
Submitted:
14 November 2025
Posted:
19 November 2025
You are already at the latest version
Abstract
The effects of 4-phenylbutyrate (4-PBA) on particle-induced macrophage apoptosis in vitro have not been previously documented. This study aims to investigate the thera-peutic potential of 4-PBA in alleviating titanium nanoparticle (TiNP)-induced in-flammatory injury mediated by macrophages. Our findings demonstrate that TiNPs exhibit specific properties capable of inciting inflammatory injury to macrophages. Furthermore, the extent of inflammatory injury inflicted by TiNPs on macrophages progressively increases in correlation with both the concentration of particles and du-ration of exposure. Exposure to TiNPs significantly reduces macrophage viability, promotes apoptosis, enhances the expression of inflammatory cytokines, and exacer-bates the production of reactive oxygen species (ROS). Co-treatment with 4-PBA sig-nificantly alleviates the severity of particle-induced inflammatory injury while simul-taneously facilitating apoptosis in stressed macrophages. Pharmacological intervention with 4-PBA facilitates TiNP-induced macrophage apoptosis by increasing the expres-sion of pro-apoptotic proteins such as Caspase-3 and Bax, while decreasing the levels of anti-apoptotic protein Bcl-2. When used as an independent treatment, 4-PBA does not provoke a pronounced inflammatory response nor induce significant changes in mac-rophage apoptosis, underscoring its potential as a safe therapeutic option. These find-ings substantiate the hypothesis that employing a specific approach to facilitate mac-rophage apoptosis could serve as an effective therapeutic strategy for managing aseptic loosening. Significantly, this approach demonstrates a favorable safety profile, posi-tioning it as a promising avenue for future clinical applications in this domain.
Keywords:
macrophage
; aseptic loosening
; osteolysis
; wear particles
; 4-phenylbutyrate
; apoptosis
; inflammation
1. Introduction
Metal alloys have been utilized for over five decades as a substitute for joint segments in prosthetic applications. The prevalence of TiAl6V4 alloy implants has steadily increased worldwide, with the durability and lifespan of a prosthetic joint being determined by the wear rate of the materials involved in its moving components [1]. Wear particles generated from artificial implants during movement, which subsequently disperse into the surrounding periprosthetic area and synovial fluid [2]. The continuous release of particle from various implant materials is recognized to eventually elicit an inflammatory response in the peri-prosthetic bone [3]. This phenomenon is of significant concern in the field of orthopedics, as the presence of wear particles can initiate a cascade of biological reactions that may compromise the integrity of the surrounding bone tissue over time [4]. Aseptic osteolysis is believed to stem from the cellular response of the prosthetic microenvironment to these wear particles, and is frequently viewed as the detrimental consequence of artificial implants that determines their longevity and functionality [4].
TiAl6V4 prostheses are recognized for their excellent biocompatibility; however, the byproducts associated with titanium, particularly TiAl6V4 nanoparticles (TiNPs), are far from being bio-inert. Despite achieving high success rates during the initial 10 years, issues such as periprosthetic loosening and orthopedic peri-implantitis have emerged as significant contributors to implant failure, posing substantial health concerns for both clinicians and the medical community. The reported risk of periprosthetic osteolysis occurring after 10 years ranges from 20% to 56% [5]. Unfortunately, there are currently no widely accepted or standardized treatment protocols for addressing these complications, leading to persistently high rates of recurrence and revision among affected patients. Given these challenges, extending the lifespan of metal prostheses has become a critical objective within orthopedic rehabilitation. Therefore, it is essential for researchers to explore new and innovative approaches that could serve as enhancements to, or replacements for, traditional revision surgeries. By doing so, they can significantly improve patient outcomes and increase the durability of prosthetic devices, ultimately fostering a better quality of life for individuals reliant on these implants.
Considerable evidence exists indicating that the process of osteolysis is characterized by the overactivity of various cell types, including neutrophils, monocytes, macrophages, osteoblasts, osteocytes, fibroblasts, and lymphocytes [3,6]. Furthermore, research has demonstrated that macrophages predominantly drive the inflammatory response to wear particles and are identified as the primary pathophysiological contributor associated with aseptic osteolysis [7]. Wear particle-induced macrophage-mediated inflammatory injury and tissue damage exacerbate the osteolytic response through various biological functions, including the transformation of macrophages into osteoclasts, the induction of apoptosis in both macrophages and neighboring cells, and the secretion of additional cytokines and proteolytic enzymes [14]. The interconnected nature of these interactions contributes to the continuation and intensification of both inflammatory injury and bone resorption, significantly influencing whether the inflammatory injury initiated by wear particles is successfully resolved or, conversely, worsened, potentially escalating into irreversible osteolysis [7,8,9]. However, current interventions focused on managing inflammation, inhibiting cytokines or enzymes, or reducing osteoclast activity for the prevention and treatment of aseptic osteolysis fall short of being ideal solutions [3,6]. Notably, the presence of apoptotic macrophages has been identified within the osteolytic interface membrane, indicating that the processes associated with macrophage apoptosis are interconnected with periprosthetic osteolysis. This relationship may unveil specific targets for developing advanced pharmacological therapies that focus on macrophage activity and viability, aiming to provide a more precise and effective strategy for managing osteolytic conditions.
The endoplasmic reticulum stress (ERS) represents a crucial response to the accumulation of improperly folded or unfolded proteins within the cellular environment [10,11]. This stress response is essential for re-establishing equilibrium and maintaining cellular homeostasis by improving the cell’s ability to manage and eliminate these dysfunctional proteins, thereby minimizing the potential damage associated with their accumulation [10,11]. When the protein-folding capacity of the ER is exceeded or excessive accumulation of misfolded proteins occurs, the ER transitions into a state of stress (ERS) and activates the unfolded protein response (UPR) to rescue cells from this stress. The recovery of ER functions in cells that have suffered stress-induced damage can result in either survival through the restoration of normal cellular functions or programmed cell death via uncontrolled alternative pathways distinct from the ERS apoptotic pathway [12]. This dual potential underscores the critical balance that cells are required to uphold in the face of stress, further emphasizing the significance of ERS response as a mechanism of cellular protection.
ERS is implicated in various diseases and pathological conditions. As a survival or adaptation mechanism, chaperone molecules are upregulated to manage ERS and maintain cellular homeostasis. In the quest for drug candidates for ERS-related diseases, chemical versions of chaperones, known as chemical chaperones, have been developed, with 4-phenylbutyrate (4-PBA) being one of the most widely recognized examples [10]. This compound influence various cellular processes through several critical mechanisms that are essential for maintaining cell health and functionality, such as inhibiting histone deacetylase, acting as an alternative metabolite in urea cycle disorders (UCDs), and reducing the accumulation of misfolded or excess proteins within the ER [10]. Evidence from animal studies demonstrates that 4-PBA reduces ERS and facilitates recovery from inflammatory injury. This highlights the correlation between ERS and various physiological and pathological conditions, including inflammation [13], infection [12], cancer [14], degenerative musculoskeletal conditions [15], metabolic diseases [16], cardiovascular disease [17], genetic disorders [16], lysosomal storage disorders [18], neurodegenerative disorders [19,20], and autoimmune disorders [21]. These findings suggest that ERS and chemical chaperones may underlie the pathology of these conditions and present a potential therapeutic alternative. Although only a few chemical chaperones have been approved as pharmaceutical drugs, the literature has witnessed a dramatic surge in recent decades, reflecting the substantial efforts made by numerous researchers. Such dedicated endeavors necessitate a clearer understanding of ERS and its associated diseases, thereby providing a promising platform for drug discovery with chaperone activity. This could potentially lead to innovative therapeutic options for patients suffering from related health conditions.
Recent studies have emphasized the significance of ERS as a crucial element in deciphering the intricate mechanisms contributing to aseptic osteolysis [22,23]. The activation of ERS has been documented in both in vivo and in vitro experiments related to particle-induced inflammatory osteolysis. Notably, the use of 4-PBA as a pharmacological treatment has demonstrated efficacy in reducing the severity of osteolysis by inhibiting the inflammatory response mediated through ERS pathways [12,22]. Nonetheless, current studies regarding the influence of 4-PBA on aseptic osteolysis primarily focus on its anti-inflammatory properties [12,22]. Previous research conducted by our team has demonstrated that 4-PBA ameliorate the severity of particle-induced osteolysis by suppressing the inflammatory response and facilitating macrophage apoptosis in an in vivo murine calvaria resorption model. Despite these findings, the effects of 4-PBA on macrophage activity, viability, and apoptosis induced by wear particles in vitro have not been adequately addressed or documented in the existing literature. There remains a notable gap in the exploration of how the regulation of ERS response specifically influences wear particle-induced macrophage-mediated inflammatory injury, particularly regarding its broader implications for macrophage apoptosis.
The ongoing investigation into the therapeutic potential of 4-PBA raises a crucial question: can it effectively alleviate particle-induced inflammatory injury by facilitating macrophage apoptosis while avoiding the activation of an inflammatory response? This question has not yet been answered, underscoring the necessity for further research to elucidate the effects of 4-PBA in modulating macrophage behavior when confronted with wear particles. Such an investigation is significant for understanding the specific pathological conditions that contribute to bone resorption and destruction, as well as for elucidating the specific therapeutic effects of 4-PBA. In the present study, we aimed to investigate the therapeutic potential of the chemical chaperone 4-PBA in modulating TiNP-induced macrophage-mediated inflammatory injury, with a particular emphasis on its role in regulating macrophage apoptosis and inflammation. Our research endeavors are poised to provide valuable insights into how 4-PBA may affect the viability and behavior of macrophages exposed to specific particles, potentially elucidating the underlying mechanisms by which pharmacological treatments can be employed to alleviate such conditions. By thoroughly exploring this relationship, we can enhance the overall understanding of macrophage dynamics and the implications of 4-PBA as a potential co-intervention agent.
2. Materials and Methods
2.1. Nanoparticles Preparation
TiNPs with an average diameter of 51.7 nm were generously provided by Dr. Zhenzhong Zhang from the College of Materials Science and Engineering at Nanjing University of Technology [7,22,24,25]. To ensure the integrity and sterility of the TiNPs, they were meticulously processed under controlled conditions within a cell culture hood, thereby minimizing any risk of contamination. Various solution concentrations were dispersed in the medium for a duration of 10 minutes using a Shumei KQ218 Ultrasonic Cleaner (Kunshan Ultrasonic Instruments Co., Ltd., Jiangsu, China). To further validate the suitability of these particles for subsequent experimental use, a quantitative limulus amebocyte lysate (LAL) assay (Charles River, Grand Island, UK) was performed to confirm the absence of endotoxins. This assay exhibited a detection threshold of 0.25% EU/mL, ensuring that the TiNPs met the necessary standards for reliability and safety prior to their application in further experiments.
2.2. Cell Culture
The murine macrophage-like Raw264.7 cell, obtained from the China Center for Type Culture Collection (Shanghai, China), was cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) to provide essential nutrients for cell growth. Antibiotics were incorporated to maintain a sterile environment during cultivation; specifically, penicillin was added at a concentration of 100 IU/mL, and streptomycin was included at 100 mg/mL. Cells were maintained in a humidified incubator at a temperature of 37 °C, with a controlled atmosphere comprised of 5% carbon dioxide and 95% air to simulate physiological conditions.
To optimize the experimental conditions for subsequent procedures, the cells were subjected to serum deprivation for 16 to 18 hours before initiating the formal experimental protocols. This step was essential to minimize the serum’s potential impact on the outcomes and help the cells achieve a relatively stable metabolic state, thereby providing an optimal environment for the interventions to be applied. In situations where 4-PBA (Sigma-Aldrich, St. Louis, MO, USA) was employed, this compound was administered either as a standalone therapy or in combination with other interventions. Prior to the main experimental procedures, the cells underwent a preparatory phase, during which they were pre-treated with 4-PBA at concentrations of 2 or 10 mmol/L for 8 hours. This pretreatment was crucial for adequately preparing the cells for the subsequent experimental manipulations, thereby ensuring optimal conditions for the intended procedures.
RAW264.7 cells were divided into four distinct culture groups using a randomization technique based on a digital table method. Each group consisted of three samples to ensure the replicability of results. In the Control group, cells were maintained in serum-free DMEM to establish a baseline for comparison. The TiNPs group was exposed to a range of TiNP concentrations (10, 20, 50, 100, 200, 500, 1000, 2000, and 5000 µg/mL) to evaluate the effects of increasing TiNP dosages on the cells. Concurrently, the 4-PBA group received treatments with varying levels of 4-PBA (0, 0.5, 1, 2, 5, 10, 20, and 50 mmol/L) to assess how different levels of 4-PBA influenced RAW264.7 cells. Lastly, the TiNPs + 4-PBA group was treated with 1000 µg/mL of TiNPs in conjunction with different concentrations of 4-PBA. This combination treatment sought to investigate the interactions between TiNPs and 4-PBA, thereby providing insights into their cumulative effects on cell behavior.
2.3. Cell Viability
The viability of Raw264.7 cells was assessed following a culture duration of 24 and/or 48 hours. The influence of TiNPs, 4-PBA, and their combined synergistic effects was evaluated using an MTT assay (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazolium bromide, supplied by Sigma-Aldrich), adhering strictly to the manufacturer’s guidelines.
2.4. Macrophage Apoptosis
The assessment of macrophage apoptosis was conducted using the ANXA5 (Annexin V)-FITC/PI apoptosis detection kit (4A Biotech Co. Ltd., FXP018–100) following the manufacturer’s instructions. Subsequent analysis was performed using FACScan Cellquest software (Becton Dickinson, San Jose, CA, USA). In the staining analysis quadrants, Annexin V-/PI- cells in the lower left region represent viable cells, while Annexin V+/PI- cells in the lower right region indicate early-stage apoptotic cells. Annexin V+/PI+ cells in the upper right region represent late-stage apoptotic cells, and cells designated as Annexin V-/PI+ in the upper left corner are considered necrotic cells.
2.5. ROS Production
The production of reactive oxygen species (ROS) by macrophages was quantified through flow cytometry, in accordance with the established protocols detailed in prior studies [26]. This assessment was conducted immediately after the cells were resuspended, employing FACScan Cellquest software for the analysis of the flow cytometry results.
2.6. Caspase-3 Activities
The supernatant obtained from cell lysis was prepared to assess the activity of Caspase-3 (Caspase-3 colorimetric assays from BioSource, Belgium). Each procedural step was executed in strict adherence to the manufacturer’s guidelines. The results were normalized based on the total protein content of the cells, which was quantified using a commercially available kit, and reported as U/gprot.
2.7. Inflammatory Cytokines
Cytokines that promote inflammation and respond to inflammatory injury, including tumor necrosis factor-alpha (TNF-α), interleukin-1 beta (IL-1β), and interleukin-6 (IL-6), were quantitatively assessed using enzyme-linked immunosorbent assay (ELISA) kits (Jonin Biotech, Shanghai, China). All procedures were conducted in accordance with the manufacturer’s protocols.
2.8. Western Blotting (WB)
Equal amounts of total protein extracted from cell lysis supernatants were subjected to separation using 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) (1.5 M TRIS·HCl, pH 8.8, 30% acrylamide, 10% SDS, AP, TEMED). Following electrophoresis, the proteins were transferred to nitrocellulose membranes (Millipore, Billerica, MA, USA) to inhibit nonspecific interactions. The membranes were subsequently blocked with nonfat milk and then incubated and probed with rabbit polyclonal antibodies against Bcl-2, Bax (Proteintech, Wuhan, China), and Caspase-3 (Abway, Shanghai, China). Afterward, a secondary antibody conjugated with horseradish peroxidase (HRP) against rabbit IgG (Seracare, Los Angeles, USA) was applied. The molecular weight of the purified proteins was assessed via SDS-PAGE, and the corresponding bands were excised for further identification using a chemiluminescence detection system (Shenhua Science Technology, Hangzhou, China). The density of the protein bands was quantitatively analyzed using ImageJ 1.41 (National Institutes of Health, Bethesda, MD, USA).
2.9. Statistical Analysis
Data are presented as the mean ± standard error of the mean (SEM), providing a comprehensive overview of the variability and central tendency of the measurements taken. To assess the differences in the mean values of the experimental parameters across various groups, analysis of variance (ANOVA) was employed. This method allows for an effective comparison of the means from multiple groups to determine whether any significant differences exist. Post hoc testing for differences between groups was conducted using Duncan’s test when ANOVA indicated significance. A threshold P-value of less than 0.05 was established to signify a statistically significant difference, suggesting that the observed differences were unlikely to have occurred by chance. No adjustments for multiple testing were made, as the overall approach was exploratory in nature, indicating a primary focus on generating insights rather than stringent correction methodologies. All statistical analyses were performed using SPSS v17.0 (SPSS Inc, Chicago, IL, USA).
3. Results
3.1. The Effect of 4-PBA on TiNP-Induced Macrophage Viability
The impact of TiNPs and/or 4-PBA on the viability of Raw-264.7 macrophages was evaluated through MTT assays, as depicted in Figure 1A–D. A notable correlation was identified between macrophage viability and stimulation of TiNPs (P<0.01), suggesting that wear particles have a considerable influence on the survival of macrophages. During the macrophage culture process, an incremental increase in both the concentration of TiNPs and the duration of exposure resulted in a marked decline in macrophage viability, demonstrating both dose- and time-dependent responses (Figure 1A,B). Furthermore, a substantial reduction in the number of macrophages exposed to TiNPs over a 48-hour period was observed. This decrease in cell count was significant enough that it failed to meet the required cell count for the subsequent experiments. Consequently, a stimulation duration of 24 hours would be more appropriate for conducting the follow-up experimentation. Additionally, during the 24-hour exposure window, significant alterations in macrophage viability were observed when cells were exposed to TiNPs at a concentration of 100 µg/mL (P<0.01). However, when the concentration of TiNPs exceeded 1000 µg/mL, a notable reduction in the variability of the effects on macrophage viability was recorded. When conducting investigations under in vitro conditions, it is essential to consider not only the concentration of stimuli employed but also the practical feasibility of the experiments. High concentrations of metal wear particle suspensions can pose substantial challenges related to cleanliness; increased washing frequency may negatively impact the attached cells, leading to potential complications and confounding factors that could compromise the reliability of the results. In light of these considerations, we evaluated various concentration levels and ultimately determined that a stimulation concentration of 1000 µg/mL would be the most appropriate for our experiments. To establish a clear comparative baseline for our studies, we also set a concentration of 100 µg/mL as a low concentration control. This deliberate selection was made to ensure that our results would be both robust and credible. Finally, the concentrations of TiNPs selected for subsequent experimentation were determined to be 100 and 1000 µg/mL.
To investigate the potential effects of 4-PBA on macrophage viability, various concentrations of 4-PBA, ranging from 0 to 50 mmol/L, were administered to macrophages both in isolation and in conjunction with 1000 µg/mL TiNPs for a duration of 24 hours. The application of 4-PBA as a standalone treatment resulted in a dose-dependent decrease in macrophage quantity (Figure 1C). Specifically, significant alterations in macrophage viability began to manifest after 24 hours of exposure to 2 mmol/L of 4-PBA (P<0.01). Notably, at a concentration of 10 mmol/L of 4-PBA, a discernible reduction in the variability of the effects on macrophage viability was observed, indicating a more consistent response at this dosage. Furthermore, the transient inhibitory effect of TiNPs on macrophage viability was further amplified by the concurrent application of 4-PBA in a dose-responsive manner (Figure 1D). When macrophages were exposed to TiNPs at a concentration of 1000 μg/mL, significant changes in their viability were observed upon the combined treatment with 2 mmol/L of 4-PBA (P<0.01). However, as the concentration of 4-PBA exceeded 10 mmol/L, the rate of change in macrophage viability began to decelerate. Consequently, the concentrations of 4-PBA chosen for the subsequent experiment were established at 2 and 10 mmol/L, reflecting the critical thresholds observed in macrophage responses. This selection aims to enhance the analysis of how 4-PBA influences macrophage activity and response, thereby providing a clearer understanding of its biological impact.
3.2. The Suppressive Effect of 4-PBA on TiNP-Induced Inflammation in Macrophages
As illustrated in Figure 1E–G, the investigation into the influence of TiNPs and/or 4-PBA on inflammation in macrophages was conducted. The introduction of TiNPs to macrophages resulted in a significant increase in the levels of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6), with the extent of this increase directly correlating to the dosage of TiNPs administered (P<0.05). Furthermore, when examining the modulatory impact of 4-PBA on TiNP-induced inflammation in macrophages, it was observed that 4-PBA effectively diminished the expression of these inflammatory factors elevated by TiNPs exposure (P<0.05). Notably, this suppressive effect exhibited a dose-dependent characteristic, indicating that higher concentrations of 4-PBA corresponded to more pronounced reductions in the expression of pro-inflammatory cytokines. However, when macrophages were treated solely with 4-PBA in the absence of TiNPs, its impact on the expression of pro-inflammatory cytokines was found to be relatively minimal (P>0.05).
3.3. The Alleviative Effect of 4-PBA on TiNP-Induced ROS Production in Macrophages
As illustrated in Figure 1H and Figure 2A–C, notable increases in intracellular ROS levels were observed following exposure to TiNPs, with the magnitude of this increase being influenced by both the concentration and duration of exposure (P<0.05). Specifically, higher concentrations of TiNPs correlated with a more pronounced elevation in ROS production, while prolonged exposure times further exacerbated ROS generation. To evaluate the sustained impact of 4-PBA on ROS levels, this compound was employed as a co-intervention strategy to assess its regulatory effect on TiNP-induced ROS production in macrophages. A distinct dose-dependent reduction in ROS levels was observed when treated with different concentrations of 4-PBA in the presence of TiNPs over exposure durations of 6 hours (Figure 2A), 12 hours (Figure 2B), and 24 hours (Figure 2C) (P<0.05). This clear trend indicates that 4-PBA is capable of significantly alleviating the oxidative stress response in macrophages triggered by TiNPs.
3.4. The Facilitatory Effect of 4-PBA on TiNP-Induced Macrophage Apoptosis
To investigate whether the observed transient decrease in macrophage viability, along with the reduction of inflammation and oxidative stress responses caused by 4-PBA, results from apoptosis, we employed Annexin V-FITC/PI double-labeling to evaluate the externalization of phosphatidylserine (PS), a recognized marker of early-stage apoptosis. The results presented in Figure 3A–C and Figure 4A demonstrate a notable increase in the proportion of apoptosis induced by TiNPs, which follows a dose-dependent pattern (P<0.05). Significant differences compared to the Control group were observed at various time intervals: 6 hours (Figure 3A), 12 hours (Figure 3B), and 24 hours (Figure 3C) (P<0.05). Furthermore, the combined application of 4-PBA significantly amplified the effects of TiNPs on macrophage apoptosis in a dose-dependent manner (P<0.05), indicating that 4-PBA intensifies the apoptotic response elicited by TiNPs in macrophages. Interestingly, when the presence of 4-PBA was in conjunction with TiNPs, its impact on apoptosis was pronounced; however, in the absence of TiNPs, its influence on macrophage apoptosis was minimal (P>0.05). Additionally, our findings revealed that 4-PBA primarily facilitates the early-stage apoptosis of macrophages induced by wear particles, rather than influencing late-stage apoptosis or necrosis.
3.5. The Effect of 4-PBA on TiNP-Induced Apoptotic Regulatory Proteins in Macrophage Apoptosis
Given the crucial role of Caspase-3 in regulating the apoptosis process, we investigated the enzyme activity of Caspase-3 in macrophages (Figure 4B–D). Significant increases in Caspase-3 activity were observed when macrophages were exposed to TiNPs, demonstrating both dose- and time-dependent characteristics (P<0.05). Specifically, as the concentration of TiNPs increased and the duration of exposure extended, there was a consistent upward trend in Caspase-3 activity, indicating that the influence of TiNPs on Caspase-3 activity in macrophages is intricately linked to both concentration and exposure duration. Furthermore, following co-treatment with a high concentration of TiNPs at 1000 µg/mL, a time-dependent increase in Caspase-3 activity was observed with the combined intervention of 4-PBA. Notably, a dose-dependent response was evident after 12 and 24 hours of exposure (P<0.05). However, when the duration of the combined intervention with 4-PBA was reduced to 6 hours, the activation effect on Caspase-3 was significantly diminished (P>0.05). It is particularly noteworthy that the influence of 4-PBA on Caspase-3 activity exhibited conditional dependence: in scenarios where TiNPs were present, 4-PBA had a pronounced effect, further enhancing Caspase-3 activity (P<0.05). Conversely, in the absence of TiNPs, the impact of 4-PBA on Caspase-3 activity in macrophages was minimal (P>0.05).
To investigate the involvement of apoptotic regulatory proteins in macrophage apoptosis induced by 4-PBA, we evaluated the expression levels of Caspase-3 (Figure 4E,F), Bax (Figure 4E,G), and Bcl-2 (Figure 4E,H) under various conditions. This assessment also aimed to determine whether the activity of Caspase-3 correlated with the expression levels of these apoptosis-related proteins. Following exposure to TiNPs, we observed a significant increase in the expression of Caspase-3 and Bax, while the levels of Bcl-2 exhibited a marked decrease (P<0.05). Furthermore, after the combination treatment of TiNPs and 4-PBA, there was a substantial increase in the expressions of Caspase-3 and Bax, whereas the expression of Bcl-2 was suppressed (P<0.05). Notably, the application of 4-PBA as a standalone treatment yielded only a marginal effect on the expression of apoptotic regulatory proteins (P>0.05).
4. Discussion
As a fundamental basis for artificial joints, metal implants find extensively utilized in orthopedic surgery [1]. However, the inevitable wear and corrosion that occur within the alloy during the lifespan of a joint prosthesis pose significant challenges. This degradation results in the release of metal wear particles into the surrounding periprosthetic tissues, which can have detrimental consequences [4,27]. The biological response to these wear particles is multifaceted, encompassing a variety of harmful effects, including cytotoxicity, compromised cell viability, and inflammatory injury [4,27]. Over time, these reactions can culminate in aseptic osteolysis, a condition characterized by bone resorption in the absence of infection, ultimately leading to the loosening and eventual failure of the prosthetic joint [28].
In patients experiencing aseptic osteolysis of joint prostheses, TiNPs have been detected in the membranes located at the periprosthetic interface [1,29,30]. Generally, the predominant ultrastructural features of these particles found within the affected periprosthetic interface membranes typically fall within a size range of 30 to 60 nm, with these nanoparticles primarily being phagocytosed by macrophages [23,31]. Understanding the interactions between metal wear particles and macrophage-mediated inflammatory injury, as well as the biological tissue response, is critical for informing future strategies to mitigate the adverse effects associated with metal implants. Furthermore, TiNP-induced macrophage-mediated inflammatory injury serves as a prevalent in vitro model for assessing pharmaceutical effects on aseptic osteolysis within the field of cellular molecular research. The mechanisms underlying this model bear similarities to those associated with other types of wear particles, making it a valuable reference point for further investigation. To synchronize with our research objectives and build upon earlier transmission electron microscopy findings [23,31], we opted to incorporate TiNPs, which possess an average particle size of 51.7 nm, into the in vitro cellular studies.
4.1. The Role of Wear Particle in Macrophage-Mediated Inflammatory Injury
The findings from our study demonstrate that TiNPs possess properties that can induce inflammatory injury to macrophages, suggesting their potential to damage cellular structures. Specifically, TiNPs impair the functionality and activity of macrophages that manage to survive exposure, resulting in heightened inflammatory responses and increased oxidative stress within these immune cells. Furthermore, our study reveals that TiNPs contribute to a reduction in cell population by promoting apoptosis in macrophages. These conclusions are consistent with results from previous studies [32], thereby reinforcing the understanding of the detrimental effects of TiNPs on immune cell dynamics. Additionally, our research indicates that macrophages exhibit a relatively weak stress reaction when subjected to low-dose TiNPs exposure, suggesting a certain level of tolerance and a comparatively sluggish condition. In contrast, exposure to high doses of TiNPs elicits a positively response from macrophages, leading to a subdued physiological state characterized by a pronounced stress reaction, significant cellular dysfunction, and ultimately detrimental effects on cell viability and integrity. Importantly, the stress reaction in macrophages due to wear particles not only enhances with increasing doses but also demonstrates escalating intensity over time. This highlights that the inflammatory injury caused by TiNPs on macrophages accumulates progressively with both the amount of exposure and the duration of exposure. These observations align with conclusions drawn in previous studies, reinforcing our understanding of how TiNPs impact macrophage function.
Our research, supported by earlier studies, provides compelling evidence that the effect of wear particles on macrophages is significantly influenced by two primary factors: the concentration of particles and the duration of exposure. The introduction of wear particles to macrophages resulted in a marked increase in inflammatory injury levels. The extent of wear particle-induced macrophage-mediated inflammatory injury is largely contingent upon both the duration of particle presence and the particle dosage received by the cells (i.e., the concentration of phagocytosable particles per cell), which is determined by the total volume/mass. Therefore, the concentration and exposure time of wear particles in macrophages, alongside the overall quantity of stress-damaged macrophages that have survived without undergoing apoptosis, are critical determinants of macrophage-mediated inflammatory injury. By elucidating the interplay between these variables, we can gain deeper insights into the mechanisms underlying macrophage-mediated aseptic osteolysis, which may ultimately inform therapeutic strategies or risk assessments regarding materials that generate wear particles.
4.2. The Role of Macrophages in Wear Particle-Induced Aseptic Osteolysis
Macrophages are essential innate immune effector cells, capable of rapid, non-specific responses to threats without prior antigen exposure [6,7,8,9,33]. They maintain tissue homeostasis and are primarily tasked with providing rapid, front-line defense against the invasion of noxious stimuli that could disrupt physiological equilibrium [6,8,9,33]. As immune “scavengers,” they phagocytose dead cells and debris, mediate host responses to infections and tumors, initiation inflammation, and regulate tissue repair through four innate capabilities: phagocytosis, sensing, chemotaxis, and adaptive stimulation [6,8,9,33].
As multifunctional sentinels, macrophages play a central role in recognizing wear particles and triggering inflammatory cascades leading to aseptic osteolysis [6,7,8,9,27,33,34,35,36,37,38]. While various cell types—including neutrophils, monocytes, macrophages, osteoblasts, osteocytes, fibroblasts, and lymphocytes—are overactivated and contribute to particle-induced osteolysis, macrophages are established as the primary drivers of the inflammatory response to wear particles and are considered a key pathophysiological factor in aseptic osteolysis [3,6,39,40,41,42,43]. They constitute 60–80% of the overall cell composition in osteolytic interface membranes around loosened prostheses [44].
The biological response to wear particles in aseptic osteolysis is intricate, involving interactions among prosthetic biomaterials, tissues, and cells, as well as between particles and tissues/cells, and among cells and tissues, alongside various signaling pathways [34]. At the heart of this concept is the fact that extremely small particles stimulate periprosthetic macrophages to produce inflammatory injury. The mediators, resulting from inflammatory injury, orchestrate increased formation, accumulation, activity, and survival of osteoclasts while simultaneously suppressing the osteogenic functions of osteoblasts [6]. This tilts the balance toward bone destruction over regeneration, ultimately leading to net bone destruction and osteolysis at the implant interface [36].
Inflammatory injury represents a fundamental and universal response of the immune system to various external and internal threats, including foreign substances, necrotic tissues, and unfavorable mechanical or metabolic stimuli [6,8,9,33]. Wear particles, continuously generated at the bone-prosthesis interface, are actively phagocytosed by resident tissue macrophages [45]. These macrophages perform their essential phagocytic function by ingesting wear particles; however, this persistent interaction may also initiate innate immunity (nonspecific immunity) [24]. This sustained state of stimulation can further exacerbate the inflammatory injury, leading to increased tissue damage [8,27,39]. The presence of macrophages in conjunction with wear particles indicates an active immune response aimed at addressing the chronic inflammation triggered by the continuous presence of implant-related wear particles [6,8,9,33]. In cases of prosthetic loosening, the immune response is typically characterized by the formation of interface membranes, where wear particles are co-localized with macrophages. Furthermore, wear particles and macrophages elicit an inflammatory response and facilitates the formation of osteoclasts, thereby contributing to the development of aseptic osteolysis [22,23,34].
Periprosthetic macrophages originate from blood and bone marrow monocytes, playing a crucial role in immune surveillance [6,8,9,33]. Failed implant tissues exhibit dense macrophage infiltration in interface membranes, with these migratory cells and resident macrophages possess the capacity to differentiate into osteoclasts [34,44]. The accumulation of particles triggers the migration of monocyte/macrophage precursors to the localized particle sites, leading to systemic activation [6,8,9,33]. This activation does not solely occur in the immediate vicinity of the prosthesis; rather, it also stimulates circulating monocytes and other immune cell types [39]. Failed implant tissues exhibit dense macrophage infiltration in interface membranes, with these migratory cells and resident macrophages possess the capacity to differentiate into osteoclasts [34,44]. Such expanded activation indicates that the role of macrophages in the immune response is multifaceted, extending into the bloodstream and impacting not only the localized environment but also the systemic immune responses [9]. The ability of macrophages to communicate and collaborate with various subsets of immune cells highlights their essential function in orchestrating efficient immune responses across diverse environments within the organism [9,39]. This intricate interaction guarantees the immune system to adapt and respond efficiently to various challenges it encounters.
Therefore, it is reasonable to propose that macrophages play a dual role in the inflammatory cascade triggered by wear particles, as well as in the subsequent process of aseptic osteolysis [6,7,8,9,27,33,34,35,36,37,38]. They serve not only as the primary “initiators” of the pathological cascade resulting in bone resorption but also as “catalysts” and “amplifiers” of this detrimental process [39]. This indicates that their involvement is critical at multiple stages of the inflammatory response. Consequently, the dynamics of the interaction between macrophages and wear particles at the interface membrane are likely to be a key determinant in the outcome of this condition [9]. The nature of this interaction will significantly influence whether the inflammatory injury triggered by wear particles is effectively resolved or, conversely, exacerbated, potentially escalating into permanent osteolysis [7,8,9].
4.3. The Crosstalk Between Macrophage Apoptosis and Inflammation in Periprosthetic Microenvironment
In vitro studies have confirmed that wear particles induce macrophage apoptosis within the periprosthetic microenvironment, particularly in periprosthetic tissues of patients experiencing aseptic loosening [23,34,35,44,46,47]. A significant correlation has been established between the severity of osteolysis progression and an increased incidence of macrophage apoptosis [23,34,44]. Furthermore, the processes of macrophage apoptosis in periprosthetic microenvironment involves activation of all major pathways: death receptor pathway, ERS pathway, and mitochondrial pathway [23,34].
Apoptosis is typically regarded as a “clean” mechanism for the elimination of dead cells, which generally does not provoke significant inflammatory responses [34,44]. This characteristic is crucial as it facilitates the efficient clearance of cellular debris without causing undue harm to surrounding tissues [44]. However, the continuous use of prostheses and the ongoing generation of wear particles can overwhelm the critical threshold of apoptosis and stress reactions, thereby impairing the macrophage clearance mechanism [44]. This failure in clearance promotes the accumulation of apoptotic debris and unresolved tissue inflammation. Consequently, this accumulation leads to the migration and aggregation of systemic macrophages, forming a vicious cycle of wear particle-induced macrophage-mediated inflammatory injury. Furthermore, this cycle is exacerbated by a decline in the phagocytic capacity, scavenging mechanisms, and the ability of these stressed and damaged macrophages to undergo apoptosis. As these macrophages fail to undergo normal programmed cell death and instead persist in a state of inflammatory injury, they secrete additional inflammatory mediators, which further aggravate the inflammatory damage to surrounding cells and accelerate the process of osteolysis. Consequently, the ineffective elimination of these stressed and damaged macrophages may complicate tissue management and inadvertently provoke an inflammatory response in the affected area.
The existing literature has extensively detailed the mechanisms underlying macrophage apoptosis and the remission of inflammation [34,44]. Macrophage apoptosis generally suppresses pro-inflammatory signals and is regarded as an indicator of inflammation resolution, signaling a shift towards normal tissue healing in particles-induced osteolysis [44]. This phenomenon has also been observed in various other inflammation conditions, such as impaired wound healing [48,49], pulmonary inflammation and fibrosis [50], alcohol-induced hepatic fibrosis [51], and Alzheimer’s disease [19]. In these cases, the extent and intensity of apoptotic reactions are often so pronounced that the remnants of stress-damaged cells cannot be completely eliminated from the tissue [44]. Consequently, this incomplete removal of cellular debris contributes to an el increased evated release of fibrogenic mediators, which may be responsible for enhanced fibroblast proliferation despite the elevated levels of apoptosis [44]. Moreover, this process of ineffective elimination results in elevated inflammatory factors that may trigger the activation of osteoclasts and the recruitment of systematic macrophages, potentially exacerbating the progression of inflammatory injury [44]. As previously mentioned [22,23,24], the progression of periprosthetic osteolysis correlates with both increased macrophage apoptosis and elevated local inflammatory factors. In this study, wear particles alone induced significant macrophage apoptosis alongside concurrent inflammatory escalation.
4.4. The Protective Role of Chemical Chaperone 4-PBA in Wear Particle-Induced Macro-Phage-Mediated Inflammatory Injury
Given the important role of macrophages in the development of wear particle-induced aseptic osteolysis, the transient and selective depletion of macrophages in periprosthetic tissues may benefit patients at high risk for developing osteolysis. This study aimed to determine whether treatment with 4-PBA reduces the severity of wear particle-induced macrophage-mediated inflammatory injury. Our results demonstrate that the ER chemical chaperone 4-PBA significantly alleviates the inflammatory response and oxidative stress in macrophages triggered by wear particles, consistent with findings from previous studies [12,22,24,25,52,53,54,55,56]. Notably, our investigation revealed a critical aspect: when 4-PBA was administered in the presence of wear particles, it not only mitigated the inflammatory response associated with the condition but also promoted an increase in macrophage depletion and apoptosis. 4-PBA has the potential to further impair the viability and activity of stress-damaged macrophages experiencing inflammatory injury due to wear particles. This dual action underscores the potential therapeutic benefits of 4-PBA in managing aseptic osteolysis by addressing both inflammation and macrophage lifespan.
The compound 4-PBA, a low-molecular-weight chemical chaperone, has been identified as a potential therapeutic agent for ERS-related pathologies. It plays a crucial role in maintaining cellular homeostasis and alleviating the detrimental effects of ERS, while also exhibiting anti-inflammatory properties [12,22,24,25,52,53,54,55,56]. Furthermore, 4-PBA acts as a small aromatic fatty acid and a histone deacetylase inhibitor, rendering it a compelling compound capable of inhibiting cell growth, promoting anti-proliferative effects, and inducing apoptosis in stress-damaged cells [14,16,57,58,59,60,61,62]. These attributes position 4-PBA as an attractive candidate for further investigation in therapeutic applications related to osteolytic conditions. Given that 4PBA is already being utilized in human patients and is currently undergoing various clinical trials, it is essential to comprehensively understand its impact on osteolysis and the mechanisms through which it exerts its effects [12].
The evident occurrence of ERS in particles-induced osteolysis, coupled with the anti-inflammatory effects of 4-PBA [12,22,24,25,52,53,54,55,56], supports the hypothesis that employing 4-PBA to inhibit ERS may offer therapeutic benefits in alleviating bone resorption and decelerating the progression of osteolysis. The apoptotic pathway triggered by 4-PBA has been progressively elucidated across various cell types, particularly in terms of its initiation and termination. The initiation phase is characterized by the inhibition of histone deacetylases, which are crucial for regulating gene expression and cellular functions. Meanwhile, the termination phase is believed to involve the activation of caspases [63]. Although the precise mechanism underlying macrophage depletion due to intracellular accumulation of 4-PBA remains unclear, especially in particle-induced osteolytic environments, it is hypothesized that this accumulation generates signals from 4-PBA that promote macrophage apoptosis.
The response to ERS can facilitate apoptosis and enhance inflammation [64,65,66]. Notably, our investigation revealed that the application of 4-PBA, aimed at inhibiting this stress response, results in a decrease in inflammation while concurrently elevating apoptosis levels. The suppression of ERS signaling leads to a reduction in inflammation, which may be beneficial in various pathological situations where chronic inflammation is a concern [24]. Nonetheless, this reduction in inflammation is accompanied by an undesirable consequence: an increase in apoptosis. Thus, while the inhibition of ERS signaling via 4-PBA can be advantageous by alleviating inflammation, it also carries the significant trade-off of elevating apoptotic activity within cells. This phenomenon highlights the complex interplay that the ERS response maintains between promoting cell survival and triggering cell death in macrophages subjected to stress.
The presence of unremoved apoptotic bodies and remnants generated during macrophage apoptosis, alongside stress-damaged cells that have survived without undergoing apoptosis, may undergo secondary necrosis [39]. This process can lead to the release of various inflammatory and cytotoxic substances into the surrounding environment, contributing to particles-induced osteolysis [39]. Such a release poses a risk of further damaging nearby cells and may exacerbate tissue inflammation and proliferation [34,44]. This underscores the critical need for effective pharmacological intervention to maintain cellular homeostasis, prevent the accumulation of stress-damaged cells, and alleviate the associated inflammatory responses. Pharmacological intervention with therapeutic chaperones represents a promising strategy for managing macrophage health, especially in cases where these immune cells are adversely affected by exposure to particulate matter [24]. Therapeutic chaperones demonstrate promising potential in facilitating apoptosis and, in conjunction with their ability to modulate the activation of cells that have experienced stress-induced damage, may play a crucial role in restoring immune balance [61]. By addressing the detrimental effects of these particles on the function and viability of macrophages, therapeutic chaperones can enhance immune responses, ultimately leading to improved health outcomes for individuals exposed to such environmental stressors [22,25].
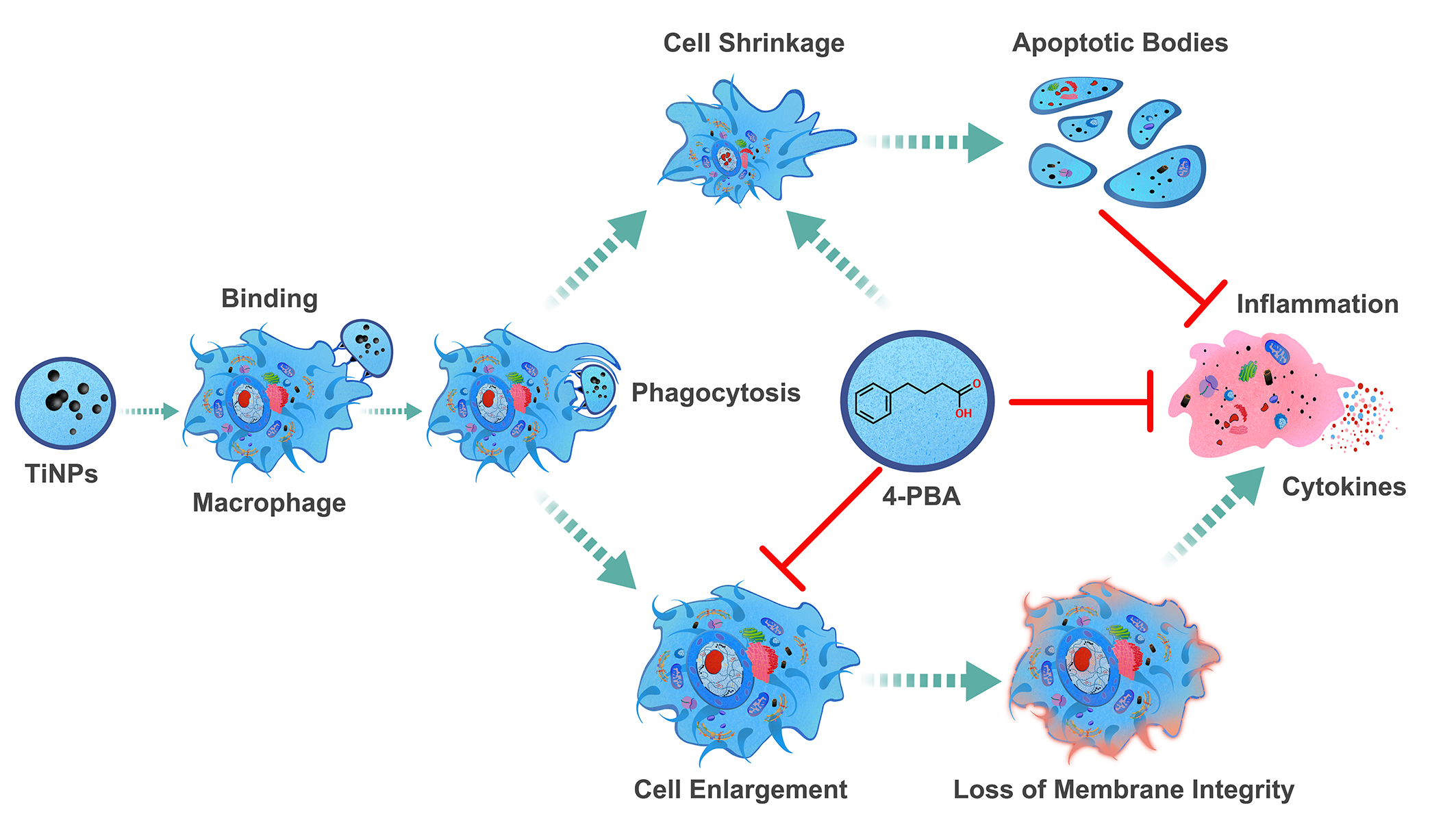
Research has indicated that promoting apoptosis in stress-damaged macrophages—which can survive without undergoing programmed cell death and exhibit characteristics distinct from typical macrophages—could be a valuable therapeutic strategy. Targeting this apoptosis could significantly alleviate issues related to aseptic osteolysis and help prevent the loosening of prosthetic devices. The unique behavior of these altered macrophages underscores an important avenue for developing treatments aimed at combating these complications. Fortunately, the compound 4-PBA, a crucial element of therapeutic chaperones, possesses a variety of beneficial properties and functionalities that render it particularly effective in tackling the challenges identified in our investigation of the particle-induced macrophage-mediated inflammatory injury model, as well as in our prior earlier study involving the particle-induced murine calvaria resorption model [13]. The remarkable versatility and effectiveness of 4-PBA underscore its potential as a significant therapeutic agent in addressing inflammatory injuries resulting from particle exposure (Figure 5). Moreover, its effectiveness extends beyond this context, as it has also demonstrated effectiveness in various other inflammatory conditions explored in prior research [14,57,63]. This broad applicability reinforces the importance of further investigating 4-PBA’s therapeutic capabilities across different inflammatory scenarios.
The 4-PBA approach is designed to leverage the inherent biological mechanisms of immune cells to achieve two primary objectives: inhibiting undesirable apoptotic processes and promoting beneficial ones [21,61]. By adopting this dual strategy, the approach aims to enhance the overall functionality of immune cells, which is crucial for improving health outcomes. Through the implementation of the 4-PBA process, the body gains the ability to precisely regulate the number of immune cells within the system [57,67]. This regulation is vital as it facilitates the effective identification and removal of cells that have been damaged due to stress, ensuring that only healthy and functional cells remain to support the immune response [14,57]. Moreover, 4-PBA is characterized by relatively low toxicity levels, which is a significant advantage for its therapeutic applications. Its stability further enhances its appeal, as a stable compound is crucial for effective treatment. Additionally, 4-PBA is generally well-accepted in clinical contexts, having undergone rigorous safety evaluations. One of its remarkable features is its ability to be administered safely on a daily basis, which can extend throughout a person’s lifetime without adverse effects. Importantly, the body naturally synthesizes 4-PBA within the intestinal tracts of both humans and animals, suggesting that it is a familiar compound with a well-integrated biological presence [57]. In particular, the findings highlight that using 4-PBA as a standalone treatment, without wear particles, exhibited minimal toxicity and was deemed safe based on experimental results. Furthermore, it is significant that this treatment did not induce notable alterations in the inflammatory mediators produced by macrophages, nor did it provoke a pronounced inflammatory response, irrespective of whether the assessments occurred in vivo or in vitro. These findings are particularly significant, suggesting that 4-PBA may be effectively utilized in various therapeutic scenarios while minimizing health risks, at least during the preliminary evaluation phases. The experimental findings support the conclusion that 4-PBA is safe for use in medical treatments.
The findings from this research, in conjunction with the identified dual function of the ER chemical chaperone 4-PBA [14,57,63,67], highlight the intricate relationship between 4-PBA and the regulation of ERS, the inflammatory response, and the survival and proliferation of macrophages in osteolytic disease conditions. The complexity of these relationships suggests that interventions targeting ERS and inflammation may have varied and intricate effects on cellular outcomes and the overall progression of the disease (Figure 5). This underscores the necessity for a detailed understanding of these processes when developing effective therapeutic approaches for diseases where both ERS and inflammation play significant roles in progression. Furthermore, the dynamic interplay between cellular stress responses and the regulatory processes that determine cell survival and apoptosis warrants further exploration to elucidate the fundamental mechanisms involved. Investigating these interactions may provide crucial insights into the regulatory systems that dictate cell fate under stress conditions. By delving deeper into these connections, researchers may uncover critical insights into the regulatory frameworks that govern cell fate in response to stress.
The TiNP-induced macrophage-mediated inflammatory injury model employed in this study offers a sensitive, rapid, economical, and reproducible approach to elucidate the cellular and molecular mechanisms underlying particle-induced osteolysis. However, several limitations must be acknowledged when interpreting the results. Firstly, the characteristics of wear particles—such as size, shape, and material composition—significantly influence the severity of osteolysis [68]. Our research specifically concentrated on nanoscale TiAl6V4 metal particles. Nonetheless, the mechanisms of osteolysis induced by TiNPs are comparable to those associated with various other wear particles, including CoCrMo, polyethylene, and ceramic particles. The TiNP-induced inflammatory injury model is the most widely used framework for evaluating the impact of pharmaceuticals on osteolysis, thereby facilitating meaningful comparisons of our results with existing literature. Therefore, further exploration into the biological responses elicited by different sources and types of wear particles is essential for a thorough validation of our conclusions. Secondly, given that wear particles can interact with various cell types and provoke complex and detrimental local responses at the cellular and molecular levels, future investigations should explore the effects of 4-PBA on other critical cell types, including osteoblasts, osteoclasts, osteocytes, and fibroblasts. This exploration will provide a comprehensive understanding of the potential of 4-PBA in modulating biological responses within the periprosthetic microenvironment. Thirdly, 4-PBA, known for its low toxicity and selectivity towards stressed cells, may provide a novel therapeutic strategy for inflammatory osteolysis that is refractory to traditional anti-inflammatory agents. This compound could be regarded as a potential option for treating osteolysis, which presents a significant unmet medical need, however, further data regarding its efficacy in clinical trials with long-term safety profile considerations, as well as comparative trials with existing therapies prior to clinical application, are necessary. Furthermore, our findings provide initial evidence that the apoptosis of macrophages induced by 4-PBA may positively influence inflammatory osteolysis. Nevertheless, the precise downstream mechanisms facilitating this process remain unclear and warrant further investigation. A more in-depth exploration of these mechanisms is crucial to enhance our understanding of how 4-PBA confers its protective benefits in the context of inflammatory osteolysis. Currently, 4-PBA is undergoing clinical trials in conjunction with other therapeutic agents to assess potential synergistic effects [20,69,70]. Future research should also focus on the potential integration of 4-PBA with additional drugs for the treatment of inflammatory osteolysis. Identifying the molecules and pathways that 4-PBA activates to induce apoptosis in stressed macrophages is of therapeutic significance for the classification of inflammatory osteolysis and the management of patients with aseptic loosening using 4-PBA, whether as a monotherapy or in combination with other agents.
5. Conclusions
The introduction of TiNPs to macrophages resulted in a significant increase in inflammatory injury levels. The duration of particle presence, the dosage received by macrophages, and the overall quantity of stress-damaged macrophages that have survived without undergoing apoptosis are critical determinants of macrophage-mediated inflammatory injury. Macrophages contribute to the severity of TiNP-induced inflammatory injury, indicating their potential as therapeutic targets for preventing and treating patients with total joint replacements who are at high risk of developing osteolysis. Furthermore, intervention with pharmacological chaperone 4-PBA alleviates the severity of particle-induced macrophage-mediated inflammatory injury by suppressing the inflammatory response and encouraging macrophage apoptosis. This suggests that by focusing on the temporary depletion or programmed cell death of macrophages may yield an effective strategy for preventing or alleviating chronic inflammatory injuries mediated by these immune cells, particularly those triggered by wear particles in periprosthetic tissue. Such an approach appears especially beneficial during the early stages of osteolysis development, where managing the inflammatory response and apoptosis of stress-damaged macrophages could significantly influence the progression of tissue damage.
The successful pharmacological intervention employing chemical chaperones to prevent wear particle-induced inflammatory injury in macrophages not only underscores the critical role of macrophages in the pathological process of aseptic loosening but also highlights potential therapeutic strategies for achieving inflammation-free apoptosis in the management of osteolysis. The implications of this research are profound, as it paves the way for developing innovative treatment strategies that could target macrophage viability, ultimately improving patient outcomes in conditions related to osteolysis. Further exploration of 4-PBA as a potential therapeutic alternative holds considerable promise for enhancing the biocompatibility of artificial implants and minimizing the occurrence of osteolysis by addressing the challenges posed by particle-induced macrophage-mediated inflammation and apoptosis.
Author Contributions
Conceptualization, Guoyin Liu, Ruigang Jia, Ying Xue and Peng He; Data curation, Jie Ding, Yahui Fu and Jingyan Mao; Formal analysis, Tianting Bai and Minghai Wu; Funding acquisition, Guoyin Liu and Ying Xue; Investigation, Guoyin Liu; Methodology, Tianting Bai; Project administration, Jianmin Chen and Peng He; Resources, Jie Ding, Jianmin Chen, Ruigang Jia and Ying Xue; Software, Minghai Wu, Jingyan Mao and Peng He; Validation, Guoyin Liu, Tianting Bai, Minghai Wu, Ruigang Jia, Ying Xue and Peng He; Visualization, Peng He; Writing—original draft, Guoyin Liu; Writing—review & editing, Tianting Bai, Jie Ding, Yahui Fu and Ruigang Jia. All authors reviewed the manuscript.
Funding
This work was supported by National Natural Science Foundation of China (NSFC. 82102547, awarded to Guoyin Liu), Key Grant Project of Chinese Medicine Education Assciation (No. 2024KTM021, awarded to Guoyin Liu), Research Project of National Association of Health Industry and Enterprise Management (No. ZWB2023001, awarded to Guoyin Liu), and Nanjing Medical University Project Fund (NO. NMUB20220144, awarded to Ying Xue).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusion of this article will be made available from the first author.
Abbreviations
The following abbreviations are used in this manuscript:
| 4-PBA | Multidisciplinary Digital Publishing Institute |
| TiNP | Titanium Nanoparticle |
| TiNPs | Titanium Nanoparticles |
| ERS | Endoplasmic Reticulum Stress |
| ER | Endoplasmic Reticulum |
| ROS | Reactive Oxygen Species |
| UPR | Unfolded Protein Response |
| UCDs | Urea Cycle Disorders |
References
- Gibon, E.; Amanatullah, D.F.; Loi, F.; Pajarinen, J.; Nabeshima, A.; Yao, Z.; Hamadouche, M.; Goodman, S.B. The biological response to orthopaedic implants for joint replacement: Part I: Metals. J. Biomed. Mater. Res. Part B 2017, 105, 2162–2173. [Google Scholar] [CrossRef]
- Gallo, J.; Vaculova, J.; Goodman, S.B.; Konttinen, Y.T.; Thyssen, J.P. Contributions of human tissue analysis to understanding the mechanisms of loosening and osteolysis in total hip replacement. Acta Biomater. 2014, 10, 2354–2366. [Google Scholar] [CrossRef]
- Goodman, S.B.; Gallo, J. Periprosthetic Osteolysis: Mechanisms, Prevention and Treatment. J. Clin. Med. 2019, 8. [Google Scholar] [CrossRef]
- Chavan, A.; Avula, I.; Sahoo, S.N.; Biswal, S.; Mandal, S.; Musthafa, M.; Roy, S.; Nandi, S.K.; Mukherjee, S.; Roy, M. Functional medium entropy alloys for joint replacement: An atomistic perspective of material deformation and a correlation to wear, corrosion, and biocompatibility. Acta Biomater. 2024, 187, 451–470. [Google Scholar] [CrossRef] [PubMed]
- Eger, M.; Hiram-Bab, S.; Liron, T.; Sterer, N.; Carmi, Y.; Kohavi, D.; Gabet, Y. Mechanism and Prevention of Titanium Particle-Induced Inflammation and Osteolysis. Front. Immunol. 2018, 9, 2963. [Google Scholar] [CrossRef]
- Panez-Toro, I.; Heymann, D.; Gouin, F.; Amiaud, J.; Heymann, M.F.; Cordova, L.A. Roles of inflammatory cell infiltrate in periprosthetic osteolysis. Front. Immunol. 2023, 14, 1310262. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Chen, S.; Wang, J.; Liang, S.; Chen, M.; Liu, Q.; Zhang, Z.; Liu, G.; Yang, Y.; Hu, Y.; et al. Enhanced Osteolysis Targeted Therapy through Fusion of Exosomes Derived from M2 Macrophages and Bone Marrow Mesenchymal Stem Cells: Modulating Macrophage Polarization. Small 2024, 20, e2303506. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Gong, G.; Liu, X.; Yin, J. Mechanism of regulating macrophages/osteoclasts in attenuating wear particle-induced aseptic osteolysis. Front. Immunol. 2023, 14, 1274679. [Google Scholar] [CrossRef]
- Hodges, N.A.; Sussman, E.M.; Stegemann, J.P. Aseptic and septic prosthetic joint loosening: Impact of biomaterial wear on immune cell function, inflammation, and infection. Biomaterials 2021, 278, 121127. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, G.; Ha, D.P.; Wang, J.; Xiong, M.; Lee, A.S. ER chaperone GRP78/BiP translocates to the nucleus under stress and acts as a transcriptional regulator. Proc. Natl. Acad. Sci. U. S. A. 2023, 120, e2303448120. [Google Scholar] [CrossRef]
- Liu, G.; Wu, J.; Wang, Y.; Xu, Y.; Xu, C.; Fang, G.; Li, X.; Chen, J. The Differential Expressions and Associations of Intracellular and Extracellular GRP78/Bip with Disease Activity and Progression in Rheumatoid Arthritis. Bioengineering-Basel 2025, 12. [Google Scholar] [CrossRef]
- Jeon, J.H.; Im, S.; Kim, H.S.; Lee, D.; Jeong, K.; Ku, J.M.; Nam, T.G. Chemical Chaperones to Inhibit Endoplasmic Reticulum Stress: Implications in Diseases. Drug Des Devel Ther 2022, 16, 4385–4397. [Google Scholar] [CrossRef]
- Liu, G.; Gong, H.; Bai, T.; Fu, Y.; Li, X.; Lu, J.; Zhao, J.; Chen, J. Pharmacological Intervention with 4-Phenylbutyrate Ameliorates TiAl6V4 Nanoparticles-Induced Inflammatory Osteolysis by Promoting Macrophage Apoptosis. Bioengineering-Basel 2025, 12, 701. [Google Scholar] [CrossRef]
- Xu, Y.; Zheng, S.; Chen, B.; Wen, Y.; Zhu, S. Sodium phenylbutyrate antagonizes prostate cancer through the induction of apoptosis and attenuation of cell viability and migration. OncoTargets Ther. 2016, 9, 2825–2833. [Google Scholar] [CrossRef]
- Tang, Y.H.; Yue, Z.S.; Zheng, W.J.; Shen, H.F.; Zeng, L.R.; Hu, Z.Q.; Xiong, Z.F. 4-Phenylbutyric acid presents therapeutic effect on osteoarthritis via inhibiting cell apoptosis and inflammatory response induced by endoplasmic reticulum stress. Biotechnol. Appl. Biochem. 2018, 65, 540–546. [Google Scholar] [CrossRef] [PubMed]
- de Pablo, S.; Rodriguez-Comas, J.; Diaz-Catalan, D.; Alcarraz-Vizan, G.; Castano, C.; Moreno-Vedia, J.; Montane, J.; Parrizas, M.; Servitja, J.M.; Novials, A. 4-Phenylbutyrate (PBA) treatment reduces hyperglycemia and islet amyloid in a mouse model of type 2 diabetes and obesity. Sci. Rep. 2021, 11, 11878. [Google Scholar] [CrossRef] [PubMed]
- Lynn, E.G.; Lhotak, S.; Lebeau, P.; Byun, J.H.; Chen, J.; Platko, K.; Shi, C.; O’Brien, E.R.; Austin, R.C. 4-Phenylbutyrate protects against atherosclerotic lesion growth by increasing the expression of HSP25 in macrophages and in the circulation of Apoe(-/-) mice. Faseb. J. 2019, 33, 8406–8422. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Cuesta, M.; Herrera-Gonzalez, I.; Garcia-Moreno, M.I.; Ashmus, R.A.; Vocadlo, D.J.; Garcia Fernandez, J.M.; Nanba, E.; Higaki, K.; Ortiz Mellet, C. sp(2)-Iminosugars targeting human lysosomal beta-hexosaminidase as pharmacological chaperone candidates for late-onset Tay-Sachs disease. J. Enzym. Inhib. Med. Chem. 2022, 37, 1364–1374. [Google Scholar] [CrossRef]
- Arnold, S.E.; Hendrix, S.; Nicodemus-Johnson, J.; Knowlton, N.; Williams, V.J.; Burns, J.M.; Crane, M.; Mcmanus, A.J.; Vaishnavi, S.N.; Arvanitakis, Z.; et al. Biological effects of sodium phenylbutyrate and taurursodiol in Alzheimer’s disease. Alzheimers Dement.-Transl. Res. Clin. Interv. 2024, 10, e12487. [Google Scholar] [CrossRef]
- Bowser, R.; An, J.; Mehta, L.; Chen, J.; Timmons, J.; Cudkowicz, M.; Paganoni, S. Effect of sodium phenylbutyrate and taurursodiol on plasma concentrations of neuroinflammatory biomarkers in amyotrophic lateral sclerosis: results from the CENTAUR trial. J. Neurol. Neurosurg. Psychiatry. 2024, 95, 605–608. [Google Scholar] [CrossRef]
- Choi, Y.; Lee, E.G.; Jeong, J.H.; Yoo, W.H. 4-Phenylbutyric acid, a potent endoplasmic reticulum stress inhibitor, attenuates the severity of collagen-induced arthritis in mice via inhibition of proliferation and inflammatory responses of synovial fibroblasts. Kaohsiung J. Med. Sci. 2021, 37, 604–615. [Google Scholar] [CrossRef]
- Liu, G.; Liu, N.; Xu, Y.; Ti, Y.; Chen, J.; Chen, J.; Zhang, J.; Zhao, J. Endoplasmic reticulum stress-mediated inflammatory signaling pathways within the osteolytic periosteum and interface membrane in particle-induced osteolysis. Cell. Tissue. Res. 2016, 363, 427–447. [Google Scholar] [CrossRef]
- Liu, G.; Guo, T.; Zhang, Y.; Liu, N.; Chen, J.; Chen, J.; Zhang, J.; Zhao, J. Apoptotic pathways of macrophages within osteolytic interface membrane in periprosthestic osteolysis after total hip replacement. Apmis. 2017, 125, 565–578. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Z.; Ma, Y.; Liu, G.; Shi, H.; Chen, J.; Dong, L.; Zhao, J.; Zhang, J. Particle-induced osteolysis mediated by endoplasmic reticulum stress in prosthesis loosening. Biomaterials 2013, 34, 2611–2623. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, N.; Shi, T.; Zhou, G.; Wang, Z.; Gan, J.; Guo, T.; Qian, H.; Bao, N.; Zhao, J. ER Stress Mediates TiAl6V4 Particle-Induced Peri-Implant Osteolysis by Promoting RANKL Expression in Fibroblasts. PLoS One 2015, 10, e0137774. [Google Scholar] [CrossRef]
- Niemietz, I.; Brown, K.L. Hyaluronan promotes intracellular ROS production and apoptosis in TNFalpha-stimulated neutrophils. Front. Immunol. 2023, 14, 1032469. [Google Scholar] [CrossRef]
- Bijukumar, D.R.; Salunkhe, S.; Zheng, G.; Barba, M.; Hall, D.J.; Pourzal, R.; Mathew, M.T. Wear particles induce a new macrophage phenotype with the potential to accelerate material corrosion within total hip replacement interfaces. Acta Biomater. 2020, 101, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhang, W.; Lyu, Z.; Long, T.; Wang, Y. ZnO nanoparticles attenuate polymer-wear-particle induced inflammatory osteolysis by regulating the MEK-ERK-COX-2 axis. J. Orthop. Transl. 2022, 34, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gibon, E.; Cordova, L.A.; Lu, L.; Lin, T.H.; Yao, Z.; Hamadouche, M.; Goodman, S.B. The biological response to orthopedic implants for joint replacement. II: Polyethylene, ceramics, PMMA, and the foreign body reaction. J. Biomed. Mater. Res. Part B 2017, 105, 1685–1691. [Google Scholar] [CrossRef]
- Goodman, S.B.; Gallo, J.; Gibon, E.; Takagi, M. Diagnosis and management of implant debris-associated inflammation. Expert Rev. Med. Devices 2020, 17, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Billi, F.; Campbell, P. Nanotoxicology of metal wear particles in total joint arthroplasty: a review of current concepts. J Appl Biomater Biomech 2010, 8, 1–6. [Google Scholar]
- Dalal, A.; Pawar, V.; Mcallister, K.; Weaver, C.; Hallab, N.J. Orthopedic implant cobalt-alloy particles produce greater toxicity and inflammatory cytokines than titanium alloy and zirconium alloy-based particles in vitro, in human osteoblasts, fibroblasts, and macrophages. J. Biomed. Mater. Res. Part A 2012, 100, 2147–2158. [Google Scholar] [CrossRef]
- Ivanovski, S.; Bartold, P.M.; Huang, Y.S. The role of foreign body response in peri-implantitis: What is the evidence? Periodontol. 2000 2022, 90, 176–185. [Google Scholar] [CrossRef]
- Yang, F.; Wu, W.; Cao, L.; Huang, Y.; Zhu, Z.; Tang, T.; Dai, K. Pathways of macrophage apoptosis within the interface membrane in aseptic loosening of prostheses. Biomaterials 2011, 32, 9159–9167. [Google Scholar] [CrossRef] [PubMed]
- Catelas, I.; Petit, A.; Zukor, D.J.; Marchand, R.; Yahia, L.; Huk, O.L. Induction of macrophage apoptosis by ceramic and polyethylene particles in vitro. Biomaterials 1999, 20, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wang, Y.; Tang, Z.; Peng, X.; Li, C.; Dang, Y.; Ma, R. Calycosin alleviates titanium particle-induced osteolysis by modulating macrophage polarization and subsequent osteogenic differentiation. J. Cell. Mol. Med. 2024, 28, e18157. [Google Scholar] [CrossRef]
- Lin, S.; Wen, Z.; Li, S.; Chen, Z.; Li, C.; Ouyang, Z.; Lin, C.; Kuang, M.; Xue, C.; Ding, Y. LncRNA Neat1 promotes the macrophage inflammatory response and acts as a therapeutic target in titanium particle-induced osteolysis. Acta Biomater. 2022, 142, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Zaveri, T.D.; Dolgova, N.V.; Lewis, J.S.; Hamaker, K.; Clare-Salzler, M.J.; Keselowsky, B.G. Macrophage integrins modulate response to ultra-high molecular weight polyethylene particles and direct particle-induced osteolysis. Biomaterials 2017, 115, 128–140. [Google Scholar] [CrossRef]
- Connors, J.P.; Stelzer, J.W.; Garvin, P.M.; Wellington, I.J.; Solovyova, O. The Role of the Innate Immune System in Wear Debris-Induced Inflammatory Peri-Implant Osteolysis in Total Joint Arthroplasty. Bioengineering-Basel 2022, 9. [Google Scholar] [CrossRef]
- Xie, Y.; Peng, Y.; Fu, G.; Jin, J.; Wang, S.; Li, M.; Zheng, Q.; Lyu, F.J.; Deng, Z.; Ma, Y. Nano wear particles and the periprosthetic microenvironment in aseptic loosening induced osteolysis following joint arthroplasty. Front. Cell. Infect. Microbiol. 2023, 13, 1275086. [Google Scholar] [CrossRef]
- Deans, C.F.; Buckner, B.C.; Garvin, K.L. Wear, Osteolysis, and Aseptic Loosening Following Total Hip Arthroplasty in Young Patients with Highly Cross-Linked Polyethylene: A Review of Studies with a Follow-Up of over 15 Years. J. Clin. Med. 2023, 12. [Google Scholar] [CrossRef]
- Marmotti, A.; Messina, D.; Cykowska, A.; Beltramo, C.; Bellato, E.; Colombero, D.; Agati, G.; Mangiavini, L.; Bruzzone, M.; Dettoni, F.; et al. Periprosthetic osteolysis: a narrative review. J. Biol. Regul. Homeost. Agents. 2020, 34, 405–417. [Google Scholar]
- Eger, M.; Hiram-Bab, S.; Liron, T.; Sterer, N.; Carmi, Y.; Kohavi, D.; Gabet, Y. Mechanism and Prevention of Titanium Particle-Induced Inflammation and Osteolysis. Front. Immunol. 2018, 9, 2963. [Google Scholar] [CrossRef]
- Landgraeber, S.; von Knoch, M.; Loer, F.; Wegner, A.; Tsokos, M.; Hussmann, B.; Totsch, M. Extrinsic and intrinsic pathways of apoptosis in aseptic loosening after total hip replacement. Biomaterials 2008, 29, 3444–3450. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, N.; Liu, K.; Zhou, G.; Gan, J.; Wang, Z.; Shi, T.; He, W.; Wang, L.; Guo, T.; et al. Autophagy mediated CoCrMo particle-induced peri-implant osteolysis by promoting osteoblast apoptosis. Autophagy 2015, 11, 2358–2369. [Google Scholar] [CrossRef]
- Sun, K.; Li, Y.; Lu, Z.; Zhang, L.; Gao, Z.; Jin, Q. Suppression of titanium particle-induced TNF-alpha expression and apoptosis in human U937 macrophages by siRNA silencing. Int. J. Artif. Organs. 2013, 36, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Fang, Y.; Wang, Q.; Sun, Y.; Xiong, C.; Cao, L.; Wang, B.; Bao, N.; Zhao, J. Tumor necrosis factor-like weak inducer of apoptosis regulates particle-induced inflammatory osteolysis via the p38 mitogen-activated protein kinase signaling pathway. Mol. Med. Rep. 2015, 12, 1499–1505. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Wu, Y.; Tang, Y.; Ge, Z.; Wang, D.; Zheng, C.; Zhao, R.; Lin, W.; Wang, G. Microenvironment-responsive smart hydrogels with antibacterial activity and immune regulation for accelerating chronic wound healing. J. Control. Release 2024, 368, 518–532. [Google Scholar] [CrossRef]
- Riwaldt, S.; Corydon, T.J.; Pantalone, D.; Sahana, J.; Wise, P.; Wehland, M.; Kruger, M.; Melnik, D.; Kopp, S.; Infanger, M.; et al. Role of Apoptosis in Wound Healing and Apoptosis Alterations in Microgravity. Front. Bioeng. Biotechnol. 2021, 9, 679650. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Antonini, J.M.; Rojanasakul, Y.; Castranova, V.; Scabilloni, J.F.; Mercer, R.R. Potential role of apoptotic macrophages in pulmonary inflammation and fibrosis. J. Cell. Physiol. 2003, 194, 215–224. [Google Scholar] [CrossRef]
- Purohit, V.; Brenner, D.A. Mechanisms of alcohol-induced hepatic fibrosis: a summary of the Ron Thurman Symposium. Hepatology. 2006, 43, 872–878. [Google Scholar] [CrossRef]
- Yu, X.; Ding, H.; Wang, D.; Ren, Z.; Chen, B.; Wu, Q.; Yuan, T.; Liu, Y.; Zhang, L.; Zhao, J.; et al. Particle-induced osteolysis is mediated by endoplasmic reticulum stress-associated osteoblast apoptosis. Chem. Biol. Interact. 2023, 383, 110686. [Google Scholar] [CrossRef]
- Yu, X.; Hu, J.; Yang, X.; Xu, Q.; Chen, H.; Zhan, P.; Zhang, B. Sesamin inhibits RANKL-induced osteoclastogenesis and attenuates LPS-induced osteolysis via suppression of ERK and NF-kappaB signalling pathways. J. Cell. Mol. Med. 2024, 28, e18056. [Google Scholar] [CrossRef]
- Zhang, L.; Bao, D.; Li, P.; Lu, Z.; Pang, L.; Chen, Z.; Guo, H.; Gao, Z.; Jin, Q. Particle-induced SIRT1 downregulation promotes osteoclastogenesis and osteolysis through ER stress regulation. Biomed. Pharmacother. 2018, 104, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tian, Z.; Li, W.; Wang, X.; Man, Z.; Sun, S. Inhibitory effect of quercetin on titanium particle-induced endoplasmic reticulum stress (ERS)-related apoptosis and in vivoosteolysis. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Endoplasmic reticulum stress and the inflammatory basis of metabolic disease. Cell 2010, 140, 900–917. [Google Scholar] [CrossRef]
- Mostoufi, A.; Baghgoli, R.; Fereidoonnezhad, M. Synthesis, cytotoxicity, apoptosis and molecular docking studies of novel phenylbutyrate derivatives as potential anticancer agents. Comput. Biol. Chem. 2019, 80, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Alqallaf, A.; Cates, D.W.; Render, K.P.; Patel, K.A. Sodium Phenylbutyrate and Taurursodiol: A New Therapeutic Option for the Treatment of Amyotrophic Lateral Sclerosis. Ann. Pharmacother. 2024, 58, 165–173. [Google Scholar] [CrossRef]
- Tiwari, S.; Gupta, P.; Singh, A.; Chaturvedi, S.; Wahajuddin, M.; Mishra, A.; Singh, S. 4-Phenylbutyrate Mitigates the Motor Impairment and Dopaminergic Neuronal Death During Parkinson’s Disease Pathology via Targeting VDAC1 Mediated Mitochondrial Function and Astrocytes Activation. Neurochem. Res. 2022, 47, 3385–3401. [Google Scholar] [CrossRef]
- Iannitti, T.; Palmieri, B. Clinical and experimental applications of sodium phenylbutyrate. Drugs R&D 2011, 11, 227–249. [Google Scholar] [CrossRef]
- Stein, D.; Slobodnik, Z.; Tam, B.; Einav, M.; Akabayov, B.; Berstein, S.; Toiber, D. 4-phenylbutyric acid-Identity crisis; can it act as a translation inhibitor? Aging Cell 2022, 21, e13738. [Google Scholar] [CrossRef]
- Saito, Y.; Liang, G.; Egger, G.; Friedman, J.M.; Chuang, J.C.; Coetzee, G.A.; Jones, P.A. Specific activation of microRNA-127 with downregulation of the proto-oncogene BCL6 by chromatin-modifying drugs in human cancer cells. Cancer Cell 2006, 9, 435–443. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, L.; Yang, Y.; Yu, Q. Sodium 4-phenylbutyrate induces apoptosis of human lung carcinoma cells through activating JNK pathway. J. Cell. Biochem. 2004, 93, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.; Xu, W.; Reed, J.C. Cell death and endoplasmic reticulum stress: disease relevance and therapeutic opportunities. Nat. Rev. Drug Discov. 2008, 7, 1013–1030. [Google Scholar] [CrossRef] [PubMed]
- Kapuy, O.; Marton, M.; Banhegyi, G.; Vinod, P.K. Multiple system-level feedback loops control life-and-death decisions in endoplasmic reticulum stress. FEBS Lett. 2020, 594, 1112–1123. [Google Scholar] [CrossRef]
- Xu, C.; Bailly-Maitre, B.; Reed, J.C. Endoplasmic reticulum stress: cell life and death decisions. J. Clin. Invest. 2005, 115, 2656–2664. [Google Scholar] [CrossRef]
- Cao, Y.; Liu, Z.; Xiao, W.; Gu, Z.; Xiao, G.; Yuan, F.; Chen, F.; Pei, Y.; Li, H.; Su, L. 4-Phenylbutyrate Prevents Endoplasmic Reticulum Stress-Mediated Apoptosis Induced by Heatstroke in the Intestines of Mice. Shock 2020, 54, 102–109. [Google Scholar] [CrossRef]
- Goodman, S.B.; Gibon, E.; Gallo, J.; Takagi, M. Macrophage Polarization and the Osteoimmunology of Periprosthetic Osteolysis. Curr. Osteoporos. Rep. 2022, 20, 43–52. [Google Scholar] [CrossRef]
- Jiang, Z.; He, Q.; Wezeman, J.; Darvas, M.; Ladiges, W. A cocktail of rapamycin, acarbose, and phenylbutyrate prevents age-related cognitive decline in mice by targeting multiple aging pathways. GeroScience 2024, 46, 4855–4868. [Google Scholar] [CrossRef] [PubMed]
- Paganoni, S.; Macklin, E.A.; Hendrix, S.; Berry, J.D.; Elliott, M.A.; Maiser, S.; Karam, C.; Caress, J.B.; Owegi, M.A.; Quick, A.; et al. Trial of Sodium Phenylbutyrate-Taurursodiol for Amyotrophic Lateral Sclerosis. N. Engl. J. Med. 2020, 383, 919–930. [Google Scholar] [CrossRef]
Figure 1.
The impact of 4-PBA on cell viability and the inflammatory factors induced by TiNPs in macrophages. (A-D) The cytotoxic effects of varying concentrations of TiNPs (A), different exposure durations to TiNPs (B), 4-PBA alone (C), and the combination of 1000 µg/mL of TiNPs with varying concentrations of 4-PBA (D) on RAW-264.7 cells. (E-G) The expression levels of pro-inflammatory cytokines TNF-α (E), IL-1β (F), and IL-6 (G) in RAW-264.7 cells. (H) The production of ROS in RAW-264.7 cells. Data represent as the means ± SEM from three independent experiments. *: P<0.05, **: P<0.01 versus Control. #: P<0.05, ##: P<0.01 versus TiNPs.
Figure 1.
The impact of 4-PBA on cell viability and the inflammatory factors induced by TiNPs in macrophages. (A-D) The cytotoxic effects of varying concentrations of TiNPs (A), different exposure durations to TiNPs (B), 4-PBA alone (C), and the combination of 1000 µg/mL of TiNPs with varying concentrations of 4-PBA (D) on RAW-264.7 cells. (E-G) The expression levels of pro-inflammatory cytokines TNF-α (E), IL-1β (F), and IL-6 (G) in RAW-264.7 cells. (H) The production of ROS in RAW-264.7 cells. Data represent as the means ± SEM from three independent experiments. *: P<0.05, **: P<0.01 versus Control. #: P<0.05, ##: P<0.01 versus TiNPs.
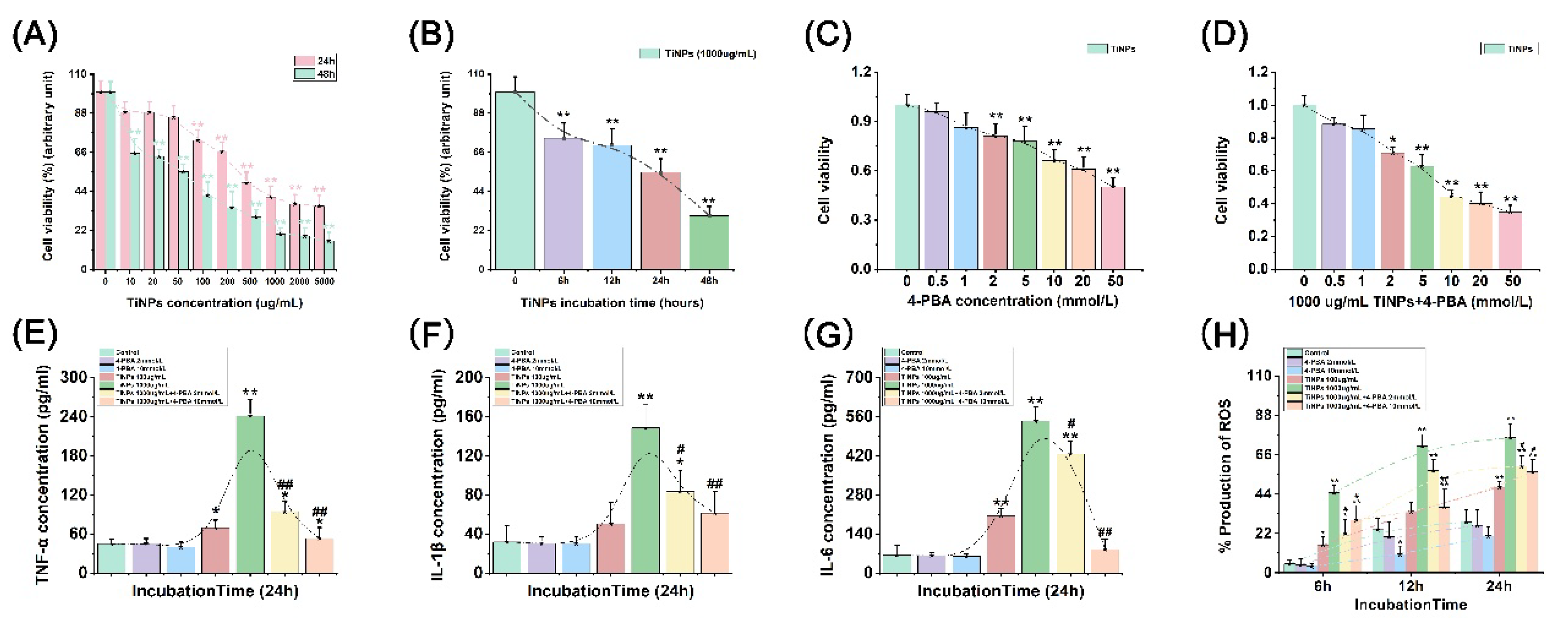
Figure 2.
The impact of 4-PBA on ROS production induced by TiNPs in macrophages. (A-C) The levels of ROS production following interventions at 6 hours (A), 12 hours (B), and 24 hours (C). Data represent as the means ± SEM from three independent experiments. *: P<0.05, **: P<0.01 versus Control. #: P<0.05, ##: P<0.01 versus TiNPs.
Figure 2.
The impact of 4-PBA on ROS production induced by TiNPs in macrophages. (A-C) The levels of ROS production following interventions at 6 hours (A), 12 hours (B), and 24 hours (C). Data represent as the means ± SEM from three independent experiments. *: P<0.05, **: P<0.01 versus Control. #: P<0.05, ##: P<0.01 versus TiNPs.
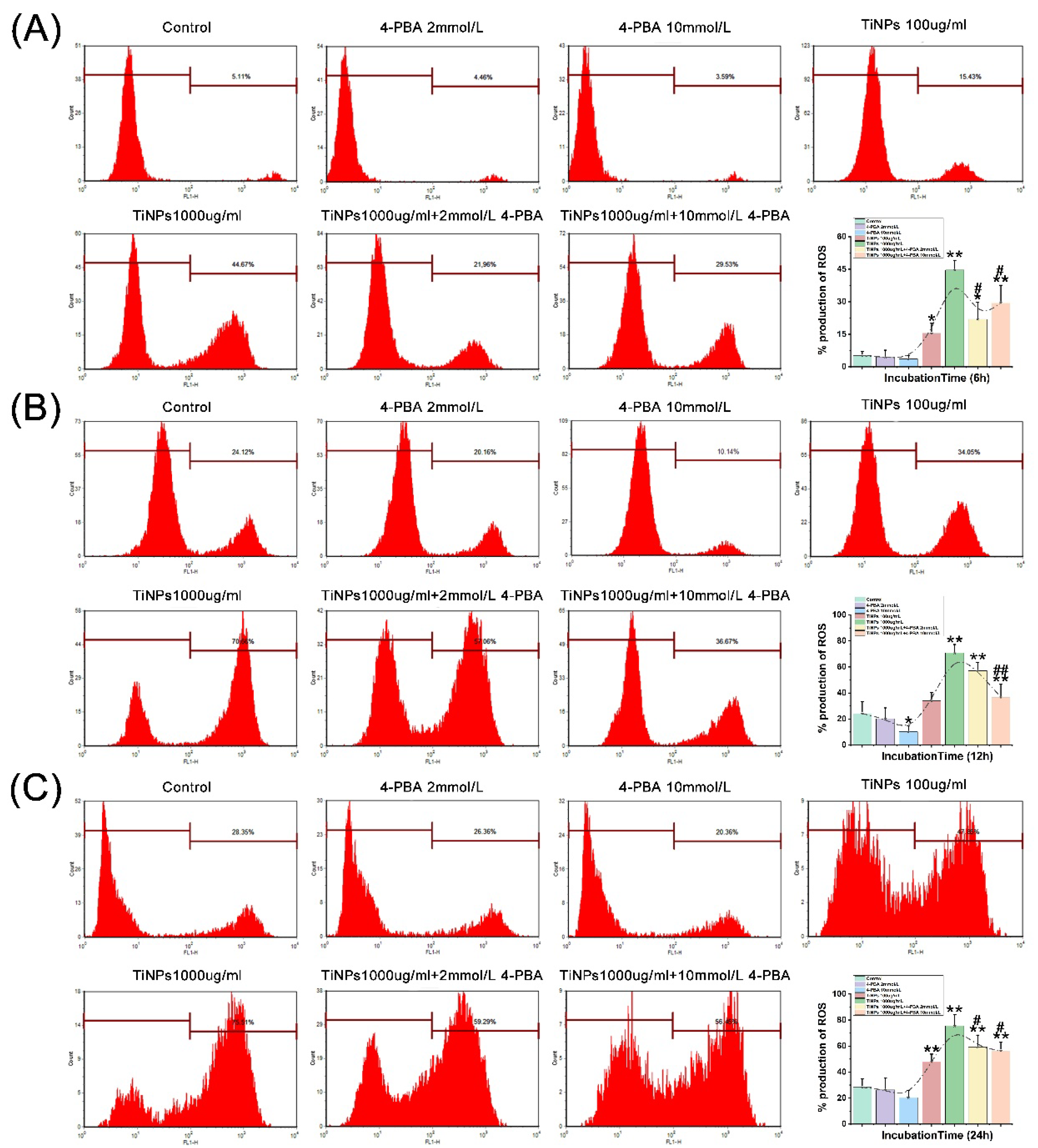
Figure 3.
The impact of 4-PBA on the apoptosis of macrophages induced by TiNPs. (A-C) The proportion of apoptotic cells following interventions at 6 hours (A), 12 hours (B), and 24 hours (C). Data represent as the means ± SEM from three independent experiments. *: P<0.05, **: P<0.01 versus Control. #: P<0.05, ##: P<0.01 versus TiNPs.
Figure 3.
The impact of 4-PBA on the apoptosis of macrophages induced by TiNPs. (A-C) The proportion of apoptotic cells following interventions at 6 hours (A), 12 hours (B), and 24 hours (C). Data represent as the means ± SEM from three independent experiments. *: P<0.05, **: P<0.01 versus Control. #: P<0.05, ##: P<0.01 versus TiNPs.
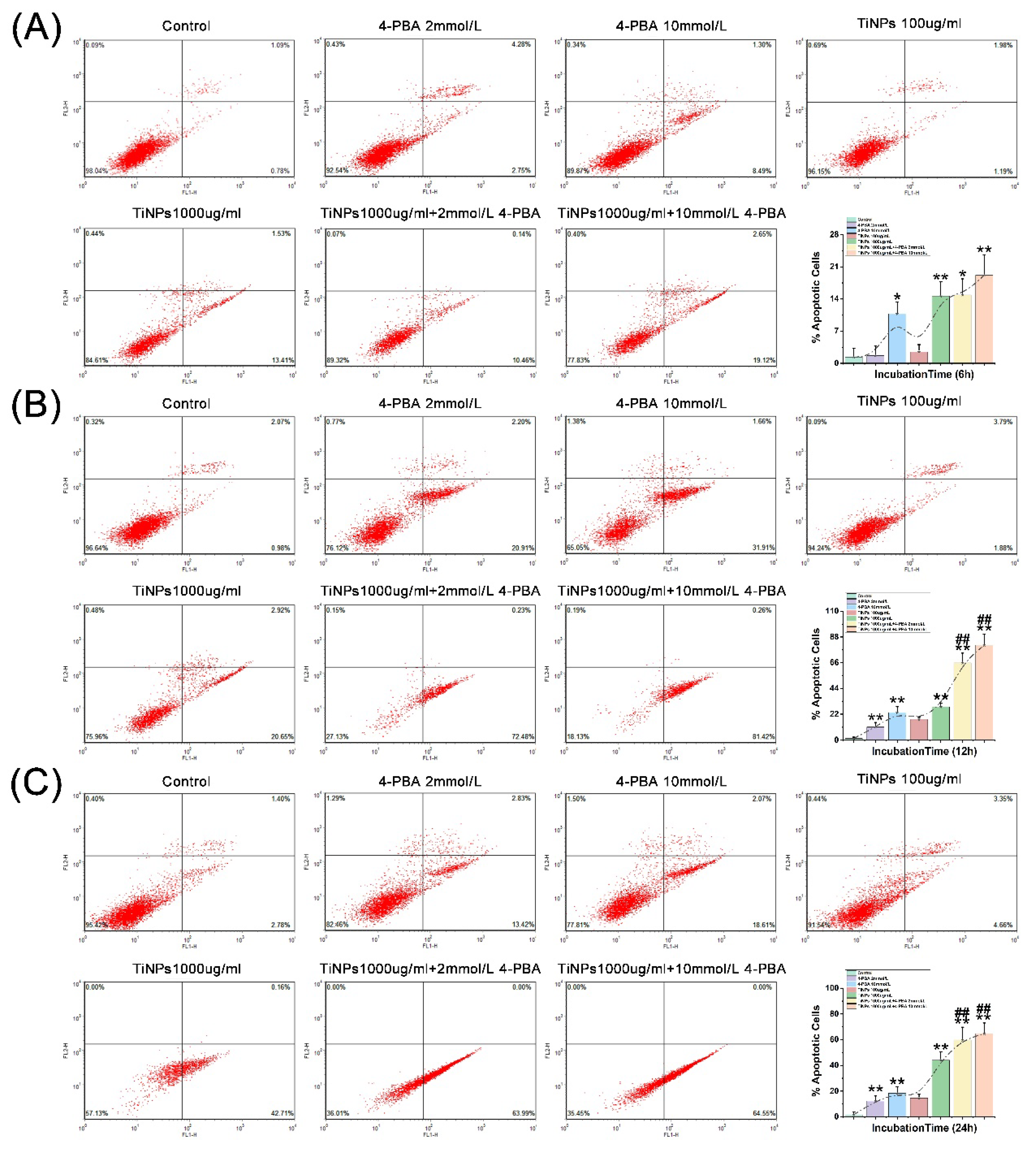
Figure 4.
The impact of 4-PBA on the apoptotic regulatory proteins induced by TiNPs in macrophages. (A) The apoptosis of macrophages after 6, 12 and 24 hours of interventions. (B-D) The levels of caspase-3 enzyme activity following interventions at 6 hours (B), 12 hours (C), and 24 hours (D). (E-H) Western blots results for Caspase-3, Bax, and Bcl-2 in macrophages. The density of the Western blot bands was quantified using ImageJ software. Data represent as the means ± SEM from three independent experiments. *: P<0.05, **: P<0.01 versus Control. #: P<0.05, ##: P<0.01 versus TiNPs.
Figure 4.
The impact of 4-PBA on the apoptotic regulatory proteins induced by TiNPs in macrophages. (A) The apoptosis of macrophages after 6, 12 and 24 hours of interventions. (B-D) The levels of caspase-3 enzyme activity following interventions at 6 hours (B), 12 hours (C), and 24 hours (D). (E-H) Western blots results for Caspase-3, Bax, and Bcl-2 in macrophages. The density of the Western blot bands was quantified using ImageJ software. Data represent as the means ± SEM from three independent experiments. *: P<0.05, **: P<0.01 versus Control. #: P<0.05, ##: P<0.01 versus TiNPs.
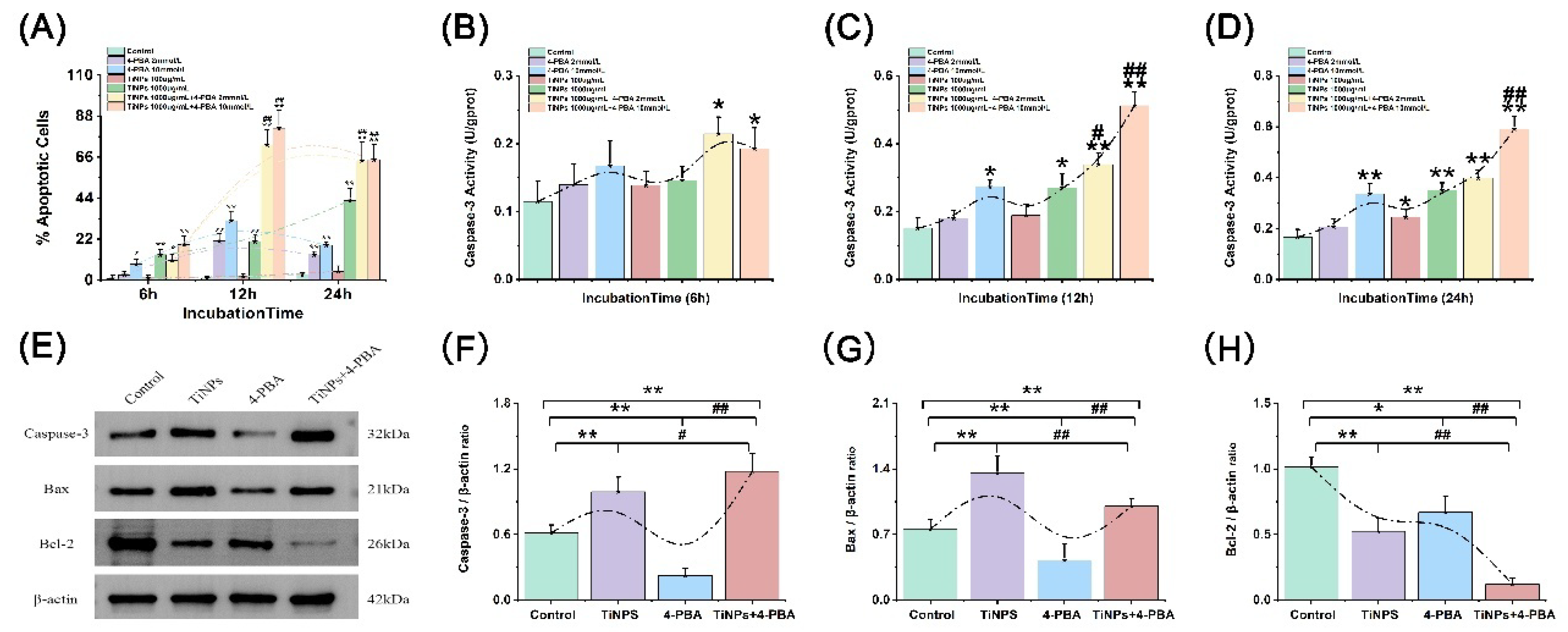
Figure 5.
4-PBA alleviates macrophage-mediated inflammatory injury induced by TiNPs by inhibiting the inflammatory response, and simultaneously facilitating the apoptosis of stress-damaged macrophages.
Figure 5.
4-PBA alleviates macrophage-mediated inflammatory injury induced by TiNPs by inhibiting the inflammatory response, and simultaneously facilitating the apoptosis of stress-damaged macrophages.
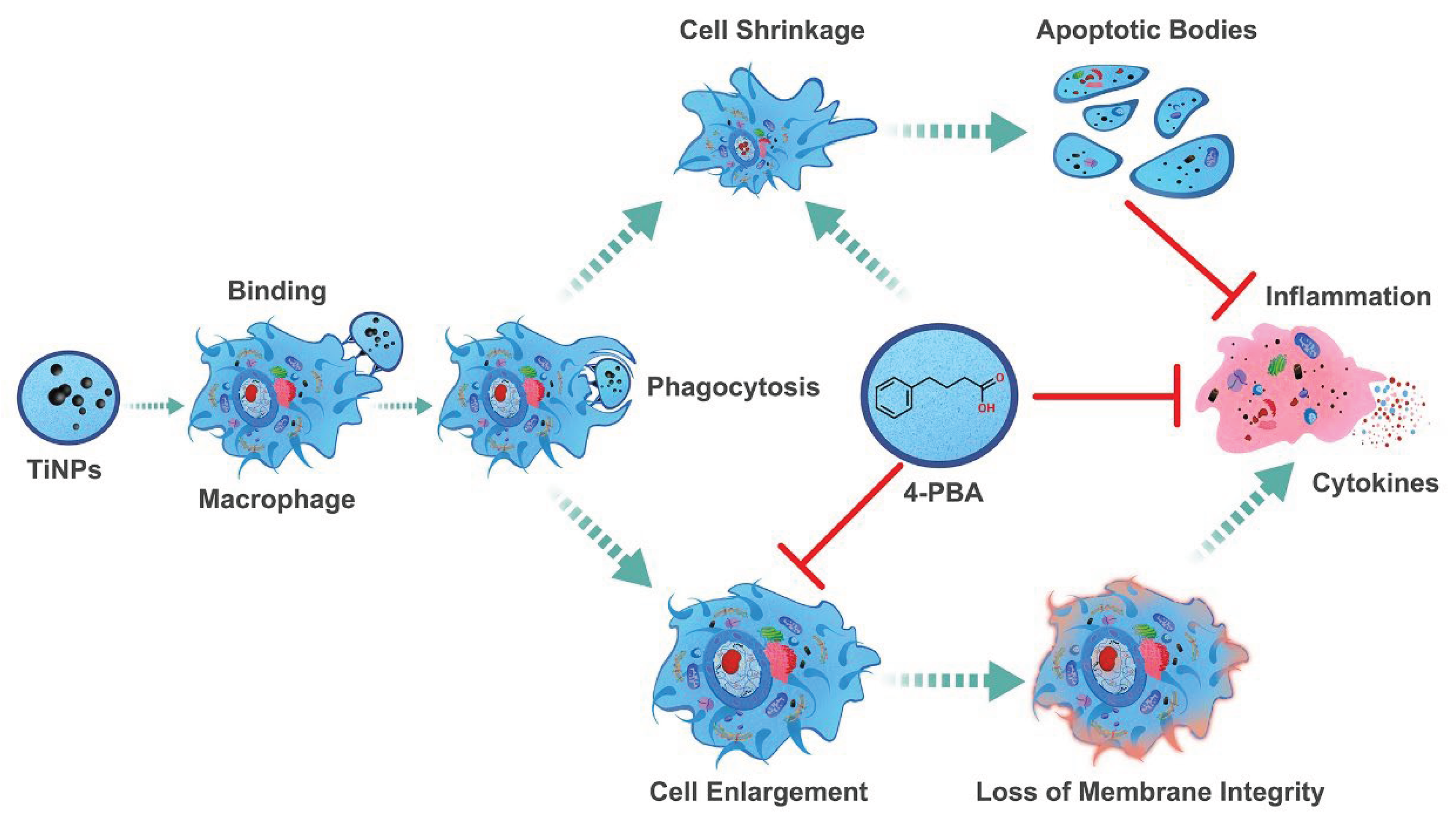
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



