
Submitted:
18 November 2025
Posted:
18 November 2025
You are already at the latest version
Abstract
Background: Breast carcinoma represents the most frequent malignancy in women in Canada. The intrinsic or acquired drug resistance significantly contributes to in-creased risk of recurrence and metastasis. Although, front-line therapy is multimodal, chemoresistance remains a major hurdle in treatment and therapy. Intake of natural compounds issued from the fermentation processes are now considered a god strategy to help overcome chemoresistance. Materials and Methods: Extracellular vehicles (EVs) from Mesenchymal Stro-mal/Stem Cells (MSCs) pretreated with Lentinula edodes cultured extract (AHCC) were used to study the effect on reducing chemoresistance and modulating mi-croRNAs in cell lines, MCF-7 and MCF-7/DOX cells by EVs derived from MSCs under AHCC treatment. Characterization of EVs was done by using nanoparticles tracking analysis. MicroRNas and the formation of cancer stem cells were studied. Results: miRNA analysis revealed that AHCC remarkably affected the expression of several microRNAs, amongst which are miR-155, miR-34a, miR-Let7 and miR-200c. In vitro experiments showed inhibition of cancer stem cell proliferation after challenging the cells with EVs pretreated with AHCC. Conclusion: Our data demonstrated that AHCC may contribute to the modulation of tumor microenvironment, thus influenc-ing the development of cancer stem cells.
Keywords:
Lentinula edodes
; extracellular vesicle
; cancer stem cells
; mesenchymal stem cell
; microRNAs
1. Introduction
Breast cancer is the most common cancer in Canadian women, except for non-melanoma skin cancer [1]. It accounts for approximately 25% of new cancer cases and 13% of all cancer deaths in Canadian women. In 2024, an estimated 310,720 women in the United States will be diagnosed with invasive breast cancer [2]. It is estimated that breast cancer ranks the second most diagnosed malignancy in Canada after lung cancer, and first in cancer related to female However, ranked as number one according to the Canadian cancer society statistics [3]. The current treatment protocol depends on the histology and stage of the cancer and consists of total resection of tumor mass followed by different modalities included: radical chemo-radiotherapy, hormone therapy, immunotherapy and targeted therapy. Patients may receive a combination or all of these modalities, and while some patients are responsive initially to this regimen, the likelihood of the recurrence is extremely high [4,5]. Moreover, metastasis and development of drug resistance are the major challenges in breast cancer therapy.
Mounting evidence, suggests that a sub-population of tumor cells with the self-renewal properties is the main culprit of the drug resistance, metastasis and recurrence in many types of cancer [6]. Breast cancer also contains these subsets of cells functionally with stemness properties named "breast cancer stem cells" (BCSCs) [7,8]. This small tumor sub-population significantly contribute to the tumor reconstitution after therapy and resistance to therapy [9]. Furthermore, chemotherapy drugs used in treating breast cancer have significant side effects since they have the potential to harm healthy cells, leading to a range of adverse effects[10].
Hence, it is plausible that breast cancer patients may potentially benefit from including nutritional interventions alongside conventional therapies through specifically targeting the regulation of cancer stem cell proliferation [11,12]. There has been an increasing focus on the potential usage of non-toxic natural compounds as chemo-preventive medicines for patients susceptible to tumor recurrence and/or have a high risk of metastasis. The possible applications of mushroom extracts in cancer prevention have garnered heightened interest in recent years [13,14].
Research has demonstrated that an extract derived from cultured Lentinula edodes mycelia exhibits inhibitory effects on several forms of cancer, including breast cancer, as observed in both animal models and clinical investigations[15,16,17,18]. There is a growing body of evidence indicating that the compound referred to as AHCC has the potential to regulate the growth of cancer stem cells by the anti-inflammatory and epigenetic processes. It was reported that AHCC is capable on increasing tumor suppressor miR-335, thus avoiding immune system evasion seen in cancer cells behavior [19,20].
Moreover, numerous studies related to AHCC’s effects on immune cells modulation have reported its effects on increased natural killer cell activity, [21] decrease tumor formation [22], enhanced T cell proliferation [23], altered cytokine release via altered T cell activity[22,24,25], anti-inflammatory, and anti-oxidant properties [26,27,28]. Furthermore, a significant body of evidences derived from clinical investigations proved safety, feasibility and efficacy of this supplement in healthy volunteers [29,30] as well as patients with malignancies [31,32]. Evidence also indicate that AHCC regulate immune response by sensitizing the intestinal epithelium, that is facilitated through recognition of non-pathogenic food-associated molecular patterns (FAMPS) by TLR4 and TLR2 receptors [33].
In addition to the AHCC’s potential impacts on immune cells, emerging evidences noted that the AHCC’s may also play a role in reprogramming tumor microenvironment [34]. This includes impacting on miRNAs, small non-coding RNAs that regulate genes by posttranscriptional regulation of gene expression, involved in self-renewal, progression, and metastasis [34,35]. They can be categorized based on their function, into two distinct groups: oncomiRs that promote tumor formation and progression. Studies shown that oncomiRs are differentially expressed in cancer cells and the tumor microenvironment [36], whereas miRsupps that induce apoptosis and inhibit proliferation, invasion and growth, tumor growth [37].
We have previously showed that AHCC could target CSCs via epigenetic reprograming and altering some key micro-RNAs and reduce functionally enriched stem cells growth and sphere formation [19]. The aim of this study was to analyze the impact of AHCC on modulation of MSCs by indirectly modulating sphere formation in sensitive and chemoresistance breast cancer cell lines. The auspicious features of MSCs which came from their regenerative abilities and plasticity’s into different cell lineage, make them as an good candidate to better understand the dynamic of interaction between MSCs- CSCs and tumor environment [38,39]. A glance at the clinical trial database shows that more than ten thousand trials have been registered using MSCs as the therapeutic agent in different clinical applications [40,41,42]. However, the application of these cells in cancer treatment remains controversial. While there is lot of evidence highlighting the potential roles of MSC in cancer progression and pathogenesis, emerging data reveals some beneficial effects on cancer treatment. Regarding tumor environment, MSCs can exert mutually exclusive effects on surrounding cells including cancer cells. In some cases, naïve MSCs showed tumor suppression through the activity of Wnt signaling inhibitor protein Dickkopf-related protein 1 (DKK1) [43,44,45]. This consequently leads in c-Myc and Cyclin-D2 downregulation [46]. Moreover, it has been showed that naïve MSCs can inhibit angiogenesis and promote the apoptosis of vascular endothelial cells [47]. On the other hand, reports confirming the adverse effect of these cells on tumor cells by up-regulating Akt/mTOR signaling pathway, enhancing angiogenesis and promoting metastasis [48,49]. MSCs communicate with cancer or non-cancer cells in tumor microenvironment by releasing EVs. EVs are nano-size, membranous particles which can play an important role as a bio-vehicle generated from original cells to the recipient cells and consequently changing the fate and physiology of the recipient cells [50]. The unique characteristics of the EVs have created a paradigm shift for their usage as therapeutic agent, [51,52]. EVs derived from unmodified MSCs have demonstrated significant potential in various experimental and clinical studies and showed promising therapeutic effects [53,54].
Of relevance for breast cancer is the bipartite relationship of MSC-derived EVs on the cancer development and progression. Tumor homing capacity of EVs make these vesicles as the attractive agents for potential tumor-targeted delivery vehicles [55]. Several studies reported the anti-proliferative effects of MSC-derived EVs against ovarian, bladder, hepatoma and sarcoma tumor cells [56,57,58]. Furthermore, it has been reported that MSC-derived EVs transport different kinds of proteins, metabolites and miRNAs to the cancer cells and alter the behavior and fate of the target cells through releasing of their contents [59,60,61]. Therefore, MSC-derived EV has been proposed as a promising alternative for MSCs for tackle with various kinds of cancers including breast cancer [61,62].
The present study was conducted to evaluate the epigenetic anti-proliferative effects of AHCC-treated MSCs derived EVs on MCF-7 (wild type) and MCF-7 /DOX cell lines.
Therefore, we evaluated the effect of conditioned media of MSCs exposed to AHCC on cancer stem cells and the differentially expressed miRNA profile, including oncogenic (miR-155) and tumor suppressor miRNAs (miR-34a, miR-Let7, miR-200c).
2. Materials and Methods
2.1. Chemical and Reagent
AHCC® (lot S26-0530-2; Amino Up Co., Ltd., 363-32 Shin-ei, Kiyota-ku, Sapporo, Japan) is a standardized extract of cultured Lentinula edodes mycelia, provided by Amino Up Co., Ltd. (Sapporo, Japan).The extract was prepared in Dulbecco’s modified Eagle’s (DMEM) provided from Gibco, Grand Island, NY, USA for adherent cell culture and DMEM-F12 (Invitrogen), for Sphere Culture, at the 40mg/mL concentration and filtered through a 0.22 µm Millex-GV filter.
2.2. Cell Culture
Human MSCs cultured on T75 tissue culture flask in DMEM, supplemented with 10%FBS and 1% penicillin-streptomycin (Invitrogen, Carlsbad, CA, USA) in a humidified chamber with 5%. The medium was refreshed every other day, and Cells were observed daily. Serial passages were done when confluent monolayer formed.
The MCF7wt was purchased from American Type Cell Collection (ATCC; Chicago, IL). MCF7/DOX (kindly gifted from Dr. Amado M. Parissenti). The cells were maintained in Dulbecco's Modified Eagle's medium (DMEM) containing Fetal bovine serum (FBS) (10%, v/v) (Sigma-Aldrich, Oakville, ON, Canada), penicillin/streptomycin (0.05 mg/mL) at 37° C in a humidified atmosphere with 5% CO2 inside CO2 incubator.
2.3. Spheroid Culture and Growth for Initial Assessment of AHCC
Adherent cells were detached by trypsin and single cells were counted using Countess (Invitrogen). Single cells were plated in ultralow attachment 96-well plates (Corning, NY, USA) at 0.3x105 cells/0.2 ml/well in DMEMF12 (Invitrogen), supplemented with 10 ng/ml EGF, 20 ng/ml bFGF, 5 µg/ml insulin, 1 mM sodium pyruvate, 0.5 µg/ml hydrocortisone, and penicillin/streptomycin (0.05 mg/mL). Cells were treated with 4 mg/ml AHCC and grown in as non-adherent conditions were counted after 2-3 days For sphere culture from the cell lines(MCF-7,MCF-7/Dox), 0.3×105 freshly trypsinated detached single cells were suspended and seeded in any wells of the 6-well ultra-low attached flask (Corning, NY, USA) in serum free DMEN-F12(1:1) supplemented with 250 mM L-glutamine (Sigma Aldrich, Oakville, ON), 100mM Sodium pyruvate, 10 mg/mL insulin (ThermoFisher) , 20 ug/mL bFGF (ThermoFisher) , 20 ug/mL EGF (ThermoFisher) , 1000x streptomycin-penicillin (Sigma Aldrich) and 100 ug/mL hydrocortisone (Sigma Aldrich), in accordance with our previous published experiments. Spheres were cultured in two groups: the first group were supplemented with the media as the control and the second were exposed to 4 mg/ml AHCC as the treatment group. Both groups were incubated in a humidified incubator (5% CO2 at 37 °C).
2.4. Purification Characterization and Modification of EVs
The passages 3-8 of MSCs were cultured in up to 70% cell confluency to isolate EVs. The FBS-enriched medium was replaced with MSC basal medium supplemented with EV-free FBS for the control group. The MSCs were treated with EV-free FBS enriched media plus 3 mg/ml AHCC as the experimental group. Forty-eight hours post-incubation with EV-free medium, the conditioned medium (CM) of each group was collected and stored at -80˚C to purify EVs. Both groups' condition media were collected 48 h post-incubation with EV-free FBS CM and EV-free FBS CM+AHCC and store at -80°C. To purify EVs, we used the exoEasy Maxi kit (Qiagen 76204). The isolated EVs have been homogenized, pooled, and aliquoted to be used for the experiments. A sample of EVs was used for protein content assay and nanoparticle tracking analysis (NTA). The protein concentration of EVs was measured using a BCA protein assay kit (Pierce, ThermoFisher).
2.5. Sphere Culture and Administration of Modified EVs
For EV administration in sphere culture, both cell lines cultivated in the same condition as mentioned above. Spheres were cultured in two groups: the first group treated with MSC-derived EVs as the control and the second were exposed to AHCC treated MSC-EVs as the treatment group Immediately after seeding single cells. Both groups were incubated in a humidified incubator (5% CO2 at 37 °C).
2.6. In-Vitro Assessment of the Immunostimulatory Effect of AHCC on Breast Cancer Cell Lines
Three samples of MCF-7 wild type and MCF-7/DOX cells were grown in a 6-well plate at a concentration of 0.3 × 106 cells/ml in DMEM complete medium. The cells were then exposed to concentrations of 4 mg/ml AHCC for 24 hours as we previously established. The control groups for each cell line were also grown in an equal volume of medium. Cells were harvested for micro-RNA profiling in next steps.
2.7. Quantitative RT-PCR for MicroRNA Profiling
RNAs were extracted from three samples of adherent and sphere cultures from both groups (control group and AHCC exposed sphere group) using Trizol protocol, miRNeasy mini kit according to manufacturer’s protocol (Qiagen, Toronto, ON). The quality and quantity of extracted RNA were assessed by Nano-drop (Nanodrop Technologies, Wilmington, DE, USA).
Then, complementary DNA (cDNA) was synthesized from 2ng/µl of 1:60 diluted of the total RNA using reverse transcription miRCURY LNA RT(Qiagen,USA).The quantitative RT- PCR was carried out with the Syber-Green (Qiagen, Toronto, Canada) in accordance with the RT-PCR system and standard PCR conditions in a CFX96 machine (Bio-Rad). Primers for miR-155, miR-34a, let-7a, and miR-200c were obtained from the miRCURY LNA miRNA PCR assay (Qiagen, Toronto, Canada) as described in previous studies. These were used to assess changes in expression levels relative to the internal control SNORD65 (mmu) [63,64,65,66].
2.8. Statistical Analysis
Statistical analysis of miRNA expression was performed using GraphPad Prism software (version 5). Data was presented as mean ± SEM from at least three independent experiments. We utilized unpaired t test to evaluate the statistical significance of differences between groups. Differences were statistically significant when value < 0.05.
3. Results
3.1. Regulatory Effects of AHCC on Tumor Suppressor miRNA and Oncogenic miRNA Expression in Breast Cancer Cell Lines
In this investigation, we assessed the expression of miRNA in cultures obtained from wild-type MCF-7 and MCF-7/DOX cell lines. First, these cell cultures were subjected to treatment with AHCC at a dose of 4 mg/mL for a duration of 24 hours. A distinct expression pattern of microRNAs (miRNAs), namely miR-155, miR-34a, miR-200c, and miR-Let7a, was detected in the experimental group compared to the control group. It is worth mentioning that the expression of miR-155 was dramatically reduced in the MCF-7 cells treated with AHCC in comparison to the control group, as illustrated in Figure 1a.
In contrast, there was a significant increase in the expression levels of tumor-suppressive microRNAs, including miR-34a, miR-Let7a, and miR-200c. the differentially expressed miRNAs were also analyzed in MCF7/DOX cells treated with AHCC with similar pattern expression (Figure 1b).
Quantitative RTPCR assessments of miRNA expression in wild-type MCF-7 following treatment with 4 ug/mL AHCC. Panels A, B and C demonstrate the significant upregulation of the tumor-suppressive miRNAs; miR-Let7a, miR-200c and miR-34a respectively, whereas Pan D shows the downregulation of the oncogenic miR-155 under the same conditions. Figure 1b- AHCC enhances tumor-suppressive miRNAs and downregulates miR-155 in MCF-7/DOX cells. Comparative expression analysis of miRNAs in MCF-7/DOX cell lines chronologically following treatment with 4 ug/mL dosage of AHCC. Panel A shows higher expression levels of Let7a miRNA, Panel B, shows significant higher expression of miR34-a as compared with the control group, and C depicted the increased levels of, miR-200c, panel D statistical graph show the expression of miR - 155 which significantly got down.
3.2. Purification and Characterization of EVs
The results of the BCA assay demonstrated a significant increase in the protein content of EVs formed from MSCs treated with AHCC. The protein concentration in the EVs from AHCC-treated MSCs was four times higher compared to EVs from untreated MSCs, with values of 0.66 µg/µL and 0.167 µg/µL, respectively. The observed increase was additionally supported by the utilization of nanoparticle tracking analysis (NTA), a technique that quantifies light scatters from EVs. The results of NTA demonstrated that EVs derived from MSCs treated with AHCC exhibited a nearly two-fold rise in particle count compared to EVs derived from untreated MSCs as presented in Figure 2 (1.2×10¹⁰ particles/mL for AHCC-treated EVs versus 5.7×10⁹ particles/mL for untreated MSC-derived EVs).
3.3. Inhibitory Effects of AHCC-Treated MSC-Derived EVs on Colony Formation and Sphere Growth in MCF7 and MCF7-Dox Cells
The impact of EVs derived from MSCs on the colony-forming abilities in MCF7 and MCF7-Dox cells was also assessed. The control group consisted of EVs obtained from untreated MSCs, whereas the experimental group consisted of EVs derived from MSCs treated with AHCC. The findings of the study demonstrated that the presence of AHCC-treated MSC-derived EVs had a suppressive effect on colony formation and sphere growth. This observation was made during a 24-hour timeframe. Subsequent inquiries were undertaken to unravel the underlying processes responsible for the observed inhibitory impact, focusing on the examination of miRNA profiles.
3.3. Modified EVs Delivered Their Cargo and Changed the Expression Level of Target miRNAs
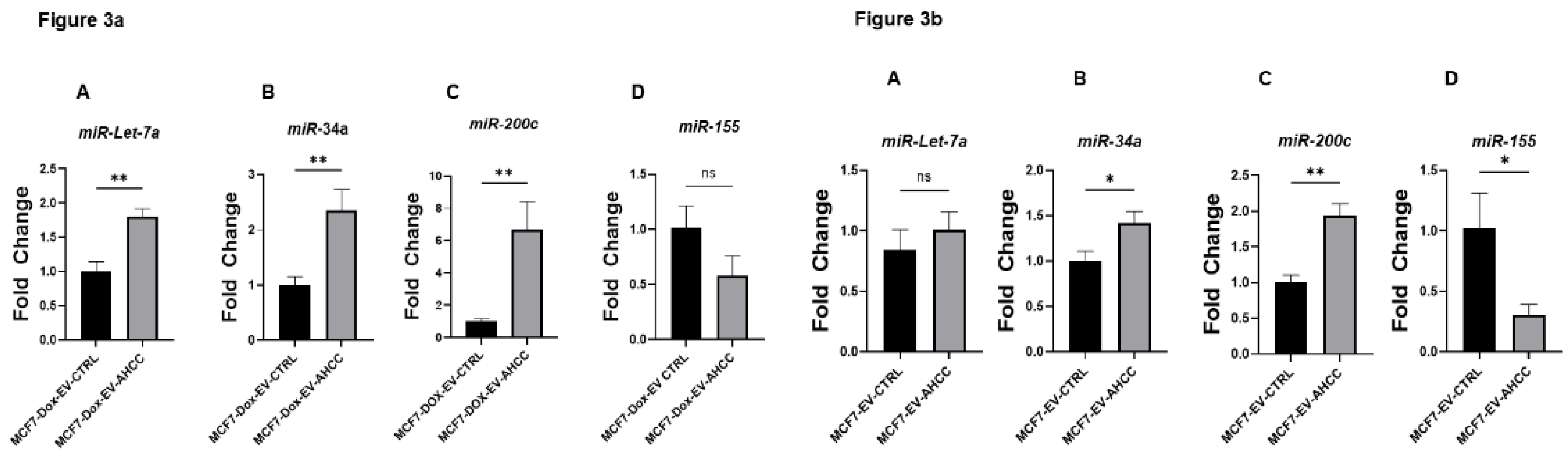
Using the sphere culture model, we investigated the effects of AHCC-modified MSC-derived EVs on miRNA expression in MCF7 WT and MCF7-Dox cell cultures. Naïve MSC-derived EVs were used as the control in our experimental design. Different quantities of EV proteins were utilized (5 µg/ml, 10 µg/ml, 15 µg/ml) to conduct miRNA profiling. The results revealed that there was differential expression observed for all the targeted miRNAs in both cell lines when they were subjected to non-attached growing circumstances. Significantly, the administration of 10 ug/ml AHCC-modified MSC-derived EVs resulted in a substantial upregulation of miR-34a, miR-200c, and miR-let7a expression levels inside the MCF7-Dox sphere cultures, in contrast to the MCF7-Dox sphere cultures treated with untreated MSC-derived EVs. Although the decrease in miR-155 expression did not reach statistical significance in the MCF7-Dox spheres, a discernible downward trend was detected at a dose of 10 µg/ml (Figure 3a). In contrast, a notable decrease in the expression of miR-155 was observed at a concentration of 10 µg/ml in the MCF7 spheres that were treated with extracellular vesicles produced from AHCC-modified MSCs, compared to the control group. Moreover, notable elevations in the expression of miR-200c and miR-34a were detected in MCF7- WT spheres (Figure 3b).
4. Discussion
In the continuous battle against cancer, primary, secondary and tertiary prevention and integrative medicine have emerged as crucial components. There is a growing trend among breast cancer patients to include evidence-based alternative and complementary medicine into their treatment protocols. In recent times, the extract of Cultured Lentinula edodes Mycelia has garnered considerable attention as as evidence-based supported component in the field of preventative medicine. A significant proportion of breast cancer patients are embracing its utilization due to its potential to regulate the immune system and function as a chemo-preventive agent [67,68]. The extract is particularly enriched with the acetylated form of the α-1,4 glucan and other bioactive compounds released following the process of culture or fermentation [67,69]. Of interest, we have previously shown that it can influence CSC via modulating miR-335 [19].
miRNAs play important functions in gene regulation expression in normal and disease conditions [70]. Several studies have shown that dysregulation in miRNA expression results in tumor development through mechanisms involved in gene expression manipulation, defects in miRNA biogenesis, and dysregulated cell microenvironment [71]. MicroRNAs may act as either tumor suppressors or oncogenes [72]. In this study, four miRNAs were targeted:oncogenes miR-155, and tumor suppressor categories: miR-34a, miR-200c, and miR-Let7a.
miR-155 is an oncomir known to act as an oncogene by inhibiting anti-tumoral activity of p53 [63]. miR-155 is significantly upregulated in breast cancer and positively correlated with high tumor grade, progressive stage and metastases and it is inversely correlated with overall and disease-free survival [73]. miR-155 plays an important role in the immunopathogenesis of breast cancer. Expression of miR-155 is also promoted by factors that induce tumor inflammation, like interleukin IL6 and interferon IFN-γ [74]. AHCC was able to downregulate this oncomiR in CSC of chemo-resistance cell line. The results showed that modified EVs can effectively decrease oncomiR-155. The miR-34a is a tumor suppressor linked to cancer progression prevention [64]. Therefore, down-regulation of miR-34a expression correlates with cancer progression. For instance, aberrant CpG methylation of miR-34a promoter showed a significant reduction its expression in several cancers [75] . A growing body of evidence has pointed to the role of mir-34a in cell cycle, apoptosis, and mitotic signaling [76] via the regulation of several genes involved in stemness phenotype maintenance, chemo-resistance, and metastasis [75,77]. In this regard, it has been shown that oncogenic pathways (Wnt, Notch) and transcription factors such as c-Myc, Oct4, and Klf4 downregulate miR-34a expression in many types of cancers [78,79,80]. Moreover, computational prediction and some experimental studies represent c-Myc, as a target for miR-34a [81]. In the current study, treating tumourspheres with AHCC led to significant upregulation of miR-34a in MCF-7 and MCF-7/DOX spheres as depicted in figures (1,2,4,5).
The let-7 family is a family of miRNAs first studied [82]. Down-regulation of let-7 expression has shown to have a prognostic effect on lung cancer patients' survival after surgery [82]. Many members of the let-7 family are considered as the tumor suppressor; however, let-7a is the most studied miRNA, with impact on oncogenic c-Myc [83]. Our data indicates that AHCC upregulates let-7 expression in a dose-dependent manner and affects sphere formation effectively in both wild and chemo resistant cell lines cell lines (1,2,4,5).
The miR-200c is a member of the miRNAs family, which has shown to be a tumor suppressor in some cancers, including breast cancer [65]. It regulates epithelial-mesenchymal transition (EMT) via ZEB1/2 downregulation and E-cadherin upregulation [66]. There is also evidence that miR-200c is a novel regulator of TLR4 and HMBG-1 signaling and plays essential roles in cell cycle control, proliferation, invasion, apoptosis, and metastasis in different cancer types [84]. We assessed the expression of miR-200c in MCF-7, and MCF-7/ Dox treated with AHCC after 24 hours. Our data confirms the AHCC effect on miR-200c expression, as shown in figures 3-6.
AHCC is recognized for its system modulation and preventive effects against cancer. Our study emphasizes its influence on oncomiRs and tumor suppressing miRNAs like miR 155, miR 34a, miR-let7a and miR 200c. It shows that AHCC reduces the expression of oncomiR miR-155, which hinders the tumor suppressor p53 and is linked to tumor stages and spread. Moreover, AHCC boosts the levels of tumor suppressing miRNAs such as miR-34a, miR-let7a and miR-200c which play roles in impeding cancer advancement, epithelial mesenchymal transition and inhibiting cancer growth.
In summary, the research highlights the impact of AHCC in reducing chemoresistance and modulating miRNAs in breast cancer. Through the MSC derived EVs.
5. Future Investigation
Exploring how AHCC interacts with pathways related to cancer progression or inhibition; Future investigations should aim at unraveling the connections between AHCC and other cancer-related pathways, like p53 signaling, Wnt/β catenin, Notch and Hedgehog.
Investigating the impact of AHCC on regulation in breast cancer cells involves studying how AHCC influences DNA methylation/demethylation patterns and histone modifications on transcription factors. Additionally exploring how AHCC affects the characteristics of CSCs includes examining its influence on CSC maintenance, differentiation and self-renewal abilities in relation, to related transcription factors. In-vivo modeling of AHCC treatment: Exploit animal models to assess the therapeutic efficacy of AHCC, on tumor growth and metastasis prevention. Evaluate the potential synergic effect with check point inhibitors like PD1/PDL1 and CTLA4 to obtain the best results of immunotherapy.
Author Contributions
Conceptualization, Farzaneh Sharifzad and Chantal Matar; methodology, Farzaneh Sharifzad; validation, Farzaneh Sharifzad, Hamed Yasavoli, Roqia Shahbazi, and Dylen Burger; formal analysis, Farzaneh Sharifzad; investigation, Farzaneh Sharifzad, Hamed Yasavoli, Roqia Shahbazi, and Dylen Burger; NTA analysis and related experiments, Farzaneh Sharifzad and Nawal Alsadi; writing-original draft preparation, Farzaneh Sharifzad and Nawal Alsadi; writing-review and editing, Chantal Matar; supervision, Chantal Matar; project administration, Farzaneh Sharifzad. All authors have read and agreed to the published version of the manuscript. Chantal Matar is the corresponding author. All authors have read and agreed to the published version of the manuscript.”
Funding
This research received no external funding. The APC was funded by Chantal Matar.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The data presented in this study are available on request from the corresponding author.
The authors have reviewed and edited the output and take full responsibility for the content of this publication.”
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| EVs | Extracellular vehicles |
| MSCs | Mesenchymal Stromal/Stem Cells |
| BCSCs | Breast cancer stem cells |
| FAMPS | Food-associated molecular patterns |
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef]
- society, A.c. Cancer Facts & Figures 2024. Cancer Facts & Figures 2024 2024 [cited 2024 2024-10-12]; Available from: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/2024-cancer-facts-figures.html.
- in, C.C.S.A.C. , et al. Canadian statistic 2023 [cited 2024-02-17; Available from: https://cdn.cancer.ca/-/media/files/research/cancer-statistics/2023-statistics/2023_pdf_en.pdf?rev=7e0c86ef787d425081008ed22377754d&hash=DBD6818195657364D831AF0641C4B45C&_gl=1*1scrz89*_gcl_au*NDE0NDE3MTI1LjE3MDc3OTc3NTU.
- Agarwal, G. and P. J.B.c. Ramakant, Breast cancer care in India: the current scenario and the challenges for the future. 2008, 3, 21–27. [Google Scholar]
- Ho, P.J.; Ow, S.G.W.; Sim, Y.; Liu, J.; Lim, S.H.; Tan, E.Y.; Tan, S.-M.; Lee, S.C.; Tan, V.K.-M.; Yap, Y.-S.; et al. Impact of deviation from guideline recommended treatment on breast cancer survival in Asia. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Zhou, Y.; Xie, T.; Yuan, Y.; Li, H.; Shi, W.; Zheng, L.; Li, X.; Zhang, W. Tumor microenvironment of cancer stem cells: Perspectives on cancer stem cell targeting. Genes Dis. 2024, 11, 101043. [Google Scholar] [CrossRef]
- Kakarala, M. and M. S.J.J.o.c.o.o.j.o.t.A.S.o.C.O. Wicha, Implications of the cancer stem-cell hypothesis for breast cancer prevention and therapy. 2008, 26, 2813. [Google Scholar]
- Gooding, A.J.; Schiemann, W.P. Epithelial–Mesenchymal Transition Programs and Cancer Stem Cell Phenotypes: Mediators of Breast Cancer Therapy Resistance. Mol. Cancer Res. 2020, 18, 1257–1270. [Google Scholar] [CrossRef] [PubMed]
- Sharifzad, F.; Ghavami, S.; Verdi, J.; Mardpour, S.; Sisakht, M.M.; Azizi, Z.; Taghikhani, A.; Łos, M.J.; Fakharian, E.; Ebrahimi, M.; et al. Glioblastoma cancer stem cell biology: Potential theranostic targets. Drug Resist. Updat. 2019, 42, 35–45. [Google Scholar] [CrossRef]
- Hass, H.G.; Seywald, M.; Wöckel, A.; Muco, B.; Tanriverdi, M.; Stepien, J. Psychological distress in breast cancer patients during oncological inpatient rehabilitation: incidence, triggering factors and correlation with treatment-induced side effects. Arch. Gynecol. Obstet. 2022, 307, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Telang, N.T. Natural products as drug candidates for breast cancer (Review). Oncol. Lett. 2023, 26, 1–8. [Google Scholar] [CrossRef]
- Xiao, Y.-L.; Gong, Y.; Qi, Y.-J.; Shao, Z.-M.; Jiang, Y.-Z. Effects of dietary intervention on human diseases: molecular mechanisms and therapeutic potential. Signal Transduct. Target. Ther. 2024, 9, 1–34. [Google Scholar] [CrossRef]
- Rock, E. and A. J.T.J.o.n. DeMichele, Nutritional approaches to late toxicities of adjuvant chemotherapy in breast cancer survivors. 2003, 133, 3785S–3793S. [Google Scholar]
- Jayedi, A.; Emadi, A.; Khan, T.A.; Abdolshahi, A.; Shab-Bidar, S. Dietary Fiber and Survival in Women with Breast Cancer: A Dose-Response Meta-Analysis of Prospective Cohort Studies. Nutr. Cancer 2020, 73, 1570–1580. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Kitagawa, T.; Baron, B.; Kuhara, K.; Nagayasu, H.; Kobayashi, M.; Chiba, I.; Kuramitsu, Y. A standardized extract of cultured Lentinula edodes mycelia downregulates cortactin in gemcitabine-resistant pancreatic cancer cells. Oncol. Lett. 2021, 22, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, A.A.; Rakhmetova, V.; Kapanova, G.; Mussakhanova, A.; Tashenova, G.; Tulebayeva, A.; Akhenbekova, A.; Xu, B. Suppressive effects of bioactive herbal polysaccharides against different cancers: From mechanisms to translational advancements. Phytomedicine 2022, 110, 154624. [Google Scholar] [CrossRef]
- Mathew, L.; Gaikwad, A.; Gonzalez, A.; Nugent, E.K.; Smith, J.A. Evaluation of Active Hexose Correlated Compound (AHCC) in Combination With Anticancer Hormones in Orthotopic Breast Cancer Models. Integr. Cancer Ther. 2017, 16, 300–307. [Google Scholar] [CrossRef]
- Paganelli, F.; Chiarini, F.; Palmieri, A.; Martinelli, M.; Sena, P.; Bertacchini, J.; Roncucci, L.; Cappellini, A.; Martelli, A.M.; Bonucci, M.; et al. The Combination of AHCC and ETAS Decreases Migration of Colorectal Cancer Cells, and Reduces the Expression of LGR5 and Notch1 Genes in Cancer Stem Cells: A Novel Potential Approach for Integrative Medicine. Pharmaceuticals 2021, 14, 1325. [Google Scholar] [CrossRef] [PubMed]
- Graham, É.A.; Mallet, J.-F.; Jambi, M.; Nishioka, H.; Homma, K.; Matar, C. MicroRNA signature in the chemoprevention of functionally-enriched stem and progenitor pools (FESPP) by Active Hexose Correlated Compound (AHCC). Cancer Biol. Ther. 2017, 18, 765–774. [Google Scholar] [CrossRef]
- Shan, N.L.; Shin, Y.; Yang, G.; Furmanski, P.; Suh, N. Breast cancer stem cells: A review of their characteristics and the agents that affect them. Mol. Carcinog. 2021, 60, 73–100. [Google Scholar] [CrossRef]
- Ritz, B.W.; Nogusa, S.; A Ackerman, E.; Gardner, E.M. Supplementation with Active Hexose Correlated Compound Increases the Innate Immune Response of Young Mice to Primary Influenza Infection. J. Nutr. 2006, 136, 2868–2873. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, D.; Sun, B.; Fujii, H.; Kosuna, K.-I.; Yin, Z. Active hexose correlated compound enhances tumor surveillance through regulating both innate and adaptive immune responses. Cancer Immunol. Immunother. 2005, 55, 1258–1266. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.S.; Park, H.-J.; Maeda, T.; Nishioka, H.; Fujii, H.; Kang, I. The Effects of AHCC®, a Standardized Extract of Cultured Lentinura edodes Mycelia, on Natural Killer and T Cells in Health and Disease: Reviews on Human and Animal Studies. J. Immunol. Res. 2019, 2019, 1–7. [Google Scholar] [CrossRef]
- Yin, Z.; Fujii, H.; Walshe, T. Effects of active hexose correlated compound on frequency of CD4+ and CD8+ T cells producing interferon-γ and/or tumor necrosis factor–α in healthy adults. Hum. Immunol. 2010, 71, 1187–1190. [Google Scholar] [CrossRef]
- Chowdhury, A.H. , et al., Modulation of T regulatory and dendritic cell phenotypes following ingestion of Bifidobacterium longum, active hexose correlated compound (AHCC®) and azithromycin in healthy individuals. 11(10).
- OYEDEPO, T.A. E.J.H.P.D.F. MORAKINYO, and Applications, Medicinal Mushrooms. 2020: p. 167.
- Matsui, K.; Ozaki, T.; Oishi, M.; Tanaka, Y.; Kaibori, M.; Nishizawa, M.; Okumura, T.; Kwon, A.-H. Active Hexose Correlated Compound Inhibits the Expression of Proinflammatory Biomarker iNOS in Hepatocytes. Eur. Surg. Res. 2011, 47, 274–283. [Google Scholar] [CrossRef]
- Tanaka, Y.; Ohashi, S.; Ohtsuki, A.; Kiyono, T.; Park, E.Y.; Nakamura, Y.; Sato, K.; Oishi, M.; Miki, H.; Tokuhara, K.; et al. Adenosine, a hepato-protective component in active hexose correlated compound: Its identification and iNOS suppression mechanism. Nitric Oxide 2014, 40, 75–86. [Google Scholar] [CrossRef]
- Terakawa, N.; Matsui, Y.; Satoi, S.; Yanagimoto, H.; Takahashi, K.; Yamamoto, T.; Yamao, J.; Takai, S.; Kwon, A.-H.; Kamiyama, Y. Immunological Effect of Active Hexose Correlated Compound (AHCC) in Healthy Volunteers: A Double-Blind, Placebo-Controlled Trial. Nutr. Cancer 2008, 60, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Spierings, E. , et al. , A phase 1 study of the safety of the nutritional supplement, active hexose correlated compound, AHCC, in healthy volunteers. 2008, 13, 81–82. [Google Scholar]
- Cowawintaweewat, S.; Manoromana, S.; Sriplung, H.; Khuhaprema, T.; Tongtawe, P.; Tapchaisri, P.; Chaicumpa, W. Prognostic improvement of patients with advanced liver cancer after active hexose correlated compound (AHCC) treatment. Journal of Immunotherapy 2006, 24, 33–45. [Google Scholar]
- Parida, D.K.; Wakame, K.; Nomura, T. Integrating Complimentary and Alternative Medicine in Form of Active Hexose Co-Related Compound (AHCC) in the Management of Head & Neck Cancer Patients. Int. J. Clin. Med. 2011, 02, 588–592. [Google Scholar] [CrossRef]
- Mallet, J.-F.; Graham, É.; Ritz, B.W.; Homma, K.; Matar, C. Active Hexose Correlated Compound (AHCC) promotes an intestinal immune response in BALB/c mice and in primary intestinal epithelial cell culture involving toll-like receptors TLR-2 and TLR-4. Eur. J. Nutr. 2015, 55, 139–146. [Google Scholar] [CrossRef]
- Miura, T. D.J.N. Kulkarni, Immunity,, and Infection, 25 AHCC Nutritional Supplement. 2017: p. 427.
- Olamigoke, L.T. , Multifactorial Effects of Biological Response Modifiers and Anti-Neoplastic Induced Immune Cell Activation and Triple Negative Breast Cancer Cell Inhibition. 2017, Texas Southern University.
- Dhawan, A.; Scott, J.G.; Harris, A.L.; Buffa, F.M. Pan-cancer characterisation of microRNA across cancer hallmarks reveals microRNA-mediated downregulation of tumour suppressors. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Bedi, A. , Breast cancer invasion by microRNAs aid. Authorea Preprints 2023. [Google Scholar]
- Ocansey, D.K.W.; Pei, B.; Yan, Y.; Qian, H.; Zhang, X.; Xu, W.; Mao, F. Improved therapeutics of modified mesenchymal stem cells: an update. J. Transl. Med. 2020, 18, 42. [Google Scholar] [CrossRef]
- Slama, Y.; Ah-Pine, F.; Khettab, M.; Arcambal, A.; Begue, M.; Dutheil, F.; Gasque, P. The Dual Role of Mesenchymal Stem Cells in Cancer Pathophysiology: Pro-Tumorigenic Effects versus Therapeutic Potential. Int. J. Mol. Sci. 2023, 24, 13511. [Google Scholar] [CrossRef]
- Yuan, Q.; Zhang, Y.; Chen, Q. Mesenchymal Stem Cell (MSC)-Derived Extracellular Vesicles: Potential Therapeutics as MSC Trophic Mediators in Regenerative Medicine. Anat. Rec. 2019, 303, 1735–1742. [Google Scholar] [CrossRef]
- Tsiapalis, D. and L.J. C. O’Driscoll, Mesenchymal stem cell derived extracellular vesicles for tissue engineering and regenerative medicine applications. Cells 2020, 9, 991. [Google Scholar] [CrossRef]
- Mardpour, S.; Hamidieh, A.A.; Taleahmad, S.; Sharifzad, F.; Taghikhani, A.; Baharvand, H. Interaction between mesenchymal stromal cell-derived extracellular vesicles and immune cells by distinct protein content. J. Cell. Physiol. 2018, 234, 8249–8258. [Google Scholar] [CrossRef]
- Jaschke, N.; Hofbauer, L.C.; Göbel, A.; Rachner, T.D. Evolving functions of Dickkopf-1 in cancer and immunity. Cancer Lett. 2020, 482, 1–7. [Google Scholar] [CrossRef]
- Zhu, G.; Song, J.; Chen, W.; Yuan, D.; Wang, W.; Chen, X.; Liu, H.; Su, H.; Zhu, J. Expression and Role of Dickkopf-1 (Dkk1) in Tumors: From the Cells to the Patients. Cancer Manag. Res. 2021, 13, 659–675. [Google Scholar] [CrossRef]
- Wang, H.; Ashton, R.; Hensel, J.A.; Lee, J.H.; Khattar, V.; Wang, Y.; Deshane, J.S.; Ponnazhagan, S. RANKL-Targeted Combination Therapy with Osteoprotegerin Variant Devoid of TRAIL Binding Exerts Biphasic Effects on Skeletal Remodeling and Antitumor Immunity. Mol. Cancer Ther. 2020, 19, 2585–2597. [Google Scholar] [CrossRef]
- Qiao, L.; Xu, Z.-L.; Zhao, T.-J.; Ye, L.-H.; Zhang, X.-D. Dkk-1 secreted by mesenchymal stem cells inhibits growth of breast cancer cells via depression of Wnt signalling. Cancer Lett. 2008, 269, 67–77. [Google Scholar] [CrossRef]
- Otsu, K.; Das, S.; Houser, S.D.; Quadri, S.K.; Bhattacharya, S.; Bhattacharya, J. Concentration-dependent inhibition of angiogenesis by mesenchymal stem cells. Blood 2009, 113, 4197–4205. [Google Scholar] [CrossRef]
- Jafari, A.; Rezaei-Tavirani, M.; Farhadihosseinabadi, B.; Zali, H.; Niknejad, H. Human amniotic mesenchymal stem cells to promote/suppress cancer: two sides of the same coin. Stem Cell Res. Ther. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Aravindhan, S.; Ejam, S.S.; Lafta, M.H.; Markov, A.; Yumashev, A.V.; Ahmadi, M. Mesenchymal stem cells and cancer therapy: insights into targeting the tumour vasculature. Cancer Cell Int. 2021, 21, 1–15. [Google Scholar] [CrossRef]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular Vesicles: Unique Intercellular Delivery Vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef]
- Pan, S.; Zhang, Y.; Natalia, A.; Lim, C.Z.J.; Ho, N.R.Y.; Chowbay, B.; Loh, T.P.; Tam, J.K.C.; Shao, H. Extracellular vesicle drug occupancy enables real-time monitoring of targeted cancer therapy. Nat. Nanotechnol. 2021, 16, 734–742. [Google Scholar] [CrossRef]
- Crescitelli, R.; LäsSer, C.; LötVall, J. Isolation and characterization of extracellular vesicle subpopulations from tissues. Nat. Protoc. 2021, 16, 1548–1580. [Google Scholar] [CrossRef]
- Taghikhani, A.; Farzaneh, F.; Sharifzad, F.; Mardpour, S.; Ebrahimi, M.; Hassan, Z.M. Engineered Tumor-Derived Extracellular Vesicles: Potentials in Cancer Immunotherapy. Front. Immunol. 2020, 11, 221. [Google Scholar] [CrossRef]
- Mardpour, S. , et al., Hydrogel-Mediated Sustained Systemic Delivery of Mesenchymal Stem Cell-Derived Extracellular Vesicles Improves Hepatic Regeneration in Chronic Liver Failure. ACS Applied Materials & Interfaces 2019, 11, 37421–37433. [Google Scholar]
- Zhang, X.; Yang, Y.; Zhang, L.; Lu, Y.; Zhang, Q.; Fan, D.; Zhang, Y.; Zhang, Y.; Ye, Z.; Xiong, D. Mesenchymal stromal cells as vehicles of tetravalent bispecific Tandab (CD3/CD19) for the treatment of B cell lymphoma combined with IDO pathway inhibitor d-1-methyl-tryptophan. J. Hematol. Oncol. 2017, 10, 1–14. [Google Scholar] [CrossRef]
- Lin, S.; Zhu, B.; Huang, G.; Zeng, Q.; Wang, C. Microvesicles derived from human bone marrow mesenchymal stem cells promote U2OS cell growth under hypoxia: the role of PI3K/AKT and HIF-1α. Hum. Cell 2018, 32, 64–74. [Google Scholar] [CrossRef]
- Bruno, S.; Collino, F.; Deregibus, M.C.; Grange, C.; Tetta, C.; Camussi, G. Microvesicles Derived from Human Bone Marrow Mesenchymal Stem Cells Inhibit Tumor Growth. Stem Cells Dev. 2013, 22, 758–771. [Google Scholar] [CrossRef]
- Wu, S.; Ju, G.-Q.; Du, T.; Zhu, Y.-J.; Liu, G.-H. Microvesicles Derived from Human Umbilical Cord Wharton’s Jelly Mesenchymal Stem Cells Attenuate Bladder Tumor Cell Growth In Vitro and In Vivo. PLOS ONE 2013, 8, e61366. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Nahand, J.S.; Vandchali, N.R.; Darabi, H.; Doroudian, M.; Banafshe, H.R.; Moghoofei, M.; Babaei, F.; Salmaninejad, A.; Mirzaei, H. Exosomal MicroRNAs: Novel Players in Cervical Cancer. Epigenomics 2020, 12, 1651–1660. [Google Scholar] [CrossRef]
- Moradi-Chaleshtori, M.; Bandehpour, M.; Heidari, N.; Mohammadi-Yeganeh, S.; Hashemi, S.M. Exosome-mediated miR-33 transfer induces M1 polarization in mouse macrophages and exerts antitumor effect in 4T1 breast cancer cell line. Int. Immunopharmacol. 2021, 90, 107198. [Google Scholar] [CrossRef]
- Yang, E.; Wang, X.; Gong, Z.; Yu, M.; Wu, H.; Zhang, D. Exosome-mediated metabolic reprogramming: the emerging role in tumor microenvironment remodeling and its influence on cancer progression. Signal Transduct. Target. Ther. 2020, 5, 1–13. [Google Scholar] [CrossRef]
- Hemmatzadeh, M.; Mohammadi, H.; Jadidi-Niaragh, F.; Asghari, F.; Yousefi, M. The role of oncomirs in the pathogenesis and treatment of breast cancer. Biomed. Pharmacother. 2016, 78, 129–139. [Google Scholar] [CrossRef]
- Li, L.; Yuan, L.; Luo, J.; Gao, J.; Guo, J.; Xie, X. MiR-34a inhibits proliferation and migration of breast cancer through down-regulation of Bcl-2 and SIRT1. Clin. Exp. Med. 2012, 13, 109–117. [Google Scholar] [CrossRef]
- Rahimi, M.; Sharifi-Zarchi, A.; Firouzi, J.; Azizmi, M.; Zarghami, N.; Alizadeh, E.; Ebrahimi, M. An integrated analysis to predict micro-RNAs targeting both stemness and metastasis in breast cancer stem cells. J. Cell. Mol. Med. 2019, 23, 2442–2456. [Google Scholar] [CrossRef]
- MicroRNA-200c Increases Radiosensitivity of Human Cancer Cells with Activated EGFR-associated Signaling. 2018, 서울대학교 대학원.
- Venturella, G.; Ferraro, V.; Cirlincione, F.; Gargano, M.L. Medicinal Mushrooms: Bioactive Compounds, Use, and Clinical Trials. Int. J. Mol. Sci. 2021, 22, 634. [Google Scholar] [CrossRef]
- Hou, Y.-N., G. Deng, and J.J. Mao, Practical Application of "About Herbs" Website: Herbs and Dietary Supplement Use in Oncology Settings. Cancer journal (Sudbury, Mass.) 2019, 25, 357–366. [Google Scholar] [CrossRef]
- Oyedepo, T.A. and A.E. Morakinyo, Medicinal Mushrooms, in Herbal Product Development. 2020, Apple Academic Press. p. 167-203.
- Chakraborty, C.; Chin, K.-Y.; Das, S. miRNA-regulated cancer stem cells: understanding the property and the role of miRNA in carcinogenesis. Tumor Biol. 2016, 37, 13039–13048. [Google Scholar] [CrossRef]
- Yoshida, K.; Yamamoto, Y.; Ochiya, T. miRNA signaling networks in cancer stem cells. Regen. Ther. 2021, 17, 1–7. [Google Scholar] [CrossRef]
- Takahashi, R.; Prieto-Vila, M.; Kohama, I.; Ochiya, T. Development of miRNA-based therapeutic approaches for cancer patients. Cancer Sci. 2019, 110, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Zhang, H.-W.; Lu, M.-H.; He, X.-H.; Li, Y.; Gu, H.; Liu, M.-F.; Wang, E.-D. MicroRNA-155 Functions as an OncomiR in Breast Cancer by Targeting theSuppressor of Cytokine Signaling 1Gene. Cancer Res. 2010, 70, 3119–3127. [Google Scholar] [CrossRef] [PubMed]
- Mattiske, S.; Suetani, R.J.; Neilsen, P.M.; Callen, D.F. The Oncogenic Role of miR-155 in Breast Cancer. Cancer Epidemiology Biomarkers Prev. 2012, 21, 1236–1243. [Google Scholar] [CrossRef]
- Ma, Y.; Chai, N.; Jiang, Q.; Chang, Z.; Chai, Y.; Li, X.; Sun, H.; Hou, J.; Linghu, E. DNA methyltransferase mediates the hypermethylation of the microRNA 34a promoter and enhances the resistance of patient-derived pancreatic cancer cells to molecular targeting agents. Pharmacol. Res. 2020, 160, 105071. [Google Scholar] [CrossRef]
- Peng, X.; Chang, H.; Chen, J.; Zhang, Q.; Yu, X.; Mi, M. 3,6-Dihydroxyflavone regulates microRNA-34a through DNA methylation. BMC Cancer 2017, 17, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Wang, W. A Systemic Review on the Regulatory Roles of miR-34a in Gastrointestinal Cancer. OncoTargets Ther. 2020, 13, 2855–2872. [Google Scholar] [CrossRef]
- Bonetti, P.; Climent, M.; Panebianco, F.; Tordonato, C.; Santoro, A.; Marzi, M.J.; Pelicci, P.G.; Ventura, A.; Nicassio, F. Dual role for miR-34a in the control of early progenitor proliferation and commitment in the mammary gland and in breast cancer. Oncogene 2018, 38, 360–374. [Google Scholar] [CrossRef]
- Deng, J.; Yang, M.; Jiang, R.; An, N.; Wang, X.; Liu, B. Long Non-Coding RNA HOTAIR Regulates the Proliferation, Self-Renewal Capacity, Tumor Formation and Migration of the Cancer Stem-Like Cell (CSC) Subpopulation Enriched from Breast Cancer Cells. PLOS ONE 2017, 12, e0170860. [Google Scholar] [CrossRef]
- Zare, M.; Bastami, M.; Solali, S.; Alivand, M.R. Aberrant miRNA promoter methylation and EMT-involving miRNAs in breast cancer metastasis: Diagnosis and therapeutic implications. J. Cell. Physiol. 2017, 233, 3729–3744. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.-. .-G.; Shi, J.-.-C.; Shang, J.; Hao, J.-.-G.; Du, X. Effect of miR-34a on resistance to sunitinib in breast cancer by regulating the Wnt/β-catenin signaling pathway. European Review for Medical & Pharmacological Sciences 2019, 23, 1151–1157. [Google Scholar] [CrossRef]
- Shi, Y.; Cao, T.; Huang, H.; Lian, C.; Yang, Y.; Wang, Z.; Ma, J.; Xia, J. Arsenic trioxide inhibits cell growth and motility via up-regulation of let-7a in breast cancer cells. Cell Cycle 2017, 16, 2396–2403. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Fan, J.J.; Dong, C.; Li, H.-T.; Ma, B.-L. Inhibition effect of exosomes-mediated Let-7a on the development and metastasis of triple negative breast cancer by down-regulating the expression of c-Myc. European Review for Medical & Pharmacological Sciences 2019, 23, 5301–5314. [Google Scholar] [CrossRef]
- Bandyopadhayaya, S.; Mandal, C.C. A Differential Role of miRNAs in Regulation of Breast Cancer Stem Cells, in Cancer Stem Cells: New Horizons in Cancer Therapies. 2020, Springer. p. 87-109.
Figure 1.
1a- AHCC induces tumor-suppressive miRNA expression (miR-Let-7a, miR-200c, and miR-34a) and reduces oncogenic miR-155 in wild-type MCF-7 cells.
Figure 1.
1a- AHCC induces tumor-suppressive miRNA expression (miR-Let-7a, miR-200c, and miR-34a) and reduces oncogenic miR-155 in wild-type MCF-7 cells.
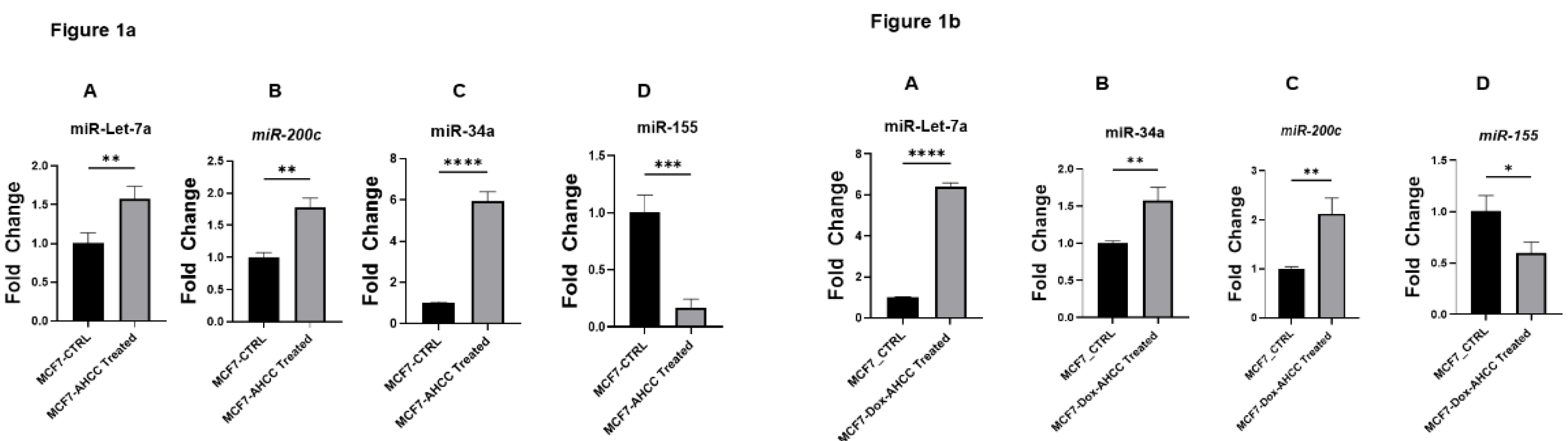
Figure 2.
2-Nanoparticle tracking analysis of EV concentrations from AHCC-treated versus untreated MSCs. Panel A shows the Concentration measurement results of (MSCs) treated with AHCC EVs: 1.2E+10 Particles / mL. Panel B shows MSCs that were not treated with AHCC produced EVs with a concentration of 5.7E+9 particles/mL.
Figure 2.
2-Nanoparticle tracking analysis of EV concentrations from AHCC-treated versus untreated MSCs. Panel A shows the Concentration measurement results of (MSCs) treated with AHCC EVs: 1.2E+10 Particles / mL. Panel B shows MSCs that were not treated with AHCC produced EVs with a concentration of 5.7E+9 particles/mL.
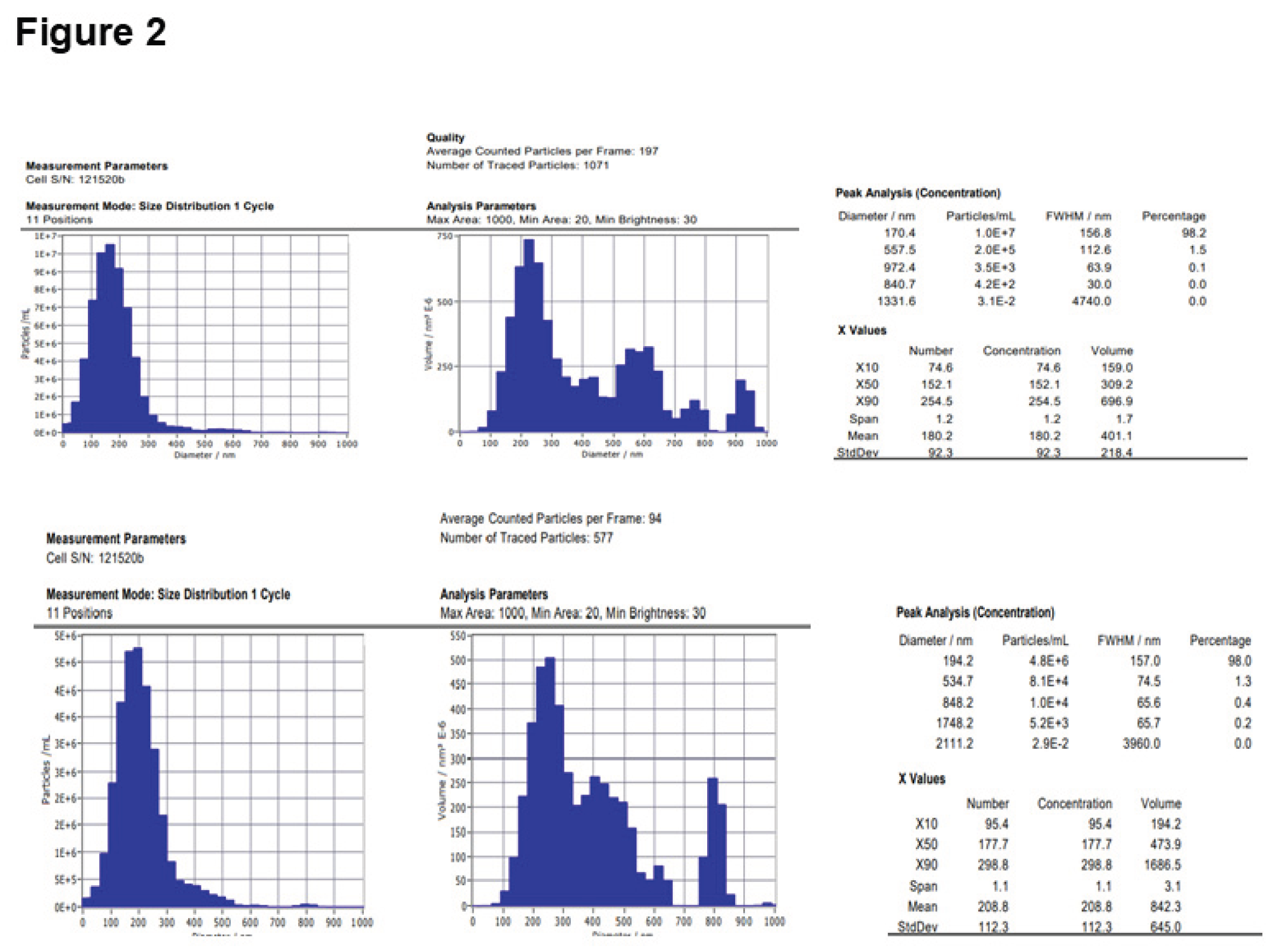
Figure 3.
Figure 3a. AHCC-treated MSC-derived EVs upregulate tumor-suppressive miRNAs in MCF-7/DOX spheres.Quantitative RTPCR assessments of miRNA expression in MCF-7/DOX spheres, following treatment with AHCC-treated MSC-derived EVs and non-treated MSC-derived EVs as control group. Panels A, B and C demonstrate the significant upregulation of the tumor-suppressive miRNAs such as miR-34a, miR-Let7a, and miR-200c respectively. Panel D No significant difference was observed between the control and AHCC treated MSC-derived EVs treatment groups. Figure 3b. Effects of AHCC-treated MSC-derived EVs on miRNA expression in MCF-7 cell spheres. Quantitative RT-PCR analysis of miRNA expression in wild-type MCF-7 cell sphere after treatment with AHCC-treated MSCs derived EVs and Naïve MSCs derived EVs as control group. A The expressions level of Let-7a represented no significant difference vs control. B, Significant upregulation of miR-34a. C shows the expression levels of tumor-suppressive miRNAs which are significantly upregulated after 24 hours of exposure to AHCC-treated MSC-derived EVs. Panel D illustrates the reduction of the oncogenic miR-155 in the same sphere culture at the same condition.
Figure 3.
Figure 3a. AHCC-treated MSC-derived EVs upregulate tumor-suppressive miRNAs in MCF-7/DOX spheres.Quantitative RTPCR assessments of miRNA expression in MCF-7/DOX spheres, following treatment with AHCC-treated MSC-derived EVs and non-treated MSC-derived EVs as control group. Panels A, B and C demonstrate the significant upregulation of the tumor-suppressive miRNAs such as miR-34a, miR-Let7a, and miR-200c respectively. Panel D No significant difference was observed between the control and AHCC treated MSC-derived EVs treatment groups. Figure 3b. Effects of AHCC-treated MSC-derived EVs on miRNA expression in MCF-7 cell spheres. Quantitative RT-PCR analysis of miRNA expression in wild-type MCF-7 cell sphere after treatment with AHCC-treated MSCs derived EVs and Naïve MSCs derived EVs as control group. A The expressions level of Let-7a represented no significant difference vs control. B, Significant upregulation of miR-34a. C shows the expression levels of tumor-suppressive miRNAs which are significantly upregulated after 24 hours of exposure to AHCC-treated MSC-derived EVs. Panel D illustrates the reduction of the oncogenic miR-155 in the same sphere culture at the same condition.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



