Submitted:
18 November 2025
Posted:
18 November 2025
You are already at the latest version
Abstract
Background: Ammonium nitrogen (NH₄⁺) serves as a vital nitrogen source, playing pivotal regulatory roles in plant growth, development, and high-yield formation. Ammonium transporters (AMTs), encoded by the AMT gene family, are central to NH4+ transport. However, the functional roles of AMT genes in wheat remain poorly understood.Methods: In this study, a total of 21 TaAMT members were identified. A comprehensive genome-wide analysis was conducted, encompassing investigations into gene structure, protein motif composition, gene duplication events, collinearity relationships, and cis-acting regulatory elements. Furthermore, the expression patterns of distinct TaAMT members were examined under varying ammonium supply conditions and pathogen stress. Results: All TaAMT proteins exhibited hydrophobic characteristics and localized to the plasma membrane. Phylogenetic analysis clustered these genes into four distinct subgroups. Comparative analyses of gene structure and conserved motifs revealed conserved domain composition and motif organization within each subgroup. Interspecific synteny analysis highlighted evolutionary conservation across species. Promoter region analysis identified multiple cis-regulatory elements associated with hormone signaling, light responsiveness, and abiotic stress adaptation. Expression profiling demonstrated that TaAMT members exhibit both tissue-specific and constitutive expression patterns across developmental stages. RT-qPCR further revealed that the expression of TaAMT members responds to varying concentrations of ammonium nitrogen supply, as well as infection stresses caused by stripe rust and powdery mildew. Conclusions: Collectively, this study provides the useful molecular information into the roles of TaAMT members in nitrogen use efficiency and disease resistance, offering valuable genetic resources for wheat breeding programs aimed at enhancing agronomic traits.
Keywords:
wheat
; ammonium nitrogen
; Puccinia striiformis
; Blumeria graminis
1. Introduction
Nitrogen (N) is an indispensable macronutrient for plant growth and development, playing a pivotal role in various physiological processes, such as protein synthesis, nucleic acid metabolism, and photosynthetic efficiency [1,2]. Among the diverse forms of nitrogen available in the soil, ammonium (NH₄⁺) serves as a primary nitrogen source for many plants, especially in environments where nitrification is inhibited, such as acidic soils or waterlogged conditions [3]. The efficient acquisition and assimilation of ammonium in plants predominantly rely on ammonium transporters (AMTs), which facilitate the selective movement of NH₄⁺ across cellular membranes [4,5]. Given their critical role in nitrogen acquisition and homeostasis, AMT genes have garnered significant attention in plant physiology and molecular biology research.
To date, genome-wide identification and characterization of the AMT gene family have been conducted in several plant species [6,7,8,9], such as Arabidopsis thaliana, rice (Oryza sativa) [10], and maize (Zea mays) [11,12], revealing the diversity, structural features, and functional roles of these genes. In Arabidopsis thaliana, six AtAMTs were identified and divided into two subfamilies (AMT1 and AMT2), of which five belong to the AMT1 subfamily (AtAMT1.1-AMT1.5) and only one (AtAMT2) belongs to that of AMT2. AMT1 proteins primarily involved in high-affinity NH₄⁺ transport, while AMT2 members function in low-affinity transport and metabolic regulation [13,14]. The different AMT subtypes exhibit variations in ammonium ion substrate affinity and transport activity [15]. The AMT1 subfamily plays a dominant role in high-affinity NH₄⁺ absorption [16]. For example, AtAMT1.1 and AtAMT1.3 respond to changes in NH₄⁺ concentration via phosphorylation, forming a dynamic regulatory loop characterized by low-nitrogen-induced activation and high-nitrogen-suppressed degradation [13,17,18].
Wheat (Triticum aestivum L.) is one of the most important staple crops globally, providing a major source of calories and protein for human consumption. Nitrogen (N) nutrition is a critical determinant of crop productivity and grain quality. The first “Green Revolution” in the 1960s [19,20], marked by the breeding of new dwarf wheat varieties, led to a dramatic increase in global wheat production and resolved the food crisis caused by explosive population growth worldwide [21]. Dwarf crops, with their optimized plant height, significantly enhanced lodging resistance while efficiently utilizing sunlight and water resources, ultimately boosting yields [22,23]. However, dwarf varieties generally exhibit low nitrogen use efficiency (NUE), making their yields heavily dependent on chemical fertilizer inputs [24,25]. Excessive nitrogen fertilization not only increases cultivation costs but also causes environmental pollution. How to break through the breeding bottleneck of coordinating high yield and nitrogen efficiency, and achieve increased crop yields with reduced nitrogen inputs, has become a critical challenge for sustainable agricultural development [26,27]. Notably, compared to ammonium-preferred crops like rice, wheat exhibits significantly lower affinity for ammonium nitrogen (NH₄⁺), and the activity of its key nitrogen assimilation enzyme (e.g., glutamine synthetase, GS) is highly susceptible to environmental fluctuations [28,29]. These characteristics make wheat’s NUE even more complex to improve[30]. Thus, systematically dissecting the molecular regulatory mechanisms underlying wheat nitrogen absorption and breaking through its “high-input, low-efficiency” utilization bottleneck have become core scientific issues for achieving the agricultural goal of “reduced nitrogen input with enhanced efficiency” [27,31]. Furthermore, previous studies have shown that AMT1 can positively influence sheath blight (ShB) resistance by regulating ammonium uptake and assimilation [32]. This raises an interesting question of whether TaAMT gene family members may play a role in pathogen defense response [33], and their expression dynamics under NH₄⁺ stress and pathogen challenge remain poorly understood [34].
This study performed a genome-wide identification of the AMT gene family in wheat, and investigated their phylogenetic relationships, gene structures, and expression profiles under ammonium nutrition and pathogen infection. The results may contribute to a deeper understanding of the biological functions of TaAMT members, and provide novel insights into the dual roles of TaAMT members in N nutrition and biotic stress adaptation, offering potential targets for for breeding wheat varieties with improved NUE and disease resistance [35].
2. Materials and Methods
2.1. Experimental Materials and Treatment
The experimental materials included the wheat varieties Kenong 199 (KN199), 7182, and the wheat-Thinopyrum ponticum disomic alien substitution line CH10A5 [36]. The seeds were disinfected with 1% sodium hypochlorite for 10 minutes, rinsed with distilled water, and then and germinated at 25 °C in the dark for 24 hours. Uniformly germinated seedlings (with radicles 1-2cm long) were selected and transferred to hydroponic boxes (containing 2L of wheat nutrient solution, pH = 5.5, Coolaber NSP1070). They were then cultivated in a light incubator (16h light / 8h dark, light intensity 300 μmol·m⁻²·s⁻¹, temperature 22 ± 2 °C).
For ammonium salt stress treatment, three-leaf seedlings were exposed to nutrient solutions with 0.04 mM (low nitrogen), 0.4 mM (normal nitrogen, CK), and 40 mM (high nitrogen) NH₄+. Samples (0.5 g of the second top leaf) were collected at specified times (0, 6, 12, 24, 36, 48, 72, 96, 144, 168, and 216 hours), frozen in liquid nitrogen, and stored at -80 °C, with three biological replicates per treatment.
For pathogen inoculation, seedlings at the two-leaf and one-heart stage were inoculated with stripe rust race CYR34 and powdery mildew race E09, respectively. Samples were taken at 0, 24 and 48 h post-inoculation, frozen in liquid nitrogen, and stored at -80 °C, with three biological replicates per treatment.
2.2. Identification and Physicochemical Property Analysis of the AMT Gene Family in Wheat
Reference genomes, protein sequences, and GFF3 annotations for Arabidopsis thaliana (TAIR 10), rice (MSU 7.0), and wheat (RefSeq v1.1) were downloaded from Ensembl Plants [37]. Arabidopsis thaliana AMT proteins (https://www.arabidopsis.org/) were used as queries in local BLAST (E-value < 1e-5) against the wheat proteome to identify candidate TaAMT sequences. The AMT family HMM profile (PF00909) from Pfam (https://pfam-legacy.xfam.org/) was searched in wheat proteins using HMMER 3.3 (HMMsearch, E-value < 1e-10). Candidates were validated for conserved AMT domains (PF00909) via CDD (https://www.ncbi.nlm.nih.gov/cdd) and SMART (http://smart.embl-heidelberg.de/). Physicochemical properties (Mw, pI, instability index, aliphatic index) of TaAMT proteins were predicted using ExPASy ProtParam (https://web.expasy.org/protparam/). TaAMT members were renamed based on chromosomal positions.
2.3. Phylogenetic Analysis of TaAMT Proteins
AMT protein sequences from Arabidopsis, rice, and wheat were aligned using Clustal X 2.1 (default parameters: Gap Opening Penalty=15, Gap Extension=6.66, DNA Weight Matrix=IUB) [38]. A phylogenetic tree was constructed with MEGA 11.0 via the Maximum Likelihood (ML) method, using the JTT+G+I substitution model selected by Model Test (1,000 bootstrap replicates) [39]. The tree was visualized and annotated with species origins and gene names using iTOL (https://itol.embl.de/upload.cgi) [40].
2.4. Analysis of Gene Structure, Protein Features, and Cis-Regulatory Elements
TaAMT members structures were analyzed using TBtools v2.225 to extract exon/intron counts, lengths, and positions from wheat GFF3 annotations and generate schematic diagrams. Conserved motifs in TaAMT proteins were identified with MEME Suite v5.0.5 (motif width: 6-50 aa; max motifs: 15; default EM algorithm settings, 100 iterations) [41]. Promoter sequences (2,000 bp upstream) were retrieved from WheatOmics 1.0 [42] (http://wheatomics.sdau.edu.cn/) and analyzed for cis-elements using PlantCARE (https://bioinformatics.psb.ugent.be/webtools/plantcare/). The results were visualized with TBtools v2.225 [43].
2.5. Chromosomal Distribution, Gene Duplication Events, and Synteny Analysis of TaAMTs
The chromosomal physical positions of TaAMT members were extracted from the wheat GFF3 annotation file, and their distribution maps on chromosomes were generated using TBtools v2.225 [44]. Gene duplication events (tandem and segmental) within the TaAMT family were identified via MCScanX (http://chibba.pgml.uga.edu/mcscan2/) using wheat RefSeq v1.1 synteny data (E-value < 1e-10. The synteny analysis results were visualized using the built-in module of MCScanX.
2.6. Analysis of Tissue Expression Patterns of TaAMTs
Transcriptome data of TaAMTs genes across stages and tissues were downloaded from ExpVIP (http://www.wheat-expression.com/). The selected tissues and developmental stages for expression analysis included: roots (seven stages ), leaves (six stages), stems (five stages ), spikes (one stages ), and grains (five stages). The expression data of TaAMT members were standardized using Z-score normalization with TBtools v2.225, and hierarchical clustering heatmaps were generated with log₂(FPKM+1) as the metric. Colors in heatmaps indicate expression levels (red: high; blue: low), with Euclidean distance used for clustering.
2.7. Analysis of Expression Patterns of TaAMTs Under Different Stress Conditions
Ten representative TaAMT members (covering AMT1 and AMT2 subfamilies) with tissue-specific expression were selected to analyze their expression dynamics under ammonium stress, stripe rust, and powdery mildew infection via RT-qPCR. TaActin served as the reference gene, with amplification on the QuantStudio TM6 Flex system using TBGreen® Premix ExTaqTM II (Tli RNaseH Plus, RR820A, TaKaRa). Three biological replicates were set per time point. Relative expression was calculated by 2−ΔΔCT method, and significance of differences among treatments was tested using One-way ANOVA and Duncan’s test (P < 0.01) in SPSS 26.0. The primers used in this study were synthesized by Shanghai Sangon Biological Engineering Technology Co., Ltd., and the primer sequences are listed in Table A1.
3. Results
3.1. Identification and Basic Physicochemical Properties of TaAMT Members
Based on the Chinese Spring reference genome data, a total of 21 TaAMT family members were identified, and then named according to their chromosomal locations (Supplementary Table S1) Physicochemical property analysis showed that The TaAMT proteins range in length from 466 to 563 amino acids, with relative molecular masses of 49,910.41 to 59,889.76 Da and theoretical isoelectric points (pI) ranging from 6.30 to 9.13, averaging 7.52. The total grand average of hydrophobicity (GRAVY) ranged from 0.420 to 0.630, indicating that all members are hydrophobic (Table 1). All TaAMT members were localized on the plasma membrane, indicating that these genes play regulatory roles in the plasma membrane.
3.2. Phylogenetic Tree Analysis, Conserved Domains, and Gene Structural Features
A phylogenetic tree was constructed using homologous proteins from Arabidopsis thaliana, rice and wheat, which categorized the TaAMT family members into four distinct groups ( Figure 1). Notably, Groups Ⅰ and Ⅱ clustered with AtAMT1, whereas groups Ⅲ and Ⅳ aligned with AtAMT2, mirroring the classification pattern observed in OsAMTs. We identified 15 conserved motifs within the TaAMT proteins via MEME Suite 5.0.5, with proteins belonging to the same group or subgroup displaying similar motif compositions (Figure 2A, 2B). Specifically, motifs 1, 2, 3, 6, 9, 10, 12, and 13 emerge as the core motifs conserved across the family. Group Ⅰ, encompassing six members, exhibits high homology to OsAMT1, encompassing 11 motifs, including the distinctive motifs 12 and 14. Conversely, groups Ⅱ (five members), groups Ⅲ (five members), and groups Ⅳ (three members) show homology to OsAMT2, OsAMT3, and OsAMT4, respectively. Gene structure analysis revealed that members of group Ⅰ and group Ⅳ harbor a single exon, group Ⅳ contains three exons, and group Ⅲ possesses either one or two exon (Figure 2D). These findings suggest that these structural discrepancies may potentially arise from gene duplication events, subsequently leading to functional divergence.
3.3. Chromosomal Localization, Synteny, and Interspecific Evolution
Chromosomal localization analysis revealed an uneven distribution of 21 TaAMT gene family members across 15 wheat chromosomes, all located on the long arms ( Figure 3). Notably, the first homology group harbored merely 2 genes (9.5%), whereas the third homology group exhibited the highest gene count, with 7 genes (33.3%). Synteny analysis identified 34 gene pairs within wheat genome (Figure 4). Among these, paralogous gene pairs situated on homologous chromosomes predominated. Interspecific analysis reveals 1 gene pair between Arabidopsis thaliana and wheat (3A), and 30 between wheat and rice, with wheat chromosome 3 and rice chromosome 1 showing high linear consistency in AMT gene evolution, providing clues for cross-species origin studies. Inter-species synteny analysis (Figure 5) showed evolutionary conservation, with only one syntenic pair between Arabidopsis thaliana and wheat, but 30 between wheat and rice, especially a high degree of linear consistency between wheat chromosome three and rice chromosome one in AMT gene evolution, which providing key clues for studying the cross-species origin of the AMT family.
3.4. Cis-Acting Element Analysis
The prediction of cis-acting elements extraction from TaAMT members revealed that all TaAMT gene family members collectively encompass a rich diversity of elements(Figure 6).These elements are associated with hormone responses and abiotic stresses, with hormone-responsive elements being the most prevalent, covering 10 distinct types. Among them, methyl jasmonate (MeJA)-related elements (CGTCA-motif and TGACG-motif) were particularly abundant, totaling 124, closely followed by abscisic acid (ABA)-responsive elements (ABRE) with 114 occurrences, while elements responsive to auxin, gibberellin, and salicylic acid were present in smaller numbers. Notably, all TaAMT members contain the ABA-responsive element ABRE, suggesting their universal involvement in ABA-mediated stress responses or developmental regulation. Additionally, five types of abiotic stress-related elements were identified, including drought-responsive MBS, low-temperature-responsive LTR, hypoxia-responsive ARE, and GC-motif elements, along with a limited number of metal ion stress-related MRE, general stress-responsive MBSI, and NON-box elements. The presence of these elements strongly implies that the TaAMT family genes may dynamically regulate ammonium nitrogen uptake in response to environmental cues such as drought, low temperature, and hypoxia, thereby enhancing the plants’ ability to adapt and thrive under adverse conditions.
3.5. Tissue-Specific Expression Pattern Analysis
Based on the transcriptome data from the ExpVIP database, a heatmap depicting the expression patterns of TaAMT members was generated (Figure 7) . The results revealed that the 21 TaAMT members exhibited notable tissue-specific expression patterns, which could be categorized into three groups: Firstly, there were three constitutively expressed genes showing expression across all examined tissues, likely participating in the fundamental nitrogen metabolism processes in wheat. Secondly, seven genes displayed constitutively low expression, meaning their expression levels were extremely low in all tissues, and their functions require further verification. Additionally, 11 genes exhibited tissue-specific expression, with significant variations in expression levels across different tissues. Among them, TaAMT1, TaAMT2, TaAMT6, TaAMT9, TaAMT11, TaAMT20 and TaAMT21 were highly expressed in roots, suggesting their potential involvement in ammonium nitrogen uptake by roots. TaAMT1, TaAMT2, TaAMT13 and TaAMT16 showed high expression in leaves, possibly related to nitrogen transport or assimilation in the above-ground parts. Notably, TaAMT21 had significantly higher expression levels in stems and spikes compared to other tissues, indicating its crucial role in nitrogen allocation during the reproductive growth stage. The diversity of these tissue-specific expression patterns provides a molecular basis for the coordinated regulation of nitrogen utilization in wheat by the TaAMT gene family members at different developmental stages.
3.6. Analysis of Expression Patterns of TaAMT Members Under Three Stress Conditions
To explore TaAMT members responses to ammonium nitrogen stress, four genes (TaAMT9, TaAMT11, TaAMT13, TaAMT16) were studied via RT-qPCR under high (40 mmol/L NH4+) and low (0.04 mmol/L NH4+) ammonium stresses.TaAMT13 and TaAMT16 showed delayed upregulation under low ammonium, with roots upregulated 2.3-fold and 1.8-fold at 72h, suggesting a role in low-nitrogen absorption. Under high ammonium, their expression reversed, downregulated to 45% and 52% of control at 48h, possibly avoiding cytotoxicity (Figure 8). Conversely, TaAMT9 and TaAMT11 were downregulated by 30%-40% at 24h under low ammonium, but rebounded to 1.6-1.8 times control at 48h under high ammonium, indicating dynamic ammonium sensing and nitrogen homeostasis regulation.
The expression of four TaAMT member was analyzed in the wheat varieties 7182 and CH10A5 under stripe rust stress. At 24h post-inoculation, TaAMT1, TaAMT2, and TaAMT17 were significantly upregulated, likely linked to defensive nitrogen metabolism. At 48h, TaAMT15 expression peaked at 2.1-2.4 times control, coinciding with hyphal expansion, suggesting its role in disease resistance nitrogen signaling (Figure 9).
Under powdery mildew stress induced by E09 infection, nine members of the TaAMT gene family exhibited time-course-specific expression patterns. Specifically, TaAMT1 peaked at 24h (5-fold), while TaAMT2 was transiently induced at 24h. TaAMT6 surged 13-fold at 24h. TaAMT9 and TaAMT13 had transient peaks at 24h (10-15-fold). TaAMT11 mirrored TaAMT9 but with a weaker peak. TaAMT15 was upregulated only at 24h, TaAMT16 remained high at 24h and 48h, and TaAMT18 peaked at 48h (Figure 10).
Over all, TaAMT gene family members show gene-specific, time-dependent expression under ammonium stress and coordinated responses to stripe rust and powdery mildew infections, suggesting their key roles in ammonium homeostasis regulation and disease resistance via pathogen signal sensing, offering vital candidates for exploring “nitrogen-disease” interaction mechanisms.
4. Discussion
Nitrogen is an essential component of important organic compounds within plants [45]. Different wheat germplasm resources [46] exhibit varying nitrogen fertilizer uptake capacities under different environmental factors [47,48,49]. Key components of signal transduction pathways, such as ubiquitin ligases and transcription factors, play a central role in the processes by which plants perceive and respond to nutrient changes [50,51,52,53]. Epigenetic modifications also participate in regulating the efficiency of nitrogen transport and utilization in plants [54]. This study conducted a systematic examination of the evolutionary traits, expression patterns, and stress response mechanisms within the wheat AMT gene family members, highlighting its pivotal role in nitrogen homeostasis and disease resistance. The findings bridge gaps in TaAMT gene family members functional research and lay a theoretical foundation for molecularly designing wheat varieties with “reduced nitrogen input, enhanced efficiency.”
4.1. Evolutionary Dynamics and Functional Divergence of TaAMT Gene Family Members
The evolutionary trajectory of the TaAMT gene family members were closely tied to wheat polyploidization. Phylogenetic analysis categorizes TaAMT into four clusters, aligning with Arabidopsis thaliana and rice classifications, suggesting conserved functional divergence in monocots. Compared to Arabidopsis thaliana (6) and rice (8), wheat boasts a larger repertoire with 21 AMT members, likely due to polyploidy-driven gene expansion. Chromosomal mapping reveals a high concentration on homologous group Ⅲ, with collinearity analysis indicating segmental and tandem duplications as key expansion mechanisms [55,56]. Motif and gene structure analyses underscore functional differentiation, with GroupⅠ members potentially mediating high-affinity transport, while Group Ⅲ members may be involved in low-affinity or adaptive regulation. All TaAMT members localize to the plasma membrane, reinforcing their role as ammonium transporters [57,58].The synteny analysis strongly suggest that the expansion of the TaAMT family primarily hinges on gene duplication events occurring between homologous chromosomes and may have undergone truncated duplication during the course of evolution.
4.2. Cis-Regulatory Elements and Stress Response Mechanisms
Analysis of cis-acting elements in the promoter regions revealed a regulatory network governing TaAMT members responses to diverse signals. All TaAMT gene family members harbor ABA-responsive elements (ABREs). Given their expression patterns under abiotic stresses such as drought and low temperature (as evidenced by the presence of MBS and LTR elements) [59], it is hypothesized that ABA may serve as a key signaling molecule for TaAMT members in response to environmental stresses. Specifically, ABA activates the transcription of TaAMT members by binding to ABREs, thereby promoting ammonium uptake to maintain nitrogen homeostasis under stress conditions [60]. NRT1.1B has been confirmed to function as an ABA receptor [61], and when plants utilize nitrate (NO3−) as a nitrogen source, ABA levels decrease compared to those when ammonium (NH4+) is used [62,63]. Consequently, there exists an interplay and mutual influence between the regulatory networks of AMTs and ABA. Additionally, MeJA-related elements (CGTCA-motif and TGACG-motif) are the most abundant (124 elements), suggesting that jasmonic acid signaling may be involved in the defensive regulation of TaAMT members. Previous studies have demonstrated that jasmonic acid can enhance plant disease resistance by inducing the expression of defense-related genes [64,65], and the upregulation of TaAMT members (e.g., TaAMT1 and TaAMT2) during the early stages of stripe rust infection may act synergistically with this pathway.
4.3. Tissue-Specific Expression and Dynamic Regulation of Nitrogen Uptake and Utilization
The tissue expression heatmap revealed a distinct tissue-specific division of labor among TaAMT members: TaAMT1, TaAMT2, TaAMT6, and others exhibited high expression in roots, suggesting their dominant role in root ammonium uptake. TaAMT13 and TaAMT16 showed high expression in leaves, potentially participating in nitrogen translocation or assimilation in the shoot. TaAMT21 was specifically highly expressed in stems and spikes, implying its regulatory role in nitrogen allocation during reproductive growth stages. This tissue specificity is highly conserved compared to the patterns of AtAMT1;1 and OsAMT1;1 [15,66], reflecting the central role of AMT genes in plant nitrogen uptake.
Notably, TaAMT9 and TaAMT11 were highly expressed in roots but significantly downregulated within 24h under low ammonium stress, while their expression rebounded to 1.6-1.8 times the control level at 48h under high ammonium stress. This dynamic pattern of “low ammonium inhibition-high ammonium activation” resembles the phosphorylation-mediated regulation mechanism of AtAMT1;3. It likely involves perceiving extracellular NH4+ concentration changes to dynamically adjust transport activity, thereby avoiding energy waste under low nitrogen conditions or toxicity accumulation under high nitrogen conditions [18]. In contrast, TaAMT13 and TaAMT16 exhibited a compensatory pattern of “delayed upregulation under low ammonium-rapid downregulation under high ammonium”. This implies that they may synergistically maintain nitrogen homeostasis by delaying the activation of ammonium uptake pathways under low ammonium concentrations and implementing negative feedback inhibition under high ammonium concentrations. Research have shown that early low nitrogen pretreatment can enhance nitrogen uptake and assimilation capacity in wheat seedlings, ensuring normal crop growth and high yield [67]. Under appropriate high ammonium conditions, plants can utilize complex regulatory systems to ensure adequate nitrogen uptake while avoiding ammonium toxicity [68,69].
4.4. Pathogen Infection Responses and the “Nitrogen-Disease” Nexus
The expression of TaAMT1, TaAMT2, and TaAMT17 was significantly upregulated 24 hours after infection with the stripe rust fungus, which indicating their potential involvement in nitrogen metabolism reprogramming during early defense responses [70,71]. Pathogen invasion induces nitrogen uptake in the host to support the synthesis of defensive compounds [72]. Meanwhile, TaAMT15 was upregulated after 48 hours, with its peak expression coinciding with a critical phase of hyphal expansion (48-72 hours), suggesting its role in activating downstream defense genes through nitrogen signaling. Under stripe rust stress, some genes exhibited marked upregulation, potentially linked to defensive nitrogen metabolism responses and disease resistance-related nitrogen signaling [70].
Following powdery mildew infection, the significant upregulation of TaAMT1 and TaAMT6 aligns with reports that powdery mildew secretes effector proteins to interfere with host nitrogen uptake, implying that the elevated expression of TaAMT may represent a compensatory strategy by the host to counteract pathogen nitrogen exploitation [71]. Additionally, the sustained high expression of TaAMT16 and TaAMT18 suggests their involvement in long-term regulation of disease resistance, such as maintaining nitrogen metabolism balance to support continuous synthesis of pathogenesis-related proteins. Notably, distinct temporal patterns and intensities of responses among different TaAMT members after powdery mildew infection indicate their potentially diverse functional roles in ammonium homeostasis regulation or defense responses during wheat pathogen infection. However, significant research gaps remain regarding the specific mechanisms of AMT genes in powdery mildew, how they perceive pathogen signals, and their involvement in wheat disease resistance. Further investigations are urgently needed to elucidate the molecular mechanisms underlying the “nitrogen-disease” interaction.
5. Conclusions
In summary, this study systematically characterized the TaAMT gene family members, identifying 21 TaAMT members distributed across 15 chromosomes, with distinct physicochemical properties and plasma membrane localization. Phylogenetic analysis grouped TaAMTs into four clades, mirroring the evolutionary divergence of AMT1/AMT2 subfamilies in Arabidopsis thaliana and rice, suggesting functional specialization in high and low-affinity ammonium uptake. Cis-element analysis revealed enrichment of hormone-responsive (e.g., ABA, MeJA) and abiotic stress-related motifs in their promoters, aligning with tissue-specific expression patterns, indicating roles in nitrogen homeostasis under stress. Under ammonium stress, TaAMT13 and TaAMT16 showed delayed upregulation under low-N conditions and downregulation under high-N, while TaAMT9 and TaAMT11 exhibited dynamic responses, suggesting adaptive nitrogen sensing. During stripe rust and powdery mildew infections, TaAMT1,TaAMT2, and TaAMT15 were significantly upregulated, implying dual roles in nitrogen metabolism and disease resistance. These findings provide genetic resources and mechanistic insights for improving nitrogen use efficiency and stress resilience in wheat through targeted breeding.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/doi/s1, Table S1: TaAMT members rename.
Author Contributions
Conceptualization, Y.W. and C.G.; methodology, Y.W.; software, J.L.; validation, X.L., and R.H., ; formal analysis,Y.W.; resources, M.L., X.L. and C.G.; writing—original draft preparation, C.G. and Y.W.; writing—review and editing, R.H.; supervision, Y.Z.; project administration, Y.W.; funding acquisition, X.L., Y.W., and M.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by Youth Fund of Basic Scientific Research of Shanxi Province (202203021212433), the outstanding doctoral program of Shanxi Agricultural University (2021BD119 & 2023BQ96), Scientific Research Project of Shanxi Province Work Award Fund (SXBYKY2022089), Scientific and Technological Innovation Promotion Project of Shanxi Agricultural University (CXGC202446), Central guidance for local scientific and technological development funding projects (YDZJSX20231C011), The project of shanxi Province key labconstruction (Z135050009017-1-12), and Wheat Seed Industry Innovation and Joint Breeding Project (NYGG26-01).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the first author.
Acknowledgments
Thanks to the Peking University Institute of Advanced Agricultural Sciences for providing the stripe rust fungus CYR34.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AMT | Ammonium transporter |
| RT-qPCR | Reverse Transcription Quantitative Polymerase Chain Reaction |
| GRAVY | Grand Average of Hydropathy |
| pI | isoelectric points |
| ABA | abscisic acid |
| MBS | Matrix Binding Site |
| LTR | Long Terminal Repeat |
| MRE | Metal Response Element |
| MBSI | Matrix Binding Site I |
Appendix A

Table A1.
Primer sequences used in this study.
| Gene | Forward primer sequence | Reverse primer sequence |
| TaAMT1 | GGGTGGGACCGAGGCTGAAG | CGTGATGTTGGCGGCGTAGC |
| TaAMT2 | CAGTTCGAGTTCGCCGCCATC | TGTAGGAGAGCATGAGCCAGAGC |
| TaAMT6 | GGTCCGCCCTCCTCCAGAAG | TGGGTGTGCAGGGTGGTGAG |
| TaAMT9 | CCCACGCAGCCCTACTACCC | TGAACGCCATCCACGCCTTG |
| TaAMT11 | CTGCTCCTCTCCTACACCGTCTG | ATGACGTAGCCGCCGGAGTAG |
| TaAMT13 | CGTCGTCATCCGCTTAGTTGTCC | GTGTCATCGTAGTGCTCGCCATC |
| TaAMT15 | CAGTTCGTCTTCGCCGCCATC | CCGACGGTGTAGGAGAAGGTGAG |
| TaAMT16 | TCTGTGTTGTCATCCGCCITGTC | GTGTCATCGTAGTGCTCGCCATC |
| TaAMT17 | CAGTTCGTCTTCGCCGCCATC | CCGACGGTGTAGGAGAAGGTGAG |
| TaAMT18 | GTCGTCATCCGCCTTGTCGTC | GTGTCATCGTAGTGCTCGCCATC |
| TaActin | ACCTTCAGTTGCCCAGCAAT | CAGAGTCGAGCACAATACCAGTTG |
References
- Wei, W.; Aifu, L.; Zhihua, Z.; Chengcai, C. Posttranslational Modifications: Regulation of Nitrogen Utilization and Signaling. Plant Cell Physiol 2021, 62. [CrossRef]
- Fujita, T.; Beier, M.P.; Tabuchi-Kobayashi, M.; Hayatsu, Y.; Nakamura, H.; Umetsu-Ohashi, T.; Sasaki, K.; Ishiyama, K.; Murozuka, E.; Kojima, M.; et al. Cytosolic Glutamine Synthetase GS1;3 Is Involved in Rice Grain Ripening and Germination. Front Plant Sci 2022, 13, 835835. [CrossRef]
- Oldroyd, G.E.D.; Leyser, O. A plant’s diet, surviving in a variable nutrient environment. Science 2020, 368. [CrossRef]
- Hao, D.L.; Zhou, J.Y.; Yang, S.Y.; Qi, W.; Yang, K.J.; Su, Y.H. Function and Regulation of Ammonium Transporters in Plants. Int J Mol Sci 2020, 21. [CrossRef]
- Ma, L.; Qin, D.B.; Sun, L.; Zhang, K.; Yu, X.; Dang, A.K.; Hou, S.; Zhao, X.; Yang, Y.; Wang, Y.; et al. Salt overly sensitive2 and ammonium transporter1;1 contribute to plant salt tolerance by maintaining ammonium uptake. Plant Cell 2025, 37. [CrossRef]
- Fang, L.; Wang, M.; Chen, X.; Zhao, J.; Wang, J.; Liu, J. Analysis of the AMT gene family in chili pepper and the effects of arb uscular mycorrhizal colonization on the expression patterns of CaAMT2 genes. BMC genomics 2023, 24, 158. [CrossRef]
- Chen, M.; Zhu, K.; Xie, J.; Liu, J.; Tan, P.; Peng, F. Genome-wide Identification and expression analysis of AMT and NRT gene family in Pecan (Carya illinoinensis) Seedlings Revealed a Preference for NH4+-N. Int J Mol Sci 2022, 23, 13314. [CrossRef]
- Wu, X.; Yang, H.; Qu, C.; Xu, Z.; Li, W.; Hao, B.; Yang, C.; Sun, G.; Liu, G. Sequence and expression analysis of the AMT gene family in poplar. Front Plant Sci 2015, 6, 337. [CrossRef]
- Wang, X.; Wu, H.; Manzoor, N.; Dongcheng, W.; Su, Y.; Liu, Z.; Lin, C.; Mao, Z. The identification of AMT family genes and their expression, function, and regulation in Chenopodium quinoa. Plants (Basel) 2024, 13, 3524. [CrossRef]
- Sonoda, Y.; Ikeda, A.; Saiki, S.; Yamaya, T.; Yamaguchi, J. Feedback regulation of the ammonium transporter gene family AMT1 by glutamine in rice. Plant Cell Physiol 2003, 44, 1396-1402. [CrossRef]
- Gu, R.; Duan, F.; An, X.; Zhang, F.; von Wirén, N.; Yuan, L. Characterization of AMT-mediated high-affinity ammonium uptake in roots of maize (Zea mays L.). Plant Cell Physiol 2013, 54, 1515-1524. [CrossRef]
- Hui, J.; An, X.; Li, Z.; Neuhäuser, B.; Ludewig, U.; Wu, X.; Schulze, W.X.; Chen, F.; Feng, G.; Lambers, H.; et al. The mycorrhiza-specific ammonium transporter ZmAMT3;1 mediates mycorrhiza-dependent nitrogen uptake in maize roots. Plant Cell 2022, 34, 4066-4087. [CrossRef]
- Loque, D.; Yuan, L.; Kojima, S.; Gojon, A.; Wirth, J.; Gazzarrini, S.; Ishiyama, K.; Takahashi, H.; von Wiren, N. Additive contribution of AMT1;1 and AMT1;3 to high-affinity ammonium uptake across the plasma membrane of nitrogen-deficient Arabidopsis roots. Plant J 2006, 48, 522-534. [CrossRef]
- Wang, Q.; Zhao, Y.; Luo, W.; Li, R.; He, Q.; Fang, X.; Michele, R.D.; Ast, C.; von Wirén, N.; Lin, J. Single-particle analysis reveals shutoff control of the Arabidopsis ammonium transporter AMT1;3 by clustering and internalization. Proc Natl Acad Sci U S A 2013, 110, 13204-13209. [CrossRef]
- Yuan, L.; Loque, D.; Kojima, S.; Rauch, S.; Ishiyama, K.; Inoue, E.; Takahashi, H.; von Wiren, N. The organization of high-affinity ammonium uptake in Arabidopsis roots depends on the spatial arrangement and biochemical properties of AMT1-type transporters. Plant Cell 2007, 19, 2636-2652. [CrossRef]
- von Wiren, N.; Gazzarrini, S.; Gojon, A.; Frommer, W.B. The molecular physiology of ammonium uptake and retrieval. Curr Opin Plant Biol 2000, 3, 254-261. PMID: 10837267.
- Loque, D.; Lalonde, S.; Looger, L.L.; von Wiren, N.; Frommer, W.B. A cytosolic trans-activation domain essential for ammonium uptake. Nature 2007, 446, 195-198. [CrossRef]
- Wu, X.; Liu, T.; Zhang, Y.; Duan, F.; Neuhauser, B.; Ludewig, U.; Schulze, W.X.; Yuan, L. Ammonium and nitrate regulate NH4+ uptake activity of Arabidopsis ammonium transporter AtAMT1;3 via phosphorylation at multiple C-terminal sites. J Exp Bot 2019, 70, 4919-4930. [CrossRef]
- Sasaki, A.; Ashikari, M.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Swapan, D.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.S.; et al. Green revolution: a mutant gibberellin-synthesis gene in rice. Nature 2002, 416, 701-702. [CrossRef]
- Peng, J.; Richards, D.E.; Hartley, N.M.; Murphy, G.P.; Devos, K.M.; Flintham, J.E.; Beales, J.; Fish, L.J.; Worland, A.J.; Pelica, F.; et al. ‘Green revolution’ genes encode mutant gibberellin response modulators. Nature 1999, 400, 256-261. [CrossRef]
- Li, S.; Tian, Y.; Wu, K.; Ye, Y.; Yu, J.; Zhang, J.; Liu, Q.; Hu, M.; Li, H.; Tong, Y.; et al. Modulating plant growth-metabolism coordination for sustainable agriculture. Nature 2018, 560, 595-600. [CrossRef]
- Ray, D.K.; Sloat, L.L.; Garcia, A.S.; Davis, K.F.; Ali, T.; Xie, W. Crop harvests for direct food use insufficient to meet the UN’s food security goal. Nat Food 2022, 3, 367-374. [CrossRef]
- Wang, C.; Shen, Y.; Fang, X.T.; Xiao, S.Q.; Liu, G.Y.; Wang, L.G.; Gu, B.J.; Zhou, F.; Chen, D.L.; Tian, H.Q.; et al. Reducing soil nitrogen losses from fertilizer use in global maize and wheat production. Nature Geoscience 2024, 17, 1008-1015. [CrossRef]
- Wang, F.; Yoshida, H.; Matsuoka, M. Making the ‘Green Revolution’ truly green: improving crop nitrogen use efficiency. Plant Cell Physiol 2021, 62, 942-947. [CrossRef]
- Islam, S.; Zhang, J.; Zhao, Y.; She, M.; Ma, W. Genetic regulation of the traits contributing to wheat nitrogen use efficiency. Plant Sci 2021, 303, 110759. [CrossRef]
- Liu, Q.; Wu, K.; Song, W.; Zhong, N.; Wu, Y.; Fu, X. Improving crop nitrogen use efficiency toward sustainable green revolution. Annu Rev Plant Biol 2022, 73, 523-551. [CrossRef]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu Rev Plant Biol 2012, 63, 153-182. [CrossRef]
- Vidal, E.A.; Alvarez, J.M.; Araus, V.; Riveras, E.; Brooks, M.D.; Krouk, G.; Ruffel, S.; Lejay, L.; Crawford, N.M.; Coruzzi, G.M.; et al. Nitrate in 2020: Thirty tears from transport to signaling networks. Plant cell 2020, 32, 2094-2119. [CrossRef]
- Li, L.; Liu, K.H.; Sheen, J. Dynamic nutrient signaling networks in plants. Annu Rev Cell Dev Biol 2021, 37, 341-367. [CrossRef]
- Shanks, C.M.; Rothkegel, K.; Brooks, M.D.; Cheng, C.Y.; Alvarez, J.M.; Ruffel, S.; Krouk, G.; Gutiérrez, R.A.; Coruzzi, G.M. Nitrogen sensing and regulatory networks: it’s about time and space. Plant Cell 2024, 36, 1482-1503. [CrossRef]
- Shi, H.W.; Wang, W.C.; Gao, L.F.; Wu, J.R.; Hu, C.M.; Yan, H.S.; Shi, Y.G.; Li, N.; Ma, Y.Z.; Zhou, Y.B.; et al. Genome-wide association study of seedling nitrogen-use efficiency-associated traits in common wheat. Crop J 2024, 12, 222-231. [CrossRef]
- Wu, X.X.; Yuan, P.; Chen, H.; Kumar, V.; Kang, S.M.; Jia, B.; Xuan, Y.H. Ammonium transporter 1 increases rice resistance to sheath blight by promoting nitrogen assimilation and ethylene signalling. Plant Biotechnol J 2022, 20, 1085-1097. [CrossRef]
- Li, T.; Liao, K.; Xu, X.; Gao, Y.; Wang, Z.; Zhu, X.; Jia, B.; Xuan, Y. Wheat Ammonium Transporter (AMT) gene gamily: diversity and possible role in Host-Pathogen interaction with stem rust. Front Plant Sci 2017, 8, 1637. [CrossRef]
- Jiang, J.; Zhao, J.; Duan, W.; Tian, S.; Wang, X.; Zhuang, H.; Fu, J.; Kang, Z. TaAMT2;3a, a wheat AMT2-type ammonium transporter, facilitates the infection of stripe rust fungus on wheat. BMC Plant Biol 2019, 19, 239. [CrossRef]
- Muratore, C.; Espen, L.; Prinsi, B. Nitrogen Uptake in Plants: The plasma membrane root transport systems from a physiological and proteomic perspective. Plants (Basel) 2021, 10. [CrossRef]
- Wang Y, Cao Q, Zhang J, Wang S, Chen C, Wang C, Zhang H, Wang Y, Ji W. Cytogenetic Analysis and Molecular Marker Development for a new wheat-Thinopyrum ponticum 1Js (1D) disomic substitution line with resistance to stripe rust and powdery mildew. Front Plant Sci 2020, 11, 1282. [CrossRef]
- Bolser, D.; Staines, D.M.; Pritchard, E.; Kersey, P. Ensembl Plants: integrating tools for visualizing, mining, and analyzing plant genomics data. Methods Mol Bio 2016, 1374, 115-40. [CrossRef]
- Chenna, R.; Sugawara, H.; Koike, T.; Lopez, R.; Gibson, T.J.; Higgins, D.G.; Thompson, J.D. Multiple sequence alignment with the Clustal series of programs. Nucleic Acids Res 2003, 31, 3497-3500. [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol Biol Evol 2016, 33, 1870-1874. [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v6: recent updates to the phylogenetic tree display and annotation tool. Nucleic Acids Res 2024, 52, W78-W82. [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.Y.; Li, W.W.; Noble, W.S. MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res 2009, 37, W202-W208. [CrossRef]
- Ma, S.; Wang, M.; Wu, J.; Guo, W.; Chen, Y.; Li, G.; Wang, Y.; Shi, W.; Xia, G.; Fu, D.; et al. WheatOmics: A platform combining multiple omics data to accelerate functional genomics studies in wheat. Mol Plant 2021, 14, 1965-1968. [CrossRef]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res 2002, 30, 325-327. [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol Plant 2020, 13, 1194-1202. [CrossRef]
- Zorb, C.; Ludewig, U.; Hawkesford, M.J. Perspective on wheat yield and quality with reduced nitrogen supply. Trends Plant Sci 2018, 23, 1029-1037. [CrossRef]
- Hawkesford, M.J. Genetic variation in traits for nitrogen use efficiency in wheat. J Exp Bot 2017, 68, 2627-2632. [CrossRef]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu Rev Plant Biol 2012, 63, 153-182. [CrossRef]
- Li, H.; Zhu, X.; Wang, J.; Wei, Y.; Nai, F.; Yu, H.; Wang, X. Unraveling differential characteristics and mechanisms of nitrogen uptake in wheat cultivars with varied nitrogen use efficiency. Plant Physiol Biochem 2024, 206, 108278. [CrossRef]
- Keerthana, K.; Ramakrishnan, M.; Ahmad, Z.; Amali, P.; Vijayakanth, V.; Wei, Q. Root-derived small peptides: Key regulators of plant development, stress resilience, and nutrient acquisition. Plant Sci 2025, 354, 112433. [CrossRef]
- Tsay, Y.-F.; Ho, C.-H.; Chen, H.-Y.; Lin, S.-H. Integration of Nitrogen and Potassium Signaling. Annual Review of Plant Biology 2011, 62, 207-226. [CrossRef]
- Wang, H.; Ma, Q.; Shan, F.; Tian, L.; Gong, J.; Quan, W.; Yang, W.; Hou, Q.; Zhang, F.; Zhang, S. Transcriptional regulation mechanism of wheat varieties with different nitrogen use efficiencies in response to nitrogen deficiency stress. BMC Genomics 2022, 23, 727. [CrossRef]
- Zhang, Y.; Zhao, Y.; Sun, L.; Han, P.; Bai, X.; Lin, R.; Xiao, K. The N uptake-associated physiological processes at late growth stage in wheat (Triticum aestivum) under N deprivation combined with deficit irrigation condition. Plant Physiol Biochem 2021, 164, 160-172. [CrossRef]
- Liu, B.; Xu, W.; Niu, Y.; Li, Q.; Cao, B.; Qi, J.; Zhao, Y.; Zhou, Y.; Song, L.; Cui, D.; et al. TaTCP6 is required for efficient and balanced utilization of nitrate and phosphorus in wheat. Nat Commun 2025, 16, 1683. [CrossRef]
- Zhang, H.; Jin, Z.; Cui, F.; Zhao, L.; Zhang, X.; Chen, J.; Zhang, J.; Li, Y.; Li, Y.; Niu, Y.; et al. Epigenetic modifications regulate cultivar-specific root development and metabolic adaptation to nitrogen availability in wheat. Nat Commun 2023, 14, 8238. [CrossRef]
- International Wheat Genome Sequencing, C. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [CrossRef]
- Li, T.; Kong, C.; Deng, P.; Li, C.; Zhao, G.; Li, H.; Gao, L.; Cui, D.; Jia, J. Intra-Varietal diversity and its contribution to wheat evolution, domestication, and improvement in wheat. Int J Mol Sci 2023, 24. [CrossRef]
- Howitt, S.M.; Udvardi, M.K. Structure, function and regulation of ammonium transporters in plants. Biochim Biophys Acta 2000, 1465, 152-170. [CrossRef]
- Hao, D.L.; Yang, S.Y.; Liu, S.X.; Zhou, J.Y.; Huang, Y.N.; Very, A.A.; Sentenac, H.; Su, Y.H. Functional Characterization of the Arabidopsis Ammonium Transporter AtAMT1;3 With the Emphasis on Structural Determinants of Substrate Binding and Permeation Properties. Front Plant Sci 2020, 11, 571. [CrossRef]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313-324. [CrossRef]
- Chinnusamy, V.; Schumaker, K.; Zhu, J.K. Molecular genetic perspectives on cross-talk and specificity in abiotic stress signalling in plants. J Exp Bot 2004, 55, 225-236. [CrossRef]
- Ma, X.; Wang, W.; Zhang, J.; Jiang, Z.; Xu, C.; Zhu, W.; Shi, B.; Yang, W.; Su, H.; Wang, X.; et al. NRT1.1B acts as an abscisic acid receptor in integrating compound environmental cues for plants. Cell 2025. [CrossRef]
- Garnica, M.; Houdusse, F.; Zamarreño, A.M.; Garcia-Mina, J.M. The signal effect of nitrate supply enhances active forms of cytokinins and indole acetic content and reduces abscisic acid in wheat plants grown with ammonium. J Plant Physiol 2010, 167, 1264-1272. [CrossRef]
- Oka, M.; Shimoda, Y.; Sato, N.; Inoue, J.; Yamazaki, T.; Shimomura, N.; Fujiyama, H. Abscisic acid substantially inhibits senescence of cucumber plants (Cucumis sativus) grown under low nitrogen conditions. J Plant Physiol 2012, 169, 789-796, doi:/10.1016/j.jplph.2012.02.001.
- Liu, H.; Carvalhais, L.C.; Kazan, K.; Schenk, P.M. Development of marker genes for jasmonic acid signaling in shoots and roots of wheat. Plant Signal Behav 2016, 11, e1176654. [CrossRef]
- Fernandez-Milmanda, G.L.; Crocco, C.D.; Reichelt, M.; Mazza, C.A.; Kollner, T.G.; Zhang, T.; Cargnel, M.D.; Lichy, M.Z.; Fiorucci, A.S.; Fankhauser, C.; et al. A light-dependent molecular link between competition cues and defence responses in plants. Nat Plants 2020, 6, 223-230. [CrossRef]
- Kumar, A.; Silim, S.N.; Okamoto, M.; Siddiqi, M.Y.; Glass, A.D. Differential expression of three members of the AMT1 gene family encoding putative high-affinity NH4+ transporters in roots of Oryza sativa subspecies indica. Plant Cell Environ 2003, 26, 907-914. [CrossRef]
- Lu, Y.; Gao, L.; Hu, J.; Liu, X.; Jiang, D.; Cao, W.; Dai, T.; Tian, Z. Low nitrogen priming improves nitrogen uptake and assimilation adaptation to nitrogen deficit stress in wheat seedling. Planta 2024, 259, 107. [CrossRef]
- Liu, Y.; von Wiren, N. Ammonium as a signal for physiological and morphological responses in plants. J Exp Bot 2017, 68, 2581-2592. [CrossRef]
- Zhao, R.; Cao, Y.; Ge, Y.; Xu, J.; Li, R.; Yang, M.; Chen, Y.; Wu, D.; Xiao, J.; Li, R. Single-Molecule and Vesicle Trafficking Analysis of Ubiquitination Involved in the Activity of Ammonium Transporter AMT1;3 in Arbidopsis under high ammonium Stress. Cells-Basel 2022, 11. [CrossRef]
- Devadas, R.; Simpfendorfer, S.; Backhouse, D.; Lamb, D.W. Effect of stripe rust on the yield response of wheat to nitrogen. Crop J 2014, 2, 201-206. [CrossRef]
- Bolton, M.D.; Thomma, B.P.H.J. The complexity of nitrogen metabolism and nitrogen-regulated gene expression in plant pathogenic fungi. Physiol Mol Plant P 2008, 72, 104-110. [CrossRef]
- Sun, Y.; Wang, M.; Mur, L.A.J.; Shen, Q.; Guo, S. Unravelling the roles of nitrogen nutrition in plant disease defences. Int J Mol Sci 2020, 21. [CrossRef]
Figure 1.
Phylogenetic analysis of AMT proteins were from Arabidopsis thaliana (At), Oryza sativa Japonica (Os), and Triticum aestivum (Ta). The tree was constructed using the Maximum Likelihood (ML) method implemented in MEGA 11 (version 11.0.13).
Figure 1.
Phylogenetic analysis of AMT proteins were from Arabidopsis thaliana (At), Oryza sativa Japonica (Os), and Triticum aestivum (Ta). The tree was constructed using the Maximum Likelihood (ML) method implemented in MEGA 11 (version 11.0.13).
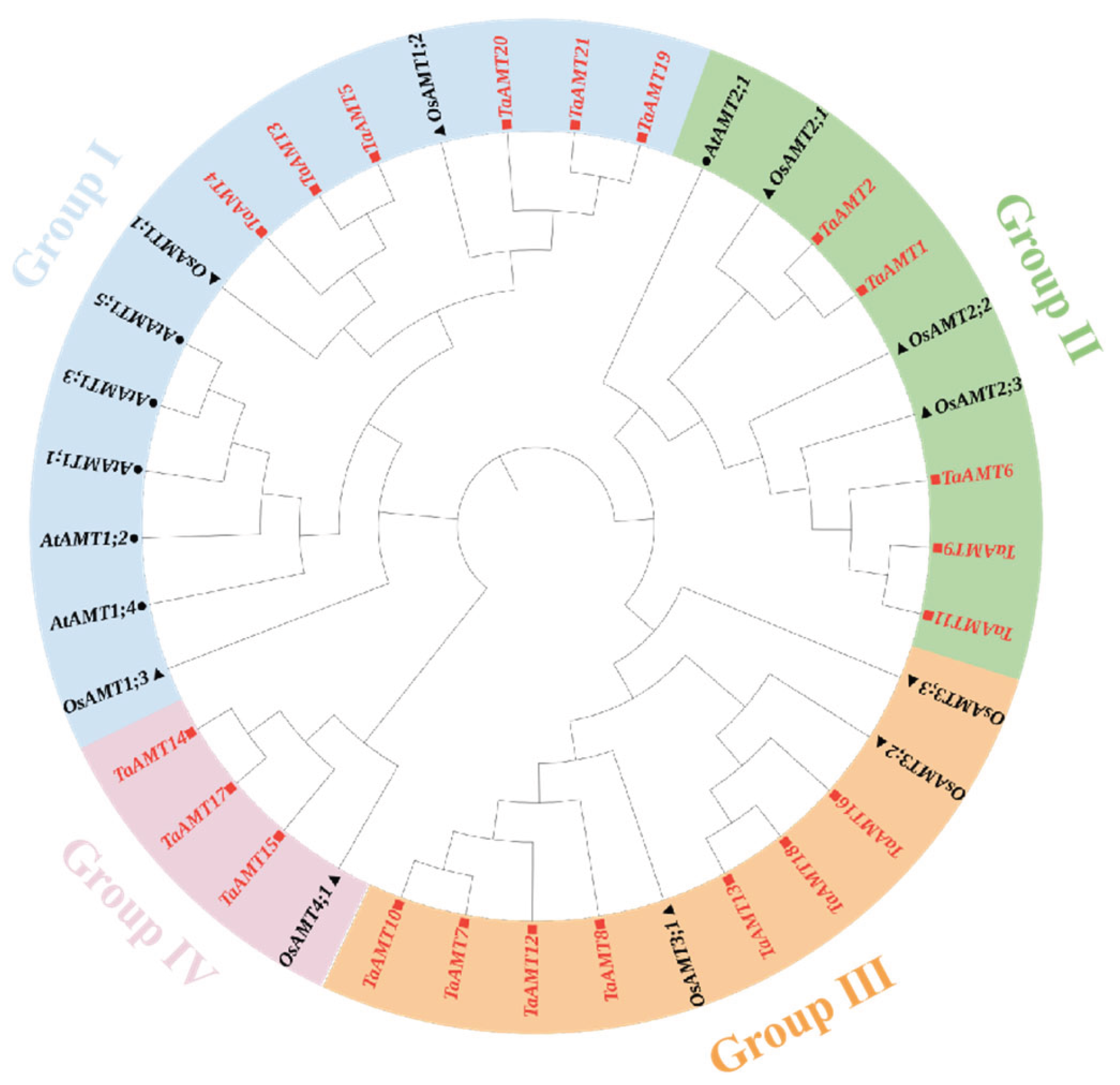
Figure 2.
Comprehensive analysis of TaAMT gene family members. (A) Phylogenetic tree showing evolutionary relationships. (B) MEME-based visualization of conserved motifs, with colored boxes representing specific conserved amino acid sequences. (C) Conserved domain architecture inferred from NCBI-CDD analysis, with color-coded boxes indicating phylogenetically conserved functional modules. (D) Gene structure analysis depicting exon-intron organization of TaAMT members.
Figure 2.
Comprehensive analysis of TaAMT gene family members. (A) Phylogenetic tree showing evolutionary relationships. (B) MEME-based visualization of conserved motifs, with colored boxes representing specific conserved amino acid sequences. (C) Conserved domain architecture inferred from NCBI-CDD analysis, with color-coded boxes indicating phylogenetically conserved functional modules. (D) Gene structure analysis depicting exon-intron organization of TaAMT members.
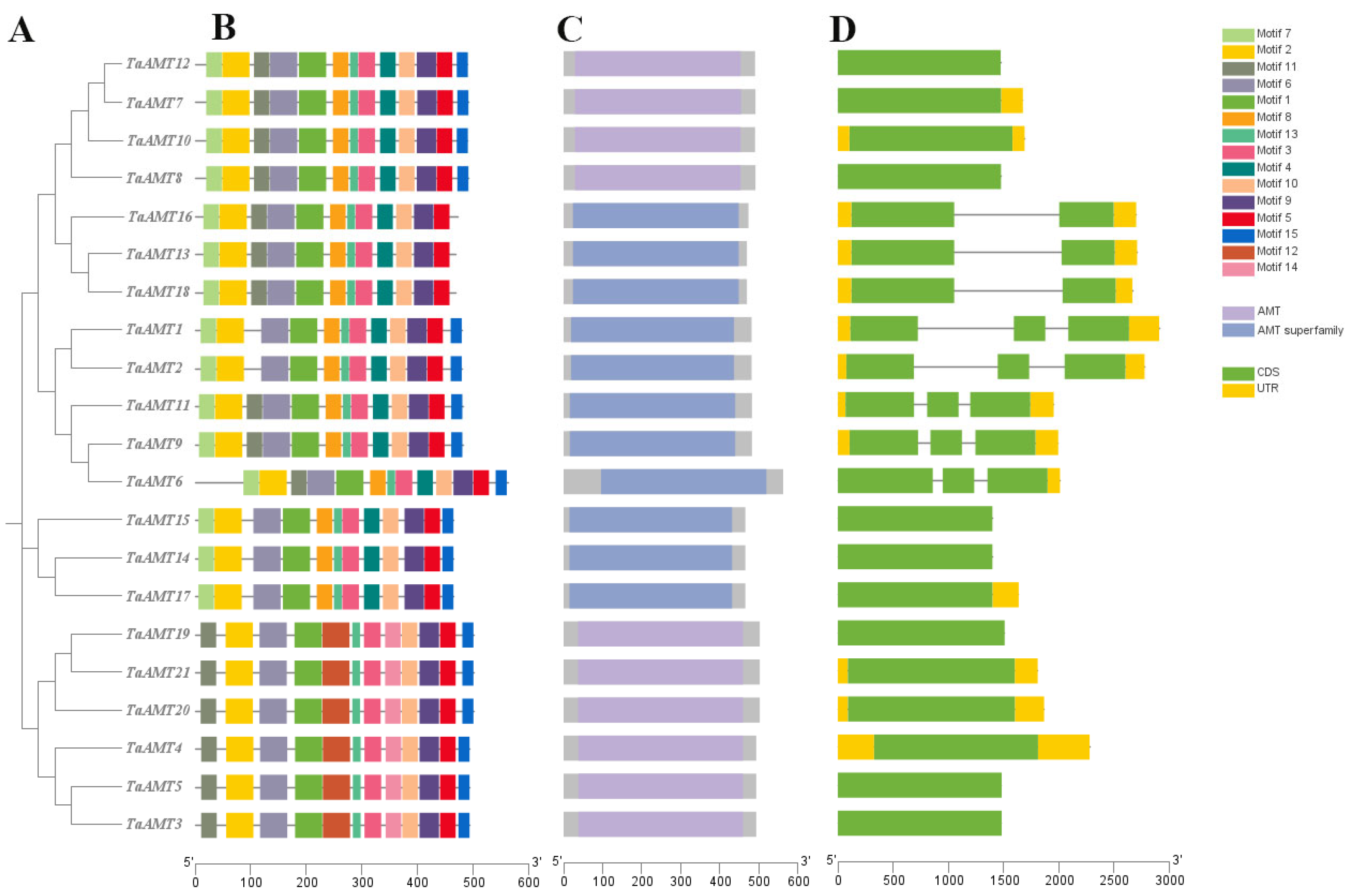
Figure 3.
Chromosome distribution of TaAMT members in wheat genome.
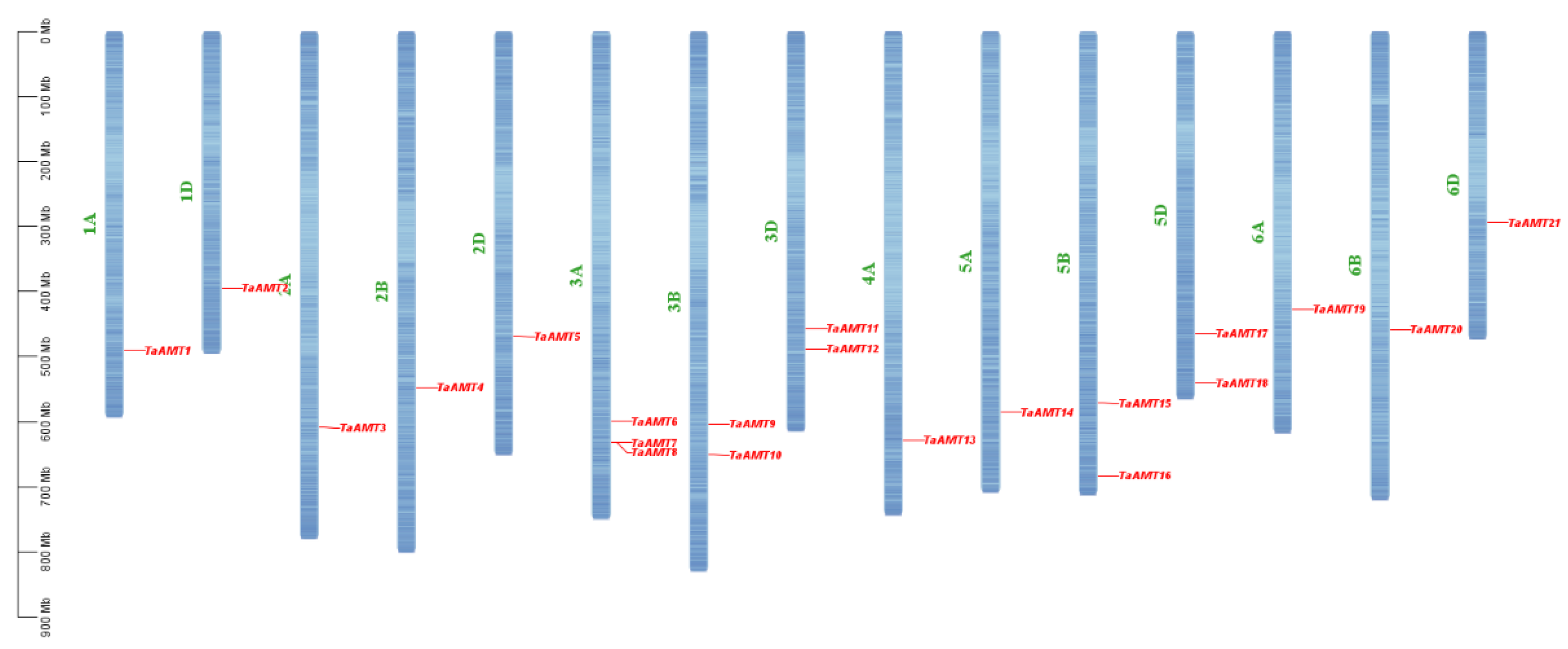
Figure 4.
Genomic collinearity analysis of TaAMT gene family members. Red lines indicate gene pairs with collinearity relationships,numbers within the boxes represent chromosomes, and light blue boxes denote gene density.
Figure 4.
Genomic collinearity analysis of TaAMT gene family members. Red lines indicate gene pairs with collinearity relationships,numbers within the boxes represent chromosomes, and light blue boxes denote gene density.
Figure 5.
Interspecific collinearity analysis of AMT gene family members. Red lines indicate collinear gene pairs between wheat (Ta) and other plant species, with chromosomal positions shown in circular layout. Species abbreviations: Ta, Triticum aestivum (wheat); At, Arabidopsis thaliana; Os, Oryza sativa (rice).
Figure 5.
Interspecific collinearity analysis of AMT gene family members. Red lines indicate collinear gene pairs between wheat (Ta) and other plant species, with chromosomal positions shown in circular layout. Species abbreviations: Ta, Triticum aestivum (wheat); At, Arabidopsis thaliana; Os, Oryza sativa (rice).
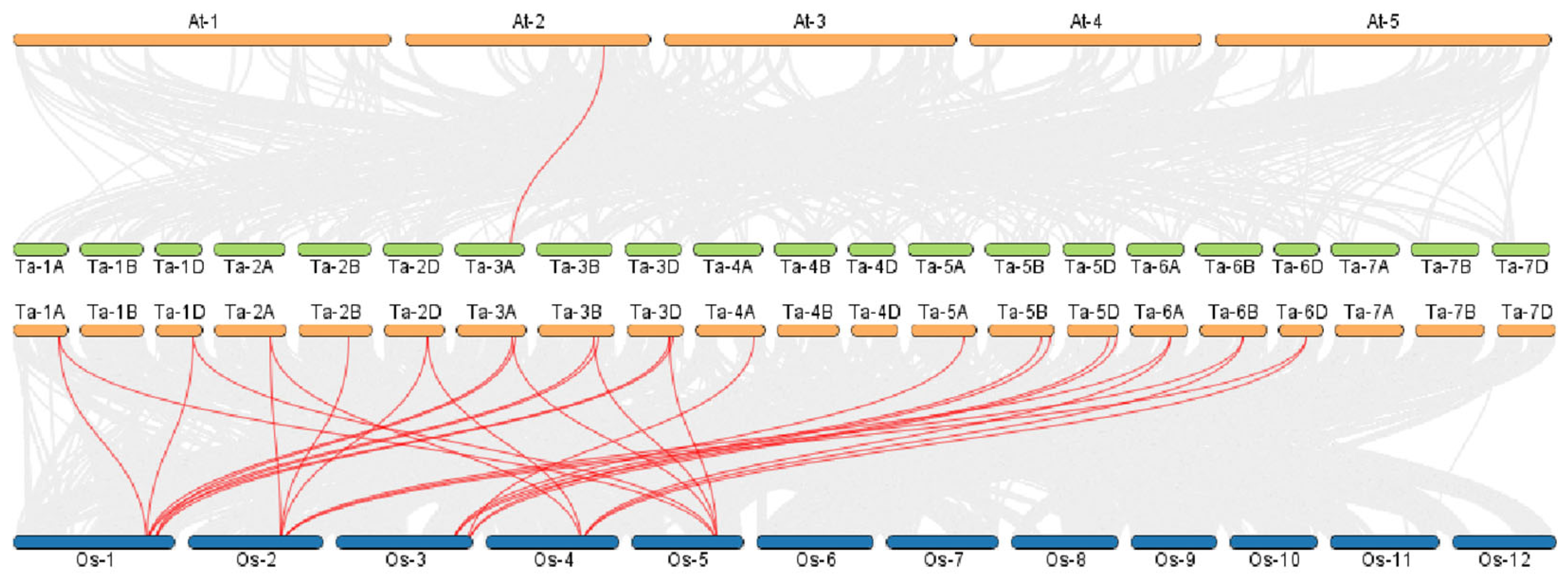
Figure 6.
Analysis of cis-acting elements in TaAMT members. Boxes of different colors represent distinct cis-acting elements, with their names listed on the right.
Figure 6.
Analysis of cis-acting elements in TaAMT members. Boxes of different colors represent distinct cis-acting elements, with their names listed on the right.
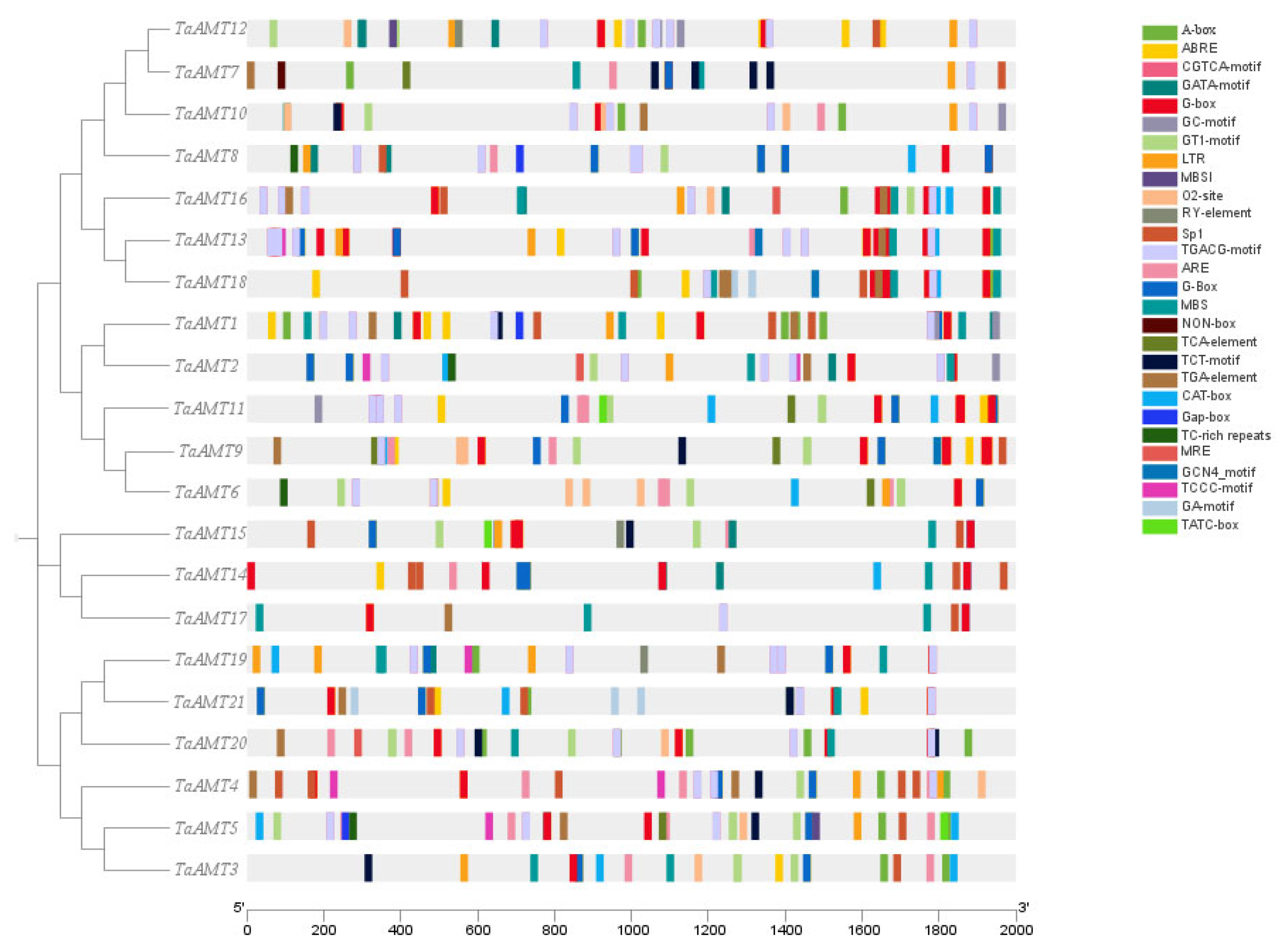
Figure 7.
Heatmap of TaAMT members expression across different growth stages from the ExpVIP database.
Figure 7.
Heatmap of TaAMT members expression across different growth stages from the ExpVIP database.
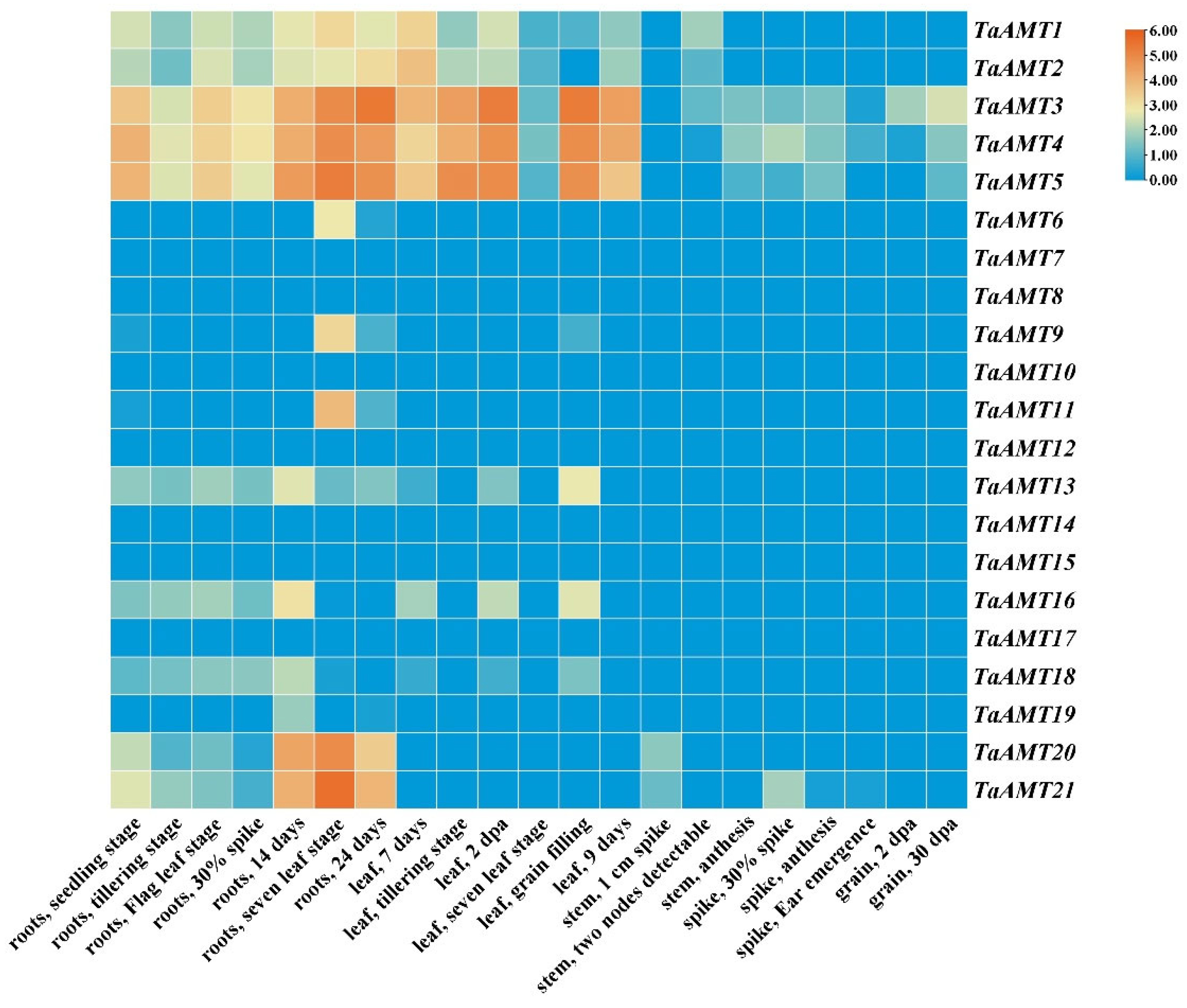
Figure 8.
Analysis of expression patterns of four TaAMT members under high-ammonium (A, Blue, 40 mM) and low-ammonium (B, Red, 0.04 mM) stress conditions. The numbers 1 to 4 represent the genes TaAMT13, TaAMT16, TaAMT9, and TaAMT11, respectively. Values shown are the means ± SD of three replicates. For each datum, bars with different letters indicate significant differences ( Duncan’s test, at P < 0.01 ).
Figure 8.
Analysis of expression patterns of four TaAMT members under high-ammonium (A, Blue, 40 mM) and low-ammonium (B, Red, 0.04 mM) stress conditions. The numbers 1 to 4 represent the genes TaAMT13, TaAMT16, TaAMT9, and TaAMT11, respectively. Values shown are the means ± SD of three replicates. For each datum, bars with different letters indicate significant differences ( Duncan’s test, at P < 0.01 ).
Figure 9.
Expression patterns of four TaAMT members under stripe rust stress (CYR34). **,***, and **** indicate the significant correlations at the levels of P < 0.01, P < 0.001, and P < 0.0001 respectively.
Figure 9.
Expression patterns of four TaAMT members under stripe rust stress (CYR34). **,***, and **** indicate the significant correlations at the levels of P < 0.01, P < 0.001, and P < 0.0001 respectively.
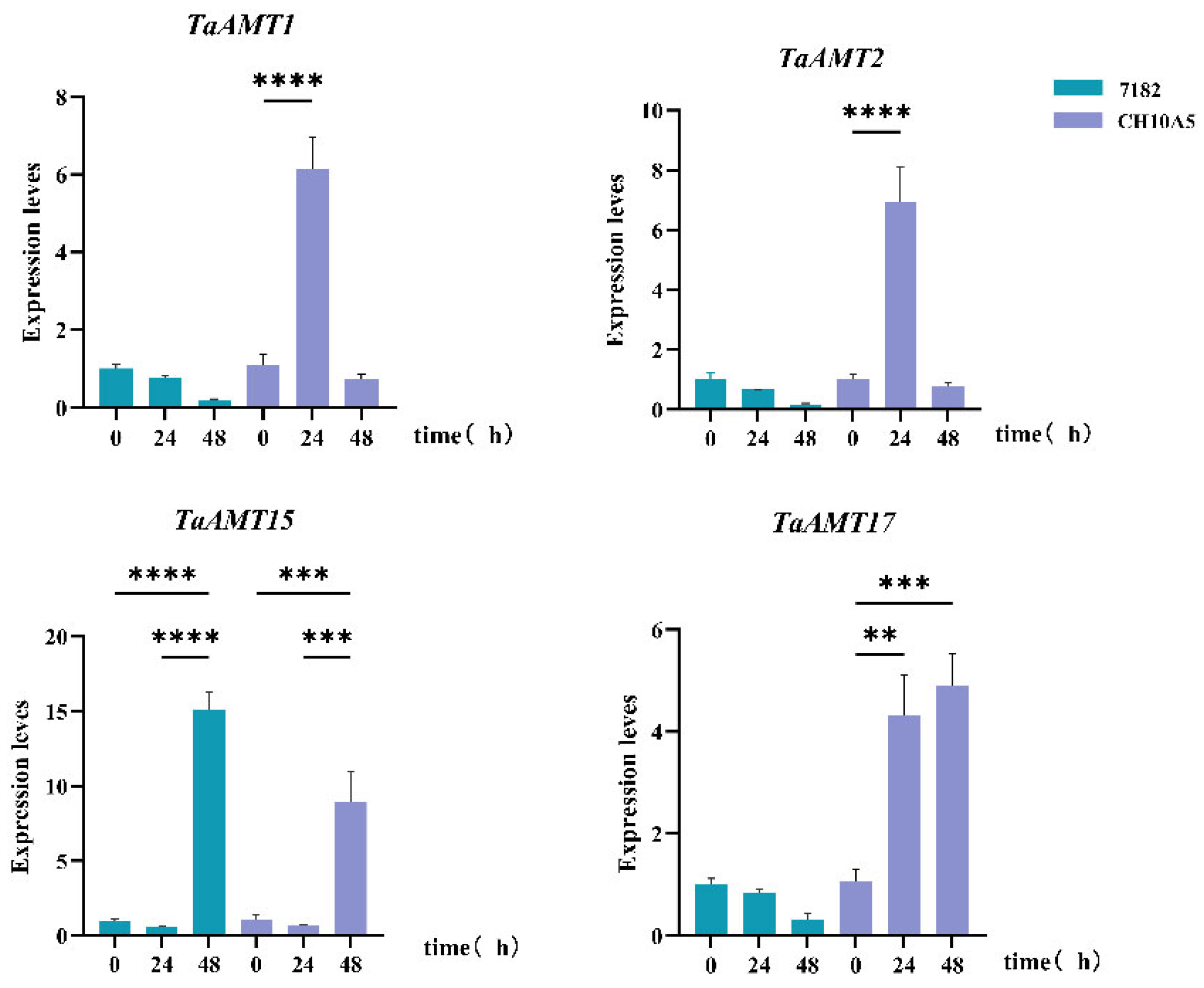
Figure 10.
Gene expression patterns of nine TaAMT members under powdery mildew stress (E09). **,***, and **** indicate the significant correlations at the levels of P < 0.01, P < 0.001, and P < 0.0001, respectively; ns indicate no significant difference.
Figure 10.
Gene expression patterns of nine TaAMT members under powdery mildew stress (E09). **,***, and **** indicate the significant correlations at the levels of P < 0.01, P < 0.001, and P < 0.0001, respectively; ns indicate no significant difference.
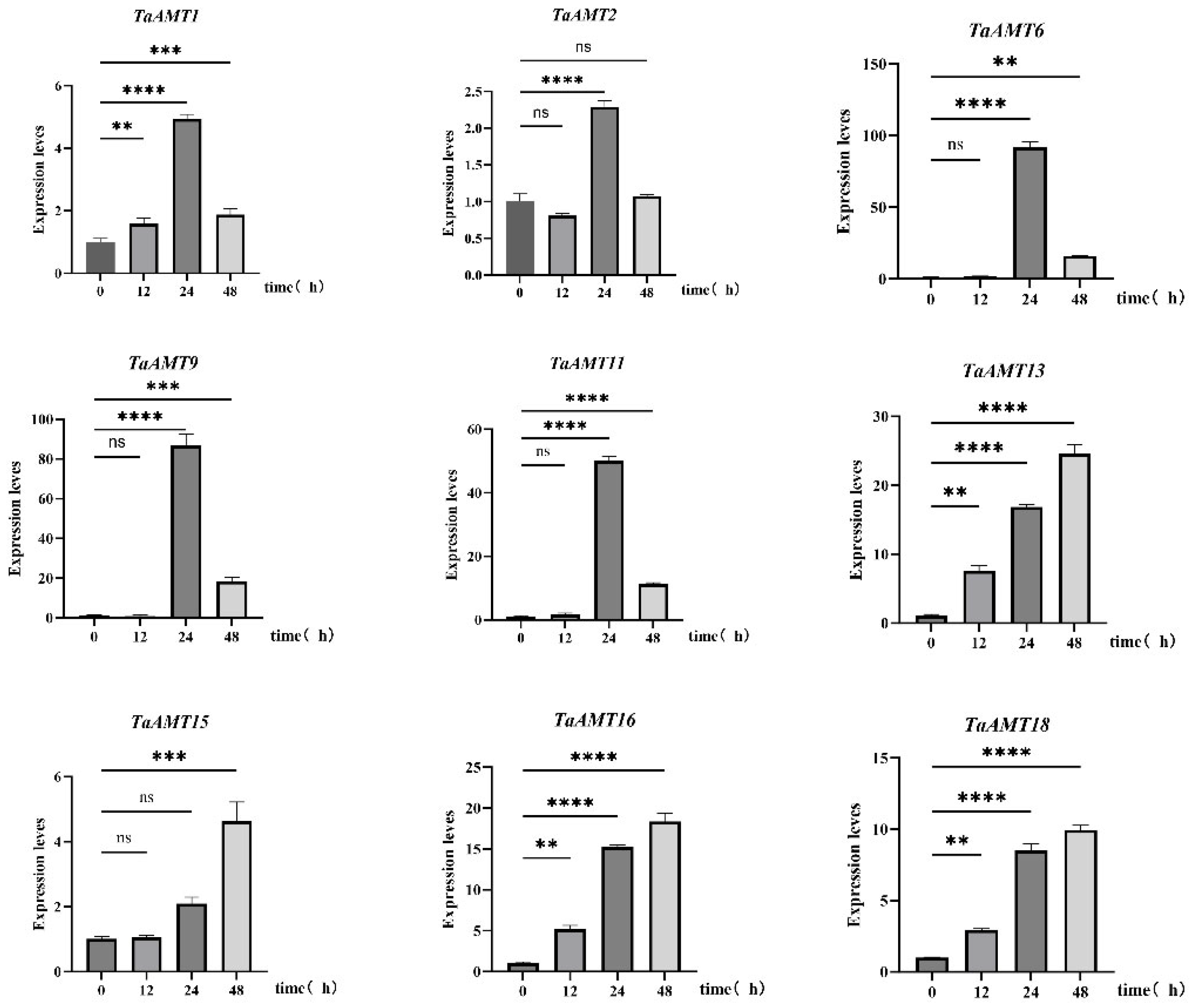
Table 1.
Physicochemical parameters of TaAMT members.
| Name | Number of Amino Acid | Molecular Weight | Theoretical pI | Instability Index | Aliphatic Index | Grand Average of Hydropathicity | Subcellular Localization |
| TaAMT1 | 482 | 51273.1 | 7.75 | 28.77 | 102.45 | 0.541 | Plasma Membrane |
| TaAMT2 | 482 | 51336.2 | 8.67 | 29.15 | 101.43 | 0.537 | Plasma Membrane |
| TaAMT3 | 494 | 52190.17 | 7.15 | 19.04 | 94.11 | 0.520 | Plasma Membrane |
| TaAMT4 | 494 | 52298.33 | 7.15 | 19.59 | 93.32 | 0.523 | Plasma Membrane |
| TaAMT5 | 494 | 52224.19 | 7.15 | 19.04 | 93.32 | 0.518 | Plasma Membrane |
| TaAMT6 | 563 | 59889.76 | 9.13 | 38.08 | 100.36 | 0.427 | Plasma Membrane |
| TaAMT7 | 492 | 53474.26 | 7.63 | 34.41 | 96.30 | 0.420 | Plasma Membrane |
| TaAMT8 | 492 | 53458.29 | 7.10 | 34.55 | 97.13 | 0.452 | Plasma Membrane |
| TaAMT9 | 483 | 51327.01 | 8.28 | 31.13 | 107.52 | 0.583 | Plasma Membrane |
| TaAMT10 | 491 | 53372.22 | 7.63 | 33.25 | 96.90 | 0.434 | Plasma Membrane |
| TaAMT11 | 483 | 51280.94 | 8.28 | 31.53 | 107.70 | 0.580 | Plasma Membrane |
| TaAMT12 | 491 | 53385.21 | 7.63 | 34.29 | 96.90 | 0.428 | Plasma Membrane |
| TaAMT13 | 470 | 50042.43 | 6.49 | 34.26 | 103.11 | 0.625 | Plasma Membrane |
| TaAMT14 | 466 | 50040.55 | 7.61 | 33.81 | 102.62 | 0.613 | Plasma Membrane |
| TaAMT15 | 466 | 49917.42 | 7.07 | 32.81 | 104.72 | 0.630 | Plasma Membrane |
| TaAMT16 | 474 | 50497.9 | 6.30 | 32.68 | 102.05 | 0.596 | Plasma Membrane |
| TaAMT17 | 466 | 49910.41 | 7.61 | 32.72 | 102.83 | 0.626 | Plasma Membrane |
| TaAMT18 | 470 | 50056.46 | 6.49 | 34.08 | 103.11 | 0.625 | Plasma Membrane |
| TaAMT19 | 503 | 52851.72 | 7.62 | 23.94 | 91.87 | 0.514 | Plasma Membrane |
| TaAMT20 | 503 | 52865.74 | 7.62 | 23.94 | 92.07 | 0.514 | Plasma Membrane |
| TaAMT21 | 503 | 52865.1 | 7.62 | 24.10 | 91.67 | 0.509 | Plasma Membrane |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



