
Submitted:
14 November 2025
Posted:
17 November 2025
You are already at the latest version
Abstract
Plankton community respiration (PCR) plays a central role in aquatic ecosystems, driving the breakdown of organic matter and influencing global carbon cycling through its contribution to the production and consumption of carbon and oxygen. Coastal areas, which serve as critical interfaces between terrestrial and marine ecosystems, are regarded as metabolic hotspots in the oceans, due to their intense biological and biogeochemical activities. Additionally, they are particularly sensitive to the impacts of global climate change. In this regard, this review synthesizes experimental evidence to explore how environmental constraints and climate drivers affect PCR in European coastal waters. In total, 46 studies were found in which PCR was measured during experiments testing the effects of one or multiple global climate change drivers in European coastal waters. Among them, the majority of experiments focused on changes in temperature, nutrient concentrations and stoichiometry, and/or pH, while other stressors were less studied. Many experiments confirmed theoretical predictions, notably regarding the predicted positive effects of increased temperature and nutrient concentrations on metabolism, but more complex responses, often linked to trophic cascade mechanisms and thresholds between positive and negative feedbacks were also often reported. Overall, this review, the first comprehensive synthesis of experimental evidence on PCR in European coastal waters, highlights critical knowledge gaps, notably regarding non- and understudied areas and understudied interactions between stressors that occurs jointly in natural ecosystems. Future research should aim to integrate controlled experiments, long-term monitoring, and modeling approaches to deepen our understanding of PCR dynamics under changing environmental conditions and to predict potential feedbacks to global climate processes.
Keywords:
plankton
; respiration
; climate change
; experiments
1. Introduction
Plankton community respiration (PCR) plays a fundamental role in marine ecosystems by driving the breakdown of organic matter and facilitating carbon cycling. As both autotrophic and heterotrophic plankton organisms metabolize organic compounds, they consume dissolved oxygen (O2) and release carbon dioxide (CO2) through aerobic respiration (Robinson and Williams, 2005; Robinson 2019). In contrast, phytoplankton, as the primary producers, convert dissolved CO2 into organic matter via photosynthesis, producing dissolved oxygen (O2) as a by-product (Falkowski 1994). These metabolic activities influence the overall efficiency of the ocean’s biological pump, a critical mechanism through which carbon is sequestered from the atmosphere and stored in deep ocean waters, mitigating global climate change (Field et al., 1998; Jiao and Azam 2011). Indeed, the balance between plankton production and respiration regulates carbon fluxes within marine ecosystems, and indicates whether a system acts as a net source of O2, and so a net sink of CO2, for the atmosphere, or if, conversely, the system acts as a net sink of O2, and a net source of CO2 for the atmosphere (López-Urrutia et al., 2006; Serret et al., 2015). Therefore, PCR is a vital component in determining how coastal and open ocean waters act as carbon sinks or sources. Given the sensibility of planktonic processes to environmental changes (Hays et al., 2005; Litchman et al., 2012), studying PCR and its influence on global biogeochemical cycling appears to be crucial, particularly in the context of global climate change and increased anthropogenic pressures on aquatic ecosystems.
Coastal waters are among the most dynamic and productive regions of the ocean, characterized by complex interactions between terrestrial and marine environments (Cloern et al., 2014; Carstensen et al., 2015). These areas are influenced by a variety of local environmental conditions, such as freshwater inflows from rivers (Fredston-Hermann et al., 2016), nutrient loading from agricultural and terrestrial runoff (Beman et al., 2005), and physical factors like tides, currents, and wind-driven mixing (Lizon et al., 1995). These interactions create highly variable environments where physical, chemical, and biological conditions can shift rapidly. Coastal waters are also particularly sensitive to global climate change drivers (Harley et al., 2006; Rabalais et al., 2009). Notably, warming temperatures, ocean acidification, increasing extreme weather event frequency, rising sea levels and changing precipitation patterns, resulting in intensified freshwater and nutrient inputs from land, are altering the structure and function of coastal ecosystems (Harley et al., 2006). Finally, coastal waters are key sites of biogeochemical cycling and biodiversity, as their metabolism contributes more to the global biogeochemical cycles than spatially larger but less productive regions of the open ocean, such as oligotrophic gyres (Aranguren-Gassis et al., 2013; Serret et al., 2015). Therefore, understanding how these dynamic systems respond to both local pressures and global climate change is crucial for predicting future impacts on global biogeochemical cycles and the resilience of marine environments.
Plankton communities are influenced by many of the environmental factors to which coastal waters are sensitive (e.g., Chou et al., 2012; Forsblom et al., 2019). Therefore, PCR can be highly variable in coastal waters, displaying strong seasonal variations and relationships to environmental conditions (e.g., Agusti et al., 2018; García-Martín et al., 2019a; Gomez-Castillo et al., 2023; Mantikci et al., 2024; Prichett et al., 2024) and plankton community biomass (Olesen et al., 1999; Lozano et al., 2021) and structure (Bas-Silvestre et al., 2024). In this regard, coastal PCR may be particularly prone to climate change and anthropogenic pressures (del Giorgio and Williams, 2005). However, while monitoring primary production is relatively straightforward, resulting in hundreds of thousands of observations since half a decade and primary production being well-parametrized and integrated into Earth system models (Laufkötter et al., 2015), monitoring PCR remains challenging. Budget methods, which rely on in situ changes in dissolved O2 concentration, require expensive and carefully calibrated equipment and complex calculations (Staehr et al., 2010; Soulié et al., 2021). Direct methods, such as in vitro incubations during which dissolved O2 concentration changes are measured are labor-intensive and extremely time-consuming, and are susceptible to manipulation biases and “bottle effects” (Robinson and Williams, 2005; García-Martín et al., 2011). Consequently, PCR is rarely monitored, and the few available observations are often sparse and not always fully reliable (Arístegui and Harrison, 2002; Regaudie-de-Gioux and Duarte, 2013). This has led to a limited understanding, both temporally and spatially, of climate change impacts on PCR, as well as the parameterization of its drivers and their incorporation into predictive models (Wikner et al., 2023). Accordingly, respiration and its related processes have been listed as a highly important knowledge gap in quantifying the effect of climate change on biological carbon storage in the ocean (Grégoire et al., 2023; Henson et al., 2024).
In this context, controlled-environment experiments could play a valuable role in elucidating the mechanisms behind observed trends (Wernberg et al., 2012). While direct measurements are crucial for capturing the complexity of natural systems, experiments remain essential to verify and parameterize causal relationships between key components in coastal ecosystems. Observations typically highlight correlations between environmental conditions and biological processes but cannot confirm causality. Experiments, on the other hand, can uncover underlying mechanisms, though they inevitably simplify the complexities of the systems being studied (Stewart et al., 2013).
In regard of the above, the objective of this review is to provide a comprehensive synthesis of experimental research, examining how environmental constraints and climate change drivers shape PCR in European coastal ecosystems. By focusing on results from controlled experiments, this review aims to elucidate the mechanistic relationships between PCR and environmental factors such as temperature, nutrient availability, and light penetration, alongside climate stressors like ocean warming, acidification, and marine heatwaves. Through an analysis of experimental findings, the review will highlight how these environmental drivers interact to influence PCR, revealing regional patterns and system-specific responses. This experimental perspective will provide critical insights into how future climate conditions may reshape metabolic processes in coastal waters, while identifying key gaps in the studied disturbances and areas and proposing directions for future research to advance our understanding of PCR in the context of global change in Europe.
2. Methods
As this review is focusing on the experimental evaluations of the effects of global climate change on PCR, it is first necessary to define certain terms used in this review. The term "global climate change" refers here to the shifts in global climate driven by anthropogenic emissions of CO2 and other greenhouse gases, along with its impacts on climatic, oceanographic, and biogeochemical processes. Such impacts encompass global phenomena, such as the ongoing rise in water temperature and decrease in pH, as well as extreme weather phenomena, such as the increase in frequency of marine heatwaves and terrestrial runoffs. In addition, some environmental drivers reflect both global climate change and direct anthropogenic pressures, and are included in the present review. For example, nutrient concentrations and stoichiometry are related to both global climate change (e.g., changes in upwelling processes due to climate change) and direct anthropogenic pressures (e.g., eutrophication due to agricultural waste runoffs). However, stressors related only to anthropogenic activities (e.g., inputs of pesticides) were not taken into account in the present review as they are not directly linked to global climate change.
In addition, this review focusses on scientific peer-reviewed publications. In one instance, the abstract from a presentation at an international conference was used (Serret et al., 2024), as it dealt with a very understudied process (ocean alkalinization) and I was able to access the data from the main author. Additionally, this review only focuses on experiments in which PCR was measured. No limits in the volume of the experimental units were considered, as long as the respiration rate of natural communities was measured. Hence, the experiments in which only one biological compartment (e.g., only phytoplankton) was included were not considered in the present review. Similarly, to ensure that a maximum of studies was considered in the review, no limits were taken into account regarding how PCR was measured, i.e., studies with different approaches and different methodologies (incubations vs. sensor-based). However, it should be noted that both the volume of the experimental units and the method used to measure PCR could play an important role in the comparison of the results between studies. Nonetheless, the goal of the present review was not to try to explain potential discrepancies in results related to methodological questions, but rather to give a global overview on the global climate change effects on PCR in coastal European waters.
Finally, the present review focusses on coastal waters of Europe. Particularly, coastal waters were not here defined from a certain maximum distance from the shore, but rather as marine waters under some influence from the land. As a result, some experiments were included in the present review while they were performed in offshore environments (e.g., in the Tyrrhenian and Ionian Seas, Gazeau et al., 2021) because they specifically tested disturbances arising from the land, e.g., inputs of Saharan dust). Additionally, Europe was considered here only as the geographical region.
In line with all of this, the search for relevant articles was done mainly using the Web of Science (WoS) website (https://apps.webofknowledge.com/, last access: 15 March 2025) maintained by Clarivate. The search was done using several terms in various combinations. These terms included “plankton respiration”, “climate change”, “global climate change”, “warming”, “temperature”, “acidification”, “pCO2”, “runoff”, “light”, “nutrients”, “ultraviolet”, “UV”, “salinity”, “alkalinization”, “metabolism”, “plankton metabolism”, “ecosystem metabolism”, “experiments”, “coastal waters”. Some papers from 2024 and 2025 were found with an unstructured search in Google Scholar (https://scholar.google.com/, last access: 11 November 2025), as this website includes more recent articles than WoS.
These various searches resulted in over 100 papers, of which many were not suitable for the present review, as they do not explicitly present PCR data, and/or were not focused on the effects of global climate change, and/or did not present results from experiments, and/or were not performed with coastal European waters. All the relevant studies were saved in a specific Zotero (https://www.zotero.org/) library. At the end, 46 studies meeting the review criteria were included. A table summarizing the main information regarding these studies can be found in Table 1.
3. Global Change Effects on Plankton Community Respiration
3.1. Temperature Sensitivity and Effect of Warming and Marine Heatwaves
Temperature is a primary driver of metabolic rates, with a direct relationship between temperature and metabolism as described by the Arrhenius Law and which serves as the foundation for the Metabolic Theory of Ecology (MTE) (Brown et al., 2004). The MTE uses the fundamental principles of the Arrhenius Law to explain how temperature regulates metabolic processes in ecosystems, influencing ecological functions across all levels of organization (Yvon-Durocher et al., 2012; Boscolo-Galazzo et al., 2018). According to this theory, PCR is expected to rise with increasing temperature. This was verified at the global ocean scale by Regaudie-de-Gioux and Duarte (2012) and Garcia-Corral et al. (2017), who compiled a large dataset of PCR measurements in various regions of the ocean and found a temperature sensitivity of PCR in accordance with the predictions of the MTE. However, this analysis is only based on concurrent measurements of PCR and water temperature, preventing from disentangling the variability in PCR related to temperature to that related to other concomitant environmental forcings. Indeed, temperature effects on PCR can also be nonlinear, as, for example, extreme heat can exceed the thermal tolerance of plankton communities (Chen 2015), disrupting ecosystem functioning. It can especially be the case in coastal areas, where many factors, and notably exchanges with the sediment and the land, can simultaneously affect plankton communities in addition to temperature. Accordingly, some experimental studies showed contrasted effects of changes in temperature on PCR in European coastal waters, sometimes even in contradiction to the predictions of the PCR (Vidussi et al., 2011; Soulié et al., 2022a).
Warming of the sea surface, either in the form of long-term warming (IPCC 2019; Cheng et al., 2022) or episodic marine heatwaves (Frölicher et al., 2018), is one the most known consequences of global climate change for the oceans (Smith et al., 2023), and also one of the most studied stressors regarding PCR responses toward global climate change (e.g., Cavan and Boyd, 2018; Latorre et al., 2023). Coastal waters are considered highly vulnerable to global warming, with sea surface temperatures projected to rise by 3°C by the end of the century in some regions (IPCC 2019), and particularly threatened by marine heatwaves (Garrabou et al., 2022; Pastor and Khodayar, 2023; Bashiri et al., 2024; Darmaraki et al., 2024). In this regard, many studies have experimentally investigated the effect of increased temperature on PCR in coastal waters. Albeit having used different experimental set-ups and studied different systems and communities, most of them highlighted a positive response of coastal PCR to warming, notably in the North Western Mediterranean Sea (Thau Lagoon (Soulié et al., 2022b; 2023), Bay of Blanes (Vázquez-Domínguez et al., 2007), and Majorca (Vaquer-Sunyer and Duarte, 2013)), in the Baltic Sea (Kiel Bight (Hoppe et al., 2008; Breithaupt 2009; Wohlers et al., 2009), Öre estuary (Panigrahi et al., 2013), South East Baltic Sea (Vaquer-Sunyer et al., 2015)), in the North East Atlantic Ocean (Bay of Biscay (Huete-Stauffer et al., 2018), Irish Sea (Lefèvre et al., 1994), Ría de Vigo (López-Sandoval et al., 2025; Soulié et al., 2025)), and in the Arctic (Svalbard archipelago (Vaquer-Sunyer et al., 2010; Holding et al., 2013; Wolf et al., 2024)). All the results from these studies tend therefore to confirm the predictions of the MTE, even if they were conducted in complex coastal environments.
However, other studies reported a lack of response, or even a negative effect of warming on PCR, thus contradicting with theoretical predictions and global observations. In the Mediterranean coastal Thau Lagoon, Vidussi et al. (2011) reported no significant effect of a +3°C increase on PCR during a in situ mesocosm experiment in spring, and suggested a trophic cascade induced by warming and favoring microzooplankton grazing on bacteria, ultimately reducing bacterial abundance and contribution to PCR. In the same location and with similar mesocosm set-ups, Soulié et al. (2022a) reported a strong negative response of PCR to a +3°C warming related to a 50% decrease in phytoplankton biomass due to enhanced grazing under elevated temperature during a spring bloom, and no significant effect of +3°C on PCR during of the same year, supposedly due to enhanced competition between heterotrophic bacteria and cyanobacteria under warmed conditions. In another Mediterranean coastal lagoon (Cabras Lagoon, Italy), Pulina et al. (2020) reported a decrease in PCR under +3°C and +6°C conditions, which could also have been related to warming effects on heterotrophic bacteria and their grazers. In addition, Maugendre et al. (2015) found no significant effect of +3°C temperature increase on PCR in the Bay of Villefranche, NW Mediterranean Sea, potentially due to nutrient-depleted conditions preventing the response of PCR. In the Baltic Sea, +3°C warming, applied either constantly or with fluctuating pulses, was shown to decrease PCR Sea (Cabrerizo et al., 2021), certainly due to a strong decrease in phytoplankton biomass under warmed conditions. Lewandowska et al. (2014) also reported no significant effect of +3°C warming on Baltic Sea PCR, and suggested a potential trophic cascade mechanism which could have prevented from a stronger response of PCR. Overall, the results from the above-mentioned studies highlight the importance of warming indirect effects, such as changes in the trophic structure of the plankton communities and in nutrient availabilities, which could override the direct positive response of metabolism toward warming in complex systems such as coastal environments. This stresses out the need of experimental studies which incorporates the full natural complexity of coastal ecosystems, and suggests that indirect effects of warming, often neglected, are taken into consideration in global models of the effects of climate change on PCR.
3.2. pH Sensitivity and Effect of Acidification
Ocean acidification, driven by the increasing absorption of atmospheric CO₂ into seawater (Guinotte and Fabry, 2008), has significant implications for coastal ecosystems (Matear and Lenton, 2014), particularly for plankton community respiration (PCR). As CO₂ dissolves in seawater, it forms carbonic acid, lowering the pH and altering the carbonate chemistry of marine environments. As a result of ongoing anthropogenic activities, it is predicted that the global average of the world’s ocean pH will continue to decrease, by up to 0.4 by 2100, and 0.77 by 2300 (Turley and Findlay, 2016). This decrease can disrupt metabolic processes in both autotrophic and heterotrophic plankton, impacting PCR directly and indirectly. For phytoplankton, the increase in inorganic carbon availability may directly fuel photosynthesis, and so, indirectly enhances autotrophic respiration (e.g., Qu et al., 2021), while acidification can also affect nutrient uptake, and cellular processes (Das and Mangwani, 2015; Bermúdez et al., 2016). Heterotrophic organisms may experience changes in metabolic efficiency and energy demands, often leading to changes in respiration rates (Motegi et al., 2013; Cripps et al., 2016; Davis et al., 2017; James et al., 2017). However, very few studies have investigated the effect of acidification on respiration in coastal waters at the community level.
Few mesocosm and microcosm experiments conducted in diverse coastal environments (e.g., Mediterranean Sea, Baltic Sea, Svalbard fjords) investigated the effects of elevated CO2 on PCR. Across these experiments, elevated CO2 levels generally had no direct impacts on plankton metabolic processes in oligotrophic areas. For example, studies conducted in low-nutrient, low-chlorophyll waters of the coastal Mediterranean Sea (Bay of Villefranche and Bay of Calvi) revealed no significant changes in PCR in response to increased CO2 (up to 1327 µatm) (Maugendre et al., 2017). Similarly, elevated CO2 (up to 700 µatm), simulating predicted pH conditions of year 2100, did not affect significantly PCR in a mesocosm experiment in Raunefjorden (South of Norway, Atlantic Ocean), even if other metabolic processes, such as the calcification rate of coccolithophorids, were significantly affected (Delille et al., 2005). In the same system, increased pCO2 (1050 µatm) did not affect PCR during another experiment, in which nutrients were added from the start of the experiment (Egge et al., 2009). Additionally, in the nutrient-limited Kongsfjorden (Arctic coastal waters, Svalbard archipelago), no significant relationship was found between PCR and 7 levels of pCO2 (up to 1420 µatm) in a mesocosm experiment, even after nutrient addition (Tanaka et al., 2013). Similarly, in coastal Mediterranean waters (North Western Alboran Sea, Southern Spain), elevated pCO2 (1000 µatm) and nutrient concentrations did not alter significantly PCR during a 7-d microcosm experiment, while light availability appeared to be the main controlling factor PCR in this system (Mercado et al., 2014).
In contrast, only two experimental studies revealed significant effects of simulated acidification on PCR in coastal European waters. In coastal Atlantic waters of the Canary Islands, increased pCO2 significantly enhanced PCR after nutrient amendment, while no significant effects were found before adding the nutrients (Filella et al., 2018), suggesting a positive effect of acidification on PCR in highly eutrophic conditions. Conversely, PCR was significantly reduced by almost 40% under high CO2 levels (up to ~ 1330 µatm) during a mesocosm experiment with nitrogen-limited coastal waters from the Gulf of Finland (Baltic Sea) (Spilling et al., 2016), maybe due to a shift toward smaller plankton groups. Overall, the contrasted results obtained from the few studies that have experimentally investigated the effects of ocean acidification on PCR at the community level suggest limited consequences of this stressor, but, most importantly, emphasizes the need of further studies, notably in coastal waters of various nutrient status, to expand our limited knowledge regarding the long-term effect of acidification. It should be noted that ocean alkalinity enhancement, by artificially increasing carbonate ion concentration, is growingly proposed as a nature-based solution to counteract acidification. The environmental consequences of this solution still need to be evaluated at a broad spatial and temporal scales; the first results of Marín-Samper et al. (2024) indicated no significant effect of a gradient of alkalinization on PCR during a mesocosm experiment in coastal waters of the Canary Islands (North Eastern Atlantic Ocean), while preliminary results from Serret et al. (2024) indicated a reduction of PCR in response to the addition of calcium hydroxide in coastal productive Atlantic waters (Ría de Vigo, Spain) and in the ultraoligotrophic eastern Mediterranean Sea (Crete, Greece).
3.3. Nutrient Sensitivity and Effect of Eutrophication
Nutrient availability, both inorganic (such as nitrate (NO₃⁻), orthophosphate (PO₄³⁻), and silicate (SiO₃²⁻)) and organic (including dissolved organic carbon (DOC), nitrogen (DON), and phosphorus (DOP)), is a key driver of plankton metabolism, influencing both autotrophic and heterotrophic metabolism (Legendre and Rassoulzadegan, 1995). Inorganic nutrients are essential for both autotrophic and heterotrophic metabolism while organic nutrients play a critical role in supporting the microbial loop and heterotrophic processes (Jones et al., 2024; Vanharanta et al., 2024). Therefore, the interaction between these inorganic and organic nutrient sources, and the nutrient limitation patterns, can significantly alter the structure and function of planktonic communities (Caron et al., 2000; Weber and Deutsch, 2010). Nutrient enrichment, for example from the land, either directly from terrestrial runoffs or from watershed and groundwater discharge (e.g., Palma Bay, Mediterranean Sea (Basterretxea et al., 2024)), can significantly change the structure of planktonic communities, and by doing so, increase or decrease PCR (e.g., Wilson et al., 2024). Nutrient inputs also often result in blooms of larger phytoplankton like diatoms (Spatharis et al., 2007), which have high metabolic rates and significantly contribute to carbon fixation (Coggins et al., 2023). However, this can also enhance PCR, as observed during spring blooms in temperate coastal waters (the Celtic Sea, North Atlantic Ocean (García-Martín et al., 2019b), San Francisco Bay, North Pacific Ocean (Caffrey et al., 1998)). Overall, due to these important roles, nutrient availability is often considered as the main factor regulating plankton metabolism (Prichett et al., 2024), often surpassing the effects of other parameters, notably temperature (Staehr and Sand-Jensen, 2006; Marañón et al., 2018).
In European coastal waters, nutrient availability is either dictated by hydrographic processes, such as coastal upwellings, and anthropogenic inputs, often leading in eutrophication. Eutrophication is the process by which water bodies become enriched with excess nutrients, particularly nitrogen (N) and phosphorus (P), often due to agricultural runoff, wastewater discharge, and other anthropogenic activities (Clarke et al., 2006; Rabalais et al., 2009). Eutrophication has become an important environmental issue in many coastal waters of Europe, and notably in the Baltic Sea, where it is associated with bad ecological status since several decades (Murray et al., 2019). Eutrophication can lead to algal blooms, enhancing PCR, such as reported in a Danish estuary (Jensen et al., 1990), particularly when large blooms die off and decompose, fueling heterotrophic bacteria and depleting dissolved oxygen in the water column. This often results in hypoxia (low oxygen levels), which can stress or kill marine organisms and alter the structure of planktonic communities, also changing PCR (Murrell et al., 2013; Zhu et al., 2016). However, some other coastal waters of southern Europe have experienced oligotrophication – a reduction in nutrient inputs - in the last decades (Mozetič et al., 2010; Derolez et al., 2020). In addition, global climate change, particularly the increase in temperature, is expected to have profound impacts on the water column stratification—the layering of water due to differences in temperature and density— of water column, which is anticipated to be intensified, especially during the summer months (Li et al., 2020). This increased stratification can reduce vertical mixing, limiting the exchange of nutrients between surface and deeper layers and potentially impairing nutrient availability for plankton metabolism in surface waters (Mena et al., 2019). Similarly to eutrophication, increase stratification and oligotrophication can significantly alter PCR, as observed for example in a coastal Mediterranean system (Mallorca, Spain) in which oligotrophication resulted in a significant decline of PCR (Agustí et al., 2017).
In light of the crucial role eutrophication and oligotrophication – changes in nutrient inputs in general - play in regulating marine ecosystems, many experiments have been conducted to investigate the effects of changes in nutrients on coastal plankton communities. However, differences in the chemical forms of added nutrients, as well as variations in the magnitude and frequency of nutrient inputs, make it challenging to compare results across experiments. Three in situ mesocosm experiments, with similar designs and undertaken simultaneously, were performed to assess the effects of added nutrients (nitrate and ammonium) on coastal plankton communities from the Baltic Sea (Tvärminne archipelago, Gulf of Finland), the Mediterranean Sea (Bay of Blanes, North Western Mediterranean), and the Atlantic Sea (Hopavågen Bay, central Norway) (Olsen et al., 2006). In all three locations, PCR responded linearly with nutrient loadings, suggesting a common response of PCR toward eutrophication regardless of different environments. Accordingly, positive effects of nutrient enrichment, regardless of the magnitude, frequency or type of loading performed, were found in coastal Mediterranean waters: inputs of phosphorus, silicate, and nitrogen in the Bay of Blanes (Duarte et al., 2004), inputs of phosphorus-rich dust in the Bay of Blanes (Lekunberri et al., 2010), inputs of nitrogen- and phosphorus-rich dust in oligotrophic waters of the Mediterranean (Gazeau et al., 2021), and inputs of ammonium and orthophosphate in the phosphorus-depleted Ionian basin (Lagaria et al., 2011). Similarly, positive effects of nutrient enrichment were also found in coastal Atlantic waters: inputs of nitrogen, phosphate and silicate-enriched deep water in the Canary Islands (Baños et al., 2022; Ortiz et al., 2022), inputs of nitrogen, phosphate and silicate-enriched deep water at varying silicate:nitrogen ratios in the Canary Islands (Ortiz et al., 2024), and, in the Ría de Vigo, with inputs of nitrogen-rich rainwater (Martínez-García et al., 2014) or with direct inputs of NH4NO3, Na2SiO3, and Na2HPO4 (López-Sandoval et al., 2025).
However, inorganic nitrogen and phosphorus inputs were found to decrease PCR in microcosms carried out with phosphorus-limited coastal Atlantic waters, while no effect were observed with the same experimental set-up in coastal Mediterranean waters, with lower in situ phosphorus-limitation (Cabrerizo et al., 2022), suggesting that the abiotic conditions to which plankton communities play a role in the observed response of PCR to nutrient loadings. Similarly, no significant effects of dust addition, and inputs of nitrogen, phosphate and organic matter, respectively, on PCR were found in coastal waters from the South Western (Cabrerizo et al., 2016) and North Western (Mercado et al., 2014) Mediterranean Sea. Furthermore, nitrogen-rich wastewater dissolved organic matter inputs decreased PCR from the coastal Baltic Sea, maybe due to the high lability of the added organic matter that could have fueled bacterial production without enhancing respiration (Vaquer-Sunyer et al., 2016). In contrast, the input of organic nutrients enhanced PCR in coastal Atlantic waters (Ría de Vigo, Spain) while the input of inorganic nutrients did not significantly affect PCR during the same experiment (Martínez-García et al., 2013). Overall, these results suggest that while increased nutrient availability generally has a positive effect on PCR, the unexpected absence or even negative response of PCR to nutrient enrichment in some cases may indicate complex mechanisms related to shifts in community structure and interactions among organisms. This complexity makes it difficult to predict the effects of climate change-related changes in nutrient availabilities on PCR in coastal waters on a global scale in the future.
3.4. Light and Ultraviolet Sensitivity
Light availability is a fundamental driver of plankton metabolism, directly influencing photosynthesis in phytoplankton (Striebel et al., 2023), and indirectly increasing overall plankton community respiration (PCR) through photosynthesis providing organic substrates that support respiratory processes (Pringault et al., 2009; Mesa et al., 2017). However, exposure to ultraviolet (UV) radiation, particularly UV-B (280-320 nm), can have both inhibitory and stimulatory effects on different components of plankton (Belzile et al., 2006; Moreau et al., 2014). UV radiation can impair photosynthetic efficiency and damage cellular structures, especially in surface-dwelling plankton, leading to stress-induced increases in respiration rates as cells repair damage (Agustí et al., 2014). Therefore, the balance between beneficial and harmful effects of light and UV exposure makes irradiance a complex yet essential factor in determining the metabolic balance of planktonic ecosystems. While Regaudie-de-Gioux et al. (2014) and Agustí et al. (2014) showed higher respiration rates and shifts toward heterotrophy at the global scale when comparing metabolism estimated in bottles that do not remove UV wavelengths (quartz bottles) versus bottles that do (glass borosilicate bottles), the only study that actually assessed the effects of increased UV radiations on PCR in larger volumes (2260 L in situ mesocosms) in European coastal waters reported no significant effects of a 20% increase in UVB radiations on PCR in coastal Mediterranean waters (Thau Lagoon, Vidussi et al., 2011).
In many coastal European waters, climate change and anthropogenic activities are increasing the frequency and intensity of terrestrial runoffs, lowering light availability and changing the light spectrum of the water column by bringing colored light-absorbing dissolved organic matter (the so called ‘brownification’ or ‘darkening’ phenomenon) (Frigstad et al., 2023; Opdal et al., 2023; Aigars et al., 2024; Solås et al., 2024). In Arctic coastal waters, this phenomenon is of extreme importance, notably due to the intensification of glacier melting and consequent huge runoffs of glacier meltwater (van Pelt et al., 2019). Accordingly, riverine runoffs were suggested to be the most important factor controlling PCR in some regions of the European Arctic (Vaquer-Sunyer et al., 2013). Many experiments have reported significant effects of terrestrial runoffs on plankton community structure in European coastal waters (e.g., Courboulès et al., 2023; Garnier et al., 2023; Ktistaki et al., 2024), however, only few experiments have investigated the effects of such phenomenon on PCR. In Norwegian coastal waters, brownification, simulated by the addition of a highly recalcitrant humic substance, decreased PCR by 27%, while light availability was reduced by 23% (Soulié et al., 2022c). Accordingly, an artificial decrease of light (to 32% of the full solar irradiance) significantly decreased PCR in coastal Mediterranean waters (Alboran Sea (Mercado et al., 2014)). In contrast, simulated terrestrial runoffs significantly enhanced PCR by 53 to 73% in coastal Mediterranean waters (Liess et al., 2016; Soulié et al., 2024), despite significant reduction in light availability. However, in both studies, the simulated terrestrial runoff added particulate or dissolved nutrients that fueled plankton metabolism to an extent that suppressed the negative effect due to light attenuation. These studies suggest the existence of a threshold of nutrient enrichment versus light reduction brought by terrestrial runoffs upon which the fate of PCR would depend, making it even more difficult to predict the effects of the increases in intensity and frequency of terrestrial runoffs due to climate change on the long term. In this regard, the limited number of existing studies in which PCR response to experimental light manipulation in natural communities calls for further investigation and experiments.
4. Knowledge Gaps in Studied Disturbances and Areas
Because measuring PCR can be time-consuming, labor-intensive, and expensive, PCR is usually not included in monitoring programs of coastal environments and in climate-change experiments (Breitburg et al., 2018; Wikner et al., 2023). Hence, available literature on the climate change effects on PCR remains quite scarce, notably in some understudied systems. A recent review on the contribution of PCR on ocean global deoxygenation highlighted the need of better parametrization of the influence of environmental and biological parameters, such as nutrient concentrations and stoichiometry or plankton community structure, on microbial respiration (Robinson 2019). Similarly, current models predicting ocean deoxygenation are still lacking a mechanistic understanding of respiratory oxygen demand to accurately reproduce observe ocean deoxygenation (Oschlies et al., 2018). In this regard, experiments can be used to characterize this mechanistic understanding and help resolve the influence of regional processes on global biogeochemical cycles.
The Figure 1 represents the location of all the experiments testing climate change disturbance that were included in the present review. A clear difference in the extent systems have been studied can be seen from this map, with the Mediterranean and Baltic seas being more studied than other European coastal systems. Notably, no experiments testing the effect of climate change-related disturbances on PCR were published for the Black Sea, some parts of the Mediterranean Sea (e.g., the Adriatic Sea), and some parts of the Atlantic Ocean (e.g., the North Sea) (Figure 1). While common responses were identified regarding the effect of certain disturbances on PCR, some system-specific were also highlighted, emphasizing the need of more experimental studies in the understudied and unstudied systems. Regarding the complex relationships between PCR and environmental conditions, these experiments are needed to better our capacity to understand and predict the consequences of the global climate change on biogeochemical cycles.
Change in temperature is the most studied stressor (24 studies) among all experiments included in the present study (Figure 2), in accordance with the importance of water temperature in the regulation of metabolism at a global scale (Regaudie-de-Gioux and Duarte 2012; Boscolo-Galazzo et al., 2018). Even if the positive impact of warming described in global ocean datasets and theoretical predictions was confirmed by experiments in many coastal systems, some studies highlighted contradictory responses of PCR to warming, often related to warming-induced trophic cascade phenomenon altering the biomass of phyto- and/or bacterioplankton. Typically, observations and long-term monitoring cannot parametrize such causal relationships, emphasizing on the need of experimental studies in diverse coastal systems. Changes in nutrient inputs and ratios, including here inputs of nutrients of any sort (terrestrial runoff, artificial inputs, dust inputs..) were the second most studied stressor (18 studies). Global climate change has been proved to greatly affect nutrient dynamics and cycling in the ocean (Hutchins and Capone, 2022), hence the importance of experimentally testing the effects of nutrient changes on PCR. As for warming, while the positive effect of nutrient inputs on metabolism, and so PCR, is straightforward, changes in nutrient ratios can alter community dynamics and interactions, ultimately impacting PCR. This calls for experiments in which nutrient inputs are closely mimicking naturally occurring processes and regional effects of climate change, such as done with Saharan dust inputs in the Mediterranean or changes in upwelling dynamics in coastal Atlantic waters. Finally, ocean acidification is also among the most studied stressors (13 studies) within all the experiments included in the present study. In contrast to temperature and nutrients, the effect of ocean acidification on metabolism at the global scale is difficult to predict. Importance of experimentations testing the effect of acidification on PCR is highlighted by the negative and positive effects found in coastal waters of the Baltic Sea (Spilling et al., 2016) and the Canary Islands (Filella et al., 2018), respectively, while all other studies found no significant effects on PCR. In this regard, other experiments are needed to confirm or infirm such results, and gain understanding in why significant effects were found in such systems but not in other. Other stressors, and notably changes in light and UV availability and spectrum, were not tested in many experiments (7 studies). Therefore, knowledge is still scarce regarding such consequences of climate change; however, certain coastal systems in northern Europe are now increasingly darker (Opdal et al., 2024), and knowledge of the effect of such disturbance on PCR is essential to be able to fully understand and predict the response of such systems in the future. Furthermore, experiments in which light is isolated from associated stressors, notably nutrients, are needed to be able to fully characterize the effects of light and disentangle them from potential effects of other environmental factors, as for now, only one study studied the response of coastal PCR to the input of recalcitrant humic substance to simulate coastal darkening so that light reduction and change in water color are simulated without nutrient enrichment (Soulié et al., 2022c). In addition to stressors included in the experiments testing PCR in European coastal waters, the effect of some disturbances related to climate change on PCR were never studied to my knowledge, while these disturbances can be locally very important in shaping the ecosystem. For example, salinity has decreased and is expected to continue to decrease in coastal waters of the Baltic Sea, due to increased precipitations and freshwater runoffs (Lehmann et al., 2022). Such salinity changes have been shown to shift zooplankton (Hall and Lewandowska, 2022) and phytoplankton (Pilkaitytë et al., 2004) community composition, and induce changes in copepod respiration (Soulié et al., 2022d), but no study have still investigated the effects of salinity changes on PCR in the Baltic Sea. Similarly, intensification of glacier meltwater runoffs (increased by 11% since the 1980-2000 period) in the coastal Arctic Ocean have been shown to increase and decrease phytoplankton productivity with marine- and land-terminating glaciers, respectively (Ardyna and Arrigo, 2020), but no experimental study has ever been performed to assess the response of PCR toward glacier meltwater, even if such response may play a crucial role in the future Arctic Ocean biogeochemical cycles.
While studying individual drivers like temperature, pH, and nutrient concentrations provides valuable insights in to the response of PCR and can inform specific mitigation strategies, it's essential to study these factors simultaneously as they often act in combination in natural settings. These simultaneous changes can lead to complex, interactive effects on planktonic communities that might not be predictable based solely on the isolated impact of each driver (e.g., Moreno et al., 2022). Among all experiments included in the present review, only 7 studies tested different drivers simultaneously in full factorial designs. These studies can help understand the response of PCR toward simultaneous stressors, similarly to what occurs naturally in coastal waters, and if these disturbances will act antagonistically or synergistically on PCR. Many experiments highlighted complex interactions between stressors, and the effect of stressors applied simultaneously often differs from the addition of single-stressor effects. For example, Cabrerizo et al. (2022) showed that the effects of +3°C warming on PCR acted additively to nutrient enrichment in coastal Mediterranean and Atlantic waters, but only under future conditions of pH and UV irradiance. Overall, a more holistic approach, using multi-driver experiments, is crucial for developing effective mitigation strategies that reflect the multifaceted reality of global change.
The present review listed a number of studies assessing the complex responses of PCR in European coastal waters towards various climate change effects, this number following an increasing trend since the early 2000’s (Figure 2), suggesting that an increasing number of scientists and projects acknowledge the great importance of PCR in the functioning of coastal ecosystems and global biogeochemical cycles. However, this review mainly stresses out the scarcity of studies in which PCR is actually monitored, putting into light understudied regions and stressors, and calling for a better integration of PCR in both experimental studies and long-term monitoring programs. Future research on the effects of climate change on PCR should consider integrating approaches that combine experimental studies, long-term monitoring, and modeling techniques. While experimental studies offer controlled environments to test specific hypotheses on PCR and isolate the effects of stressors, long-term monitoring programs can help capture natural variability and identify trends in PCR while taking into account the full complexity of natural systems. Coupling these approaches with predictive modeling can help bridge the gap between small-scale, short-term experiments and the complex, large-scale dynamics of coastal ecosystems. Such an integrated framework is vital for informing conservation strategies and sustainable management of coastal waters in the face of global climate change.
Author Contributions
T. S. designed and wrote the article and produced the figures.
Data Availability Statement
This review article only refers to published research articles and their data. The reader is referred to the cited literature to access the data.
Acknowledgements
The author would like to thank Dr. Justine Courboulès for useful discussions and constructive comments. The writing of this review was not supported by any specific financial support.
References
- Agustí, S., A. Regaudie-de-Gioux, J. M. Arrieta, and C. M. Duarte. 2014. Consequences of UV-enhanced community respiration for plankton metabolic balance. Limnol. Oceanogr. 59(1):223-232. [CrossRef]
- Agustí, S., J. Martinez-Ayala, A. Regaudie-de-Gioux, and C. M. Duarte. 2017. Oligotrophication and metabolic slowing-down of a NW Mediterranean coastal ecosystem. Front. Mar. Sci. 4:432. [CrossRef]
- Agustí, S., L. Vigoya, and C. M. Duarte. 2018. Annual plankton community metabolism in estuarine and coastal waters in Perth (Western Australia). PeerJ 6:e5081. [CrossRef]
- Aigars, J., N. Suhareva, D. Cepite-Frisfelde, I. Kokorite, A. Iital, M. Skudra, and M. Viska. 2024. From green to brown: two decades of darkening coastal water in the Gulf of Riga, the Baltic Sea. Front. Mar. Sci. 11:1369537. [CrossRef]
- Aranguren-Gassis, M., P. Serret, E. Fernández, J. L. Herrera, J. F. Domínguez, V. Pérez, and J. Escanez. 2013. Balanced plankton net community metabolism in the oligotrophic North Atlantic subtropical gyre from Lagrangian observations. Deep Sea Res. I: Oceanogr. Res. Pap. 68:116-122. [CrossRef]
- Ardyna, M., and K. R. Arrigo. 2020. Phytoplankton dynamics in a changing Arctic Ocean. Nat. Clim. Change 10:892-903. [CrossRef]
- Arístegui, J., and W. G. Harrison. 2002. Decoupling of primary production and community respiration in the ocean: implications for regional carbon studies. Aquat. Microb. Ecol. 29:199-209. [CrossRef]
- Baños, I., J. Arístegui, M. Benavides, M. Gómez-Letona, M. F. Montero, J. Ortiz, K. G. Schulz, A. Ludwig, and U. Riebesell. 2022. Response of plankton community respiration under variable simulated upwelling events. Front. Mar. Sci. 9:1006010. [CrossRef]
- Bashiri, B., A. Barzandeh, A. Männik, and U. Raudsepp. 2024. Variability of marine heatwaves’ characteristics and assessment of their potential drivers in the Baltic Sea over the last 42 years. Sci. Rep. 14:22419. [CrossRef]
- Bas-Silvestre, M., M. Antón-Pardo, D. Boix, S. Gascón, J. Compte, J. Bou, B. Obrador, and X. D. Quintana. 2024. Phytoplankton composition in Mediterranean confined coastal lagoons: testing the use of ecosystem metabolism for the quantification of community-related variables. Aquat. Sci. 86:71. [CrossRef]
- Basterretxea, G., J. S. Font-Muñoz, M. Kane, A. Regaudie-de-Gioux, C. T. Satta, and I. Tuval. 2024. Pulsed wind-driven control of phytoplankton biomass at a groundwater-enriched nearshore environment. Sci. Tot. Env. 955:177123. [CrossRef]
- Belzile, C., S. Demers, G. A. Ferreyra, I. Schloss, C. Nozais, K. Lacoste, and others. 2006. UV effects on marine planktonic food webs: A synthesis of results from mesocosm experiments. Photochem. Photobiol. 82:850-856. [CrossRef]
- Beman, J. M., K. R. Arrigo, and P. A. Matson. 2005. Agricultural runoff fuels large phytoplankton blooms in vulnerable areas of the ocean. Nature 434:211-214. [CrossRef]
- Bermúdez, J. R., U. Riebesell, A. Larsen, and M. Winder. 2016. Ocean acidification reduces transfer of essential biomolecules in a natural plankton community. Sci. Rep. 6:27749. [CrossRef]
- Breitburg, D., L. A. Levin, A. Oschlies, M. Grégoire, F. P. Chavez, D. J. Conley, and others. 2018. Declining oxygen in the global ocean and coastal waters. Science 359:eaam7240. [CrossRef]
- Breithaupt, P. 2009. The impact of climate change on phytoplankton-bacterioplankton interactions. PhD Thesis. Christian-Albrechts-Universität zu Kiel, 210p.
- Boscolo-Galazzo, F., K. A. Crichton, S. Barker, and P. N. Pearson. 2018. Temperature dependency of metabolic rates in the upper ocean: A positive feedback to global climate change? Global Planet. Change 170:201-212. [CrossRef]
- Brown, J. H., J. F. Gillooly, A. P. Allen, V. M. Savage, and G. B. West. 2004. Toward a metabolic theory of ecology. Ecology 85(7):1771-1789. [CrossRef]
- Cabrerizo, M. J., J. M. Medina-Sánchez, J. M. González-Olalla, M. Vila-Argaiz, and P. Carrillo. 2016. Saharan dust inputs and high UVR levels jointly alter the metabolic balance of marine oligotrophic ecosystems. Sci. Rep. 6:35892. [CrossRef]
- Cabrerizo, M. J., E. Marañón, C. Fernández-González, A. Alonso-Núñez, H. Larsson, and M. Aranguren-Gassis. 2021. Temperature fluctuation attenuates the effects of warming in estuarine microbial plankton communities. Front. Mar. Sci. 8:656282. [CrossRef]
- Cabrerizo, M. J., J. M. Medina-Sánchez, J. M. González-Olalla, D. Sánchez-Gómez, and P. Carrillo. 2022. Microbial plankton responses to multiple environmental drivers in marine ecosystems with different phosphorus limitation degrees. Sci. Tot. Env. 816:151491. [CrossRef]
- Caffrey, J. M., J. E. Cloern, and C. Grenz. 1998. Changes in production and respiration during a spring phytoplankton bloom in San Francisco Bay, California, USA: implications for net ecosystem metabolism. Mar. Ecol. Prog. Ser. 172:1-12. [CrossRef]
- Caron, D. A., E. L. Lim, R. W. Sanders, M. R. Dennett, and U.-G. Berninger. 2000. Responses of bacterioplankton and phytoplankton to organic carbon and inorganic nutrient additions in contrasting oceanic ecosystems. Aquat. Microb. Ecol. 22:175-184. [CrossRef]
- Carstensen, J., R. Klais, and J. E. Cloern. 2015. Phytoplankton blooms in estuarine and coastal waters: Seasonal patterns and key species. Est. Coast. Shelf Sci. 162:98-109. [CrossRef]
- Cavan, E. L., and P. W. Boyd. 2018. Effect of anthropogenic warming on microbial respiration and particulate organic carbon export rates in the Sub-Antarctic Southern Ocean. Aquat. Microb. Ecol. 82:11-127. [CrossRef]
- Chen, B. 2015. Patterns of thermal limits of phytoplankton. J. Plankt. Res. 37(2):285-292. [CrossRef]
- Cheng, L., K. von Schuckmann, J. P. Abraham, K. E. Trenberth, M. E. Mann, L. Zanna, and others. 2022. Past and future ocean warming. Nat. Rev. Earth Environ. 3:776-794. [CrossRef]
- Chou, W. R., L. S. Fang, W. H. Wang, and K. S. Tew. 2012. Environmental influence on coastal phytoplankton and zooplankton diversity: a multivariate statistical model analysis. Environ. Monit. Assess. 184:5679-5688. [CrossRef]
- Clarke, A. L., K. Weckström, D. J. Conley, N. J. Anderson, F. Adser, E. Andrén, and others. 2006. Long-term trends in eutrophication and nutrients in the coastal zone. Limnol. Oceanogr. 51(1):385-397. [CrossRef]
- Cloern, J. E., S. Q. Foster, and A. E. Kleckner. 2014. Phytoplankton primary production in the world’s estuarine-coastal ecosystems. Biogeosciences 11: 2477-2501. [CrossRef]
- Coggins, A., A. J. Watson, U. Schuster, N. Mackay, B. King, E. McDonagh, and A. J. Poulton. 2023. Surface ocean carbon budget in the 2017 South Georgia diatom bloom: Observations and validation of profiling biogeochemical Argo floats. Deep Sea Res. II: Top. Stud. Oceanogr. 209:105275. [CrossRef]
- Courboulès, J., F. Vidussi, T. Soulié, E. Nikiforakis, M. Heydon, S. Mas, F. Joux, and B. Mostajir. 2023. Effects of an experimental terrestrial runoff on the components of the plankton food web in a Mediterranean coastal lagoon. Front. Mar. Sci. 10:1200757. [CrossRef]
- Cripps, G., K. J. Flynn, and P. K. Lindeque. 2016. Ocean acidification affects the phyto-zoo plankton trophic transfer efficiency. PLoS ONE 11(4):e0151739. [CrossRef]
- Darmaraki, S., D. Denaxa, I. Theodorou, E. Livanou, D. Rigatou, D. E. Raitsos, and others. 2024. Marine heatwaves in the Mediterranean Sea: A literature review. Medit. Mar. Sci. 25(3):586-620. [CrossRef]
- Das, S., and N. Mangwani. 2015. Ocean acidification and marine microorganisms: responses and consequences. Oceanologia 57(4):349-361. [CrossRef]
- Davis, C. V., E. B. Rivest, T. M. Hill, B. Gaylord, A. D. Russell, and E. Sandford. 2017. Ocean acidification compromises a planktic calcifier with implications for global carbon cycling. Sci. Rep. 7:2225. [CrossRef]
- del Giorgio, P. A., and P. J. le B. Williams. 2005. The global significance of respiration in aquatic ecosystems: from single cells to the biosphere. In: P. A. del Giorgio, P. J. le B. Williams, and B. LE. (Eds) Respiration in aquatic ecosystems. Oxford University Press, 267-303.
- Delille, B., J. Harlay, I. Zondervan, S. Jacquet, L. Chou, R. Wollast, and others. 2005. Response of primary production and calcification to changes of pCO2 during experimental blooms of the coccolithophorid Emiliania huxleyi. Global Biogeochem. Cycles 19(2). [CrossRef]
- Derolez, V., D. Soudant, N. Malet, C. Chiantella, M. Richard, E. Abadie, C. Aliaume, and B. Bec. 2020. Two decades of oligotrophication: Evidence for a phytoplankton community shift in the coastal lagoon of Thau (Mediterranean Sea, France). Est. Coast. Shelf Sci. 241:106810. [CrossRef]
- Duarte, C. M., S. Agustí, and D. Vaqué. 2004. Controls on planktonic metabolism in the Bay of Blanes, northwestern Mediterranean littoral. Limnol. Oceanogr. 49(6):2162-2170. [CrossRef]
- Egge, J. K., T. F. Thingstad, A. Larsen, A. Engel, J. Wohlers, R. G. J. Bellerby, and U. Riebesell. 2009. Primary production during nutrient-induced blooms at elevated CO2 concentrations. Biogeosciences 6:877-885. [CrossRef]
- Falkowski, P. G. 1994. The role of phytoplankton photosynthesis in global biogeochemical cycles. Photosynth. Res. 39: 235-258. [CrossRef]
- Field, C. B., M. J. Behrenfeld, J. T. Randerson, and P. Falkowski. 1998. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 281: 237-240. [CrossRef]
- Filella, A., I. Baños, M. F. Montero, N. Hernández-Hernández, A. Rodríguez-Santos, A. Ludwig, U. Riebesell, and J. Aristegui. 2018. Plankton community respiration and ETS activity under variable CO2 and nutrient fertilization during a mesocosm study in the subtropical North Atlantic. Front. Mar. Sci. 5:310. [CrossRef]
- Forsblom, L., J. Engström-Öst, S. Lehtinen, I. Lips, and A. Lindén. 2019. Environmental variables driving species and genus level changes in annual plankton biomass. J. Plankt. Res. 41(6):925-938. [CrossRef]
- Fredston-Hermann, A., C. J. Brown, S. Albert, C. J. Klein, S. Mangubhai, J. L. Nelson, and others. 2016. Where does river runoff matter for coastal marine conservation? Front. Mar. Sci. 3:273. [CrossRef]
- Frigstad, H., G. S. Andersen, H. C. Trannum, M. McGovern, L.-J. Naustvoll, Ø. Kaste, A. Deininger, and D. Ø. Hjermann. 2023. Three decades of change in the Skagerrak coastal ecosystem, shaped by eutrophication and coastal darkening. Est. Coast. Shelf Sci. 283:108193. [CrossRef]
- Frölicher, T. L., E. M. Fischer, and N. Gruber. 2018. Marine heatwaves under global warming. Nature 560:360-364. [CrossRef]
- Garcia-Corral, L. S., J. M. Holding, P. Carrillo-de-Albornoz, A. Steckbauer, M. Pérez-Lorenzo, P. Serret, and others. 2017. Temperature dependance of plankton community metabolism in the subtropical and tropical oceans. Global Biogeochem. Cycles 31:1141-1154. [CrossRef]
- García-Martín, E. E., P. Serret, and M. Pérez-Lorenzo. 2011. Testing potential bias in marine plankton respiration rates by dark bottle incubations in the NW Iberian shelf: incubation time and bottle volume. Cont. Shelf Res. 31(5):496-506. [CrossRef]
- García-Martín, E. E., C. J. Daniels, K. Davidson, C. E. Davis, C. Mahaffey, K. M. J. Mayers, and others. 2019a. Seasonal changes in plankton respiration and bacterial metabolism in a temperate shelf sea. Prog. Oceanogr. 177:101884. [CrossRef]
- García-Martín, E. E., C. J. Daniels, K. Davidson, J. Lozano, K. M. J. Mayers, S. McNeill, and others. 2019b. Plankton community respiration and bacterial metabolism in a North Atlantic shelf sea during spring bloom development (April 2015). Prog. Oceanogr. 177:101873. [CrossRef]
- Garnier, A., Ö. Östman, J. Ask, O. Bell, M. Berggren, M. P. D. Rulli, H. Younes, and M. Huss. 2023. Coastal darkening exacerbates eutrophication symptoms through bottom-up and top-down control modification. Limnol. Oceanogr. 68:678-691. [CrossRef]
- Garrabou, J., D. Gómez-Gras, A. Medrano, C. Cerrano, M. Ponti, R. Schlegel, and others. 2022. Marine heatwaves drive recurrent mass mortalities in the Mediterranean Sea. Glob. Change Biol. 28(19):5708-5725. [CrossRef]
- Gazeau, F., F. Van Wambeke, E. Marañón, M. Pérez-Lorenzo, S. Alliouane, C. Stolpe, and others. 2021. Impact of dust addition on the metabolism of Mediterranean plankton communities and carbon export under present and future conditions of pH and temperature. Biogeosciences 18:5423-5446. [CrossRef]
- Gomez-Castillo, A. P., A. Panton, and D. A. Purdie. 2023. Temporal variability of phytoplankton biomass and net community production in a macrotidal temperate estuary. Est. Coast. Shelf Sci. 280:108182. [CrossRef]
- Grégoire, M., A. Oschlies, D. Canfield, C. Castro, I. Ciglenečki, P. Croot, K. Salin, B. Schneider, P. Serret, C.P. Slomp, T. Tesi, and M. Yücel. 2023. Ocean Oxygen: the role of the Ocean in the oxygen we breathe and the threat of deoxygenation. In: Rodriguez Perez, A., Kellett, P., Alexander, B., Muñiz Piniella, Á., Van Elslander, J., Heymans, J. J., [Eds.] Future Science Brief No. 10 of the European Marine Board, Ostend, Belgium. ISSN: 2593-5232. ISBN: 9789464206180. [CrossRef]
- Guinotte, J. M., and V. J. Fabry. 2008. Ocean acidification and its potential effects on marine ecosystems. Annals of the New York Academy of Sciences 1134:320-342. [CrossRef]
- Hall, C. A. M., and A. M. Lewandowska. 2022. Zooplankton dominance shift in response to climate-driven salinity change: A mesocosm study. Front. Mar. Sci. 9:861297. [CrossRef]
- Harley, C. D. G., A. R. Hughes, K. M. Hultgren, B. G. Milner, C. J. B. Sorte, C. S. Thornber, and others. 2006. The impacts of climate change in coastal marine systems. Ecol. Lett. 9(2):228-241. [CrossRef]
- Hays, G. C., A. J. Richardson, and C. Robinson. 2005. Climate change and marine plankton. Trends Ecol. Evol. 20(6): 337-344. [CrossRef]
- Henson, S., C. A. Baker, P. Halloran, A. McQuatters-Gollop, S. Painter, A. Planchat, and A. Tagliablue. 2024. Knowledge gaps in quantifying the climate change response of biological storage of carbon in the ocean. Earth’s future 12:e2023EF004375.
- Holding, J. M., C. M. Duarte, J. M. Arrieta, R. Vaquer-Sunyer, A. Coello-Camba, P. Wassmann, and S. Agustí. 2013. Experimentally determined temperature thresholds for Arctic plankton community metabolism. Biogeosciences 10:357-370. [CrossRef]
- Hoppe, H.-G., P. Breithaupt, K. Walther, R. Koppe, S. Bleck, U. Sommer, and K. Jürgens. 2008. Climate warming in winter affects the coupling between phytoplankton and bacteria during the spring bloom: a mesocosm study. Aquat. Microb. Ecol. 51:105-115. [CrossRef]
- Huete-Stauffer, T. M., N. Arandia-Gorostidi, N. González-Benítez, L. Díaz-Pérez, A. Calvo-Díaz, and X. A. G. Morán. 2018. Large plankton enhance heterotrophy under experimental warming in a temperate coastal ecosystem. Ecosystems 21:1139-1154. [CrossRef]
- Hutchins, D. A., and D. G. Capone. 2022. The marine nitrogen cycle: new developments and global change. Nat. Rev. Microbiol. 20:401-414. [CrossRef]
- IPCC, 2019. IPCC Special Report on the Ocean and Cryosphere in a Changing Climate [H.-O. Pörtner, D.C. Roberts, V. Masson-Delmotte, P. Zhai, M. Tignor, E. Poloczanska, K. Mintenbeck, A. Alegría, M. Nicolai, A. Okem, J. Petzold, B. Rama, N.M. Weyer (eds.)]. Cambridge University Press, Cambridge, UK and New York, NY, USA, 755 pp. [CrossRef]
- James, A. K., U. Passow, M. A. Brzezinski, R. J. Parsons, J. N. Trapani, and C. A. Carlson. 2017. Elevated pCO2 enhances bacterioplankton removal of organic carbon. PLoS ONE 12:e0173145. [CrossRef]
- Jensen, L. M., K. Sand-Jensen, S. Marcher, and M. Hansen. 1990. Plankton community respiration along a nutrient gradient in a shallow Danish estuary. Mar. Ecol. Prog. Ser. 61:75-85. [CrossRef]
- Jiao, N., and F. Azam. 2011. Microbial carbon pump and its significance for carbon sequestration in the ocean. In: Jiao N., Azam F., Sanders S. (Eds.) Microbial carbon pump in the Ocean 10:43-45, Science/AAAS, Washington DC.
- Jones, K., A. Liess, and J. Sjöstedt. 2024. Microbial carbon utilization in a boreal lake under the combined pressures of brownification and eutrophication: insights from a field experiment. Hydrobiologia. [CrossRef]
- Ktistaki, G., I. Magiopoulos, G. Corno, J. Courboulès, E. M. Eckert, J. González, and others. 2024. Brownification in the Eastern Mediterranean Sea: effect of simulated terrestrial input on the planktonic microbial food web in an oligotrophic sea. Front. Mar. Sci. 11:1343415. [CrossRef]
- Lagaria, A., S. Psarra, D. Lefèvre, F. Van Wambeke, C. Courties, M. Pujo-Pay, L. Oriol, T. Tanaka, and U. Christaki. 2011. The effects of nutrient additions on particulate and dissolved primary production and metabolic state in surface waters of three Mediterranean eddies. Biogeosciences 8:2595-2607. [CrossRef]
- Latorre, M. P., C. M. Iachetti, I. R. Schloss, J. Antoni, A. Malits, F. de la Rosa, and others. 2023. Summer heatwaves affect coastal Antarctic plankton metabolism and community structure. J. Exp. Mar. Biol. Ecol. 567:151926. [CrossRef]
- Laufkötter, C., M. Vogt, N. Gruber, M. Aita-Noguchi, O. Aumont, L. Bopp, and others. 2015.. Drivers and uncertainties of future global marine primary production in marine ecosystem models. Biogeosciences 12:6955-6984. [CrossRef]
- Lefèvre, D., T. L. Bentley, C. Robinson, S. P. Blight, and P. J. le B. Williams. 1994. The temperature response of gross and net community production and respiration in time-varying assemblages of temperate marine micro-plankton. J. Exp. Mar. Biol. Ecol. 184(2):201-215. [CrossRef]
- Legendre, L., and F. Rassoulzadegan. 1995. Plankton and nutrient dynamics in marine waters. Ophelia 41(1):153-172. [CrossRef]
- Lehmann, A., K. Myrberg, P. Post, I. Chubarenko, I. Dailidiene, H.-H. Hinrichsen, and others. 2022. Salinity dynamics of the Baltic Sea. Earth Syst. Dynam. 13:373-392. [CrossRef]
- Lekunberri, I., T. Lefort, E. Romero, E. Vázquez-Domínguez, C. Romera-Castillo, C. Marrasé, F. Peters, M. Weinbauer, and J. M. Gasol. 2010. Effects of a dust deposition event on coastal marine microbial abundance and activity, bacterial community structure and ecosystem function. J. Plankt. Res. 32(4):381-396. [CrossRef]
- Lewandowska, A. M., D. G. Boyce, M. Hofmann, B. Matthiessen, U. Sommer, and B. Worm. 2014. Effects of sea surface warming on marine plankton. Ecol. Lett. 17(5):614-623. [CrossRef]
- Li, G., L. Cheng, J. Zhu, K. E; Trenberth, M. E. Mann, and J. P. Abraham. 2020. Increasing ocean stratification over the past half-century. Nat. Clim. Change 10:1116-1123. [CrossRef]
- Liess, A., O. Rowe, S. N. Francoeur, J. Guo, K. Lange, A. Schröder, and others. 2016. Terrestrial runoff boosts phytoplankton in a Mediterranean coastal lagoon, but these effects do not propagate to higher trophic levels. Hydrobiologia 766:275-291. [CrossRef]
- Litchman, E., K. F. Edwards, C. A. Klausmeier, and M. K. Thomas. 2012. Phytoplankton niches, traits and eco-evolutionary responses to global environmental change. Mar. Ecol. Prog. Ser. 470: 235-248. [CrossRef]
- Lizon, F. Y. Lagadeuc, C. Brunet, D. Aelbrecht, and D. Bentley. 1995. Primary production and photoadaptation of phytoplankton in relation with tidal mixing in coastal waters. J. Plankt. Res. 17(5):1039-1055. [CrossRef]
- López-Sandoval, D. C., C. Fernández-González, C. González-García, and E. Marañón. 2025. Warming accelerates phytoplankton bloom dynamics and differentially affects the fluxes of carbon, nitrogen and oxygen through a coastal microbial community. Microb. Ecol. 88:117. [CrossRef]
- López-Urrutia, A., E. San Martin, R. P. Harris, and X. Irigoien. 2006. Scaling the metabolic balance of the oceans. Proc. Natl. Acad. Sci. U. S. A. 103: 8739-8744. [CrossRef]
- Lozano, J., M. Aranguren-Gassis, E. E. García-Martín, J. González, J. L. Herrera, B. Hidalgo-Robatto, D. Mártinez-Castrillón, M. Pérez-Lorenzo, R. A. Varela, and P. Serret. 2021. Seasonality of phytoplankton cell size and the relation between photosynthesis and respiration in the Ría de Vigo (NW Spain). Mar. Ecol. Prog. Ser. 664:43-58. [CrossRef]
- Mantikci, M., M. Bentzon-Tilia, S. J. Traving, H. Knudsen-Leerbeck, L. Riemann, and others. 2024. Plankton community metabolism variations in two temperate coastal waters of contrasting nutrient richness. JGR Biogeosci. 129(6):e2023JG007919. [CrossRef]
- Marañón, E., M. P. Lorenzo, P. Cermeño, and B. Mouriño-Carballido. 2018. Nutrient limitation suppresses the temperature dependence of phytoplankton metabolic rates. ISME J. 12(7):1836-1845. [CrossRef]
- Marín-Samper, L., J. Arístegui, N. Hernández-Hernández, J. Ortiz, S. D. Archer, A. Ludwig, and U. Riebesell. 2024. Assessing the impact of CO2-equilibrated ocean alkalinity enhancement on microbial metabolic rates in an oligotrophic system. Biogeosciences 21:2859-2876. [CrossRef]
- Martínez-García, S., E. Fernández, A. Calvo-Díaz, P. Cermeño, E. Marañón, X. A. G. Morán, and E. Teira. 2013. Differential response of microbial plankton to nutrient inputs in oligotrophic versus mesotrophic waters of the North Atlantic. Mar. Biol. Res. 9(4):358-370. [CrossRef]
- Martínez-García, S., B. Arbones, E. E. García-Martín, I. G. Teixeira, P. Serret, E. Fernández, F. G. Figueiras, E. Teira, and X. A. Álvarez-Salgado. 2014. Impact of atmospheric deposition on the metabolism of coastal microbial communities. Est. Coast. Shelf Sci. 153:18-28. [CrossRef]
- Matear, R. J., and A. Lenton. Quantifying the impact of ocean acidification on our future climate. Biogeosciences 11(14):3965-3983. [CrossRef]
- Maugendre, L., J.-P. Gattuso, J. Louis, A. de Kluijver, S. Marro, K. Soetaert, and F. Gazeau. 2015. Effect of ocean warming and acidification on a plankton community in the NW Mediterranean Sea. ICES J. Mar. Sci. 72(6):1744-1755. [CrossRef]
- Maugendre, L., J.-P. Gattuso, A. J. Poulton, W. Dellisanti, M. Gaubert, C. Guieu, and F. Gazeau. 2017. No detectable effect of ocean acidification on plankton metabolism in the NW oligotrophic Mediterranean Sea: Results from two mesocosm studies. Est. Coast. Shelf Sci. 186(A):89-99. [CrossRef]
- Mena, C., P. Reglero, M. Hidalgo, E. Sintes, R. Santiago, M. Martín, G. Moyà, and R. Balbín. 2019. Phytoplankton community structure is driven by stratification in the oligotrophic Mediterranean Sea. Front. Microbiol. 10:1698. [CrossRef]
- Mercado, J. M., C. Sobrino, P. J. Neale, M. Segovia, A. Reul, A. L. Amorim, and others. 2014. Effect of CO2, nutrients and light on coastal plankton. II. Metabolic rates. Aquat. Biol. 22:43-57. [CrossRef]
- Mesa, E., A. Delgado-Huertas, P. Carillo-de-Albornoz, L. S. García-Corral, M. Sanz-Martín, P. Wassmann, and others. 2017. Continuous daylight in the high-Arctic summer supports high plankton respiration rates compared to those supported in the dark. Sci. Rep. 7:1247. [CrossRef]
- Moreno, H. D., M. Köring, J. Di Pane, N. Tremblay, K. H. Wiltshire, M. Boersma, and C. L. Meunier. 2022. An integrated multiple driver mesocosm experiment reveals the effect of global change on planktonic food web structure. Commun. Biol. 5:179. [CrossRef]
- Motegi, C., T. Tanaka, J. Piontek, C. P. D. Brussaard, J.-P. Gattuso, and M. G. Weinbauer. 2013. Effect of CO2 enrichment on bacterial metabolism in an Arctic fjord. Biogeosciences 10:3285-3296. [CrossRef]
- Mozetič, P., C. Solidoro, G. Cossarini, G. Socal, R. Precali, J. Francé, and others. 2010. Recent trends towards oligotrophication of the Northern Adriatic: Evidence from chlorophyll a time series. Est. Coasts 33:362-375. [CrossRef]
- Murray, C. J., B. Müller-Karulis, J. Carstensen, D. J. Conley, B. G. Gustafsson, and J. H. Andersen. 2019. Past, present and future eutrophication status of the Baltic Sea. Front. Mar. Sci. 6:2. [CrossRef]
- Murrell, M. C., R. S. Stanley, J. C; Lehrter, and J. D. Hagy III. 2013. Plankton community respiration, net ecosystem metabolism, and oxygen dynamics on the Louisiana continental shelf: Implications for hypoxia. Cont. Shelf. Res. 52:27-38. [CrossRef]
- Olesen, M., C. Lundsgaard, and A. Andrushaitis. 1999. Influence of nutrients and mixing on the primary production and community respiration in the Gulf of Riga. J. Mar. Sys. 23:127-143. [CrossRef]
- Olsen, Y., S. Agustí, T. Andersen, C. M. Duarte, J. M. Gasol, I. Gismervik, and others. 2006. A comparative study of responses in planktonic food web structure and function in contrasting European coastal waters exposed to nutrient addition. Limnol. Oceanogr. 51(1):488-503. [CrossRef]
- Opdal, A. F., T. Andersen, D. O. Hessen, C. Lindemann, and D. L. Aksnes. 2023. Tracking freshwater browning and coastal water darkening from boreal forests to the Arctic Ocean. Limnol. Oceanogr. Lett. 8(4):611-619. [CrossRef]
- Opdal, A. F., C. Lindemann, T. Andersen, D. O. Hessen, Ø. Fiksen, D. L. Aksnes. 2024. Land use change and coastal water darkening drive synchronous dynamics in phytoplankton and fish phenology on centennial timescales. Glob. Change Biol. 30(5):e17308. [CrossRef]
- Ortiz, J., J. Arístegui, J. Taucher, and U. Riebesell. 2022. Artificial upwelling in singular and recurring mode: Consequences for net community production and metabolic balance. Front. Mar. Sci. 8:743105. [CrossRef]
- Ortiz, J., J. Arístegui, S. U. Goldenberg, M. Fernández-Méndez, J. Taucher, S. D. Archer, M. Baumann, and U. Riebesell. 2024. Phytoplankton physiology and functional traits under artificial upwelling with varying Si:N. Front. Mar. Sci. 10:1319875. [CrossRef]
- Oschlies, A., P. Brandt, L. Stramma, and S. Schmidtko. 2018. Drivers and mechanisms of ocean deoxygenation. Nat. Geosci. 11:467-473. [CrossRef]
- Panigrahi, S., A. Nydahl, P. Anton, and J. Wikner. 2013. Strong seasonal effect of moderate experimental warming on plankton respiration in a temperate estuarine plankton community. Est. Coast. Shelf Sci. 135:269-279. [CrossRef]
- Pastor, F., and S. Khodayar. 2023. Marine heat waves: Characterizing a major climate impact in the Mediterranean. Sci. Tot. Env. 861:160621. [CrossRef]
- Pilkaitytë, R., A. Schoor, and H. Schubert. 2004. Response of phytoplankton communities to salinity changes – a mesocosm approach. Hydrobiologia 513:27-38. [CrossRef]
- Prichett, D., J. M. Bonilla Pagan, C. L. S. Hodgkins, and J. M. Testa. 2024. Controls on water-column respiration rates in a coastal plan estuary: Insights from long-term time-series measurements. Est. Coasts 47:2542-2551. [CrossRef]
- Pringault, O., S. Tesson, and E. Rochelle-Newall. 2009. Respiration in the light and bacterio-phytoplankton coupling in a coastal environment. Microb. Ecol. 57:321-334. [CrossRef]
- Pulina, S., S. Suikkanen, B. M. Padedda, A. Brutemark, L. M. Grubisic, C. T. Satta, T. Caddeo, P. Farina, and A. Lugliè. 2020. Responses of a Mediterranean coastal lagoon plankton community to experimental warming. Mar. Biol. 167:22. [CrossRef]
- Qu, L., D. A. Campbell, and K. Gao. 2021. Ocean acidification interacts with growth light to suppress CO2 acquisition efficiency and enhance mitochondrial respiration in a coastal diatom. Mar. Poll. Bull. 163:112008. [CrossRef]
- Rabalais, N. N., R. E. Turner, R. J. Díaz, and D. Justić. 2009. Global change and eutrophication of coastal waters. 2009. ICES J. Mar. Sci. 66(7):1528-1537. [CrossRef]
- Regaudie-de-Gioux, A., and C. M. Duarte. 2012. Temperature dependance of planktonic metabolism in the ocean. Glob. Biogeochem. Cycles 26(1). [CrossRef]
- Regaudie-de-Gioux, A., and C. M. Duarte. 2013. Global patterns in oceanic planktonic metabolism. Limnol. Oceanogr. 58(3):977-986. [CrossRef]
- Regaudie-de-Gioux, A., S. Agustí, and C. M. Duarte. 2014. UV sensitivity of planktonic net community production in ocean surface waters. JGR Biogeosciences 119(5):929-936. [CrossRef]
- Robinson, C., and P. J. Le B. Williams. 2005. Respiration and its measurement in surface marine waters. In: P. A. del Giorgio, P. J. le B. Williams, and B. LE. (Eds) Respiration in aquatic ecosystems. Oxford University Press, 147-180.
- Robinson, C. 2019. Microbial respiration: the engine of ocean deoxygenation. Front. Mar. Sci. 5:533. [CrossRef]
- Serret, P., C. Robinson, M. Aranguren-Gassis, and others. 2015. Both respiration and photosynthesis determine the scaling of plankton metabolism in the oligotrophic ocean. Nat. Commun. 6: 6961. [CrossRef]
- Serret, P., D. Basso, P. Pitta, I. Magiopoulos, P. Alcaraz, A. Penin, and others. 2024. The impact of ocean liming on phytoplankton size-structure and the balance of photosynthesis and respiration in two contrasting environments. EGU General Assembly 2024, Vienna, Austria. EGU24-13093. [CrossRef]
- Smith, K. E., M. T. Burrows, A. J. Hobday, N. G. King, P. J. Moore, A. Sen Gupta, and others. 2023. Biological impacts of marine heatwaves. Ann. Rev. Mar. Sci. 15:119-145. [CrossRef]
- Solås, M. R., A. G. V. Salvanes, and D. L. Aksnes. 2024. Association between water darkening and hypoxia in a Norwegian fjord. Est. Coast. Shelf Sci. 310:108988. [CrossRef]
- Soulié, T., S. Mas, D. Parin, F. Vidussi, and B. Mostajir. 2021. A new method to estimate planktonic oxygen metabolism using high-frequency sensor measurements in mesocosm experiments and considering daytime and nighttime respirations. Limnol. Oceanogr. Methods 19(5):303-316. [CrossRef]
- Soulié, T., F. Vidussi, J. Courboulès, S. Mas, and B. Mostajir. 2022a. Metabolic responses of plankton to warming during different productive seasons in coastal Mediterranean waters revealed by in situ mesocosm experiments. Sci. Rep. 12:9001. [CrossRef]
- Soulié, T., F. Vidussi, S. Mas, and B. Mostajir. 2022b. Functional stability of a coastal Mediterranean plankton community during an experimental marine heatwave. Front. Mar. Sci. 9:831496. [CrossRef]
- Soulié, T., H. Stibor, S. Mas, B. Braun, J. Knechtel, J. C. Nejstgaard, and others. 2022c. Brownification reduces oxygen gross primary production and community respiration and changes the phytoplankton community composition: An in situ mesocosm experiment with high-frequency sensor measurements in a North Atlantic bay. Limnol. Oceanogr. 67(4):874-887. [CrossRef]
- Soulié, T., J. Engström-Öst, and O. Glippa. 2022d. Copepod oxygen consumption along a salinity gradient. Mar. Fresh. Behav. Physiol. 55(5-6):107-119. [CrossRef]
- Soulié, T., F. Vidussi, S. Mas, and B. Mostajir. 2023. Functional and structural responses of plankton communities toward consecutive experimental heatwaves in Mediterranean coastal waters. Sci. Rep. 13:8050. [CrossRef]
- Soulié, T., F. Vidussi, J. Courboulès, M. Heydon, S. Mas, F. Voron, C. Cantoni, F. Joux, and B. Mostajir. 2024. Simulated terrestrial runoff shifts the metabolic balance of a coastal Mediterranean plankton community towards heterotrophy. Biogeosciences 21:1887-1902. [CrossRef]
- Soulié, T., J. Gonzalez, P. Serret, I. G. Teixeira, C. G. Castro, S. Mas, D. Parin, F. Vidussi, and B. Mostajir. 2025. Warming enhances primary production and respiration and changes plankton community structure in an estuarine upwelling system. Limnol. Oceanogr. [CrossRef]
- Spatharis, S., G. Tsirtsis, D. B. Danielidis, T. Do Chi, and D. Mouillot. 2007. Effects of pulsed nutrient inputs on phytoplankton assemblage structure and blooms in an enclosed coastal area. Est. Coast. Shelf Sci. 73(3-4):807-815. [CrossRef]
- Spilling, K., A. J. Paul, N. Virkkala, T. Hastings, S. Lischka, A. Stuhr, and others. 2016. Ocean acidification decreases plankton respiration: evidence from a mesocosm experiment. Biogeosciences 13:4707-4719. [CrossRef]
- Staehr, P. A., and K. Sand-Jensen. 2006. Seasonal changes in temperature and nutrient control of photosynthesis, respiration and growth of natural phytoplankton communities. Fresh. Biol. 51:249-262. [CrossRef]
- Staehr, P. A., D. Bade, M. C. Van de Bogert, G. R. Koch, C. Williamson, P. Hanson, J. J. Cole, and T. Kratz. 2010. Lake metabolism and the diel oxygen technique: State of the science. Limnol. Oceanogr. Methods 8(11):628-644. [CrossRef]
- Stewart, R. I. A., M. Dossena, D. A. Bohan, E. Jeppesen, R. L. Kordas, M. E. Ledger, and others. 2013. Chapter Two -Mesocosm experiments as a tool for ecological climate-change research. Adv. Ecol. Res. 48:71-181. [CrossRef]
- Striebel, M., L. Kallajoki, C. Kunze, J. Wollschläger, A. Deininger, and H. Hillebrand. 2023. Marine primary producers in a darker future: a meta-analysis of light effects on pelagic and benthic autotrophs. Oikos 2023:e09501. [CrossRef]
- Tanaka, T., S. Alliouane, R. G. B. Bellerby, J. Czerny, A. de Kluijver, U. Riebesell, K. G. Schulz, A. Silyakova, and J.-P. Gattuso. 2013. Effect of increased pCO2 on the planktonic metabolic balance during a mesocosm experiment in an Arctic fjord. Biogeosciences 10:315-325. [CrossRef]
- Turley, C., and H. S. Findlay. 2016. Chapter 18 – Ocean acidification. In: T. M. Letcher (Ed) Climate Change (Second Edition). Elsvier, 271-293. [CrossRef]
- van Pelt, W., V. Pohjola, R. Pettersson, S. Marchenko, J. Kohler, B. Luks, and others. 2019. A long-term dataset of climatic mass balance, snow conditions, and runoff in Svalbard (1957-2018). The Cryosphere 13:2259-2280. [CrossRef]
- Vanharanta, M., M. Santoro, C. Villena-Alemany, J. Piiparinen, K. Piwosz, H.-P. Grossart, M. Labrenz, and K. Spilling. 2024. Microbial remineralization processes during postspring-bloom with excess phosphate available in the northern Baltic Sea. FEMS Microb. Ecol. 100:fiae103. [CrossRef]
- Vaquer-Sunyer, R., C. M. Duarte, R. Santiago, P. Wassmann, and M. Reigstad. 2010. Experimental evaluation of planktonic respiration response to warming in the European Arctic sector. Polar Biol. 33:1661-1671. [CrossRef]
- Vaquer-Sunyer, R., and C. M. Duarte. 2013. Experimental evaluation of the response of coastal Mediterranean planktonic and benthic metabolism to warming. Est. Coasts 36:697-707. [CrossRef]
- Vaquer-Sunyer, R., C. M. Duarte, A. regaudie-de-Gioux, J. Holding, L. S. Garcia-Corral, M. Reigstad, and P. Wassmann. 2013. Seasonal patterns in Arctic planktonic metabolism (Fram Strait - Svalbard region). Biogeosciences 10:1451-1469. [CrossRef]
- Vaquer-Sunyer, R., D. J. Conley, S. Muthusamy, M. V. Lindh, J. Pinhassi, and E. S. Kritzberg. 2015. Dissolved organic nitrogen inputs from wastewater treatment plant effluents increase responses of planktonic metabolic rates to warming. Environ. Sci. Technol. 49:11411-11420. [CrossRef]
- Vaquer-Sunyer, R., H. E. Reader, S. Muthusamy, M. V. Lindh, J. Pinhassi, D. J. Conley, and E. S. Kritzberg. 2016. Effects of wastewater treatment plant effluent inputs on planktonic metabolic rates and microbial community composition in the Baltic Sea. Biogeosciences 13:4751-4765. [CrossRef]
- Vázquez-Domínguez, E., D. Vaqué, and J. M. Gasol. 2007. Ocean warming enhances respiration and carbon demand of coastal microbial plankton. Glob. Change Biol. 13(7):1327-1334. [CrossRef]
- Vidussi, F., B. Mostajir, E. Fouilland, E. Le Floc’h, J. Nouguier, C. Roques, P. Got, D. Thibault-Botha, T. Bouvier, and M. Troussellier. 2011. Effects of experimental warming and increased ultraviolet B radiation on the Mediterranean plankton food web. Limnol. Oceanogr. 56(1):206-218. [CrossRef]
- Weber, T. S., and C. Deutsch. 2010. Ocean nutrient ratios governed by plankton biogeography. Nature 467:550-554. [CrossRef]
- Wernberg, T., D. A. Smale, and M. S. Thomsen. 2012. A decade of climate change experiments on marine organisms: procedures, patterns and problems. Glob. Change Biol. 18(5):1491-1498. [CrossRef]
- Wikner, J., K. Vikström, and A. Verma. 2023. Regulation of marine plankton respiration: A test of models. Front. Mar. Sci. 10:1134699. [CrossRef]
- Wilson, J. M., S. S. Abboud, and J. M. Beman. 2024. Effects of experimental nutrient enrichment and eutrophication on microbial community structure and function in “marine lakes”. Elem. Sci. Anth. 12:1. [CrossRef]
- Wohlers, J., A. Engel, A. Zöllner, P. Breithaupt, K. Jürgens, H.-G. Hoppe, and others. 2009. Changes in biogenic carbon flow in response to sea surface warming. Proc. Natl. Acad. Sci. USA 106:7067-7072. [CrossRef]
- Wolf, K. K. E., C. J. M. Hoppe, L. Rehder, E. Schaum, U. John, and B. Rost. 2024. Heatwave responses of Arctic phytoplankton communities are driven by combined impacts of warming and cooling. Sci. Adv. 10:eadl5904. [CrossRef]
- Yvon-Durocher, G., J. M. Caffrey, A. Cescatti, M. Dossena, P. del Giorgio, J. M. Gasol, and others. 2012. Reconciling the temperature dependence of respiration across timescales and ecosystem types. Nature 487:472-476. [CrossRef]
- Zhu, Z.-Y., J. Hu, G.-D. Song, Y. Wu, J. Zhang, and S.-M. Liu. 2016. Phytoplankton-driven dark plankton respiration in the hypoxic zone off the Changjiang Estuary, revealed by in vitro incubations. J. Mar. Sys. 154(A):50-56. [CrossRef]
Figure 1.
Location of the experiments testing the effects of climate change disturbances on plankton community respiration (PCR) in coastal waters of Europe. The ‘multiple drivers’ category encompasses locations in which multiple disturbances have been tested (either simultaneously or during different experiments).
Figure 1.
Location of the experiments testing the effects of climate change disturbances on plankton community respiration (PCR) in coastal waters of Europe. The ‘multiple drivers’ category encompasses locations in which multiple disturbances have been tested (either simultaneously or during different experiments).
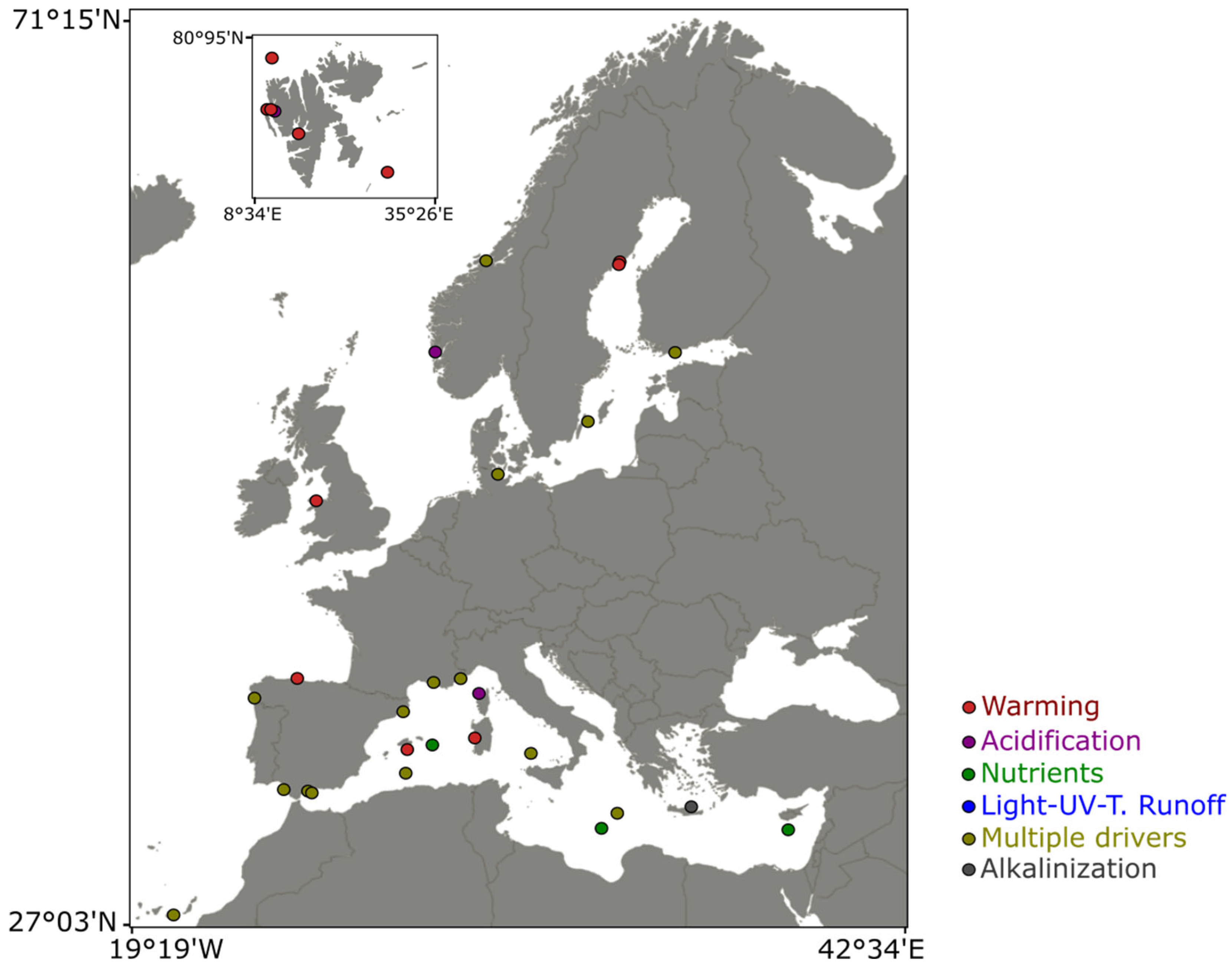
Figure 2.
Number of studies describing experiments in which the effects of climate change disturbances on plankton community respiration (PCR) were investigated in coastal waters of Europe, separated based on tested disturbance (A.), studied area (B.), or year of publications (C.).
Figure 2.
Number of studies describing experiments in which the effects of climate change disturbances on plankton community respiration (PCR) were investigated in coastal waters of Europe, separated based on tested disturbance (A.), studied area (B.), or year of publications (C.).
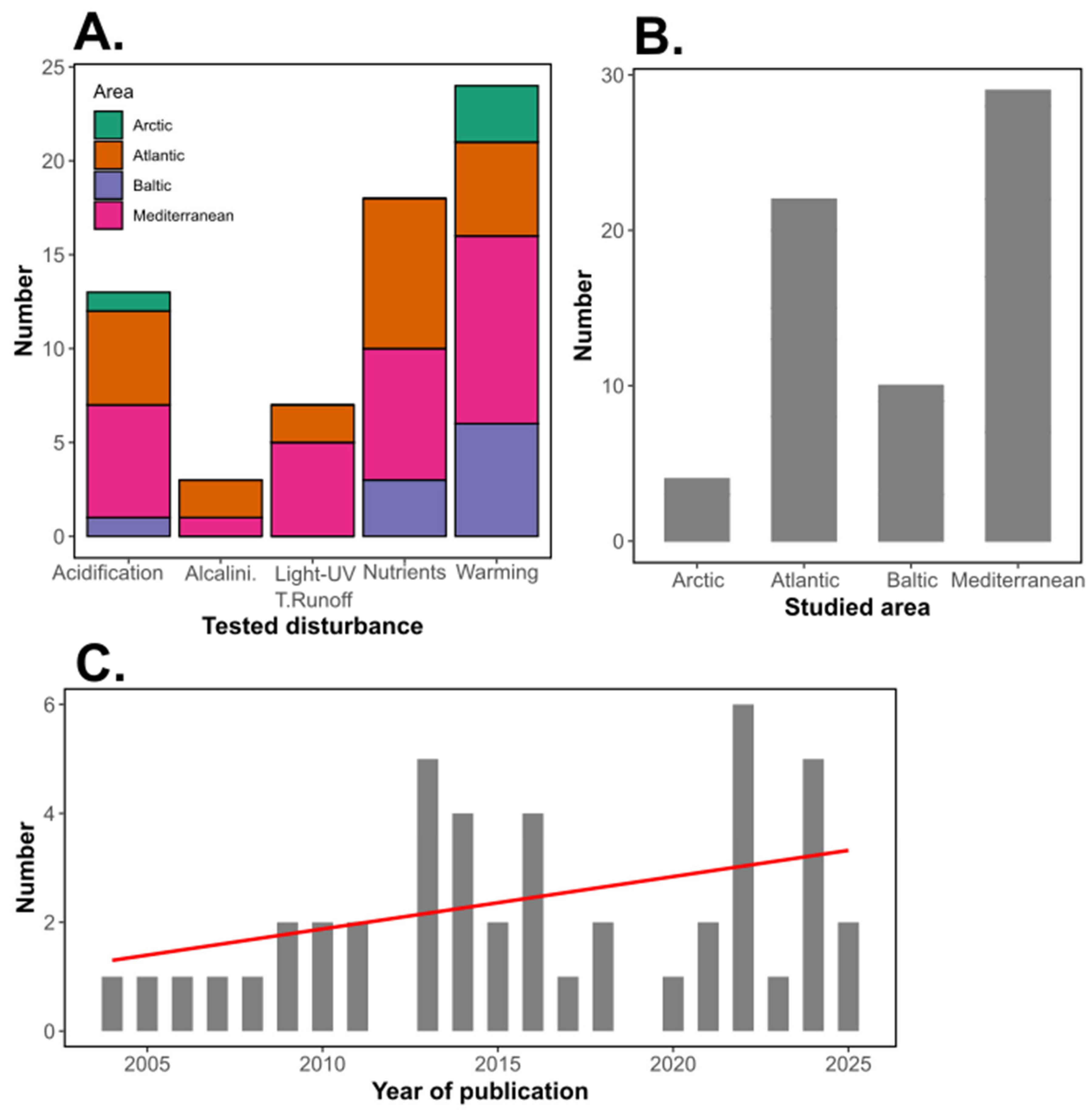

Table 1.
Summary of all the studies included in the present review, with their type of disturbance studied, their study site and the broader area of their study site.
Table 1.
Summary of all the studies included in the present review, with their type of disturbance studied, their study site and the broader area of their study site.
| Study | Disturbance | Study site | Area | |
| 1 | Vaquer-Sunyer et al., (2010) | Warming | Svalbard | Arctic |
| 2 | Holding et al., (2013) | Warming | Svalbard | Arctic |
| 3 | Wolf et al., (2024) | Warming | Svalbard | Arctic |
| 4 | Hoppe et al., (2008) | Warming | Kiel fjord | Baltic |
| 5 | Cabrerizo et al., (2021) | Warming | Gulf of Bothnia | Baltic |
| 6 | Panigrahi et al., (2013) | Warming | Bothnian Sea | Baltic |
| 7 | Lewandowska et al., (2014) | Warming | Baltic Sea | Baltic |
| 8 | Wohlers et al., (2009) | Warming | Western Baltic Sea | Baltic |
| 9 | Soulié et al., (2022a) | Warming | Thau Lagoon | Mediterranean |
| 10 | Vázquez-Domínguez et al., (2007) | Warming | Bay of Blanes | Mediterranean |
| 11 | Vaquer-Sunyer & Duarte (2013) | Warming | Majorca | Mediterranean |
| 12 | Soulié et al., (2022b) | Warming | Thau Lagoon | Mediterranean |
| 13 | Soulié et al., (2023) | Warming | Thau Lagoon | Mediterranean |
| 14 | Pulina et al., (2020) | Warming | Cabras lagoon | Mediterranean |
| 15 | Maugendre et al., (2015) | Warming; Acidification | Bay of Villefranche | Mediterranean |
| 16 | Huete-Stauffer et al., (2018) | Warming | Bay of Biscay | Atlantic |
| 17 | Soulié et al., (2025) | Warming | Ría de Vigo | Atlantic |
| 18 | López-Sandoval et al., (2025) | Warming; Nutrients | Ría de Vigo | Atlantic |
| 19 | Vaquer-Sunyer et al, (2015) | Warming; Nutrients | Baltic Proper | Baltic |
| 20 | Tanaka et al., (2013) | Acidification | Svalbard | Arctic |
| 21 | Spilling et al., (2016) | Acidification | Storfjärden, Finland | Baltic |
| 22 | Maugendre et al., (2017) | Acidification | Bay of Villefranche; Bay of Calvi | Mediterranean |
| 23 | Delille et al., (2005) | Acidification | Raunefjporden, Norway | Atlantic |
| 24 | Egge et al., (2009) | Acidification | Raunefjporden, Norway | Atlantic |
| 25 | Filella et al., (2018) | Acidification; Nutrients | Taliarte, Gran Canaria | Atlantic |
| 26 | Martínez-García et al., (2013) | Nutrients | Ría de Vigo | Atlantic |
| 27 | Vaquer-Sunyer et al., (2016) | Nutrients | Baltic Sea | Baltic |
| 28 | Olsen et al., (2006) | Nutrients | Tvarminne, Blanes Bay, Hopavågen Bay | Baltic, Mediterranean, Atlantic |
| 29 | Lagaria et al., (2011) | Nutrients | Mediterranean | Mediterranean |
| 30 | Duarte et al., (2004) | Nutrients | Bay of Blanes | Mediterranean |
| 31 | Cabrerizo et al., (2022) | Warming; Nutrients | Coastal waters of the Mediterranean and the Atlantic | Mediterranean, Atlantic |
| 32 | Martínez-García et al., (2014) | Nutrients | Ría de Vigo | Atlantic |
| 33 | Baños et al., (2022) | Nutrients | Gando Bay, Canary | Atlantic |
| 34 | Ortiz et al., (2022) | Nutrients | Gando Bay, Canary | Atlantic |
| 35 | Cabrerizo et al., (2016) | UV; Nutrients | SW Mediterreanean Sea | Mediterranean |
| 36 | Lekunberri et al., (2010) | Nutrients | Bay of Blanes | Mediterranean |
| 37 | Ortiz et al., (2024) | Nutrients | Gando Bay, Canary | Atlantic |
| 38 | Soulié et al., (2024) | Terrestrial runoff | Thau Lagoon | Mediterranean |
| 39 | Liess et al., (2016) | Terrestrial runoff | Thau Lagoon | Mediterranean |
| 40 | Soulié et al., (2022c) | Terrestrial runoff | Hopavågen Bay | Atlantic |
| 41 | Agustí et al., (2014) | Light, UV | Majorca | Mediterranean |
| 42 | Marín-Samper et al., (2024) | Alkalinity | Taliarte, Gran Canria | Atlantic |
| 43 | Serret et al., (2024) | Alkalinity | Ría de Vigo, Cretan Sea | Atlantic; Mediterranean |
| 44 | Mercado et al., (2014) | Acidification; Nutrients; Light | Fuengirola, southern Spain | Mediterranean |
| 45 | Gazeau et al., (2021) | Warming, Acidification, Nutrients | Mediterranean | Mediterranean |
| 46 | Vidussi et al., (2011) | Warming; UV | Thau Lagoon | Mediterranean |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



