Submitted:
21 November 2025
Posted:
24 November 2025
You are already at the latest version
Abstract
Porcine cytomegalovirus, more accurately classified as porcine roseolovirus (PCMV/PRV), was shown pathogenic in the context of xenotransplantation. Transmission of PCMV/PRV to non-human primates receiving hearts or kidneys from virus-positive pigs significantly reduced the survival time of the recipients. PCMV/PRV was also transmitted to the first human recipient of a pig heart transplant and contributed to the patient’s death. PCMV/PRV is highly prevalent in all pig breeds and wild boars, including slaughterhouse pigs, but there were no reports of infection or disease in either healthy, ill or immunocompromised humans. This indicates that this virus is not zoonotic and must be classified as xenozoonotic. Moreover, it remains unclear whether PCMV/PRV is capable of infecting human cells in vitro. To address this question, human 293T cells resistant to hygromycin were co-cultured with porcine fallopian tube (PFT) cells producing PCMV/PRV. After hygromycin selection, the remaining human cells showed no evidence of infection. Because herpesviruses are generally considered to be species-specific—a notion that has been shown to be not entirely correct—it was also investigated whether PCMV/PRV can infect mouse cells using the same approach. Similarly, no infection was observed. Since the target cells employed in both assays had a reduced capacity to resist viral infection, the findings strongly suggest that PCMV/PRV is unable to infect human or mouse cells, which are equipped with functional antiviral mechanisms.
Keywords:
porcine cytomegalovirus/porcine roseolovirus (PCMV/PRV)
; herpesviruses
; xenotransplantation
; infection assays
1. Introduction
Xenotransplantation using genetically modified pigs offers a promising solution to the shortage of allogeneic donor organs and is steadily advancing toward clinical application. Long-term survival has been achieved in non-human primates transplanted with hearts and kidneys from genetically modified pigs, and the first transplantations of pig hearts, kidneys and livers into human patients and brain-dead individuals have been reported [1]. However, xenotransplantation carries the potential risk of transmitting porcine microorganisms, particularly viruses, to the recipient, and preventing such transmission is of critical importance [2,3].
The transmission of a porcine herpesvirus, porcine cytomegalovirus, more accurately classified as porcine roseolovirus (PCMV/PRV) [4], to the first human recipient of a pig heart transplant, where it contributed to the patient’s death [5], highlights the critical importance of ensuring viral safety in xenotransplantation.
PCMV/PRV (the official name is suid herpesvirus 2, SuHV-2), is widespread in pigs, being detected in both wild boars [6,7] and slaughterhouse pigs [8,9], with nearly all slaughterhouse animals carrying the virus. Despite this broad distribution, no cases of PCMV/PRV transmission to healthy, ill, or immunocompromised humans have been reported, indicating that the virus is not zoonotic. This contrasts with hepatitis E virus genotype 3 (HEV3), which is zoonotic and causes chronic HEV infections, hepatitis and manifestations of extrahepatic diseases in infected individuals [10,11]. The risks associated with acquiring HEV3 include occupational exposure to infected pigs and consumption of undercooked pork or pork products. Pig caretakers and swine veterinarians are at an increased risk of HEV infection [12].
At present, it remains unclear whether PCMV/PRV can infect human cells. One study reported infection of human fibroblasts [13], whereas another showed that co-cultivation of PCMV-infected pig macrophages with two human cell lines (293 and Raji) did not result in virus transmission [14]. In our own preliminary infection studies, using PBMCs from PCMV/PRV-positive pigs and human 293 cells, no evidence of infection was observed.
Therefore, additional experiments are needed to determine whether PCMV/PRV can infect human cells. It is also of interest to assess whether mouse cells are susceptible to PCMV/PRV. Although herpesviruses are generally considered species-specific, this notion has been shown to be not entirely correct [15]. For this reason, we used hygromycin-resistant human and mouse cells, co-incubated them with porcine PFT cells producing PCMV/PRV, and subsequently eliminated the pig cells by adding hygromycin selection medium. Whereas uninfected PFT cells could be infected with PCMV/PRV produced by infected PFT cells, either through co-cultivation or exposure to cell-free virus-containing supernatant, human and mouse cells could not be infected.
2. Materials and Methods
2.1. PCMV/PRV Producing PFT Cells
PFT cells infected with PCMV/PRV were kindly supplied by Dr. N.J. Mueller, University Hospital Zurich, University of Zurich, Switzerland. PFT cells producing PCMV/PRV have been derived from the inner lining of the pig oviduct, also evident by the presence of Barr bodies in these cells. They were adherent and had both epithelial- and fibroblast-like morphology. These cells were characterized by increased nuclear-cytoplasmic ratio as well as increased size and number of nucleoli [16,17,18,19]. Seventeen of 24 RNA viruses and eight of nine DNA viruses replicate in this cell line [20]. They were cultured in Dulbecco’s modified Eagle medium (DMEM) (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% heat inactivated fetal bovine serum (FBS) (PAN Biotech, Aidenbach, Germany), 2mM L-Glutamine (PAN Biotech, Aidenbach, Germany), 1x non-essential amino acids (NEA) (Biochrom GmbH, Berlin, Germany), 100 µg/ml streptomycin and 60 µg/ml penicillin (PAN Biotech, Aidenbach, Germany) in an incubator (Heracell, Thermo Fisher Scientific, Waltham, MA, USA) with 5% CO2 at 37 °C. Upon confluency, cells were detached using Accutase (Invitrogen, Waltham, MA, USA) and split for expansion.
2.2. Hygromycin-Resistant Human Cells
Human embryonic kidney 293T cell line harboring a construct expressing p15E-NHR-His, a part of the transmembrane envelope protein of the porcine endogenous retrovirus (PERV) together with the hygromycin resistance gene (293Thyg) [21] were cultured in DMEM with 10% heat inactivated FBS, 100 µg/ml streptomycin and 60 µg/ml penicillin in 5% CO2 at 37 °C.
2.3. Hygromycin-Resistant Mouse Cells
Embryo fibroblasts from double-stranded RNA-dependent protein kinase (PKR) knockout mice were treated according to the 3T3 protocol [22]. These 3T3-like cells are immortal. In these cells the induction of type I interferon (IFN) as well as the activation of NF-kB by poly(I)-poly(C) (pIC) were strongly impaired [23]. The cells were transfected with a plasmid pY3 having a hygromycin resistance gene. One clone, MI4Y-3/8, was cultured in low-glucose DMEM GlutaMAX (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% heat-inactivated FBS and 1% penicillin/streptomycin (PAN Biotech, Aidenbach, Germany) in 5% CO2 at 37 °C incubator.
2.4. Transfection of PFT Cells
PFT cells were seeded at a cell density of 2×105 per well in a 6-well plate in 2 ml of serum free DMEM media, a day before transfection. 1 µg of pVITRO-eGFP plasmid [24] DNA was mixed with sterile water up to 20 µl in a microfuge tube. The tube was gently flicked for mixing. 200 µl of Opti-MEM (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) was added to the tube containing plasmid and vortexed mildly, followed by a short spin. 8 µl of polyethylenimine (PEI; Sigma-Aldrich, St. Louis, Missouri, USA) were added to the tube and vortexed mildly again, followed by incubation at room temperature for 20 min. The transfection mixture was added to the cells overnight. Next day, the medium was removed and replaced with fresh serum containing DMEM. After 2 days, 400 µg/ml hygromycin was added. Cells were monitored for the expression of eGFP under a fluorescence microscope (Zeiss Axio, Oberkochen, Germany). Upon enrichment of transfected cells and removal of non-transfected ones, they were expanded for subsequent experiments.
2.5. Co-Cultivation and Selection
PFT cells producing PCMV/PRV were co-cultured with 293Thyg cells and MI4Y-3/8 cells (Figure 1), respectively at a cell density of 0.5×106 cells of each cell line/ 5 ml in a T25 flask (Sarstedt AG & Co. KG, Numbrecht, Germany) at 37 °C in a 5% CO2 incubator. At day 3, when cells became confluent, they were split into two T25 flasks, followed by another round of splitting the cells upon reaching confluency. Post attachment of the cells to the flask surface, 400 µg/ml hygromycin was added to the flask for selection of 293Thyg and MI4Y-3/8 cells. After 2 days, cells were split again and maintained in 400 µg/ml hygromycin (Carl Roth, Karlsruhe, Germany) containing complete DMEM until confluency. Co-cultured cells with 293Thyg became confluent in two days whereas with MI4Y-3/8 cells in 4 days. Cells were washed twice with PBS to remove any remaining PFT producing PCMV/PRV cells. Cells were detached using Accutase (Invitrogen, Waltham, MA, USA) and processed for DNA isolation and real-time PCR for testing PCMV/PRV, human glyceraldehyde-3-phosphate-dehydrogenase (hGAPDH) as well as short interspersed nuclear elements (SINE) sequences.
As a positive control for the assay, PFT cell transfected with the pVITRO-eGFP vector were also co-cultured with PFT cells producing PCMV/PRV. After 5 days, 400 µg/ml hygromycin was added to the flask for selection of PFT-pVITRO-eGFP cells. They were monitored under fluorescence microscope and real-time-PCR for PCMV/PRV.
2.6. Cell-Free Infection
Supernatant from PFT cells producing PCMV/PRV was centrifuged twice (300 rpm, 10 min, and 500 rpm, 15 min, at room temperature), to remove any remaining cells. It was filtered using a 0.2 µm filter (Millipore, Burlington, MA, USA) and added repeatedly to the uninfected PFT cells after splitting.
2.7. Virus Pelleting
Supernatant from PFT cells producing PCMV/PRV was collected in a similar way as described in section 2.6. and ultracentrifuged at 30,000 rpm, 4 °C for 1.5 h using SW32 Ti-rotor (Beckmann Coulter, Brea, CA, USA). The pellet was resuspended in 200 µl PBS, DNA was isolated and analyzed by PCR.
2.8. DNA Isolation
DNA was isolated from the cell lines and the virus pellet using DNeasy Blood & Tissue kit (Qiagen, Hilden, Germany) following manufacturer’s instructions. DNA was quantified using a NanoDrop ND-1000 (Thermo Fisher Scientific Inc., Worcester, MA, USA).
2.9. Real-Time PCR for PCMV/PRV, GAPDH and SINE
Real-time PCRs using specific primers and probes (Table 1) were performed with DNA isolated from the respective cell lines to test for PCMV/PRV infection. It was performed using the SensiFAST Probe No-ROX kit (Meridian Bioscience, Cincinnati, OH, USA) in a reaction volume of 16 µl plus 4 µl (100 ng) of DNA template. All real-time PCRs were carried out as duplex PCRs testing simultaneously the virus gene of interest and hGAPDH / murine SINE (SINE B1) / porcine SINE (PRE-1) / porcine GAPDH (pGAPDH) as internal control for each sample. Real-time PCR reactions were carried out with a qTOWER3 G qPCR cycler (Analytik Jena, Jena, Germany).
For PCMV/PRV, hGAPDH, pGAPDH and murine SINE, the temperature-time profile consisted of an initial inactivation step at 50 °C for 2 min, followed by denaturation at 95 °C for 10 min, and 45 cycles of denaturation at 95 °C for 15 s and annealing/extension at 60 °C for 1 min. For porcine SINE, the temperature-time profile consisted of an initial inactivation step at 95 °C for 5 min, followed by 45 cycles of denaturation at 95 °C for 15 s, annealing at 60 °C for 30 s, and extension at 72 °C for 30 s.
2.10. Microscopy
1x106 cells were seeded in T25 cell culture flask (Sarstedt AG & Co。KG, Numbrecht,Germany) and imaged using Zeiss Axio microscope (Oberkochen, Germany) with 10x objective. Brightfield and fluorescent images were acquired. Images were processed using ImageJ [25].
3. Results
3.1. Characterization of PCMV/PRV Used in the Infection Studies
PFT cells and PFT cells producing PCMV/PRV are well characterized, they were found positive for porcine endogenous retrovirus (PERV) respectively for both PERV and PCMV/PRV [16,17,18,19,20]. The cell line was originally derived from the inner lining of the pig oviduct, also known as porcine fallopian tube (PFT) or uterine tubes [16]. This line was susceptible to 17 of 24 porcine RNA viruses and eight of nine porcine DNA viruses, among them was PCMV/PRV [20]. PCMV/PRV-infected cells showed a cytomegalic cytopathogenic effect, and intranuclear inclusion [17]. Recently, virus particles were isolated from the culture supernatant and characterized by electron microscopy [32]. Virus released from PFT cells was shown to infect uninfected PFT cells [18,19,32] as well as porcine aortic endothelial cells (PAEC) in vitro, leading to an activation of the cells as demonstrated by increased expression of porcine tissue factor [33].
PFT cells used here were PCMV/PRV-negative and the PFT cells producing PCMV/PRV (Figure 2) were positive by real-time PCR analysis (Table 2). Both cell lines were as all pig cells positive for porcine endogenous retroviruses PERV-A and PERV-B, but not for PERV-C (Table 2).
In the PCMV/PRV-producing cell line large round cells were visible (Figure 2b), which should represent PCMV/PRV-producing cells as has been shown previously by Fischer et al. [32] using immunological methods.
To determine whether PCMV/PRV is released into the supernatant of PCMV/PRV-positive PFT cells, DNA was repeatedly isolated directly from the culture supernatant and analyzed by real-time PCR for the presence of viral DNA. In all cases, PCMV/PRV was not detected. Therefore, the supernatant was first centrifuged to remove cells, filtrated and then ultracentrifuged to pellet viral particles. PCMV/PRV was subsequently detected in the virus pellet (Table 3). 50 ng DNA were extracted from the virus pellet, the measured 102.4 viral copies correspond to 5 ml supernatant.
3.2. PCMV/PRV Infection of Hygromycin-Resistant PFT Cells
In order to obtain hygromycin-resistant PFT cells, PFT cells were transfected with pVITRO-eGFP and selection was performed with hygromycin. The selected cells showed expression of GFP in all cells and hygromycin resistance (PFThyg) (Figure 3). PFThyg cells were co-cultivated with PFT cells producing PCMV/PRV as shown in Figure 1. Upon selection with hygromycin, virus producing PFT cells were eliminated, followed by screening for PCMV/PRV and porcine SINE by real-time PCR (Table 4). The presence of the SINE sequence indicates the porcine origin of the cells. The PFThyg cells were PCMV/PRV positive, demonstrating proof-of-concept for PCMV/PRV infection experiments.
In both the PCMV/PRV-producing PFT cells as well as the PFT cells freshly infected with PCMV/PRV, we found by real-time PCR 10⁴ copies per 100 ng DNA. Since 100 ng DNA correspond to 1.7 × 10⁴ cells (assuming 6 pg DNA per cell), approximately 58.8% of the cells were infected, assuming that each infected cell carries only one viral copy.
3.3. Failure to Infect Human Cells
Human 293T cells were made resistant to hygromycin (293Thyg) transfecting them with a construct expressing in addition to hygromycin a fragment of the transmembrane envelope protein p15E of PERV used for other purposes [21]. These cells were co-cultured with PFT cells producing PCMV/PRV with splitting the cells in between upon confluency, followed by screening for PCMV/PRV by real-time PCR. A schematic representation of the co-culture experimental set-up is shown in Figure 1. At day 21, both PCR assays—the one detecting PCMV and the one detecting pig SINE—were negative, indicating that no pig cells remained and that the human cells were not infected (Table 4). To note, brightfield microscopy images of 293Thyg cells after co-cultivation and selection (Figure 2) demonstrated the absence of morphologically PFT-PCMV/PRV specific cells.
3.4. Failure to Infect Mouse Cells
Mouse cells derived from RNA-dependent protein kinase (PKR) knockout mice, which were made hygromycin-resistant [23] (Figure 2), were co-cultivated with PFT cells producing PCMV/PRV in a similar manner as with 293Thyg cells (Figure 1). After removal of the pig cells by adding selection medium containing 400 µg/ml hygromycin, the mouse cells were tested for PCMV/PRV and pig SINE, both PCR were negative (Table 4). As an additional control murine SINE B1 was detected in the treated cells.
3.5. Infection of PFT Cells by Supernatant from PCMV/PRV-Producing PFT Cells
To assess whether PFT cells can be infected by cell-free PCMV/PRV, supernatant from PCMV/PRV-positive PFT cells was centrifuged to remove residual cells, filtered through a 0.2 µm filter, and repeatedly added to uninfected PFT cells at each passage together with fresh medium. This procedure was performed every four days over a period of 25 days. At each passage, the cells were washed with PBS and part of them was analyzed by real-time PCR for the presence of PCMV/PRV. Although PCMV/PRV was not detected directly in the supernatant, likely due to a low viral copy number, it was present in the pelleted virus fraction (Table 3) and was capable of infecting PFT cells (Table 5). This indicates that PCMV/PRV can establish a cell-free infection, although the infection rate is lower compared with the rate after co-culture (15% vs. 58,8%).
4. Discussion
PCMV/PRV has been shown to reduce the survival time of xenotransplants when transmitted by an organ from virus-infected donor pigs. This was shown in the case of pig thymokidneys in baboons [33], pig kidneys in baboons [34], pig kidneys in cynomolgus monkeys [35] and pig hearts in baboons [36]. Notably, PCMV/PRV was transmitted to the first human recipient of a pig heart and contributed to the patient’s death [5].
In baboon recipients, PCMV-expressing cells were found in all organs by immunohistochemistry indicating the presence of cell expressing viral proteins [37]. High levels of PCMV/PRV DNA detected by PCR in all baboon organs confirmed this finding [37]. The concurrent identification of porcine SINE sequences in the same samples [38] suggests that these corresponded to disseminated pig cells.
Since humans have not been reported to be infected with PCMV/PRV under natural conditions, and the virus causes disease only in the context of xenotransplantation, it is not considered zoonotic but should instead be classified as xenozoonotic [39]. In baboons, the virus induced consumptive coagulopathy and thrombocytopenia [33]. Analyzing the cytokine levels in the blood, an increase in IL-6 and TNFα in the baboons with PCMV/PRV-positive hearts was observed [36]. Furthermore, high levels of tissue plasminogen activator (tPA) and plasminogen activator inhibitor 1 (tPA-PAI-1) complexes were found, suggesting a complete loss of the pro-fibrinolytic properties of the endothelial cells [36].
Only two studies have addressed the question whether PCMV/PRV can infect human cells. In one, human nasal fibroblasts were inoculated with supernatants from persistently infected porcine turbinate cells (PT-K75 cells) and infection was demonstrated by a cytopathic effect by 7 days post-infection, and reverse-transcriptase polymerase chain reaction sequencing identified PCMV/PRV RNA polymerase transcripts in these infected cells [13]. However, this study lacked critical controls, and the possibility of transmission of virus-producing pig cells could not be excluded. Surprisingly, a monoclonal antibody to human CMV (HCMV) glycoprotein B was used to detect PCMV/PRV. In our studies no cross-rection between PCMV/PRV and HCMV has been observed using the recombinant gB protein of PCMV/PRV [40]. A second study used persistently infected primary porcine alveolar macrophages (PAM) as a source of PCMV/PRV, which were co-cultivated for up to 15 passages with human B (RAJI) and 293 cells. No PCMV/PRV infection was observed by PCR [14]. Consistently, our preliminary infection experiments using PBMCs from PCMV/PRV-positive pigs and human 293 cells also revealed no infection.
Previous work has demonstrated that PCMV/PRV failed to replicate in rabbits, mice, hamsters, chick embryos, or cattle [41], reinforcing its species-specificity.
Herpesviruses were traditionally considered strictly species-specific and incapable of crossing species barriers. However, this view has changed with accumulating evidence demonstrating transspecies transmission in several herpesviruses [42]. For example, baboon cytomegalovirus (BaCMV) has been shown to infect human cells in vitro and was detected in human recipients of baboon liver transplants [43,44]. Similarly, human cytomegalovirus (HCMV) has been reported to infect pig cells [45]. In contrast, PCMV/PRV appears to remain strictly species-specific.
We deliberately selected cell lines with impaired antiviral defense mechanisms for our infection experiments. In 293T cells, a downregulation of several genes involved in viral sensing and restriction has been reported, including apolipoprotein B mRNA editing enzyme catalytic subunit 3G (APOBEC3G), stimulator of interferon genes (STING), Janus kinase 3 (JAK3), IFI16, guanylate-binding protein 5 (GBP5), bone marrow stromal cell antigen 2 (BST2), and MX2 [46,47]. Moreover, 293 cells are deficient in Toll-like receptors (TLRs) and Nod2 [48]. These cells are profoundly defective in cytosolic DNA-sensing and interferon induction, and multiple HEK293 cell lines—regardless of their source—naturally lack detectable cGAS or STING protein expression [49,50].
The functions of these factors are well characterized. APOBEC3G functions as a cytidine deaminase that introduces cytidine-to-uridine mutations, thereby compromising viral coding sequences and replication capacity. STING detects viral nucleic acids and triggers the production of interferons α and β. JAK3, a member of the Janus family of tyrosine kinases associated with cytokine receptors, mediates downstream signaling upon cytokine binding; its inhibition is known to exert immunosuppressive effects, for instance, in organ transplantation. The interferon-inducible protein IFI16 contributes to viral DNA sensing [51], whereas GBP5, a dynamin-like GTPase, promotes NLRP3 inflammasome activation and plays an important role in innate immunity. BST2, better known as tetherin, prevents the release of budding viral particles from the cell surface. The interferon-induced GTP-binding protein Mx2 displays antiviral activity against HIV-1 and herpesviruses [52]. Finally, cyclic GMP–AMP synthase (cGAS) acts as a cytosolic DNA sensor that activates type I interferon responses; notably, cGAS-deficient mice are more susceptible to lethal infections caused by both DNA and RNA viruses [53].
Mouse target cells were derived from mice lacking functional double-stranded RNA-dependent protein kinase (PKR) [23]. PKR has been implicated in the induction of interferons (IFNs) and in the antiviral response. In these mice, the antiviral activity induced by IFN-γ and poly(I:C) (pIC) was reduced; however, the induction of type I IFN genes by pIC and viral infection remained unimpaired. In contrast, embryonic fibroblasts derived from PKR knockout mice exhibited a pronounced impairment in both type I IFN induction and NF-κB activation following pIC stimulation.
Most interferon-stimulated genes (ISGs) encode signaling molecules that enhance cellular responsiveness to pathogens and propagate immune signals from localized sites of infection. PKR serves as an effector molecule that mediates antiviral responses through the phosphorylation of protein substrates, thereby activating signaling pathways involved in homeostasis, immune regulation, and—under sustained activation—apoptosis [54]. IFN signaling induces PKR expression; however, PKR remains inactive until activated by binding to double-stranded RNA (dsRNA) [55,56]. Interaction with dsRNA triggers PKR dimerization and autophosphorylation, leading to kinase activation. Activated PKR then catalyzes serine/threonine phosphorylation of various substrates, most notably the α-subunit of eukaryotic translation initiation factor 2 (eIF2α) [57,58]. Phosphorylation of eIF2α by PKR inhibits both host and viral mRNA translation, effectively suppressing viral replication [58]. The absence of PKR in the mouse cells used for our infection experiments therefore resulted in a substantial reduction of antiviral activity.
Since the target cells used in both assays exhibited a reduced capacity to mount antiviral defenses, our findings strongly indicate that PCMV/PRV is inherently unable to infect human or mouse cells, which are equipped with functional antiviral mechanisms. This observation is consistent with previous reports showing that PCMV/PRV does not infect human 293 or B cells [14], as well as with findings from non-human primate and human recipients of PCMV/PRV-positive xenotransplants, in whom no evidence of infection has been detected [5,36,37].
For the infection assay, a highly efficient co-incubation method was applied in which PCMV/PRV-producing cells are directly mixed with target cells that are resistant to a specific selection marker [59]. The resistance gene enables selective elimination of virus-producing cells upon addition of the appropriate selection medium. This assay supports both infection by cell-free virus and cell-to-cell transmission, while allowing complete removal of the virus-producing pig cells. To verify full elimination of pig cells, detection of the porcine SINE PRE1 sequence was employed as a highly sensitive confirmation method. In Sus scrofa, approximately 1.7 million SINE copies have been reported [60], of which more than one million are PRE1 elements [61]. In Chinese pig breeds, 1.98 million PRE1 copies have been detected [62]. Our finding of 108 SINE PRE1 copies in the PFT cells (Table 4) falls within a comparable range. Similar to the porcine SINEs, very high copy numbers have also been reported in mice, with approximately 564,000 B1 copies in the mouse genome when sequencing the genome of a female C57BL/6J mouse [63,64]. We found 106.9, i.e., 7,940,000 cells in the immortalized MI4Y-3/8 cells from PKR knockout mice with a 129/Sv(ev)XC57BL/6J genetic background, indicating that the copy number may be even higher. It is well known that sequencing analyses usually underestimate the true number of repetitive sequences [65].
5. Conclusions
PCMV/PRV has been transmitted in numerous preclinical xenotransplantation studies and in the first transplantation of a pig heart into a human patient, where it contributed to the death of the recipients. Here, we demonstrate that PCMV/PRV does not infect human or mouse cells. Given that these cells exhibit a markedly reduced antiviral defense capacity, it can be concluded that the virus is intrinsically unable to infect human or mouse cells, which are equipped with functional antiviral mechanisms. Consequently, PCMV/PRV likely acts indirectly on recipient immune and endothelial cells to induce pathological changes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Standard curve of the real-time PCR using primers and probes for the murine SINE B1 sequence.
Author Contributions
Conceptualization, J.D.; methodology, H.J., R.S., and J.D.; validation, H.J., R.S., B.K. and J.D.; formal analysis, H.J., R.S., B.K. and J.D.; investigation, H.J.; resources, R.S.; data curation, H.J., R.S., B.K. and J.D; writing—original draft preparation, J.D. and H.J.; writing—review and editing, H.J., R.S., B.K. and J.D.; supervision, J.D. and B.K.; project administration, J.D. and B.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are present in this manuscript.
Acknowledgments
We would like to thank Antonia Godehardt for discussion of previous infection experiments and Nicolas Mueller for the PFT cells.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| PERV | Porcine endogenous retrovirus |
| PCMV/PRV | Porcine cytomegalovirus/porcine reoseolovirus |
| PFT | Porcine fallopian tube |
| SINE | Short Interspersed Nuclear Element |
| SuHV-2 | Suid herpesvirus 2 |
References
- Cooper, D.K.C.; Pierson, R.N. 3rd. Milestones on the path to clinical pig organ xenotransplantation. Am. J. Transplant. 2023, 23, 326–335. [Google Scholar] [CrossRef]
- Denner, J. Virus Safety of Xenotransplantation. Viruses. 2022, 14. [Google Scholar] [CrossRef]
- Fishman, J.A. Prevention of infection in xenotransplantation: Designated pathogen-free swine in the safety equation. Xenotransplantation. 2020, 27. [Google Scholar] [CrossRef]
- Denner, J.; Bigley, T.M.; Phan, T.L.; Zimmermann, C.; Zhou, X.; Kaufer, B.B. Comparative Analysis of Roseoloviruses in Humans, Pigs, Mice, and Other Species. Viruses. 2019, 11. [Google Scholar] [CrossRef]
- Griffith, B.P.; Grazioli, A.; Singh, A.K.; Tully, A.; Galindo, J.; Saharia, K.K.; Shah, A.; Strauss, E.R.; Odonkor, P.N.; Williams, B.; Silverman, H.J.; Burke, A.; Drachenberg, C.B.; Wells, C.L.; Dickfeld, T.; Hong, S.N.; Hicks, A.J. 3rd; Ananthram, M.; Gupta, A.; Christenson, R.H.; Tamburro, L.; Zhang, T.; Hershfeld, A.; Lewis, B.; Feller, E.D.; Kuravi, K.; Sorrells, L.; Morgand, E.; Mezine, F.; Goutaudier, V.; Rothblatt, M.; Lau, C.L.; Taylor, B.; Perrin, S.; Loupy, A.; Ayares, D.; Mohiuddin, M.M. Transplantation of a genetically modified porcine heart into a live human. Nat. Med. 2025, 31, 589–598. [Google Scholar] [CrossRef]
- Jezdimirović, N.; Savić, B.; Milovanović, B.; Glišić, D.; Ninković, M.; Kureljušić, J.; Maletić, J.; Aleksić Radojković, J.; Kasagić, D.; Milićević, V. Molecular Detection of Porcine Cytomegalovirus, Porcine Parvovirus, Aujeszky Disease Virus and Porcine Reproductive and Respiratory Syndrome Virus in Wild Boars Hunted in Serbia during 2023. Vet. Sci. 2024, 11. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.; Menandro, M.L.; Franzo, G.; Krabben, L.; Marino, S.F.; Kaufer, B.; Denner, J. Presence of porcine cytomegalovirus, a porcine roseolovirus, in wild boars in Italy and Germany. Arch. Virol. 2023, 168. [Google Scholar] [CrossRef] [PubMed]
- Jhelum, H.; Kaufer, B.; Denner, J. Application of Methods Detecting Xenotransplantation-Relevant Viruses for Screening German Slaughterhouse Pigs. Viruses. 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Jhelum, H.; Papatsiros, V.; Papakonstantinou, G.; Krabben, L.; Kaufer, B.; Denner, J. Screening for Viruses in Indigenous Greek Black Pigs. Microorganisms. 2024, 12. [Google Scholar] [CrossRef]
- Wang, B.; Meng, X.J. Hepatitis E virus: host tropism and zoonotic infection. Curr. Opin. Microbiol. 2021, 59, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. Xenotransplantation and Hepatitis E virus. Xenotransplantation. 2015, 22, 167–173. [Google Scholar] [CrossRef]
- Wu, J.Y.; Lau, E.H.Y.; Lu, M.L.; Guo, C.; Guo, Z.M.; Yuan, J.; Lu, J.H. An occupational risk of hepatitis E virus infection in the workers along the meat supply chains in Guangzhou, China. One Health. 2022, 14, 100376. [Google Scholar] [CrossRef]
- Whitteker, J.L.; Dudani, A.K.; Tackaberry, E.S. Human fibroblasts are permissive for porcine cytomegalovirus in vitro. Transplantation. 2008, 86, 155–162. [Google Scholar] [CrossRef]
- Tucker, A.W.; Galbraith, D.; McEwan, P.; Onions, D. Evaluation of porcine cytomegalovirus as a potential zoonotic agent in xenotransplantation. Transplant. Proc. 1999, 31, 915. [Google Scholar] [CrossRef]
- Denner, J. Reduction of the survival time of pig xenotransplants by porcine cytomegalovirus. Virol. J. 2018, 15. [Google Scholar] [CrossRef] [PubMed]
- Bouillant, A.M.; Genest, P.; Greig, A.S. Growth characteristics of a cell line derived from the pig oviduct. Can. J. Microbiol. 1975, 21, 2094–2097. [Google Scholar] [CrossRef] [PubMed]
- Bouillant, A.M.; Greig, A.S. Type C virus production by a continuous line of pig oviduct cells (PFT). J. Gen. Virol. 1975, 27, 173–180. [Google Scholar] [CrossRef]
- Kawamura, H.; Tajima, T.; Hironao, T.; Kajikawa, T.; Kotani, T. Replication of porcine cytomegalovirus in the 19-PFT cell line. J. Vet. Med. Sci. 1992, 54, 1209–1211. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, H.; Matsuzaki, S. Influence of 12-O-tetradecanoylphorbol 13-acetate on replication of porcine cytomegalovirus in the 19-PFT-F cell line. J. Vet. Med. Sci. 1996, 58, 263–265. [Google Scholar] [CrossRef]
- Bouillant, A.M.; Dulac, G.C.; Willis, N.; Girard, A.; Greig, A.S.; Boulanger, P. Viral susceptibility of a cell line derived from the pig oviduct. Can. J. Comp. Med. 1975, 39, 450–456. [Google Scholar]
- Denner, J.; Schwinzer, R.; Pokoyski, C.; Kaufer, B.B.; Dierkes, B.; Lovlesh, L. Further evidence for the immunosuppressive activity of the transmembrane envelope protein p15E of the porcine endogenous retrovirus (PERV). Research Square. 2025. [Google Scholar] [CrossRef]
- Todaro, G.J.; Green, H. Quantitative studies of the growth of mouse embryo cells in culture and their development into established lines. J. Cell Biol. 1963, 17, 299–313. [Google Scholar] [CrossRef]
- Yang, Y.L.; Reis, L.F.; Pavlovic, J.; Aguzzi, A.; Schäfer, R.; Kumar, A.; Williams, B.R.; Aguet, M.; Weissmann, C. Deficient signaling in mice devoid of double-stranded RNA-dependent protein kinase. EMBO J. 1995, 14, 6095–6106. [Google Scholar] [CrossRef]
- Schippers, T. Functional Characterization of the Potential Immune Evasion Proteins pUL49.5 and p012 of Marek’s Disease Virus (MDV). Ph.D. Thesis, Free University Berlin, Freie Universität, Berlin, Germany.
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Mueller, N.J.; Barth, R.N.; Yamamoto, S.; Kitamura, H.; Patience, C.; Yamada, K.; Cooper, D.K.; Sachs, D.H.; Kaur, A.; Fishman, J.A. Activation of cytomegalovirus in pig-to-primate organ xenotransplantation. J. Virol. 2002, 76, 4734–4740. [Google Scholar] [CrossRef]
- Behrendt, R.; Fiebig, U.; Norley, S.; Gürtler, L.; Kurth, R.; Denner, J. A neutralization assay for HIV-2 based on measurement of provirus integration by duplex real-time PCR. J. Virol. Methods 2009, 159, 40–46. [Google Scholar] [CrossRef]
- Duvigneau, J.C.; Hartl, R.T.; Groiss, S.; Gemeiner, M. Quantitative simultaneous multiplex real-time PCR for the detection of porcine cytokines. J. Immunol. Methods 2005, 306, 16–27. [Google Scholar] [CrossRef]
- Walker, J.A.; Hughes, D.A.; Anders, B.A.; Shewale, J.; Sinha, S.K.; Batzer, M.A. Quantitative intra-short interspersed element PCR for species-specific DNA identification. Anal. Biochem. 2003, 316, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Gualtieri, A.; Andreola, F.; Sciamanna, I.; Sinibaldi-Vallebona, P.; Serafino, A.; Spadafora, C. Increased expression and copy number amplification of LINE-1 and SINE B1 retrotransposable elements in murine mammary carcinoma progression. Oncotarget 2013, 4, 1882–1893. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; Patience, C.; Magre, S.; Weiss, R.A.; Banerjee, P.T.; Le Tissier, P.; Stoye, J.P. Host range and interference studies of three classes of pig endogenous retrovirus. J. Virol. 1998, 72, 9986–9991. [Google Scholar] [CrossRef] [PubMed]
- Fischer, N.; Gulich, B.; Keßler, B.; Längin, M.; Fishman, J.A.; Wolf, E.; Boller, K.; Tönjes, R.R.; Godehardt, A.W. PCR and peptide based PCMV detection in pig - development and application of a combined testing procedure differentiating newly from latent infected pigs. Xenotransplantation. 2023, 30. [Google Scholar] [CrossRef] [PubMed]
- Gollackner, B.; Mueller, N.J.; Houser, S.; Qawi, I.; Soizic, D.; Knosalla, C.; Buhler, L.; Dor, F.J.; Awwad, M.; Sachs, D.H.; Cooper, D.K.; Robson, S.C.; Fishman, J.A. Porcine cytomegalovirus and coagulopathy in pig-to-primate xenotransplantation. Transplantation. 2003, 75, 1841–1847. [Google Scholar] [CrossRef]
- Yamada, K.; Tasaki, M.; Sekijima, M.; Wilkinson, R.A.; Villani, V.; Moran, S.G.; Cormack, T.A.; Hanekamp, I.M.; Hawley, R.J.; Arn, J.S.; Fishman, J.A.; Shimizu, A.; Sachs, D.H. Porcine cytomegalovirus infection is associated with early rejection of kidney grafts in a pig to baboon xenotransplantation model. Transplantation 2014, 98, 411–418. [Google Scholar] [CrossRef]
- Sekijima, M.; Waki, S.; Sahara, H.; Tasaki, M.; Wilkinson, R.A.; Villani, V.; Shimatsu, Y.; Nakano, K.; Matsunari, H.; Nagashima, H.; Fishman, J.A.; Shimizu, A.; Yamada, K. Results of life-supporting galactosyltransferase knockout kidneys in cynomolgus monkeys using two different sources of galactosyltransferase knockout swine. Transplantation 2014, 98, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Denner, J.; Längin, M.; Reichart, B.; Krüger, L.; Fiebig, U.; Mokelke, M.; Radan, J.; Mayr, T.; Milusev, A.; Luther, F.; Sorvillo, N.; Rieben, R.; Brenner, P.; Walz, C.; Wolf, E.; Roshani, B.; Stahl-Hennig, C.; Abicht, J.M. Impact of porcine cytomegalovirus on long-term orthotopic cardiac xenotransplant survival. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Fiebig, U.; Abicht, J.M.; Mayr, T.; Längin, M.; Bähr, A.; Guethoff, S.; Falkenau, A.; Wolf, E.; Reichart, B.; Shibahara, T.; Denner, J. Distribution of Porcine Cytomegalovirus in Infected Donor Pigs and in Baboon Recipients of Pig Heart Transplantation. Viruses 2018, 10. [Google Scholar] [CrossRef]
- Jhelum, H.; Bender, M.; Reichart, B.; Mokelke, M.; Radan, J.; Neumann, E.; Krabben, L.; Abicht, J.M.; Kaufer, B.; Längin, M.; Denner, J. Evidence for Microchimerism in Baboon Recipients of Pig Hearts. Viruses 2023, 15. [Google Scholar] [CrossRef]
- Denner, J. Zoonosis and xenozoonosis in xenotransplantation: A proposal for a new classification. Zoonoses Public Health 2023, 70, 578–579. [Google Scholar] [CrossRef]
- Fiebig, U.; Holzer, A.; Ivanusic, D.; Plotzki, E.; Hengel, H.; Neipel, F.; Denner, J. Antibody Cross-Reactivity between Porcine Cytomegalovirus (PCMV) and Human Herpesvirus-6 (HHV-6). Viruses 2017, 9. [Google Scholar] [CrossRef]
- Mettenleiter, T.C.; Ehlers, B.; Müller, T.; Yoon, K.-J.; Teifke, J.P. Herpesviruses in, Diseases of Swine, 10th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Eds.; John Wiley & Sons, Inc.: Ames, Iowa, USA, 2012. [Google Scholar]
- Azab, W.; Dayaram, A.; Greenwood, A.D.; Osterrieder, N. How Host Specific Are Herpesviruses? Lessons from Herpesviruses Infecting Wild and Endangered Mammals. Annu. Rev. Virol. 2018, 5, 53–68. [Google Scholar] [CrossRef]
- Michaels, M.G.; Alcendor, D.J.; St George, K.; Rinaldo, C.R. Jr.; Ehrlich, G.D.; Becich, M.J.; Hayward, G.S. Distinguishing baboon cytomegalovirus from human cytomegalovirus: Importance for xenotransplantation. J. Infect. Dis. 1997, 176, 1476–1483. [Google Scholar] [CrossRef] [PubMed]
- Michaels, M.G.; Jenkins, F.J.; St George, K.; Nalesnik, M.A.; Starzl, T.E.; Rinaldo, C.R. Jr. Detection of infectious baboon cytomegalovirus after baboon-to-human liver xenotransplantation. J. Virol. 2001, 75, 2825–2828. [Google Scholar] [CrossRef] [PubMed]
- Degré, M.; Ranneberg-Nilsen, T.; Beck, S.; Rollag, H.; Fiane, A.E. Human cytomegalovirus productively infects porcine endothelial cells in vitro. Transplantation 2001, 72, 1334–1337. [Google Scholar] [CrossRef] [PubMed]
- Rausell, A.; Muñoz, M.; Martinez, R.; Roger, T.; Telenti, A.; Ciuffi, A. Innate immune defects in HIV permissive cell lines. Retrovirology 2016, 13. [Google Scholar] [CrossRef]
- Piroozmand, A.; Yamamoto, Y.; Khamsri, B.; Fujita, M.; Uchiyama, T.; Adachi, A. Generation and characterization of APOBEC3G-positive 293T cells for HIV-1 Vif study. J. Med. Invest. 2007, 54, 154–158. [Google Scholar] [CrossRef]
- Panda, D.; Dinh, P.X.; Beura, L.K.; Pattnaik, A.K. Induction of interferon and interferon signaling pathways by replication of defective interfering particle RNA in cells constitutively expressing vesicular stomatitis virus replication proteins. J. Virol. 2010, 84, 4826–4831. [Google Scholar] [CrossRef]
- Ding, S.; Diep, J.; Feng, N.; Ren, L.; Li, B.; Ooi, Y.S.; Wang, X.; Brulois, K.F.; Yasukawa, L.L.; Li, X.; Kuo, C.J.; Solomon, D.A.; Carette, J.E.; Greenberg, H.B. STAG2 deficiency induces interferon responses via cGAS-STING pathway and restricts virus infection. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef]
- Ferreira, C.B.; Sumner, R.P.; Rodriguez-Plata, M.T.; Rasaiyaah, J.; Milne, R.S.; Thrasher, A.J.; Qasim, W.; Towers, G.J. Lentiviral Vector Production Titer Is Not Limited in HEK293T by Induced Intracellular Innate Immunity. Mol. Ther. Methods Clin. Dev. 2019, 17, 209–219. [Google Scholar] [CrossRef]
- Unterholzner, L.; Keating, S.E.; Baran, M.; Horan, K.A.; Jensen, S.B.; Sharma, S. IFI16 is an innate immune sensor for intracellular DNA. Nat. Immunol. 2010, 11, 997–1004. [Google Scholar] [CrossRef]
- Staeheli, P.; Haller, O. Human MX2/MxB: a Potent Interferon-Induced Postentry Inhibitor of Herpesviruses and HIV-1. J. Virol. 2018, 92, e00709–18. [Google Scholar] [CrossRef]
- Li, X.D.; Wu, J.; Gao, D.; Wang, H.; Sun, L.; Chen, Z.J. Pivotal roles of cGAS-cGAMP signaling in antiviral defense and immune adjuvant effects. Science 2018, 341, 1390–1394. [Google Scholar] [CrossRef]
- Pindel, A.; Sadler, A. The role of protein kinase R in the interferon response. J. Interferon Cytokine Res. 2011, 31, 59–70. [Google Scholar] [CrossRef]
- Hovanessian, A.G. On the discovery of interferon-inducible, double-stranded RNA activated enzymes: the 2′-5′oligoadenylate synthetases and the protein kinase PKR. Cytokine Growth Factor Rev. 2007, 18, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Krug, R.M. Viral proteins that bind double-stranded RNA: countermeasures against host antiviral responses. J. Interferon Cytokine Res. 2014, 34, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.A.; Meurs, E.F.; Esteban, M. The dsRNA protein kinase PKR: virus and cell control. Biochimie 2007, 89, 799–811. [Google Scholar] [CrossRef]
- Munir, M.; Berg, M. The multiple faces of protein kinase R in antiviral defense. Virulence 2013, 4, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. Co-Cultivation Assays for Detecting Infectious Human-Tropic Porcine Endogenous Retroviruses (PERVs). Int J Mol Sci. 2025, 26(15), 7111. [Google Scholar] [CrossRef]
- Zhao P, Gu L, Gao Y, Pan Z, Liu L, Li X, Zhou H, Yu D, Han X, Qian L, Liu GE, Fang L, Wang Z. Young SINEs in pig genomes impact gene regulation, genetic diversity, and complex traits. Commun Biol. [CrossRef]
- Zheng M, Guo T, Yang B, Zhang Z, Huang L. Origin, evolution, and tissue-specific functions of the porcine repetitive element 1. Genet Sel Evol. [CrossRef]
- Fang X, Mou Y, Huang Z, Li Y, Han L, Zhang Y, Feng Y, Chen Y, Jiang X, Zhao W, Sun X, Xiong Z, Yang L, Liu H, Fan D, Mao L, Ren L, Liu C, Wang J, Li K, Wang G, Yang S, Lai L, Zhang G, Li Y, Wang J, Bolund L, Yang H, Wang J, Feng S, Li S, Du Y. The sequence and analysis of a Chinese pig genome. Gigascience. 2012, 1(1), 16. [Google Scholar] [CrossRef]
- Ichiyanagi K, Li Y, Watanabe T, Ichiyanagi T, Fukuda K, Kitayama J, Yamamoto Y, Kuramochi-Miyagawa S, Nakano T, Yabuta Y, Seki Y, Saitou M, Sasaki H. Locus- and domain-dependent control of DNA methylation at mouse B1 retrotransposons during male germ cell development. Genome Res. 2058. [CrossRef]
- Waterston RH, Lindblad-Toh K, Birney E, Rogers J, Abril JF, Agarwal P,Agarwala R, Ainscough R, Alexandersson M, An P, et al. . Initial sequencing and comparative analysis of the mouse genome. Nature 2002, 420, 520–562. [Google Scholar] [CrossRef]
- Denner, J. What does the PERV copy number tell us? Xenotransplantation. 2022 Mar;29(2):e12732. [CrossRef]
Figure 1.
Schematic overview of the cocultivation experiments with PCMV/PRV-producing PFT cells. The timeline refers to co-cultivation with human 293hyg cells; for the other cell lines, small variations in the number of days were applied.
Figure 1.
Schematic overview of the cocultivation experiments with PCMV/PRV-producing PFT cells. The timeline refers to co-cultivation with human 293hyg cells; for the other cell lines, small variations in the number of days were applied.
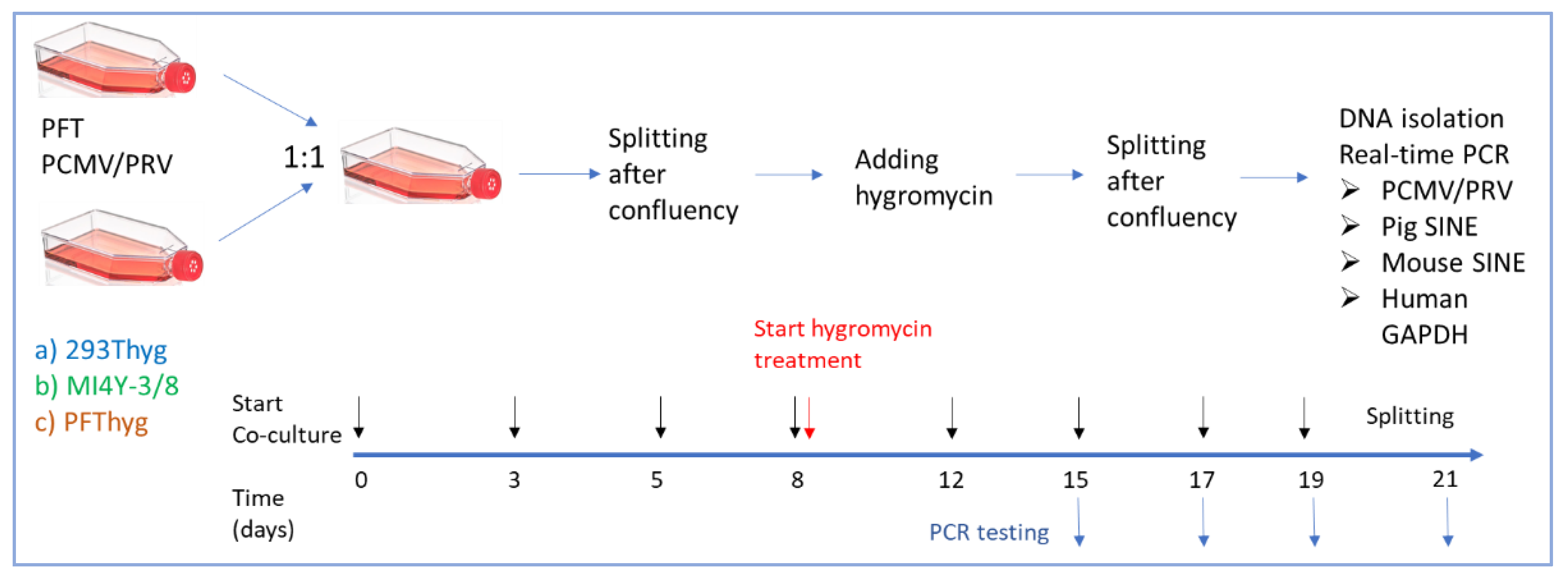
Figure 2.
Morphology of the used cell lines before and after co-cultivation. a, uninfected PFT cells; b, PFT cells producing PCMV/PRV; c, untreated 293T cells; d, 293 cells after co-culture with PFT cells producing PCMV/PRV and selection; e, untreated MI4Y-3/8 cells; f, MI4Y-3/8 cells after co-culture with PFT cells producing PCMV/PRV and selection. Scale bar indicated is 100 µm and magnification is 10x. Large round PCMV/PRV producing cells were indicated by arrow.
Figure 2.
Morphology of the used cell lines before and after co-cultivation. a, uninfected PFT cells; b, PFT cells producing PCMV/PRV; c, untreated 293T cells; d, 293 cells after co-culture with PFT cells producing PCMV/PRV and selection; e, untreated MI4Y-3/8 cells; f, MI4Y-3/8 cells after co-culture with PFT cells producing PCMV/PRV and selection. Scale bar indicated is 100 µm and magnification is 10x. Large round PCMV/PRV producing cells were indicated by arrow.

Figure 3.
Morphology of the PFThyg cells before, during and after co-cultivation with PFT cells producing PCMV/PRV. a, uninfected PFThyg cells; b, PFThyg cells co-cultured with PFT cells producing PCMV/PRV; c, PFThyg cells after selection by hygromycin. Scale bar is 100 µm and magnification is 10x.
Figure 3.
Morphology of the PFThyg cells before, during and after co-cultivation with PFT cells producing PCMV/PRV. a, uninfected PFThyg cells; b, PFThyg cells co-cultured with PFT cells producing PCMV/PRV; c, PFThyg cells after selection by hygromycin. Scale bar is 100 µm and magnification is 10x.
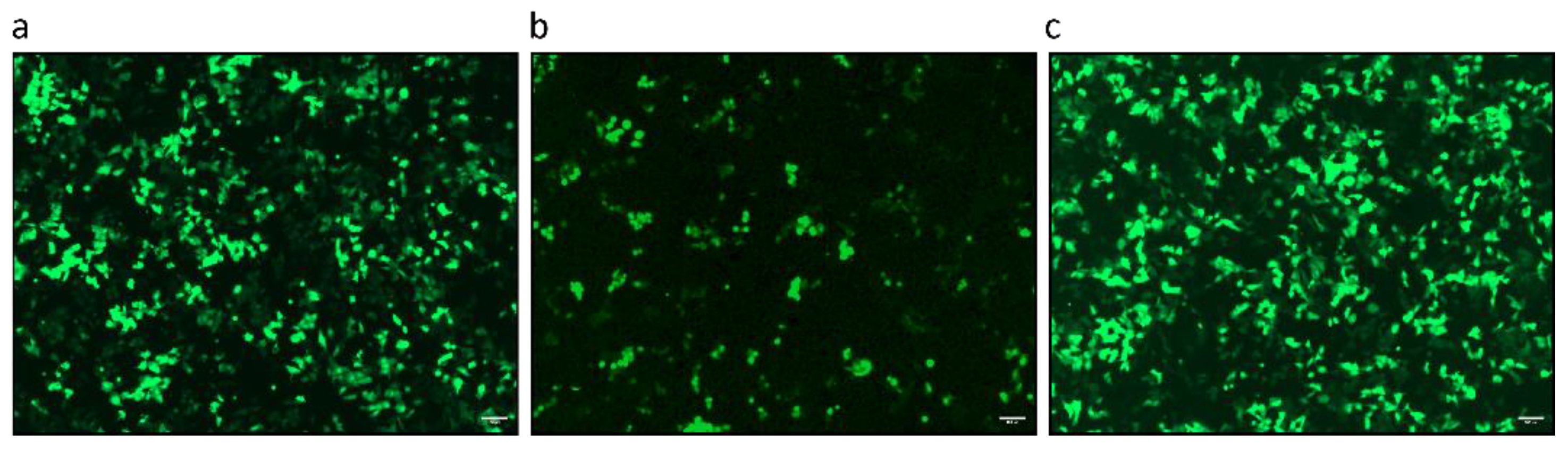

Table 1.
Primers and probes.
| Primers and Probes used for real time-PCR | Sequence 5′-3′ | Nucleotide Position | Accession Number | Reference |
| PCMV/PRV fw PCMV/PRV rev PCMV/PRV probe |
ACTTCGTCGCAGCTCATCTGA GTTCTGGGATTCCGAGGTTG 6FAM-CAGGGCGGCGGTCGAGCTC-BHQ1 |
45206 – 45226 45268 – 45249 45247 - 45229 |
AF268039 | Mueller et al., 2002 [26] |
| hGAPDH fw hGAPDH rev hGAPDH probe |
GGCGATGCTGGCGCTGAGTAC TGGTTCACACCCATGACGA HEX-CTTCACCACCATGGAGAAGGCTGGG-BHQ1 |
3568–3587 3803–3783 3655–3678 |
AF261085 | Behrendt et al., 2009 [27] |
| pGAPDH fw pGAPDH rev pGAPDH probe |
GATCGAGTTGGGGCTGTGACT ACATGGCCTCCAAGGAGTAAGA HEX-CCACCAACCCCAGCAAGAG-BHQ |
1083–1104 1188–1168 1114–1137 |
NM_001206359.1 | Duvigneau et al., 2005 [28] |
| PRE-1 fw PRE-1 rev PRE-1 probe |
GACTAGGAACCATGAGGTTGCG AGCCTACACCACAGCCACAG FAM-TTTGATCCCTGGCCTTGCTCAGTGG-BHQ1 |
37–58 61–85 151–170 |
Y00104 |
Walker et al., 2003 [29] |
| SINE B1 fw SINE B1 rev SINE B1 probe |
TGGCGCACGCCTTTAATC TGGCCTCGAACTCAGAATCC 6FAM-ACTCGGGAGGCAGAGG-BHQ1 |
n.a. | n.a. | Gualtieri et al., 2013 [30] |
| PERV-C fw PERV-C rev |
CTGACCTGGATTAGAACTGG ATGTTAGAGGATGGTCCTGG |
6606-6625 6867-6886 |
AM229312 | Takeuchi et al., 1998 [31] |
n.a., not applicable.
Table 2.
Detection of PCMV/PRV, and PERV-C in PFT cell lines (gene copy number per 100 ng DNA in the case of PCMV/PRV and negative as tested by conventional PCR in the case of PERV-C).
Table 2.
Detection of PCMV/PRV, and PERV-C in PFT cell lines (gene copy number per 100 ng DNA in the case of PCMV/PRV and negative as tested by conventional PCR in the case of PERV-C).
| Cell line | PCMV/ PRV | PERV-C | pGAPDH |
| PFT | n.d. | - | 105.4 |
| PFT-PCMV/PRV | 104 | - | 105.4 |
n.d., not detected.
Table 3.
Screening for PCMV/PRV in the supernatant and in the pelleted virus fraction (gene copy number per 50 ng DNA).
Table 3.
Screening for PCMV/PRV in the supernatant and in the pelleted virus fraction (gene copy number per 50 ng DNA).
| Supernatant | PCMV/ PRV |
| Direct | n.d. |
| Virus pellet | 102.4 |
n.d., not detected.
Table 4.
Real-time screening for PCMV/PRV infection in co-culture experiments after 21 days (gene copy number per 100 ng DNA).
Table 4.
Real-time screening for PCMV/PRV infection in co-culture experiments after 21 days (gene copy number per 100 ng DNA).
| Cell lines |
PCMV PRV |
PRE-1 (porcine SINE) |
SINE B1 (murine SINE) |
Human GAPDH |
| PFT PCMV/PRV cells | 104 | 108 | n.a. | n.a. |
| PFThyg cells after co-culture with PFT PCMV/PRV cells and subsequent selection | 104 | 108 | n.a. | n.a. |
| 293Thyg cells after co-culture with PFT PCMV/PRV cells and subsequent selection | n.d. | n.d. | n.a. | 105.2 |
| MI4Y-3/8 cells after co-culture with PFT PCMV/PRV cells and subsequent selection | n.d. | n.d. | 106.9 | n.a. |
Table 5.
Detection of PCMV/PRV in PFT cell line treated with infected culture supernatant after 25 days (gene copy number per 100 ng DNA).
Table 5.
Detection of PCMV/PRV in PFT cell line treated with infected culture supernatant after 25 days (gene copy number per 100 ng DNA).
| Cell line | PCMV/ PRV | pGAPDH |
| PFT cells after treatment with supernatant from PCMV/PRV-producing cells | 103.4 | 104.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



