
Submitted:
07 November 2025
Posted:
07 November 2025
You are already at the latest version
Abstract
The CXC chemokine receptor 2 (CXCR2) is a member of the G-protein-coupled receptor superfamily and regulates a diverse range of immune responses and tumor progression. CXCR2 is expressed on immune cells, especially neutrophils, and is involved in various immune responses by interacting with its chemokine ligands. Therefore, the development of sensitive monoclonal antibodies (mAbs) for CXCR2 has been desired for treatment and diagnosis. This study established a novel sensitive anti-mouse CXCR2 (mCXCR2) mAb; Cx2Mab-5 (rat IgG2a, kappa), using the mCXCR2 synthetic N-terminus peptide immunization method. In flow cytometry, Cx2Mab-5 recognized mCXCR2-overexpressed Chinese hamster ovary-K1 cells (CHO/mCXCR2) and WEHI-3B (murine myelomonocytic leukemia cell) cells, which express endogenous mCXCR2. Cx2Mab-5 did not cross-react with other mouse CC, CXC, CX3C, and XC chemokine receptors. Cx2Mab-5 showed a moderate binding affinity for both CHO/mCXCR2 and WEHI-3B. Further-more, Cx2Mab-5 detected mCXCR2 in western blot and immunohistochemistry in CHO/mCXCR2 cells. Hence, Cx2Mab-5 can be a valuable tool for analyzing mCXCR2-expressing cells, such as immune cells and tumors.
Keywords:
chemokine receptor
; CXCR2
; monoclonal antibody
; flow cytometry
1. Introduction
Chemokines are small chemoattractant proteins secreted by various cells, including immune-related cells, that play essential roles in a myriad physiological activities, such as cellular migration, invasion, and inflammatory responses, by activating chemokine receptors [1,2]. Chemokine receptors belong to the superfamily of G-protein-coupled receptors (GPCRs), and can be classified into four different subfamilies: CC, CXC, CX3C, and XC, depending on the number and position of cysteine residues in their N-terminus. Chemokine receptors display the seven transmembrane structures characteristic of typical GPCRs [3]. Chemokines-chemokine receptors signal contributes to various aspects of our immune response, and their aberrant signaling is implicated in multiple diseases, including allergy [4], autoimmune diseases [5], and cancer [6].
Chemokine concentration gradients enhance the migration of immune cells. Several chemokines, including CXCL1, CXCL2, CXCL3, CXCL5, CXCL6, CXCL7 in mice and CXCL8 in humans, have been identified as ligands for CXCR2 [7,8]. Although the chemokines for CXCR2 have low sequence identity, with less than 25%, they share structural similarities [9]. CXCR2 and CXCR1 share common ligands, CXCL6 and CXCL8 in humans and CXCL5 and CXCL6 in mice [7]. CXCR2 contributes to tumorigenesis through recruitment of immune cells, such as myeloid-derived suppressor cells (MDSCs) and tumor-associated neutrophils, autocrine effects on the tumor, and angiogenesis by interacting with these chemokines [10,11]. CXCR2 is considered a key regulator of the tumor microenvironment (TME). CXCR2 also plays a crucial role in antibacterial defense in normal tissues. The higher expression of CXCR2 is confirmed in neutrophils in both humans and mice. Cxcr2 knockout (KO) mice decreased mature neutrophils in the spleen. Many parameters of fine splenic neutrophils decline in Cxcr2 KO mice, including the phagocytic ability, reactive oxygen species production, and the level of F-actin and α-tubulin [12]. Single-cell RNA sequencing analysis revealed the eight clusters of neutrophil maturity in infected mice. Among these clusters, the most mature neutrophil showed higher CXCR2 expression in peripheral blood and spleen [13]. Interestingly, neutrophil function is influenced by circadian rhythms. Bmal1, a well-known clock gene, upregulates the CXCL2 expression to activate CXCR2, followed by diurnal changes of neutrophils correlated with anti-microbial defense [14]. Furthermore, CXCR2-mediated epithelial and endothelial CXCL1 transcytosis contributes to the bacterial clearance by recruitment of neutrophils into the lung [15]. Overall, CXCR2 plays a key role in antibacterial responses.
Numerous reports have identified CXCR2 and its chemokine ligands as significant regulators of tumor growth in various cancers. The high expression of CXCR2 is associated with worse tumor grade and overall survival in triple-negative breast cancer (TNBC) [16]. M2 macrophages-secreting CXCL1 and CXCL5 promote the expression of PD-L1 in TNBC [17] and epithelial-mesenchymal transition (EMT) in prostate cancer [18], respectively. Renal cell carcinoma secreting transforming growth factor-β transforms normal fibroblasts into cancer-associated fibroblasts (CAFs). Transformed CAFs provide CXCL3 to RCC and induce EMT and cell stemness [19]. Not only does the function of CXCR2 expressed on cancer cells, but also cancer cells evade immune attack by regulating CXCR2 on immune cells. In pancreatic cancer, CXCL7 secreted by macrophages binds to CXCR2 on CD8+ T cells resulting in exhaustion through AKT/mTOR pathway [20]. CXCL6 recruits CXCR2+ mast cells with high levels of PD-L2 expression to osteosarcoma tissue, thereby creating an immunosuppressive TME [21]. CXCR2 is intricately involved in the formation of the TME through its ligand combinations and functions in cancer and immune cells. Therefore, CXCR2 is considered a promising target for cancer treatment and diagnosis. For tumor therapy, CXCR2 is often expressed in chimeric antigen receptor-T (CAR-T) cells, enabling them to home to CXCR2 ligand-secreting solid tumors [22,23].
We have previously developed many monoclonal antibodies (mAbs) against chemokine receptors using the peptide immunization method and the Cell-Based Immunization and Screening method [24,25,26,27,28,29,30,31]. Applying these methods enables efficient isolation of antibodies with diverse binding modes, including conformational and linear epitopes. In this study, we successfully developed novel anti-mouse CXCR2 (mCXCR2) mAbs using the synthetic peptide immunization method.
2. Materials and Methods
2.1. Antibodies
The anti-mCXCR2 mAb (clone SA045E1) was purchased from BioLegend (San Diego, CA, USA). The anti-DYKDDDDK mAb (clone 1E6) was purchased from FUJIFILM Wako Pure Chemical Corporation (Wako; Osaka, Japan). An anti-isocitrate dehydrogenase 1 (IDH1) mAb (clone RcMab-1) was developed previously in our lab [32]. The secondary Alexa Fluor 488-conjugated anti-rat IgG was purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA). Secondary horseradish peroxidase-conjugated anti-mouse IgG and anti-rat IgG were obtained from Agilent Technologies Inc. (Santa Clara, CA, USA) and Merck KGaA (Darmstadt, Germany), respectively.
2.2. Preparation of Cell Lines
Chinese hamster ovary (CHO)-K1 and P3X63Ag8U.1 (P3U1) cells were obtained from the American Type Culture Collection (Manassas, VA, USA). The WEHI-3B (murine myelomonocytic leukemia cell) cell line was obtained from the Cell Resource Center for Biomedical Research, Institute of Development, Aging and Cancer, Tohoku University (Miyagi, Japan). Furthermore, the cDNAs of mouse CXCR2 (mCXCR2; pCMV6neo-mCXCR2-Myc-DDK, accession No.: NM_009909.3, catalog No.: MR227587) were purchased from OriGene Technologies, Inc. (Rockville, MD, USA). The mCXCR2 expression plasmid was transfected into the cell line using a Neon transfection system (Thermo Fisher Scientific Inc., Waltham, MA, USA). Subsequently, CHO-K1 which stably overexpressed mCXCR2 with C-terminal Myc-DDK tags (CHO/mCXCR2) was established through cell sorting, using an anti-mCXCR2 mAb (clone SA045E1; BioLegend, San Diego, CA, USA) and a cell sorter (SH800; Sony Corp., Tokyo, Japan), following cultivation in a medium containing 0.5 mg/mL G418 (Nacalai Tesque, Inc., Kyoto, Japan). These cells and other mouse chemokine receptor-expressed CHO-K1 cells (e.g., CHO/mCXCR1) were established and cultured as previously reported [33].
2.3. Production of Hybridomas
The animal experiments were conducted in accordance with relevant guidelines and regulations to minimize animal suffering and distress in the laboratory. The Animal Care and Use Committee of Tohoku University approved animal experiments (Permit number: 2022MdA-001). A 6-week-old female Sprague–Dawley (Jcl:SD) rat (CLEA, Japan Tokyo, Japan) was immunized via the intraperitoneal route with keyhole limpet hemocyanin (KLH)-conjugated mCXCR2 peptide (MGEFKVDKFNIEDFFSGDLC, 100 μg of peptide/time, Eurofins Genomics KK, Tokyo, Japan) as previously reported [33]. The hybridoma supernatants were screened using enzyme-linked immunosorbent assay (ELISA) and flow cytometric analysis.
2.4. ELISA
A synthesized mCXCR2 peptide (MGEFKVDKFNIEDFFSGDLC, Eurofins Genomics KK)-immobilized immunoplates were incubated with hybridoma supernatants, followed by peroxidase-conjugated anti-rat immunoglobulins (1:2000 dilution; Agilent Technologies Inc., Santa Clara, CA, USA). Afterward, enzymatic reactions were conducted using the ELISA POD substrate TMB kit (Nacalai Tesque, Inc.), and the optical density was measured as previously reported [33].
2.5. Flow Cytometry
CHO-K1, CHO/mCXCR2, and WEHI-3B cells were harvested after brief exposure to 1 mM ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque, Inc.). The cells were washed with 0.1% BSA in PBS and treated with primary mAbs for 30 min at 4 °C. For the peptide-blocking assay, Cx2Mab-5 or SA045E1 were pre-incubated with mCXCR2 peptide or dimethyl sulfoxide (DMSO) for 15 min at room temperature, and further incubated with CHO/mCXCR2 and WEHI-3B for 30 min at 4 °C.Afterward, cells were treated with Alexa Fluor 488-conjugated anti-rat IgG (1:1000 dilution), and fluorescence data were collected using an SA3800 Cell Analyzer (Sony Corp., Tokyo, Japan).
2.6. Determination of the Binding Affinity by Flow Cytometry
CHO/mCXCR2 and WEHI-3B cells were suspended in 100 μL serially diluted anti-mCXCR2 mAbs, after which Alexa Fluor 488-conjugated anti-rat IgG (1:200 dilution) was added. The dissociation constant (KD) was using GraphPad PRISM 6 (GraphPad Software, Inc., La Jolla, CA, USA) as previously reported [33].
2.7. Western Blot Analysis
Western blot analysis were performed using 5 μg/mL of Cx2Mab-5, 5 μg/mL of SA045E1, 1 μg/mL of an anti-IDH1 mAb (clone RcMab-1), or 0.5 μg/mL anti-DYKDDDDK (clone 1E6) mAb as described previously [25].
2.8. Immunohistochemical Analysis
The CHO/mCXCR2 and CHO-K1 cell blocks were prepared using iPGell (Genostaff Co., Ltd., Tokyo, Japan). The staining was performed using 5 μg/mL of Cx2Mab-5 and 5 μg/mL of SA045E1 as described previously [34].
3. Results
3.1. Development of Anti-mCXCR2 mAbs
To develop anti-mCXCR2 mAbs, we employed the mCXCR2 N-terminus peptide immunization method. Hybridomas were screened by ELISA and flow cytometric methods (Figure 1). Briefly, a rat was immunized with KLH-conjugated mouse CXCR2 synthetic peptide by intraperitoneal injection. Two days after the final boost injection, rat spleen and myeloma cells were fused using PEG1500 to generate hybrida. Subsequently, hybridomas were seeded into 96-well plates, and an ELISA assay was used to identify positive wells for the mCXCR2 peptide. This was followed by the selection of CHO/mCXCR2-reactive and parental CHO-K1-nonreactive supernatants using flow cytometry. Afterward, we obtained several CHO/mCXCR2-reactive supernatants following the establishment of Cx2Mab-5 (rat IgG2a, kappa) by cloning via limiting dilution and additional evaluations.
3.2. Flow Cytometry Using Cx2Mab-5
Flow cytometry was conducted using Cx2Mab-5 and commercially available anti-mCXCR2 mAb SA045E1 against CHO/mCXCR2, CHO-K1, and WEHI-3B. Results showed that Cx2Mab-5 dose-dependently recognized CHO/mCXCR2 (Figure 2A), but not CHO-K1 (Figure 2B). Another anti-mCXCR2 mAb, SA045E1 from BioLegend, also dose-dependently recognized CHO/mCXCR2 (Figure 2A), but not CHO-K1 (Figure 2B). Both Cx2Mab-5 and SA045E1 also reacted dose-dependently with endogenous mCXCR2-expressing WEHI-3B (Figure 2C). Thus, Cx2Mab-5 could recognize exogenously and endogenously expressing mCXCR2 in flow cytometry.
3.3. The Specificity of Cx2Mab-5
We have established anti-mouse CC, CXC, CX3C, and XC chemokine receptor-expressed CHO-K1 cells in previous study [33]. Using these eighteen cell lines, the specificity of Cx2Mab-5 was evaluated in Figure 3. The results described that Cx2Mab-5 recognized only CHO/mCXCR2, but not other chemokine receptors (Figure 3A). We confirmed the expression of each chemokine receptors by treatment with corresponding mAbs (Figure 3B).
We next evaluated antibody reactivity using a peptide-blocking assay with the mCXCR2 peptide. As shown in Figure 4, Both Cx2Mab-5 and SA045E1 reacted with CHO/mCXCR2 and WEHI-3B cells. The reactivity of Cx2Mab-5 was completely blocked by the mCXCR2 peptide, indicating that its recognition is mediated through binding to the N-terminus of mCXCR2. Similarly, SA045E1’s reactivity was entirely neutralized by the mCXCR2 peptide. These findings demonstrate that the N-terminus of mCXCR2 is the epitope recognized by SA045E1. Taken together, Cx2Mab-5 is an anti-mCXCR2 specific mAb that recognizes the N-terminus of mCXCR2.
3.4. Determination of the Binding Affinity of Cx2Mab-5
The binding affinity of Cx2Mab-5 was assessed with CHO/mCXCR2 and WEHI-3B using flow cytometry. Results showed that the KD of Cx2Mab-5 and SA045E1 for CHO/mCXCR2 were 3.0 × 10−8 M and 1.5 × 10−8 M, respectively (Figure 5A). The KD of Cx2Mab-5 and SA045E1 for WEHI-3B were 9.3 × 10−9 M and 1.3 × 10−8 M, respectively (Figure 5B). The experiment was conducted at least twice, yielding similar results each time (Supplemental Figure 1). These results indicated that Cx2Mab-5 had a moderate affinity for CHO/mCXCR2 and WEHI-3B cells, almost equivalent to that of SA045E1.
3.5. Western Blot Analyses Using Cx2Mab-5
We investigated whether Cx2Mab-5 can be used for western blot analysis by analyzing CHO-K1 and CHO/mCXCR2 cell lysates. As shown in Figure 6, Cx2Mab-5 could clearly detect mCXCR2 as around a 48-63 kDa band in CHO/mCXCR2 cell lysates, while no band around the predicted size was detected in parental CHO-K1 cells. In contrast, another anti-mCXCR2 mAb, SA045E1, could not detect any signals on the membrane. An anti-DYKDDDDK mAb (1E6) was used as a positive control and was also able to detect a band in CHO/mCXCR2 cell lysates. An anti-IDH1 mAb (RcMab-1) was used for internal control. These results indicate that Cx2Mab-5 can detect mCXCR2 in western blot analysis.
3.6. Immunohistochemistry Using Cx2Mab-5
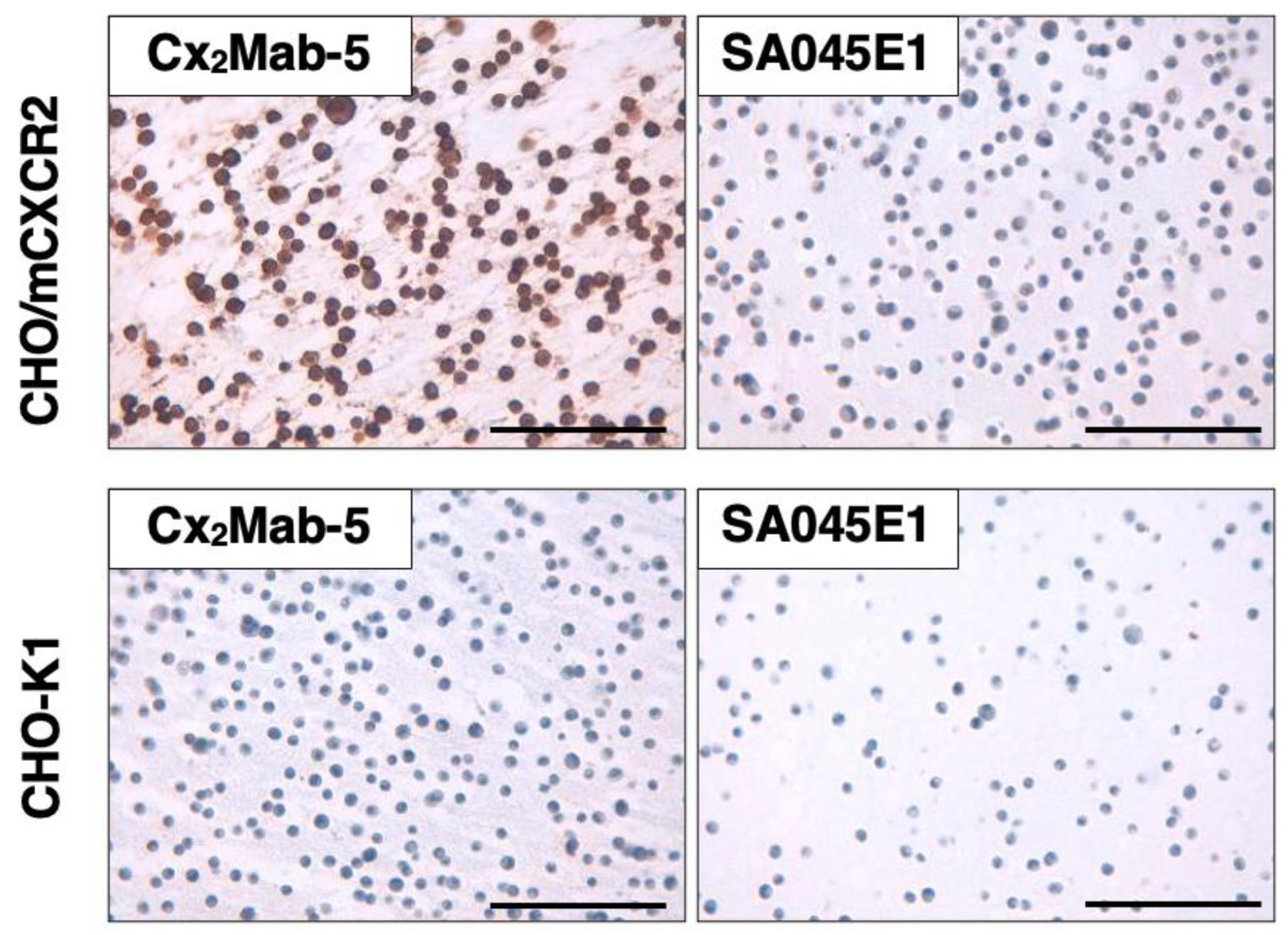
To investigate whether Cx2Mab-5 can be used for immunohistochemistry (IHC), paraffin-embedded CHO-K1 and CHO/mCXCR2 sections were stained with Cx2Mab-5. Apparent membranous staining by Cx2Mab-5 was observed in CHO/mCXCR2 (Figure 7, left upper). The SA045E1, another anti-mCXCR2 mAb, did not stain CHO/mCXCR2 sections (Figure 7, right upper). Neither Cx2Mab-5 nor SA045E1 reacted with the CHO-K1 section. (Figure 7 lower). Additionally, Cx2Mab-5 slightly stained the WEHI-3B section (Supplemental Figure 2). These results indicate that Cx2Mab-5 is suitable for IHC detection of mCXCR2-positive cells in paraffin-embedded tissue samples.
4. Discussion
Myeloid expansion in peripheral blood, as indicated by a high neutrophil-to-lymphocyte ratio, associates with shorter survival and treatment resistance in cancer patients [35,36]. The myeloid chemotaxis inhibition by CXCR2 blockade has an impact on reversing therapy resistance in a subset of patients with metastatic castration-resistant prostate cancer [37]. Since CXCR2 binds multiple chemokines, it is thought to govern a variety of immune responses that are deeply involved in cancer-related inflammation [38].
An immunosuppressive neutrophil subpopulation has been observed within human and mouse glioma, one of the most aggressive tumors [39,40]. The CXCR2+ neutrophils, which are recruited by glioma-secreting CXCL1, establish the immunosuppressive TME. Genetic depletion of these neutrophils improves antitumor immunity and prolongs survival [40]. MDSCs are also remarkable immunosuppressive cells in the TME [41]. The inhibition of CXCR2 is reportedly a promising treatment for controlling polymorphonuclear-MDSCs accumulation associated with tumor progression [42,43]. Furthermore, the blockade of granulocytic MDSCs by CXCR2 inhibition restores macrophage capacity for tumor clearance [44]. Thus, CXCR2 is a key immunomodulatory factor with potential for next-generation cancer therapy.
To date, the development of CXCR2 inhibitors has progressed, and several clinical trials have been conducted for the treatment of cancer and immune diseases. Blocking CXCR2 (and CXCR1) itself or its ligands is considered a therapeutic target. SB225002, a CXCR2-selective inhibitor, decreases cell viability in colorectal cancer cells, but not reparixin, a CXCR1/CXCR2 noncompetitive allosteric inhibitor [45]. The treatments with SB225002 and anti-PD-1 mAb improve the therapeutic index of radiotherapy in mice xenograft models [46]. A fully humanized anti-CXCL8 mAb BMS-986253, which aims to inhibit CXCR2 ligand, is undergoing clinical trials in metastatic or unresectable solid tumors [47]. The diverse range of options, including compounds and antibodies targeting CXCR2 and its ligands, increases the potential for developing therapeutic drugs. In fact, numerous clinical trials have been conducted for various diseases, including tumors, viral infections, diabetes, and chronic obstructive pulmonary disease [8]. Treatment of anti-CXCR2 antibodies may show dramatic therapeutic effects, especially when combined with other immune checkpoint inhibitors, such as anti-PD-1 mAb [42,43].
We will investigate whether the established Cx2Mab-5 exerts antitumor effects by inhibiting CXCR2-expressing cancers or immune cells, such as neutrophils, in mouse models. Moreover, the N-terminus of both the chemokines and the receptors is necessary for their mutual binding [7,48,49]. Antitumor effects have also been observed with the administration of the CXCR2 antibody alone in a human pancreatic cancer xenograft model, which was generated to target the N-terminus of human CXCR2 by phage display technology [50]. We developed Cx2Mab-5 by N-terminus mCXCR2 peptide immunization (Figure 1). It is anticipated that future studies utilizing mouse models will determine whether Cx2Mab-5 inhibits CXCR2 activation by chemokine ligands and exhibits antitumor activity.
Tumor therapy with CAR-T cells has been developed, primarily for B-cell malignancies [51,52]. However, CAR-T cell therapy for solid tumors has limited effectiveness and low antitumor activity in early-phase clinical studies [53,54]. To address this problem, modifications are being made to introduce chemokine receptors, such as CXCR2 and CXCR6, into CAR-T cells to enhance their homing ability to solid tumors, including B7-H3 CAR-T for pediatric sarcoma [22,23], glypican 2 CAR-T for neuroblastoma [55], claudin 18.2 CAR-T for pancreatic ductal adenocarcinoma [56], and HER2 CAR-T [57]. Furthermore, CXCR2-armed CAR-natural killer cells targeting glypican 3 demonstrate antitumor efficacy against hepatocellular carcinoma in combination with radiotherapy [58]. Similar to CXCR2 as a target for cancer treatment, further applications exploiting its unique function are expected in the future. Therefore, more fundamental research on CXCR2 is necessary. Cx2Mab-5 is suitable for various experiments, including flow cytometry (Figure 2, Figure 3, Figure 4 and Figure 5), Western blotting (Figure 6), and immunohistochemistry (Figure 7), and will be a valuable tool for elucidating the function of CXCR2 in preclinical models.
Funding
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP25am0521010 (to Y.K.), JP25ama121008 (to Y.K.), JP25ama221339 (to Y.K.), and JP25bm1123027 (to Y.K.), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) grant no. 25K10553 (to Y.K.).
Author Contributions
Honoka Taruta Investigation Tomohiro Tanaka: Investigation, Funding acquisition, Writing – original draft Mika K. Kaneko: Conceptualization Hiroyuki Suzuki: Writing – review and editing Yukinari Kato: Conceptualization, Funding acquisition, Project administration, Writing – review and editing All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
The animal study protocol was approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2022MdA-001) for studies involving animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
All related data and methods are presented in this paper. Additional inquiries should be addressed to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest involving this article.
References
- López-Cotarelo, P.; Gómez-Moreira, C.; Criado-García, O.; Sánchez, L.; Rodríguez-Fernández, J.L. Beyond Chemoattraction: Multifunctionality of Chemokine Receptors in Leukocytes. Trends in Immunology 2017, 38, 927-941. [CrossRef]
- Nagarsheth, N.; Wicha, M.S.; Zou, W. Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy. Nat Rev Immunol 2017, 17, 559-572. [CrossRef]
- Rajagopalan, L.; Rajarathnam, K. Structural basis of chemokine receptor function--a model for binding affinity and ligand selectivity. Biosci Rep 2006, 26, 325-339. [CrossRef]
- Kaplan, A.P. Chemokines, chemokine receptors and allergy. Int Arch Allergy Immunol 2001, 124, 423-431. [CrossRef]
- Shachar, I.; Karin, N. The dual roles of inflammatory cytokines and chemokines in the regulation of autoimmune diseases and their clinical implications. J Leukoc Biol 2013, 93, 51-61. [CrossRef]
- O’Hayre, M.; Salanga, C.L.; Handel, T.M.; Allen, S.J. Chemokines and cancer: migration, intracellular signalling and intercellular communication in the microenvironment. Biochem J 2008, 409, 635-649. [CrossRef]
- Saha, S.; Sano, F.K.; Sharma, S.; Ganguly, M.; Mishra, S.; Dalal, A.; Akasaka, H.; Kobayashi, T.A.; Zaidi, N.; Tiwari, D.; et al. Molecular basis of promiscuous chemokine binding and structural mimicry at the C-X-C chemokine receptor, CXCR2. Mol Cell 2025, 85, 976-988.e979. [CrossRef]
- Lazennec, G.; Rajarathnam, K.; Richmond, A. CXCR2 chemokine receptor - a master regulator in cancer and physiology. Trends Mol Med 2024, 30, 37-55. [CrossRef]
- Martin, P.; Kurth, E.A.; Budean, D.; Momplaisir, N.; Qu, E.; Simien, J.M.; Orellana, G.E.; Brautigam, C.A.; Smrcka, A.V.; Haglund, E. Biophysical characterization of the CXC chemokine receptor 2 ligands. PLoS One 2024, 19, e0298418. [CrossRef]
- Lazennec, G.; Richmond, A. Chemokines and chemokine receptors: new insights into cancer-related inflammation. Trends in Molecular Medicine 2010, 16, 133-144. [CrossRef]
- Yang, C.; Geng, H.; Yang, X.; Ji, S.; Liu, Z.; Feng, H.; Li, Q.; Zhang, T.; Zhang, S.; Ma, X.; et al. Targeting the immune privilege of tumor-initiating cells to enhance cancer immunotherapy. Cancer Cell 2024, 42, 2064-2081.e2019. [CrossRef]
- Delobel, P.; Ginter, B.; Rubio, E.; Balabanian, K.; Lazennec, G. CXCR2 intrinsically drives the maturation and function of neutrophils in mice. Front Immunol 2022, 13, 1005551. [CrossRef]
- Xie, X.; Shi, Q.; Wu, P.; Zhang, X.; Kambara, H.; Su, J.; Yu, H.; Park, S.-Y.; Guo, R.; Ren, Q.; et al. Single-cell transcriptome profiling reveals neutrophil heterogeneity in homeostasis and infection. Nature Immunology 2020, 21, 1119-1133. [CrossRef]
- Adrover, J.M.; Del Fresno, C.; Crainiciuc, G.; Cuartero, M.I.; Casanova-Acebes, M.; Weiss, L.A.; Huerga-Encabo, H.; Silvestre-Roig, C.; Rossaint, J.; Cossío, I.; et al. A Neutrophil Timer Coordinates Immune Defense and Vascular Protection. Immunity 2019, 50, 390-402.e310. [CrossRef]
- Thomas, K.; Rossaint, J.; Ludwig, N.; Mersmann, S.; Kötting, N.; Grenzheuser, J.; Schemmelmann, L.; Oguama, M.; Margraf, A.; Block, H.; et al. Alveolar epithelial and vascular CXCR2 mediates transcytosis of CXCL1 in inflamed lungs. Nat Commun 2025, 16, 4846. [CrossRef]
- Chu, H.; Li, W.; Li, H. C-X-C motif chemokine receptor type 2 correlates with higher disease stages and predicts worse prognosis, and its downregulation enhances chemotherapy sensitivity in triple-negative breast cancer. Translational Cancer Research 2020, 9, 840-848.
- Zhang, L.; Gu, S.; Wang, L.; Zhao, L.; Li, T.; Zhao, X.; Zhang, L. M2 macrophages promote PD-L1 expression in triple-negative breast cancer via secreting CXCL1. Pathol Res Pract 2024, 260, 155458. [CrossRef]
- Wu, T.; Wang, W.; Shi, G.; Hao, M.; Wang, Y.; Yao, M.; Huang, Y.; Du, L.; Zhang, X.; Ye, D.; et al. Targeting HIC1/TGF-β axis-shaped prostate cancer microenvironment restrains its progression. Cell Death & Disease 2022, 13, 624. [CrossRef]
- Wang, Y.; Ding, W.; Hao, W.; Gong, L.; Peng, Y.; Zhang, J.; Qian, Z.; Xu, K.; Cai, W.; Gao, Y. CXCL3/TGF-β-mediated crosstalk between CAFs and tumor cells augments RCC progression and sunitinib resistance. iScience 2024, 27, 110224. [CrossRef]
- Zheng, C.; Wang, J.; Zhou, Y.; Duan, Y.; Zheng, R.; Xie, Y.; Wei, X.; Wu, J.; Shen, H.; Ye, M.; et al. IFNα-induced BST2(+) tumor-associated macrophages facilitate immunosuppression and tumor growth in pancreatic cancer by ERK-CXCL7 signaling. Cell Rep 2024, 43, 114088. [CrossRef]
- Wang, C.; Lei, Z.; Zhang, C.; Hu, X. CXCL6-CXCR2 axis-mediated PD-L2(+) mast cell accumulation shapes the immunosuppressive microenvironment in osteosarcoma. Heliyon 2024, 10, e34290. [CrossRef]
- Lake, J.A.; Woods, E.; Hoffmeyer, E.; Schaller, K.L.; Cruz-Cruz, J.; Fernandez, J.; Tufa, D.; Kooiman, B.; Hall, S.C.; Jones, D.; et al. Directing B7-H3 chimeric antigen receptor T cell homing through IL-8 induces potent antitumor activity against pediatric sarcoma. J Immunother Cancer 2024, 12. [CrossRef]
- Talbot, L.J.; Chabot, A.; Ross, A.B.; Beckett, A.; Nguyen, P.; Fleming, A.; Chockley, P.J.; Shepphard, H.; Wang, J.; Gottschalk, S.; et al. Redirecting B7-H3.CAR T Cells to Chemokines Expressed in Osteosarcoma Enhances Homing and Antitumor Activity in Preclinical Models. Clin Cancer Res 2024, 30, 4434-4449. [CrossRef]
- Asano, T.; Tanaka, T.; Suzuki, H.; Li, G.; Nanamiya, R.; Tateyama, N.; Isoda, Y.; Okada, Y.; Kobayashi, H.; Yoshikawa, T.; et al. Development of a Novel Anti-Mouse CCR6 Monoclonal Antibody (C(6)Mab-13) by N-Terminal Peptide Immunization. Monoclon Antib Immunodiagn Immunother 2022, 41, 343-349. [CrossRef]
- Kitamura, K.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Cx(6)Mab-1: A Novel Anti-Mouse CXCR6 Monoclonal Antibody Established by N-Terminal Peptide Immunization. Monoclon Antib Immunodiagn Immunother 2022, 41, 133-141. [CrossRef]
- Li, G.; Tanaka, T.; Ouchida, T.; Kaneko, M.K.; Suzuki, H.; Kato, Y. Cx(1)Mab-1: A Novel Anti-mouse CXCR1 Monoclonal Antibody for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2024, 43, 59-66. [CrossRef]
- Ouchida, T.; Isoda, Y.; Nakamura, T.; Yanaka, M.; Tanaka, T.; Handa, S.; Kaneko, M.K.; Suzuki, H.; Kato, Y. Establishment of a Novel Anti-Mouse CCR1 Monoclonal Antibody C(1)Mab-6. Monoclon Antib Immunodiagn Immunother 2024, 43, 67-74. [CrossRef]
- Ouchida, T.; Suzuki, H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Cx(4)Mab-1: A Novel Anti-Mouse CXCR4 Monoclonal Antibody for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2024, 43, 10-16. [CrossRef]
- Tanaka, T.; Li, G.; Asano, T.; Saito, M.; Kaneko, M.K.; Suzuki, H.; Kato, Y. Development of a Novel Anti-Mouse CCR2 Monoclonal Antibody (C(2)Mab-6) by N-Terminal Peptide Immunization. Monoclon Antib Immunodiagn Immunother 2022, 41, 80-86. [CrossRef]
- Tanaka, T.; Nanamiya, R.; Takei, J.; Nakamura, T.; Yanaka, M.; Hosono, H.; Sano, M.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of Anti-Mouse CC Chemokine Receptor 8 Monoclonal Antibodies for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2021, 40, 65-70. [CrossRef]
- Tanaka, T.; Suzuki, H.; Isoda, Y.; Asano, T.; Nakamura, T.; Yanaka, M.; Handa, S.; Takahashi, N.; Okuno, S.; Yoshikawa, T.; et al. Development of a Sensitive Anti-Human CCR9 Monoclonal Antibody (C(9)Mab-11) by N-Terminal Peptide Immunization. Monoclon Antib Immunodiagn Immunother 2022, 41, 303-310. [CrossRef]
- Ikota, H.; Nobusawa, S.; Arai, H.; Kato, Y.; Ishizawa, K.; Hirose, T.; Yokoo, H. Evaluation of IDH1 status in diffusely infiltrating gliomas by immunohistochemistry using anti-mutant and wild type IDH1 antibodies. Brain Tumor Pathol 2015, 32, 237-244. [CrossRef]
- Ishikawa, K.; Suzuki, H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Establishment of a high-affinity anti-mouse CXCR5 monoclonal antibody for flow cytometry. MI 2024, 2. [CrossRef]
- Yamamoto, H.; Suzuki, H.; Tanaka, T.; Satofuka, H.; Kaneko, M.K.; Kato, Y. C7Mab-2: A novel monoclonal antibody against mouse CCR7 established by immunization of the extracellular loop domain. MI 2025. [CrossRef]
- Howard, R.; Kanetsky, P.A.; Egan, K.M. Exploring the prognostic value of the neutrophil-to-lymphocyte ratio in cancer. Sci Rep 2019, 9, 19673. [CrossRef]
- Valero, C.; Lee, M.; Hoen, D.; Weiss, K.; Kelly, D.W.; Adusumilli, P.S.; Paik, P.K.; Plitas, G.; Ladanyi, M.; Postow, M.A.; et al. Pretreatment neutrophil-to-lymphocyte ratio and mutational burden as biomarkers of tumor response to immune checkpoint inhibitors. Nat Commun 2021, 12, 729. [CrossRef]
- Guo, C.; Sharp, A.; Gurel, B.; Crespo, M.; Figueiredo, I.; Jain, S.; Vogl, U.; Rekowski, J.; Rouhifard, M.; Gallagher, L.; et al. Targeting myeloid chemotaxis to reverse prostate cancer therapy resistance. Nature 2023, 623, 1053-1061. [CrossRef]
- Korbecki, J.; Kupnicka, P.; Chlubek, M.; Gorący, J.; Gutowska, I.; Baranowska-Bosiacka, I. CXCR2 Receptor: Regulation of Expression, Signal Transduction, and Involvement in Cancer. Int J Mol Sci 2022, 23. [CrossRef]
- Maas, R.R.; Soukup, K.; Fournier, N.; Massara, M.; Galland, S.; Kornete, M.; Wischnewski, V.; Lourenco, J.; Croci, D.; Álvarez-Prado, Á.F.; et al. The local microenvironment drives activation of neutrophils in human brain tumors. Cell 2023, 186, 4546-4566.e4527. [CrossRef]
- Zhao, J.; Wu, D.; Liu, J.; Zhang, Y.; Li, C.; Zhao, W.; Cao, P.; Wu, S.; Li, M.; Li, W.; et al. Disease-specific suppressive granulocytes participate in glioma progression. Cell Rep 2024, 43, 115014. [CrossRef]
- Alshetaiwi, H.; Pervolarakis, N.; McIntyre, L.L.; Ma, D.; Nguyen, Q.; Rath, J.A.; Nee, K.; Hernandez, G.; Evans, K.; Torosian, L.; et al. Defining the emergence of myeloid-derived suppressor cells in breast cancer using single-cell transcriptomics. Sci Immunol 2020, 5. [CrossRef]
- Highfill, S.L.; Cui, Y.; Giles, A.J.; Smith, J.P.; Zhang, H.; Morse, E.; Kaplan, R.N.; Mackall, C.L. Disruption of CXCR2-mediated MDSC tumor trafficking enhances anti-PD1 efficacy. Sci Transl Med 2014, 6, 237ra267. [CrossRef]
- Nishinakamura, H.; Shinya, S.; Irie, T.; Sakihama, S.; Naito, T.; Watanabe, K.; Sugiyama, D.; Tamiya, M.; Yoshida, T.; Hase, T.; et al. Coactivation of innate immune suppressive cells induces acquired resistance against combined TLR agonism and PD-1 blockade. Sci Transl Med 2025, 17, eadk3160. [CrossRef]
- Banuelos, A.; Zhang, A.; Berouti, H.; Baez, M.; Yılmaz, L.; Georgeos, N.; Marjon, K.D.; Miyanishi, M.; Weissman, I.L. CXCR2 inhibition in G-MDSCs enhances CD47 blockade for melanoma tumor cell clearance. Proc Natl Acad Sci U S A 2024, 121, e2318534121. [CrossRef]
- Bazzichetto, C.; Di Martile, M.; Del Bufalo, D.; Milella, M.; Conciatori, F. Induction of cell death by the CXCR2 antagonist SB225002 in colorectal cancer and stromal cells. Biomed Pharmacother 2025, 188, 118203. [CrossRef]
- Bergeron, P.; Dos Santos, M.; Sitterle, L.; Tarlet, G.; Lavigne, J.; Liu, W.; Gerbé de Thoré, M.; Clémenson, C.; Meziani, L.; Schott, C.; et al. Non-homogenous intratumor ionizing radiation doses synergize with PD1 and CXCR2 blockade. Nat Commun 2024, 15, 8845. [CrossRef]
- Bilusic, M.; Heery, C.R.; Collins, J.M.; Donahue, R.N.; Palena, C.; Madan, R.A.; Karzai, F.; Marté, J.L.; Strauss, J.; Gatti-Mays, M.E.; et al. Phase I trial of HuMax-IL8 (BMS-986253), an anti-IL-8 monoclonal antibody, in patients with metastatic or unresectable solid tumors. J Immunother Cancer 2019, 7, 240. [CrossRef]
- Prado, G.N.; Suetomi, K.; Shumate, D.; Maxwell, C.; Ravindran, A.; Rajarathnam, K.; Navarro, J. Chemokine Signaling Specificity: Essential Role for the N-Terminal Domain of Chemokine Receptors. Biochemistry 2007, 46, 8961-8968. [CrossRef]
- Urvas, L.; Kellenberger, E. Structural Insights into Molecular Recognition and Receptor Activation in Chemokine–Chemokine Receptor Complexes. Journal of Medicinal Chemistry 2023, 66, 7070-7085. [CrossRef]
- Rahimizadeh, P.; Kim, S.; Yoon, B.J.; Jeong, Y.; Lim, S.; Jeon, H.; Lim, H.J.; Park, S.H.; Park, S.I.; Kong, D.H.; et al. Novel CXCR2 antibodies exhibit enhanced anti-tumor activity in pancreatic cancer. Biomed Pharmacother 2025, 185, 117966. [CrossRef]
- Gardner, R.A.; Finney, O.; Annesley, C.; Brakke, H.; Summers, C.; Leger, K.; Bleakley, M.; Brown, C.; Mgebroff, S.; Kelly-Spratt, K.S.; et al. Intent-to-treat leukemia remission by CD19 CAR T cells of defined formulation and dose in children and young adults. Blood 2017, 129, 3322-3331. [CrossRef]
- Park, J.H.; Rivière, I.; Gonen, M.; Wang, X.; Sénéchal, B.; Curran, K.J.; Sauter, C.; Wang, Y.; Santomasso, B.; Mead, E.; et al. Long-Term Follow-up of CD19 CAR Therapy in Acute Lymphoblastic Leukemia. N Engl J Med 2018, 378, 449-459. [CrossRef]
- Ahmed, N.; Brawley, V.S.; Hegde, M.; Robertson, C.; Ghazi, A.; Gerken, C.; Liu, E.; Dakhova, O.; Ashoori, A.; Corder, A.; et al. Human Epidermal Growth Factor Receptor 2 (HER2) -Specific Chimeric Antigen Receptor-Modified T Cells for the Immunotherapy of HER2-Positive Sarcoma. J Clin Oncol 2015, 33, 1688-1696. [CrossRef]
- Hou, B.; Tang, Y.; Li, W.; Zeng, Q.; Chang, D. Efficiency of CAR-T Therapy for Treatment of Solid Tumor in Clinical Trials: A Meta-Analysis. Dis Markers 2019, 2019, 3425291. [CrossRef]
- Giudice, A.M.; Roth, S.L.; Matlaga, S.; Cresswell-Clay, E.; Mishra, P.; Schürch, P.M.; Boateng-Antwi, K.A.M.; Samanta, M.; Pascual-Pasto, G.; Zecchino, V.; et al. Reprogramming the neuroblastoma tumor immune microenvironment to enhance GPC2 CAR T cells. Mol Ther 2025, 33, 4552-4569. [CrossRef]
- Dai, Z.; Lin, X.; Wang, X.; Zou, X.; Yan, Y.; Wang, R.; Chen, Y.; Tasiheng, Y.; Ma, M.; Wang, X.; et al. Ectopic CXCR2 expression cells improve the anti-tumor efficiency of CAR-T cells and remodel the immune microenvironment of pancreatic ductal adenocarcinoma. Cancer Immunol Immunother 2024, 73, 61. [CrossRef]
- Hosking, M.P.; Shirinbak, S.; Omilusik, K.; Chandra, S.; Kaneko, M.K.; Gentile, A.; Yamamoto, S.; Shrestha, B.; Grant, J.; Boyett, M.; et al. Preferential tumor targeting of HER2 by iPSC-derived CAR T cells engineered to overcome multiple barriers to solid tumor efficacy. Cell Stem Cell 2025, 32, 1087-1101.e1084. [CrossRef]
- Lin, X.; Liu, Z.; Dong, X.; Wang, K.; Sun, Y.; Zhang, H.; Wang, F.; Chen, Y.; Ling, J.; Guo, Y.; et al. Radiotherapy enhances the anti-tumor effect of CAR-NK cells for hepatocellular carcinoma. J Transl Med 2024, 22, 929. [CrossRef]
Figure 1.
A schematic illustration of the production of anti-mCXCR2 mAbs. The rat was intraperitoneally immunized with KLH-conjugated N-terminus mCXCR2 peptide (five times in total). Hybridoma screening was then conducted by ELISA, using naked-mCXCR2 peptide, followed by the flow cytometric analysis using mCXCR2-overexpressed CHO-K1 cells and parental cells. To obtain monoclonal clone, single cell cloning was performed by limitting dilution.
Figure 1.
A schematic illustration of the production of anti-mCXCR2 mAbs. The rat was intraperitoneally immunized with KLH-conjugated N-terminus mCXCR2 peptide (five times in total). Hybridoma screening was then conducted by ELISA, using naked-mCXCR2 peptide, followed by the flow cytometric analysis using mCXCR2-overexpressed CHO-K1 cells and parental cells. To obtain monoclonal clone, single cell cloning was performed by limitting dilution.
Figure 2.
Flow cytometric analysis using anti-mCXCR2 mAbs. CHO/mCXCR2 cells (A), CHO-K1 cells (B), and WEHI-3B cells (C) were treated with 0.01–10 µg/mL of Cx2Mab-5 and SA045E1, followed by treatment with Alexa Fluor 488-conjugated anti-rat IgG. Black line represents the negative control (no primary mAb treatment).
Figure 2.
Flow cytometric analysis using anti-mCXCR2 mAbs. CHO/mCXCR2 cells (A), CHO-K1 cells (B), and WEHI-3B cells (C) were treated with 0.01–10 µg/mL of Cx2Mab-5 and SA045E1, followed by treatment with Alexa Fluor 488-conjugated anti-rat IgG. Black line represents the negative control (no primary mAb treatment).
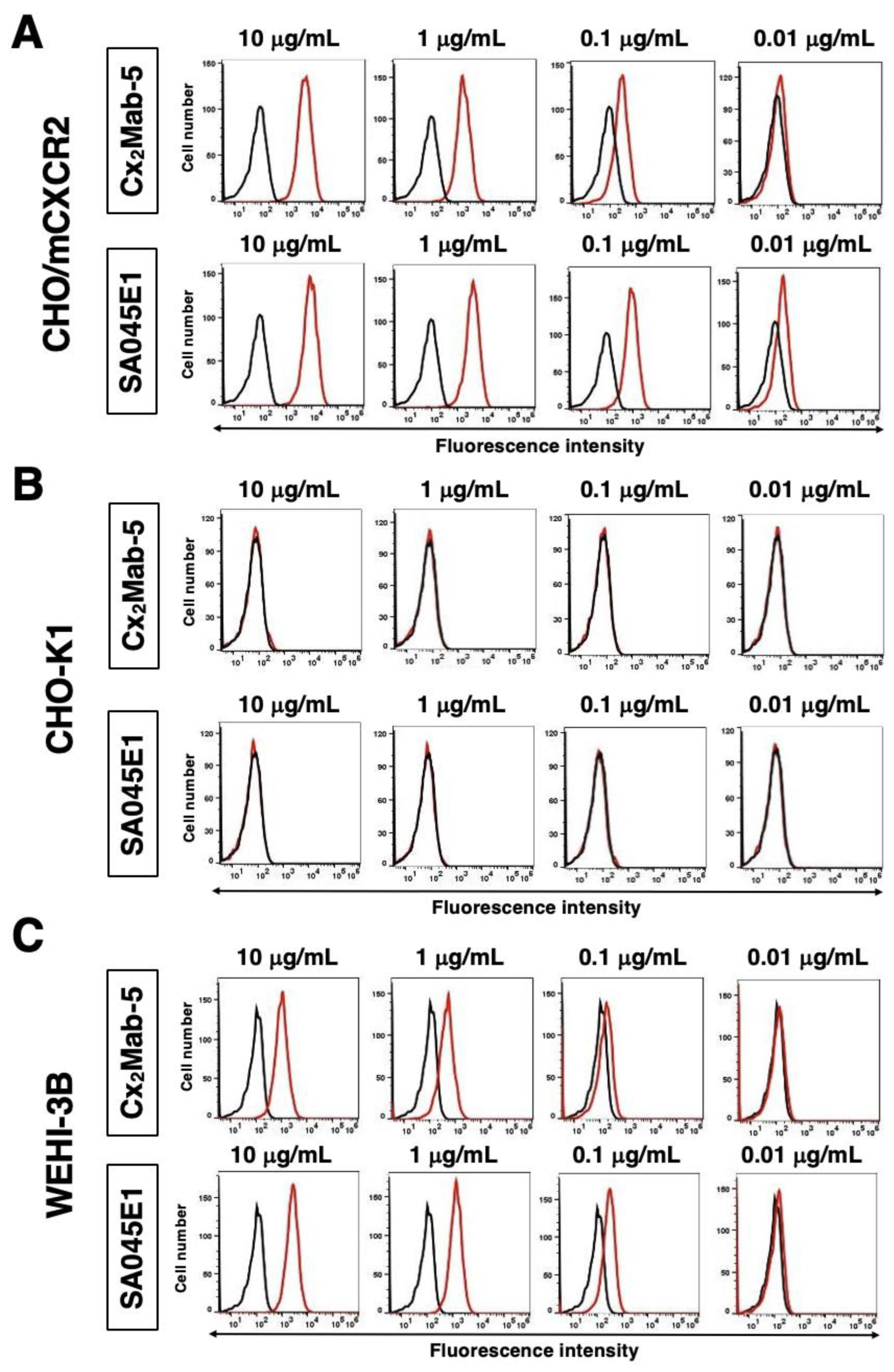
Figure 3.
Cross-reactivity of Cx2Mab-5 in in mouse CC, CXC, CX3C, and XC chemokine receptor-expressed CHO-K1 cells. (A) The mouse CC, CXC, CX3C, and XC chemokine receptor-expressed CHO-K1 cells were treated with 1 μg/mL of Cx2Mab-5 (red line) or control blocking buffer (black line), followed by the treatment with an Alexa Fluor 488-conjugated anti-rat IgG. (B) The cells were treated with 1 μg/mL of corresponding mAbs, including anti-mCCR1 mAb (clone C1Mab-6), anti-mCCR2 mAb (clone C2Mab-6), anti-mCCR3 mAb (clone C3Mab-4), anti-mCCR5 mAb (clone C5Mab-2), anti-mCCR7 mAb (clone 4B12, BioLegend,), anti-mCCR8 mAb (clone C8Mab-2), anti-mCCR9 mAb (clone C9Mab-24), anti-mCXCR1 mAb (clone Cx1Mab-1), anti-mCXCR2 mAb (clone SA045E1), anti-mCXCR3 mAb (clone Cx3Mab-4), anti-mCXCR4 mAb (clone Cx4Mab-1), anti-mCXCR5 mAb (clone Cx5Mab-3), anti-mCXCR6 mAb (clone Cx6Mab-1), anti-mCX3CR1 mAb (clone SA011F11, BioLegend,), anti-mXCR1 mAb (clone ZET, BioLegend,), and anti-PA tag mAb (NZ-1), followed by the treatment with corresponding secondary antibodies conjugated with Alexa Fluor 488.
Figure 3.
Cross-reactivity of Cx2Mab-5 in in mouse CC, CXC, CX3C, and XC chemokine receptor-expressed CHO-K1 cells. (A) The mouse CC, CXC, CX3C, and XC chemokine receptor-expressed CHO-K1 cells were treated with 1 μg/mL of Cx2Mab-5 (red line) or control blocking buffer (black line), followed by the treatment with an Alexa Fluor 488-conjugated anti-rat IgG. (B) The cells were treated with 1 μg/mL of corresponding mAbs, including anti-mCCR1 mAb (clone C1Mab-6), anti-mCCR2 mAb (clone C2Mab-6), anti-mCCR3 mAb (clone C3Mab-4), anti-mCCR5 mAb (clone C5Mab-2), anti-mCCR7 mAb (clone 4B12, BioLegend,), anti-mCCR8 mAb (clone C8Mab-2), anti-mCCR9 mAb (clone C9Mab-24), anti-mCXCR1 mAb (clone Cx1Mab-1), anti-mCXCR2 mAb (clone SA045E1), anti-mCXCR3 mAb (clone Cx3Mab-4), anti-mCXCR4 mAb (clone Cx4Mab-1), anti-mCXCR5 mAb (clone Cx5Mab-3), anti-mCXCR6 mAb (clone Cx6Mab-1), anti-mCX3CR1 mAb (clone SA011F11, BioLegend,), anti-mXCR1 mAb (clone ZET, BioLegend,), and anti-PA tag mAb (NZ-1), followed by the treatment with corresponding secondary antibodies conjugated with Alexa Fluor 488.
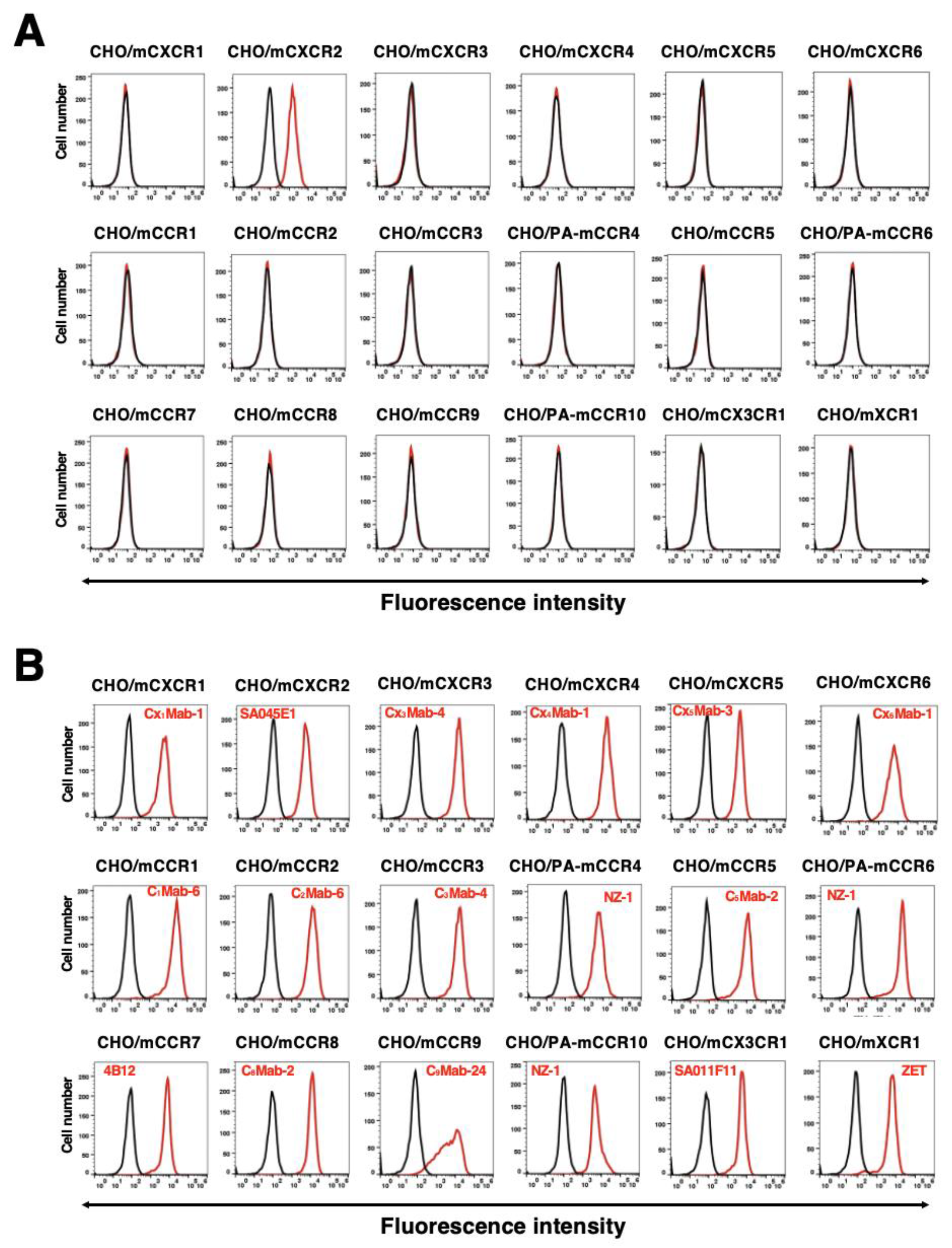
Figure 4.
Peptide-blocking analysis using anti-mCXCR2 mAbs with mCXCR2 peptide. CHO/mCXCR2 and WEHI-3B cells were incubated with Cx2Mab-5 (1 μg/mL) plus control (0.1% DMSO in blocking buffer), Cx2Mab-5 plus mCXCR2 peptide (10 μg/mL), SA045E1 (1 μg/mL) plus control (0.1% DMSO in blocking buffer), or SA045E1 plus mCXCR2 peptide (10 μg/mL) for 30 min at 4 °C. Cells were then treated with Alexa Fluor 488-conjugated anti-rat IgG. Fluorescence data were collected using the SA3800 Cell Analyzer. The black line represents the negative control (blocking buffer).
Figure 4.
Peptide-blocking analysis using anti-mCXCR2 mAbs with mCXCR2 peptide. CHO/mCXCR2 and WEHI-3B cells were incubated with Cx2Mab-5 (1 μg/mL) plus control (0.1% DMSO in blocking buffer), Cx2Mab-5 plus mCXCR2 peptide (10 μg/mL), SA045E1 (1 μg/mL) plus control (0.1% DMSO in blocking buffer), or SA045E1 plus mCXCR2 peptide (10 μg/mL) for 30 min at 4 °C. Cells were then treated with Alexa Fluor 488-conjugated anti-rat IgG. Fluorescence data were collected using the SA3800 Cell Analyzer. The black line represents the negative control (blocking buffer).
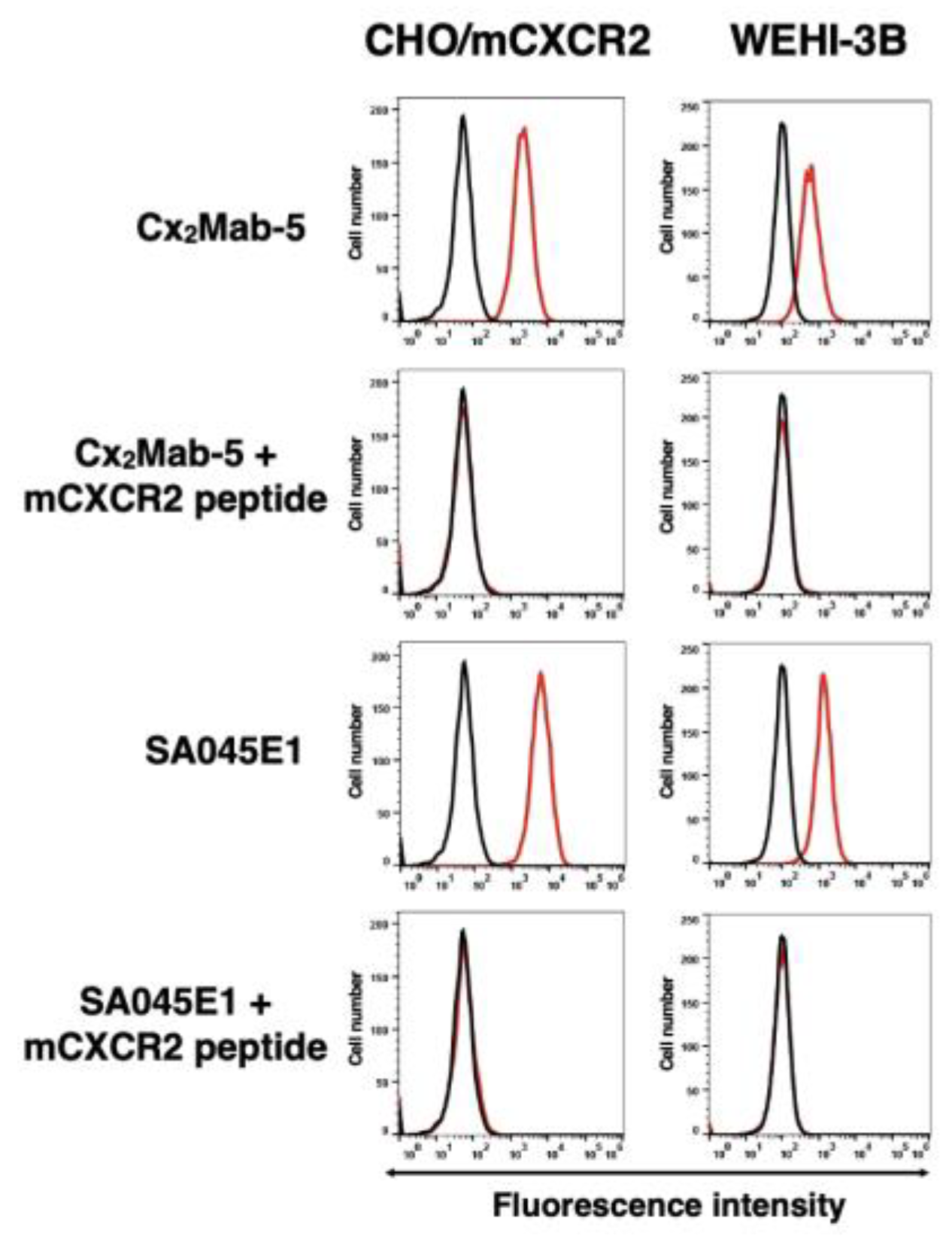
Figure 5.
The determination of the dissociation constant of Cx2Mab-5. CHO/mCXCR2 (A) or WEHI-3B (B) cells were suspended in 100 µL serially diluted Cx2Mab-5 (100 to 0.006 µg/mL) and SA045E1 (10 to 0.0006 µg/mL for CHO/mCXCR2, 50 to 0.003 µg/mL for WEHI-3B). Then, cells were treated with Alexa Fluor 488-conjugated anti-rat IgG. Fluorescence data were subsequently collected using a SA3800 Cell Analyzer, following the calculation of the dissociation constant (KD) by GraphPad PRISM 6.
Figure 5.
The determination of the dissociation constant of Cx2Mab-5. CHO/mCXCR2 (A) or WEHI-3B (B) cells were suspended in 100 µL serially diluted Cx2Mab-5 (100 to 0.006 µg/mL) and SA045E1 (10 to 0.0006 µg/mL for CHO/mCXCR2, 50 to 0.003 µg/mL for WEHI-3B). Then, cells were treated with Alexa Fluor 488-conjugated anti-rat IgG. Fluorescence data were subsequently collected using a SA3800 Cell Analyzer, following the calculation of the dissociation constant (KD) by GraphPad PRISM 6.
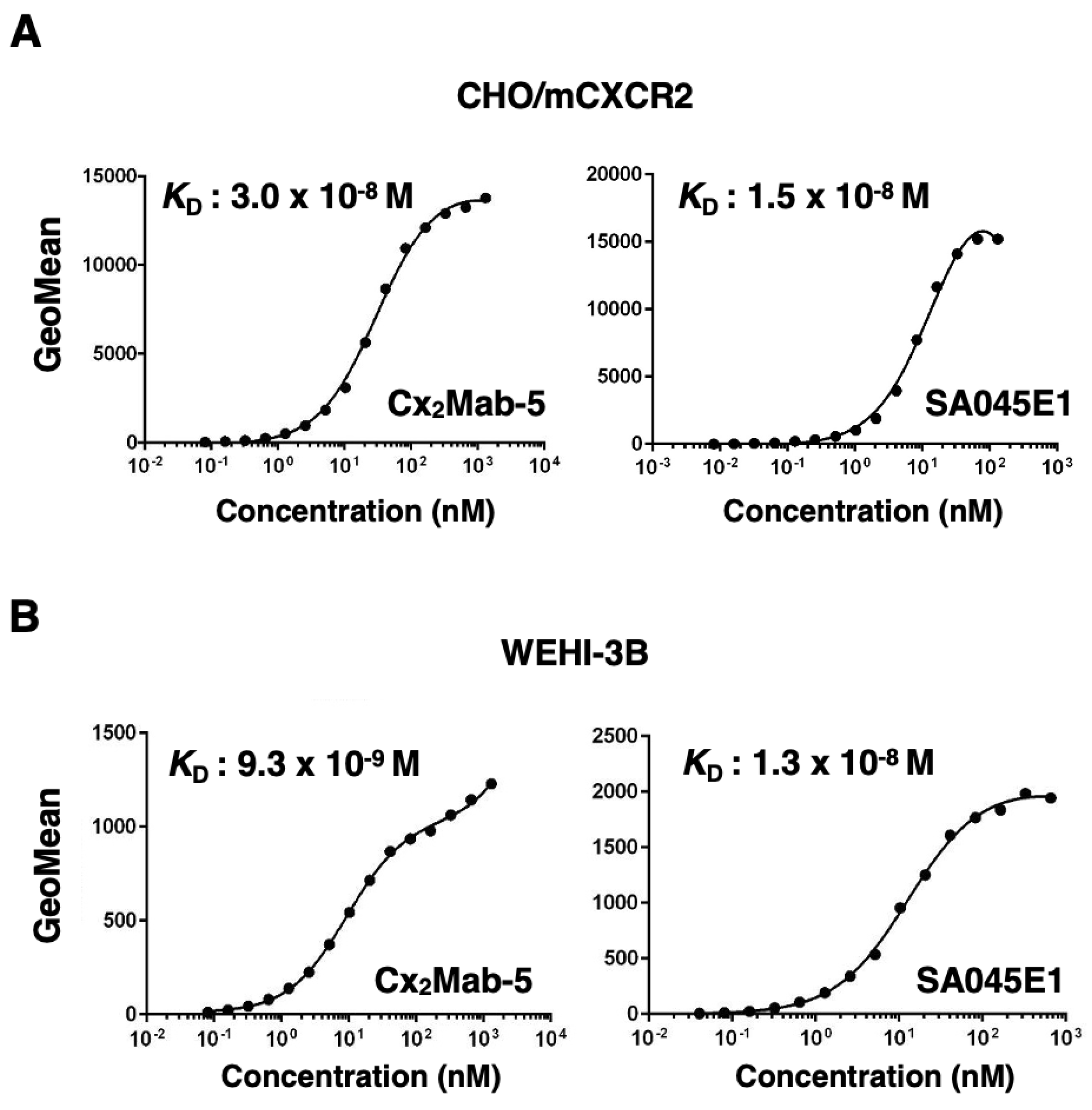
Figure 6.
The detection of mCXCR2 by western blot analysis. Cell lysates of CHO-K1 and CHO/mCXCR2 were electrophoresed and transferred onto polyvinylidene difluoride membranes. The membranes were incubated with 5 μg/mL of Cx2Mab-5, 5 μg/mL of SA045E1, 0.5 μg/mL of 1E6 (an anti-DYKDDDDK mAb), and 1 μg/mL of RcMab-1 and subsequently with horseradish peroxidase-conjugated anti-mouse or anti-rat immunoglobulins. Blots were developed using PierceTM ECL Plus Western Blotting Substrate or ImmunoStar LD and imaged with a Sayaca-Imager.
Figure 6.
The detection of mCXCR2 by western blot analysis. Cell lysates of CHO-K1 and CHO/mCXCR2 were electrophoresed and transferred onto polyvinylidene difluoride membranes. The membranes were incubated with 5 μg/mL of Cx2Mab-5, 5 μg/mL of SA045E1, 0.5 μg/mL of 1E6 (an anti-DYKDDDDK mAb), and 1 μg/mL of RcMab-1 and subsequently with horseradish peroxidase-conjugated anti-mouse or anti-rat immunoglobulins. Blots were developed using PierceTM ECL Plus Western Blotting Substrate or ImmunoStar LD and imaged with a Sayaca-Imager.
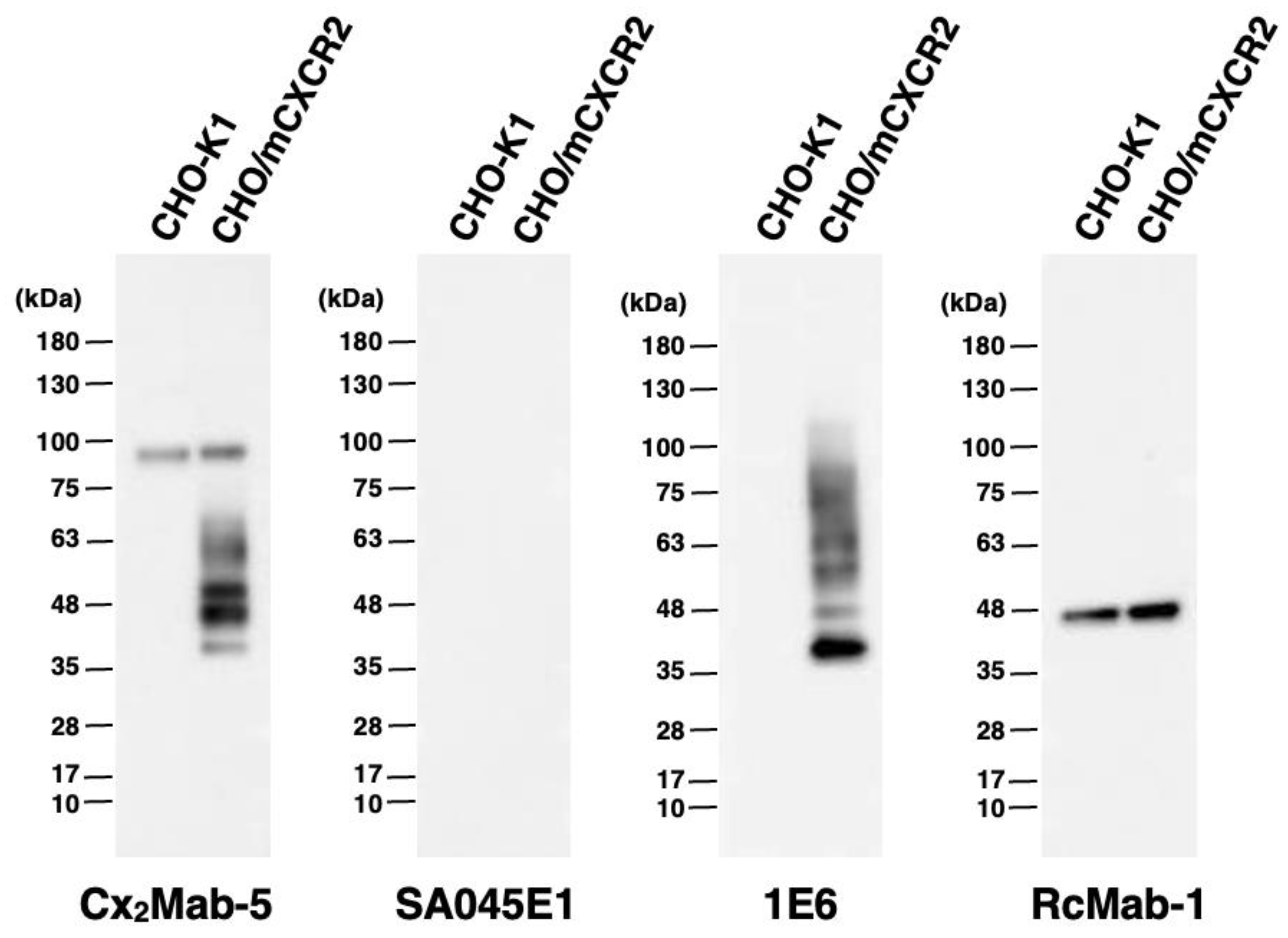
Figure 7.
Immunohistochemical staining of paraffin-embedded section of CHO/mCXCR2 and CHO-K1. The sections of CHO/mCXCR2 and CHO-K1 cells were treated with 5 μg/mL of Cx2Mab-5 or 5 μg/mL of SA045E1, followed by that with the Histofine Simple Stain Mouse MAX PO (Rat). Color was developed using DAB, and counterstaining was performed using hematoxylin (Merck KGaA). Scale bar = 100 μm.
Figure 7.
Immunohistochemical staining of paraffin-embedded section of CHO/mCXCR2 and CHO-K1. The sections of CHO/mCXCR2 and CHO-K1 cells were treated with 5 μg/mL of Cx2Mab-5 or 5 μg/mL of SA045E1, followed by that with the Histofine Simple Stain Mouse MAX PO (Rat). Color was developed using DAB, and counterstaining was performed using hematoxylin (Merck KGaA). Scale bar = 100 μm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



