
Submitted:
06 November 2025
Posted:
07 November 2025
You are already at the latest version
Abstract
Beans are widely consumed worldwide and are a good source of amino acids and micronutrients; however, they contain anti-nutrients, such as lectins, tannins, protein inhibitors, saponins, and phytic acid, among others, which can reduce the food’s quality and cause adverse health effects. In this study, we analyzed the genotoxic and cytotoxic effects of a protein extract from Phaseolus acutifolius (TBE) seeds. The extract contained some antinutritional compounds, with a higher lectin content, with an activity of 2701.85 HU. The acute toxicity test in mice showed that the extract was non-toxic at the concentrations tested, as it did not cause any mortality. The in vitro cytotoxicity study on small intestinal epithelial cells indicated that the lectin-rich extract was cytotoxic in both assays, with IC50 values of 10.08 µg/mL and 108.93 µg/mL for the free cell and intestinal fragment assays, respectively. In the in vivo study, an erythropoiesis-stimulatory effect was observed, with significant genotoxic damage noted at 48 h, evidenced by 11 micronucleated erythrocytes at 1000 mg/kg TBE. However, no genotoxicity was detected with prolonged treatment times. These results indicate that TBE is cytotoxic within the tested concentration range, and genotoxic damage is influenced by both concentration and exposure time.
Keywords:
proteins
; toxicity
; cytotoxicity
; genotoxic
; Phaseolus acutifolius
1. Introduction
Legumes play a significant role in human diets, particularly in developing countries [1]. Preferences for bean consumption vary by region within each country. In Mexico, various domesticated species are consumed depending on the region, including Phaseolus coccineus L., Phaseolus acutifolius A. Gray, Phaseolus lunatus L., and Phaseolus vulgaris, with colors such as black, pink, sulfur, pinto, or white. The per capita consumption of beans in México is approximately nine kilograms per year [2].
Beans provide essential nutrients, including protein, vitamin B, minerals, and fiber; they are gluten-free, have a low glycemic index, and exhibit significant antioxidant capacity. Beans are particularly important for populations with limited access to animal protein due to high costs or for those with health conditions that restrict their consumption of animal products, contributing approximately 11% of daily protein intake and 6% of daily energy in the human diet. The bean Phaseolus acutifolius, commonly known in northern México as the Tepary bean, is a species well-adapted to arid and semi-arid growing conditions. It is a valuable source of nutrients, including protein, carbohydrates, lipids, amino acids, minerals, phenols, and antioxidants. The nutritional significance of the tepary bean lies in its protein content, which varies by variety from 13% to 32%.[3–6]
Consuming this type of legume provides not only essential nutrients for a balanced diet but also confers physiological benefits that support health. Studies suggest that it may help prevent metabolic diseases such as diabetes, heart disease, and certain types of cancer [7].
Like other legumes, beans contain compounds with anti-nutritional properties that can affect quality by interfering with food digestibility and metabolism, and potentially causing physiological harm to consumers. These compounds include lectins, protease inhibitors, tannins, saponins, and phytates [7–9].
The quality of a food’s protein is primarily determined by its digestibility, the essential amino acids it provides, and the availability of these amino acids [8]. Despite their excellent nutritional profile, beans have some disadvantages due to anti-nutritional compounds, which significantly reduce protein and amino acid digestibility, thereby affecting nutrient bioavailability [8,10,11].
To minimize potential adverse effects, it is essential to reduce anti-nutritional compounds through processing methods such as thermal extrusion, boiling, pH adjustments, or ultra-high temperature processing [11–16].
Given the potentially toxic effects of certain beans, it is essential to carried out toxicological tests, such as cytotoxicity and genotoxicity assays, be conducted to assess the potential harm of the compounds or substances present in this food.
One method for assessing potential genotoxic effects is the micronucleus (MN) assay, which detects fragments of chromosomes that are excluded from the nucleus during mitosis [17]. Additionally, performing the MN test on peripheral blood from mice provides a straightforward, rapid, and reliable method to evaluate the cytotoxic effects of the tested sample [18–23] .
The genus Phaseolus is a promising source of proteins and peptides with therapeutic potential [24]. Protein isolates derived from beans have been increasingly produced as they offer functional ingredients that enhance food quality. Thus, beans can be utilized to prepare protein isolates rich in essential amino acids and functional compounds, contributing to the nutritional enrichment of products [25–30].
Given the growing interest in the use of protein isolates and the possible presence of compounds that may pose health risks, it is crucial to evaluate their cytotoxic and genotoxic effects to mitigate any associated hazards. Accordingly, this study aimed to evaluate the cytotoxic and genotoxic effects of a protein extract derived from tepary bean.
2. Materials and Methods
4.1. Animals
Fifty-nine male CD-1 mice, with a mean weight of 13 g, were used in the micronucleus assay. They were obtained from the Institute of Health Sciences-UAEH (Pachuca, Hidalgo, México) and maintained under standard conditions in metallic cages at 23 ± 2°C and 50 ± 10% humidity, with food (TEKLAD Global 2018S, Harlan, México City) and water provided ad libitum on a 12-hour light–dark cycle. The animals underwent a one-week acclimatization period before treatments.
Nine male Wistar rats, weighing 200–250 g, were used to obtain intestinal tissue. These animals were also obtained from the Institute of Health Sciences-UAEH and maintained under the same conditions as the mice. Prior to the assays, the rats were fasted for 12 h with water provided ad libitum.
4.2. Human Erythrocytes
Peripheral blood erythrocytes (Type A) were obtained from a male volunteer donor in accordance with Official Mexican Standards NOM-007-SSA3-2011 and NOM-253-SSA1-2012. The donor provided signed informed consent, and the study was approved by the Ethics and Research Committee of the Autonomous University of the State of Hidalgo.
4.3. Plant Seeds
White seeds of Phaseolus acutifolius (tepary bean seeds, TB) used in the experiment were sourced from a local market in Hermosillo, Sonora, México. The seeds were cleaned and stored at 4°C until use.
4.4. Protein Extraction of Phaseolus acutifolius Seeds (TBE)
Proteins were extracted from unprocessed white tepary beans following the methodology proposed by [30]. Briefly, a protein solution was prepared by shaking seed powder for 16 h at 4°C in 10 mM phosphate-buffered saline (PBS), pH 7.4, at a ratio of 1:10 (w/v). The mixture was then centrifuged at 15,000 × g for 60 minutes to obtain the protein extract (TBE). The extract was dialyzed overnight against deionized water at 4°C, lyophilized, and stored at −20°C.
The protein concentration of TBE was determined using the Bradford method, with bovine serum albumin (Sigma Chemical Co., Burlington,MA, USA) as the standard (98) .
4.5. Antinutritional Compounds
4.5.1. Lectins Assay
The serial dilution method was employed [102]. Ninety-six-well microplates were used, with 50 µL of phosphate-buffered saline (PBS) added to each well. A 50 µL sample of the protein solution was serially diluted, and then 50 µL of type A erythrocyte suspension was introduced. The mixture was incubated at room temperature for 1 h, after which the highest dilution that showed hemagglutination was recorded. The results were expressed as hemagglutination units.
4.5.2. Trypsin Inhibitors Determination
Trypsin inhibitory activity was measured using the synthetic substrate BAPNA (N-benzoyl-DL-arginine p-nitroanilide). The trypsin inhibitor was extracted with NaOH (0.01 N) at pH 9.6 for 2.5 h under stirring, and the supernatant containing the trypsin inhibitors was separated by centrifugation for 30 min at 5000 rpm. A trypsin solution (20 µg/mL in 0.001 N HCl) was added to the supernatant containing trypsin inhibitors and incubated for 10 min at 37 °C. Subsequently, BAPNA (0.4 mg/mL, pH 8.2) (Sigma Chemical Co., St. Louis, MO, USA) was added and allowed to react for 10 min at 37 °C. The reaction was stopped by adding 30% acetic acid, and the absorbance at 410 nm was measured (Perkin Elmer Lambda 40). Total trypsin inhibitory activity was expressed as trypsin inhibitor units (TIU) per milligram of sample (TIU/mg) [31].
4.5.3. Tannins Determination
Tannins were quantified in the protein extract using the modified vanillin–HCl method [32], with (+)-catechin (Sigma Chemical Co., St. Louis, MO, USA) as the reference standard. Briefly, tannins were extracted in methanol for 30 min at room temperature, followed by centrifugation at 3500 rpm. The supernatant was then reacted with vanillin in acidified methanol (0.5:8%), and absorbance was measured at 500 nm using a Perkin Elmer Lambda 40 UV/Vis spectrophotometer. Tannin content was expressed as milligrams of catechin equivalents per gram of sample (mg CE/g).
4.5.4. Saponins Determination
Saponin content was determined following the methodology described by Valadez (2022) [33], with some modifications. Briefly, saponins were extracted from the protein extract using a methanol: water solution (85:15%) for 1 h. The solvents were removed by evaporation, and the saponins were diluted in 0.9% NaCl. For saponins identification, the serial dilution method with type O human erythrocytes was employed. Results were expressed as hemolytic units per milligram of sample and represent the mean of three independent measurements.
4.6. Lethal Median Dose (LD50)
The experiment was carried out in two phases. In the first phase, 12 male CD-1 mice were divided into four groups and administered TBE intraperitoneally at doses of 0, 10, 100, and 1000 mg/kg. Since no mortality was observed, in the second phase, four groups of three mice were given TBE at doses of 0, 1600, 2900, and 5000 mg/kg to assess animal mortality. The average lethal dose (LD50) was then calculated [30,99].
4.7. Evaluation of the Genotoxic Capacity of TBE by the Micronucleus Assay and Cytotoxicity to Bone Marrow
To assess the genotoxic potential of TBE, 35 male CD-1 mice were divided into five groups and administered single intraperitoneal doses of TBE at 250, 500, and 1000 mg/kg. A negative control group received NaCl solution (0.9%), and a positive control group was treated with daunorubicin (DAU) (Sigma Chemical Co., Burlington, MA, USA) at 3 mg/kg. Peripheral blood samples were collected from the tail of each animal at 0, 24, 48, 72, and 94 h post-treatment. Smears were prepared, fixed with methanol for 5 min, and stained with 4% Giemsa for 20 minutes [30,100]. To determine the frequency of micronucleated polychromatic erythrocytes (MNCE), 1000 erythrocytes were counted from each sample. Bone marrow cytotoxicity was assessed by scoring 1000 erythrocytes per animal and calculating the ratio of polychromatic erythrocytes (PE) to normochromatic erythrocytes (PE/NE ratio).
The Ethics and Biosecurity Committee of the Institute of Health Sciences approved the protocol for the micronucleus assay.
4.8. In Vitro Cytotoxicity Assay in Rat Intestinal Epithelial Cells
Intestinal Smear Assay
Four male Wistar rats (200–250 g) were fasted for 12 h prior to the experiment. The small intestine was dissected, and 3 cm segments were collected, washed with NaCl (0.9%) containing streptomycin (1.5%), and inverted on sterile glass rods. The intestinal fragments were then maintained in modified McCoy 5A medium (Gibco, Grand Island, NY, USA), supplemented with 7% fetal bovine serum (Gibco, Grand Island, NY, USA) and 1.5% streptomycin (Gibco, Grand Island, NY, USA). The tissue fragments were incubated for 15 min at 37°C with the extracts at concentrations of 0, 5, 10, 50, 100, 200, 500, and 1000 µg/mL, with gentle agitation. Epithelial cell smears were prepared by lightly pressing the intestinal mucosa onto a slide, which was then slid to release the cells. The cells were stained with 0.05% trypan blue (Sigma Chemical Co., Burlington, MA, USA) to assess cell viability using the exclusion technique, where unstained cells were considered viable [58,62].
4.9. Epithelial Cell Assay
Five male Wistar rats (200–250 g) were fasted for 12 h before use. The small intestine was removed, inverted on a glass rod, and washed as described previously. To isolate the epithelial cells, the intestinal fragments were incubated in a 0.25% bovine pancreatic trypsin solution (Fluka Biochemika) with constant agitation at 37°C for 25 min. The cells were then recovered by centrifugation at 2500 rpm, washed three times with modified McCoy 5A medium (Gibco, Grand Island, NY, USA), and resuspended in the same medium supplemented with 7% fetal bovine serum (Gibco, Grand Island, NY, USA) and 1.5% streptomycin (Gibco, Grand Island, NY, USA) [4,58].
An aliquot of 600 µL of the cell suspension, containing 8.5 × 10⁷ cells, was transferred to tubes containing 900 µL of the protein extract at concentrations of 0, 1, 2, 5, 10, 50, 100, 200, 500, and 1000 µg/mL, and incubated for 15 min at 37°C. The cells were then recovered by centrifugation at 2500 rpm and resuspended in modified McCoy 5A medium (Gibco, Grand Island, NY, USA) supplemented with 7% fetal bovine serum (Gibco, Grand Island, NY, USA) and 1.5% streptomycin (Gibco, Grand Island, NY, USA). An aliquot of cells was taken and stained with trypan blue (Sigma Chemical Co., Burlington, MA, USA) to assess cell viability using the exclusion method [62].
3. Results
2.1. Protein Extract and Antinutritional Compounds
The protein extract from powder tepary bean, unprocessed, had a concentration of 3.97 mg/mL. Regarding antinutritional compounds, the protein extract exhibited the presence of lectins, trypsin inhibitors, tannins, and saponins, as shown in Table 1.
Antinutrient compounds were quantified in the protein extract of tepary bean.. HAU: hemagglutinin activity units/mg protein, TIU: Trypsin inhibitory units/ mg, mg CE /g: mg catechin equivalents/mg sample, HU/mg: hemolysis units/mg. Results represent the mean ± standard deviation values of triplicate experiments.
2.2. Epithelial Cell Cytotoxicity
2.2.1. Smear Assay
Figure 1 shows the cytotoxic effect observed in the epithelial cells from smears prepared from intestinal fragments exposed to the tepary bean protein extract, revealing a dose-dependent response. Starting from the lowest dose, a decrease in cell viability was observed, with a continuous reduction noted across all tested doses. The mean inhibitory concentration (IC₅₀) was reached at 108.97 µg/mL, while at the highest dose tested, cell viability was inhibited by 85.3%.
2.2.2. Free Epithelial Cells
The cytotoxic effects of the tepary bean protein extract on epithelial cells, isolated from the small intestine, are shown in Figure 1. The results indicate that the extract was toxic to the cells, significantly reducing viability at low concentrations. At 10 µg/mL, the extract inhibited cell viability by 50.2%, with an IC₅₀ of 10.08 µg/mL. A consistent decrease in viability of approximately 10% was observed across the tested concentrations, reaching 83.6% at a concentration of 200 µg/mL. This reduction in viability continued at higher concentrations, with a maximum inhibition of 92.3% at the highest concentration tested.
2.4. Evaluation of the Genotoxic Capacity of TBE by the Micronucleus Assay and Cytotoxicity to Bone Marrow
The acute toxicity study of the tepary bean protein extract (TBE) in CD-1 mice demonstrated that the extract was not toxic at doses up to 5000 mg/kg, as no mortality was observed at any of the administered concentrations.
Figure 2 shows the evaluation of the genotoxic effects of the tepary bean extract, daunorubicin (DAU),used as a positive control, and physiological saline solution (0.9% NaCl), used as a negative control.
As expected, the mice treated with the negative control (NaCl) showed no genotoxicity, with mean values of 1.6, 1.8, 2.4, and 2.0 for the number of EPCMN at 24, 48, 72, and 96 h, respectively. In contrast, DAU treatment resulted in a significant increase in EPCMN after 24 h, with a mean of 28.6. The most strongest genotoxic effect was observed at 48 h, where EPCMN levels increased by 33.5-fold compared to the negative control. Genotoxicity persisted up to 96 h, but with a considerable decline in EPCMN levels, averaging 14.2, which was 4.7 times lower than the peak observed at 48 h.
Regarding the number of EPCMN induced by the white tepary bean protein extract, genotoxic effects were observed at doses of 250 mg/kg (5.75 EPCMN), 500 mg/kg (6.25 EPCMN), and 1000 mg/kg (11 EPCMN) at 48 h post-administration. At later time points, EPCMN counts decreased to the point where no genotoxic damage was detectable.
Figure 3 shows the results of the cytotoxicity evaluation. DAU treatment led to an increase in the number of polychromatic erythrocytes (EPC) over time, reaching an average of 34.8 at 96 h. The saline solution had no effect, maintaining an average of 14 EPC throughout the study. In contrast, TBE exhibited a stimulatory effect on erythropoiesis beginning at 24 h, with this effect persisting throughout the study. The most significant effect was observed at 48 h with a dose of 250 mg/kg, which was four times greater than the negative control. At 72 h, a dose of 500 mg/kg produced a similar response, 4.2 times greater than that of NaCl.
4. Discussion
Beans are a widely consumed legume globally, and in México, they are a staple in the diets of the population [31]. México produces a wide variety of beans, with preferences varying by region. The most commonly cultivated varieties are sulfur, pinto, and black beans, with the latter being predominantly consumed in the central and southern parts of the country [2,31]. In contrast, lighter-colored varieties, such as the tepary bean (Phaseolus acutifolius), are preferred in northern México.
The protein content in various tepary bean varieties has been reported to range from 18.9% to 21.6% [3]; however, when proteins are extracted using saline solutions, concentrations have been reported between 7.9 and 30.40 mg/mL, with values higher than those observed in this study (3.97 mg/mL).
In comparison to other bean species, the protein concentration in the white tepary bean extract was lower (77.4–85.7%) than that of some other species [34,35], but 74.6% higher than that of mung bean extracts [34]. These differences may be attributed to variations in species, bean variety, and the methods used for protein extraction [4,37,38].
The presence of antinutritional compounds in grain legumes, such as beans, has been widely reported. Accordingly, their occurrence in protein extracts is expected and is influenced by the extraction method employed. The biological effects of these compounds depend on multiple factors, including their concentration, chemical nature, the species examined, and the physiological status of the animal [39];.
Although white tepary bean seeds were used in this study, tannins were detected in the protein extract; however, their concentration was considerably lower than that previously reported for both white and colored seeds of the same species [4];. It is well established that the content of these polyphenolic compounds is associated with the color of the seed coat, such that more intensely pigmented or darker coats exhibit higher concentration [40]; As the analysis in this study was conducted on a protein extract derived from white seed flour, a low concentration of these compounds was expected.
Saponins were analyzed using the hemolysis method, and their concentration in the protein extract was found to be higher than that previously reported for protein extracts (0.0161 HU/mg) and flours (0.0078–0.0126 HU/mg) of certain P. vulgaris varieties [41];. The differences observed can be attributed to both species-specific variation and the methods employed for protein extraction and saponin determination.
Trypsin inhibitors were detected in the protein extract; although their concentration was low, these antinutritional compounds are known to impair protein digestion. Consequently, their inactivation is essential to prevent adverse effects. It can be achieved through treatments such as cooking, roasting, extrusion, ultrasound, ultrafiltration, reducing agents, acids, bases, or biological processes, including germination and fermentation [39,42,43];. In protein extracts from legumes such as beans, lentils, and soybeans, concentrations of these compounds have been reported to range from 11.69 to 89.78 HU/mg, which are substantially higher than those observed in tepary bean protein extracts [41]. Moreover, it has been reported that trypsin inhibitors are preferentially extracted under alkaline conditions with NaOH at pH 8.4–10 [39]. Thus, the lower levels observed in TBE compared with other samples can be attributed to the extraction method, since in the present study the protein extract was obtained using a phosphate buffer at pH 7.4, which likely resulted in the extraction of fewer inhibitors.
Among the proteins found in beans are lectins, which have a globular structure and can be easily extracted with saline solutions [30,44,45]. Lectins are known for their ability to agglutinate erythrocytes, exhibit cytotoxic effects on various cell types (39–44), and cause adverse effects in laboratory animals [52-53].
For lectin extraction, a buffered saline solution was used in this study, which proved to be effective for isolating these proteins. A significant concentration of lectins was observed, with an activity of 2701.85 hemagglutination units (HU). Previous studies have demonstrated that lectins from different bean species or varieties exhibit varying levels of hemagglutinating activity.
In tepary beans, lectin-rich extracts have been reported to have 8,618 HU, while standard bean extracts can show values ranging from 1.5 to 1.9 × 10¹⁰ HU [38,54,55]. The differences in hemagglutinating activity are primarily attributed to the source of the lectins and the type of erythrocytes used in the assays. It has been shown that lectin affinity is influenced by the oligosaccharides present on the erythrocyte membrane, as well as the origin of the cells. Since previous studies have employed erythrocytes from different sources, the observed differences in activity are likely due to these variables.
The results of this study indicated a cytotoxic effect of the tepary bean extract on epithelial cells, both in free cell assays and in intestinal fragments. A slightly greater effect (~10%) was observed in free cells, as indicated by the differences in the IC50 values. This disparity may be due to the increased exposure of free cells to the protein extract, whereas cells attached to tissue are somewhat protected from the full effect.
It has been reported that protein extracts from bean seeds can cause damage to the intestines of laboratory animals, resulting in a reduction in the activity of enzymes such as maltase, saccharase, alkaline phosphatase, and hydrolases [56,57], this damage is thought to be caused by anti-nutritional factors, such as lectins, which are present in high concentrations in several bean varieties. These lectins have been linked to adverse effects, including alterations and abnormal development of the microvilli in the small intestine [49,58].
In vivo studies have demonstrated that feeding laboratory animals beans containing biologically active lectins results in an increase in intestinal size and weight, thickening of the intestinal wall, and alterations in the size, metabolism, and function of the entire digestive tract [59,60]. Lafont et al. reported that the extent of intestinal damage caused by lectins in laboratory animals is dose-dependent, leading to irregularly sized, shortened, and spaced microvilli, as well as microvilli bleeding and even cell death [56].
The damage observed in epithelial cells after exposure to the protein extract can be attributed mainly to the lectins present in the extract, as this is the antinutritional compound found in the highest quantity (2701.85 HU). Previous studies have reported that tepary beans contain significant concentrations of lectins, which are cytotoxic to malignant colon and cervical cells, as well as to T lymphocytes and 3T3 fibroblasts [45,61,62].
The genotoxicity study revealed that, at 24 h post-treatment, the tepary bean extract did not induce genotoxic effects at any of the administered doses. However, at 48 h, a significant, dose-dependent increase in the number of micronuclei was observed. The animals appeared to recover by 72 h, as indicated by a reduction in the number of micronuclei, with no significant difference (p < 0.05) compared to the negative control.
Similar results have been reported in studies on laboratory animals, as well as through the Comet and Ames tests, which showed that seeds from the black and white kidney varieties of Phaseolus vulgaris, as well as from the Lady Joy bean, did not cause genotoxic damage, suggesting that these seeds are safe for consumption [63-64]. Analysis of human lymphocytes revealed that the aqueous extract of white kidney beans did not induce genotoxicity at the concentrations tested [66]. Additionally, the protein hydrolysate from Psophocarpus tetragonolobus (winged bean) seeds, as analyzed by the Ames test, did not induce damage [67]. Furthermore, proteins such as Sambucus nigra lectin have demonstrated DNA-protective and repairing effects [68], and the protein extract from Spirulina maxima has shown a genoprotective effect in mice [69].
Other natural proteins, such as those derived from Bauhinia monandra [64], Sambucus nigra [68], Phaseolus vulgaris L.,Vigna radiata, Arachis hypogaea L., Phaseolus vulgaris L. [48], Sapindus saponaria [65], Microgramma vacciniifolia [72], have also shown no genotoxic effects.
These effects are often attributed to the ability of certain compounds present in these extracts to trap free radicals, thereby reducing oxidative stress, a process likely mediated by phenolic compounds found in the extracts[48,65]. Some authors have demonstrated that lectins from of plant sources can protect and induce DNA repair processes [68, 70, 73] and prevent the mutagenicity of toxic agents [74].These processes could be stimulated by lymphokines or cytokines secreted by activated leukocytes[75]. The findings in the present study are consistent with these reports, as significant concentrations of biologically active lectins were found. While these lectins may have caused some cellular damage, their effects were significantly lower than those on genetic material, and were also dose- and time-dependent.
In contrast, proteins derived from various plants have demonstrated dose-dependent genotoxic effects, as observed with those from Amaranthus hypochondriacus [30], Syzygium cumini [76], Sapindus saponaria [71], Moringa oleifera Lam. [77], and Phaseolus coccineus [24].
The in vivo and in vitro cytotoxicity assay showed that the damage caused by bean proteins was dose-dependent. In the cytotoxicity study in mice, we found that, as visualized by the ratio of polychromatic to normochromatic erythrocytes (PCE/NCE), it was rising, which is an indicator of inhibition in erythrocyte division and maturation [78], indicating that the extract is related to bone marrow damage and cell production [72,7,80].
The highest cytotoxic effect was observed in animals exposed to lower doses; with the most significant damage at 48 h. Similar results were observed with Phaseolus coccineus extracts, where the highest toxicity of the extracts was at 48 h [24]. Regarding the in vitro assay, it was observed that the tepary bean extract exhibits a dose-dependent effect, with greater cytotoxicity at high doses. This was established as a mean inhibitory dose of 108.97 µg/mL in the study using intestinal fragments, while in free epithelial cells, it was 10.08 µg/m. These results show that the toxicity of the extract to the cells depends not only on the composition of the extract but also on the protection that the tissue provides to the cells as they are more exposed and susceptible to damage when they are released from the tissue. It is essential to note that either way the extract presents a risk, as consuming the proteins in their bioactive form; they can cause damage to the intestine.
Studies conducted in both cells and laboratory animals support the findings of our research. In vitro studies demonstrated that protein extracts from various varieties of Phaseolus acutifolius and Phaseolus vulgaris are cytotoxic to the epithelial cells of the small intestine in rats [4,50,58]. Also, animal studies, to which bean protein extracts were administered, reported that the proteins were toxic to organs, causing changes in intestinal permeability, alterations in the morphology of the microvilli, morphological distortion in the jejunum, alterations in the hydroelectrolytic flow of the intestine, and increased intestinal vascular permeability [51,81–87].
Other studies involving protein extracts from seeds have indicated that these extracts can be toxic to various cell types, including lymphocytes and intestinal cells [58,67,88]. Additionally, they can cause damage to organs and even lead to the death of animals when the extracts are administered directly or mixed into their feed [51,82,85,89].
The results from our studies provide important insights into the damage caused by protein extracts on intestinal cells and bone marrow, as well as the risks associated with consuming these beans if they are not adequately cooked. It has been reported that protein extracts from seeds may contain anti-nutritional compounds, such as lectins and protease inhibitors, that can cause cellular damage and harm to animals. These compounds have been shown to damage various organs and tissues and may even result in animal death [50,61,62,90–95].
Lectins, a type of protein found in these extracts, have been identified in protein extracts from several bean species, including Phaseolus vulgaris, Phaseolus coccineus, and Phaseolus acutifolius [25, 48, 61–63]. The concentration of lectins depends on various factors, including the variety, growing conditions, and extraction methods. Lectins pose a potential risk if consumed, as they have been associated with adverse effects, including damage to animal organs, cytotoxicity to both normal and malignant cells, and even death in mice. Therefore, it is crucial to ensure that these proteins are inactivated before consuming these legumes [13,16,61,96-102].
5. Conclusions
Our results indicate that TBE did not cause mortality in mice across the range of concentrations tested. However, it exhibited genotoxic effects, which were most pronounced at 48 h and decreased significantly over time. TBE also showed cytotoxicity to bone marrow, with a stimulatory effect on erythropoiesis. Additionally, it caused dose-dependent damage to rat intestinal epithelial cells, with a more pronounced effect observed when the cells were in their free form. The observed toxic effects of TBE are likely attributable to the presence of anti-nutritional compounds, such as lectins, and other substances extracted from it.
Future studies should focus on exploring the specific molecular mechanisms by which lectins and other proteins in the TBE exert their toxic effects. This could include research on protein-receptor interactions, cellular signaling pathways, and changes in gene expression in response to exposure. Additional in vivo studies are needed to evaluate long-term toxicity, organ-specific effects, and dose-response relationships. Such research will help establish safe levels and provide critical insights into the safety and potential therapeutic applications of TBE.
Author Contributions
Conceptualization, C.V.-V. and O.L.M..; methodology, L.O.-F., B.P.V.-M., G.B.-C., V.M.M.-P. and J.A.M.-G.; software, R.V.-A. and D.E.-L.; validation, R.V.-A., V.M.M.-P., L.G.-C. and A.B.-N.; formal analysis, L.G.-C. and A.B.-N.; investigation, V.M.M.-P., C.V.-V., L.O.-F., B.P.V.-M. and J.A. M.-G..; resources, L.O.-F., L.G.-C. and D.E.-L.; data curation, D.E.-L. and L.O.-F.; writing—original draft preparation, C.V.-V. L.O.-F., B.P.V.-M., O.L.-M. and G.B.-C..; writing—review and editing, All authors reviewed the written manuscript and editing was made by C.V.-V., L.O.-F., B.P.V.-M. and G.B.-C.; visualization, A.B.-N., R.V.-A. and V.M.M.-P.; supervision, C.V.-V.; project administration, C.V.-V., O.L.-M. and G.B.-C.; funding acquisition C.V.-V. and O.L.-M.. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Informed Consent Statement
The donor provided written informed consent, and the protocol was approved by the Ethics and Research Committee of the Autonomous University of Hidalgo State.
Data Availability Statement
Data are contained within the article.
Acknowledgments
Thanks to Autonomous University of Hidalgo State for the technical services during this study.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
TBE Tepay bean protein extract
MNPE micronucleated polychromatic erythrocytes
EPC polychromatic erythrocytes
PE polychromatic erythrocytes and the number
NE normochromatic erythrocytes
References
- Vikas, R.; Kalamvrezos, N.D. The Global Economy of pulses. Knowledge and Power in the Global Economy: The Effects of School Reform in a Neoliberal/Neoconservative Age: Second Edition. 2019. 91–99 p.
- Panorama Agroalimentario frijol 2016. Panorama Agroalimentario. México; 2016 Nov. chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/https://www.gob.mx/cms/uploads/attachment/file/200638/Panorama_Agroalimentario_Frijol_2016. 27 February 2025.
- Scheerens, J.C.; Tinsley, A.M; Abbas, I.R.; Weber, C.W. The Nutritional Significance of Tepary Bean Consumption. Desert Plants. 1983;5(1):11–4 y 50–6. https://repository.arizona. 1015. [Google Scholar]
- De Mejia, E.G.; Valadez-Vega, M.D.C.; Reynoso-Camacho, R.; Loarca-Pina, G. Tannins, Trypsin Inhibitors and Lectin Cytotoxicity in Tepary (Phaseolus acutifolius) and Common (Phaseolus vulgaris) Beans. Plant Foods Hum. Nutr. 2005, 60, 137–145. [Google Scholar] [CrossRef]
- Salas-López, F.; Gutiérrez-Dorado, R.; Milán-Carrillo, J.; Cuevas-Rodríguez, E.O.; Canizalez-Roman, V.A.; León-Sicairos C., R.; Reyes-Moreno, C. Nutritional and antioxidant potential of a desert underutilized legume - tepary bean (Phaseolus acutifolius). Optimization of germination bioprocess. Food Sci. Technol (Brazil). 2018;38:254–62. https://www.scielo.br/j/cta/a/XYLNRcWKDM4vkhnBTM4DjKz/?
- Michaels T.E. Grain Legumes and Their Dietary Impact: Overview [Internet]. 2nd ed. Reference Module in Food Science. Elsevier.; 2016. 1–8 p. Available from: http://dx.doi.org/10.1016/B978-0-08-100596-5.00040-8.
- Tharanathan, R.; Mahadevamma, S. Grain legumes—a boon to human nutrition. Trends Food Sci. Technol. 2003, 14, 507–518. [Google Scholar] [CrossRef]
- Gilani, G.S.; Xiao, C.W.; Cockell, K.A. Impact of Antinutritional Factors in Food Proteins on the Digestibility of Protein and the Bioavailability of Amino Acids and on Protein Quality. Br. J. Nutr. 2012, 108, S315–S332. [Google Scholar] [CrossRef]
- Cominelli, E.; Sparvoli, F.; Lisciani, S.; Forti, C.; Camilli, E.; Ferrari, M.; Le Donne, C.; Marconi, S.; Vorster, B.J.; Botha, A.-M.; et al. Antinutritional factors, nutritional improvement, and future food use of common beans: A perspective. Front. Plant Sci. 2022, 13, 992169. [Google Scholar] [CrossRef]
- Gilani, G.S.; A Cockell, K.; Sepehr, E. Effects of Antinutritional Factors on Protein Digestibility and Amino Acid Availability in Foods. J. AOAC Int. 2005, 88, 967–987. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Guzmán, N.; Gallegos-Infante, J.; González-Laredo, R.; Castillo-Antonio, P.; Delgado-Licon, E.; Ibarra-Pérez, F. Functional properties of three common bean (Phaseolus vulgaris) cultivars stored under accelerated conditions followed by extrusion. LWT 2006, 39, 6–10. [Google Scholar] [CrossRef]
- Grant, G.; More, L.J.; McKenzie, N.H.; Pusztai, A. The effect of heating on the haemagglutinating activity and nutritional properties of bean (Phaseolus vulgaris) seeds. J. Sci. Food Agric. 1982, 33, 1324–1326. [Google Scholar] [CrossRef] [PubMed]
- Arija, I.; Centeno, C.; Viveros, A.; Brenes, A.; Marzo, F.; Illera, J.C.; Silvan, G. Nutritional Evaluation of Raw and Extruded Kidney Bean (Phaseolus vulgaris L. var. Pinto) in Chicken Diets. Poult. Sci. 2006, 85, 635–644. [Google Scholar] [CrossRef]
- Yuan, S.; Chang, S.K.C.; Liu, Z.; Xu, B. Elimination of Trypsin Inhibitor Activity and Beany Flavor in Soy Milk by Consecutive Blanching and Ultrahigh-Temperature (UHT) Processing. J. Agric. Food Chem. 2008, 56, 7957–7963. [Google Scholar] [CrossRef]
- Batista, K.A.; Prudêncio, S.H.; Fernandes, K.F. Changes in the Functional Properties and Antinutritional Factors of Extruded Hard-to-Cook Common Beans ( Phaseolus vulgaris, L. ). J. Food Sci. 2010, 75, C286–C290. [Google Scholar] [CrossRef] [PubMed]
- Batista, K.A.; Prudêncio, S.H.; Fernandes, K.F. Changes in the Functional Properties and Antinutritional Factors of Extruded Hard-to-Cook Common Beans ( Phaseolus vulgaris, L. ). J. Food Sci. 2010, 75, C286–C290. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, C.; Cordoba-Diaz, D.; Cordoba-Diaz, M.; Girbés, T.; Jiménez, P. Effects of temperature, pH and sugar binding on the structures of lectins ebulin f and SELfd. Food Chem. 2017, 220, 324–330. [Google Scholar] [CrossRef]
- Demma, J.; Engidawork, E.; Hellman, B. Potential genotoxicity of plant extracts used in Ethiopian traditional medicine. J. Ethnopharmacol. 2009, 122, 136–142. [Google Scholar] [CrossRef]
- Schmid, W. The micronucleus test. Mutat. Res. Mutagen. Relat. Subj. 1975, 31, 9–15. [Google Scholar] [CrossRef]
- Alice, C.; Vargas, V.; Silva, G.; de Siqueira, N.; Schapoval, E.; Gleye, J.; Henriques, J.; Henriques, A. Screening of plants used in south Brazilian folk medicine. J. Ethnopharmacol. 1991, 35, 165–171. [Google Scholar] [CrossRef]
- Krishna, G.; Hayashi, M. In vivo rodent micronucleus assay: protocol, conduct and data interpretation. Mutat. Res. Mol. Mech. Mutagen. 2000, 455, 155–166. [Google Scholar] [CrossRef]
- Zúñiga-González, G.; Gómez-Meda, B.; Zamora-Perez, A.; Ramos-Ibarra, M.; Batista-González, C.; Espinoza-Jiménez, S.; Gallegos-Arreola, M.; Álvarez-Moya, C.; Torres-Bugarín, O. Induction of micronuclei in proestrus vaginal cells from colchicine- and cyclophosphamide-treated rats. Environ. Mol. Mutagen. 2003, 42, 306–310. [Google Scholar] [CrossRef]
- Gómez-Meda, B.C.; Zúñiga-González, G.M.; Zamora-Perez, A.; Ramos-Ibarra, M.L.; Batista-González, C.M.; Torres-Mendoza, B.M. Folate supplementation of cyclophosphamide-treated mothers diminishes micronucleated erythrocytes in peripheral blood of newborn rats. Environ. Mol. Mutagen. 2004, 44, 174–178. [Google Scholar] [CrossRef]
- Zamora-Perez, A.L.; Lazalde-Ramos, B.P.; Sosa-Macías, M.G.; Gómez-Meda, B.C.; Torres-Bugarín, O.; Zúñiga-González, G.M. Methylphenidate lacks genotoxic effects in mouse peripheral blood erythrocytes. Drug Chem. Toxicol. 2011, 34, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Teniente-Martínez, G.; Bernardino-Nicanor, A.; Cariño-Cortés, R.; Valadez-Vega, M.d.C.; Montañez-Soto, J.L.; Acosta-García, G.; González-Cruz, L. Cytotoxic and genotoxic activity of protein isolate of ayocote beans and anticancer activity of their protein fractions. J. Food Meas. Charact. 2019, 13, 1040–1048. [Google Scholar] [CrossRef]
- Ringe, M.L.; Love, M.H. Kinetics of Protein Quality Change in an Extruded Cowpea-Corn Flour Blend under Varied Steady-State Storage Conditions. J Food Sci. 1988;53(2):584–8. [CrossRef]
- Sánchez-Vioque, R.; Clemente, A.; Vioque, J.; Bautista, J.; Millán, F. Protein isolates from chickpea (Cicer arietinum L.): chemical composition, functional properties and protein characterization. Food Chem. 1999, 64, 237–243. [Google Scholar] [CrossRef]
- Lqari, H.; Vioque, J.; Pedroche, J.; Millán, F. Lupinus angustifolius protein isolates: chemical composition, functional properties and protein characterization. Food Chem. 2002, 76, 349–356. [Google Scholar] [CrossRef]
- Horax, R.; Hettiarachchy, N.S.; Chen, P.; Jalaluddin, M. Preparation and Characterization of Protein Isolate from Cowpea ( Vigna unguiculata L. Walp.). J. Food Sci. 2004, 69, fct114–fct118. [Google Scholar] [CrossRef]
- León, J.C.M.; Vázquez-Mata, N.; Torres, N.; Gil-Zenteno, L.; Bressani, R. Preparation and Characterization of Protein Isolate from Fresh and Hardened Beans ( Phaseolus vulgaris L.). J. Food Sci. 2007, 72, C96–C102. [Google Scholar] [CrossRef]
- León, J.C.M.; Vázquez-Mata, N.; Torres, N.; Gil-Zenteno, L.; Bressani, R. Preparation and Characterization of Protein Isolate from Fresh and Hardened Beans ( Phaseolus vulgaris L.). J. Food Sci. 2007, 72, C96–C102. [Google Scholar] [CrossRef]
- Valadez-Vega, C.; Lugo-Magaña, O.; Morales-González, J.A.; Delgado-Olivares, L.; Izquierdo-Vega, J.A.; Sánchez-Gutiérrez, M.; López-Contreras, L.; Bautista, M.; Velázquez-González, C. Phytochemical, cytotoxic, and genotoxic evaluation of protein extract ofAmaranthus hypochondriacusseeds. CyTA - J. Food 2021, 19, 701–709. [Google Scholar] [CrossRef]
- Mercado del Frijol situación y prospectiva. Mercado del frijol, situación y prospectiva. Palacio Legislativo de San Lázaro, Ciudad de México. Febrero 2020. Available from: http://www.cedrssa.gob. 22 January 2025.
- Feeny, P. Inhibitory effect of oak leaf tannins on the hydrolysis of proteins by trypsin. Phytochemistry 1969, 8, 2119–2126. [Google Scholar] [CrossRef]
- Valadez-Vega, C.; Lugo-Magaña, O.; Figueroa-Hernández, C.; Bautista, M.; Betanzos-Cabrera, G.; Bernardino-Nicanor, A.; González-Amaro, R.M.; Alonso-Villegas, R.; Morales-González, J.A.; González-Cruz, L. Effects of Germination and Popping on the Anti-Nutritional Compounds and the Digestibility of Amaranthus hypochondriacus Seeds. Foods 2022, 11, 2075. [Google Scholar] [CrossRef] [PubMed]
- Sgarbieri, V.C.; Galeazzi, M.A.M. QUANTIFICATION AND SOME CHEMICAL AND BIOCHEMICAL CHARACTERIZATION OF NITROGENOUS SUBSTANCES FROM VARIETIES OF COMMON BEANS (PHASEOLUS VULGARIS, L.). J. Food Biochem. 1990, 14, 233–247. [Google Scholar] [CrossRef]
- Jafari, M.; Rajabzadeh, A.R.; Tabtabaei, S.; Marsolais, F.; Legge, R.L. Physicochemical characterization of a navy bean (Phaseolus vulgaris) protein fraction produced using a solvent-free method. Food Chem. 2016, 208, 35–41. [Google Scholar] [CrossRef]
- Rizvi, N.B.; Aleem, S.; Khan, M.R.; Ashraf, S.; Busquets, R. Quantitative Estimation of Protein in Sprouts of Vigna radiate (Mung Beans), Lens culinaris (Lentils), and Cicer arietinum (Chickpeas) by Kjeldahl and Lowry Methods. Molecules 2022, 27, 814. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Albores, F.; de la Fuente, G.; Agundis, C.; Córdoba, F. Purification and Characterization of A Lectin from Phaseolus Acu-Tifolius Var. Latifolius. Prep. Biochem. 1987, 17, 379–396. [Google Scholar] [CrossRef] [PubMed]
- Valadez-Vega, C.; Guzmán-Partida, A.M.; Soto-Cordova, F.J.; Álvarez-Manilla, G.; Morales-González, J.A.; Madrigal-Santillán, E.; Villagómez-Ibarra, J.R.; Zúñiga-Pérez, C.; Gutiérrez-Salinas, J.; Becerril-Flores, M.A. Purification, Biochemical Characterization, and Bioactive Properties of a Lectin Purified from the Seeds of White Tepary Bean (Phaseolus Acutifolius Variety Latifolius). Molecules 2011, 16, 2561–2582. [Google Scholar] [CrossRef]
- Avilés-Gaxiola, S.; Chuck-Hernández, C.; Saldívar, S.O.S. Inactivation Methods of Trypsin Inhibitor in Legumes: A Review. J. Food Sci. 2017, 83, 17–29. [Google Scholar] [CrossRef]
- De Bock, P.; Daelemans, L.; Selis, L.; Raes, K.; Vermeir, P.; Eeckhout, M.; Van Bockstaele, F. Comparison of the Chemical and Technological Characteristics of Wholemeal Flours Obtained from Amaranth (Amaranthus sp.), Quinoa (Chenopodium quinoa) and Buckwheat (Fagopyrum sp.) Seeds. Foods 2021, 10, 651. [Google Scholar] [CrossRef] [PubMed]
- Elkowicz, K.; Sosulski, F.W. Antinutritive Factors in Eleven Legumes and Their Air-classified Protein and Starch Fractions. J. Food Sci. 1982, 47, 1301–1304. [Google Scholar] [CrossRef]
- Olloqui, E.J.; Castañeda-Ovando, A.; Evangelista-Lozano, S.; Alanís-García, E.; Ramírez-Moreno, E.; Valadez-Vega, C.; Añorve-Morga, J. Measurement of nutrients and minor components of a non-toxic variety of Jatropha curcas. J. Food Meas. Charact. 2021, 16, 1029–1037. [Google Scholar] [CrossRef]
- Valadez-Vega, C.; Lugo-Magaña, O.; Figueroa-Hernández, C.; Bautista, M.; Betanzos-Cabrera, G.; Bernardino-Nicanor, A.; González-Amaro, R.M.; Alonso-Villegas, R.; Morales-González, J.A.; González-Cruz, L. Effects of Germination and Popping on the Anti-Nutritional Compounds and the Digestibility of Amaranthus hypochondriacus Seeds. Foods 2022, 11, 2075. [Google Scholar] [CrossRef]
- González-Cruz, L.; Valadez-Vega, C.; Juárez-Goiz, J.M.S.; Flores-Martínez, N.L.; Montañez-Soto, J.L.; Bernardino-Nicanor, A. Partial Purification and Characterization of the Lectins of Two Varieties of Phaseolus coccineus (Ayocote Bean). Agronomy 2022, 12, 716. [Google Scholar] [CrossRef]
- Valadez-Vega, C.; Lugo-Magaña, O.; Betanzos-Cabrera, G.; Villagómez-Ibarra, J.R. Partial Characterization of Lectins Purified from the Surco and Vara (Furrow and Rod) Varieties of Black Phaseolus vulgaris. Molecules 2022, 27, 8436. [Google Scholar] [CrossRef]
- Moreno-Celis, U.; López-Martínez, J.; Blanco-Labra, A.; Cervantes-Jiménez, R.; Estrada-Martínez, L.E.; García-Pascalin, A.E.; Guerrero-Carrillo, M.D.J.; Rodríguez-Méndez, A.J.; Mejía, C.; Ferríz-Martínez, R.A.; et al. Phaseolus acutifolius Lectin Fractions Exhibit Apoptotic Effects on Colon Cancer: Preclinical Studies Using Dimethilhydrazine or Azoxi-Methane as Cancer Induction Agents. Molecules 2017, 22, 1670. [Google Scholar] [CrossRef]
- Chel-Guerrero, L.; Gallegos-Tintoré, S.; Martínez-Ayala, A.; Castellanos-Ruelas, A.; Betancur-Ancona, D. Functional Properties of Proteins from Lima Bean (Phaseolus lunatus L.) Seeds. Food Sci. Technol. Int. 2011, 17, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Inthachat, W.; Suttisansanee, U.; Kruawan, K.; On-Nom, N.; Chupeerach, C.; Temviriyanukul, P. Evaluation of Mutagenicity and Anti-Mutagenicity of Various Bean Milks Using Drosophila with High Bioactivation. Foods 2022, 11, 3090. [Google Scholar] [CrossRef] [PubMed]
- King, T.; Pusztai, A.; Clarke, E. Kidney bean (Phaseolus vulgaris) lectin-induced lesions in rat small intestine: 1. Light microscope studies. J. Comp. Pathol. 1980, 90, 585–595. [Google Scholar] [CrossRef]
- Sousa, V.; Sotelo, A.; González-Garza, M.T. Differential cytotoxicity of the isolated protein fraction of Escumite bean (Phaseolus acutifolius). Plant Foods Hum. Nutr. 1982, 31, 319–325. [Google Scholar] [CrossRef]
- Moreira, R.d.A.; Ainouz, I.L.; de Oliveira, J.T.A.; Cavada, B.S. Plant lectins, chemical and biological aspects. Mem. Do Inst. Oswaldo Cruz 1991, 86, 211–218. [Google Scholar] [CrossRef]
- Muramoto, K. Lectins as Bioactive Proteins in Foods and Feeds. Food Sci. Technol. Res. 2017, 23, 487–494. [Google Scholar] [CrossRef]
- Greer, F.; Pusztai, A. Toxicity of Kidney Bean (Phaseolus vulgaris) in Rats: Changes in Intestinal Permeability. Digestion 1985, 32, 42–46. [Google Scholar] [CrossRef]
- He, S.; Simpson, B.K.; Sun, H.; Ngadi, M.O.; Ma, Y.; Huang, T. Phaseolus vulgaris lectins: A systematic review of characteristics and health implications. Crit. Rev. Food Sci. Nutr. 2017, 58, 70–83. [Google Scholar] [CrossRef]
- Santiago, J.; Levy-Benshimol, A.; Carmona, A. Effect of Phaseolus vulgaris lectins on glucose absorption, transport, and metabolism in rat everted intestinal sacs. J. Nutr. Biochem. 1993, 4, 426–430. [Google Scholar] [CrossRef]
- Alcázar-Valle, M.; Lugo-Cervantes, E.; Mojica, L.; Morales-Hernández, N.; Reyes-Ramírez, H.; Enríquez-Vara, J.N.; García-Morales, S. Bioactive Compounds, Antioxidant Activity, and Antinutritional Content of Legumes: A Comparison between Four Phaseolus Species. Molecules 2020, 25, 3528. [Google Scholar] [CrossRef]
- Lafont, J.; Rouanet, J.; Gabrion, J.; Assouad, J.; Infante, Z.; Besançon, P. Duodenal Toxicity of Dietary Phaseolus vulgaris Lectins in the Rat: An Integrative Assay. Digestion 1988, 41, 83–93. [Google Scholar] [CrossRef]
- Nciri, N.; Cho, N. New research highlights: Impact of chronic ingestion of white kidney beans (Phaseolus vulgaris L. var. Beldia) on small-intestinal disaccharidase activity in Wistar rats. Toxicol. Rep. 2018, 5, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Sotelo, A.; Arteaga, M.E.; Frías, M.I.; González-Garza, M.T. Cytotoxic effect of two legumes in epithelial cells of the small intestine. Plant Foods Hum. Nutr. 1980, 30, 79–85. [Google Scholar] [CrossRef]
- Huisman, J.; Van Der Poel, A.F.B.; Mouwen, J.M.V.M.; Weerden, J.M.V.M. Effect of variable protein contents in diets containing Phaseolus vulgaris beans on performance, organ weights and blood variables in piglets, rats and chickens. Br. J. Nutr. 1990, 64, 755–764. [Google Scholar] [CrossRef]
- Bardocz, S.; Grant, G.; Ewen, S.W.; Duguid, T.J.; Brown, D.S.; Englyst, K.; Pusztai, A. Reversible effect of phytohaemagglutinin on the growth and metabolism of rat gastrointestinal tract. Gut 1995, 37, 353–360. [Google Scholar] [CrossRef]
- Castillo-Villanueva, A.; Caballero-Ortega, H.; Abdullaev-Jafarova, F.; Garfias, Y.; Jiménez-Martínez, M.d.C.; Bouquelet, S.; Martínez, G.; Mendoza-Hernández, G.; Zenteno, E. Lectin fromPhaseolus acutifoliusvar. escumite: Chemical Characterization, Sugar Specificity, and Effect on Human T-Lymphocytes. J. Agric. Food Chem. 2007, 55, 5781–5787. [Google Scholar] [CrossRef]
- Valadez-Vega, C.; Alvarez-Manilla, G.; Riverón-Negrete, L.; García-Carrancá, A.; Morales-González, J.A.; Zuñiga-Pérez, C.; Madrigal-Santillán, E.; Esquivel-Soto, J.; Esquivel-Chirino, C.; Villagómez-Ibarra, R.; et al. Detection of Cytotoxic Activity of Lectin on Human Colon Adenocarcinoma (Sw480) and Epithelial Cervical Carcinoma (C33-A). Molecules 2011, 16, 2107–2118. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, L.; Gomes, J.; Stringheta, P.; Gontijo, Á.; Padovani, C.; Ribeiro, L.; Salvadori, D. Black bean (Phaseolus vulgaris L.) as a protective agent against DNA damage in mice. Food Chem. Toxicol. 2003, 41, 1671–1676. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-T.; Chiu, C.-Y.; Huang, C.-F.; Peng, F.-C.; Liu, S.-H. Genotoxicity and 28-day oral toxicity studies of a functional food mixture containing maltodextrin, white kidney bean extract, mulberry leaf extract, and niacin-bound chromium complex. Regul. Toxicol. Pharmacol. 2018, 92, 67–74. [Google Scholar] [CrossRef]
- Frassinetti, S.; Gabriele, M.; Caltavuturo, L.; Longo, V.; Pucci, L. Antimutagenic and Antioxidant Activity of a Selected Lectin-free Common Bean (Phaseolus vulgaris L.) in Two Cell-based Models. Plant Foods Hum. Nutr. 2015, 70, 35–41. [Google Scholar] [CrossRef]
- Pereira, L.L.S.; Marcussi, S.; Sátiro, L.C.; Pereira, C.A.; Andrade, L.F.; Davide, L.C.; dos Santos, C.D. Application of Comet assay to assess the effects of white bean meal on DNA of human lymphocytes. Braz. J. Pharm. Sci. 2012, 48, 103–108. [Google Scholar] [CrossRef]
- Kawanding, O.J.; Abdullah, N.; Noor, Z.M.; Hashim, N.; Saari, N.; Yahya, M.F.Z.; Abdullah, M.F.F. Genotoxicity and Cytotoxicity Evaluation of Winged Bean (Psophocarpus Tetragonolobus) Protein Hydrolysate. Malaysian Applied Biology. 2015;44(3):6–11.ttps://www.academia. 1698. [Google Scholar]
- Lylo, V.; Karpova, I.; Kotsarenko, K.; Macewicz, L.; Ruban, T.; Lukash, L. LECTINS OF SAMBUCUS NIGRA IN REGULATION OF CELLULAR DNA-PROTECTIVE MECHANISMS. Acta Hortic. 2015, 103–108. [Google Scholar] [CrossRef]
- Álvarez-González, I.; Vázquez-Sánchez, J.; Chamorro-Cevallos, G.; Madrigal-Bujaidar, E. Effect of Spirulina maxima and Its Protein Extract on Micronuclei Induction by Hydroxyurea in Pregnant Mice and Their Fetuses. J. Med. Food 2013, 16, 992–996. [Google Scholar] [CrossRef] [PubMed]
- Sisenando, H.A.A.A.C.N.; Macedo, M.F.S.; Saturnino, A.C.R.D.; Coelho, L.C.B.B.; de Medeiros, S.R.B. Evaluation of the genotoxic potential of Bauhinia monandra leaf lectin (BmoLL). Food Chem. Toxicol. 2009, 47, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Tavares, G.D.B.; Aiub, C.A.F.; Felzenszwalb, I.; Dantas, E.K.C.; Araújo-Lima, C.F.; Júnior, C.L.S. In vitro biochemical characterization and genotoxicity assessment of Sapindus saponaria seed extract. J. Ethnopharmacol. 2021, 276, 114170. [Google Scholar] [CrossRef] [PubMed]
- da Silva, G.C.; de Oliveira, A.M.; Machado, J.C.B.; Ferreira, M.R.A.; de Medeiros, P.L.; Soares, L.A.L.; de Souza, I.A.; Paiva, P.M.G.; Napoleão, T.H. Toxicity assessment of saline extract and lectin-rich fraction from Microgramma vacciniifolia rhizome. Toxicon 2020, 187, 65–74. [Google Scholar] [CrossRef]
- Karpova, I.; Lylo, V.; Macewicz, L.; Kotsarenko, K.; Palchykovska, L.; Ruban, T.; Lukash, L. LECTINS OF SAMBUCUS NIGRA AS BIOLOGICALLY ACTIVE AND DNA-PROTECTIVE SUBSTANCES. Acta Hortic. 2015, 93–102. [Google Scholar] [CrossRef]
- Hong, C.; Lyu, S. The Antimutagenic Effect of Mistletoe Lectin (Viscum album L. var. coloratum agglutinin). Phytotherapy Res. 2011, 26, 787–790. [Google Scholar] [CrossRef]
- Hoessli, D.C.; Ahmad, I. Mistletoe Lectins: Carbohydrate-Specific Apoptosis Inducers and Immunomodulators. Curr. Org. Chem. 2008, 12, 918–925. [Google Scholar] [CrossRef]
- H.S. Nandini, N.; Prabhudas, A. V. M. H.S. Nandini, N.; Prabhudas, A. V. M.; Krutty P.R.K.; P.K. Partial characterization, cytotoxic and genotoxic properties of glycoproteins from Syzygium cumini Lam. Skeel (Black plum). Int J of Pharmaceutical Sci and Res. 2020;1(12):6333–41. [CrossRef]
- De Barros, M.C.; Videres, L.C.C.d.A.; Batista, A.M.; Guerra, M.M.P.; Coelho, L.C.B.B.; Da Silva, T.G.; Napoleão, T.H.; Paiva, P.M.G. Cytotoxicity and genotoxicity assessment of the extract and lectins from Moringa oleifera Lam. Seeds / Avaliação da citotoxicidade e genotoxicidade do extrato e lectinas das sementes de Moringa oleifera Lam. Braz. J. Dev. 2021, 7, 94854–94869. [Google Scholar] [CrossRef]
- Suzuki, Y.; Nagae, Y.; Li, J.; Sakaba, H.; Mozawa, K.; Takahashi, A.; Shimizu, H. The micronucleus test and erythropoiesis. Effects of erythropoietin and a mutagen on the ratio of polychromatic to normochromatic erythrocytes (P/N ratio). Mutagenesis 1989, 4, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Sung, J.H.; Ji, J.H.; Song, K.S.; Lee, J.H.; Kang, C.S.; Yu, I.J. In vivo Genotoxicity of Silver Nanoparticles after 90-day Silver Nanoparticle Inhalation Exposure. Saf. Heal. Work. 2011, 2, 34–38. [Google Scholar] [CrossRef]
- Araujo-Espino, D.I.; Zamora-Perez, A.L.; Zúñiga-González, G.M.; Gutiérrez-Hernández, R.; Morales-Velazquez, G.; Lazalde-Ramos, B.P. Genotoxic and cytotoxic evaluation of Jatropha dioica Sessé ex Cerv. by the micronucleus test in mouse peripheral blood. Regul. Toxicol. Pharmacol. 2017, 86, 260–264. [Google Scholar] [CrossRef]
- Myer, R.O.; Froseth, J.A.; Coon, C.N. Protein Utilization and Toxic Effects of Raw Beans (Phaseolus Vulgaris) for Young Pigs2. J. Anim. Sci. 1982, 55, 1087–1098. [Google Scholar] [CrossRef]
- Bender, A.E.; Reaidi, G.B. Toxicity of Kidney Beans (Phaseolus Vulgaris) With Particular Reference to Lectins. J. Plant Foods 1982, 4, 15–22. [Google Scholar] [CrossRef]
- Kumar, S.; Verma, A.K.; Das, M.; Jain, S.; Dwivedi, P.D. Clinical complications of kidney bean (Phaseolus vulgaris L.) consumption. Nutrition 2013, 29, 821–827. [Google Scholar] [CrossRef]
- Nciri, N.; Cho, N.; Bergaoui, N.; El Mhamdi, F.; Ben Ammar, A.; Trabelsi, N.; Zekri, S.; Guémira, F.; Ben Mansour, A.; Sassi, F.H.; et al. Effect of White Kidney Beans (Phaseolus vulgarisL. var. Beldia) on Small Intestine Morphology and Function in Wistar Rats. J. Med. Food 2015, 18, 1387–1399. [Google Scholar] [CrossRef] [PubMed]
- Nciri, N.; Cho, N.; El Mhamdi, F.; Ben Ismail, H.; Ben Mansour, A.; Sassi, F.H.; Ben Aissa-Fennira, F. Toxicity Assessment of Common Beans (Phaseolus vulgarisL.) Widely Consumed by Tunisian Population. J. Med. Food 2015, 18, 1049–1064. [Google Scholar] [CrossRef]
- Luna-Vital, D.A.; de Mejía, E.G.; Loarca-Piña, G. Dietary Peptides from Phaseolus vulgaris L. Reduced AOM/DSS-Induced Colitis-Associated Colon Carcinogenesis in Balb/c Mice. Plant Foods Hum. Nutr. 2017, 72, 445–447. [Google Scholar] [CrossRef]
- Nciri, N.; Cho, N. New research highlights: Impact of chronic ingestion of white kidney beans (Phaseolus vulgaris L. var. Beldia) on small-intestinal disaccharidase activity in Wistar rats. Toxicol. Rep. 2018, 5, 46–55. [Google Scholar] [CrossRef]
- Lu, X. Cytotoxicity evaluation of the whole protein extract from Bar-transgenic rice on Mus musculus lymphocytes. Genet. Mol. Res. 2017, 16. [Google Scholar] [CrossRef] [PubMed]
- Idouraine, A.; Sathe, S.K.; Weber, C.W. Biological evaluation of flour and protein extract of tepary bean (Phaseolus acutifolius). J. Agric. Food Chem. 1992, 40, 1856–1859. [Google Scholar] [CrossRef]
- Zambre M.; Montagu M Van.; Angenon G.; Terryn N. Tepary Beans (Phaseolus acutifolius). Methods Mol. Biol. 2006;343:407–14.
- Osman, M.A.; Reis, P.M.; Weber, C.W. The Effect of Feeding Tepary Bean (Phaseolus acutifolius) Proteinase Inhibitors on the Growth and Pancreas of Young Mice. Pak.J. Nutr. 2003;2(3):111–5. https://scialert.net/abstract/?doi=pjn.2003.111.
- Sharon, N. Lectins: past, present and future1. Biochem. Soc. Trans. 2008, 36, 1457–1460. [Google Scholar] [CrossRef] [PubMed]
- Van Damme E.J.M; Rouge P, Peumans W.J. 3.26 plant lectins. In 2007. p. 563–99.
- Wati, R.K.; Theppakorn, T.; Benjakul, S.; Rawdkuen, S. Trypsin Inhibitor from 3 Legume Seeds: Fractionation and Proteolytic Inhibition Study. J. Food Sci. 2010, 75, C223–C228. [Google Scholar] [CrossRef]
- Faheina-Martins, G.V.; da Silveira, A.L.; Ramos, M.V.; Marques-Santos, L.F.; Araujo, D.A.M. Influence of fetal bovine serum on cytotoxic and genotoxic effects of lectins in MCF-7 cells. J. Biochem. Mol. Toxicol. 2011, 25, 290–296. [Google Scholar] [CrossRef]
- Ren, J.; Shi, J.; Kakuda, Y.; Kim, D.; Xue, S.J.; Zhao, M.; Jiang, Y. Phytohemagglutinin isolectins extracted and purified from red kidney beans and its cytotoxicity on human H9 lymphoma cell line. Sep. Purif. Technol. 2008, 63, 122–128. [Google Scholar] [CrossRef]
- Sharma, A.; Wong, J.H.; Lin, P.; Chan, Y.S.; Ng, T.B. Purification and Characterization of a Lectin from the Indian Cultivar of French Bean Seeds. Protein Pept. Lett. 2010, 17, 221–227. [Google Scholar] [CrossRef]
- Saha, S.; Singh, G.; Mahajan, V.; Gupta, H.S. Variability of Nutritional and Cooking Quality in Bean (Phaseolus vulgaris L) as a Function of Genotype. Plant Foods Hum. Nutr. 2009, 64, 174–180. [Google Scholar] [CrossRef]
- R.A. Ynalves L.M.F.; C.V Sanchez. Comparison and Temperature Study of Lectin Activities in Texas Live Oak (Quercus fusiformis) Crude Extract. J Plant Sci. 2011;6(3):124–34. https://scialert.net/abstract/?doi=jps.2011.124.
- Shi, L.; Mu, K.; Arntfield, S.D.; Nickerson, M.T. Changes in levels of enzyme inhibitors during soaking and cooking for pulses available in Canada. J. Food Sci. Technol. 2017, 54, 1014–1022. [Google Scholar] [CrossRef]
- Wainaina, I.; Wafula, E.; Sila, D.; Kyomugasho, C.; Grauwet, T.; Van Loey, A.; Hendrickx, M. Thermal treatment of common beans (Phaseolus vulgaris L.): Factors determining cooking time and its consequences for sensory and nutritional quality. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3690–3718. [Google Scholar] [CrossRef] [PubMed]
- Valadez-Vega, C.; Lugo-Magaña, O.; Figueroa-Hernández, C.; Bautista, M.; Betanzos-Cabrera, G.; Bernardino-Nicanor, A.; González-Amaro, R.M.; Alonso-Villegas, R.; Morales-González, J.A.; González-Cruz, L. Effects of Germination and Popping on the Anti-Nutritional Compounds and the Digestibility of Amaranthus hypochondriacus Seeds. Foods 2022, 11, 2075. [Google Scholar] [CrossRef]
- Waterborg, J.H.; Matthews, H.R. Method for Quantitation. Protein Protocols Handbook. 1984;1(173):7–9. [CrossRef]
- Valadez-Vega, C.; Lugo-Magaña, O.; Betanzos-Cabrera, G.; Villagómez-Ibarra, J.R. Partial Characterization of Lectins Purified from the Surco and Vara (Furrow and Rod) Varieties of Black Phaseolus vulgaris. Molecules 2022, 27, 8436. [Google Scholar] [CrossRef]
- Lorke, D. A new approach to practical acute toxicity testing. Arch. Toxicol. 1983, 54, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Madrigal-Santillán, E.; Álvarez-González, I.; Márquez-Márquez, R.; Velázquez-Guadarrama, N.; Madrigal-Bujaidar, E. Inhibitory Effect of Mannan on the Toxicity Produced in Mice Fed Aflatoxin B1 Contaminated Corn. Arch. Environ. Contam. Toxicol. 2007, 53, 466–472. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Cytotoxicity of TBE in epithelial cells and intestinal fragments. Cells and intestinal fragments were exposed to the indicated concentration of TBE for 15 min. Cell viability was determinated as described in the experimental section. The results are presented as the percentage of viable cells relative to untreated cells, which are considered 100% viable. Each value represents the mean±standard deviation.
Figure 1.
Cytotoxicity of TBE in epithelial cells and intestinal fragments. Cells and intestinal fragments were exposed to the indicated concentration of TBE for 15 min. Cell viability was determinated as described in the experimental section. The results are presented as the percentage of viable cells relative to untreated cells, which are considered 100% viable. Each value represents the mean±standard deviation.
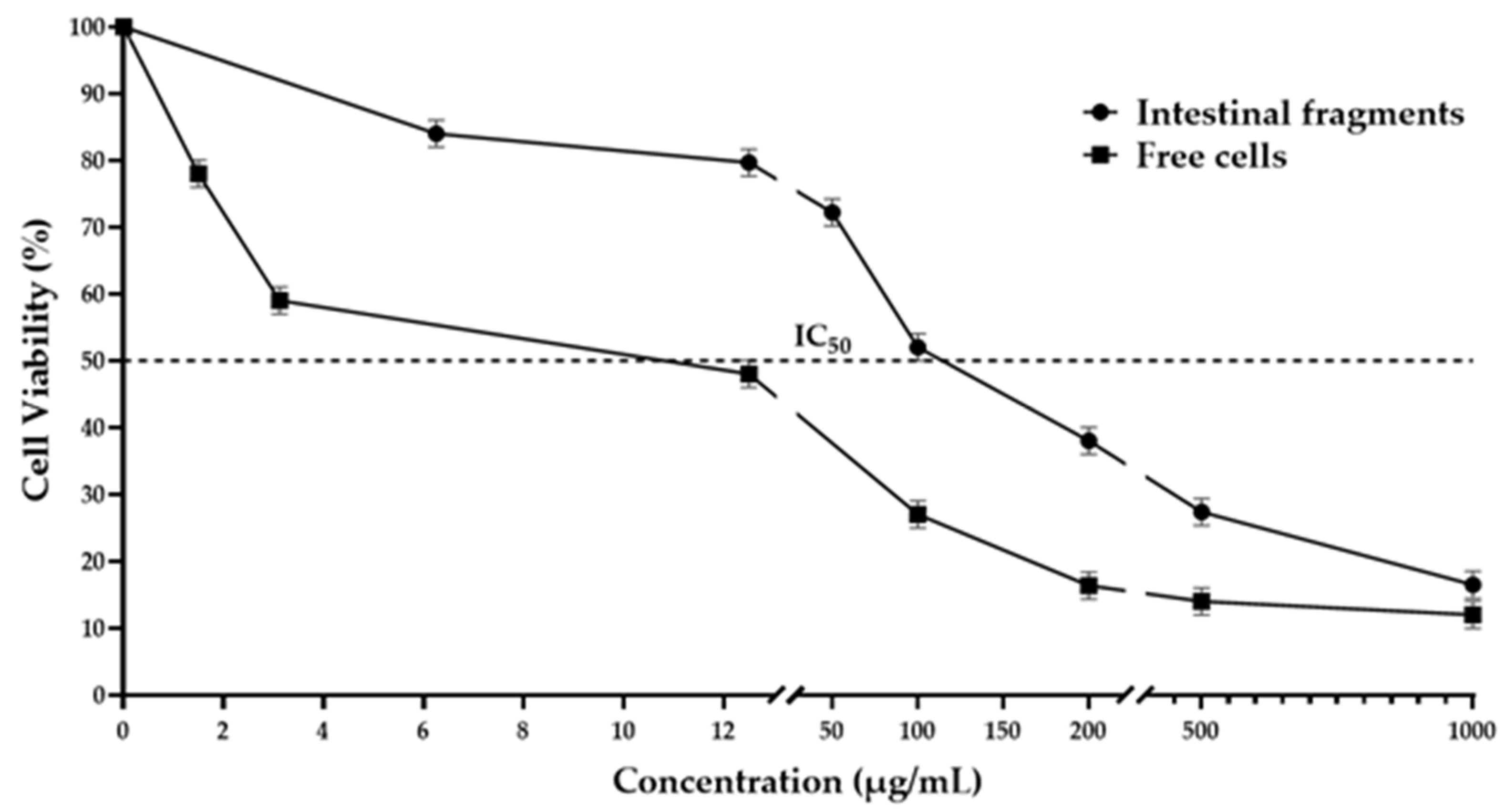
Figure 2.
Frequency of micronucleated polychromatic erythrocytes (MNPE) in mice treated with the protein extract from Phaseolus acutifolius seeds (TBE) and daunorubicin (DAU). Values represent the mean ± standard deviation of seven mice per group. * indicates a statistically significant difference from the negative control. ANOVA and Tukey tests (p < 0.05).
Figure 2.
Frequency of micronucleated polychromatic erythrocytes (MNPE) in mice treated with the protein extract from Phaseolus acutifolius seeds (TBE) and daunorubicin (DAU). Values represent the mean ± standard deviation of seven mice per group. * indicates a statistically significant difference from the negative control. ANOVA and Tukey tests (p < 0.05).
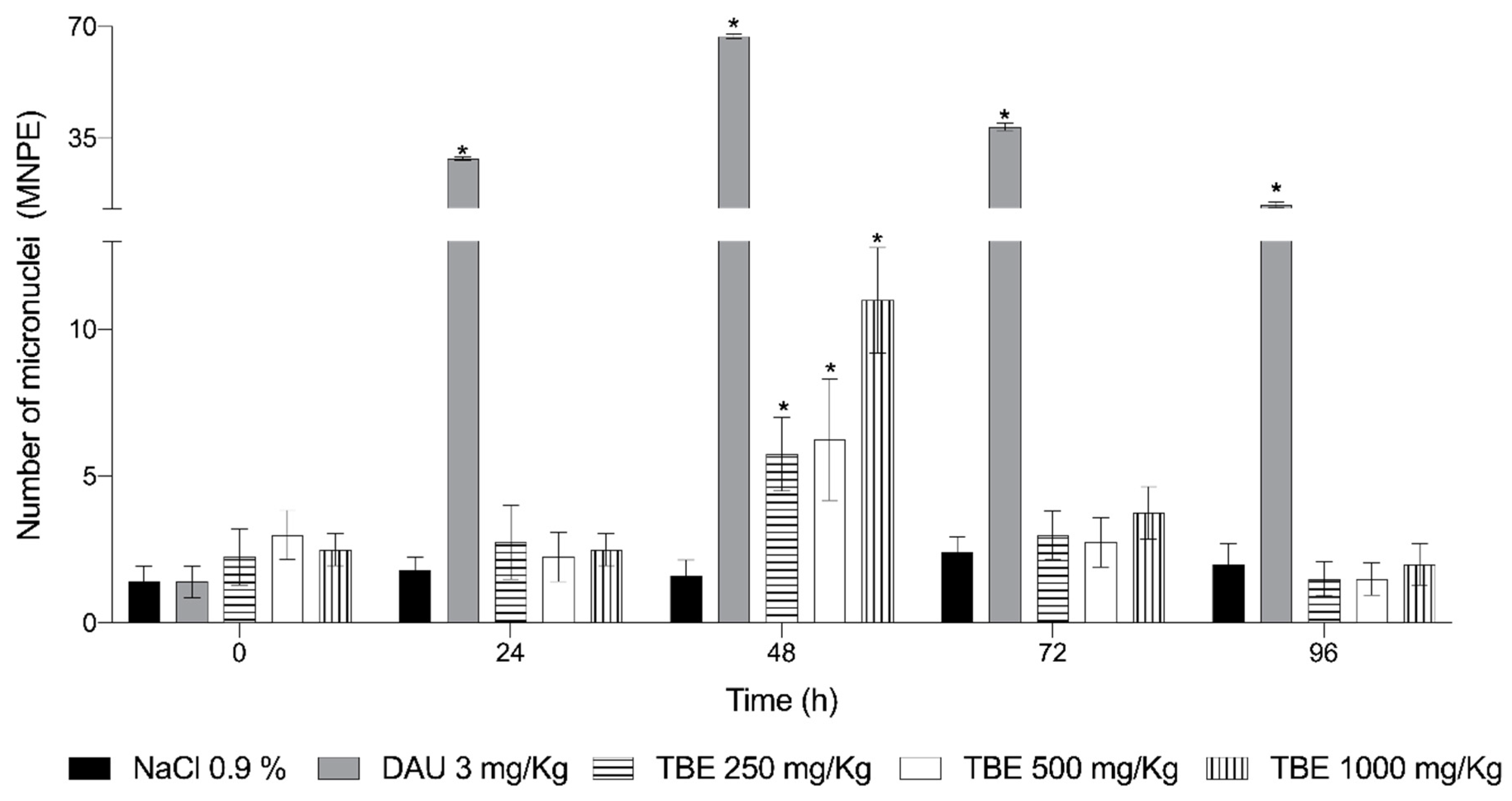
Figure 3.
Cytotoxicity of the protein extract from Phaseolus acutifolius. Relationship between the number of polychromatic erythrocytes and the number of normochromatic erythrocytes (PE/NE index). Values represent the mean ± standard deviation of seven mice per group. * indicates a statistically significant difference from the negative control. ANOVA and Tukey tests (p < 0.05).
Figure 3.
Cytotoxicity of the protein extract from Phaseolus acutifolius. Relationship between the number of polychromatic erythrocytes and the number of normochromatic erythrocytes (PE/NE index). Values represent the mean ± standard deviation of seven mice per group. * indicates a statistically significant difference from the negative control. ANOVA and Tukey tests (p < 0.05).
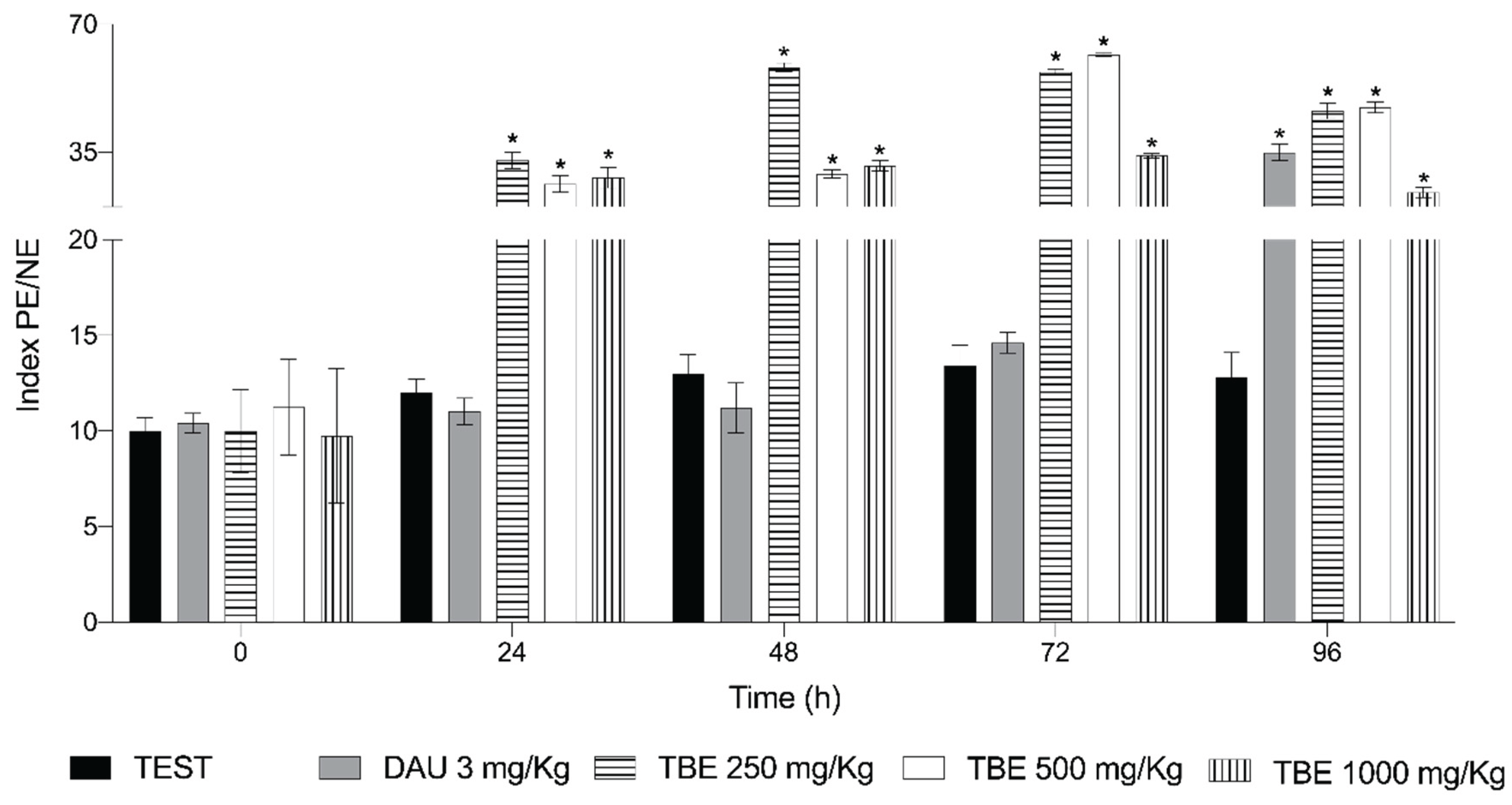

Table 1.
Antinutrient compounds present in Tepary bean extract.
| Lectins (HAU/mg) | Tripsin Inhibitors (TIU/mg) | Taninns (mg eq Cat/g) | Saponins (HU/mg) |
| 2701.85±0.0 | 6.86±0.55 | 0.1757±0.0002 | 20.16±0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



