sentinel REACT Nonrandomized Clinical Trial. JAMA Netw Open. 2025 July 1;8(7):e2522740–e2522740.
- Macareño-Castro J, Solano-Salazar A, Dong LT, Mohiuddin M, Espinoza JL. Fecal microbiota transplantation for Carbapenem-Resistant Enterobacteriaceae: A systematic review. Journal of Infection. 2022 June 1;84(6):749–59.
- Singh R, de Groot PF, Geerlings SE, Hodiamont CJ, Belzer C, Berge IJM ten, et al. Fecal microbiota transplantation against intestinal colonization by extended spectrum beta-lactamase producing Enterobacteriaceae: a proof of principle study. BMC Research Notes. 2018 Mar 22;11(1):190.
- Woodworth MH, Hayden MK, Young VB, Kwon JH. The Role of Fecal Microbiota Transplantation in Reducing Intestinal Colonization With Antibiotic-Resistant Organisms: The Current Landscape and Future Directions. Open Forum Infect Dis [Internet]. 2019 July 1 [cited 2025 Sept 12];6(7). Available from: https://dx.doi.org/10.1093/ofid/ofz288. [CrossRef]
- Crum-Cianflone NF, Sullivan E, Ballon-Landa G. Fecal Microbiota Transplantation and Successful Resolution of Multidrug-Resistant-Organism Colonization. Journal of Clinical Microbiology [Internet]. 2015 Apr 15 [cited 2025 Sept 12]; Available from: https://journals.asm.org/doi/10.1128/jcm.00820-15.
- Keskey R, Cone JT, DeFazio JR, Alverdy JC. The use of fecal microbiota transplant in sepsis. Translational Research. 2020 Dec 1;226:12–25.
- Kim SM, DeFazio JR, Hyoju SK, Sangani K, Keskey R, Krezalek MA, et al. Fecal microbiota transplant rescues mice from human pathogen mediated sepsis by restoring systemic immunity. Nat Commun. 2020 May 11;11(1):2354.
- Barua N, Buragohain AK. Therapeutic Potential of Curcumin as an Antimycobacterial Agent. Biomolecules. 2021 Sept;11(9):1278.
- Fatima S, Kumari A, Dwivedi VP. Advances in adjunct therapy against tuberculosis: Deciphering the emerging role of phytochemicals. MedComm (2020). 2021 Aug 5;2(4):494–513.
- Nakamura M, Urakawa D, He Z, Akagi I, Hou DX, Sakao K. Apoptosis Induction in HepG2 and HCT116 Cells by a Novel Quercetin-Zinc (II) Complex: Enhanced Absorption of Quercetin and Zinc (II). International Journal of Molecular Sciences. 2023 Jan;24(24):17457.
- Jeon J, Kim JH, Lee CK, Oh CH, Song HJ. The Antimicrobial Activity of (-)-Epigallocatehin-3-Gallate and Green Tea Extracts against Pseudomonas aeruginosa and Escherichia coli Isolated from Skin Wounds. Ann Dermatol. 2014 Oct;26(5):564–9.
- DeDiego ML, Portilla Y, Daviu N, López-García D, Villamayor L, Mulens-Arias V, et al. Iron oxide and iron oxyhydroxide nanoparticles impair SARS-CoV-2 infection of cultured cells. Journal of Nanobiotechnology. 2022 July 30;20(1):352.
- Lu Z, Yu D, Nie F, Wang Y, Chong Y. Iron Nanoparticles Open Up New Directions for Promoting Healing in Chronic Wounds in the Context of Bacterial Infection. Pharmaceutics. 2023 Sept 15;15(9):2327.
- Ikhazuagbe IH, Ofoka EA, Odofin OL, Erumiseli O, Edoka OE, Ezennubia KP, et al. Antibacterial activity and mechanistic insights of gallium-based nanoparticles: an emerging frontier in metal-based antimicrobials. RSC Adv. 2025 Aug 29;15(38):31122–53.
- Ramesh G, Kaviyil JE, Paul W, Sasi R, Joseph R. Gallium–Curcumin Nanoparticle Conjugates as an Antibacterial Agent against Pseudomonas aeruginosa: Synthesis and Characterization. ACS Omega [Internet]. 2022 Feb 17 [cited 2025 Sept 12]; Available from: https://pubs.acs.org/doi/full/10.1021/acsomega.1c06398.
- Paladini F, Pollini M. Antimicrobial Silver Nanoparticles for Wound Healing Application: Progress and Future Trends. Materials. 2019 Jan;12(16):2540.
- Holubnycha V, Husak Y, Korniienko V, Bolshanina S, Tveresovska O, Myronov P, et al. Antimicrobial Activity of Two Different Types of Silver Nanoparticles against Wide Range of Pathogenic Bacteria. Nanomaterials. 2024 Jan;14(2):137.
- Baveloni FG, Meneguin AB, Sábio RM, Camargo BAF de, Trevisan DPV, Duarte JL, et al. Antimicrobial effect of silver nanoparticles as a potential healing treatment for wounds contaminated with Staphylococcus aureus in wistar rats. Journal of Drug Delivery Science and Technology. 2025 Jan 1;103:106445.
- Khalifa HO, Oreiby A, Mohammed T, Abdelhamid MAA, Sholkamy EN, Hashem H, et al. Silver nanoparticles as next-generation antimicrobial agents: mechanisms, challenges, and innovations against multidrug-resistant bacteria. Front Cell Infect Microbiol [Internet]. 2025 Aug 14 [cited 2025 Sept 12];15. Available from: https://www.frontiersin.org/journals/cellular-and-infection-microbiology/articles/10.3389/fcimb.2025.1599113/full.
- Bergeron RJ, Wiegand J, McManis JS, Bharti N. Desferrithiocin: A Search for Clinically Effective Iron Chelators. J Med Chem. 2014 Nov 26;57(22):9259–91.
- Gaynor RB, McIntyre BN, Lindsey SL, Clavo KA, Shy WE, Mees DE, et al. Steric Effects on the Chelation of Mn2+ and Zn2+ by Hexadentate Polyimidazole Ligands: Modeling Metal Binding by Calprotectin Site 2. Chemistry. 2023 July 3;29(37):e202300447.
- Horonchik L, Wessling-Resnick M. The Small-Molecule Iron Transport Inhibitor Ferristatin/NSC306711 Promotes Degradation of the Transferrin Receptor. Chemistry & Biology. 2008 July 21;15(7):647–53.
- Juttukonda LJ, Beavers WN, Unsihuay D, Kim K, Pishchany G, Horning KJ, et al. A Small-Molecule Modulator of Metal Homeostasis in Gram-Positive Pathogens. mBio. 2020 Oct 27;11(5):10.1128/mbio.02555-20.
- Ito A, Nishikawa T, Matsumoto S, Yoshizawa H, Sato T, Nakamura R, et al. Siderophore Cephalosporin Cefiderocol Utilizes Ferric Iron Transporter Systems for Antibacterial Activity against Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2016 Nov 21;60(12):7396–401.
- Chan DCK, Guo I, Burrows LL. Forging New Antibiotic Combinations under Iron-Limiting Conditions. Antimicrob Agents Chemother. 2020 Feb 21;64(3):e01909-19.
- Li X qin, Zhang W xian. Sequestration of Metal Cations with Zerovalent Iron NanoparticlesA Study with High Resolution X-ray Photoelectron Spectroscopy (HR-XPS). J Phys Chem C. 2007 May 1;111(19):6939–46.
- Khatun S, Putta CL, Hak A, Rengan AK. Immunomodulatory nanosystems: An emerging strategy to combat viral infections. Biomater Biosyst. 2023 Jan 30;9:100073.
- Mitchell MJ, Billingsley MM, Haley RM, Wechsler ME, Peppas NA, Langer R. Engineering precision nanoparticles for drug delivery. Nat Rev Drug Discov. 2021 Feb;20(2):101–24.
- Clark MA, Goheen MM, Cerami C. Influence of host iron status on Plasmodium falciparum infection. Front Pharmacol [Internet]. 2014 May 6 [cited 2025 Aug 31];5. Available from: https://www.frontiersin.org/journals/pharmacology/articles/10.3389/fphar.2014.00084/full.
- Traore O, Carnevale P, Kaptue-Noche L, M’Bede J, Desfontaine M, Elion J, et al. Preliminary report on the use of desferrioxamine in the treatment of Plasmodium falciparum malaria. Am J Hematol. 1991 July;37(3):206–8.
- Gordeuk VR, Biemba G, Thuma PE. Clinical Studies of Iron-Chelating Treatment in Malaria. In: Abraham NG, Asano S, Brittinger G, Maestroni GJM, Shadduck RK, editors. Molecular Biology of Hematopoiesis 5 [Internet]. Boston, MA: Springer US; 1996 [cited 2025 Aug 31]. p. 685–91. Available from. [CrossRef]
- Smith HJ, Meremikwu MM. Iron-chelating agents for treating malaria. Cochrane Database Syst Rev. 2003 Apr 22;2003(2):CD001474.
- Theriault ME, Pisu D, Wilburn KM, Lê-Bury G, MacNamara CW, Michael Petrassi H, et al. Iron limitation in M. tuberculosis has broad impact on central carbon metabolism. Commun Biol. 2022 July 9;5(1):685.
- Parihar SP, Guler R, Khutlang R, Lang DM, Hurdayal R, Mhlanga MM, et al. Statin therapy reduces the mycobacterium tuberculosis burden in human macrophages and in mice by enhancing autophagy and phagosome maturation. J Infect Dis. 2014 Mar 1;209(5):754–63.
- Jin X, Zhang M, Lu J, Duan X, Chen J, Liu Y, et al. Hinokitiol chelates intracellular iron to retard fungal growth by disturbing mitochondrial respiration. Journal of Advanced Research. 2021 Dec 1;34:65–77.
- Garnacho-Montero J, Barrero-García I, León-Moya C. Fungal infections in immunocompromised critically ill patients. Journal of Intensive Medicine. 2024 July 1;4(3):299–306.
- Ali Zaidi SS, Fatima F, Ali Zaidi SA, Zhou D, Deng W, Liu S. Engineering siRNA therapeutics: challenges and strategies. Journal of Nanobiotechnology. 2023 Oct 18;21(1):381.
- Munteanu C, Schwartz B. The relationship between nutrition and the immune system. Front Nutr. 2022 Dec 8;9:1082500.
- Sejersen K, Eriksson MB, Larsson AO. Calprotectin as a Biomarker for Infectious Diseases: A Comparative Review with Conventional Inflammatory Markers. Int J Mol Sci. 2025 July 4;26(13):6476.
- Jukic A, Bakiri L, Wagner EF, Tilg H, Adolph TE. Calprotectin: from biomarker to biological function. Gut. 2021 Oct;70(10):1978–88.
- Bjarnason, I. The Use of Fecal Calprotectin in Inflammatory Bowel Disease. Gastroenterol Hepatol (N Y). 2017 Jan;13(1):53–6.
- Xu MJ, Feng D, Wu H, Wang H, Chan Y, Kolls J, et al. The liver is the major source of elevated serum lipocalin-2 levels after bacterial infection or partial hepatectomy: a critical role for IL-6/STAT3. Hepatology. 2015 Feb;61(2):692–702.
- Stuart T, Satija R. Integrative single-cell analysis. Nat Rev Genet. 2019 May;20(5):257–72.
- Ladomersky E, Petris MJ. Copper tolerance and virulence in bacteria. Metallomics. 2015 June 1;7(6):957–64.
- Karlsson EA, Beck MA, MacIver NJ. Editorial: Nutritional Aspects of Immunity and Immunometabolism in Health and Disease. Front Immunol [Internet]. 2020 Oct 7 [cited 2025 Sept 4];11. Available from: https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2020.595115/full.
- Su Y, Chen D, Yuan D, Lausted C, Choi J, Dai CL, et al. Multi-Omics Resolves a Sharp Disease-State Shift between Mild and Moderate COVID-19. Cell. 2020 Dec 10;183(6):1479-1495.e20.
- Preza GC, Ruchala P, Pinon R, Ramos E, Qiao B, Peralta MA, et al. Minihepcidins are rationally designed small peptides that mimic hepcidin activity in mice and may be useful for the treatment of iron overload. J Clin Invest. 2011 Dec 1;121(12):4880–8.
- Casu C, Nemeth E, Rivella S. Hepcidin agonists as therapeutic tools. Blood. 2018 Apr 19;131(16):1790–4.
- Katsarou A, Pantopoulos K. Hepcidin Therapeutics. Pharmaceuticals (Basel). 2018 Nov 21;11(4):127.
- Nairz M, Haschka D, Demetz E, Weiss G. Iron at the interface of immunity and infection. Front Pharmacol [Internet]. 2014 July 16 [cited 2025 Sept 4];5. Available from: https://www.frontiersin.org/journals/pharmacology/articles/10.3389/fphar.2014.00152/full.
- Gallant CJ, Malik S, Jabado N, Cellier M, Simkin L, Finlay BB, et al. Reduced in vitro functional activity of human NRAMP1 (SLC11A1) allele that predisposes to increased risk of pediatric tuberculosis disease. Genes & Immunity. 2007 Dec;8(8):691–8.
- Schaible UE, Kaufmann SHE. Malnutrition and Infection: Complex Mechanisms and Global Impacts. PLOS Medicine. 2007 May 1;4(5):e115.
- Hambidge M. Human zinc deficiency. J Nutr. 2000 May;130(5S Suppl):1344S-9S.
- Trivellone V, Hoberg EP, Boeger WA, Brooks DR. Food security and emerging infectious disease: risk assessment and risk management. Royal Society Open Science. 2022 Feb 16;9(2):211687.
- Tong MX, Hansen A, Hanson-Easey S, Cameron S, Xiang J, Liu Q, et al. Infectious Diseases, Urbanization and Climate Change: Challenges in Future China. International Journal of Environmental Research and Public Health. 2015 Sept;12(9):11025–36.
- Brestoff JR, Artis D. Commensal bacteria at the interface of host metabolism and the immune system. Nature Immunology. 2013 July;14(7):676–84.
- Kortman GAM, Raffatellu M, Swinkels DW, Tjalsma H. Nutritional iron turned inside out: intestinal stress from a gut microbial perspective. FEMS Microbiol Rev. 2014 Nov 1;38(6):1202–34.
- Ellermann M, Arthur JC. Siderophore-mediated iron acquisition and modulation of host-bacterial interactions. Free Radic Biol Med. 2017 Apr;105:68–78.
- Fusco W, Lorenzo MB, Cintoni M, Porcari S, Rinninella E, Kaitsas F, et al. Short-Chain Fatty-Acid-Producing Bacteria: Key Components of the Human Gut Microbiota. Nutrients. 2023 May 6;15(9):2211.
- Sankarganesh P, Bhunia A, Ganesh Kumar A, Babu AS, Gopukumar ST, Lokesh E. Short-chain fatty acids (SCFAs) in gut health: Implications for drug metabolism and therapeutics. Medicine in Microecology. 2025 Sept 1;25:100139.
- Trompette A, Pernot J, Perdijk O, Alqahtani RAA, Domingo JS, Camacho-Muñoz D, et al. Gut-derived short-chain fatty acids modulate skin barrier integrity by promoting keratinocyte metabolism and differentiation. Mucosal Immunol. 2022 May;15(5):908–26.
- Koh A, Vadder FD, Kovatcheva-Datchary P, Bäckhed F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell. 2016 June 2;165(6):1332–45.
- Shen Y, Fan N, Ma S, Cheng X, Yang X, Wang G. Gut Microbiota Dysbiosis: Pathogenesis, Diseases, Prevention, and Therapy. MedComm (2020). 2025 Apr 18;6(5):e70168.
- Kim CH. Complex regulatory effects of gut microbial short-chain fatty acids on immune tolerance and autoimmunity. Cell Mol Immunol. 2023 Apr;20(4):341–50.
- Sousa Gerós A, Simmons A, Drakesmith H, Aulicino A, Frost JN. The battle for iron in enteric infections. Immunology. 2020 Nov;161(3):186–99.
- Lim HJ, Shin HS. Antimicrobial and Immunomodulatory Effects of Bifidobacterium Strains: A Review. J Microbiol Biotechnol. 2020 Dec 28;30(12):1793–800.
- Vlasova AN, Kandasamy S, Chattha KS, Rajashekara G, Saif LJ. Comparison of probiotic lactobacilli and bifidobacteria effects, immune responses and rotavirus vaccines and infection in different host species. Vet Immunol Immunopathol. 2016 Apr;172:72–84.
- Toledo A, Benach JL. Hijacking and Use of Host Lipids by Intracellular Pathogens. Microbiology Spectrum [Internet]. 2015 Dec 21 [cited 2025 Sept 16]; Available from: https://journals.asm.org/doi/10.1128/microbiolspec.vmbf-0001-2014.
- Russell DG, Huang L, VanderVen BC. Immunometabolism at the interface between macrophages and pathogens. Nat Rev Immunol. 2019 May;19(5):291–304.
- Bosch M, Sánchez-Álvarez M, Fajardo A, Kapetanovic R, Steiner B, Dutra F, et al. Mammalian lipid droplets are innate immune hubs integrating cell metabolism and host defense. Science [Internet]. 2020 Oct 16 [cited 2025 Sept 16]; Available from: https://www.science.org/doi/10.1126/science.aay8085.
- Allen PE, Martinez JJ. Modulation of Host Lipid Pathways by Pathogenic Intracellular Bacteria. Pathogens. 2020 Aug;9(8):614.
- Monson EA, Trenerry AM, Laws JL, Mackenzie JM, Helbig KJ. Lipid droplets and lipid mediators in viral infection and immunity. FEMS Microbiol Rev [Internet]. 2021 Aug 17 [cited 2025 Sept 16];45(4). Available from. [CrossRef]
- Nazarova EV, Montague CR, La T, Wilburn KM, Sukumar N, Lee W, et al. Rv3723/LucA coordinates fatty acid and cholesterol uptake in Mycobacterium tuberculosis. Kana BD, editor. eLife. 2017 June 27;6:e26969.
- Qin C, Xie T, Yeh WW, Savas AC, Feng P. Metabolic Enzymes in Viral Infection and Host Innate Immunity. Viruses. 2024 Jan;16(1):35.
- Drakesmith H, Prentice A. Viral infection and iron metabolism. Nat Rev Microbiol. 2008 July;6(7):541–52.
- Wessling-Resnick M. Crossing the Iron Gate: Why and How Transferrin Receptors Mediate Viral Entry. Annu Rev Nutr. 2018 Aug 21;38:431–58.
- Girelli D, Marchi G, Busti F, Vianello A. Iron metabolism in infections: Focus on COVID-19. Semin Hematol. 2021 July;58(3):182–7.
- Read SA, Obeid S, Ahlenstiel C, Ahlenstiel G. The Role of Zinc in Antiviral Immunity. Advances in Nutrition. 2019 July 1;10(4):696–710.
- Velthuis AJW te, Worm SHE van den, Sims AC, Baric RS, Snijder EJ, Hemert MJ van. Zn2+ Inhibits Coronavirus and Arterivirus RNA Polymerase Activity In Vitro and Zinc Ionophores Block the Replication of These Viruses in Cell Culture. PLOS Pathogens. 2010 Nov 4;6(11):e1001176.
- Warnes SL, Keevil CW. Inactivation of Norovirus on Dry Copper Alloy Surfaces. PLOS ONE. 2013 Sept 9;8(9):e75017.
- Wang C, Guan Y, Lv M, Zhang R, Guo Z, Wei X, et al. Manganese Increases the Sensitivity of the cGAS-STING Pathway for Double-Stranded DNA and Is Required for the Host Defense against DNA Viruses. Immunity. 2018 Apr 17;48(4):675-687.e7.
- Nakano R, Nakano A, Sasahara T, Suzuki Y, Nojima Y, Yano H. Antiviral effects of copper and copper alloy and the underlying mechanisms in severe acute respiratory syndrome coronavirus 2. Journal of Hazardous Materials Advances. 2025 Feb 1;17:100589.
- Purniawan A, Lusida MI, Pujiyanto RW, Nastri AM, Permanasari AA, Harsono AAH, et al. Synthesis and assessment of copper-based nanoparticles as a surface coating agent for antiviral properties against SARS-CoV-2. Sci Rep. 2022 Mar 22;12(1):4835.
- Wessels I, Rolles B, Rink L. The Potential Impact of Zinc Supplementation on COVID-19 Pathogenesis. Front Immunol [Internet]. 2020 July 10 [cited 2025 Sept 17];11. Available from: https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2020.01712/full.
- Vallboehmer F, Schoofs H, Rink L, Jakobs J. Zinc supplementation among zinc-deficient vegetarians and vegans restores antiviral interferon-α response by upregulating interferon regulatory factor 3. Clinical Nutrition. 2025 Aug 1;51:161–73.
- Dalamaga M, Karampela I, Mantzoros CS. Commentary: Could iron chelators prove to be useful as an adjunct to COVID-19 Treatment Regimens? Metabolism. 2020 July;108:154260.
- Nemeth E, Ganz T. Hepcidin and Iron in Health and Disease. Annu Rev Med. 2023 Jan 27;74:261–77.
- Ganz T. Iron and infection. Int J Hematol. 2018 Jan 1;107(1):7–15.
- Das S, Saqib M, Meng RC, Chittur SV, Guan Z, Wan F, et al. Hemochromatosis drives acute lethal intestinal responses to hyperyersiniabactin-producing Yersinia pseudotuberculosis. Proceedings of the National Academy of Sciences. 2022 Jan 11;119(2):e2110166119.
- Kramer J, Özkaya Ö, Kümmerli R. Bacterial siderophores in community and host interactions. Nat Rev Microbiol. 2020 Mar;18(3):152–63.
- Kontoghiorghe CN, Kontoghiorghes GJ. New developments and controversies in iron metabolism and iron chelation therapy. World J Methodol. 2016 Mar 26;6(1):1–19.
- Kontoghiorghe CN, Kontoghiorghes GJ. Efficacy and safety of iron-chelation therapy with deferoxamine, deferiprone, and deferasirox for the treatment of iron-loaded patients with non-transfusion-dependent thalassemia syndromes. Drug Des Devel Ther. 2016 Jan 29;10:465–81.
- Vamathevan J, Clark D, Czodrowski P, Dunham I, Ferran E, Lee G, et al. Applications of machine learning in drug discovery and development. Nat Rev Drug Discov. 2019 June;18(6):463–77.
- Stokes JM, Yang K, Swanson K, Jin W, Cubillos-Ruiz A, Donghia NM, et al. A Deep Learning Approach to Antibiotic Discovery. Cell. 2020 Feb 20;180(4):688-702.e13.
- Ikoba U, Peng H, Li H, Miller C, Yu C, Wang Q. Nanocarriers in therapy of infectious and inflammatory diseases. Nanoscale. 2015 Feb 26;7(10):4291–305.
- Salouti M, Ahangari A, Salouti M, Ahangari A. Nanoparticle based Drug Delivery Systems for Treatment of Infectious Diseases. In: Application of Nanotechnology in Drug Delivery [Internet]. IntechOpen; 2014 [cited 2025 Sept 14]. Available from: https://www.intechopen.com/chapters/47150.
- Armstead AL, Li B. Nanomedicine as an emerging approach against intracellular pathogens. Int J Nanomedicine. 2011;6:3281–93.
- Hosseini SM, Taheri M, Nouri F, Farmani A, Moez NM, Arabestani MR. Nano drug delivery in intracellular bacterial infection treatments. Biomedicine & Pharmacotherapy. 2022 Feb 1;146:112609.
- Russell DG, Huang L, VanderVen BC. Immunometabolism at the interface between macrophages and pathogens. Nat Rev Immunol. 2019 May;19(5):291–304.
Figure 1.
Schematic illustration of nutritional immunity mechanisms. Illustrated here are the various strategies used by the host to disrupt pathogen nutrition. a, metal sequestration; the host sequesters essential metals like iron (Fe), zinc (Zn), manganese (Mn), and copper (Cu) that pathogens need to grow and multiply. b, metal toxicity; host intentionally delivers toxic levels of certain transition metals into pathogen-containing compartments, which overwhelms microbial detoxification systems, and may have pathogens incorporate incorrect metals into their enzymes, such as copper instead of iron, leading to dysfunctional proteins and impaired microbial survival. c, nutrient competition; host cells and immune cells compete with pathogens for key nutrients like glucose, lipids, and amino acids. d, metabolic reprogramming; the host alters its own metabolism during infection to reduce the availability of nutrients like glucose and amino acids to pathogens. e, hormonal regulation; inflammatory cytokines and hormones like hepcidin regulate iron homeostasis by decreasing iron absorption and trapping iron in storage sites. f, production of antimicrobial peptides; molecules like calprotectin bind and sequester multiple metals, including zinc and manganese, but also have direct antimicrobial effects. g, siderophore interference; hosts produce proteins like lipocalin-2 that bind bacterial siderophores, which are iron-scavenging molecules, preventing pathogens from retrieving iron.
Figure 1.
Schematic illustration of nutritional immunity mechanisms. Illustrated here are the various strategies used by the host to disrupt pathogen nutrition. a, metal sequestration; the host sequesters essential metals like iron (Fe), zinc (Zn), manganese (Mn), and copper (Cu) that pathogens need to grow and multiply. b, metal toxicity; host intentionally delivers toxic levels of certain transition metals into pathogen-containing compartments, which overwhelms microbial detoxification systems, and may have pathogens incorporate incorrect metals into their enzymes, such as copper instead of iron, leading to dysfunctional proteins and impaired microbial survival. c, nutrient competition; host cells and immune cells compete with pathogens for key nutrients like glucose, lipids, and amino acids. d, metabolic reprogramming; the host alters its own metabolism during infection to reduce the availability of nutrients like glucose and amino acids to pathogens. e, hormonal regulation; inflammatory cytokines and hormones like hepcidin regulate iron homeostasis by decreasing iron absorption and trapping iron in storage sites. f, production of antimicrobial peptides; molecules like calprotectin bind and sequester multiple metals, including zinc and manganese, but also have direct antimicrobial effects. g, siderophore interference; hosts produce proteins like lipocalin-2 that bind bacterial siderophores, which are iron-scavenging molecules, preventing pathogens from retrieving iron.
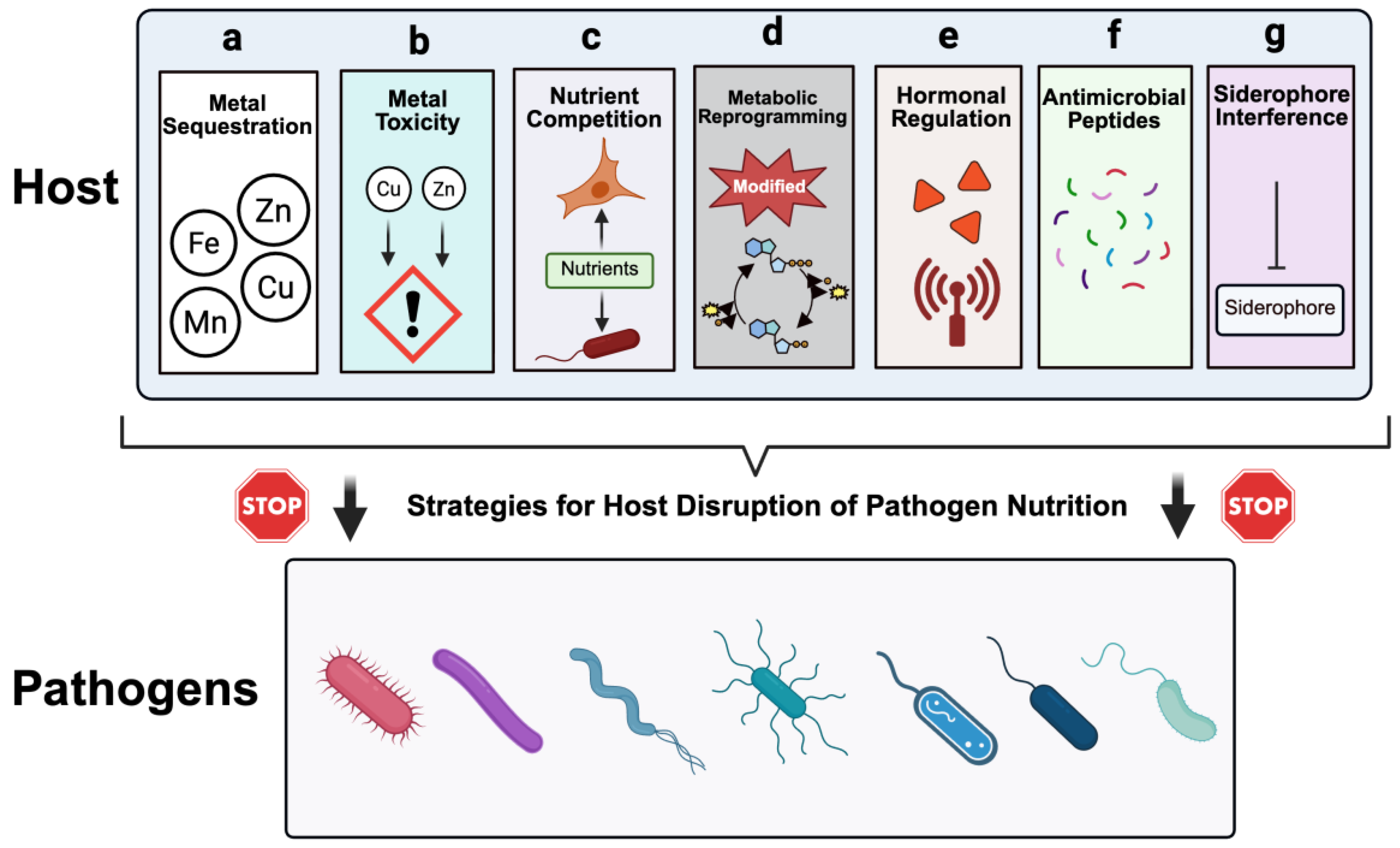
Figure 2.
Host-pathogen tug of war for nutrient trace metals. Upon pathogen entry, the host deploys mechanisms to hide nutrient trace metals from the pathogen. These mechanisms include host proteins like transferrin, which binds Fe in blood, limiting extracellular Fe; lactoferrin, a high-affinity Fe-binding glycoprotein in mucosal secretions and neutrophil granules; ferritin, an intracellular Fe storage, that restricts cytoplasmic Fe availability; hemopexin, which binds free heme, limiting bacterial access to the Fe in heme; haptoglobin, which binds free hemoglobin (Hb), preventing bacterial Hb uptake; lipocalin-2, which binds bacterial siderophores, thereby, blocking Fe scavenging; hepcidin, a hormone that degrades ferroportin, reducing Fe export from cells into circulation; calprotectin, a neutrophil protein that sequesters Zn and Mn; ZIP/ZnT transporters, which control intracellular Zn flux to deprive intracellular or extracellular pathogens; and NRAMP1, which pumps Fe and Mn out of phagosomes to deprive intracellular bacteria. In response, the pathogen, also deploy counter mechanisms that allow them to still access these critical nutrients, including bacterial siderophores like enterobactin, pyoverdine, yersiniabactin, staphyloferrin, mycobactin, which are secreted molecules that chelate Fe with very high affinity. Bacterial pathogens also deploy high-affinity transporters like the ZnuABC system, a high-affinity Zn transporter; MntABC transporter, a high-affinity Mn uptake system; the CbiMNQO transporters that import Co for cobalamin biosynthesis; and lactoferrin receptors, which bind lactoferrin and facilitate pathogen acquisition of Fe. Fe: Iron, Cu: Copper, Zn: Zinc, Mn: Manganese, Co: Cobalt, Ni: Nickel, ZIP: Zrt, Irt-like proteins, ZnT: Zinc transporter, NRAMP1: Natural Resistance-Associated Macrophage Protein 1.
Figure 2.
Host-pathogen tug of war for nutrient trace metals. Upon pathogen entry, the host deploys mechanisms to hide nutrient trace metals from the pathogen. These mechanisms include host proteins like transferrin, which binds Fe in blood, limiting extracellular Fe; lactoferrin, a high-affinity Fe-binding glycoprotein in mucosal secretions and neutrophil granules; ferritin, an intracellular Fe storage, that restricts cytoplasmic Fe availability; hemopexin, which binds free heme, limiting bacterial access to the Fe in heme; haptoglobin, which binds free hemoglobin (Hb), preventing bacterial Hb uptake; lipocalin-2, which binds bacterial siderophores, thereby, blocking Fe scavenging; hepcidin, a hormone that degrades ferroportin, reducing Fe export from cells into circulation; calprotectin, a neutrophil protein that sequesters Zn and Mn; ZIP/ZnT transporters, which control intracellular Zn flux to deprive intracellular or extracellular pathogens; and NRAMP1, which pumps Fe and Mn out of phagosomes to deprive intracellular bacteria. In response, the pathogen, also deploy counter mechanisms that allow them to still access these critical nutrients, including bacterial siderophores like enterobactin, pyoverdine, yersiniabactin, staphyloferrin, mycobactin, which are secreted molecules that chelate Fe with very high affinity. Bacterial pathogens also deploy high-affinity transporters like the ZnuABC system, a high-affinity Zn transporter; MntABC transporter, a high-affinity Mn uptake system; the CbiMNQO transporters that import Co for cobalamin biosynthesis; and lactoferrin receptors, which bind lactoferrin and facilitate pathogen acquisition of Fe. Fe: Iron, Cu: Copper, Zn: Zinc, Mn: Manganese, Co: Cobalt, Ni: Nickel, ZIP: Zrt, Irt-like proteins, ZnT: Zinc transporter, NRAMP1: Natural Resistance-Associated Macrophage Protein 1.
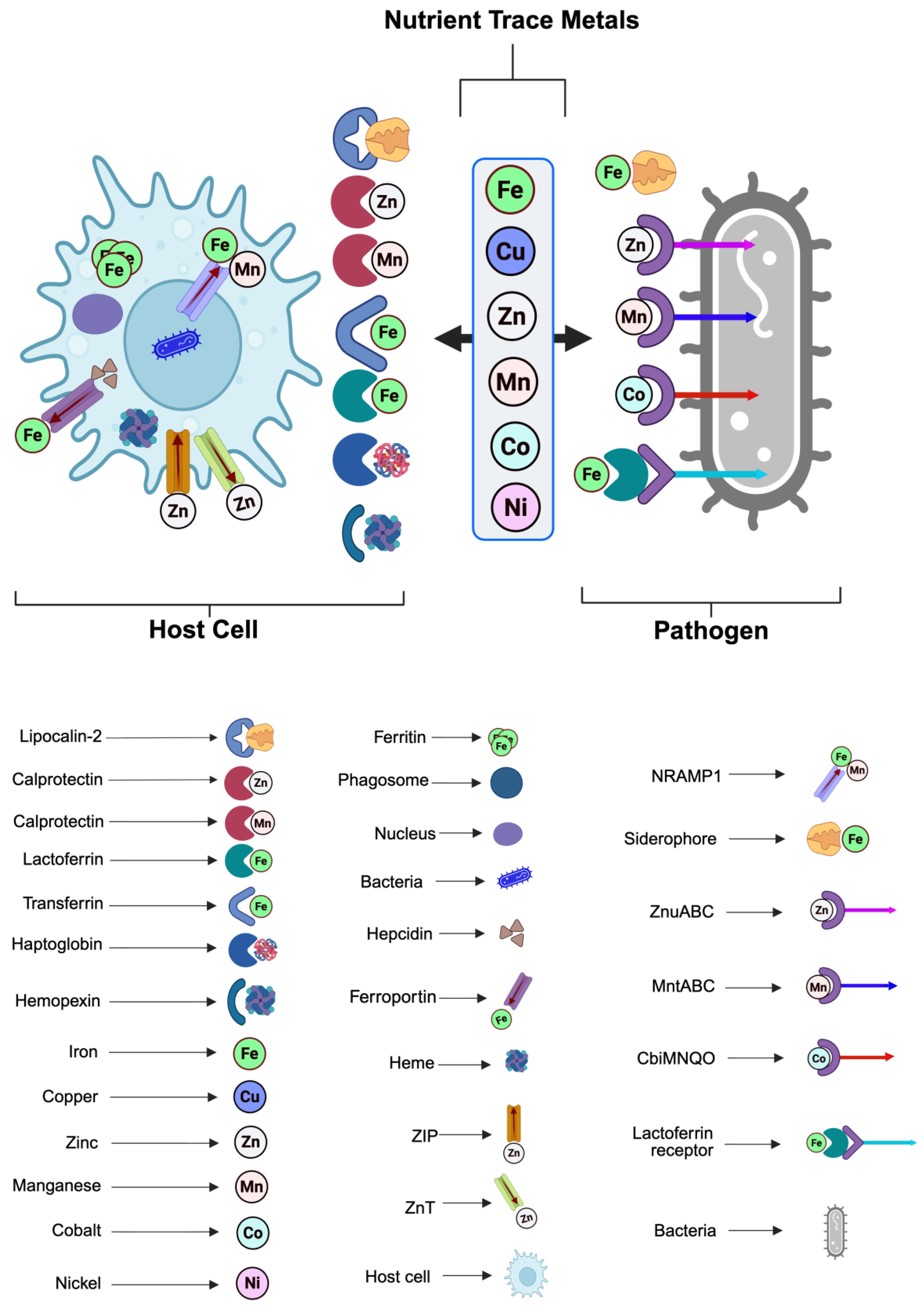
Table 1.
Summary of nutritional immunity mechanisms.
| S/n | Mechanism | Description | Main host factors | Targeted nutrients | Impact on pathogen | Examples | References |
|---|---|---|---|---|---|---|---|
| 1 | Metal sequestration | Host proteins bind essential transition metals firmly, restricting their availability to invading microbes. | Ferritin, lactoferrin, transferrin, ceruloplasmin, hemopexin, haptoglobin. | Fe, Mn, Cu, Zn. | Limits cofactor accessibility for bacterial enzymes such as superoxide dismutase and ribonucleotide reductase, thereby impairing replication. | Neutrophil lactoferrin reduce iron accessibility to Staphylococcus aureus, calprotectin sequesters zinc and manganese from Candida albicans | [5,18,19,20] |
| 2 | Metal Toxicity | Host intentionally delivers toxic levels of certain transition metals into pathogen-containing compartments. | Copper transporters (ATP7A, ATP7B), Zinc transporters (ZIP8, ZnT family), NRAMP1 (SLC11A1). | Excess Cu or Zn causing toxicity and/or replacing Fe/Mn. | Overwhelms microbial detoxification systems, causes mis-metalation, and interferes with critical metabolic processes. | Copper toxicity against Mycobacterium tuberculosis, Zinc-mediated intoxication of Streptococcus pneumoniae and Mycobacterium tuberculosis | [5,17,21,22] |
| 3 | Nutrient competition | Host cells outcompete pathogens for limited nutrients in inflamed tissues. | Activated macrophages, neutrophils, T cells. | Fe, Glucose, fatty acids, amino acids like tryptophan and arginine. | Stress induced by starvation, forcing bacteria into alternative metabolic pathways or non-replicating forms. | Gut commensals competing with Salmonella enterica for iron and carbon sources decrease pathogen colonization | [4,23,24] |
| 4 | Metabolic reprogramming | Host cells shift metabolic flux to starve pathogens, such as Warburg-like glycolysis, and reduced amino acid availability | HIF-1α, mTOR, autophagy pathways. | Glucose, glutamine, arginine, serine. | Limits access to central carbon and nitrogen sources | Macrophage glycolytic reprogramming restricting Mycobacterium tuberculosis growth and altering CCV environment for Coxiella burnetii. | [4,25,26] |
| 5 | Hormonal regulation | Hormones and inflammatory cytokines regulate systemic nutrient availability. | Hepcidin, TNF-α, IL-6. | Iron via regulation of ferroportin. | Reduces plasma iron, trapping it in macrophages and hepatocytes. | IL-6–induced hepcidin release suppressing iron availability during Salmonella infection. | [10,14] |
| 6 | Production of antimicrobial peptides (AMPs) | AMPs bind essential metals and exert direct antimicrobial effects. | Psoriasin (S100A7), calprotectin (S100A8/A9), defensins. | Zn, Mn. | Blocks metalloenzyme activity, triggers oxidative stress. | Calprotectin limiting Staphylococcus aureus growth in neutrophil extracellular traps (NETs). | [20,27,28] |
| 7 | Siderophore interference | Host proteins arrest or neutralize bacterial siderophores to block iron retrieval. | Lipocalin-2 (NGAL), siderocalin. | Iron via siderophore sequestration. | Suppresses iron uptake despite active siderophore production. | Lipocalin-2 binding Escherichia coli enterobactin, blocking bacterial iron acquisition. | [29,30,31,32] |
Table 2.
Therapeutic approaches inspired by nutritional immunity.
| S/N | Therapeutic strategy | Mechanism of action | Examples/clinical trials | Challenges | Translational outlook |
|---|---|---|---|---|---|
| 1 | Iron chelation therapy | Starves pathogens of iron by binding free iron, mimicking host nutritional immunity | Deferasirox and its derivatives used against Candida albicans, Candida glabrata, and Cryptococcus neoformans [192,193]; Deferiprone as an adjunctive therapy against Mycobacterium abscessus [194,195]; Lactoferrin supplementation in trials studies for neonatal sepsis [196,197]. | Risk of host iron depletion, impaired immune function, and anemia. | Early / late clinical (iron chelators already approved for other indications; infection-specific use experimental). |
| 2 | Metal supplementation to cause toxicity | Deliver toxic levels of metals such as zinc, copper, or manganese into pathogen-containing compartments | Zinc oxide nanoparticles antibacterial activity against Staphylococcus aureus, Escherichia coli, Pseudomonas aeruginosa, and Bacillus subtilis [198,199,200]; Copper ionophores such as disulfiram, elesclomol studied for antimicrobial activity against Mycobacterium tuberculosis, Chlamydia trachomatis and Neisseria gonorrhoeae [201,202,203] | Balancing pathogen killing with minimal host toxicity | Preclinical / early clinical. |
| 3 | Siderophore decoys and mimics | Neutralize bacterial siderophores or prevent their uptake by pathogens | Gallium potency against chronic Pseudomonas aeruginosa airway infections in clinical trials [204]; Lipocalin-2 facilitating host defense against Klebsiella pneumoniae [205] and Escherichia coli [206] | Toxicity at high doses and pathogen adaptation | Early clinical (gallium); other decoy approaches preclinical. |
| 4 | Siderophore–antibiotic conjugates as “Trojan horse” drugs | Antibiotics fused to siderophores leverage bacterial uptake systems to facilitate intracellular drug delivery | Cefiderocol, an FDA-approved drug against multidrug resistant Gram-negative bacteria and nosocomial pneumonia and complicated urinary tract infections [207]; siderophore-monobactam (BAL30072) potency against multidrug-resistant Acinetobacter and multidrug resistant gram negative bacteria [208,209]; synthetic sideromycin conjugates under development [207]. | Resistance through altered siderophore receptors and high cost | FDA approved (cefiderocol); siderophore-monobactam and synthetic sideromycin conjugates in preclinical. |
| 5 | Immunometabolic modulation | Remodel host metabolism to limit key nutrients such as glucose, amino acids, and vitamins from pathogens | IDO activators for tryptophan depletion in viral and bacterial infections [210,211]; Methionine restriction shown to limit intestinal barrier dysfunction and inflammation caused by Salmonella typhimurium [212]; Vitamin D supplementation enhances therapeutic outcomes in tuberculosis [210,211]. | Systemic metabolic adverse effects and pathogen metabolic flexibility | Early clinical (IDO, vitamin D). |
| 6 | Probiotic-pathogen competition | Beneficial microbes compete with pathogens for nutrients and synthesize siderophores to limit pathogen growth | Lactobacillus rhamnosus GG in clinical trials for treating pediatric infections [213,214]; Bifidobacterium longum modulates iron metabolism, reduces pathogen colonization [215]; Escherichia coli Nissle 1917 competes for iron and prevents pathogen expansion [216,217]. | Variability in microbiome responses and context-dependent efficacy | Late clinical / approved (some strains widely marketed, regulated as supplements). |
| 7 | Fecal microbiota transplant (FMT) | Restores gut microbial diversity and competitive exclusion against pathogens, restricting access to critical nutrients | FMT for recurrent Clostridium difficile infection in Phase III RCTs showing >85% efficacy [218,219,220,221]; studies indicating FMT efficacy and safety against multidrug-resistant Enterobacteriaceae colonization (222,222–227); emerging FMT trials in sepsis and systemic infections [228,229]. | Regulatory challenges, donor variability, and risk of pathogen transfer. | Late clinical / approved (FDA-approved for recurrent Clostridium difficile). |
| 8 | Phytobiotics (plant-derived compounds) | Plant-derived polyphenols, flavonoids, and alkaloids chelate metals, impair microbial metabolism, and modulate host immunity | Curcumin anti-Mycobacterium activity via iron chelation [230,231]; Quercetin enhances zinc uptake and modulates immune responses [232]; Antimicrobial activity of Epigallocatechin gallate (EGCG) from green tea against Pseudomonas aeruginosa and Escherichia coli [233]. | Bioavailability, limited clinical validation, and variability in plant extracts. | Preclinical/early clinical. |
| 9 | Nanotechnology-based nutrient modulation | Engineered nanocarriers deliver metals, chelators, or metabolic modulators directly to infection sites | Iron oxide nanoparticles investigated for iron sequestration and pathogen restriction [234,235]; Antibacterial activity and mechanistic insights of gallium-based nanoparticles [236,237]; Silver nanoparticles as next-generation antimicrobial agents [238,239,240,241]. | Safety, targeted delivery, and long-term stability concerns | Preclinical |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



