
Submitted:
14 November 2025
Posted:
17 November 2025
You are already at the latest version
Abstract
Background: Seasonal influenza remains a significant public health problem, and the constant antigenic drift of viruses requires regular vaccine updates. mRNA vaccines offer a promising platform for the development of new, effective influenza vaccines. Administration of the naked mRNA vaccine using a needle-free jet injection system further enhances its safety, reduces cost, and eliminates the need for lipid nanoparticles, which are traditionally used for mRNA delivery. Lyophilization of naked mRNA allows for longterm storage at +4°C. Methods: We designed and produced an mRNA vaccine against seasonal influenza, designated mRNA-Vector-Flu, encoding the hemagglutinin (HA) of the A/Wisconsin/67/2022(H1N1)pdm09, A/Darwin/9/2021(H3N2), and B/Austria/1359417/2021 strains. The vaccine was lyophilized and stored for 1 month in a refrigerator (+4°C). A comparative immunogenicity study was conducted between synthesized immediately before use prepared and lyophilized naked mRNA-Vector-Flu. The preparations were administered to BALB/c mice using a jet needleless injection twice, 3 weeks apart. Immunogenicity was assessed on day 35 of the study. Results: A comparative immunogenicity study of naked mRNA-Vector-Flu demonstrated that both the synthesized immediately before use prepared formulation and the lyophilized form, stored at +4°C for a month, induced similar levels of virus-specific antibodies and generated a pro-nounced T-cell immune response. Conclusions: Delivery of the naked mRNA vaccine us-ing a needle-free jet injection ensures a high-level immune response, which improves its safety, reduces its cost, and eliminates the need for lipid nanoparticles traditionally used for mRNA delivery. At the same time, lyophilization of the naked mRNA vaccine pre-serves its biological activity and ensures its long-term storage at above-zero temperatures. Our results demonstrate that our proposed approach can be considered a promising di-rection for the development and improvement of the mRNA vaccine platform.
Keywords:
mRNA-vaccine
; seasonal influenza vaccine
; mRNA delivery
; lipid nanoparticles
; jet injection
1. Introduction
Seasonal influenza remains a significant public health problem, with over 3 million influenza-related hospitalizations worldwide annually [1,2] and up to half a million deaths from this disease recorded annually [3]. Vaccination against seasonal influenza is an effective means of eliciting immunity, helping to reduce the significant burden of annual influenza epidemics [4,5].
Various types of influenza vaccines exist, including live attenuated, inactivated (whole-virion, split, subunit), and recombinant vaccines [6,7,8]. However, the constant antigenic drift of circulating influenza viruses renders seasonal influenza vaccines ineffective, necessitating annual re-releases of these vaccines. In this case, mRNA-based vaccine platforms offer advantages over standard influenza vaccine technologies. The low cost of producing mRNA vaccines compared to conventional split subunit vaccines developed using RNA cells and the ability to quickly and easily replace the target gene in mRNA vaccines without changing the production technology itself allows for a timely response to the emergence of new virus strains [9]. mRNA vaccine technology has enabled the rapid creation of effective and safe drugs for the prevention of coronavirus. In total, approximately one billion people worldwide have been vaccinated with mRNA-based SARS-CoV-2 vaccines, and their effectiveness has been demonstrated [10,11]. The production of mRNA vaccines does not require the use of chicken embryos, which avoids mutations in the immunogen sequence during the production of vaccine batches. mRNA vaccines do not induce an unwanted immune response in the recipient, as is typical with vector vaccines, and therefore can be administered multiple times. Another important advantage is the efficient activation of both humoral and cellular virus-specific immunity, since the target protein is expressed endogenously from mRNA [12,13].
Currently, several teams are developing mRNA vaccines against seasonal influenza [14,15,16], and several vaccines, including Moderna’s mRNA-1010, are undergoing clinical trials [17,18,19,20]. Lipid nanoparticles are typically used to deliver mRNA vaccines. Lipid nanoparticles consist of four lipid components that ensure effective encapsulation and release of mRNA, prolonged circulation in the body, and complex stability [21]. However, certain limitations have been identified, related to both the complexity of storage (down to -80 °C) and, consequently, the logistics of such vaccines, and adverse post-vaccination effects. These effects are believed to be largely related to the nature of the lipids coating the mRNA [22,23].
Therefore, alternative delivery methods have recently attracted increasing attention. Thus, the recently published [24,25,26,27] jet injection (JI) method is a promising alternative to LNPs for mRNA delivery.
Jet injection is a physical delivery method in which vaccines and other therapeutic drugs are administered in fractions of a second using a high-speed jet through a nozzle under high pressure. This allows for efficient drug delivery intradermally, intramuscularly, or subcutaneously, without the need for a needle [28]. The most attractive feature of this method is that the mRNA vaccine is administered as a naked molecule, completely eliminating the negative effects of the lipid components encapsulating the mRNA molecule.
However, in this case, the issue of storage and stability of naked mRNA molecule preparations becomes relevant, so we decided to use lyophilization.
The aim of this study was to compare the immunogenicity of the mRNA-Vector-Flu trivalent mRNA vaccine against seasonal influenza virus, synthesized immediately before use and its lyophilized form, administered via jet injection.
2. Materials and Methods
2.1. Bacterial and Viral Strains, Cell Cultures, and Plasmids.
The E. coli Neb Stable strain (New England Biolabs Inc., USA) was used to obtain and produce DNA templates for mRNA synthesis. For in vitro analysis of mRNA functionality, the continuous human embryonic kidney cell line HEK293, provided by the Microorganism Collection Department of the State Research Center of Virology and Biotechnology Vector (Rospotrebnadzor, Koltsovo, Russia), was used.
To obtain DNA templates for mRNA synthesis, plasmids encoding the hemagglutinins of the corresponding influenza virus strains were used: pVAX-H1-24, pVAX-H3-24, and pVAX-HB-24 (FBRI SRC VB Vector, Rospotrebnadzor). To obtain mRNA-GFP, the previously obtained pVAX-C3-GFP matrix (FBRI SRC VB “Vector”, Rospotrebnadzor) was used.
2.2. Templates for mRNA Synthesis.
Hemagglutinins of influenza viruses recommended by WHO for the Northern Hemisphere for the 2023-2024 season [29] were used as antigens for the mRNA vaccine: A/Wisconsin/67/2022(H1N1)pdm09 (EPI_ISL_15928538), A/Darwin/9/2021(H3N2) (EPI_ISL_20142977), B/Austria/1359417/2021 (EPI_ISL_983345) without the transmembrane and cytoplasmic domains. To stabilize the trimeric structure of hemagglutinins, a T4 trimerizing domain was added to the C-terminus [30,31]. To maintain the uncleaved form of the rHA/H1 and rHA/H3 proteins, the following amino acid substitutions were introduced into the pH switch region of the HA2 subunit: H26 and H106, where histidine was replaced by tryptophan (W) and arginine (R), and amino acids K51 and E103 were replaced by isoleucine [32]. The codon composition of the sequence was optimized using the codon adaptation tool (https://www.jcat.de/, accessed September 1, 2025). The genes were cloned into a previously developed DNA cassette to obtain a template for pVAX-C3-PolyA mRNA vaccines [33], carrying the target gene under the control of the T7 promoter modified for efficient incorporation of the AG-Cap analogue during mRNA synthesis. The cassette also contains the 5’- and 3’-untranslated regions of human ChM and β-globin, respectively, as well as a 100-nucleotide poly(A) tail. Cloning was performed using the CciNI and BamHI restriction endonuclease recognition sites (SibEnzyme, Novosibirsk, Russia). The constructs were named pVAX-C3-H1-24, pVAX-C3-H3-24, and pVAX-C3-HB-24. The DNA sequences of the resulting plasmids were confirmed by restriction analysis and Sanger sequencing.
DNA templates were purified using the HiPure Plasmid Mini Kit (Magen, Guangzhou, China) and linearized using the restriction endonuclease Bso31I (SibEnzyme, Novosibirsk, Russia).
2.3. In Vitro mRNA Synthesis
mRNA synthesis was performed using linearized pVAX-C3-H1-24, pVAX-C3-H3-24, pVAX-C3-HB-24, and pVAX-C3-GFP templates and a commercial kit (Yeasen, Shanghai, China). The reaction mixture included 1 μg of linearized plasmid DNA, T7 polymerase with buffer, AG-Cap analog (m7GmAmG cap analog (Biolabmix, Novosibirsk, Russia) (10 mM), ribonucleotide triphosphate mixture (10 mM) with uridine replaced by N1-methylpseudouridine (Biolabmix, Novosibirsk, Russia), RNase inhibitor (BelBioLab, Moscow, Russia) and nuclease-free water. The synthesis protocol was described by us previously [33], which included enzymatic synthesis of mRNA and its treatment with DNase followed by purification. The obtained mRNAs were named mRNA-H1, mRNA-H3, mRNA-HB, mRNA-GFP.
2.4. PCR to Determine the Presence of Template DNA Residues in the mRNA Preparation
For PCR, primers complementary to the 5’- and 3’-UTR sequences of the mRNA encoded in the template DNA were used. The BioMaster HS-Taq PCR kit (2×) (Biolabmix, Russia), primers, and 1 μg of synthesized mRNA were used. Linearized template DNA was used as a control.
2.5. Capillary Electrophoresis
The purity, homogeneity, and size of the synthetic mRNAs were assessed using an Agilent 2100 BioAnalyser (Agilent Technologies, USA) by microarray electrophoresis with the Agilent RNA 6000 Pico reagent kit (Agilent Technologies, Lithuania) according to the manufacturer’s protocol.
2.6. Encapsulation of mRNA in Lipid NANOPARTICLES
Encapsulation of mRNA in lipid nanoparticles and their characterization were performed as described by Yakovlev et al., 2025 [34]. Phase mixing was performed using an automated NP system (Dolomite Microfluidics, Royston, UK). Mixing was performed in a microfluidic chip of a hex herringbone micromixer (Dolomite Microfluidics, Royston, UK). The aqueous phase consisted of an mRNA solution in 100 mM citrate buffer, pH 4. The ethanol phase consisted of a mixture of ionizable lipids: phospholipids: helper lipids: PEG lipids in a molar percentage ratio of 50:10:38.5:1.5, respectively; all lipids were dissolved in 96% ethanol. The lipid composition included: the ionizable lipid SM-102 (heptadecan-9-yl 8-((2-hydroxyethyl) (6-oxo-6-(undecyloxy)hexyl)amino)octanoate); the phospholipid DSPC (1,2-distearoyl-sn-glycero-3-phosphocholine); the auxiliary lipid cholesterol; and the PEG lipid DMG-PEG2000 (1-monomethoxypolyethyleneglycol-2,3-dimyristylglycerol with polyethyleneglycol with an average molecular weight of 2000).
The resulting particles were characterized using DLS method. The DLS method showed that the mRNA-LNP complexes are monodisperse suspensions of nanoparticles (PdI = 0.158 ± 0.013 for mRNA-H1, 0.178 ± 0.006 for mRNA-H3, and 0.239 ± 0.034 for mRNA-HB). The average hydrodynamic diameter of LNPs was 94.42 ± 2.48 nm, 95.34 ± 0.38 nm, and 97.56 ± 2.44 nm, respectively. The ζ-potential of mRNA-LNP particles in three series of measurements was 0.35 ± 0.12 mV, 0.54 ± 0.42 mV, and 0.05 ± 0.62 mV, respectively, which is consistent with theoretical calculations. The percentage of mRNA encapsulation in LNPs was confirmed using Quant-iT RiboGreen and was greater than 90%.
2.7. mRNA Lyophilization
Lyophilization of mRNA sample solutions was performed in an ED-DF21A-K freeze-drying chamber (ERSTVAK, China) in automatic mode with a pneumatic capping option. Three cryoprotectant variants were used for lyophilization: the first was based on sucrose, the second was based on mannose, and the third was based on trehalose. The sterile mRNA preparation with one of three cryoprotectants was dispensed into 0.5 ml vials of 3 ml and lyophilized for 24 hours at 22±2ºC. The lyophilization process was carried out under standard conditions: freezing samples at -50±2ºC for 8 hours, and lyophilizing samples at 22±2ºC for the remainder of the time. The lyophilized mass was a white tablet. The dried mRNA was stored at temperatures of -20 °C, +4 °C, and +20 °C.
2.8. Immunization of BALB/c Mice
Animal studies were conducted in accordance with the “Guide for the Care and Use of Laboratory Animals.” The protocols were approved by the Laboratory Animal Care and Use Committee of the Federal Research Center of Virology and Biotechnology “Vector” of Rospotrebnadzor (Protocol of the Bioethics Committee No. 3, 2024). Mice were maintained under a 12-hour light cycle with free access to food and water.
The study was conducted on inbred BALB/c mice with an initial weight of 16–18 g. mRNA-Vector-Flu-LNP preparations in 100 μl of PBS were administered into the quadriceps muscle of the left hind paw using insulin syringes with a 29G needle. The mRNA-H1, mRNA-H3, mRNA-HB, and mRNA-Vector-Flu preparations, dissolved in 50 μl of saline, were administered into the left hind paw using the IM method as described previously [27]. The commercial seasonal influenza vaccine Flu-M (FSUE SPbNIIVS FMBA of Russia, Russia, 2025) was administered into the quadriceps muscle of the left hind paw of mice using a syringe.
The experiment was divided into 3 stages. In the first stage, a comparative assessment of the immunogenicity of the developed mRNA vaccine was carried out. Animals were divided into seven groups of 6 individuals each and immunized on days 0 and 21: Group 1 was immunized with 30 μg mRNA-H1; Group 2 was immunized with 30 μg mRNA-H3; Group 3 was immunized with 30 μg mRNA-HB; Group 4 was immunized with the trivalent mRNA vaccine mRNA-Vector-Flu (30 μg of each immunogen, 90 μg in total); Group 5 was immunized with the trivalent mRNA vaccine mRNA-Vector-Flu-LNP (10 μg of each immunogen, 30 μg in total); Group 6 was immunized with 250 μl of the commercial Flu-M vaccine (SPbSRIVS FMBA of Russia, 2025); Group 7 consisted of non-immunized animals. Animals from groups 1, 2, 3 and 4 were immunized using JI, animals from groups 5 and 6 were immunized using a syringe. A dose of 30 μg mRNA was chosen based on the results of our previous studies [27,33] and the results of studies on JI described by other authors [26]. On day 35, blood was collected from the retroorbital sinus. Animals were humanely euthanized by cervical dislocation.
In the second stage, the animals were immunized to analyze the dose-dependent effect of the mRNA vaccine. The animals were divided into 3 groups of 6 animals each and immunized on days 0 and 21: Group 1 was immunized with the trivalent mRNA vaccine mRNA-Vector-Flu, containing all its components (mRNA-H1, mRNA-H3, mRNA-B) at 10 μg of each mRNA (30 μg in total); Group 2 was immunized with mRNA-Vector-Flu, containing 30 μg of each component (90 μg in total); Group 3 was immunized with mRNA-Vector-Flu, containing 50 μg of each component (150 μg in total). Animals were immunized using JI. On day 35, blood was collected from the retro-orbital sinus for humoral immune response analysis.
In the third stage, animals were immunized to analyze the efficacy of the mRNA vaccine lyophilization. The animals were divided into two groups of six animals each and immunized on days 0 and 21. Group 1 was immunized with the lyophilized trivalent mRNA vaccine mRNA-Vector-Flu, containing 15 μg of each component (mRNA-H1, mRNA-H3, mRNA-B) (45 μg total). Group 2 was immunized with mRNA-Vector-Flu, containing 15 μg of each component (45 μg total). Animals were immunized using JI. On day 35, blood was collected from the retro-orbital sinus to assess the humoral response. For T-cell response analysis, animals were humanely sacrificed by cervical dislocation, after which the spleen was collected.
2.9. Enzyme-Linked Immunosorbent Assay (ELISA)
Enzyme-linked immunosorbent assay (ELISA) was performed as previously described [35]. Eukaryotic recombinant hemagglutinin protein H1 (A/Wisconsin/67/2022(H1N1)pdm09), H3 (A/Darwin/9/2021(H3N2)) and HB (B/Austria/1359417/2021), previously obtained in the Bioengineering Department of the State Research Center of Virology and Biotechnology Vector, Rospotrebnadzor, were used as antigens. Rabbit anti-mouse IgG antibodies conjugated with horseradish peroxidase (Sigma-Aldrich, St. Louis, MO, USA) were used as secondary antibodies. TMB substrate (Amresco LLC, USA) was used as a chromogenic substrate. After incubation, the reaction was stopped with stop solution (1 M hydrochloric acid), and the optical density of the solutions was measured at 450 nm using a Varioskan Lux plate reader (Thermo FisherScientific, USA).
2.10. Evaluation of the Cellular Immune Response by ELISpot
Splenocytes were isolated by individually pressing the spleens through nylon cell strainers with 70 and 40 μm pore diameters (BD FalconTM, USA). Red blood cells were removed using red blood cell lysis buffer (Sigma, USA). The intensity of the T-cell immune response in immunized mice was determined by the number of splenocytes producing IFN-γ using the IFN-γ ELISpot assay. The assay was performed using IFN-γ ELISpot kits (MABTECH, Sweden) according to the manufacturer’s instructions. To stimulate cells, a mixture of specific peptides of 10-20 aa in size corresponding to conserved T-cell epitopes of the hemagglutinin proteins of influenza viruses A/Wisconsin/67/2022(H1N1)pdm09, A/Darwin/9/2021(H3N2), and B/Austria/1359417/2021, synthesized at AtaGenix Laboratories (Wuhan, China), was used. The concentration of each peptide was 20 μg/ml. The number of IFNγ-producing cells was counted using an ELISpot reader from Carl Zeiss (Germany).
2.11. Transfection of HEK293 Cells with mRNA.
HEK293 cells were grown in 24-well culture plates (Corning, USA) with DMEM (Sigma-Aldrich, USA) supplemented with 10% FBS (HyClone, USA) and 50 mg/ml gentamicin. A 70–80% confluent cell monolayer was transfected with mRNA encoding the GFP protein using Kit for transfection of nucleic acids into eukaryotic cells using PEI (BIOSPECIFICA, Russia). The transfection agent was mixed with 1 μg of mRNA, incubated for 15 minutes at room temperature, and then added to the cells. The cell plates were then incubated in a CO2 incubator for 24 hours.
2.12. Statistical Analysis
Statistical analysis was performed using GraphPad Prism 10.0 (GraphPad Software, San Diego, CA, USA). Quantitative data are presented as mean values with a range or as a range from lowest to highest and were analyzed using nonparametric tests. Between-group differences were assessed using the nonparametric one-way Kruskal-Wallis analysis of variance with correction for multiple comparisons and Dunn’s hypothesis testing.
3. Results
3.1. Production of the Experimental Trivalent mRNA Influenza Vaccine mRNA-Vector-Flu
Previously, we developed a DNA template cassette for mRNA vaccine synthesis, called pVAX-C3-PolyA [33]. The modified chimeric β-globin sequence used in Moderna’s studies and the human β-globin sequence were included in the cassette as the 5’-UTR and 3’-UTR, respectively. These elements are necessary for increasing mRNA stability and translation efficiency. The initiator nucleotides GG in the T7 promoter were replaced with AG. This modification allows the use of the AG-Cap analog during in vitro transcription to create a “cap” at the 5’-end of mRNA, which is critical for its stability and translation. The plasmid contains a 100-nucleotide poly(A) tail containing an internal linker of 10 random nucleotides (e.g., 30(A)GCATATGACT70(A)). The poly(A) tail is important for mRNA stability and translation initiation. The poly(A) tail sequence ends with a Bso31I restriction endonuclease site. This element ensures that during DNA hydrolysis, the transcription template ends with an adenine, ensuring the precise termination of the synthesized mRNA.
The pVAX-H1-24 plasmid, carrying the hemagglutinin gene of the influenza A/Wisconsin/67/2022 (H1N1)pdm09 virus, was used as the source of the target gene for the trivalent mRNA components of the influenza vaccine. pVAX-H3-24, carrying the hemagglutinin gene of the influenza A/Darvin/9/2021 (H3N2) virus, and pVAX-HB-24, carrying the hemagglutinin gene of the influenza B/Austria/1359417/2021 virus (B/Victoria lineage). In all hemagglutinins, the transmembrane and cytoplasmic domains were removed, and the trimerizing domain of phage T4 was added to enhance immunogenicity (Figure 1a).
After inserting the hemagglutinin gene into the pVAX-C3-PolyA cassette, the pVAX-C3-H1-24, pVAX-C3-H3-24, and pVAX-C3-HB-24 constructs were obtained. These constructs were then used to synthesize the corresponding mRNAs.
3.2. mRNA Synthesis and Characterization
mRNA-C3-H1-24, mRNA-C3-H3-24, and mRNA-C3-HB-24 were synthesized using the method described in Materials and Methods 2.3. The integrity and purity of the mRNA were verified by electrophoresis in a 2% agarose gel (Figure 1b) and capillary electrophoresis (Figure S1). The mRNA transcript size was expected to be approximately 1900 base pairs, consistent with the DNA template. The absence of a high-molecular-weight signal indicates complete removal of the DNA template from the preparation. PCR was also performed for confirmation (Figure 1c).
3.3. Immunogenicity Assessment of the mRNA-Vector-Flu Vaccine
To assess the immunogenicity of the seasonal influenza mRNA vaccine, BALB\c mice (n=6) were immunized with each mRNA individually and in a mixture. A needle-free jet injection method was used for immunization, as described previously [33,34]. The mRNA dose was 30 μg of each immunogen per animal. As a control, one group of animals was immunized with the commercial Flu-M vaccine (SPbSRIVS FMBA of Russia) of the 2024-2025 formulation. Lipid nanoparticles, the gold standard of mRNA delivery, were also used as a control. Animals were immunized with a mixture of 10 μg of each mRNA encapsulated in LNPs, as described previously [25,27,34].
Animal sera were collected on day 28 of the experiment. To set up the ELISA, individual recombinant proteins of hemagglutinin H1, H3 and HB were used as the antigen, as described in section 2.9. The results showed that all nonvariant’s of the trivalent mRNA vaccine against influenza mRNA-Vector-Flu generate an immune response against the corresponding antigen (Figure 2). The average titer for mRNA-H1 was 1: 400950, and for mRNA-H3 1: 510300, mRNA-HB 1: 225000. The commercial vaccine Flu-M showed less pronounced antibody titers, and in the case of hemagglutinin H3 the titer was practically equal to 0. The antibody titer in the group of animals immunized with the trivalent mRNA vaccine in LNP was slightly higher than in the group of animals immunized with jet injection, but the differences were not significant (Figure 2).
3.4. Comparison of the Dose-Dependent Effect of the Trivalent mRNA Vaccine
For a dose-dependent comparison, BALB/c mice were immunized twice with mRNA-Vector-Flu using JI, at doses of 10, 30, and 50 μg of each immunogen. Results showed that at a 10 μg dose of trivalent mRNA, the average titer ranged from 1:100,000 to 1:200,000, depending on the antigen in the ELISA (Figure 3). At a dose of 30 μg of each immunogen, the average titer was approximately 1:500,000. At a dose of 50 μg of each immunogen, we did not observe an increase in the humoral immune response compared to the 30 μg dose. This may be due to the high immunogen dose, which resulted in immunosuppression.
3.5. mRNA Lyophilization
We used three cryoprotectants to stabilize and store naked mRNA after freeze-drying. We initially assessed mRNA integrity after lyophilization using the mRNA-GFP model. mRNA synthesis was performed as described in Section 2.3. Drying was performed as described in Section 2.7. The first cryoprotectant was sucrose-based, the second was mannose-based, and the third was trehalose-based. Samples were stored at 4 °C for one month. The lyophilized mRNA was then dissolved in nuclease-free water and used to transfect HEK293 cells. The results showed that after one month of storage, all preparations demonstrated high efficiency in GFP protein synthesis (Figure 4).
Based on the obtained results, we selected trehalose-based cryoprotectant No. 3 for further work. New mRNA-GFP preparations were prepared, lyophilized, and stored at -20 °C, +4 °C, and +20 °C for long periods. After 1 month and 3 months, lyophilized mRNA-GFP samples were dissolved in pure water and used to transfect HEK293 cells. The results showed that after both 1 month and 3 months of storage, all mRNA samples produced levels of GFP protein synthesis comparable to synthesized immediately before use prepared control mRNA; the differences were not statistically significant (Figure 4b,c). We also analyzed lyophilized mRNA using capillary electrophoresis. The results show that mRNA integrity after drying remains virtually unchanged compared to the control, synthesized immediately before use mRNA. All samples exhibited minor mRNA degradation, less than 5%, which is acceptable (Figure S2).
3.6. Immunogenicity Assessment of the Lyophilized Trivalent mRNA-Vaccine mRNA-Vector-Flu
The next stage of the study was to evaluate the immunogenic properties of the lyophilized trivalent seasonal influenza vaccine mRNA-Vector-Flu. The mRNA was lyophilized with trehalose-based cryoprotectant No. 3 and stored at +4 °C for one month.
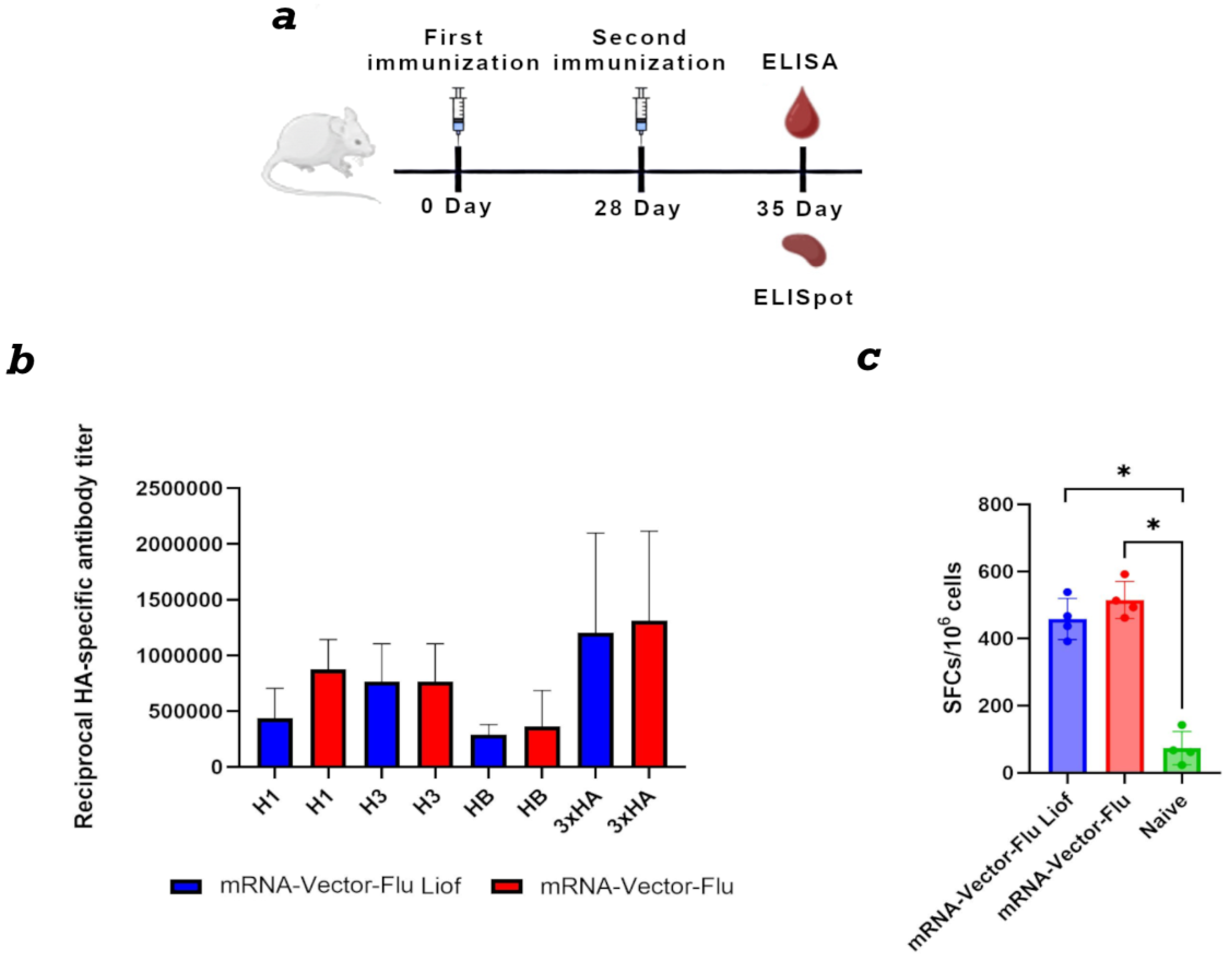
To assess the immunogenicity of the mRNA-Vector-Flu vaccine, BALB\c mice (n=6) were immunized twice with a three-week interval (Figure 5a). As a control, mice were immunized with synthesized immediately before use mRNA vaccine. Immunization was performed using the jet injection method.
Animal sera were collected on day 28 of the experiment. For ELISA, individual recombinant hemagglutinin proteins H1, H3, and HB, as well as their combination, were used as antigens. The results showed that both the synthesized immediately before use prepared and lyophilized mRNA seasonal influenza vaccine mRNA-Vector-Flu produced similar results.
The average titer for the recombinant H1 protein was 1:430,000 for the lyophilized vaccine and 1:870,000 for the synthesized immediately before use mRNA, 1:760,000 for H3, and 1:300,000 for HB. When all three recombinant hemagglutinin proteins were adsorbed in the ELISA, the average titer was 1:1,300,000 for both vaccines (Figure 5b).
For complete protection against viral infections, the development of a T-cell immune response is also necessary. This was assessed using IFN-γ-ELISpot. The results showed that, two weeks after the second immunization, T-cell immunity developed in mice immunized with both lyophilized and synthesized immediately before use mRNA vaccines in response to stimulation with a pool of specific peptides (Figure 5c). The average number of IFNγ-secreting T lymphocytes was 458 per 1 million cells in the group of animals immunized with lyophilized mRNA and 515 per 1 million cells in the group with synthesized immediately before use mRNA.
4. Discussion
mRNA vaccines offer a promising platform for developing new, effective influenza vaccines. Delivery of a naked mRNA vaccine using a needle-free jet injection system further enhances safety, reduces cost, and eliminates the need for lipid nanoparticles, which are traditionally used for mRNA delivery.
In the first phase of our work, we designed and produced a trivalent mRNA influenza vaccine, named mRNA-Vector-Flu, encoding the hemagglutinin (HA) of the seasonal influenza virus strains recommended by the WHO for the 2023-2024 season: A/Wisconsin/67/2022(H1N1)pdm09, A/Darwin/9/2021(H3N2), and B/Austria/1359417/2021. A study of the immunogenicity of the resulting constructs administered to mice using jet injection demonstrated that both the individual and trivalent mRNA-Vector-Flu mRNA influenza vaccines elicited a pronounced specific immune response in BALB/c mice (Figure 2). When immunized with the individual (monovalent) vaccines, each mRNA vaccine induced high titers of specific antibodies (mRNA-H1 – 1:400950, mRNA-H3 – 1:510300, mRNA-HB – 1:225000), as demonstrated by ELISA using recombinant hemagglutinins corresponding to the specific vaccine as antigens. This confirms their ability to induce a strong and specific humoral response.
The control group immunized with the commercial Flu-M vaccine demonstrated significantly lower antibody titers, particularly to hemagglutinin H3, where the titer was virtually zero. This is due to the difference in the composition of the seasonal influenza strains in the vaccines, as the mRNA-Vector-Flu vaccine developed in this study has a composition for the 2023-2024 season, while the Flu-M vaccine used for the control group has a composition for the 2025-2026 season. Only the H3 component changed in the vaccine compositions for these years: in the 2023 season it was A/Darvin/9/2021 (H3N2), and in the 2025 season it was A/District of Columbia/27/2023 (H3N2).
It is worth noting that the trivalent mRNA encapsulated in LNPs induced a slightly higher level of antibodies compared to jet needle-free injection; however, the differences were not statistically significant. This demonstrates that jet injection is a promising mRNA delivery method, comparable in efficacy to conventional LNP formulations.
Thus, the obtained results show that the developed mRNA vaccine mRNA-Vector-Flu elicits a strong specific immune response, and the components of the trivalent vaccine are able to maintain their immunogenicity in combination without suppressing each other’s immune responses.
Analysis of the dose-dependent immune response of the vaccine revealed varying effects of different doses on immune response formation (Figure 3). According to the obtained results, at a vaccine dose of 150 µg (50 µg of each immunogen), antibody titers were approximately equal to those observed at a 30 µg dose (10 µg of each immunogen). It is possible that high doses of mRNA molecules may lead to immunosuppression. A dose of 90 µg (30 µg of each immunogen) showed good results. Thus, the optimal dose for an mRNA vaccine against seasonal influenza is 10–30 µg of each immunogen.
It is known that in aqueous solutions, naked mRNA preparations are relatively unstable; one possible approach to stabilize the formulation is lyophilization. Therefore, the next stage of the study was to determine the feasibility of lyophilization and storage of naked mRNA.
We conducted preliminary studies on the effect of lyophilization on the biological properties of the formulation using an mRNA-GFP model. mRNA-GFP samples lyophilized with three different cryoprotectants, selected based on literature data [36,37], demonstrated comparable efficiency of GFP protein synthesis in transfected cells after one month of storage at +4 °C (Figure 4a). Based on these results, a trehalose-based cryoprotectant was selected for further work. mRNA-GFP lyophilized using trehalose and stored at –20 °C, +4 °C, and +20 °C for three months maintained GFP protein synthesis at a level comparable to synthesized immediately before use control mRNA (Figure 4b,c). The stability of lyophilized mRNA at positive temperatures may, in the future, help address the challenges of storage and transportation of mRNA vaccines.
The third stage of the study focused on evaluating the biological properties of the trivalent mRNA vaccine against seasonal influenza, mRNA-Vector-Flu, lyophilized with trehalose and stored at +4 °C for one month. A synthesized immediately before use mRNA vaccine was used as a control. Immunization with the trivalent mRNA-Vector-Flu, containing 15 µg of each component in 50 µL of saline, was performed via jet injection, and animal sera were collected on day 28 of the experiment.
To quantify the humoral response, an ELISA method was used, using both individual recombinant hemagglutinin proteins H1, H3, and HB, as well as a combination of them, as antigens. The data obtained demonstrated that the lyophilized mRNA vaccine retains a high level of immunogenicity comparable to that of the synthesized immediately before use mRNA vaccine. The average antibody titer against the H1 protein was 1:430,000 for the lyophilized vaccine and 1:870,000 for synthesized immediately before use mRNA. For H3, it reached 1:760,000, and for HB, 1:300,000. When all three proteins were simultaneously adsorbed in an ELISA, the average titer for both vaccines was 1:1,300,000, indicating the formation of a strong specific humoral response against different strains of the seasonal influenza virus. In addition to humoral immunity, the formation of a T-cell response is important for comprehensive protection against viral infection. This was assessed using an IFN-γ ELISpot assay, using a pool of specific peptides to stimulate cells. The results showed that two weeks after the second immunization, animals receiving both lyophilized and synthesized immediately before use mRNA vaccines developed a robust T-cell immune response. The average number of IFN-γ-secreting T cells was 458 per million cells in the lyophilized mRNA group and 515 per million cells in group of animals immunized with synthesized immediately before use the mRNA.
Thus, the lyophilized mRNA-Vector-Flu mRNA vaccine, stored at 4 °C for a month, demonstrates the ability to induce both a strong humoral and cellular immune response comparable to the effect of the synthesized immediately before use prepared mRNA vaccine. These data confirm the potential of the lyophilized vaccine for further use and extended shelf life without loss of immunogenic activity.
The next step in our research is to test the biological activity of lyophilized naked mRNA-Vector-Flu stored at 4 °C and 20 °C for a year.
Combining lyophilization of the naked mRNA vaccine with our developed method of delivering mRNA via jet injection solves the main problem of prophylactic mRNA vaccine platforms: delivery. Using the gold standard of delivery, lipid nanoparticles (LNPs), significantly increases the cost of vaccine production; LNPs can cause undesirable side effects; mRNA-LNPs must be stored at extremely low temperatures or a complex cryoprotectant composition must be used for their lyophilization [38].
5. Conclusions
In this study, we produced and characterized a trivalent mRNA vaccine against the seasonal influenza virus, mRNA-Vector-Flu. Delivery of the naked vaccine via jet injection was shown to elicit a high immune response in BALB/c mice. Lyophilization with a trehalose-based cryoprotectant was used for stabilization and storage. Using the mRNA-GFP model, we demonstrated that lyophilized mRNA remained stable during storage for at least 3 months at +4 °C and +20 °C. Notably, the biological activity of lyophilized mRNA-GFP was comparable to that of synthesized immediately before use prepared mRNA. A comparative study of the immunogenicity of naked mRNA-Vector-Flu showed that both the synthesized immediately before use the mRNA preparation and the lyophilized form stored at +4 °C for a month induced similar levels of virus-specific antibodies and generated a pronounced T-cell immune response.
Thus, delivery of a naked mRNA vaccine using a needle-free jet injection ensures a high-level immune response, which improves its safety, reduces its cost, and eliminates the need for lipid nanoparticles traditionally used for mRNA delivery. At the same time, lyophilization of the naked mRNA vaccine preserves its biological activity and ensures long-term storage at elevated temperatures. Our results demonstrate that our proposed approach can be considered a promising direction for the development and improvement of the mRNA vaccine platform.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Electropherogram of synthesized mRNA-H1, mRNA-H3 and mRNA-HB obtained by capillary electrophoresis; Figure S2: Electropherogram of lyophilized mRNA-GFP stored for 3 months, obtained by capillary electrophoresis; Figure S3. Evaluation of the efficiency of GFP protein synthesis in HEK293 cells transfected with lyophilized mRNA-GFP.
Author Contributions
Conceptualization, S.V.S., A.P.R., A.A.I. and L.I.K.; methodology, S.V.S., S.I.K., D.N.K., M.B.B., V.A.Y., E.V.T., E.V.S., V.R.L., L.A.K., D.I.V., K.P.M., E.A.V., K.I.I., A.A.B. and N.B.R.; validation, A.P.R., L.I.K. and A.A.I.; investigation, S.V.S., S.I.K., D.N.K., M.B.B., V.A.Y., E.V.T., E.V.S., V.R.L., L.A.K., D.I.V., K.P.M., E.A.V., K.I.I., A.A.B. and N.B.R.; resources, S.V.S., E.A.V.,V.A.Y. and S.I.K.; visualization, S.V.S.; writing—original draft preparation, S.V.S. and A.A.I.; writing—review and editing, A.P.R., A.A.I. and L.I.K; supervision, A.A.I. and L.I.K. All authors have read and agreed to the published version of the manuscript.
Funding
The research was conducted in accordance with the terms of the State Assignment of SRC Vector Rospotrebnadzor No. 25/21.
Institutional Review Board Statement
The study was conducted according to the Declaration of Helsinki and approved by the Bioethics Committee of SRC VB Vector (No. 3 Protocol of the Bioethics Committee of 29 February 2024).
Data Availability Statement
The data can be shared upon request.
Conflicts of Interest
The authors declare that they have no competing interests.
Abbreviations
The following abbreviations are used in this manuscript:
| mRNA | Messenger RNA |
| LNPs | Lipid nanoparticles |
| JI | Needle-free jet injection |
| WHO | World Health Organization |
| HA | Hemagglutinin |
| DNA | Deoxyribonucleic acid |
| ELISA | Enzyme-linked immunosorbent assay |
References
- WHO, Influenza (Seasonal). Available online: https://www.who.int/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 30 September 2025).
- Paget, J. et al. Global and national influenza-associated hospitalisation rates: estimates for 40 countries and administrative regions. J. Glob. Health 13, 04003 (2023).
- Iuliano A.D., Roguski K.M., Chang H.H., Muscatello D.J., Palekar R., Tempia S., et al. Estimates of global seasonal influenza-associated respiratory mortality: a modelling study. Lancet. 2018;391(10127):1285-300.
- Martins, J.P.; Santos, M.; Martins, A.; Felgueiras, M.; Santos, R. Seasonal Influenza Vaccine Effectiveness in Persons Aged 15–64 Years: A Systematic Review and Meta-Analysis. Vaccines 2023, 11, 1322. [CrossRef]
- World Health Organization. Vaccines against influenza: WHO position paper - May 2022. Wkly. Epidemiol. Rec. 87, 185–208 (2022).
- Chan L, Alizadeh K, Alizadeh K, Fazel F, Kakish JE, Karimi N, Knapp JP, Mehrani Y, Minott JA, Morovati S, Rghei A, Stegelmeier AA, Vanderkamp S, Karimi K, Bridle BW. Review of Influenza Virus Vaccines: The Qualitative Nature of Immune Responses to Infection and Vaccination Is a Critical Consideration. Vaccines (Basel). 2021 Sep 1;9(9):979. [CrossRef]
- Mokalla, V.R.; Gundarapu, S.; Kaushik, R.S.; Rajput, M.; Tummala, H. Influenza Vaccines: Current Status, Adjuvant Strategies, and Efficacy. Vaccines 2025, 13, 962. [CrossRef]
- Moore, K. A. et al. A research and development (R&D) roadmap for influenza vaccines: looking toward the future. Vaccine 39, 6573–6584 (2021).
- Scorza F.B., Pardi N. New kids on the block: RNA-based influenza virus vaccines // Vaccines. MDPI AG, 2018. Vol. 6, № 2.
- Walsh E.E., Frenck R.W. Jr., Falsey A.R., Kitchin N., Absalon J., Gurtman A., Lockhart S., Neuzil K., Mulligan M.J., Bailey R., Swanson K.A., Li P., Koury K., Kalina W., Cooper D., Fontes-Garfias C., Shi P.Y., Türeci Ö., Tompkins K.R., Lyke K.E., Raabe V., Dormitzer P.R., Jansen K.U., Şahin U., Gruber W.C. (2020) Safety and Immunogenicity of Two RNA-Based Covid-9 Vaccine Candidates. N Engl J Med. 383(25), 2439-2450. [CrossRef]
- Teo S.P. (2022) Review of COVID-19 mRNA Vaccines: BNT162b2 and mRNA-1273. J Pharm Pract. 35(6), 947-951. [CrossRef]
- Polack F.P., Thomas S.J., Kitchin N., Absalon J., Gurtman A., Lockhart S., et al. Safety and efficacy of the BNT162b2 mRNA Covid-19 vaccine. N Engl J Med. 2020;383(27):2603-15.
- Baden L.R., El Sahly H.M., Essink B., Kotloff K., Frey S., Novak R., et al. Efficacy and safety of the mRNA-1273 SARS-CoV-2 vaccine. N Engl J Med. 2021;384(5):403-16.
- Mazunina E.P., Gushchin V.A., Kleymenov D.A., Siniavin A.E., Burtseva E.I., Shmarov M.M., Mukasheva E.A., Bykonia E.N., Kozlova S.R., Evgrafova E.A., Zolotar A.N., Shidlovskaya E.V., Kirillova E.S., Krepkaia A.S., Usachev E.V., Kuznetsova N.A., Ivanov I.A., Dmitriev S.E., Ivanov R.A., Logunov D.Y., Gintsburg A.L. (2024) Trivalent mRNA vaccine-candidate against seasonal flu with cross-specific humoral immune response. Front Immunol. 15, 1381508. [CrossRef]
- Joe P.T., Christopoulou I., van Hoecke L., Schepens B., Ysenbaert T., Heirman C., Thielemans K., Saelens X., Aerts J.L. (2019) Intranodal administration of mRNA encoding nucleoprotein provides cross-strain immunity against influenza in mice. J Transl Med. 17(1), 242. [CrossRef]
- Zhuang X., Qi Y., Wang M., Yu N., Nan F., Zhang H., Tian M., Li C., Lu H., Jin N. (2020) mRNA Vaccines Encoding the HA Protein of Influenza A H1N1 Virus Delivered by Cationic Lipid Nanoparticles Induce Protective Immune Responses in Mice. Vaccines (Basel). 8(1), 123. [CrossRef]
- Lee I.T., Nachbagauer R., Ensz D., Gvozdjak K., Berlanda-Scorza F., Bahl J., et al. Safety and immunogenicity of a phase 1/2 randomized clinical trial of a quadrivalent, mRNA-based seasonal influenza vaccine (mRNA-1010) in healthy adults: interim analysis. Nat Commun. 2023; 14:3631.
- Rudman Spergel, A.K., Lee, I.T., Koslovsky, K. et al. Immunogenicity and safety of mRNA-based seasonal influenza vaccines encoding hemagglutinin and neuraminidase. Nat Commun 16, 5933 (2025). [CrossRef]
- Kandinov B, Soens M, Huang W, Llapur C, Ensz D, Essink B, Fierro C, Vakil J, Pucci A, Guo J, Rudden S, Hall K, Hicks B, Schaefers K, Zhou H, Ma C, Zheng L, Avanesov A, Park Y, Du E, Miller J, Ananworanich J, Nachbagauer R. An mRNA-based seasonal influenza vaccine in adults: Results of two phase 3 randomized clinical trials and correlate of protection analysis of hemagglutination inhibition titers. Hum Vaccin Immunother. 2025 Dec;21(1):2484088. [CrossRef]
- Soens M, Ananworanich J, Hicks B, Lucas KJ, Cardona J, Sher L, Livermore G, Schaefers K, Henry C, Choi A, Avanesov A, Chen R, Du E, Pucci A, Das R, Miller J, Nachbagauer R. A phase 3 randomized safety and immunogenicity trial of mRNA-1010 seasonal influenza vaccine in adults. Vaccine. 2025 Mar 19;50:126847. [CrossRef]
- Cheng, F.; Wang, Y.; Bai, Y.; Liang, Z.; Mao, Q.; Liu, D.; Wu, X.; Xu, M. Research Advances on the Stability of mRNA Vaccines. Viruses 2023, 15, 668. [CrossRef]
- Faksova, K.; Walsh, D.; Jiang, Y.; Griffin, J.; Phillips, A.; Gentile, A.; Kwong, J.C.; Macartney, K.; Naus, M.; Grange, Z.; et al. COVID-19 Vaccines and Adverse Events of Special Interest: A Multinational Global Vaccine Data Network (GVDN) Cohort Study of 99 Million Vaccinated Individuals. Vaccine 2024, 42. [CrossRef]
- Oster M.E., Shay D.K., Su J.R., Gee J., Creech C.B., Broder K.R., et al. Myocarditis Cases Reported After mRNA-Based COVID-19 Vaccination in the US From December 2020 to August 2021. JAMA. 2022;327(4):331-40.
- Mao S., Li S., Zhang Y., Long L., Peng J., Cao Y., et al. A Highly Efficient Needle-Free-Injection Delivery System for MRNA-LNP Vaccination against SARS-CoV-2. Nano Today. 2023; 48:101730.
- Kisakov D.N., Kisakova L.A., Sharabrin S.V., Yakovlev V.A., Tigeeva E.V., Borgoyakova M.B., et al. Delivery of Experimental MRNA Vaccine, Encoding the RBD of SARS-CoV-2 Using Jet Injection. Bull Exp Biol Med. 2023;176(6):751-6.
- Abbasi S., Matsui-Masai M., Yasui F., Hayashi A., Tockary T.A., Mochida Y., et al. Carrier-Free MRNA Vaccine Induces Robust Immunity against SARS-CoV-2 in Mice and Non-Human Primates without Systemic Reactogenicity. Mol Ther. 2024;32(5):1266-83.
- Kisakov D.N., Karpenko L.I., Kisakova L.A., Sharabrin S.V., Borgoyakova M.B., Starostina E.V., et al. Jet Injection of Naked mRNA Encoding the RBD of the SARS-CoV-2 Spike Protein Induces a High Level of a Specific Immune Response in Mice. Vaccines. 2025;13(1):65.
- Wang R., Bian Q., Xu Y., Xu D., Gao J. Recent Advances in Mechanical Force-Assisted Transdermal Delivery of Macromolecular Drugs. Int J Pharm. 2021; 602:120598.
- The World Health Organization (WHO). Recommendations announced for influenza vaccine composition for the 2023-2024 northern hemisphere influenza season. [Internet]. 2023 [cited 2025 October 28]. Available from: https://www.who.int/news/item/24-02-2023-recommendations-announced-for-influenza-vaccine-composition-for-the-2023-2024-northern-hemisphere-influenza-season.
- Lu Y., Welsh J.P., Swartz J.R. Production and stabilization of the trimeric influenza hemagglutinin stem domain for potentially broadly protective influenza vaccines. Proc Natl Acad Sci U S A. 2014;111(1):125-30.
- Ecker J.W., Kirchenbaum G.A., Pierce S.R., Skarlupka A.L., Abreu R.B., Cooper R.E., et al. High-Yield Expression and Purification of Recombinant Influenza Virus Proteins from Stably-Transfected Mammalian Cell Lines. Vaccines 2020;8(3):462.
- Milder F.J., Jongeneelen M., Ritschel T., Bouchier P., Bisschop I.J.M., de Man, M., et al. Universal stabilization of the influenza hemagglutinin by structure-based redesign of the pH switch regions. Proc Natl Acad Sci U S A. 2022;119(15): e2115379119.
- Sharabrin S.V., Ilyichev A.A., Kisakov D.N., et al. Development of a Needle-Free Method for Delivery of mRNA Vaccines Using a Jet Injector. Mol Biol. 2025;59(3):426-40.
- Yakovlev V.A., Litvinova V.R., Rudometova N.B., et al. Immunogenic and Protective Properties of mRNA Vaccine Encoding Hemagglutinin of Avian Influenza A/H5N8 Virus, Delivered by Lipid Nanoparticles and Needle-Free Jet Injection. Vaccines 2025;13(8):883.
- Karpenko, L.I.; Rudometov, A.P.; Sharabrin, S.V.; Shcherbakov, D.N.; Borgoyakova, M.B.; Bazhan, S.I.; Volosnikova, E.A.; Rudometova, N.B.; Orlova, L.A.; Pyshnaya, I.A.; et al. Delivery of mRNA Vaccine against SARS-CoV-2 Using a Polyglucin:Spermidine Conjugate. Vaccines 2021, 9, 76.
- Li, M., Jia, L., Xie, Y. et al. Lyophilization process optimization and molecular dynamics simulation of mRNA-LNPs for SARS-CoV-2 vaccine. npj Vaccines 8, 153 (2023). [CrossRef]
- Muramatsu H, Lam K, Bajusz C, Laczkó D, Karikó K, Schreiner P, Martin A, Lutwyche P, Heyes J, Pardi N. Lyophilization provides long-term stability for a lipid nanoparticle-formulated, nucleoside-modified mRNA vaccine. Mol Ther. 2022 May 4;30(5):1941-1951. [CrossRef]
- Khan MDFH, Baudin F, Sudalaiyadum Perumal A, Kamen AA. Freeze-Drying of mRNA-LNPs Vaccines: A Review. Vaccines (Basel). 2025 Aug 12;13(8):853. [CrossRef]
Figure 1.
(а) Schematic representation of mRNA: mRNA has a 5’-cap, poly(A) tail, 5’- and 3’-untranslated regions of ChM and human β-globin, an open reading frame encoding hemagglutinin (HA) of influenza H1N1, H3N2 and Victoria B; Schematic structure of HA genes modified by the trimerizing domain of phage T4 (T4 TD); (b) Electropherogram of mRNA synthesis products in 1% agarose gel: lane 1 - ssRNA Ladder (New England Biolabs); 2 - mRNA-H1, 3 - mRNA -H3 and 3 - mRNA-H3; (c) Electropherogram of PCR products from the mRNA preparation for the presence of a DNA template. lanes 1 - DNA marker, 2 - linearized DNA template pVAX-C3-H1-24, 2 - linearized DNA template pVAX-C3-H3-24, 3 - linearized DNA template pVAX-C3-HB-24, 4 - mRNA-H1, 5 - mRNA-H3 and 6 - mRNA-H3;.
Figure 1.
(а) Schematic representation of mRNA: mRNA has a 5’-cap, poly(A) tail, 5’- and 3’-untranslated regions of ChM and human β-globin, an open reading frame encoding hemagglutinin (HA) of influenza H1N1, H3N2 and Victoria B; Schematic structure of HA genes modified by the trimerizing domain of phage T4 (T4 TD); (b) Electropherogram of mRNA synthesis products in 1% agarose gel: lane 1 - ssRNA Ladder (New England Biolabs); 2 - mRNA-H1, 3 - mRNA -H3 and 3 - mRNA-H3; (c) Electropherogram of PCR products from the mRNA preparation for the presence of a DNA template. lanes 1 - DNA marker, 2 - linearized DNA template pVAX-C3-H1-24, 2 - linearized DNA template pVAX-C3-H3-24, 3 - linearized DNA template pVAX-C3-HB-24, 4 - mRNA-H1, 5 - mRNA-H3 and 6 - mRNA-H3;.
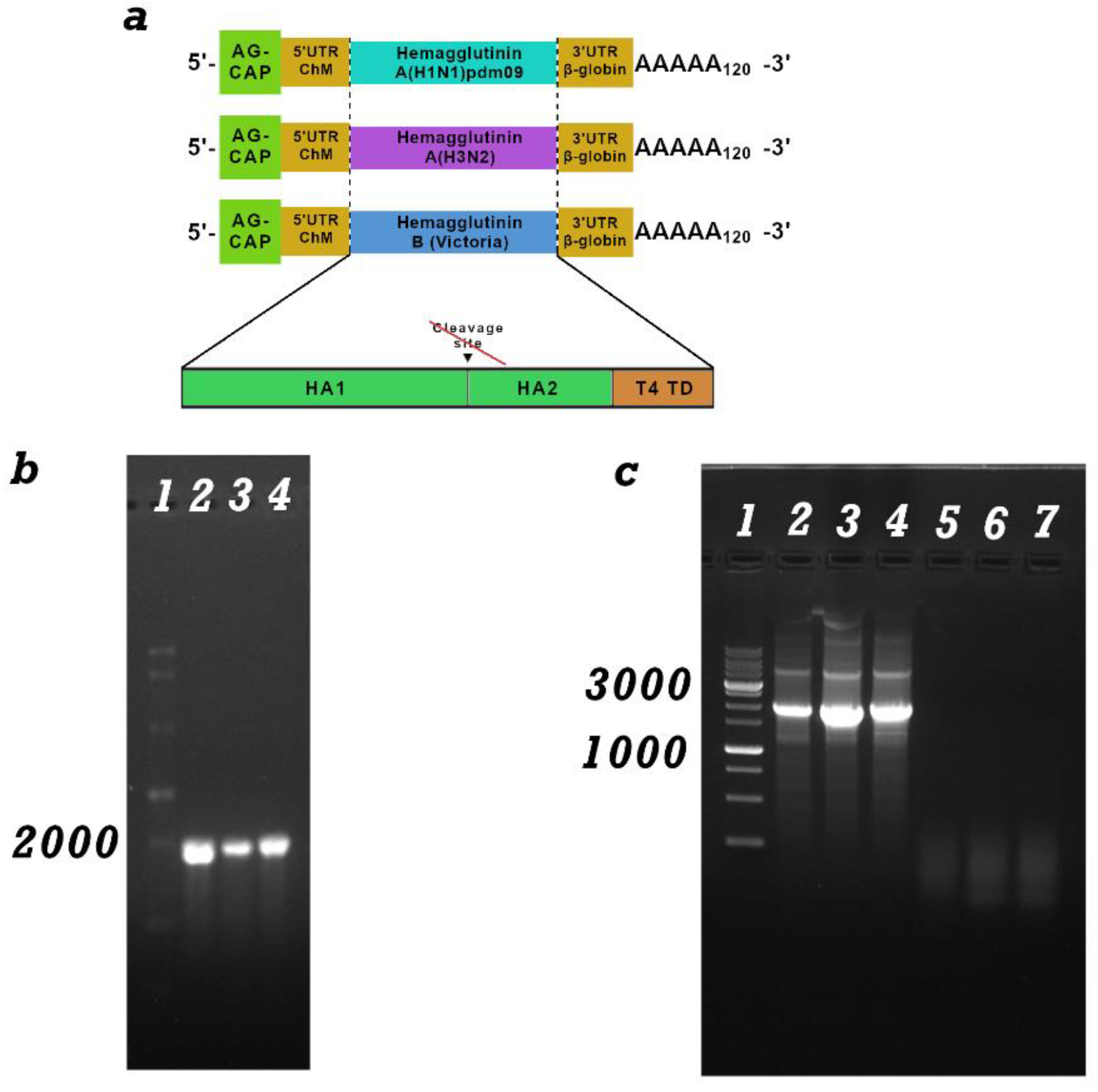
Figure 2.
Study of the humoral immune response to components of the trivalent mRNA vaccine against the seasonal influenza virus. (a) Immunization schedule. (b) H1-specific antibody titer (ELISA data) (number of animals: n= 16). (c) H3-specific antibody titer (ELISA data) (number of animals: n= 6). (d) HB-specific antibody titer (ELISA data) (number of animals: n= 6). Reciprocal titer values are provided in the plots. mRNA-H1 — a group of animals immunized with mRNA-H1; mRNA-H3 — a group of animals immunized with mRNA-H3; mRNA-HB — a group of animals immunized with mRNA-HB; mRNA-Vector-Flu JI — a group of animals immunized with a trivalent mRNA vaccine delivered by JI; mRNA-Vector-Flu LNP — a group of animals immunized with a trivalent mRNA vaccine delivered by LNP; Flu-M — a group of animals immunized with the commercial vaccine Flu-M; Naive—a group of non-immunized animals. In panels, data are provided as means with full range. * p < 0.05, ** p < 0.01, *** p < 0.001 following Kruskal–Wallis analysis of variance with correction for multiple comparisons and Dunn’s statistical hypothesis test.
Figure 2.
Study of the humoral immune response to components of the trivalent mRNA vaccine against the seasonal influenza virus. (a) Immunization schedule. (b) H1-specific antibody titer (ELISA data) (number of animals: n= 16). (c) H3-specific antibody titer (ELISA data) (number of animals: n= 6). (d) HB-specific antibody titer (ELISA data) (number of animals: n= 6). Reciprocal titer values are provided in the plots. mRNA-H1 — a group of animals immunized with mRNA-H1; mRNA-H3 — a group of animals immunized with mRNA-H3; mRNA-HB — a group of animals immunized with mRNA-HB; mRNA-Vector-Flu JI — a group of animals immunized with a trivalent mRNA vaccine delivered by JI; mRNA-Vector-Flu LNP — a group of animals immunized with a trivalent mRNA vaccine delivered by LNP; Flu-M — a group of animals immunized with the commercial vaccine Flu-M; Naive—a group of non-immunized animals. In panels, data are provided as means with full range. * p < 0.05, ** p < 0.01, *** p < 0.001 following Kruskal–Wallis analysis of variance with correction for multiple comparisons and Dunn’s statistical hypothesis test.
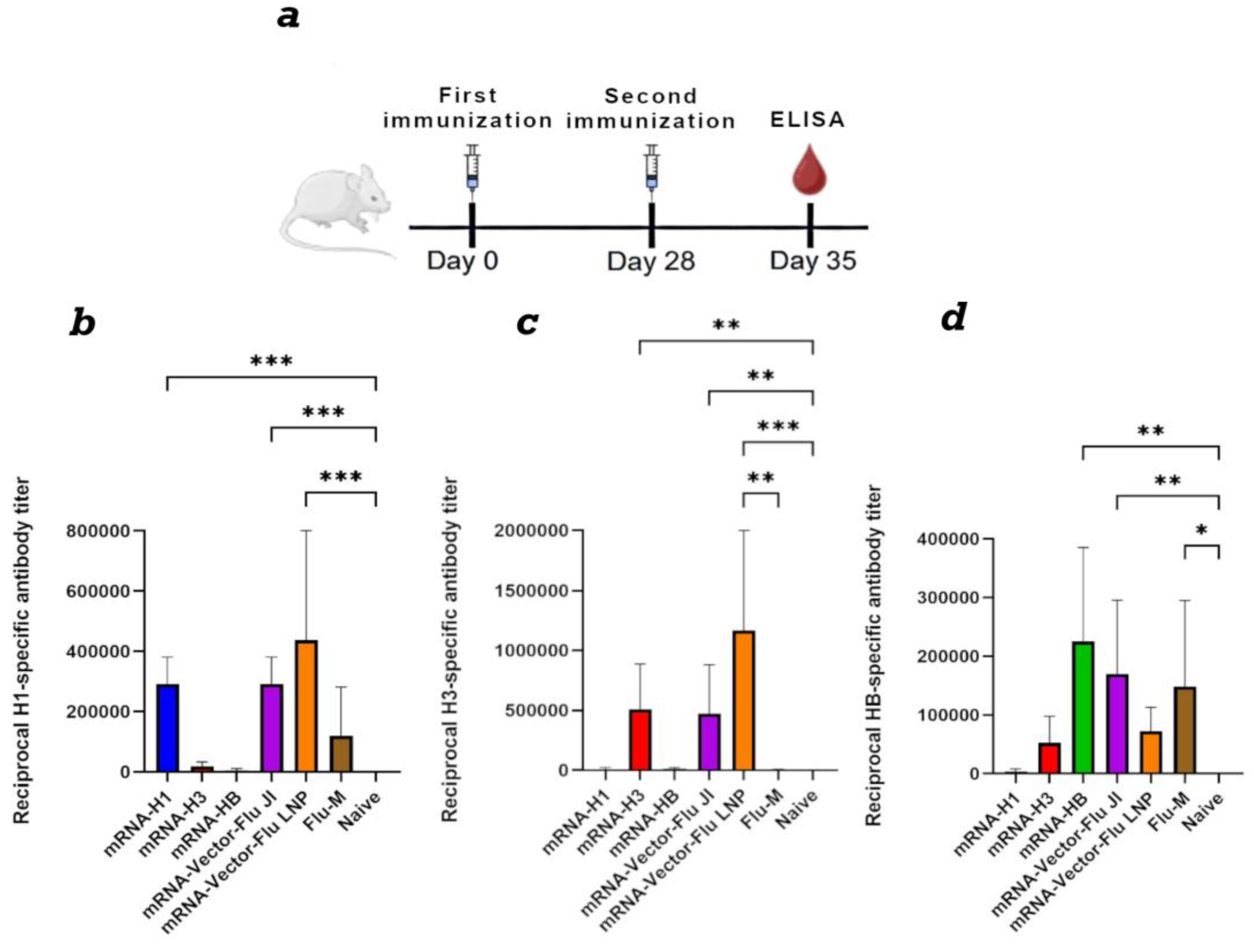
Figure 3.
Study of the humoral immune response of a trivalent mRNA vaccine against the seasonal influenza virus depending on the dose. (a) Immunization schedule. (b) H1-specific antibody titer (ELISA data) (number of animals: n= 16). (c) H3-specific antibody titer (ELISA data) (number of animals: n= 6). (d) HB-specific antibody titer (ELISA data) (number of animals: n= 6). Reciprocal titer values are provided in the plots. mRNA-Vector-Flu JI, 10 ug — a group of animals immunized with a trivalent mRNA vaccine, the dose of each component was 10 ug, a total of 30 ug; mRNA-Vector-Flu JI, 30 ug — a group of animals immunized with a trivalent mRNA vaccine, the dose of each component was 30 ug, a total of 90 ug; mRNA-Vector-Flu JI, 50 ug — a group of animals immunized with a trivalent mRNA vaccine, the dose of each component was 50 ug, a total of 150 ug. In panels, data are provided as means with full range. * p < 0.05 following Kruskal–Wallis analysis of variance with correction for multiple comparisons and Dunn’s statistical hypothesis test.
Figure 3.
Study of the humoral immune response of a trivalent mRNA vaccine against the seasonal influenza virus depending on the dose. (a) Immunization schedule. (b) H1-specific antibody titer (ELISA data) (number of animals: n= 16). (c) H3-specific antibody titer (ELISA data) (number of animals: n= 6). (d) HB-specific antibody titer (ELISA data) (number of animals: n= 6). Reciprocal titer values are provided in the plots. mRNA-Vector-Flu JI, 10 ug — a group of animals immunized with a trivalent mRNA vaccine, the dose of each component was 10 ug, a total of 30 ug; mRNA-Vector-Flu JI, 30 ug — a group of animals immunized with a trivalent mRNA vaccine, the dose of each component was 30 ug, a total of 90 ug; mRNA-Vector-Flu JI, 50 ug — a group of animals immunized with a trivalent mRNA vaccine, the dose of each component was 50 ug, a total of 150 ug. In panels, data are provided as means with full range. * p < 0.05 following Kruskal–Wallis analysis of variance with correction for multiple comparisons and Dunn’s statistical hypothesis test.
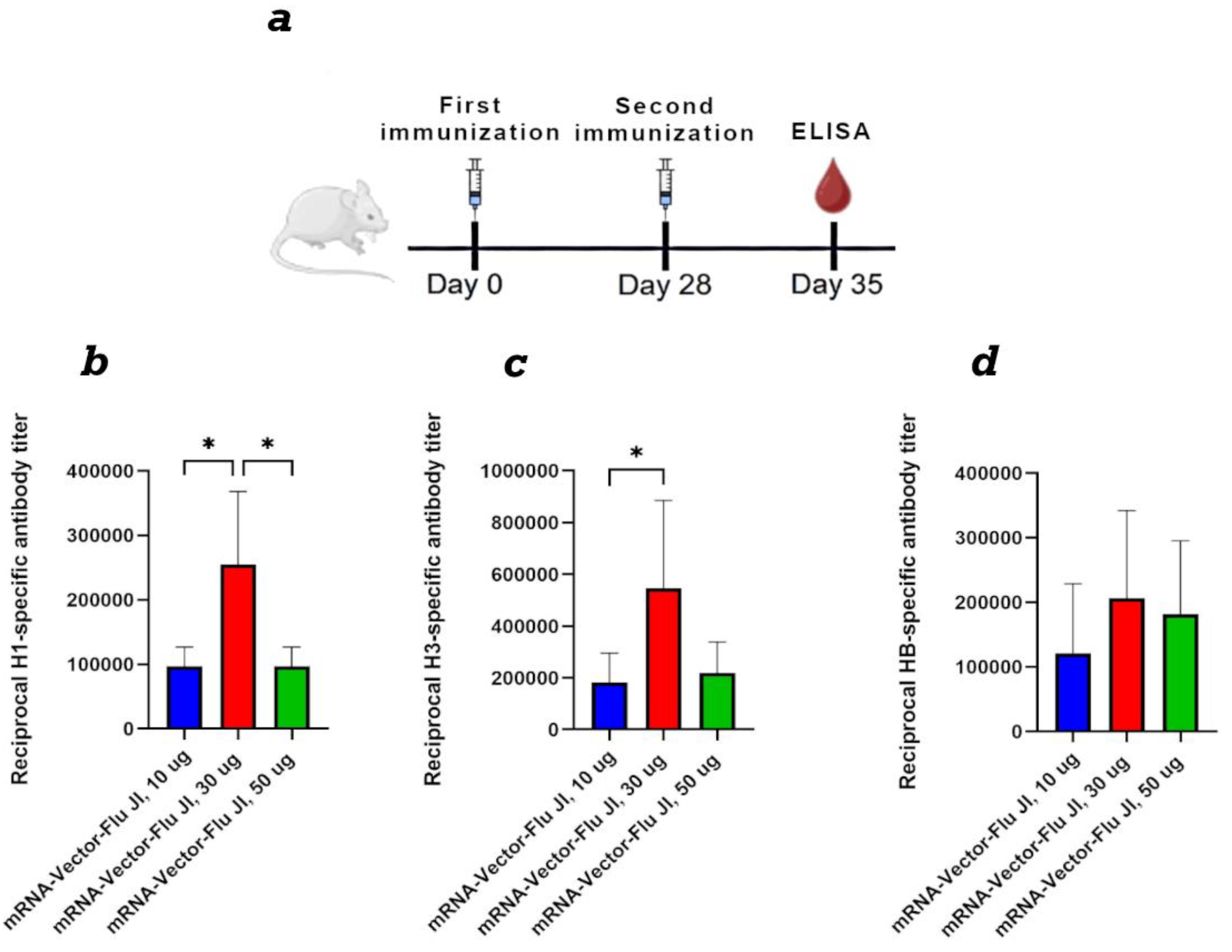
Figure 4.
Evaluation of mRNA stability after lyophilization. (a) Synthesis of GFP protein in HEK293 cells transfected with mRNA-GFP after lyophilization in different cryoprotectants. (b) The level of GFP protein synthesis in HEK293 cells transfected with lyophilized mRNA-GFP after storage for one month. (c) The level of GFP protein synthesis in HEK293 cells transfected with lyophilized mRNA-GFP after storage for three months. Data are shown as a comparison with the control synthesized immediately before use mRNA-GFP, where the control is 0. +4 °C – lyophilized mRNA-GFP stored at +4 °C; +20 °C – lyophilized mRNA-GFP stored at +20 °C; -20 °C – lyophilized mRNA-GFP stored at -20 °C.
Figure 4.
Evaluation of mRNA stability after lyophilization. (a) Synthesis of GFP protein in HEK293 cells transfected with mRNA-GFP after lyophilization in different cryoprotectants. (b) The level of GFP protein synthesis in HEK293 cells transfected with lyophilized mRNA-GFP after storage for one month. (c) The level of GFP protein synthesis in HEK293 cells transfected with lyophilized mRNA-GFP after storage for three months. Data are shown as a comparison with the control synthesized immediately before use mRNA-GFP, where the control is 0. +4 °C – lyophilized mRNA-GFP stored at +4 °C; +20 °C – lyophilized mRNA-GFP stored at +20 °C; -20 °C – lyophilized mRNA-GFP stored at -20 °C.
Figure 5.
Comparison of the immunogenicity of lyophilized and synthesized immediately before use mRNA vaccine mRNA-Vector-Flu. (a) Immunization schedule. (b) Study of the humoral immune response of the lyophilized mRNA vaccine mRNA-Vector-Flu. H1 - the recombinant protein H1 was used as the antigen in the ELISA; H3 - the recombinant protein H3 was used as the antigen in the ELISA; HB - the recombinant protein HB was used as the antigen in the ELISA; 3xHA - a mixture of recombinant HA proteins was used as the antigen in the ELISA. Lyophilized mRNA-Vector-Flu is shown in blue, synthesized immediately before use mRNA-Vector-Flu is shown in red. (c) ELISpot assay results of specific T-cell responses in immunized BALB/c mice. Number of cells expressing IFN-γ in response to stimulation with a pool of HA-specific peptides per 1 × 106 splenocytes. Data are presented as the median with a range of inverse titers. Significance was assessed using non-parametric one-factor Kruskal–Wallis analysis of variance (* p < 0.05). mRNA-Vector-Flu Liof - mice immunized with lyophilized mRNA; mRNA-Vector-Flu - mice immunized with synthesized immediately before use mRNA.
Figure 5.
Comparison of the immunogenicity of lyophilized and synthesized immediately before use mRNA vaccine mRNA-Vector-Flu. (a) Immunization schedule. (b) Study of the humoral immune response of the lyophilized mRNA vaccine mRNA-Vector-Flu. H1 - the recombinant protein H1 was used as the antigen in the ELISA; H3 - the recombinant protein H3 was used as the antigen in the ELISA; HB - the recombinant protein HB was used as the antigen in the ELISA; 3xHA - a mixture of recombinant HA proteins was used as the antigen in the ELISA. Lyophilized mRNA-Vector-Flu is shown in blue, synthesized immediately before use mRNA-Vector-Flu is shown in red. (c) ELISpot assay results of specific T-cell responses in immunized BALB/c mice. Number of cells expressing IFN-γ in response to stimulation with a pool of HA-specific peptides per 1 × 106 splenocytes. Data are presented as the median with a range of inverse titers. Significance was assessed using non-parametric one-factor Kruskal–Wallis analysis of variance (* p < 0.05). mRNA-Vector-Flu Liof - mice immunized with lyophilized mRNA; mRNA-Vector-Flu - mice immunized with synthesized immediately before use mRNA.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



