
Submitted:
06 November 2025
Posted:
07 November 2025
You are already at the latest version
Abstract
(1) Background: Carbapenem-resistant Escherichia coli (CREC) is widespread and resistant to almost all available antimicrobial agents. This study aimed to assess the phenotypic and molecular characteristics of CREC isolated from retail meats in Hat Yai, Thailand. (2) Methods: A total of 155 retail meat samples were randomly collected. A total of 412 carbapenem-non-susceptible surrogates were screened by culturing on imipenem-containing eosin methylene blue (EMB) agar. The susceptibility to imipenem and meropenem was tested using the disk diffusion method. Carbapenemase and virulence genes in CREC isolates were detected using PCR. Phylogenetic groups and genetic relatedness of carbapenemase-positive CREC isolates were analyzed using gene markers and BOX-PCR, respectively. (3) Results: The results revealed a high prevalence of carbapenem-non-susceptible E. coli (CNSEC) surrogates in beef samples. Over 89% of the CNSEC surrogates from all meat types were identified as CREC isolates. Of these, only 4.8% of CREC isolates from beef samples were positive for the blaNDM gene, and one isolate was also positive for the blaVIM gene. These isolates carried only the fimH gene as a virulence factor. The blaNDM-positive CREC isolates were classified in phylogenetic group D. (4) Conclusions: Identifying antimicrobial-resistant pathogens, particularly CREC, in food-producing animals is critical due to the potential risks to public health.
Keywords:
carbapenem-resistant Escherichia coli
; retail meat
; carbapenemase gene
; virulence gene
1. Introduction
Carbapenem resistance in Enterobacterales has become a global health problem, as it is often associated with resistance to multiple antimicrobial agents, complicating treatment options and leading to high morbidity and mortality rates [1,2]. Carbapenem resistance occurs through various mechanisms. A primary mechanism is the production of carbapenemases, which can degrade almost all β-lactams [3]. The most predominant carbapenemase-encoding genes in Enterobacterales include blaKPC, blaVIM, blaIMP, blaNDM, and blaOXA-48 [4,5,6]. These genes represent different molecular classes that determine their unique characteristics in Enterobacterales, including E. coli. The blaKPC gene is a class A serine carbapenemase, strongly inhibited by β-lactamase inhibitors, and hydrolyzes almost all β-lactams. The blaVIM, blaIMP, and blaNDM genes are class B metallo-β-lactamases (MBLs), relying on zinc for activity, and hydrolyze all β-lactams except aztreonam, and are not inhibited by traditional β-lactamase inhibitors. Finally, the blaOXA-48 gene is a class D oxacillinase that causes resistance by weakly hydrolyzing carbapenems and often sparing expanded-spectrum cephalosporins (ESBLs), which can lead to delayed detection in clinical settings. A previous study reported that the blaKPC gene was the most frequent carbapenemase gene in carbapenem-non-susceptible Enterobacterales (CNSE) isolated from patients in US hospitals, followed by blaNDM, blaIMP, blaVIM, and blaOXA-48 [7]. Crucially, ceftazidime/avibactam was the most active drug overall, inhibiting 96.9% of CNSE isolates, including almost all KPC producers. However, the discovery of a new blaKPC variant (blaKPC-58) that causes ceftazidime/avibactam resistance signals the continued evolution of resistance, despite an overall slight decline in carbapenemase prevalence during the study period. Other mechanisms involve the overexpression of efflux pumps, the loss of porin activity, and the combination of these mechanisms with the expression of blaampC and/or the production of ESBLs [4,5,6]. Most genes encode these mechanisms through the plasmids or other mobile genetic elements (MGEs) such as transposons and integrons, which can be transferred to other cells or bacterial species [8]. Horizontal gene transfer (HGT) is one of the important factors causing the spread of antimicrobial resistance (AMR) in bacteria, especially Gram-negative pathogens.
Besides the AMR, numerous virulence determinants of E. coli further complicate its management. Generally, E. coli can be categorized into diarrheagenic E. coli (DEC) and extraintestinal pathogenic E. coli (ExPEC), each group harboring distinct sets of virulence-associated genes. In the 6 categories of DEC, the major virulence determinants, namely stx1/stx2, bfpA, eae, aggR, ipaH, elt/est, and daaE, encode Shiga toxin 1/Shiga toxin 2, bundle-forming pili, intimin, the transcriptional activator AAF/I, enteroinvasive protein, heat-labile/heat-stable enterotoxins, and F1845 fimbriae, respectively [9]. These genes play crucial roles in mediating bacterial adhesion, invasion, and subsequent destruction of host epithelial cells. Furthermore, the ExPEC-associated indicator genes, namely papA, papC, sfaDE, afa, kpsMTII, and iutA, which encode the P fimbrial structural subunit, outer membrane usher protein, S fimbriae, Afa adhesin, capsular antigen, and the siderophore aerobactin, respectively, also play important roles in the pathogenesis of extraintestinal infections [10]. Additionally, fimH, cnf1, lpf, and hlyA, which encode the type I fimbrial tip, cytotoxic necrotizing factor 1, long polar fimbriae, and α-hemolysin, respectively, can further exacerbate bacterial virulence, leading to more severe disease outcomes. The presence of these virulence genes collectively enhances the pathogenic potential of E. coli, emphasizing its remarkable clinical significance.
Carbapenem-resistant Enterobacterales (CRE) were considered an urgent threat by the Centers for Disease Control and Prevention (CDC) [11]. In 2019, the CDC reported an estimated 13,100 cases of CRE infections in hospitalized patients and 1,100 related deaths in the U.S. A high estimated healthcare cost was also associated with these infections [11]. Carbapenem-resistant Klebsiella pneumoniae (CRKP) and carbapenem-resistant Escherichia coli (CREC) are the most frequent pathogens among the CRE [11]. Several studies have examined the prevalence of CRE in Thailand. National antimicrobial resistance surveillance, Thailand (NARST) reported that the resistance rates of imipenem, meropenem, and ertapenem in CREC during 2000 – 2022 were 1% to 5.3%, 0.6% to 5.3%, and 0.6% to 6.3%, respectively. A study conducted at Siriraj Hospital, one of Thailand's largest hospitals, reported a 2-year surveillance (2009 – 2011) of CRE clinical isolates. Of 12,741 non-duplicated Enterobacteriaceae isolates, 181 isolates (1.4%) were classified as CRE, with the 5 identified species, including Enterobacter cloacae, K. pneumoniae, E. coli, and Citrobacter freundii [4]. In addition to the clinical setting, CRE isolates identified in foods, animals, and environments are also of concern. Although carbapenems are not typically used in livestock, ESBL-producing CRE are common in veterinary settings [12]. The global antimicrobial use in food-producing animals is expected to reach 174,549 tons by 2030, with intensive use creating selective pressure that drives AMR [12]. This resistance can spread through the food chain and the environment, as livestock may acquire it from contaminated water or biological vectors [12]. Additionally, antibiotics can alter gut microbiota, increasing the diversity of AMR genes [12]. According to previous evidence, CRE, especially CREC, can transfer from animals to humans via food and the environment [12]. Several investigations have demonstrated a positive correlation between livestock production systems and the occurrence of CREC infections in humans. One study demonstrated the evidence of direct transmission of New Delhi metallo-β-lactamases-type carbapenem-resistant E. coli (NDM-EC) between humans and animals [13], while zooanthroponotic transmission of CREC from humans to companion animals has also been documented [14]. The presence of carbapenem-non-susceptible E. coli (CNSEC), which includes isolates exhibiting intermediate resistance or resistance to carbapenem, in animal-sourced foods is a significant concern for food safety and public health [15,16]. Thus, this study aimed to investigate the phenotypic and molecular characteristics of CREC in retail meats from an open market in Hat Yai, Thailand.
2. Materials and Methods
2.1. Sample Collection and CNSEC Screening
A total of 155 raw meat samples (beef = 53, pork = 51, and chicken =51) were collected from an open market in Hat Yai, Songkhla, Thailand, and processed following the protocol described in a previous study [17]. Briefly, 10 g of meat was soaked in 90 ml of tryptic soy broth (TSB), then the solution was mixed and incubated at 37 ℃ for 6 h. Afterwards, the solution was serially diluted 10-fold and subsequently spread on eosin methylene blue (EMB) agar supplemented with imipenem (final concentration of 2 µg/ml) to screen for CNSEC [18]. The plate was then incubated at 37℃ for 18 h. One to fifteen green metallic sheen colonies were selected to increase the probability of detecting the CNSEC surrogates and kept at -80℃ in 10% glycerol (final concentration) [19].
2.2. Antimicrobial Susceptibility Testing
Antimicrobial susceptibility in the CNSEC surrogates was examined using a disk diffusion test (Kirby-Bauer Method). Briefly, a single colony was picked, inoculated in 3 ml of Mueller-Hinton broth (MHB), and incubated at 37℃ for 3 h with shaking at 150 rpm. Subsequently, bacterial culture was adjusted to 0.5 McFarland standards with sterile normal saline solution (approximately 1.5 × 108 colony-forming units (CFU) per ml). The bacterial solution was swabbed onto the surface of Mueller-Hinton agar (MHA). Antimicrobial disks, including ceftazidime (CAZ, 30 µg), cefotaxime (CTX, 30 µg), meropenem (MEM, 10 µg), and imipenem (IPM, 10 µg), were placed on the surface of MHA and incubated at 37℃ for 18 h. E. coli ATCC 25922 was used as a quality control. The diameters of inhibition zones were measured by a vernier caliper and interpreted according to the clinical breakpoints, according to CLSI guidelines [20].
The overall proportion of CNSEC surrogates was determined by dividing the number of isolates that exhibited a non-susceptible phenotype (intermediate or resistant) to at least one tested carbapenem (imipenem or meropenem) by the total number of E. coli colonies tested from each source. The proportion was calculated using the following formula:
where:
- A: Number of isolates confirmed as CNSEC (intermediate or resistant to any carbapenem)
- B: Total number of E. coli isolates tested from that source
2.3. DNA Extraction
The bacterial DNA of each CNSEC surrogate was extracted using the boiling method [17]. Briefly, a single colony was inoculated into 1 ml of TSB and incubated at 37℃ for 3 h with shaking at 150 rpm. The bacterial culture was boiled at 100℃ for 10 min and immediately cooled on ice for 5 min. Then, it was centrifuged at 11,000 x g for 5 min. A total of 10 µl of supernatant was collected and transferred into a sterile 1.5-ml microcentrifuge tube containing 90 µl of sterile deionized water. The DNA solution of each isolate was used as a DNA template for further experiments. The eubacterial Universal primer targeting the 16S rRNA gene was employed as an internal control to assess the integrity and adequacy of the extracted genomic DNA for subsequent PCR analyses.
2.4. Confirmation of E. coli Strain
The carriage of the uidA gene, specific for the identification of E. coli, was investigated in all CNSEC surrogates using the polymerase chain reaction (PCR) method. The component of the PCR reaction mixture contained 1X of GoTaq Flexi green buffer, 0.1 mM of deoxynucleotide triphosphates (dNTPs), 3.0 mM of MgCl2, 0.4 µM of forward and reverse primers, 0.5 unit of GoTaq DNA polymerase (Promega, USA), and 2 µl of DNA template. A pair of uidA primers and the PCR conditions are shown in Tables S1 and S2, respectively. The uidA amplification was performed in a T100TM Thermal Cycler (Bio-Rad, Hercules, CA, USA). Afterwards, the amplicons were analyzed using 1% agarose gel electrophoresis under 100V for 40 min. The gel was stained with ethidium bromide and visualized under the WSE-5200 Printpraph 2M gel imaging system (ATTO Corp., Tokyo, Japan).
2.5. Detection of Carbapenemase Genes in CNSEC Isolates
To understand the primary mechanisms of carbapenem resistance in CNSEC isolates, the most common carbapenemase genes (blaKPC, blaIMP, blaVIM, blaNDM, and blaOXA-48) were investigated using the uniplex-PCR method. The uniplex-PCR reaction mixture was prepared, the genes were amplified, and the results were observed as described above. Tables S1 and S2 illustrate all pairs of primers and their PCR conditions. The representative PCR amplicons of the positive genes were further sequenced using Sanger sequencing. Afterwards, the sequences were searched against the National Center for Biotechnology Information (NCBI) database using blastn.
2.6. Detection of Virulence Genes in CNSEC Isolates
The virulence factors contributing to the pathogenicity of E. coli were assessed. In the classification of diarrheagenic E. coli (DEC), the indicator genes were identified using the PCR method. The genes were grouped into 6 pathogenic categories: category 1, stx and eae for enterohemorrhagic E. coli (EHEC); category 2, bfp and eae for typical enteropathogenic E. coli (tEPEC) or eae alone for atypical enteropathogenic E. coli (aEPEC); category 3, elt or est for enterotoxigenic E. coli (ETEC); category 4, aggR for enteroaggregative E. coli (EAEC); category 5, ipaH for enteroinvasive E. coli (EIEC); and category 6, daaE for diffusely adherent E. coli (DAEC). Meanwhile, the extraintestinal pathogenic E. coli (ExPEC) indicator genes were also investigated. Six target genes were categorized into Group 1 (papA and/or papC), Group 2 (sfaD), Group 3 (afa), Group 4 (kpsMTII), and Group 5 (iutA). For the interpretation, the CNSEC isolates with positive results for two or more groups were considered as ExPEC [21]. Furthermore, the other 6 virulence genes, astA, agn43, cnf1, hly, fimH, and lpf, were also examined. The preparation of the uniplex-PCR reaction mixture, amplification of all 20 virulence genes, and observation of the results were conducted as described above. All pairs of primers and their PCR conditions are illustrated in Tables S1 and S2, respectively.
2.7. Determination of Phylogenetic Groups
The phylogenetic groups of carbapenemase-positive CREC isolates were determined using the PCR method targeting 3 genetic markers: chuA, yjaA, and an anonymous DNA fragment known as TSPE4.C2. The PCR reaction mixture was prepared, the genes were amplified, and the results were observed as described above. Tables S1 and S2 illustrate all pairs of primers and their PCR conditions. PCR products were analyzed as described above. Bacterial phylogenetic group classification was interpreted following Clermont et al. [22], as shown in Figure S1.
2.8. Analysis of Genetic Relatedness
In addition to classifying phylogenetic groups, the DNA fingerprints of carbapenemase-positive CREC isolates were also analyzed using BOX-PCR [23]. The genomic DNA (gDNA) of all carbapenemase-positive CREC isolates was extracted using the miniprep spin column method (Geneaid, Taipei, Taiwan). The PCR reaction mixture contained 0.2 µM of BOXA1R primer (5’-CTACGGCAAGGCGACGCTGACG-3’), 3.0 mM of MgCl2, 1X GoTaq Flexi green buffer, 1.25 units of GoTaq DNA polymerase, 0.2 mM of dNTPs, and a DNA template [24]. Amplification conditions included an initial denaturation step at 95℃ for 3 min; followed by 35 cycles of denaturation at 94℃ for 30 seconds, annealing at 50℃ for 1 min, and extension at 65℃ for 8 min; and final extension at 72℃ for 5 min. Afterwards, the amplicons were observed in a 1.5% agarose gel electrophoresis under 80 V for 60 min. The gel was stained and visualized as described above.
3. Results
3.1. Prevalence of CNSEC Surrogates
The prevalence of suspected CNSEC was illustrated in Table 1. Of 155 samples, suspected CNSEC was found in beef, pork, and chicken at 56.6% (30/53), 58.8% (30/51), and 39.2% (20/51), respectively. Among these positive samples, 185, 139, and 88 suspected CNSEC isolates were obtained from beef, pork, and chicken, respectively.
3.2. Antimicrobial Resistance Profiles
Antimicrobial susceptibility profile of suspected CNSEC isolates was shown in Table 2 and Table S3. In the susceptibility to third-generation cephalosporins, it was found that 11.4%, 2.9%, and 5.7% of suspected CNSEC from beef, pork, and chicken, respectively, were resistant to cefotaxime, and 9.2% and 2.9% from beef and pork were resistant to ceftazidime. A majority of these suspected CNSEC isolates were still susceptible to cefotaxime and ceftazidime. For carbapenem susceptibility, 7.0%, 11.5%, and 12.5% of suspected CNSEC isolates from beef, pork, and chicken were intermediate-resistant to imipenem, and 23.8%, 41.0%, and 38.6% were intermediate-resistant to meropenem. Moreover, 63.2%, 86.3%, and 83.0% of the strains from beef, pork, and chicken were resistant to imipenem, and 30.8%, 36.7%, and 38.6% were resistant to meropenem. The CNSEC isolates showing resistance to at least one of the third-generation cephalosporins (cefotaxime and ceftazidime) were considered to presumptively produce ESBLs [25]. This classification was based on the fact that this phenotypic resistance profile serves as a screening marker, although we acknowledge that non-susceptibility mediated by other mechanisms (e.g., AmpC overexpression, porin loss, or efflux pump activation) may lead to false-positive presumptions. Furthermore, the suspected CNSEC isolates showing intermediate resistance or resistance to at least one carbapenem were phenotypically confirmed as CNSEC strains, and those susceptible to both imipenem and meropenem were classified as carbapenem-susceptible E. coli (CSEC). The total numbers of isolates from beef, pork, and chicken were 185, 139, and 88, respectively. Among all sample sources, the proportion of CNSEC strains (defined as non-susceptible to at least one carbapenem) was mostly found in pork (95.0%), followed by chicken (89.8%) and beef (59.5%), while 10.8%, 5.7%, and 4.3% from beef, chicken, and pork were classified as both CNSEC and ESBL-PEC isolates (Figure 1). More importantly, 90.0%, 89.6%, and 89.1% of the suspected CNSEC isolates from beef, pork, and chicken were resistant to at least one carbapenem, subsequently identified as carbapenem-resistant E. coli (CREC) (Table S3). Nevertheless, although the suspected CNSEC isolates were screened on the EMB containing imipenem, some suspected CSEC and ESBL-PEC isolates were still observed in this study (Figure 1).
3.3. Carbapenemase Genes in CREC Isolates
Primary mechanisms of carbapenem resistance were investigated in all CREC isolates (n = 284). The CREC isolates were specifically defined as the subset of CNSEC isolates that showed a definitive resistant phenotype to at least one tested carbapenem (imipenem or meropenem) across all sample sources, ensuring we focused only on those categorized as truly carbapenem-resistant for the mechanism analysis. The results demonstrated that, among 5 carbapenemase genes, the blaNDM gene was found in 15 (4.8%) CREC isolates from one beef sample (B14) (Table 3). The blaVIM gene was also detected in one blaNDM-positive CREC isolate. These 15 CREC isolates were examined for further experiments.
3.4. Molecular Relationship in blaNDM-Harboring CREC Isolates
The molecular relationship among all blaNDM-harboring CREC isolates was analyzed by detecting the genetic markers. The classification of phylogenetic groups revealed that all isolates were positive for the chuA gene and the TSPE4.C2 fragment, but negative for the yjaA gene (Table 4). Based on these genetic markers, all isolates were classified as phylogenetic group D. Furthermore, the genetic relatedness among these isolates was also analyzed using the BOX-PCR method. All isolates exhibited a similar pattern of DNA fragments (Figure S2).
4. Discussion
The contamination of pathogenic bacteria, especially antibiotic-resistant strains, in meat is a critical public health concern due to the potential for these bacteria to cause severe infections in humans [26]. With the HGT, bacteria isolated from food sources may share their AMR genes with antibiotic-resistant pathogens in humans [27]. This issue is exacerbated by the overuse of antibiotics in livestock for growth promotion and disease prevention, contributing to the development of resistance [28]. Furthermore, these pathogens in meat pose a food safety threat and have global implications, as these bacteria can spread across borders through trade and travel, worsening the global crisis of AMR [29]. Addressing this problem requires the responsible use of antibiotics in agriculture, improved surveillance of antibiotic-resistant bacteria in the food supply, and promotion of safe food handling practices. In this study, we report the prevalence of CREC in raw meats from an open market in Hat Yai, Songkhla, Thailand, and describe their phenotypic and molecular characteristics, including antimicrobial resistance profiles, carbapenemase genes, virulence genes, and phylogenetic relationships.
Our findings demonstrated that CNSEC surrogates were mostly found in beef samples, followed by pork and chicken samples. The highest CNSEC isolates from beef might be explained by specific antimicrobial usage practices in local cattle production that co-select for carbapenem resistance genes (often carried on highly mobile plasmids), or by unique contamination risks during beef processing, especially for ground beef, which disseminates intestinal flora more thoroughly [30]. A very high proportion of CNSEC surrogates were subclassified as CREC isolates. The predominance of CREC among the CNSEC isolates highlights a significant public-health concern. CNSEC refers to all isolates showing reduced carbapenem susceptibility, whereas CREC denotes confirmed resistance mechanisms and thus carries greater clinical and epidemiological relevance [31]. The high prevalence of CREC in beef and other meat types suggests the food chain as a potential reservoir for multidrug-resistant E. coli, consistent with global reports linking livestock production to the dissemination of carbapenem and broad-spectrum β-lactam resistance [32]. The antimicrobial susceptibility testing results showing resistance to imipenem and/or meropenem indicate a critical loss of efficacy for these last-line agents. The concurrent resistance to third-generation cephalosporins may support the likelihood of non-carbapenemase-mediated mechanisms [33]. Although CNSEC may include isolates with borderline susceptibility or transient adaptive mechanisms, CREC represents stable, transferable resistance with higher public-health significance. The detection of CREC in retail meat poses a risk of HGT to human commensal E. coli, facilitating the emergence of difficult-to-treat infections (Feng et al., 2021) [34]. These findings probably support the need for integrated One-Health surveillance and stricter antimicrobial stewardship across the food-production system.
In the identification of primary carbapenem resistance mechanisms, only 5.3% of the CREC isolates were positive for the blaNDM gene, while one of them was also positive for the blaVIM gene. While the detection of these MBL genes confirmed the basis of resistance in the CREC isolates from a single beef sample, the precise mechanisms driving the carbapenem resistance in the remaining isolates were not determined. The observed carbapenem resistance phenotype probably results from several factors, including, but not limited to, the co-existence of high-level AmpC and/or ESBL production combined with outer membrane porin loss and/or the enhanced activity of efflux pumps [35,36]. Future studies utilizing molecular sequencing or gene expression assays are required to definitively characterize the carbapenem resistance mechanisms in these isolates [35,36]. Several studies have reported the prevalence of this pathogen in food-producing animals. In 2018, Guo et al. screened CREC isolates from 125 duck meat samples obtained from 5 farms in China (25 meat samples per farm) [37]. They reported that 33.6% of the isolates carried the blaNDM gene with high resistance to almost all tested antimicrobial agents. Notably, their findings revealed that the structures of blaNDM-bearing plasmids were highly similar to the plasmids from human-sourced isolates. The phylogenetic tree also showed that duck-sourced isolates were clustered with human-sourced isolates from different areas in China. The study by Zhai et al. (2020) from China revealed that 14 sequence types (STs) were identified in NDM-producing E. coli isolates from chicken and their environmental samples, with ST6751 being the most frequent [38]. Besides the poultry isolates, VIM-1-producing E. coli ST88 isolated from pigs was first reported in Germany in 2011 and was also previously identified in chickens, cattle, and humans [39]. Another study from Italy showed that E. coli isolates from pigs carried the blaOXA-181, blaOXA-48, mcr-1 (mobilized colistin resistance gene), and qnrS1 (fluoroquinolone resistance gene) [40]. The findings also demonstrated that the blaOXA-181 and qnrS1 genes were detected on the IncX3 plasmid, highly similar to the plasmid from human-sourced isolates. For the study of CREC in cattle, blaOXA-48-, blaOXA-181-, blaNDM-, or blaVIM-carrying E. coli isolates were reported in Egypt in 2014, South Africa in 2020, Italy in 2021, and Spain in 2022 [30,41,42,43]. Tello et al. (2022) reported that the blaNDM-1 gene was located on the IncC plasmid that harbored aminoglycoside, sulphonamide, and trimethoprim resistance genes [43]. The evidence probably confirms the dissemination of carbapenem resistance between human- and animal-sourced isolates. However, our findings showed the presence of carbapenemase genes, blaNDM (n = 15) and blaVIM (n = 1), in the isolates from only one beef sample. The reason for a relatively low percentage of the carbapenemase genes in the CREC isolates from raw meats might be factors like geographic variability, controlled antibiotic use in agriculture, and alternative resistance mechanisms. In regions where carbapenemase production is not predominant, porin loss and efflux pump overexpression, alongside the production of ESBLs and AmpC β-lactamases, might be the frequently found mechanisms, leading to carbapenem resistance without the need for carbapenemase production in our CREC isolates [44]. We hypothesized that these mechanisms are common in hospital settings, where ESBL-producing E. coli is prevalent. Further investigation of other mechanisms would confirm this hypothesis, which was acknowledged as a limitation of the present study.
In the detection of virulence genes, the findings revealed that all blaNDM-harboring CREC isolates were positive for the fimH gene responsible for the production of type 1 fimbriae, which are adhesive structures important for attachment to host tissues, particularly in the urinary tract [45]. However, the presence of fimH alone does not classify the strain as particularly pathogenic or indicative of a specific pathotype. It is present in both pathogenic and commensal strains of E. coli [46]. In pathogenic E. coli, such as uropathogenic E. coli (UPEC), fimH plays a role in urinary tract infections (UTIs) [47]. However, if fimH is the only virulence factor presented in the studied isolates, it may not necessarily indicate high virulence.
Phylogenetic analysis exhibited that all blaNDM-harboring CREC isolates were classified in phylogenetic group D. This group is often associated with ExPEC, including strains involved in urinary tract infections and other invasive infections [48]. However, these isolates were common E. coli since they were negative for ExPEC indicator genes. In addition, the investigation of genetic relatedness revealed that all blaNDM-harboring CREC isolates were clonally related. However, further high-resolution genetic testing, such as whole-genome sequencing, may be required for confirmation at a finer scale.
Zoonotic transmission of antibiotic-resistant bacteria, particularly CREC, from animal-derived foods poses a significant public health concern. Meats from animals treated with antibiotics or exposed to resistant bacteria can act as a reservoir, infecting humans via handling or consuming undercooked meat. The presence of such bacteria in the food chain exposes weaknesses in food safety, agricultural practices, and antimicrobial stewardship, underscoring the urgent need for stricter regulations on antibiotic use in food animals, robust surveillance systems, and improved control measures to combat the growing antimicrobial resistance crisis affecting both human and veterinary medicine.
5. Conclusions
The study reported the prevalence and characteristics of CREC isolates detected in raw meats from an open market, Hat Yai, Songkhla, Thailand. The number of CREC isolates was relatively high, especially in beef samples, while a minority of them carried carbapenemase genes. The presence of CREC, particularly carbapenemase-positive isolates, in meats poses a significant threat to public health, food safety, and efforts to combat antimicrobial resistance. This underscores the urgent need for effective antimicrobial stewardship in agriculture, stringent food safety protocols, and comprehensive surveillance systems to monitor and curb the spread of these harmful bacteria along the food supply chain, from farm to fork.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Dichotomous decision tree to determine the phylogenetic group of Escherichia coli strains.; Figure S2: Genetic relatedness among 16 carbapenem-resistant Escherichia coli isolates, with at least one carbapenemase gene carriage. Lane M, 2-log DNA markers; lane P, positive control; lane 1, B14.1; lane 2, B14.2; lane 3, B14.3; lane 4, B14.4; lane 5, B14.5; lane 6, B14.6; lane 7, B14.7; lane 8, B14.8; lane 9, B14.9; lane 10, B14.10; lane 11, B14.11; lane 12, B14.12; lane 13, B14.13; lane 14, B14.14; lane 15, B14.15; Table S1: PCR primers for all detected genes in carbapenem-resistant Escherichia coli surrogates.; Table S2: PCR conditions for all detected genes in carbapenem-resistant Escherichia coli surrogates.; Table S3: Antimicrobial susceptibility patterns in carbapenem-non-susceptible Escherichia coli surrogates.
Author Contributions
Conceptualization, P.S.; methodology, P.S., A.C., W.M., P.R., and R.P.; validation, P.S. and A.C.; formal analysis, P.S. and A.C.; investigation, P.S., A.C., and W.M.; writing—original draft preparation, A.C.; writing—review and editing, P.S.; supervision, P.S.; project administration, P.S.; funding acquisition, P.S. All authors have read and agreed to the published final version of the manuscript.
Funding
This research was funded by Prince of Songkla University, grant number SCI600588S.
Data Availability Statement
The representative sequences of the detected blaNDM and blaVIM genes have been deposited to NCBI BankIt and are available under the accession numbers PX489564 and PX489565, respectively.
Acknowledgments
We thank the Division of Biological Science, Faculty of Science, Prince of Songkla University, for providing the essential facilities.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Marino, A.; Maniaci, A.; Lentini, M.; Ronsivalle, S.; Nunnari, G.; Cocuzza, S.; Parisi, F.M.; Cacopardo, B.; Lavalle, S.; La Via, L. The Global Burden of Multidrug-Resistant Bacteria. Epidemiologia 2025, 6(2), 21. [CrossRef]
- Zankere, T.; Lechiile, K.; Mokgwathi, K.; Tlhako, N.; Moorad, B.; Ntereke, T.D.; Gatonye, T.; Lautenbach, E.; Richard-Greenblatt, M.; Mokomane, M.; Mosepele, M.; Cancedda, C.; Goldfarb, D.M.; Styczynski, A.; Parra, G.; Smith, R.M.; Mannathoko, N.; Strysko, J. Admission screening for extended-spectrum cephalosporin-resistant and carbapenem-resistant Enterobacterales colonization at a referral hospital in Botswana: A one-year period-prevalence survey, 2022–2023. PLOS Global Public Health 2025, 5(10), e0005018. [CrossRef]
- Suay-García, B.; Pérez-Gracia, M.T. Present and future of carbapenem-resistant Enterobacteriaceae infections. Advances in clinical immunology, medical microbiology, COVID-19, and big data 2021, 435-456. [CrossRef]
- Netikul, T.; Kiratisin, P. Genetic characterization of carbapenem-resistant enterobacteriaceae and the spread of carbapenem-resistant klebsiella pneumonia ST340 at a university hospital in Thailand. PloS one 2015, 10(9), e0139116. [CrossRef]
- Su, Y.; Zou, G.; Huang, X.; Bi, J.; Meng, L.; Zhao, W.; Li, T. Molecular characterization and resistance mechanisms of ertapenem-non-susceptible carbapenem-resistant Klebsiella pneumoniae co-harboring ESBLs or AmpC enzymes with porin loss or efflux pump overexpression. J. Bacteriol. 2025, e00148-25. [CrossRef]
- Logan, L.K.; Weinstein, R.A. The epidemiology of carbapenem-resistant Enterobacteriaceae: the impact and evolution of a global menace. The Journal of infectious diseases 2017, 215(suppl_1), S28-S36. [CrossRef]
- Castanheira, M.; Deshpande, L.M.; Mendes, R.E.; Doyle, T.B.; Sader, H.S. Prevalence of carbapenemase genes among carbapenem-nonsusceptible Enterobacterales collected in US hospitals in a five-year period and activity of ceftazidime/avibactam and comparator agents. JAC-antimicrobial resistance 2022, 4(5), dlac098. [CrossRef]
- Hammoudi Halat, D.; Ayoub Moubareck, C. The current burden of carbapenemases: Review of significant properties and dissemination among gram-negative bacteria. Antibiotics 2020, 9(4), 186. [CrossRef]
- Nataro, J.P.; Kaper, J.B. Diarrheagenic escherichia coli. Clin. Microbiol. Rev. 1998, 11(1), 142-201.
- Johnson, J.R.; Murray, A.C.; Gajewski, A.; Sullivan, M.; Snippes, P.; Kuskowski, M.A.; Smith, K.E. Isolation and molecular characterization of nalidixic acid-resistant extraintestinal pathogenic Escherichia coli from retail chicken products. Antimicrobial agents and chemotherapy 2003, 47(7), 2161-2168. [CrossRef]
- CDC. Antibiotic Resistance Threats in the United States, 2019 (2019 AR Threats Report), Centers for Disease Control and Prevention (CDC), Atlanta, GA. 2019 [Accessed 04 January 2020]; Available from: https://www.cdc.gov/drugresistance/Biggest-Threats.html.
- Ramírez-Castillo, F.Y.; Guerrero-Barrera, A.L.; Avelar-González, F.J. An overview of carbapenem-resistant organisms from food-producing animals, seafood, aquaculture, companion animals, and wildlife. Frontiers in Veterinary Science 2023, 10(1158588. [CrossRef]
- Li, J.; Bi, Z.; Ma, S.; Chen, B.; Cai, C.; He, J.; Schwarz, S.; Sun, C.; Zhou, Y.; Yin, J. Inter-host transmission of carbapenemase-producing Escherichia coli among humans and backyard animals. Environ. Health Perspect. 2019, 127(10), 107009. [CrossRef]
- Grönthal, T.; Österblad, M.; Eklund, M.; Jalava, J.; Nykäsenoja, S.; Pekkanen, K.; Rantala, M. Sharing more than friendship–transmission of NDM-5 ST167 and CTX-M-9 ST69 Escherichia coli between dogs and humans in a family, Finland, 2015. Eurosurveillance 2018, 23(27), 1700497. [CrossRef]
- Huang, Y.-S.; Tsai, W.-C.; Li, J.-J.; Chen, P.-Y.; Wang, J.-T.; Chen, Y.-T.; Chen, F.-J.; Lauderdale, T.-L.; Chang, S.-C. Increasing New Delhi metallo-β-lactamase-positive Escherichia coli among carbapenem non-susceptible Enterobacteriaceae in Taiwan during 2016 to 2018. Scientific reports 2021, 11(1), 2609. [CrossRef]
- ECDC. Expert consensus protocol on carbapenem resistance detection and characterisation for the survey of carbapenem- and/or colistin-resistant Enterobacteriaceae – Version 3.0. Stockholm: European Centre for Disease Prevention and Control (ECDC), 2019. 2019 [30 October 2025]; Available from: https://www.ecdc.europa.eu/sites/default/files/documents/expert-consensus-protocol-carbapenem-resistance.pdf.
- Sirikaew, S.; Rattanachuay, P.; Nakaguchi, Y.; Sukhumungoon, P. Immuno-magnetic isolation, characterization and genetic relationship of Escherichia coli O26 from raw meats, Hat Yai City, Songkhla, Thailand. Southeast Asian Journal of Tropical Medicine and Public Health 2015, 46(2), 241.
- Adler, A.; Navon-Venezia, S.; Moran-Gilad, J.; Marcos, E.; Schwartz, D.; Carmeli, Y. Laboratory and clinical evaluation of screening agar plates for detection of carbapenem-resistant Enterobacteriaceae from surveillance rectal swabs. J. Clin. Microbiol. 2011, 49(6), 2239-2242. [CrossRef]
- Mosavie, M.; Blandy, O.; Jauneikaite, E.; Caldas, I.; Ellington, M.J.; Woodford, N.; Sriskandan, S. Sampling and diversity of Escherichia coli from the enteric microbiota in patients with Escherichia coli bacteraemia. BMC Research Notes 2019, 12(1), 335. [CrossRef]
- CLSI. Clinical and Laboratory Standards Institute (CLSI). Performance standards for antimicrobial susceptibility testing; Twenty-Sixth edition informational Supplement (M100S). CLSI, Wayne, PA. 2016. 2016,.
- Johnson, J.R.; Gajewski, A.; Lesse, A.J.; Russo, T.A. Extraintestinal pathogenic Escherichia coli as a cause of invasive nonurinary infections. J. Clin. Microbiol. 2003, 41(12), 5798-5802. [CrossRef]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66(10), 4555-4558. [CrossRef]
- Sukhumungoon, P.; Sae-lim, A.; Hayeebilan, F.; Rattanachuay, P. Prevalence, Virulence Profiles, and Genetic Relatedness of Escherichia coli O45 from Raw meats, Southern Thailand. Journal of Scientific and Technological Reports Online ISSN: 2773-8752 2023, 27(2), 58-71. [CrossRef]
- Versalovic, J. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Methods Mol. Cell Biol. 1994, 5(25-40.
- Tamma, P.D.; Aitken, S.L.; Bonomo, R.A.; Mathers, A.J.; Van Duin, D.; Clancy, C.J. Infectious Diseases Society of America 2022 guidance on the treatment of extended-spectrum β-lactamase producing Enterobacterales (ESBL-E), carbapenem-resistant Enterobacterales (CRE), and Pseudomonas aeruginosa with difficult-to-treat resistance (DTR-P. aeruginosa). Clin. Infect. Dis. 2022, 75(2), 187-212.
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. The lancet 2022, 399(10325), 629-655. [CrossRef]
- Rossi, F.; Rizzotti, L.; Felis, G.E.; Torriani, S. Horizontal gene transfer among microorganisms in food: current knowledge and future perspectives. Food Microbiol. 2014, 42(232-243. [CrossRef]
- Spellberg, B.; Hansen, G.R.; Kar, A.; Cordova, C.D.; Price, L.B.; Johnson, J.R. Antibiotic resistance in humans and animals. NAM Perspectives 2016,. [CrossRef]
- Authority, E.F.S.; Prevention, E.C.f.D.; Control. The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2018/2019. Efsa Journal 2021, 19(4), e06490.
- Tshitshi, L.; Manganyi, M.C.; Montso, P.K.; Mbewe, M.; Ateba, C.N. Extended spectrum beta-lactamase-resistant determinants among carbapenem-resistant Enterobacteriaceae from beef cattle in the north West Province, South Africa: a critical assessment of their possible public health implications. Antibiotics 2020, 9(11), 820. [CrossRef]
- Nasrollahian, S.; Graham, J.P.; Halaji, M. A review of the mechanisms that confer antibiotic resistance in pathotypes of E. coli. Frontiers in Cellular and Infection Microbiology 2024, 14(1387497. [CrossRef]
- Lee, K.Y.; Lavelle, K.; Huang, A.; Atwill, E.R.; Pitesky, M.; Li, X. Assessment of prevalence and diversity of antimicrobial resistant Escherichia coli from retail meats in Southern California. Antibiotics 2023, 12(4), 782. [CrossRef]
- van Boxtel, R.; Wattel, A.A.; Arenas, J.; Goessens, W.H.; Tommassen, J. Acquisition of carbapenem resistance by plasmid-encoded-AmpC-expressing Escherichia coli. Antimicrobial agents and chemotherapy 2017, 61(1), 10.1128/aac. 01413-16. [CrossRef]
- Feng, J.; Xiang, Q.; Ma, J.; Zhang, P.; Li, K.; Wu, K.; Su, M.; Li, R.; Hurley, D.; Bai, L. Characterization of carbapenem-resistant Enterobacteriaceae cultured from retail meat products, patients, and porcine excrement in China. Frontiers in Microbiology 2021, 12(743468. [CrossRef]
- Khalifa, S.M.; Abd El-Aziz, A.M.; Hassan, R.; Abdelmegeed, E.S. β-lactam resistance associated with β-lactamase production and porin alteration in clinical isolates of E. coli and K. pneumoniae. PLoS One 2021, 16(5), e0251594.
- Chang, Y.-T.; Siu, L.K.; Wang, J.-T.; Wu, T.-L.; Chen, Y.-H.; Chuang, Y.-C.; Lin, J.-C.; Lu, P.-L. Resistance mechanisms and molecular epidemiology of carbapenem-nonsusceptible Escherichia coli in Taiwan, 2012-2015. Infection and drug resistance 2019, 2113-2123. [CrossRef]
- Guo, C.-H.; Chu, M.-J.; Liu, T.; Wang, J.; Zou, M.; Liu, B.-T. High prevalence and transmission of blaNDM-positive Escherichia coli between farmed ducks and slaughtered meats: An increasing threat to food safety. Int. J. Food Microbiol. 2024, 424(110850. [CrossRef]
- Zhai, R.; Fu, B.; Shi, X.; Sun, C.; Liu, Z.; Wang, S.; Shen, Z.; Walsh, T.R.; Cai, C.; Wang, Y. Contaminated in-house environment contributes to the persistence and transmission of NDM-producing bacteria in a Chinese poultry farm. Environ. Int. 2020, 139(105715. [CrossRef]
- Fischer, J.; San José, M.; Roschanski, N.; Schmoger, S.; Baumann, B.; Irrgang, A.; Friese, A.; Roesler, U.; Helmuth, R.; Guerra, B. Spread and persistence of VIM-1 Carbapenemase-producing Enterobacteriaceae in three German swine farms in 2011 and 2012. Vet. Microbiol. 2017, 200(118-123. [CrossRef]
- Pulss, S.; Semmler, T.; Prenger-Berninghoff, E.; Bauerfeind, R.; Ewers, C. First report of an Escherichia coli strain from swine carrying an OXA-181 carbapenemase and the colistin resistance determinant MCR-1. Int. J. Antimicrob. Agents 2017, 50(2), 232-236. [CrossRef]
- Braun, S.D.; Ahmed, M.F.; El-Adawy, H.; Hotzel, H.; Engelmann, I.; Weiß, D.; Monecke, S.; Ehricht, R. Surveillance of extended-spectrum beta-lactamase-producing Escherichia coli in dairy cattle farms in the Nile Delta, Egypt. Frontiers in microbiology 2016, 7(1020. [CrossRef]
- Carfora, V.; Diaconu, E.L.; Ianzano, A.; Di Matteo, P.; Amoruso, R.; Dell'Aira, E.; Sorbara, L.; Bottoni, F.; Guarneri, F.; Campana, L. The hazard of carbapenemase (OXA-181)-producing Escherichia coli spreading in pig and veal calf holdings in Italy in the genomics era: Risk of spill over and spill back between humans and animals. Frontiers in Microbiology 2022, 13(1016895. [CrossRef]
- Tello, M.; Oporto, B.; Lavín, J.L.; Ocejo, M.; Hurtado, A. Characterization of a carbapenem-resistant Escherichia coli from dairy cattle harbouring bla NDM-1 in an IncC plasmid. J. Antimicrob. Chemother. 2022, 77(3), 843-845. [CrossRef]
- Codjoe, F.; Donkor, E. Carbapenem resistance: a review. Medical Sciences 2018, 6(1), 1.
- Connell, I.; Agace, W.; Klemm, P.; Schembri, M.; Mărild, S.; Svanborg, C. Type 1 fimbrial expression enhances Escherichia coli virulence for the urinary tract. Proceedings of the National Academy of Sciences 1996, 93(18), 9827-9832. [CrossRef]
- Sokurenko, E.V.; Chesnokova, V.; Dykhuizen, D.E.; Ofek, I.; Wu, X.-R.; Krogfelt, K.A.; Struve, C.; Schembri, M.A.; Hasty, D.L. Pathogenic adaptation of Escherichia coli by natural variation of the FimH adhesin. Proceedings of the National Academy of Sciences 1998, 95(15), 8922-8926. [CrossRef]
- Hojati, Z.; Zamanzad, B.; Hashemzadeh, M.; Molaie, R.; Gholipour, A. The FimH gene in uropathogenic Escherichia coli strains isolated from patients with urinary tract infection. Jundishapur journal of microbiology 2015, 8(2),. [CrossRef]
- Maluta, R.P.; Logue, C.M.; Casas, M.R.T.; Meng, T.; Guastalli, E.A.L.; Rojas, T.C.G.; Montelli, A.C.; Sadatsune, T.; de Carvalho Ramos, M.; Nolan, L.K. Overlapped sequence types (STs) and serogroups of avian pathogenic (APEC) and human extra-intestinal pathogenic (ExPEC) Escherichia coli isolated in Brazil. PloS one 2014, 9(8), e105016. [CrossRef]
Figure 1.
Distribution of positive isolates from beef, pork, and chicken samples. CNSEC, carbapenem-non-susceptible Escherichia coli; CSEC, carbapenem-non-susceptible Escherichia coli; ESBL-PEC, extended-spectrum β-lactamase-producing Escherichia coli.
Figure 1.
Distribution of positive isolates from beef, pork, and chicken samples. CNSEC, carbapenem-non-susceptible Escherichia coli; CSEC, carbapenem-non-susceptible Escherichia coli; ESBL-PEC, extended-spectrum β-lactamase-producing Escherichia coli.
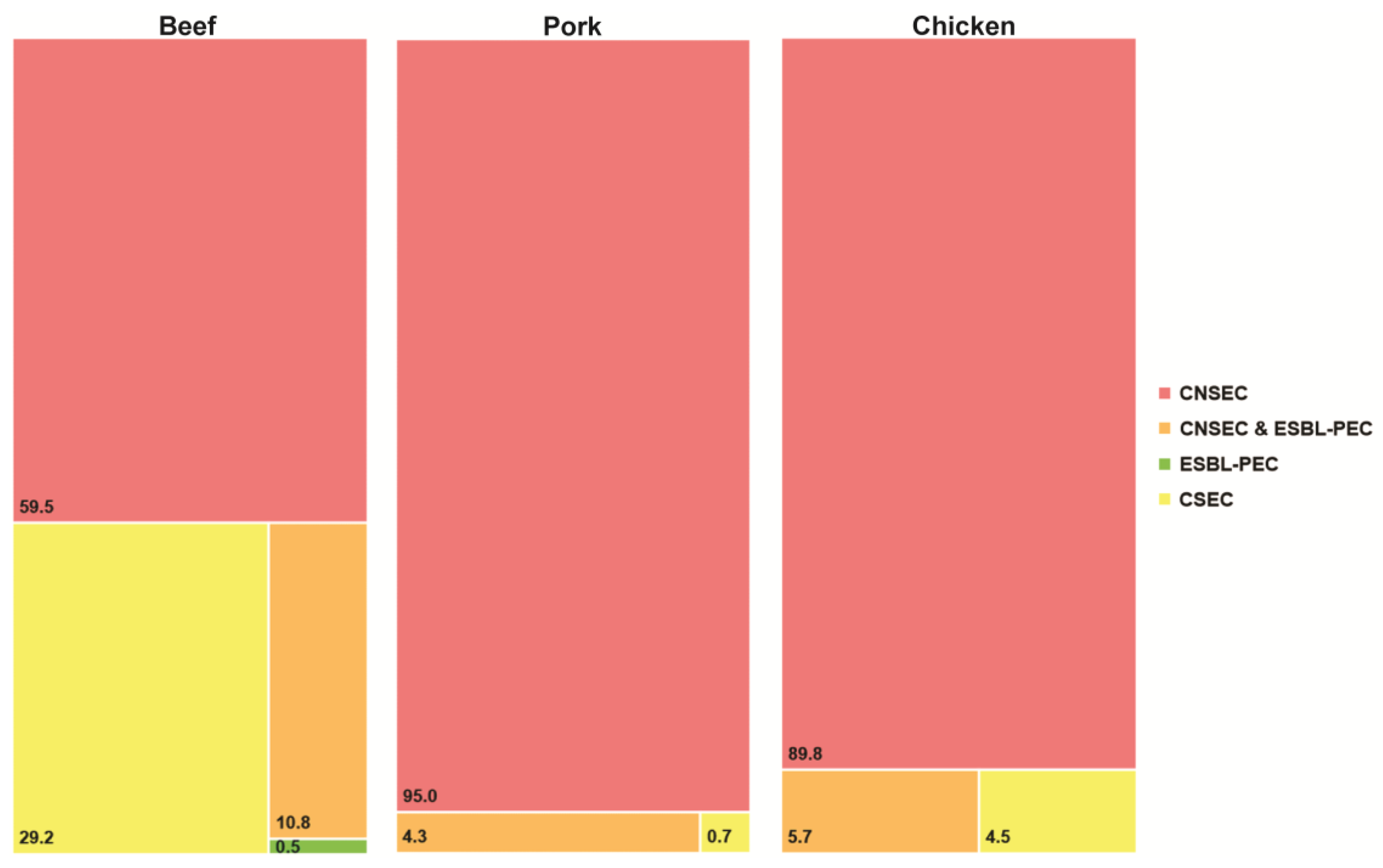

Table 1.
Prevalence of carbapenem-non-susceptible Escherichia coli surrogates in raw meats from August 2018 to May 2019.
Table 1.
Prevalence of carbapenem-non-susceptible Escherichia coli surrogates in raw meats from August 2018 to May 2019.
| Sample source | No. of positive samples/ no. of total samples (%) | No. of isolates |
|---|---|---|
| Beef | 30/53 (56.6) | 185 |
| Pork | 30/51 (58.8) | 139 |
| Chicken | 20/51 (39.2) | 88 |
| Total | 80/155 (51.6) | 412 |
Table 2.
Antimicrobial susceptibility in carbapenem-non-susceptible Escherichia coli surrogates.
| Antimicrobial agents | Beef | Pork | Chicken | ||||||
|---|---|---|---|---|---|---|---|---|---|
| S (%) | I (%) | R (%) | S (%) | I (%) | R (%) | S (%) | I (%) | R (%) | |
| Cefotaxime | 149 (80.5) | 15 (8.1) | 21 (11.4) | 106 (76.3) | 29 (20.9) | 4 (2.9) | 72 (81.8) | 11 (12.5) | 5 (5.7) |
| Ceftazidime | 155 (83.8) | 13 (7.0) | 17 (9.2) | 106 (76.3) | 29 (20.9) | 4 (2.9) | 80 (90.9) | 8 (9.1) | 0 (0.0) |
| Imipenem | 55 (29.7) | 13 (7.0) | 117 (63.2) | 3 (2.2) | 16 (11.5) | 120 (86.3) | 4 (4.5) | 11 (12.5) | 73 (83.0) |
| Meropenem | 84 (45.4) | 44 (23.8) | 57 (30.8) | 31 (22.3) | 57 (41.0) | 51 (36.7) | 20 (22.7) | 34 (38.6) | 34 (38.6) |
S, susceptible; I, intermediate-resistant; R, resistant.
Table 3.
Carbapenemase genes in carbapenem-resistant Escherichia coli isolates.
| Isolate code | uidA | Carbapenemase gene | Antimicrobial resistance pattern | ||||
|---|---|---|---|---|---|---|---|
| Class A | Class B | Class D | |||||
| blaKPC | blaNDM | blaVIM | blaIMP | blaOXA-48 | |||
| B14.1 | + | - | + | - | - | - | CTX, CAZ, IPM, MEM |
| B14.2 | + | - | + | - | - | - | CTX, CAZ, IPM, MEM |
| B14.3 | + | - | + | - | - | - | CTX, CAZ, IPM, MEM |
| B14.4 | + | - | + | - | - | - | CTX, CAZ, IPM, MEM |
| B14.5 | + | - | + | - | - | - | CTX, CAZ, IPM, MEM |
| B14.6 | + | - | + | - | - | - | CTX, CAZ, IPM, MEM |
| B14.7 | + | - | + | + | - | - | CTX, CAZ, IPM, MEM |
| B14.8 | + | - | + | - | - | - | CTX, CAZ, IPM, MEM |
| B14.9 | + | - | + | - | - | - | CTX, CAZ, IPM, MEM |
| B14.10 | + | - | + | - | - | - | CTX, CAZ, IPM, MEM |
| B14.11 | + | - | + | - | - | - | CTX, CAZ, IPM, MEM |
| B14.12 | + | - | + | - | - | - | CTX, CAZ, IPM, MEM |
| B14.13 | + | - | + | - | - | - | CTX, CAZ, IPM, MEM |
| B14.14 | + | - | + | - | - | - | CTX, CAZ, IPM, MEM |
| B14.15 | + | - | + | - | - | - | CTX, CAZ, IPM, MEM |
+, positive; - negative; CTX, cefotaxime; CAZ, ceftazidime; IPM, imipenem; MEM, meropenem.
Table 4.
Virulence genes in 16 carbapenem-resistant Escherichia coli isolates, with at least one carbapenemase gene carriage.
Table 4.
Virulence genes in 16 carbapenem-resistant Escherichia coli isolates, with at least one carbapenemase gene carriage.
| Isolate code | DEC indicator gene | ExPEC indicator gene | Other E. coli virulence gene | Phylogenetic group | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EPEC and EHEC | ETEC | EAEC | EIEC | DAEC | G1 | G2 | G3 | G4 | G5 | ||||||||||||||||
| stx1 | stx2 | eae | bfpA | elt | est | aggR | ipaH | daaE | papA | papC | sfaDE | afa | kpsMT II | iutA | agn43 | astA | cnf1 | fimH | hlyA | lpf | chuA | yjaA | TSPE4.C2 | Group | |
| B14.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
| B14.2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
| B14.3 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
| B14.4 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
| B14.5 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
| B14.6 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
| B14.7 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
| B14.8 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
| B14.9 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
| B14.10 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
| B14.11 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
| B14.12 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
| B14.13 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
| B14.14 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
| B14.15 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | D |
+, positive; -, negative; DEC, diarrheagenic Escherichia coli; EPEC, enteropathogenic Escherichia coli; EHEC, enterohemorrhagic Escherichia coli; ETEC, enterotoxigenic Escherichia coli; EAEC, enteroaggregative Escherichia coli; EIEC, enteroinvasive Escherichia coli; DAEC, diffusely adherent Escherichia coli; ExPEC, extraintestinal pathogenic Escherichia coli; G1, group 1; G2, group 2; G3, group 3; G4, group 4; G5, group 5.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



