
Submitted:
03 November 2025
Posted:
05 November 2025
You are already at the latest version
Abstract
European chestnut is an agroforest species of great ecological, economic, and cultural importance in many temperate regions. However, in recent decades, it has been seri-ously threatened by various factors, including devastating diseases such as chestnut blight and ink disease, as well as the impacts of climate change. In this context, bio-technological tools have emerged as a key alternative for the protection, improvement, and sustainable use of the species. This paper analyzes the main biotechnological strategies applied to European chestnut. First, classical and assisted breeding tech-niques are discussed, including controlled hybridization and the use of molecular markers to accelerate the selection of genotypes of interest. In the field of molecular biotechnology, studies related to the identification of key genes, the development of genetic markers (SSR, SNP), and the omics characterization of chestnut are reviewed. The use of micropropagation techniques for the clonal multiplication of elite individu-als is also included. Furthermore, advances in genetic modifications are explored, highlighting the introduction of resistance genes through transgenic and cisgenic ap-proaches, as well as emerging technologies such as CRISPR/Cas9. Finally, future per-spectives for the application of biotechnology in the recovery, improvement, and sus-tainability of chestnut in the face of current and future threats are presented.
Keywords:
blight disease
; conservation
; Castanea sativa
; CRISPR/Cas9
; gall wasp
; ink disease
; micropropagation
; molecular markers
; omics
1. Introduction
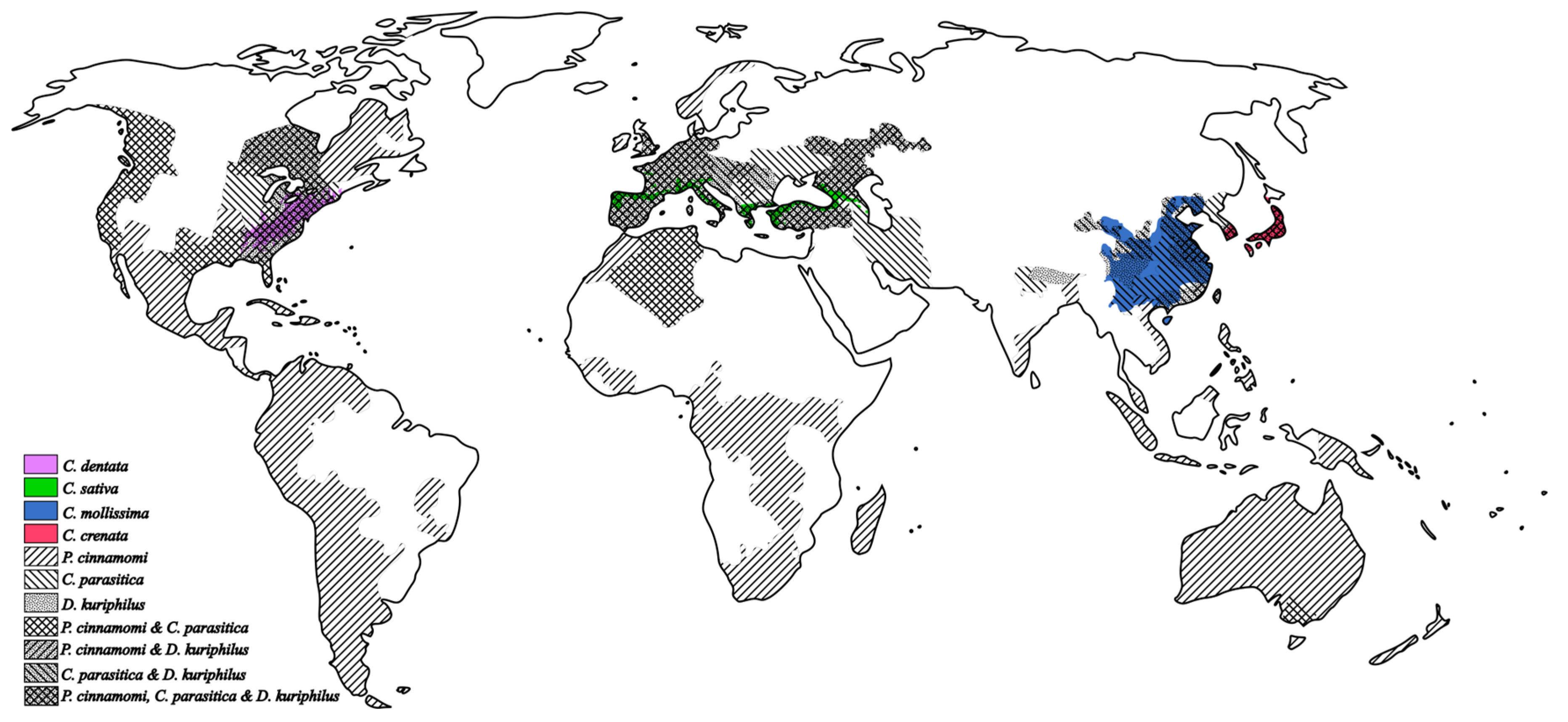
Castanea sativa Mill., also known as sweet chestnut or European chestnut, is a tree species belonging to the family Fagaceae. The genus Castanea is taxonomically divided into 13 species, the most important of which are C. sativa, C. dentata (Marsh.) Borkh. (American chestnut), and C. crenata Siebold & Zucc and C. mollissima Blume (Asian chestnuts) (Figure 1). Palynological studies indicate that the European chestnut was widely distributed during the Tertiary period, retreating to refugia in southern Europe following successive glaciations. Pollen studies also suggest a second, rapid expansion associated with human activity, particularly linked to the expansion of the Roman Empire [1]. This anthropogenic influence likely played a key role in the current distribution of the species across Europe, facilitating its spread beyond natural refugial areas through cultivation and forest management. Today, sweet chestnut is found throughout central and southern Europe including the northern Iberian Peninsula, southern France, central and northern Italy, and the southern Balkan Peninsula as well as Asia Minor, particularly western and northern Turkey and the Caucasus region [2] (Figure 1). European chestnut holds significant ecological, economic, and cultural importance. It is a multipurpose tree valued for its high-quality, durable, and rot-resistant wood, which is widely used in furniture making, fencing, and construction [3]. In addition to its timber, the species produces edible nuts and supports a variety of secondary products such as pasture, mushrooms, and berries, contributing substantially to rural livelihoods and agroforestry systems. The nutritional value of chestnuts has been recognized since ancient times, with some Greek and pre-Roman tribes even considering them superior to almonds and walnuts. According to Vieitez and Merkle [4], Montaigne wrote in his Journal du Voyage around 1570 that the Roman legions, during the Gallic Wars, survived thanks to chestnuts, which they referred to as the “bread of the forests” and called the tree Arbus panis. Currently, chestnut nuts remain highly important as a key source of income in many rural areas across Europe and are increasingly in demand for gluten-free and healthy food products. Rich in complex carbohydrates, fiber, vitamins, and antioxidants [5], chestnuts are appreciated both in traditional and modern cuisine. Finally, the European chestnut holds significant cultural value in southern Europe, historically connected to local food traditions, landscapes, and customs. It has been a vital source of sustenance during times of scarcity and features prominently in popular festivals such as Magosto in Spain, Magusto in Portugal, or Sagra in Italy. Its presence is evident in rural architecture, traditional cuisine, and agricultural landscapes, symbolizing resilience, sustainability, and a living heritage closely tied to the holistic use of forest resources.
Over the past century, two diseases have severely affected both European and American chestnut, with ink disease being one of the most destructive [6]. First reported in Spain and Portugal in the 19th century, it is now widespread across many European countries. The disease is primarily caused by Phytophthora cinnamomi Rands, prevalent in Spain, Portugal, and France, and by P. cambivora (Petri) Buisman, more common in Italy and Greece [7]. These oomycetes, whose cell walls contain cellulose and glucans instead of chitin, infect tree roots and spread through living tissues, eventually disrupting the transport of water and nutrients. Symptoms include chlorosis, reduced fruit size, wilting, and ultimately tree death [8]. The disease is identified by a dark necrotic area at the root collar and a black exudate in the surrounding soil. It spreads rapidly through zoospores and chlamydospores in moist soils, with its progression influenced by both environmental conditions and human activity. Its spread has been documented in both plantations and natural forests, and its impact is expected to increase under future climate change scenarios, as predicted by simulation models [8,9]. The distribution of this oomycete has previously been linked to climate conditions [10]. It is strongly limited by low temperatures, becoming inactive in soil below 10 °C and unable to grow in artificial media under 5 °C [11,12]. Although some isolates can produce sporangia at 7.5–10 °C, effective infection of host plants does not occur below 8–9 °C, restricting their presence in alpine and subalpine regions [13]. The distribution of P. cinnamomi shown in Figure 1 is based on data obtained from EPPO [14], which includes records from the 1990s for some regions with severe winters (i.e., some northeastern U.S. states; Canada). Therefore, some of these records may reflect temporary introductions that are no longer present and have not been recently reassessed. The other major disease affecting chestnut is blight, caused by the fungus Cryphonectria parasitica (Murr.) Barr., which infects the aerial parts of the tree through wounds [15]. C. parasitica, native to East Asia, was introduced to North America in the late 19th century, where it decimated populations of the American chestnut. It is now widespread in the eastern US and present in parts of Canada (Figure 1). In Europe, the fungus was first reported in 1938 near Genoa (Italy) on European chestnut, and has since spread throughout much of Europe, showing variable distribution and impact across different regions (Figure 1). The fungus invades the cortical tissue and develops as a saprophyte, causing necrosis that can completely girdle branches or the trunk, leading to the death of the affected organ [6]. Although the tree may partially survive, both wood quality and fruit production are severely impacted. In response, the tree produces epicormic shoots at the base of the canker. In European chestnut, the pathogen virulence is reduced due to hypovirulence caused by a hypovirus [16](and references therein); however, in recent years, severe outbreaks have occurred in Spain and northern Portugal [17,18]. Finally, chestnut trees have been increasingly affected by another threat, an insect pest known as Dryocosmus kuriphilus Yasumatsu, which causes significant damage, especially when infestations persist over successive years [19]. Native to China, it has spread to major chestnut-growing regions worldwide and is now extended across East Asia, North America, and much of Europe, including Italy, France, Spain, and Portugal (Figure 1). Galls formed by this insect develop on shoots, leaf midribs, or stipules, and following adult emergence, they substantially alter the branch architecture of chestnut trees [20]. This results in up to a 70% reduction in leaf area, fewer dormant buds, and a decline in the production of wood and flowers [21]. Infestation by the chestnut gall wasp also leads to significant reductions in nut yield, with annual losses reported between 15–30% in China and 50–75% in the United States [22]. Interestingly, Asian species such as C. crenata and C. mollissima are resistant to both ink disease and blight but are susceptible to the gall wasp [22]. The spatial distribution of the three pathogens is presented in Figure 1.
2. Biotechnological Tools Applied to Chestnut
An effective alternative for managing diseases primarily caused by P. cinnamomi is the development and deployment of resistant or tolerant chestnut plants, which can subsequently be used for reforestation in affected or susceptible areas. While tolerance could theoretically be achieved through conventional breeding programs, such long-term improvement initiatives have rarely been implemented for chestnut and other long-rotation hardwood species due to the extended timelines involved. As noted by Savill et al. [23], investments in projects with lifespans exceeding 40 years have historically been unattractive, a sentiment echoed by Vieitez et al. [24]. Among hardwoods, chestnut stands out as one of the few species where classical genetic improvement programs targeting P. cinnamomi tolerance have been actively pursued. During the early 20th century, hybridization between European chestnut and Asian resistant species (C. crenata and C. mollissima) represented the primary strategy to combat ink disease. In Spain, this effort was spearheaded by Cruz Gallástegui at the Biological Mission of Galicia (MBG-CSIC) [25]. Although the resulting hybrids exhibited enhanced tolerance to P. cinnamomi, they generally produced smaller trees and fruits of inferior size and quality compared to pure European chestnut. Currently, in the rainy northern regions of Spain, hybrid rootstocks have helped mitigate the impact of P. cinnamomi; however, these rootstocks are less suitable in southern regions due to their lower tolerance to cold and drought conditions [26].
Figure 1.
Map illustrating the geographic distribution of natural populations of Castanea sativa (European chestnut), C. dentata (American chestnut), C. mollissima (Chinese chestnut), and C. crenata (Japanese chestnut), along with the primary biotic stresses impacting each species, including chestnut blight (Cryphonectria parasitica), ink disease (Phytophthora cinnamomi), and the chestnut gall wasp (Dryocosmus kuriphilus). Map created with data from several sources [14,27,28,29,30,31,32] using Inkscape [33]. Map background available at https://commons.wikimedia.org/wiki/File:Simplified_blank_world_map_without_Antartica_(no_borders).svg.
Figure 1.
Map illustrating the geographic distribution of natural populations of Castanea sativa (European chestnut), C. dentata (American chestnut), C. mollissima (Chinese chestnut), and C. crenata (Japanese chestnut), along with the primary biotic stresses impacting each species, including chestnut blight (Cryphonectria parasitica), ink disease (Phytophthora cinnamomi), and the chestnut gall wasp (Dryocosmus kuriphilus). Map created with data from several sources [14,27,28,29,30,31,32] using Inkscape [33]. Map background available at https://commons.wikimedia.org/wiki/File:Simplified_blank_world_map_without_Antartica_(no_borders).svg.
Forest biotechnology offers promising tools to accelerate tree improvement by integrating molecular techniques into breeding programs. These include marker-assisted selection, genotyping for individual tree identification, the discovery of genes or alleles linked to pathogen resistance, as well as genetic engineering approaches, in vitro culture techniques, and cryopreservation for the development, propagation, and long-term conservation of improved genotypes [3,34,35]. In particular, genomic selection holds great potential to significantly shorten the time required for genetic evaluation of desirable traits in trees, thereby enhancing the efficiency and precision of breeding efforts [36,37].
Here, we provide a comprehensive review of the various biotechnological methods applied in the breeding, propagation, and conservation of the European chestnut. This includes traditional genetic transformation, genome editing techniques such as CRISPR/Cas9, in vitro culture, cryopreservation, and omics technologies—all integrated to enhance disease resistance (Figure 2).
2.1. Classical and Assisted Genetic Improvement
2.1.1. Controlled Hybridization
Interspecific hybridization has long underpinned the European chestnut breeding in Europe, particularly for enhancing resistance to ink disease and chestnut blight, but also for addressing other critical challenges in chestnut cultivation, such as timber production, nut quality, and grafting compatibility [38,39,40,41,42]. As mentioned above, beginning in the early 20th century, crosses between C. sativa and Asian species (C. crenata and C. mollissima) have provided the basis for breeding efforts aimed at introgressing desirable traits [43,44,45,46,47].
While C. crenata shows higher resistance to diseases, its poor climatic adaptation to early frost and drought conditions, undesirable phenotypic traits, and graft incompatibility with C. sativa cultivars have limited its direct use for nut or timber production [40,46,48]. Nonetheless, C. crenata has been the main source of ink disease resistance used in Europe, and several successful hybrid clones were derived from multiple breeding programs in France, Spain, Portugal, and Italy. These hybrids were commercialized for nut and timber production, as well as for use as rootstocks [48,49,50,51].
By contrast, C. mollissima has been particularly valued as a donor of resistance to chestnut blight and, to a lesser extent, for favourable nut traits such as ease of peeling and larger seed size. Although its direct adaptation to European environments is also limited, the incorporation of C. mollissima germplasm has complemented C. crenata in generating more balanced hybrids, combining tolerance to both pathogens with improved nut quality [45,46]. Several clones from Spanish breeding programs (111-1, 7521, 2671, and 1483) are widely used as rootstocks for their high tolerance to ink disease and high rootstock compatibility with fruit varieties [52]. In Portugal, more than fifty genotypes resistant to ink disease were selected during the first decade of the 2000s [53,54]. Following these efforts, the genetic improvement program initiated at Instituto Nacional de Investigação Agrária e Veterinária (INIAV) in 2006 placed particular emphasis on resistance to ink disease, given its continued negative impact on national orchards. This initiative also sought to modernize the plant material used in new plantations, particularly rootstocks, to ensure better adaptation to current climatic conditions, since most varieties then available still derived from breeding programs of the previous century [49,55,56]. Within this program, seven hybrid chestnut genotypes with enhanced resistance to ink disease were developed through controlled crosses (C. sativa × C. crenata/mollissima). The selection of these genotypes was based on inoculation trials conducted under controlled conditions on the progenies obtained from the crosses, allowing a rigorous characterization of their responses to P. cinnamomi infection. Disease severity was assessed using standardized symptom scales and mortality indices. Additionally, pathogenicity tests following Koch’s postulates were conducted, ensuring reliable identification of the pathogen and confirming its causal association with the observed symptoms. The robustness of these tests was reinforced through inoculations on biological replicates of each genotype, obtained via micropropagation. The results revealed marked differences among genotypes, with some exhibiting significantly attenuated symptoms, reduced root necrosis, limited lesion progression to the collar, and higher survival rates [57]. Three of these genotypes are now registered in the Portuguese National Register of Fruit Varieties, including SC1202 (Variety RIVERA), which currently has an application under review for Community Plant Variety protection at the CPVO (Community Plant Variety Office). The propagation of these genotypes was achieved through micropropagation, enabling large-scale clonal production critical for validating P. cinnamomi inoculation trials, establishing field trials, conducting graft compatibility studies, transferring technology to nurseries, and conducting more fundamental studies like the identification of candidate genes of resistance. In parallel, the Spanish public company TRAGSA selected and officially registered seven new hybrid clones (denominated C003, C004, C042, C053, P011, P042, and P043) with resistance to ink disease in the Spanish National Catalogue of Basic Plant Materials.
Other examples in France and Italy include Euro-Asian cultivars such as ‘Bouche de Bétizac’ and ‘Primato’, rootstocks such as ‘Maridonne’ and ‘Marlhac’, and rootstocks used directly as fruit producers ‘Marigoule’ and ‘Maraval’, which combine disease resistance with improved agronomic and adaptive traits [58,59,60]. More recently, some of these cultivars have also demonstrated resistance to the chestnut gall wasp [59,61,62]. Parallel breeding efforts have also been conducted outside Europe. In North America, C. mollissima and C. crenata have been used as donors of blight resistance in backcross programs with the American chestnut (C. dentata), with the aim of restoring timber chestnuts [63]. In Japan, programs have emphasized resistance to gall wasp together with nut traits such as ease of peeling [64].
Despite the success of these breeding programs, challenges remain in achieving optimal adaptation of hybrids to European climatic conditions. Over recent times, the intensification of drought and heat, acting synergistically with ink disease, has exacerbated damage in southern European chestnut stands [3,65]. Under these conditions, hybrid clones derived from C. sativa x C. crenata and C. mollissima have shown poor tolerance to these climate stressors [66,67,68]. In Spain, a breeding initiative was launched with the aim of developing trees tolerant to multiple stresses associated with global change [69,70,71]. Recently, two newly developed C. sativa clones have been reported to combine resistance to P. cinnamomi with tolerance to drought and heat, and they are currently undergoing registration at the Spanish Plant Variety Office [72].
Overall, progress in classical hybridization demonstrates the potential of interspecific breeding to provide genetic solutions for major chestnut diseases. However, the interaction of biotic and abiotic stresses under climate change highlights the need for integrated new breeding strategies. Future advances are expected to result from combining controlled hybridization with genome-assisted selection tools to accelerate the development of resilient C. sativa genotypes adapted to Mediterranean environments.
2.1.2. Perspective for Marker-Assisted Selection (MAS)
Advancements in molecular breeding must be integrated into chestnut breeding programs to support the increasing demand for elite chestnut genotypes. Thus, the application of marker-assisted selection (MAS) will be instrumental in accelerating the introgression of disease resistance and other beneficial traits such as environmental adaptability and agronomic performance. A foundational step was the development and deployment of molecular markers capable of identifying traits of interest and serving as tools for MAS programs. In C. sativa, genomic simple sequence repeats (gSSRs) were first developed approximately 20 years ago [73,74]. Since then, numerous studies have been conducted using these markers, and in perspective, they have provided the basis for MAS programs. Early applications of gSSRs demonstrated robust genotyping capacity, enabling accurate varietal discrimination [75,76,77,78]. For example, Pereira-Lorenzo et al. (2017) [79] used 24 highly polymorphic SSR markers to evaluate 271 accessions corresponding to 118 European cultivars, establishing a comprehensive European reference database for the identification and characterization of chestnut varieties.
In the Iberian Peninsula and surrounding regions, diversification and conservation of chestnut cultivars have also been explored using molecular tools. Pereira-Lorenzo et al. (2011) [80] performed a large-scale SSR-based survey on 593 grafted chestnut trees from Iberian Peninsula, and the Azores, and the Canary Islands, including more than 300-year-old individuals. They identified 356 distinct genotypes and reported a clonality rate of 33%, reflecting the widespread use of grafting. Despite clonal propagation, high genetic variability was maintained through hybridization and mutation, which emerged as the main driver of diversification. Ten cultivar groups were defined, exhibiting strong geographic structuring and evidence of long-distance dissemination to Atlantic islands, underscoring the role of human-mediated dispersal. The importance of molecular markers in distinguishing hybrids and detecting introgression from Asian species was highlighted by Pereira-Lorenzo et al. (2010) [81]. More recently, instant domestication has been highlighted to explain how traditional practices of selection and propagation rapidly shaped chestnut diversity [82]. The introduction of new sequencing techniques enabled the transformation of Single Nucleotide Polymorphisms (SNPs) into competitive allele-specific PCR (KASP) markers, successfully evaluated for varietal discrimination [83].
Moreover, information for MAS programs has also been obtained using functional markers such as Expressed Sequence Tag – Simple Sequence Repeats (EST-SSRs), which target expressed genes and therefore provide opportunities to directly associate genetic variation with adaptive traits. Their potential lies in supporting early selection for stress tolerance and disease resistance, complementing gSSR markers. Applications of EST-SSRs have included both the characterization of cultivated germplasm and of wild populations to evaluate the genetic and adaptive potential of chestnut in Europe [76,84]. Their potential has been further demonstrated through studies that revealed associations with water stress responses and tolerance to P. cinnamomi, supporting their use in the identification of drought-tolerant and disease-resistant individuals within breeding populations [26,69].
Population-level studies have further strengthened the case for functional markers. Studies using gSSRs revealed distinct gene pools across coppice and wild populations, shaped by geography and environmental gradients [85,86]. More recent work based on EST-SSRs expanded this approach, identifying loci potentially involved in stress responses. Castellana et al. [87], using the same EST-SSR markers to study variation across European chestnut populations, reported a potential association between the FIR059 allele and climatic variables, suggesting a role in abiotic stress adaptation. Similarly, Dorado et al. [70] used molecular markers associated with heat stress (VIT099 and POR016) to assess tolerance within and between populations, highlighting loci under positive selection. These markers stand out as promising candidates for early selection of heat-tolerant C. sativa individuals.
The availability of multilocus marker sets (SSRs, SNPs, double digest restriction-site associated DNA sequencing; ddRAD-seq) has also enabled the construction of high-density genetic linkage maps, which, combined with functional annotation of quantitative trait loci (QTL) intervals, provide a molecular basis for MAS and a platform for future genomic selection efforts. The first genetic linkage map for C. sativa [88], based on Random Amplified Polymorphic DNA (RAPD), Inter-Simple Sequence Repeat (ISSR), and isozyme markers, was constructed using the two-way pseudo-testcross strategy. This map facilitated the identification of QTLs associated with adaptive traits such as growth and water-use efficiency [89]. Comparative genetic and QTL mapping between Quercus robur L. and C. sativa identified homologous genomic regions, allowing putative candidate genes for bud burst to be inferred from colocations of EST-derived markers and QTLs [90].
Further refinement of genetic mapping has been achieved through bin mapping approaches, which cluster markers into cosegregating bins to improve map resolution and reduce redundancy, facilitating high-throughput genotyping and QTL discovery [91,92]. These genomic resources are now actively translated into MAS pipelines in C. sativa breeding programs. For example, breeders can pyramid resistance alleles for ink disease and gall wasp across generations using flanking SSR or SNP markers, while markers linked to heat tolerance allow early selection of individuals better adapted to increased temperatures. These approaches have already accelerated cultivar development, demonstrating how genomics-guided MAS complements classical breeding.
2.2. Molecular and Genomic Approaches
Extensive research aimed at understanding the genetic basis of resistance and susceptibility to chestnut’s main biotic stresses has been driven by the urgent need to safeguard chestnut forests and orchard health. Advances in genetics, cell and molecular biology, and bioinformatics -complemented by insights from histopathology and physiology - have collectively built a robust foundation of knowledge, largely enabled by high-throughput sequencing technologies. Over the past two decades, genomic data generated by numerous research programs has rapidly grown, providing valuable resources for the development of effective and timely control and management strategies. Building on this foundation, the following section integrates and summarizes current knowledge from transcriptomic, proteomic, and metabolomic analyses, QTL mapping, molecular marker development, and whole-genome sequencing. It offers a comprehensive overview of the molecular and genomic underpinnings of chestnut defense against P. cinnamomi, C. parasitica, and D. kuriphilus, with a particular focus on European chestnut and its hybrids. The section also highlights candidate genes and pathways that may inform future resistance breeding and genetic engineering efforts, while outlining the genetic architecture that shapes chestnut responses to these biotic stresses.
2.2.1. Molecular Mechanisms of Castanea Defence Against Phytophthora cinnamomi
Molecular studies have revealed that resistance and tolerance to P. cinnamomi in C. sativa and its hybrids with Asian chestnuts involve a complex interplay of constitutive and inducible defense mechanisms, including gene expression regulation, biochemical barriers, and cellular responses [93,94,95,96].
A fundamental aspect of plant immunity is the presence of pre-formed, constitutive defenses that provide the first barrier against pathogen invasion. In the resistant C. crenata, high basal expression of defense-related genes such as those encoding receptor-like kinases (RLKs) and antifungal proteins (Cast_Gnk2-like) may fortify the root environment, rendering it less hospitable to pathogen colonization. By contrast, the susceptible C. sativa typically exhibits lower constitutive expression levels of those genes, predisposing it to a rapid initial invasion by P. cinnamomi [95]. The absence or low expression of pattern recognition receptors (PRRs), including BAK1 orthologs, in C. sativa may compromise its ability to perceive pathogen-associated molecular patterns (PAMPs) early during infection, thereby delaying the activation of downstream immune responses [93]. The weak pre-formed defence seems to contribute to the susceptibility of C. sativa.
A recurrent theme in the response of Castanea spp. to P. cinnamomi is the critical importance of timing. Resistant C. crenata exhibits an early activation of cellular defenses (within 0.5–2 h post-inoculation), which effectively curtails pathogen colonization [97]. These early cellular responses appear to reflect constitutive rather than induced gene expression, as evidenced by previously reported high expression levels of resistance-related genes in non-inoculated plants [95] and the limited induction of only a single gene 2 h post-inoculation [93]. By contrast, C. sativa demonstrates an early but transient activation of some defense components, followed by a marked decline in gene expression at 48–72 h post-inoculation. Notably, at the same time points, genes encoding for elicitins/elicitin-like (oomycete PAMPs) and necrosis-inducing like proteins (NLPs) were more expressed by P. cinnamomi in C. sativa compared to C. crenata [93]. This temporal mismatch not only allows P. cinnamomi to bypass the initial plant defenses but also creates a window during which pathogen effectors can further suppress host immune responses, promoting disease progression. Consequently, the continuum and sustainability of the defense response in resistant genotypes are critical determinants of successful resistance, while the inability of C. sativa to maintain these defenses results in elevated susceptibility.
The rapid and robust response of C. crenata includes the upregulation of genes encoding pathogenesis-related (PR) and antifungal proteins. In contrast, susceptible C. sativa displays delayed and weaker expression of these genes, resulting in more severe symptoms and higher mortality rates [94,95]. Santos et al. [95] conducted an in-depth analysis of gene expression responses in resistant (C. crenata), susceptible (C. sativa), and interspecific hybrids inoculated with P. cinnamomi using digital PCR. Among the eight candidate genes tested, clear genotype-dependent differences were observed. Remarkably, Cast_Gnk2-like exhibited not only a high constitutive expression level but also a rapid and strong activation in C. crenata following P. cinnamomi inoculation, whereas C. sativa showed low constitutive and induced levels of expression. Therefore, Cast_Gnk2-like emerged as a potential discriminator of resistance.
Cast_Gnk2-like shares homology with the ginkbilobin-2 (Gnk2) gene from Ginkgo biloba seeds. Gnk2 is a cysteine-rich repeat secreted protein harbouring a DUF26 domain with antifungal activity [98]. Gao et al. [99] reported that Gnk2 is associated with the induction of Programmed Cell Death (PCD). Miyakawa et al. [100] suggest that Gnk2 acts as a lectin and showed that it binds with high affinity to D-mannose, which may underlie its inhibitory effect on pathogen growth. As this carbohydrate is present in the cell wall of P. cinnamomi, some authors of this review hypothesized that the protein encoded by Cast_Gnk2-like could have a direct effect on the pathogen [101]. This hypothesis led to a series of functional validation studies within the genetic improvement program for C. sativa and other Fagaceae [102,103,104].
Comparative transcriptomic studies between resistant and susceptible Castanea species have revealed that resistant genotypes, such as C. crenata, upregulate a broader array of genes involved in pathogen perception, signal transduction, transcription factor activation, and the biosynthesis of defense metabolites [93,94]. In contrast, the transcriptome of C. sativa exposed to P. cinnamomi is characterized by a more limited set of differentially expressed genes, with lower overall expression levels of key defense markers. Proteomic analyses further corroborate these findings, indicating that proteins involved in salicylic acid (SA) signaling, reactive oxygen species (ROS) metabolism, and cell wall reinforcement are less abundant in infected C. sativa tissues [105].
Plant growth regulators (PGRs) play a crucial role in modulating immune responses against pathogens. Induction of SA-mediated responses results in the activation of PR proteins, cell wall reinforcement, and hypersensitive response (HR), which are key for resistance against hemibiotrophic pathogens [106]. Moreover, the interplay between SA and other hormones such as jasmonic acid (JA), abscisic acid (ABA), and ethylene (ET) is critical for fine-tuning the immune response. ABA often antagonizes SA-mediated responses.
A recent report, using physiological and biochemical methods, found that the response to P. cinnamomi in a resistant C. sativa x C. crenata genotype was characterized by early and strong SA signaling in roots, antagonism of ABA, stable primary and secondary metabolism, and transient oxidative stress with recovery [96]. On the other hand, the susceptible C. sativa response involves delayed and weak JA signaling, lack of SA induction, high ABA accumulation in leaves, impaired carbohydrate and secondary metabolism, and fluctuating oxidative stress that is not effectively countered (insufficient antioxidant response). Indeed, Santos et al. [95] suggested that SA signaling may be activated more rapidly in resistant genotypes than in susceptible ones, based on the expression patterns of Cast_WRKY31 and Myb-related protein 4 (Cast_Myb4), transcription factors related to SAmediated responses.
Expression profiling of susceptibility genes (S genes) during P. cinnamomi infection provided further evidence supporting the delayed activation of SA-mediated defenses in the European chestnut [107]. In C. sativa, the S-genes powdery mildew resistance 4 (pmr4, encoding a callose synthase) and downy mildew resistance 6 (dmr6, repressor of the SA pathway) show early upregulation upon infection. This upregulation has been correlated with the suppression of SA-dependent defenses, thereby enhancing the susceptibility of the host. In contrast, C. crenata does not exhibit significant upregulation of these genes.
The allene oxide synthase gene from C. crenata (CcAOS), an ortholog of a key enzyme in the JA pathway, was identified from 2015 transcriptome data [94] after it was found to be significantly induced in inoculated C. crenata compared to C. sativa. The functional relevance of CcAOS in plant defense against P. cinnamomi was demonstrated by gene functional analysis, using genetic transformation to overexpress CcAOS in the susceptible Arabidopsis ecotype Ler-0, which resulted in delayed pathogen progression and enhanced tolerance [108].
Another essential mechanism of plant defense is the strengthening of the cell wall through the accumulation of phenolic compounds, which create physical barriers to pathogen cellular ingress and progression. Transcriptomic data indicate that the expression of genes linked to cell wall reinforcement such as those related to lignin biosynthesis and structural proteins is lower in C. sativa than in C. crenata [93]. This includes genes encoding cell wall-modifying enzymes such as Pectinesterase 2 (Cast_PE-2) and the TF (Cast_Myb4) [95]. These genes are upregulated promptly in resistant genotypes (C. crenata and C. sativa x C. crenata) after pathogen detection, which could lead to rapid cell wall thickening and reinforcement. Studies at the cellular level reveal these temporal differences, showing the accumulation of phenolic compounds in C. crenata and C. sativa cell walls at 30 min and 72 h after infection, respectively [97].
C. sativa can accumulate callose around intracellular hyphae as early as 24h after inoculation [97]. However, this cellular response, possibly mediated by S-gene pmr4 [107], does not constitute an effective resistance defense. The inefficient and tardy activation of structural defenses allows P. cinnamomi to advance colonization and eventually cause widespread tissue necrosis. This is supported by the area of P. cinnamomi infection [97] and the quantity of pathogen DNA present in host tissues [107], which are significantly higher in C. sativa compared to C. crenata. Moreover, the lack of sustained activation of enzymes responsible for the cross-linking of cell wall components further exacerbates the vulnerability of C. sativa to pathogen invasion [93].
The rapid production of ROS is one of the hallmark responses of plants to pathogen attack, serving both as a direct antimicrobial agent and as a secondary signal to activate further defense responses. In resistant C. crenata, transcriptomics suggests a pronounced burst of ROS in infected tissues that may initiate HR, a rapid localized pathogen-induced cell death, thereby limiting pathogen spread [93,97]. While initial ROS generation may occur in C. sativa as well, this response may not be sustained; the transcription of genes coding for enzymes such as respiratory burst oxidase homolog protein B (RBOHB) is often transient, and the subsequent detoxification by antioxidative enzymes may further diminish the antimicrobial effects. Additionally, negative regulators of programmed cell death, such as BON1-associated protein 2-like (BAP2-like), are upregulated in C. sativa, possibly contributing to suppressing HR-like cell death and allowing P. cinnamomi to shift from biotrophic to necrotrophic lifestyle. The failure to maintain a robust ROS-mediated defense, coupled with premature suppression of HR, could be a key factor underlying the inability of C. sativa to restrict pathogen proliferation.
Dorado et al. [8] studied C. sativa defense against P. cinnamomi in warming scenarios and found that it depends on both morphological resilience (growth, root biomass) and the ability to accumulate specific phenolic compounds with antioxidant and antimicrobial properties: quercetin 3-O-glucuronide, 3-feruloylquinic acid, gallic acid ethyl ester, and ellagic acid. Briefly, plants previously exposed to moderate warming were more resilient to the pathogen, while those under normal or heat wave conditions were more susceptible. On the other hand, surviving plants after infection were characterized by increased levels of the four phenolic metabolites, which are central to the adaptive response to the combined challenges of heat and pathogen attack.
The Ubiquitin/26S proteasome system is integral to the dynamic regulation of plant immune responses through the targeted degradation of key proteins involved in hormone signaling and plant defense [109]. Some components of the system may also inhibit pathogen effectors by triggering PAMP and ETI responses [109]. Transcriptomics shows that C. crenata exhibits a high differential expression of 26S proteasome regulatory subunit 4 homolog A (together with several proteases) after inoculation with P. cinnamomi, when compared to C. sativa [93]. The apparent absence of regulation of the Ubiquitin/26S proteasome system in C. sativa conceives the persistence of negative regulators that dampen defense responses. Although the precise components of this regulatory network in chestnut remain to be elucidated, current evidence underscores the importance of post-translational modifications in fine-tuning the balance between resistance and susceptibility during P. cinnamomi infection.
2.2.1.1. Genetic Basis of P. cinnamomi Resistance: Marker Development and Quantitative Trait Loci (QTL) Mapping
Building on the availability of genomic resources, including transcriptomic data [94], Santos et al. [110] developed 43 novel EST-SSR markers. These markers, designed from differentially expressed genes associated with host responses to infection, showed high amplification success and interspecific transferability across four Castanea species. The average expected heterozygosity (0.61) was higher than previously reported for chestnut EST-SSRs, confirming their utility for genetic studies.
An important advance for C. sativa breeding programs was the construction of the first interspecific genetic linkage map for C. sativa × C. crenata [111]. This map was constructed using 452 SSRs and SNPs and spans 498.9 centimorgans (cM). It enabled the detection of QTLs for ink disease resistance on linkage groups E and K, which overlap with QTLs from American x Chinese chestnut populations [112], suggesting conserved P. cinnamomi defense mechanisms across chestnut species. Additionally, QTLs on linkage group E co-localized with defense-related genes, including those putatively encoding PR proteins (NDR1/HIN1-Like protein 3), phospholipid transporters, transcriptional regulators (RNA polymerase II-associated factor 1; PAF1 homolog), and epigenetic modulators (Zinc-finger PHD-type)[111].
Genetic linkage maps and the identification of QTLs associated with ink disease resistance have revealed the polygenic nature of this trait in chestnut. These studies not only provided evidence of the genetic architecture underlying ink disease resistance in chestnut but also delivered molecular markers with strong potential for MAS. Table 1 summarizes genomic resources and key insights into the responses of susceptible and resistant chestnuts to P. cinnamomi infection.
2.2.2. Molecular Mechanisms of Castanea Defense Against Cryphonectria parasitica
Chestnut blight poses a significant threat to chestnut species worldwide. While American chestnut is highly susceptible, the European chestnut displays relatively lower susceptibility and, in some cases, tolerance. Among the Asian species, the Chinese chestnut is considered more resistant to blight than the Japanese chestnut, which is attributed to a combination of rapid immune response activation, efficient pathogen recognition, and robust structural defenses [113].
2.2.2.1. Castanea sativa: Partial Tolerance or Susceptibility to Blight
C. sativa can be susceptible to C. parasitica, particularly to virulent strains that cause extensive necrosis and canker formation. The fungus typically penetrates into the host tissue through wounds or bark cracks, often facilitated by environmental stressors like drought or mechanical damage. Once inside, the pathogen forms mycelial fans that exert physical pressure and enzymatically degrade host tissues using laccases, cellulases, and cutinases. Additionally, the pathogen produces phytotoxic secondary metabolites, including skyrin, skyrinol, rugulosin, diaportin, and nitrogen-containing compounds, which cause rapid necrosis in host tissues. Apical shoots and leaves are particularly sensitive to these toxins, with mortality observed within 8 days [114]. The host tree attempts to respond by lignifying cell walls to reinforce structural barriers and forming wound periderm, a protective layer to isolate infected tissue. However, these defenses are consistently suppressed by the advancing fungal mycelium. Instances of limited recovery of infected trees have been documented under certain environmental conditions or in specific genotypes; these occurrences are best interpreted as examples of tolerance rather than resistance [115]. Tolerance allows trees to survive and maintain some functionality despite the presence of the pathogen, but it does not preclude initial infection or the subsequent establishment of cankers.

Table 1.
Biotechnological tools to study chestnut responses to Phytophthora cinnamomi, focused on Castanea sativa (susceptible) and Castanea crenata (resistant).
Table 1.
Biotechnological tools to study chestnut responses to Phytophthora cinnamomi, focused on Castanea sativa (susceptible) and Castanea crenata (resistant).
| Methodology | Species | Main Findings | References |
|---|---|---|---|
|
Comparative transcriptomics |
C. crenata C. sativa |
C. crenata upregulates genes for pathogen perception, signaling, transcription factors, and defense metabolites. C. sativa shows limited and transient expression | [93,94] |
|
Molecular marker development |
C. sativa C. crenata |
43 EST-SSR markers identified from DEGs associated with host responses to infection | [110] supported by results in [94] |
| Genetic mapping | C. sativa x C. crenata | Interspecific linkage map enabled detection of QTLs for pathogen resistance on linkage groups E and K, co-localizing with defense-related genes | [111] supported by results in [110] |
|
Gene expression profiling |
C. crenata C. sativa C. sativa x C. crenata |
C. crenata shows high basal and induced expression of PR genes (e.g., RLKs, Cast_Gnk2-like), enabling early defense activation. C. sativa has lower expression, allowing rapid pathogen colonization | [93,95] in accordance with results in [97] |
|
Functional gene validation |
C. sativa C. dentata Quercus ilex Quercus suber Arabidopsis thaliana |
Cast_Gnk2-like relevant in Castanea and Quercus defense; CcAOS enhances tolerance in A. thaliana Ler-0 | [102,103,104,108] corroborated by results in [95] |
| Proteomics | C. sativa | C. sativa upon infection shows downregulation of proteins involved in SA signaling | [105]in accordance with results in [93,94] |
|
Histopathology and cellular studies |
C. sativa C. crenata |
C. crenata responds more efficiently than C. sativa; pathogen’s growth is restricted by early activation of callose deposition, HR-like cell death, cell wall thickening and accumulation of phenolic-like compounds. | [97] in accordance with results in [93,107] |
| Susceptibility gene expression analysis |
C. sativa C. crenata |
C. sativa upregulates pmr4 and dmr6 early in the infection, putatively contributing to suppressing SA defenses; putative callose accumulation via pmr4 is not sufficient to restrict pathogen growth | [107] in accordance with results in [97] |
|
Metabolite analysis |
C. sativa | Moderate warming enhances C. sativa resilience to pathogen. Surviving plants accumulate key phenolics (e.g., quercetin 3-O-glucuronide, ellagic acid), contributing to defense | [8] |
|
Physiological and biochemical assays |
C. sativa C. sativa x C. crenata |
C. sativa × C. crenata show early SA signaling, ABA antagonism, and oxidative stress recovery. C. sativa shows delayed JA signaling, high ABA, impaired metabolism, and weak antioxidant response | [96] in accordance with results in [95] |
ABA: Abscisic acid; CcAOS: Allene Oxide Synthase; DEGs: Differentially expressed genes; dmr6: Downy Mildew Resistant 6; EST-SSR: Expressed Sequence Tag – Simple Sequence Repeats; HR: Hypersensitive response; JA: Jasmonic acid; pmr4: Powdery Mildew Resistant 4; PR: Pathogenesis-related; QTL: Quantitative Trait Loci; RLKs: Receptor-like kinases; SA: Salicylic acid.
- Lowering of photosynthetic pigments and augmentation of antioxidant enzyme activities (Ascorbate peroxidase (APX), Guaiacol peroxidase (POD), and Superoxide dismutase (SOD)).
- Accumulation of the stress markers proline (an amino acid that in stress conditions acts as an osmolyte, stabilizes proteins, and neutralizes ROS) and malondialdehyde (a marker of lipid peroxidation caused by oxidative stress levels in infected tissues).
Despite C. sativa susceptibility to C. parasitica, the presence of hypovirulent fungal strains and the use of biological control agents like Cryphonectria hypovirus 1 (CHV1) have shown promise in mitigating disease severity in Europe [117]. These strains exhibit reduced growth and sporulation and cause superficial necrosis that may heal over time (with the development of calli), allowing the tree to compartmentalize the infection [118]. Also, hypovirulent strains produce lower levels of virulence enzymes like laccase [114]. In another set of experiments, hybrids between C. sativa and C. crenata were subjected to inoculation with both virulent and hypovirulent strains [119,120]. These studies confirmed that while the hybrids can showcase enhanced tolerance and, in some cases, genotype-dependent recovery, the pure C. sativa lines included in the trials remained susceptible to the pathogen, thereby underscoring the absence of innate immunity.
Chitinases are hydrolytic enzymes that break down chitin, a key component of fungal cell walls, and are considered crucial inducible defense proteins in plants [121]. Gene and protein expression studies in C. sativa showed systemic induction of chitinase (and β-1,3-glucanase) in response to infection, with higher activity observed in trees inoculated with hypovirulent strains [122,123]. Vannini et al. (1999) [124] purified four chitinases from C. sativa and found that three of them inhibit hyphal growth in vitro. Also, hypovirulent strains were more susceptible than virulent ones, suggesting that slower hyphal development in hypovirulent strains may be related to vulnerability to host chitinases. These findings imply that the hypovirulence-associated virus may enhance host recognition or defense responses. The endochitinase-like protein Ch3, isolated from C. sativa cotyledons, showed antifungal properties in vitro [125], and its corresponding gene was isolated for further validation as a resistance gene [126]. More details are given in section 2.4.1.
Pavese et al. (2021) [107] conducted gene expression profiling of C. sativa inoculated with C. parasitica. Their findings revealed that the upregulation of S genes such as pmr4 and dmr6 (more detailed in 2.2.1) may activate stress pathways that inadvertently facilitate the pathogen’s progression by downregulating SA-mediated responses. Also, investigations into the expression of PR proteins, including chitinases and glucanases, demonstrated significant induction upon fungal infection.
SA is a signaling molecule largely associated with plant defense, particularly in activating pathogen resistance genes [127]. Biochemical studies in C. sativa show SA accumulation following inoculation with both virulent and hypovirulent strains of C. parasitica, with significantly higher levels in trees infected by hypovirulent strains, suggesting either enhanced host response or suppression by virulent strains [123]. This pattern mirrors findings in chitinase gene expression, where hypovirulence was associated with stronger host activation [123]. Transcriptomic analyses in C. dentata and C. mollissima also revealed increased SA-related gene expression in canker tissue, supporting its role in defense across species [128]. The literature identifies SA signaling activation as a multifaceted process that involves the induction of defense genes, cell death regulation, and antagonism/fine-tuning of JA-ET signaling. This activation is both a host defensive strategy and, in some cases, a target for pathogen manipulation [127,129].
Overall, C. sativa exhibits partial and often ineffective defense responses to C. parasitica. Defense suppression by the fungus may be a hallmark of C. sativa’s limited resistance compared to Asian chestnut species. The presence of hypovirulent strains offers some hope for disease mitigation, but the species remains vulnerable without external intervention or genetic improvement.
2.2.2.2. Castanea mollissima: Robust Genetic Resistance
C. mollissima has co-evolved with the pathogen in its native range, resulting in the development of genetically encoded resistance mechanisms. This species exhibits both constitutive and inducible defenses, including rapid wound response, cell wall lignification, and the activation of resistance genes upon infection. These traits are supported by genomic studies that have identified candidate genes associated with disease resistance, particularly in the sequenced genome of the cultivar ‘Vanuxem’, which has been widely used in restoration breeding programs in North America and Europe [130].
The reference genome of C. mollissima reveals a well-structured genetic architecture, with over 36000 gene models and extensive transcriptomic data. These resources have enabled the identification of selection signatures and resistance loci that differentiate C. mollissima from susceptible species like C. dentata and C. sativa [130]. Moreover, the species shows high genetic diversity across its natural populations, especially in regions like the Qinling-Daba Mountains, which are considered reservoirs of resistance genes [131].
Additionally, the complete chloroplast genome of wild C. mollissima has been sequenced, providing further insights into its evolutionary adaptations. This genome includes 131 genes, many of which are involved in stress responses and metabolic regulation. Phylogenetic analyses confirm its close relationship with other resistant Fagaceae species, reinforcing its role as a genetic donor in breeding programs [132].
2.2.2.3. Breeding and Genomic Efforts
The stark contrast between C. sativa and C. mollissima has driven interspecific hybridization efforts, aiming to introgress resistance traits from the latter into the former (described in 2.1.1).
However, blight is most severe on the American chestnut. A review by Fernandes et al. [3] outlined decades of efforts to combat chestnut blight in the US, recurring principally to crosses between C. dentata and the cornerstone of chestnut blight resistance, C. mollissima. Hybridization efforts began in the 1920s; although first-generation hybrids showed strong resistance, they lacked the tall growth typical of C. dentata. To recover these growth traits, backcrossing to C. dentata was pursued, based on the idea that a few major alleles from C. mollissima conferred resistance to C. parasitica and P. cinnamomi [45,133]. Advanced backcross hybrids resembled C. dentata and had improved resistance, but molecular studies revealed a tradeoff between blight resistance and C. dentata ancestry. This suggests that blight resistance is a multigenic and quantitative trait, as supported by genetic mapping that identified resistance loci on all 12 chromosomes [63,134].
Westbrook et al. [135] recently provided a comprehensive overview of US breeding programs aimed at developing self-sustaining populations of American chestnut with resistance or tolerance to both C. parasitica and P. cinnamomi. The authors employed an integrative approach combining genomics, transcriptomics, and statistical modeling, establishing a robust framework to accelerate restoration efforts through both conventional breeding and biotechnological strategies. Recognizing the potential of gene editing as a precise and rapid tool, they identified candidate genes associated with resistance and susceptibility, many of which co-localize with previously identified QTLs for blight resistance. To support this, new whole-genome sequences of C. dentata ‘Ellis’ and C. mollissima ‘Mahogany’ were generated and used in genome-wide association studies (GWAS). Among the findings, four genes, including a chitinase, were highlighted as potential resistance factors, while 26 genes were proposed as susceptibility candidates.
Transgenics represents a complementary strategy to the labor-intensive process of backcross breeding. Transgenic C. dentata expressing the wheat OxO gene have shown promising results and are currently undergoing regulatory review and field trials [136,137]. The introduced oxalate oxidase enzyme enables the degradation of oxalic acid secreted by C. parasitica, producing carbon dioxide and hydrogen peroxide. This reaction mitigates the damaging effects of the pathogen by reducing the severity of fungal-induced lesions [138]. The US Department of Agriculture Animal and Plant Health Inspection Service (USDA-APHIS) completed a draft review concluding that the transgenic American chestnut is unlikely to pose a plant pest risk [139]. Other US Federal agencies (Environmental Protection Agency; Food and Drug Administration) still need to complete their reviews. Nonetheless, the potential approval of this transgenic tree will represent a major milestone in the application of transgenesis for ecological conservation.
2.2.2.4. Transcriptomic Insights into Chestnut Blight Resistance
Barakat et al. [140] used high-throughput pyrosequencing for transcriptome comparison between blight-susceptible American chestnut and blight-resistant Chinese chestnut in response to infection by C. parasitica. Researchers sequenced RNA from both healthy and infected stem tissues, generating over a million reads and assembling tens of thousands of unigenes for each species.
Key findings include: a) A large number of genes associated with stress and biotic stimulus responses were identified, with many showing differential expression between healthy and infected tissues with canker; b) Defense-related genes such as transcription factors (e.g., WRKY, zinc finger, MYB), enzymes (e.g., cinnamoyl-CoA reductase, alpha-dioxygenase), and transporters were more highly expressed in canker tissues; c) C. mollissima exhibited a broader and more robust transcriptional response to infection, consistent with its higher resistance; d) Functional annotation revealed similar overall gene function distributions between species, but with subtle differences in categories like transcription factor activity and stress response.
Sequentially, Barakat et al. [128] deepen the findings in Barakat et al. [140]. Transcriptome comparative analysis revealed that chestnut genes share greater similarity with other woody plants than with herbaceous species. The study identified hundreds of genes with differential transcript abundance between cankered and healthy stem tissues. Many of these genes are involved in key defense-related pathways, including cell wall biosynthesis, ROS production, hormone signaling (e.g., SA, JA, ET, ABA), HR, and PCD. HR and PCD are generally associated with immune responses to biotrophic organisms. However, studies have consistently shown that necrotrophic pathogens exploit the dead tissue generated by HR, thereby using the host’s defense machinery to support their own life cycles [141]. Molecular studies further complicate the picture by demonstrating that HR and PCD are not universally coupled to resistance; genetic analyses have shown that HR and PCD components can be activated independently and that resistance against necrotrophic pathogens may sometimes be achieved with limited cell death [142].
Peroxidase activity may be involved in the host response to C. parasitica, with increased transcript levels observed in canker tissue of C. mollissima [128]. Peroxidases, namely the ones of Class III, are a group of secretory plant enzymes involved in the one-electron oxidation of various substrates, producing peroxides, and are strongly associated with induced pathogen defense mechanisms [121,143]. Beyond their defensive role, they also contribute to auxin metabolism, cell wall reinforcement, lignin and suberin synthesis, and the regulation of reactive oxygen and nitrogen species [144]. In Castanea species, four distinct peroxidases have been identified in stem and leaf tissues [145], though their enzymatic properties remain poorly characterized. Although the mentioned studies suggest that peroxidase activity may play a role in the host response to C. parasitica, conflicting findings have been reported: Castanea pumila Mill., a blight-susceptible species, showed higher peroxidase transcript abundance than the resistant C. mollissima [146], and infection was found to reduce peroxidase activity in both C. dentata and C. mollissima, with a smaller decrease in the latter [145]. These findings suggest that C. parasitica may suppress host peroxidase activity, particularly in susceptible species, but the precise role of peroxidases in chestnut blight resistance remains unclear due to the lack of follow-up studies.
The findings of Barakat et al. [128,140] provide valuable genomic resources and identify candidate genes and networks potentially responsible for host resistance to C. parasitica. Although the gene expression profiles activated in canker tissues by C. parasitica were largely similar between the two species, notable differences were observed: C. dentata exhibited increased activation of housekeeping genes, whereas C. mollissima showed a stronger induction of PR transcripts. The authors hypothesized that the difference in blight tolerance between C. dentata and C. mollissima may depend more on the speed of the host’s response to infection than on the presence of specific defense-related genes.
To better understand the molecular basis of blight resistance in C. mollissima, Nie et al. [147] performed transcriptomic profiling of the resistant wild genotype ‘HBY-1’ at 0, 3, and 9 h post-inoculation with C. parasitica, after Illumina sequencing. The analysis revealed that 9 h post-infection is a critical time point for defense activation. Two hundred and eighty-three differentially expressed genes (DEGs) were identified and grouped into two major functional clusters: a) Metabolism-related pathways, including biosynthesis of secondary metabolites, phenylpropanoid biosynthesis, and photosynthesis. b) Defense-related pathways, such as plant-pathogen interaction and MAPK signal transduction. These clusters were interconnected through signaling systems involving phosphatidylinositol, phytohormones, and α-linolenic acid metabolism. Notably, genes involved in JA biosynthesis and metabolism were significantly upregulated, indicating full activation of the JA pathway early in the infection process. The study concludes that C. mollissima mounts a rapid and coordinated defense involving hormone signaling, pathogen recognition, and metabolic reprogramming.
2.2.2.5. Metabolomic Insights into Chestnut Blight Resistance
The metabolomic dynamics governing the interaction between C. parasitica and Castanea species remain largely unexplored. The review by Lovat and Donnelly [113] provides a comprehensive analysis of the mechanisms and metabolites involved in the interaction between chestnuts and C. parasitica.
In the chestnut cell walls, compounds related to the lignin barrier and wound periderm involved in blight defense have been identified. Tannins, a diverse group of polyphenolic compounds including phenolic acids, flavonoids, and sugars, play key roles in chestnut species by contributing to plant defense, antioxidant activity, and structural integrity [148]. Their interaction with the pathogen C. parasitica is multifaceted: while fungal growth may be enhanced by tannin-rich media due to tannase activity [149,150], species-specific tannin profiles influence disease resistance. Asian chestnuts like C. mollissima contain higher levels of vescalagin and castalagin, which may inhibit fungal growth more effectively than hamamelitannin, prevalent in the susceptible C. dentata and C. sativa [113]. Histological and hormonal studies suggest that esterase activity and JA signaling also modulate tannin availability and response to infection [151,152]. Moreover, tannins may inhibit fungal enzymes such as polygalacturonase, further complicating their role in pathogenesis [153]. Overall, tannins are central to the host–pathogen dynamics in chestnut blight, acting both as potential substrates and modulators of fungal virulence. A synthesis of the strategies and key findings for understanding chestnut responses to C. parasitica is presented in Table 2.
2.2.3. Molecular Mechanisms of Castanea Defense Against Dryocosmus kuriphilus
The molecular interactions between the chestnut gall wasp and the European chestnut (and its hybrids) remain poorly investigated, particularly the genetic bases of host resistance and susceptibility. Compared with research on chestnut blight and ink disease, progress has been slower, likely due to the pest’s later introduction in Europe, its initially underestimated ecological and economic impacts, and the early reliance on biological control. Most studies have focused on evaluating and implementing a biological control using the parasitoid wasp Torymus sinensis Kamijo, which is considered the most effective strategy for reducing forest infestations. Biological control has been deployed in several countries, including Japan, Korea, Italy, and the US, yet infestations continue to impact harvests and the cynipid continues to spread in Europe [21,59,154,155]. However, orchard-level containment may benefit from the use of genotypes carrying resistance or reduced susceptibility.
Identifying resistant genotypes is essential to understanding the chestnut-pest interactions for breeding programs. Resistance to D. kuriphilus has been documented in the hybrid Bouche de Bétizac (C. sativa x C. crenata). Early studies reported no infestations for 3 years [156], and this resistance was later confirmed after nearly a decade of observations [59]. Although the wasp can still lay eggs in the buds, larvae fail to develop beyond the first instar, most likely due to HR triggered by the hybrid. Dini et al. [157] demonstrated this by using diaminobenzidine (DAB) staining to detect in vivo H2O2 accumulation, an indicator of the stress-response glycoproteins germin and germin-like proteins (GLPs). GLPs, which have OxO activity, are associated with PCD and HR. Positive DAB staining was observed in Bouche de Bétizac buds but not in those of the susceptible cultivar Madonna (C. sativa), regardless of infestation state. Furthermore, strong expression of a putative GLP was detected in Bouche de Bétizac tissues during early budburst, reinforcing the presence of HR. This response was also proposed in preliminary transcriptomic studies; using Differential Display analysis between Bouche de Bétizac and the susceptible cultivar Marrone (C. sativa), Botta et al. [158] identified differentially expressed bands with sequences putatively encoding for resistance genes, mitogen-activated proteins, vesicle-associated membrane proteins, and 14-3-3 proteins.
The molecular basis of resistant and susceptible responses to D. kuriphilus infestations was investigated by Acquadro et al. [159] using transcriptomic analysis of buds (pools of several gall development stages) from Bouche de Bétizac and Madonna. The two assemblies contained 34,081 and 30,605 unigenes, respectively. For downstream analyses, the Bouche de Bétizac unigene set was functionally characterized, whereas the Madonna assembly was used mainly for RNA-seq data analysis. This work identified 1,444 putative resistance gene analogs (RGAs) and approximately 1,135 unigenes predicted as miRNA targets. Global transcriptome profiling revealed significant Gene Ontology enrichments, particularly for response to stimulus and developmental processes (e.g., post-embryonic development). Among the up-regulated genes, several were associated with attack recognition, with approximately 60 predicted to encode leucine-rich repeat (LRR) proteins; others were related to transcriptional regulator activity, including 6 APETALA2/Ethylene (AP2/ERF) and 16 WRKY (e.g., WRKY33). A putative homolog to RAV1 transcription factor was also upregulated; beyond its proposed role as a negative regulator of growth and development, Acquadro et al. (2020) [159] suggested that RAV1 regulation may also serve as a developmental adaptation to gall-induced stimuli. Additional examples of upregulated genes included protein regulators (e.g., Regulatory-Associated Protein of TOR 1b; RAPTOR1B), storage proteins (e.g., Late Embryogenesis Abundant protein D29; LEA D29), and more than 100 genes linked to death and apoptosis processes, including those involved in HR response [159]. These findings further support the HR-based mechanism previously suggested by Dini et al. [157]. In addition to clarifying molecular responses, the study produced valuable resources, including the first reference unigene catalog for the European chestnut, along with ˜7k SSR and 335k SNP/INDEL markers.
Another breakthrough in understanding the genetic resistance of Bouche de Bétizac came from high-density mapping in interspecific hybrids. A large-effect QTL, Rdk1, was mapped to linkage group K, explaining 67–69 % of the phenotypic variance [61]. Within this region, 26 candidate genes were identified, including two of particular interest - metacaspase-1b and a receptor of the Resistance to Peronospora parasitica 13 locus (RPP13) subfamily. Both genes are known to be involved in HR.
In addition to Bouche de Bétizac, Sartor et al. [59] identified six other cultivars resistant to D. kuriphilus, including two C. sativa cultivars: the Italian ‘Pugnenga’ and the French ‘Savoye’. This finding is particularly significant, as it suggests the possibility of transmitting resistant traits within the species. More recently, resistance was detected in natural populations of C. sativa in Greece [62]. A genome-wide association study provided the first genomic insights into this resistance, revealing a small region on pseudochromosome 3 (Chr3) associated with the high resistance observed in Greek provenances. This region harbors 12 candidate genes, including members of the Cytochrome P450, UDP-glycosyltransferase, and Rac-like GTP-binding protein families. Notably, 21 SNPs within this region were identified, offering promising markers for MAS in breeding programs.
Insights into resistant and susceptible responses of other chestnut species to D. kuriphilus can provide valuable guidance for European chestnut research and breeding programs. Zhu et al. [160] suggested that the peroxidase pathway may contribute to resistance in a partially resistant Chinese chestnut variety, and identified 4 transcription factors as potential players in this pathway (CmbHLH130, CmWRKY31, CmNAC50, and CmPHL12).
Although further research is needed to fully clarify the genetic basis of chestnut responses to gall wasp, current evidence consistently highlights oxidative stress signaling as a central mechanism. Several candidate resistance genes have already been identified, providing a strong foundation for future functional studies. Moreover, the recent publication of a high-quality reference genome for D. kuriphilus [161] opens new opportunities to investigate the pest’s molecular weapons and the associated host gene expression changes during infestation. Such insights will enable a more comprehensive understanding of chestnut-gall wasp interactions and inform the development of integrated management strategies (beyond the ongoing biological control with T. sinensis) to mitigate the impacts of this invasive cynipid. Table 3 provides an overview of the strategies used to understand chestnut’s resistance to chestnut gall wasp.
2.2.4. Whole Genome Sequencing
Whole genome sequencing (WGS) has become a cornerstone in advancing molecular knowledge of defense mechanisms in chestnut species, particularly in response to pathogens, pests, and climate change. By providing comprehensive insights into the genetic architecture of chestnut trees, WGS enables the identification of resistance-associated genes and regulatory elements that govern responses to biotic and abiotic stressors.
Genome assemblies differing in completeness are now available for the four chestnut species. For C. sativa, two assemblies have been published: one for the cultivar ‘Marrone di Chiusa Pesio’ using Oxford Nanopore and Illumina technologies [162] and another for the Anatolian cultivar ‘Sarı Aşılama’ [163]. The genome of C. mollissima has been sequenced for multiple cultivars, including ‘Vanuxem’, a donor of blight resistance in C. dentata restoration breeding [130]. For C. crenata, a chromosome-level genome assembly has revealed conserved chromosomal segments and a large repertoire of protein-coding genes, supporting its known resistance to diseases and pests [164].
These genomic resources facilitate comparative analyses that identify candidate genes under selection and enable MAS for resistance traits. WGS also supports landscape genomics approaches, mapping adaptive genetic variation across environmental gradients and predicting genomic offset under future climate scenarios [165]. Furthermore, genome-enabled breeding programs are incorporating adaptive diversity into backcross populations, ensuring that restored American chestnut trees are not only disease-resistant but also ecologically viable across diverse habitats [135].
Table 3.
Biotechnological strategies to understand the European chestnut’s resistance to Dryocosmus kuriphilus.
Table 3.
Biotechnological strategies to understand the European chestnut’s resistance to Dryocosmus kuriphilus.
| Methodology | Species / Genotypes | Results / Findings | References |
|---|---|---|---|
|
Biological control Torymus sinensis |
Various (wild and cultivated) |
Effective in reducing infestations, but pest continues to spread | [21,154,155] |
| Phenotypic resistance screening |
C. sativa, C. crenata and Euro japanese hybrids |
7 resistant cultivars identified: C. sativa ‘Pugnenga’ & ‘Savoye’; C. crenata ‘Idae’; Hybrids ‘BB’, ‘Marlhac’, Maridonne’, ‘Vignols’ | [59,156] |
|
Histochemistry and gene expression |
‘BB’ (R) vs. ‘Madonna’ (C. sativa, S) |
Detection of H2O2 accumulation and strong GLP expression in R hybrid linked to HR | [157] |
| RNA-seq transcriptome analysis | ‘BB’ (R) vs. ‘Madonna’ (C. sativa, S) |
1,444 RGAs, 1,135 miRNA targets; upregulation of LRRs, WRKYs, AP2/ERFs, RAV1, LEA D29, RAPTOR1B; HR-related genes | [159] |
|
C. mollissima ‘Shuhe Wuyingli’ (PR) vs. ‘HongLi’ (S) |
Peroxidase pathway implicated; 4 TFs identified (CmbHLH130, CmWRKY31, CmNAC50, CmPHL12) | [160] | |
|
Genomic resources development |
C. sativa | Reference unigene catalog; ~7k SSRs and 335k SNP/INDELs | [159] |
| QTL mapping | Interspecific hybrids ‘BB’ x ‘Madonna’ |
Rdk1 locus explains 67–69% of resistance variance; candidate genes include metacaspase-1b and RPP13 receptor | [61] |
| GWAS | Greek C. sativa provenances (R) |
Region on Chr3 with 12 candidate genes (Cytochrome P450, UDP-GT, Rac-like GTPases); 21 SNPs identified | [62] |
| Genome sequencing | D. kuriphilus (pathogen) | High-quality reference genome published; enables host-pest interaction studies | [161] |
BB: ‘Bouche de Bétizac’; GLP: germin-like protein; GWAS: genome-wide association study; HR: hypersensitive response; INDEL: insertion deletion; PR: partially resistant; QTL: quantitative trait loci; R: resistant; RGAs: resistance gene analogs; S: susceptible; SNP: single nucleotide polymorphism; SSR: simple sequence repeat; TF: transcription factor.
Importantly, WGS provides the foundation for genome editing technologies, such as CRISPR/Cas9, by enabling precise identification of target genes and regulatory sequences. In C. sativa, the first successful genome editing using CRISPR/Cas9 [166] is detailed in 2.4.2.1, demonstrating editing efficiency in protoplasts derived from somatic embryos. This DNA-free approach avoids transgene integration, offering a promising path for developing improved varieties without genetically modified organisms (GMOs) classification. The availability of reference genomes significantly enhances the accuracy and efficiency of such editing methodologies, allowing for targeted modifications that improve disease resistance, stress tolerance, and other agronomic traits.
In summary, WGS provides a foundational platform for integrating molecular genetics with ecological and evolutionary frameworks, accelerating both traditional breeding and cutting-edge genome editing approaches to develop resilient chestnut populations capable of withstanding current and emerging challenges.
2.3. Micropropagation Techniques
2.3.1. Axillary Budding Micropropagation
The initial objective of micropropagation techniques in European chestnut was to develop efficient systems for clonal propagation through axillary budding culture, using trees previously selected for their tolerance to P. cinnamomi. This method was conceived as a complementary alternative to the traditional propagation system based on ground-layering. European chestnut is a recalcitrant species, particularly with respect to rooting, a trait that persists even under in vitro conditions. To overcome these limitations, Spanish researchers initially optimized their methodology using juvenile plant material, which enabled the establishment of a reliable micropropagation system [167]. This protocol was later adapted for use with mature material, which in some cases required rejuvenation techniques (i.e., etiolation or repeated grafting) to elicit a suitable in vitro response [168,169,170]. Later on, numerous studies and reviews have been published in specialized journals on the in vitro propagation of European chestnut and its hybrids via axillary bud micropropagation [4,171,172,173,174,175]. Nowadays, micropropagation by axillary budding is an essential and strategic tool for the rapid, safe, and efficient cloning of superior chestnut genotypes with enhanced P. cinnamomi tolerance selected from European breeding programs. In Spain, a robust protocol that allows the efficient cloning of any European chestnut genotype was validated and transferred to the private sector through appropriate agreements, facilitating its application in commercial plant production [176]. For example, TRAGSA Company produced 40,000 plants in the most recent campaign, the first to be completed in the new laboratory, with the goal of testing the market’s ability to absorb increased volumes. For the upcoming campaign, which is already underway, production is expected to rise to 50,000–60,000 plants. In 2025, the company plans to scale up production to 100,000 hybrid chestnut plants, intended for use as rootstocks. Meanwhile, CULTIGAR Company has produced an average of 58,000 plants per year over the past few seasons. In Portugal, micropropagation has played a critical role in producing homogeneous experimental material, fundamental for physiological, molecular, and pathogenicity assays conducted with high technical rigor [57,93,95] and in the efficient multiplication of rootstocks tolerant to P. cinnamomi [49](return to 2.1.1. for details on registered new chestnut varieties). Adapted for the transition from the laboratory to the field, a pilot facility was established in the North Alentejo region of Portugal, Marvão, equipped to produce thousands of clonal plants, ensuring uniformity, traceability, and phytosanitary quality. The location was chosen due to its chestnut-growing tradition and optimal edaphoclimatic conditions, serving as a demonstration and dissemination platform. Tolerance to P. cinnamomi observed under controlled conditions requires validation in the field. Experimental field trials were implemented to test the performance of new genotypes, evaluating parameters such as vegetative vigor, survival rate, productivity, and health over time. These results are crucial for assessing the commercial viability of new materials and informing technical recommendations to producers. In addition to disease tolerance, ensuring graft compatibility with traditional, commercially valued chestnut varieties is essential. Graft compatibility studies are being conducted to evaluate rootstock–scion interactions, considering factors such as affinity, development, precocity, and fruit quality. This approach protects regional genetic heritage while introducing resilience into production systems.
Factors such as plant vigor, rooting uniformity, and acclimatization performance were significantly improved, marking a major step toward the large-scale, energy-efficient production of elite European chestnut genotypes. A recent advancement is the use of LED-based systems, which have emerged as powerful tools to further enhance chestnut micropropagation. In this line, Marino et al. [177] demonstrated that specific LED spectra substantially improved both the multiplication and rooting phases, doubling the proliferation index and achieving 100% rooting success compared to conventional fluorescent lighting.
2.3.2. Somatic Embryogenesis
Somatic embryogenesis (SE) has also been established in European chestnut and its hybrids. However, these procedures cannot yet be applied at a commercial level. Despite this, they have been extremely useful for developing cryopreservation, genetic transformation, and gene editing methods in this species. The first well-documented report of true somatic embryo induction and subsequent plant regeneration from immature zygotic embryos of Castanea sativa × C. crenata hybrids was published by Vieitez et al. [178]. Later studies confirmed somatic embryogenesis in both hybrid (C. sativa × C. crenata) and pure C. sativa genotypes [179,180,181,182,183]. Somatic embryo induction typically involves a two-step culture protocol: an initial induction phase using auxins in high concentration, followed by an expression medium with either low concentrations of PGRs or none at all. Among auxins, 2,4-dichlorophenoxyacetic acid (2,4-D) is most commonly used and considered essential for initiating embryogenic cultures. A cytokinin, usually benzyladenine (BA), kinetin, or zeatin, is often added at lower concentrations to support embryo development. Particulary, Sezgin and Dumanoğlu (2014) [181] successfully used indole-3-butyric acid (IBA) in combination with thidiazuron (TDZ), as an alternative induction treatment. The developmental stage of the zygotic embryo, closely tied to the collection time (weeks post-anthesis), strongly affects induction efficiency. The optimal window for harvesting embryos in northwest Spain ranges from 6 to 12 weeks post-anthesis (late July to early September), when immature zygotic embryos are most competent for somatic embryogenesis. Both whole zygotic embryos and cotyledonary segments have been used as explants. However, comparative studies indicate that embryonic axes exhibit twice the induction efficiency of cotyledonary pieces [179]. Moreover, the induction percentages were influenced by both the genotype of the mother tree and collection date, with values ranging from 2% for the hybrid material [178,183] to 11.1% for C. sativa material [179,182].
To date, SE from explants other than zygotic embryos in chestnut species has only been successfully achieved in European chestnut by Corredoira et al. [179,184]. Somatic embryos were induced from the most apical leaves excised from proliferating shoot cultures, using a three-step culture protocol. In the first step, explants were cultured on medium supplemented with 4 mg/L naphthaleneacetic acid (NAA) and 0.5 mg/L BA. This was followed by subculturing onto a medium containing reduced concentrations of both PGRs at 0.1 mg/L, and finally transferring to a PGR-free expression medium. Notably, 2,4-D and IBA regulators, commonly effective in the SE induction from zygotic embryos, were ineffective in inducing embryogenesis from non-zygotic-derived explants. Induction rates were considerably lower than those obtained from zygotic embryos, with frequencies not exceeding 1% [179]. However, subsequent studies significantly increased induction frequencies through the incorporation of 2 mg/L larch wood extract, a compound rich in arabinogalactan proteins (AGPs), resulting in SE induction rates of up to 5.3%—the highest reported to date for non-zygotic explants in European chestnut [185].
Another critical challenge in the application of SE in European chestnut, as in many other woody species, is achieving efficient plant regeneration, defined as the simultaneous development of both shoot and root systems from somatic embryos. The successful maturation and conversion of somatic embryos into viable plantlets are influenced by several factors, including the type of osmotic agents used, pre-germination treatments, and the application of PGRs [35]. In the European chestnut, the source and concentration of carbon are particularly important. Among the tested sugars, 3% maltose yielded the best results, with an overall plant recovery rate of 39%, including 6% of embryos achieving full plantlet conversion and 33% showing shoot development only [184]. In addition, increasing the agar concentration to 1.1% in the maturation medium significantly improved embryo maturation and plantlet conversion rates (10–25%) in embryogenic lines derived from zygotic embryos [180]. In contrast, ABA, which is commonly used to promote maturation in other embryogenic systems, proved largely ineffective in chestnut, showing no significant improvement in plantled development or conversion [183]. Direct transfer of somatic embryos from maturation to germination media often results in poor germination and abnormal plantlet development. Therefore, pre-germination treatments such as cold storage, desiccation, and the application of gibberellic acid (GA3) are typically required to improve conversion outcomes [35]. In European chestnut, application of cold storage of 2 months somatic embryos provided conversion rates of up to 38.9% in C. sativa lines [184] and 29–32% in hybrid material [183,186]. Desiccation, which mimics the natural drying phase in seed development, has also proven beneficial. Fast desiccation by exposing embryos for 2 h in a laminar flow hood reduced moisture content to approximately 57%, resulting in significantly enhanced plant quality [187]. Similarly, Sezgin & Dumanoğlu [182] achieved 40% plant regeneration using cold storage followed by slow desiccation, applying a desiccator with saturated salt solution. In the germination step, somatic embryos are generally cultured on germination media, often supplemented with low concentrations of cytokinins alone or in combination with auxins. For instance, the inclusion of 0.1 mg/l BA together with 0.1 m/l NAA or 0.1 mg/l IBA yielded the highest conversion rates [181,187]. Further improvements in plantlet development were achieved by adding 200–438 mg/L glutamine or 150 µM Fe-Na-EDTA to the germination medium [183,187]. Despite these advancements, plant regeneration remains a bottleneck in chestnut SE protocols, particularly due to the asynchronous or incomplete development of shoots and roots, underscoring the need for further optimization of both culture conditions and genotype-specific responses.
2.4. Genetic Engineering Strategies
One of the greatest advances in plant breeding has been achieving modified plants through genetic transformation by inserting specific genes into the plant genome. A fundamental premise for success in the production of transgenic plants is the availability of an in vitro regeneration system adequate to support the production of plants from cells, organs, or tissues susceptible to infection by Agrobacterium tumefaciens. Traditional genetic transformation and modern genome editing technologies have been employed in European chestnut for research and breeding purposes using somatic embryogenesis as regeneration system. Remarkably, it stands out as one of the first forest tree species in which gene editing has been successfully implemented, representing a significant advancement in the genetic improvement of woody plants.
2.4.1. Traditional Genetic Transformation
The first attempt at genetic transformation in European chestnut was conducted by Seabra and Pais [188], using Agrobacterium-mediated transformation of hypocotyl segments excised from in vitro-germinated seedlings. The bacterial strain LBA 4404 was employed, carrying the binary vector p35SGUSINT, which includes the nptII gene (conferring kanamycin (Kan) resistance) and the uidA (GUS) as a reporter gene, allowing detection of transgene expression through β-glucuronidase activity assays. Although transgenic shoots were regenerated, molecular analyses revealed that many were chimeric, indicating incomplete or partial transformation events. Successful stable genetic transformation was later achieved using somatic embryos as target explants, employing the same marker genes nptII and uidA [189,190]. In the initial transformation experiments, the influence of Agrobacterium strain/plasmid combinations and co-cultivation duration (3 vs. 4 days) was assessed [189]. After 12 weeks on selective medium, 112 out of 624 explants were kanamycin-resistant. Transformation efficiency, defined as the percentage of explants producing GUS-positive embryogenic cultures, was significantly affected by both the bacterial strain/plasmid and co-culture period. The highest transformation efficiency (25%) was obtained using strain EHA105 harboring plasmid pUbiGUSINT after 4 days of co-cultivation. Subsequently, the same authors investigated the effects of acetosyringone addition, bacterial density, genotype, and the developmental stage of somatic embryos on transformation efficiency [190,191]. Contrary to expectations, acetosyringone at both 100 µM and 200 µM negatively impacted transformation efficiency. All tested bacterial densities yielded kanamycin-resistant embryos, with the highest transformation efficiency (20.1%) observed when Agrobacterium was in the exponential growth phase (OD600 ≈ 0.6); however, the differences between densities were not statistically significant. Both the developmental stage of the somatic embryos and the genotype had significant impacts on transformation success. Globular and heart-shaped embryos, as well as embryo clumps, exhibited higher transformation frequencies (up to 30%) compared to cotyledonary-stage embryos, likely due to their higher proliferation potential and a greater proportion of actively embryogenic cells [192]. Genotype-dependent variability was also evident: among seven lines tested, two showed relatively high transformation efficiencies (21.7% and 33.8%), while the others ranged between 1.7% and 10%. Molecular analyses confirmed transgene integration, with all GUS-positive lines testing PCR-positive for uidA and nptII. Southern blot analysis further verified uidA presence in transgenic lines, but not in untransformed controls.
Once an efficient transformation method had been established, the next step was to introduce genes with the potential to enhance disease resistance. To this end, two PR proteins—a thaumatin-like protein (CsTL1)[193] and a chitinase (CsCH3)[125], which had been purified from mature cotyledons of European chestnut and shown to exhibit significant antifungal activity in vitro, were selected for gene overexpression (section 2.2.2.1). Somatic embryos were transformed using A. tumefaciens strain EHA105 carrying a binary plasmid containing the CsTL1 gene, the nptII selectable marker, and the egfp reporter gene [194]. The CsTL1 gene was successfully overexpressed in three somatic embryogenic lines, with transformation efficiencies ranging from 7.1% to 32.5%, resulting in the generation of 126 independent transgenic lines. Building on this work, an efficient Agrobacterium-mediated protocol was later developed to introduce the endogenous CsCH3 gene into somatic embryos, further advancing efforts to enhance disease resistance in European chestnut [126]. A total of 88 independent chitinase transgenic lines were obtained. In both studies, stable integration of CsTL1 and CsCH3 genes into the genome was confirmed by PCR and Southern blot analysis. qPCR further showed that CsTL1 expression was up to 13.5-fold higher in one transgenic line compared to its untransformed counterpart. GFP fluorescence, indicative of transgene activity, was observed not only in somatic embryos but also in shoots, leaves, and roots of transgenic plants. Importantly, no phenotypic differences were detected between transgenic and control plants, suggesting that GFP expression and transgene integration did not produce any pleiotropic effects. These molecular and phenotypic results support the potential of CsTL1 and CsCH3 as candidate genes for engineering resistance to ink disease in European chestnut. Nevertheless, further functional validation under greenhouse and field conditions is essential to confirm the effectiveness of these transgenes against Phytophthora sp and/or C. parasitica infection in planta.
Using the same transformation system, twelve independent transgenic European chestnut lines were obtained carrying the already mentioned Cast_Gnk2-like gene (section 2.2.1), which encodes a Gnk2-like antifungal protein. Transgene copy number and relative expression were analyzed by qPCR in lines with high proliferation capacity [101]. Tolerance assays against P. cinnamomi revealed that one transgenic line exhibited significantly improved tolerance compared to the non-transformed control, as evidenced by reduced root necrosis and consequently fewer disease symptoms [195].
Adapted systems of genetic transformation of somatic embryos and plant regeneration have been applied to Cast_Gnk2-like functional validation in C. dentata [101,104], Quercus ilex [102] and Q. suber [103]. Tolerance assays to P. cinnamomi are ongoing for C. dentata, while oak plantlets overexpressing Cast_Gnk2-like potentially exhibit improved tolerance to P. cinnamomi, compared to non-transformed plantlets. As oaks belong to the same family as chestnuts and both are susceptible to P. cinnamomi, these results are a promise for Fagaceae improvement. To further support Cast_Gnk2-like as a gene conferring tolerance to P. cinnamomi, the recombinant protein expressed in Escherichia coli showed activity against P. cinnamomi in laboratory pathogenicity tests, validating its potential as a protective agent and opening perspectives for the use of bioactive compounds in integrated disease management [196]. These approaches demonstrate the utility of genetic engineering for functional validation, a deeper understanding of resistance mechanisms, and the development of innovative plant protection tools.
Plant regeneration, defined as the development of both shoot and root from transgenic somatic embryos, was consistently low, irrespective of genotype. These low conversion rates significantly limit the number of plantlets available for downstream analyses. However, some somatic embryos undergo partial germination, producing shoots without roots. These shoots can be leveraged through axillary shoot proliferation and subsequently induced to root, allowing for the propagation of an unlimited number of transgenic plantlets [197]. This method provides sufficient plant material for essential molecular validation and for performing disease resistance assays. Although improving the direct conversion efficiency of somatic embryos into complete plantlets remains a key objective, axillary shoot culture currently stands as the only reliable and scalable strategy for multiplying transgenic chestnut lines for research and functional evaluation.
2.4.2. New Plant Breeding Techniques
New Plant Breeding Techniques (NPBTs) are modern biotechnological tools that enable precise modification of the plant’s DNA without introducing genes from other species. Unlike conventional breeding, which requires multiple cycles of crossing and selection, NPBTs can directly modify elite cultivars, reducing breeding timelines and minimizing the risk of losing valuable traits during hybridization [198]. This is particularly relevant for woody species, characterized by long generation times and high heterozygosity level.
Among NPBTs, the CRISPR/Cas9 (Clustered Regularly Interspaced Short Palindromic Repeats-Cas9) system is the most widely applied genome editing tool in plant biotechnology, thanks to its versatility and the ability to target multiple genes simultaneously [199]. The system takes advantage of a guide RNA (gRNA) to direct the Cas9 nuclease to a specific DNA sequence, where it generates a double-strand break (DSB). The plant cell repairs this break mainly through non-homologous end joining (NHEJ), which often introduces small insertions or deletions that disrupt the gene function, or less frequently, through homology-directed repair (HDR), which can introduce precise sequence changes if a donor DNA template is provided [200].
In woody species, CRISPR/Cas9 applications are still limited [201]. As in the Traditional transformation, the main challenge is plant regeneration: edited cells must be able to develop a whole plant, a process that is often inefficient and highly genotype-dependent [174,177]. In C. sativa, SE remains the most effective regeneration system, offering higher transformation efficiency and reducing the occurrence of chimeric plants [35,179].
An advancement over standard CRISPR/Cas9 delivery is DNA-free genome editing using ribonucleoprotein (RNP) complexes, in which purified Cas9 protein is pre-assembled with its gRNA and introduced directly into protoplasts, plant cells deprived of the cell wall [202]. In conventional vector-based systems, Cas9 can remain active for a long time, increasing the risks of off-target mutations. In contrast, RNPs act only for a short time before being degraded by the cell, limiting nuclease exposure and avoiding the stable integration of foreign DNA [198]. This non-integrative approach enables the generation of edited plants completely free of recombinant DNA, opening new opportunities for breeding and potentially improving consumer acceptance compared with traditional GMOs. RNPs can be delivered by particle bombardment, protoplast electroporation, or polyethylene glycol (PEG)-mediated uptake but their application in woody crops remains limited. The main challenge is that efficient delivery must be combined with efficient regeneration from edited cells, which continues to represent a major bottleneck for C. sativa and many other woody species [203].
2.4.2.1. CRISPR/Cas9 Genome Editing in Castanea Sativa
The first successful application of CRISPR/Cas9 in European chestnut was reported by Pavese et al. [204] as a proof-of-concept for targeted mutagenesis in this recalcitrant woody species. The phytoene desaturase (pds) gene, involved in carotenoid biosynthesis, was selected as it serves as a visible marker, producing an easily recognizable albino phenotype caused by carotenoid loss and chlorophyll degradation. Two single-guide RNAs (sgRNAs) were designed to target conserved exonic regions of pds and assembled into a binary vector containing Cas9 and the nptII selectable marker for Kan resistance. Transformation was performed on somatic embryos at globular and torpedo stages, produced according to established protocols [173,179], using A. tumefaciens strain EHA105. Following co-cultivation, embryos were cultured on Kan-containing medium to select the Kan-resistant ones. Four kan-resistant embryogenic lines were recovered, and sequencing of the target region confirmed the editing event and the presence of small insertions and deletions. Regenerated plantlets from edited lines displayed the expected albino phenotype, providing direct phenotypic validation of pds knockout. This study demonstrated that CRISPR/Cas9 can be applied in C. sativa using SE as starting material. More recently, this system has been extended to genes of agronomic relevance. In particular, susceptibility genes are being targeted to improve tolerance to P. cinnamomi [107], with the goal of developing chestnut genotypes with enhanced disease resistance, improved nut quality, and better stress tolerance.
2.4.2.2. DNA-Free Genome Editing Using Ribonucleoproteins (RNPs)
Pavese et al. (2022) [166] realized the first protocol for DNA-free genome editing in European chestnut, using pre-assembled Cas9–sgRNA ribonucleoprotein (RNP) complexes delivered into protoplasts derived from somatic embryos. High yields of viable protoplasts (~4.5 × 106/mL with 91% viability) were obtained using enzymatic digestion with cellulase (1%) and macerozyme (0.5%) for 4 h of incubation. Transfection efficiency was confirmed using a GFP marker, with ~51% of protoplasts expressing GFP 72 h post-transfection. Finally, RNPs targeting pds were directly delivered, and sequencing confirmed successful editing of the target gene. This work demonstrated that RNP-mediated editing is feasible in C. sativa and provides an important step toward precise, DNA-free editing in chestnut. The main limitation is still the regeneration of complete plants. To move from edited protoplasts to improved chestnut genotypes, RNP delivery must be combined with efficient somatic embryogenesis, which is crucial for developing traits such as pathogen resistance and stress tolerance.
2.5. Germplasm Conservation Through Cryopreservation
Cryopreservation is a technique that allows for the long-term conservation of biological tissues—such as embryos, seeds, shoot tips, or pollen at ultra-low temperatures, typically in liquid nitrogen (-196 °C, LN) [205]. At this temperature, all cellular metabolic activity comes to a complete halt, enabling the indefinite preservation of biological material. This approach is a crucial tool for conserving genetic diversity, particularly in species with recalcitrant seeds that are sensitive to extended storage periods, as is the case with chestnut species. In European chestnut, cryopreservation serves as a valuable complement to traditional field collections for the long-term conservation of germplasm, and it has been successfully applied to a wide range of materials, including zygotic embryos, somatic embryos, and axillary shoot tips excised from shoots maintained in vitro [171].
Remarkably, Corredoira et al. [206] developed a highly efficient protocol for the cryopreservation of zygotic embryos. This study involved isolating embryonic axes and subjecting them to various desiccation periods (ranging from 0 to 7 h) prior to immersion in LN. Moisture content was found to be a critical factor influencing post-cryopreservation survival. At the time of isolation, the embryonic axes had a moisture content of 65.7% (based on fresh weight), which progressively decreased to 18% after 7 h of desiccation in the laminar flow hood. The highest survival rate (100%) and the best plant regeneration rate (63%) were achieved when the moisture content was approximately 20%, reached after 5 h of desiccation. Later, Gaidamashvili et al. [207] achieved a comparable whole-plant regeneration rate of 64% using an encapsulation with activated charcoal-vitrification protocol, which is a significantly more complex and labor-intensive procedure.
Significant progress has been made in recent years in the cryopreservation of somatic embryos in woody species, including European chestnut [205]. Vitrification-based cryopreservation has been successfully applied to chestnut somatic embryos using a protocol that combines a 3-day preculture on 0.3 M sucrose medium, treatment with Plant Vitrification Solution 2 (PVS2)[208] for 60 min at 0 °C, and rapid immersion in LN. Embryo clumps at the globular or heart stage achieved a 68% recovery rate, which remained stable even after 10 years of cryostorage (Corredoira, unpublished data), confirming the long-term reliability of the method. Moreover, this approach was successfully applied to the cryopreservation of transgenic embryogenic lines carrying a thaumatin-like protein gene [194] and a chitinase gene [126], underscoring its value for preserving genetically modified material during tolerance trials. Desiccation-based cryopreservation has been investigated as an alternative to vitrification for chestnut somatic embryos; however, it has produced poorer results. While vitrification methods obtained embryo recovery rates ranging from 68% to 92%, desiccation protocols resulted in only about 33% recovery. This highlights the greater efficiency of vitrification-based techniques for cryopreserving chestnut embryogenic tissues [194,206].
Shoot tips isolated from in vitro shoot cultures were also cryopreserved following the optimization of several parameters, including explant type, PVS2 exposure duration, sucrose concentration in the preculture medium, loading pre-treatment, and the sequence of recovery media used after thawing [209]. The most effective shoot regrowth protocol utilized 0.5–1.0 mm shoot tips that had been cold-hardened for two weeks. After a two-day preculture on medium supplemented with 0.2 M sucrose, the shoot tips underwent a two-step vitrification procedure: first, 20 min at room temperature in a loading solution (2 M glycerol + 0.4 M sucrose), followed by 120 min at 0 °C in a modified PVS2 solution, before rapid immersion in LN. This protocol resulted in 38–54% shoot recovery across five chestnut clones (of both juvenile and mature origin), with successful plant regeneration achieved in all cases. Later on, this protocol enabled, for the first time, the establishment of a European chestnut germplasm bank, in collaboration with the public company TRAGSA, aimed at conserving P. cinnamomi-tolerant genotypes [210]. Of the 46 chestnut genotypes tested, 43 survived cryopreservation, although only 63% retained their capacity to regenerate new shoots.
3. Conclusions and Future Perspectives
Climate change affects European chestnut populations by altering their phenology, increasing drought and heat susceptibility, which impacts growth, fruit quality, and survival. Warmer and wetter conditions also increase the spread of diseases and pests, increasing tree vulnerability. In this context, technologies such as traditional genetic transformation, omics-based approaches, gene editing (e.g., CRISPR/Cas9), in vitro culture and cryopreservation provide a powerful and complementary toolkit for modern breeding. These biotechnological tools enable the precise introduction of disease resistance, characterization of genetic diversity, conservation and the targeted enhancement of key agronomic traits, ultimately contributing to the development of more resilient, productive, and sustainable European chestnut cultivars.
Micropropagation through axillary bud proliferation is a well-established technique, widely applied at the commercial level by numerous companies. However, the development of embryogenic systems from non-zygotic material, particularly that derived from mature genotypes, will require considerable efforts in the coming years to achieve optimization. Over the years, development of somatic embryogenic protocols has been refined through empirical approaches, involving the testing of various culture media, growth conditions, donor plant ages, and explant types. However, to further advance in vitro plant cell reprogramming and regeneration, new strategies are needed to identify novel chemical promoters that can enhance these processes [211]. Although European chestnut is one of the few species for which efficient cryopreservation protocols have been developed for various tissues, its practical application remains very limited. Given the current context of climate change and the pressing need to preserve biodiversity, efforts to establish cryobanks should be strongly promoted, especially by public institutions.
Traditional Agrobacterium-mediated transformation has been an effective tool for introducing genes into embryogenic tissues, enabling traits such as disease resistance and stress tolerance. More recently, gene editing technologies like CRISPR/Cas9 have enabled precise modifications of endogenous genes without the need to insert foreign DNA, offering a more targeted and socially acceptable alternative. Although still in the early stages of application in chestnut, gene editing holds great promise for accelerating the development of improved genotypes, particularly in addressing diseases. Looking ahead, the availability of both traditional transformation and genome editing procedures provides a comprehensive platform for the genetic breeding of European chestnut, paving the way for more resilient and sustainable cultivation systems. On the other hand, both tools are convenient for functional analysis and validation of genes of interest.
The advances in omics technologies particularly genomics, transcriptomics, and metabolomics have significantly deepened our understanding of European chestnut, enabling the identification of key genes associated with important agronomic traits such as disease resistance, fruit quality, and adaptation to abiotic stress. An integrative approach combining transcriptomics, proteomics, and metabolomics would underscore the multifaceted nature of the defense response and highlight specific molecular deficits in susceptible chestnut species that may be targeted for improvement. Besides the contribution to ongoing efforts in MAS for chestnut controlled and expedited breeding, the expanding repository of genetic and functional information creates new opportunities for applying gene editing tools with greater precision and efficiency, targeting multiple well-characterized genes. Consequently, genome editing approaches that combine resistance allele stacking in breeding strategies with the modification of susceptibility genes will accelerate the development of elite chestnut varieties adapted to the challenges of climate change, with enhanced biotic stress resilience, productivity, and sustainability. This synergy between biotechnology and omics science positions the European chestnut at the forefront of a new era of applied genomic innovation.
New chestnut varieties that result from the improvement programs to pests and diseases need to be tested on susceptibility to climatic extremes (i.e., heat waves, prolonged droughts, and late frosts) to evaluate genotype resilience in the context of climate change projections.
Taken together, the studies here reported illustrate how molecular biotechnology approaches from population-level SSR diversity analysis to transcriptome-based marker development and gene expression profiling are converging to advance chestnut research. By linking insights into genetic diversity, molecular markers, and defence mechanisms, they pave the way for more informed strategies to conserve unique germplasm and breed cultivars resilient to diseases and pests.
The integration of science, technology, and commercial application is essential to address the challenges facing chestnut cultivation in the 21st century. The research programs on chestnut conducted in Portugal, Spain, Italy, and the US (from the development of more tolerant genotypes to their validation and large-scale production in real-world conditions) exemplify knowledge transfer with direct impact on agricultural sustainability, the valorization of mountain territories, and the preservation of an emblematic resource of rural landscape and culture. These programs highlight the critical role of applied research as a driver of innovation, regional development, and climate adaptation.
Author Contributions
Conceptualization, E. Corredoira, R. Costa, P. Fernandes, A. Martín, A. Moglia and S. Serrazina; investigation, M.T. Martínez, P. Piñeiro, B. Cuenca, M. Fraga, V. Pavese, C. Mattioni, S. Serrazina, and P. Fernandes; graphics, V. Pavese, M.T. Martínez and P. Piñeiro; writing—original draft preparation, all authors; writing—review and editing, P. Fernandes, S. Serrazina and E Corredoira; supervision, E. Corredoira. All authors have read and agreed to the published version of the manuscript.
Funding
At MBG-CSIC, work was supported by Ministerio de Ciencia, Innovación y Universidades (MICIN, Spain) through the projects TED2021-129633B-I00 (funded by MCIN/AEI /10.13039/501100011033 and NextGenerationEU/PRTR), PID2020-112627RB-C33 (funded by AEI/10.13039/501100011033) and PID2024-156422OB-C33. At DIFASA, work was supported by Fondazione CRT, the BIORES project 2023.0374 and the project VALORE IN CAMPO MIPAAF-2022-0667521-Azioni di Valorizzazione e Recupero per le filiere italiane di Castagno, Mandorlo, Pistacchio e Carrubo. At BioISI, work was supported by UIDB/04046/2020 (DOI: https:// doi. org/ 10. 54499/UIDB/ 04046/ 2020) and UIDP/04046/2020 (DOI: https:// doi. org/ 10. 54499/ UIDP/ 04046/ 2020), Centre grants from Fundação para a Ciência e a Tecnologia (FCT), Portugal. At UCO, work was supported by the Consejería de Conocimiento Investigación y Universidad, Junta de Andalucía, Grant/Award Number ProyExcel_00351 and MICIN (Spain). S. Serrazina is being funded by an FCT contract in the project 2022.06990. PTDC (DOI: 10.54499/2022.06990.PTDC). P. Piñeiro was funded by a Formación de Profesorado Universitario grant (FPU23/00921) from MICIN (Spain).
Data Availability Statement
Not applicable.
Acknowledgments
The authors wish to thank their mentors and predecessors for their pioneering contributions to the application of biotechnology in chestnut improvement.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Robin, C. Francia. In Il castagno: Coltura, ambiente ed utilizazioni.; Bounous, G., Ed.; Edagricole: Bologna, Italy, 2002; pp. 232–243.
- Conedera, M.; Tinner, W.; Krebs, P.; de Rigo, D.; Caudullo, G. Castanea sativa in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., De Rigo, D., Caudullo, G., Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; pp. 78–79.
- Fernandes, P.; Colavolpe, M.B.; Serrazina, S.; Costa, R.L. European and American Chestnuts: An Overview of the Main Threats and Control Efforts. Front Plant Sci 2022, 13, 951844. [CrossRef]
- Vieitez, F.; Merkle, S. Castanea Spp. Chestnut. In Biotechnology of fruit and nut crops; Litz, R., Ed.; CAB International: Wallingford, UK, 2005; pp. 265–296.
- De Vasconcelos, M.C.; Bennett, R.N.; Rosa, E.A.; Ferreira-Cardoso, J. V Composition of European Chestnut ( Castanea Sativa Mill.) and Association with Health Effects: Fresh and Processed Products. J Sci Food Agric 2010, 90, 1578–1589. [CrossRef]
- Vannini, A.; Vettraino, A.; Anselmo, N. Patología. In Il Castagno. Coltura, Ambiente ed Utilizzazioni; Bounous, G., Ed.; Edagricole: Bologna, Italy, 2002; pp. 103–111.
- Vannini, A.; Natili, G.; Anselmi, N.; Montaghi, A.; Vettraino, A.M. Distribution and Gradient Analysis of Ink Disease in Chestnut Forests. For Pathol 2010, 40, 73–86. [CrossRef]
- Dorado, F.J.; Alías, J.C.; Chaves, N.; Solla, A. Warming Scenarios and Phytophthora cinnamomi Infection in Chestnut (Castanea sativa Mill.). Plants 2023, 12, 556. [CrossRef]
- Heinz, M.; Prospero, S. A Modeling Approach to Determine Substitutive Tree Species for Sweet Chestnut in Stands Affected by Ink Disease. J For Res (Harbin) 2025, 36, 24. [CrossRef]
- McConnell, M.E.; Balci, Y. Phytophthora Cinnamomi as a Contributor to White Oak Decline in Mid-Atlantic United States Forests. Plant Dis 2014, 98, 319–327. [CrossRef]
- McConnell, M.E.; Balci, Y. Fine Root Dynamics of Oak Saplings in Response to Phytophthora Cinnamomi Infection under Different Temperatures and Durations. For Pathol 2015, 45, 155–164. [CrossRef]
- Weste, G.; Marks, G.C. The Biology of Phytophthora cinnamomi in Australasian Forests. Annu Rev Phytopathol 1987, 25, 207–229. [CrossRef]
- Khaliq, I.; Hardy, G.E.St.J.; Burgess, T.I. Phytophthora cinnamomi Exhibits Phenotypic Plasticity in Response to Cold Temperatures. Mycol Prog 2020, 19, 405–415. [CrossRef]
- EPPO Global Database. Phytophthora cinnamomi Distribution. Available online: https://gd.eppo.int/taxon/PHYTCN/distribution (accessed on 22 October 2025).
- Rigling, D.; Prospero, S. Cryphonectria Parasitica , the Causal Agent of Chestnut Blight: Invasion History, Population Biology and Disease Control. Mol Plant Pathol 2018, 19, 7–20. [CrossRef]
- Trapiello, E.; Rigling, D.; González, A.J. Occurrence of Hypovirus-Infected Cryphonectria parasitica Isolates in Northern Spain: An Encouraging Situation for Biological Control of Chestnut Blight in Asturian Forests. Eur J Plant Pathol 2017, 149, 503–514. [CrossRef]
- Ibáñez, I.; Pereira, E.; Moura, L.; Castro, J.P.; Gouveia, E. Natural Spread of Hypovirulence in Cryphonectria parasitica. A Case Study, Sergude – Minho – Portugal. Revista de Ciências Agrárias 2015, 38, 264–274.
- Rodríguez-Molina, M. del C.; García-García, M.B.; Osuna, M.D.; Gouveia, E.; Serrano-Pérez, P. Various Population Structures of Cryphonectria parasitica in Cáceres (Spain) Determine the Feasibility of the Biological Control of Chestnut Blight with Hypovirulent Strains. Agronomy 2023, Vol. 13, Page 1208 2023, 13, 1208. [CrossRef]
- Quinto, J.; Wong, M.E.; Boyero, J.R.; Vela, J.M.; Aguirrebengoa, M. Population Dynamics and Tree Damage of the Invasive Chestnut Gall Wasp, Dryocosmus kuriphilus, in Its Southernmost European Distributional Range. Insects 2021, 12, 900. [CrossRef]
- Ugolini, F.; Massetti, L.; Pedrazzoli, F.; Tognetti, R.; Vecchione, A.; Zulini, L.; Maresi, G. Ecophysiological Responses and Vulnerability to Other Pathologies in European Chestnut Coppices, Heavily Infested by the Asian Chestnut Gall Wasp. For Ecol Manage 2014, 314, 38–49. [CrossRef]
- Gehring, E.; Bellosi, B.; Quacchia, A.; Conedera, M. Assessing the Impact of Dryocosmus kuriphilus on the Chestnut Tree: Branch Architecture Matters. J Pest Sci (2004) 2018, 91, 189–202. [CrossRef]
- EPPO Dryocosmus kuriphilus. EPPO Datasheets on Pests Recommended for Regulation. Available online: https://gd.eppo.int (accessed on 21 October 2025).
- Savill, P.; Evans, J.; Auclair, D.; Falck, J. Genetics, Silviculture and the Promotion of Wood Quality (Forestry Commission Bulletin 126); Forestry Commission: Edinburgh, UK, 2005;
- Vieitez, A.M.; Corredoira, E.; Martínez, M.T.; San-José, M.C.; Sánchez, C.; Valladares, S.; Vidal, N.; Ballester, A. Application of Biotechnological Tools to Quercus Improvement. Eur J For Res 2012, 131, 519–539.
- Vieitez, A.M.; Ballester, A. Micropropagation of Chestnut (Castanea sativa Mill.). In Cell and Tissue Culture in Forestry; Bonga, J.M., Durzan, D.J., Eds.; Springer, 1986; Vol. 1, pp. 127–146.
- Alcaide, F.; Solla, A.; Cherubini, M.; Mattioni, C.; Cuenca, B.; Camisón, Á.; Martín, M.Á. Adaptive Evolution of Chestnut Forests to the Impact of Ink Disease in Spain. J Syst Evol 2020, 58, 504–516. [CrossRef]
- Conedera, M.; Tinner, W.; Krebs, P.; de Rigo, D.; Caudullo, G. Castanea Sativa in Europe: Distribution, Habitat, Usage And. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publ. Off. EU: Luxembourg, 2016; p. e0125e0+.
- Sandercock, A.M.; Westbrook, J.W.; Zhang, Q.; Johnson, H.A.; Saielli, T.M.; Scrivani, J.A.; Fitzsimmons, S.F.; Collins, K.; Perkins, M.T.; Craddock, J.H.; et al. Frozen in Time: Rangewide Genomic Diversity, Structure, and Demographic History of Relict American Chestnut Populations. Mol Ecol 2022, 31, 4640–4655. [CrossRef]
- Tanaka, N.; Matsui, T. PRDB: Phytosociological Relevé Database, Forestry and Forest Products Research Institute Available online: http://www.ffpri.go.jp/labs/prdb/index.html (accessed on 22 October 2025).
- Dane, F.; Lang, P.; Huang, H.; Fu, Y. Intercontinental Genetic Divergence of Castanea Species in Eastern Asia and Eastern North America. Heredity (Edinb) 2003, 91, 314–321. [CrossRef]
- EPPO Global Database. Cryphonectria parasitica Distribution. Available online: https://gd.eppo.int/taxon/ENDOPA/distribution (accessed on 22 October 2025).
- EPPO Global Database. Dryocosmus Kuriphilus Distribution. Available online: https://gd.eppo.int/taxon/DRYCKU/distribution (accessed on 22 October 2025).
- Inkscape Project. Inkscape 2025 Version 1.4.2 (f4327f4, 2025/05/13). Available online: https://inkscape.org.
- Grattapaglia, D.; Plomion, C.; Kirst, M.; Sederoff, R.R. Genomics of Growth Traits in Forest Trees. Curr Opin Plant Biol 2009, 12, 148–156. [CrossRef]
- Corredoira, E.; Merkle, S.A.; Martínez, M.T.; Toribio, M.; Canhoto, J.M.; Correia, S.I.; Ballester, A.; Vieitez, A.M. Non-Zygotic Embryogenesis in Hardwood Species. CRC Crit Rev Plant Sci 2019, 38, 29–97. [CrossRef]
- Grattapaglia, D.; Resende, M.D. V. Genomic Selection in Forest Tree Breeding. Tree Genet Genomes 2011, 7, 241–255. [CrossRef]
- Pike, C.C.; Koch, J.; Nelson, C.D. Breeding for Resistance to Tree Pests: Successes, Challenges, and a Guide to the Future. J For 2021, 119, 96–105. [CrossRef]
- Gallastegui, C. Técnica de La Hibridación Artificial Del Castaño. Boletín de la Real Sociedad Española de Historia Natural 1926, 26, 88–94.
- Urquijo, P. Aspectos de La Obtención de Híbridos Resistentes a La Enfermedad Del Castaño. Boletín de Patología Vegetal y Entomología Agrícola 1944, XIII, 447–462.
- Lafitte, G. Le Châtaignier Japonais En Pays Basque. Mendionde 69 1946.
- Gomes Guerreiro, M. Acerca Do Uso Da Análise Discriminatória: Comparação Entre Duas Castas de Castanhas. – Sep. Das Publicações Da Direcção Geral Dos Serviços Estais e Aquícolas, Vol. XV, Tomo I e II. 1948, 137–151.
- Bagnaresi, U. Osservazioni Morfo-Biologiche Sulle Provenienze Di Castagno Giapponese Coltivate in Italia. Pubblicazione n. 3, Centro Studio Sul Castagno (C.N.R.), Firenze. La Ricerca Scientifica (Suppl.) 1956, 26, 7–48.
- Schad, C.; Solignat, G.; Grente, J.; Venot, P. Recherches Sur Le Chataignier a La Station de Brive. Annales de L’amélioration des plantes 1952, 3, 369–458.
- Taveira-Fernándes, C. Aspects de l’ameliorationdu Chataignier Pour La Résistance a La Maladie de l’encre. In Proceedings of the In Actas III Congr Un Fitopat. Medit; Oeiras, Portugal, 1972; pp. 313–320.
- Burnham, C.R.; Rutter, P.A.; French, D.W. Breeding Blight-Resistant Chestnuts. In Plant Breeding Reviews; Wiley, 1986; pp. 347–397.
- Pereira-Lorenzo, S.; Fernandez-Lopez, J. Propagation of Chestnut Cultivars by Grafting: Methods, Rootstocks and Plant Quality. Journal of Horticultural Science and Biotechnology 1997, 72, 731–739. [CrossRef]
- Costa, R.; Santos, C.; Tavares, F.; Machado, H.; Gomes-Laranjo, J.; Kubisiak, T.; Nelson, C.D. Mapping and Transcriptomic Approches Implemented for Understanding Disease Resistance to Phytophthora cinammomi in Castanea Sp. BMC Proc 2011, 5, O18. [CrossRef]
- Míguez-Soto, B.; López-Villamor, A.; Fernández-López, J. Additive and Non-Additive Genetic Parameters for Multipurpose Traits in a Clonally Replicated Incomplete Factorial Test of Castanea Spp. Tree Genet Genomes 2016, 12, 47. [CrossRef]
- Fernandes, P.; Amaral, A.; Colavolpe, B.; Balonas, D.; Serra, M.; Pereira, A.; Costa, R.L. Propagation of New Chestnut Rootstocks with Improved Resistance to Phytophthora cinnamomi – New Cast Rootstocks. Silva Lusit 2020, 28, 15–29. [CrossRef]
- Fernandez-Lopez, J. Identification of the Genealogy of Interspecific Hybrids between Castanea sativa, Castanea crenata and Castanea mollissima. For Syst 2011, 20, 65–80. [CrossRef]
- Hennion, B. CHESTNUT PRODUCTION IN FRANCE: REVIEW, PERSPECTIVES. Acta Hortic 2010, 866, 493–497. [CrossRef]
- Serdar, Ü.; Saito, T.; Cuenca, B.; Akyüz, B.; Laranjo, J.G.; Beccaro, G.; Bounous, G.; Costa, R.L.; Fernandes, P.; Mellano, M.G.; et al. Advances in Cultivation of Chestnuts. In; Burleigh Dodds Science Publishing Limited, 2019; pp. 349–388.
- Gomes-Laranjo, J.; Coutinho, J.P.; Peixoto, F.; Araújo Alves, J. Ecologia Do Castanheiro (C. sativa Mill.) . In Castanheiros; Gomes-Laranjo, J., Ferreira-Cardoso, J., Portela, E., G.-Abreu, C., Eds.; UTAD: Vila Real, 2007; pp. 109–149.
- Martins, L.; Anjos, R.; Costa, R.; Gomes-Laranjo, J. Colutad. Um Clone de Castanheiro Resistente À Doença Da Tinta. In Castanheiros. Técnicas e Práticas; Gomes-Laranjo, J., Peixoto, F., Ferreira-Cardoso, J., Eds.; Pulido Consulting - Indústria Criativa and Universidade Trás-os-Montes e Alto Douro: Vila Real, 2009; pp. 135–142.
- Santos, C.; Machado, H.; Serrazina, S.; Gomes, F.; Gomes-Laranjo, J.; Correia, I.; Zhebentyayeva, T.; Duarte, S.; Bragança, H.; Fevereiro, P.; et al. Comprehension of Resistance to Diseases in Chestnut. Revista de Ciências Agrárias 2016, 39, 189–193. [CrossRef]
- Fernandes, P.; Amaral, A.; Colavolpe, B.; Pereira, A.; Costa, R. Avaliação agronómica de genótipos de castanheiro selecionados do programa de melhoramento genético para a resistência à tinta. Vida Rural. 2021, pp. 76–82.
- Santos, C.; Machado, H.; Correia, I.; Gomes, F.; Gomes-Laranjo, J.; Costa, R. Phenotyping Castanea Hybrids for Phytophthora cinnamomi Resistance. Plant Pathol 2015, 64, 901–910. [CrossRef]
- Breisch, H.; Boutitie, A.; Reyne, J.; Salesses, G.; Vaysse, P. Châtaignes et Marrons; FRA : CTIFL Centre Technique des Fruits et Légumes: Paris, 1995;
- Sartor, C.; Dini, F.; Torello Marinoni, D.; Mellano, M.G.; Beccaro, G.L.; Alma, A.; Quacchia, A.; Botta, R. Impact of the Asian Wasp Dryocosmus kuriphilus (Yasumatsu) on Cultivated Chestnut: Yield Loss and Cultivar Susceptibility. Sci Hortic 2015, 197, 454–460. [CrossRef]
- Paglietta, R. ‘Lusenta’ a New Early Ripening Euro Japanese Chestnut. In Proceedings of the World Chestnut Industry Conference; Wallace, R.D., Spinella, L.G., Eds.; Chestnut Marketing Assoc., Alachua, Florida, USA: Morgantown, West Virginia, 1992; pp. 10–11.
- Torello Marinoni, D.; Nishio, S.; Valentini, N.; Shirasawa, K.; Acquadro, A.; Portis, E.; Alma, A.; Akkak, A.; Pavese, V.; Cavalet-Giorsa, E.; et al. Development of High-Density Genetic Linkage Maps and Identification of Loci for Chestnut Gall Wasp Resistance in Castanea Spp. Plants 2020, 9, 1048. [CrossRef]
- Gaudet, M.; Pollegioni, P.; Ciolfi, M.; Mattioni, C.; Cherubini, M.; Beritognolo, I. Identification of a Unique Genomic Region in Sweet Chestnut (Castanea sativa Mill.) That Controls Resistance to Asian Chestnut Gall Wasp Dryocosmus Kuriphilus Yasumatsu. Plants 2024, 13, 1355. [CrossRef]
- Westbrook, J.W.; Zhang, Q.; Mandal, M.K.; Jenkins, E. V.; Barth, L.E.; Jenkins, J.W.; Grimwood, J.; Schmutz, J.; Holliday, J.A. Optimizing Genomic Selection for Blight Resistance in American Chestnut Backcross Populations: A Trade-off with American Chestnut Ancestry Implies Resistance Is Polygenic. Evol Appl 2019, 13, 31–47. [CrossRef]
- Wang, C.; Wang, W.; Zhang, S.; Chen, Y.; Zhao, Y.; Zhu, C. Anatomical Changes during Chestnut (Castanea Mollissima BL.) Gall Development Stages Induced by the Gall Wasp Dryocosmus kuriphilus (Hymenoptera: Cynipidae). Plants 2024, Vol. 13, Page 1766 2024, 13, 1766. [CrossRef]
- Camisón, Á.; Martín, M.Á.; Flors, V.; Sánchez-Bel, P.; Pinto, G.; Vivas, M.; Rolo, V.; Solla, A. Exploring the Use of Scions and Rootstocks from Xeric Areas to Improve Drought Tolerance in Castanea sativa Miller. Environ Exp Bot 2021, 187, 104467. [CrossRef]
- Fei, S.; Liang, L.; Paillet, F.L.; Steiner, K.C.; Fang, J.; Shen, Z.; Wang, Z.; Hebard, F. V. Modelling Chestnut Biogeography for American Chestnut Restoration. Divers Distrib 2012, 18, 754–768. [CrossRef]
- Míguez-Soto, B.; Fernández-Cruz, J.; Fernández-López, J. Mediterranean and Northern Iberian Gene Pools of Wild Castanea Sativa Mill. Are Two Differentiated Ecotypes Originated under Natural Divergent Selection. PLoS One 2019, 14, e0211315. [CrossRef]
- Freitas, T.R.; Santos, J.A.; Silva, A.P.; Fraga, H. Influence of Climate Change on Chestnut Trees: A Review. Plants 2021, Vol. 10, Page 1463 2021, 10, 1463. [CrossRef]
- Alcaide, F.; Solla, A.; Mattioni, C.; Castellana, S.; Martín, M.Á. Adaptive Diversity and Drought Tolerance in Castanea Sativa Assessed through EST-SSR Genic Markers. Forestry: An International Journal of Forest Research 2019, 92, 287–296. [CrossRef]
- Dorado, F.J.; Solla, A.; Alcaide, F.; Martín, M.Á. Assessing Heat Stress Tolerance in Castanea sativa. Forestry: An International Journal of Forest Research 2022, 95, 667–677. [CrossRef]
- Perez-Rial, A.; Castro, P.; Solla, A.; Martín, M.A.; Die, J.V. Decoding Drought Tolerance from a Genomic Approach in Castanea Sativa Mill. Plant Genome 2025.
- Solla, A.; Dorado, F.J.; González, R.; Giraldo-Chaves, L.B.; Cubera, E.; Rocha, G.; Martín, C.; Martín, E.; Cuenca, B.; del Pozo, J.L.; et al. Chestnut Trees ( Castanea sativa Mill.) for Climate Change. Acta Hortic 2024, 1, 273–282. [CrossRef]
- Marinoni, D.; Akkak, A.; Bounous, G.; Edwards, K.J.; Botta, R. Development and Characterization of Microsatellite Markers in Castanea sativa (Mill.). Molecular Breeding 2003, 11, 127–136. [CrossRef]
- Buck, E.J.; Hadonou, M.; James, C.J.; Blakesley, D.; Russell, K. Isolation and Characterization of Polymorphic Microsatellites in European Chestnut ( Castanea sativa Mill.). Mol Ecol Notes 2003, 3, 239–241. [CrossRef]
- Martín, M.A.; Alvarez, J.B.; Mattioni, C.; Cherubini, M.; Villani, F.; Martin, L.M. Identification and Characterisation of Traditional Chestnut Varieties of Southern Spain Using Morphological and Simple Sequence Repeat (SSRs) Markers. Annals of Applied Biology 2009, 154, 389–398. [CrossRef]
- Martín, M.A.; Mattioni, C.; Cherubini, M.; Taurchini, D.; Villani, F. Genetic Characterisation of Traditional Chestnut Varieties in Italy Using Microsatellites (Simple Sequence Repeats) Markers. Annals of Applied Biology 2010, 157, 37–44. [CrossRef]
- Torello Marinoni, D.; Akkak, A.; Beltramo, C.; Guaraldo, P.; Boccacci, P.; Bounous, G.; Ferrara, A.M.; Ebone, A.; Viotto, E.; Botta, R. Genetic and Morphological Characterization of Chestnut (Castanea sativa Mill.) Germplasm in Piedmont (North-Western Italy). Tree Genet Genomes 2013, 9, 1017–1030. [CrossRef]
- Gobbin, D.; Hohl, L.; Conza, L.; Jermini, M.; Gessler, C.; Conedera, M. Microsatellite-Based Characterization of the Castanea sativa Cultivar Heritage of Southern Switzerland. Genome 2007, 50, 1089–1103. [CrossRef]
- Pereira-Lorenzo, S.; Ramos-Cabrer, A.M.; Barreneche, T.; Mattioni, C.; Villani, F.; Díaz-Hernández, M.B.; Martín, L.M.; Martín, Á. Database of European Chestnut Cultivars and Definition of a Core Collection Using Simple Sequence Repeats. Tree Genet Genomes 2017, 13, 114. [CrossRef]
- Pereira-Lorenzo, S.; Costa, R.M.L.; Ramos-Cabrer, A.M.; Ciordia-Ara, M.; Ribeiro, C.A.M.; Borges, O.; Barreneche, T. Chestnut Cultivar Diversification Process in the Iberian Peninsula, Canary Islands, and Azores. Genome 2011, 54, 301–315. [CrossRef]
- Pereira-Lorenzo, S.; Costa, R.M.L.; Ramos-Cabrer, A.M.; Ribeiro, C.A.M.; da Silva, M.F.S.; Manzano, G.; Barreneche, T. Variation in Grafted European Chestnut and Hybrids by Microsatellites Reveals Two Main Origins in the Iberian Peninsula. Tree Genet Genomes 2010, 6, 701–715. [CrossRef]
- Pereira-Lorenzo, S.; Ramos-Cabrer, A.M.; Barreneche, T.; Mattioni, C.; Villani, F.; Díaz-Hernández, B.; Martín, L.M.; Robles-Loma, A.; Cáceres, Y.; Martín, A. Instant Domestication Process of European Chestnut Cultivars. Annals of Applied Biology 2019, 174, 74–85. [CrossRef]
- Nunziata, A.; Ruggieri, V.; Petriccione, M.; De Masi, L. Single Nucleotide Polymorphisms as Practical Molecular Tools to Support European Chestnut Agrobiodiversity Management. International Journal of Molecular Sciences 2020, Vol. 21, Page 4805 2020, 21, 4805. [CrossRef]
- Martín, M.A.; Monedero, E.; Martín, L.M. Genetic Monitoring of Traditional Chestnut Orchards Reveals a Complex Genetic Structure. Ann For Sci 2017, 74, 15. [CrossRef]
- Mattioni, C.; Angela Martín, M.; Pollegioni, P.; Cherubini, M.; Villani, F. Microsatellite Markers Reveal a Strong Geographical Structure in European Populations of Castanea sativa (Fagaceae): Evidence for Multiple Glacial Refugia. Am J Bot 2013, 100, 951–961. [CrossRef]
- Mattioni, C.; Martin, M.A.; Chiocchini, F.; Cherubini, M.; Gaudet, M.; Pollegioni, P.; Velichkov, I.; Jarman, R.; Chambers, F.M.; Paule, L.; et al. Landscape Genetics Structure of European Sweet Chestnut (Castanea sativa Mill): Indications for Conservation Priorities. Tree Genet Genomes 2017, 13, 39. [CrossRef]
- Castellana, S.; Martin, M.Á.; Solla, A.; Alcaide, F.; Villani, F.; Cherubini, M.; Neale, D.; Mattioni, C. Signatures of Local Adaptation to Climate in Natural Populations of Sweet Chestnut (Castanea sativa Mill.) from Southern Europe. Ann For Sci 2021, 78, 27. [CrossRef]
- Casasoli, M.; Mattioni, C.; Cherubini, M.; Villani, F. A Genetic Linkage Map of European Chestnut (Castanea sativa Mill.) Based on RAPD, ISSR and Isozyme Markers. Theoretical and Applied Genetics 2001, 102, 1190–1199. [CrossRef]
- Casasoli, M.; Pot, D.; Plomion, C.; Monteverdi, M.C.; Barreneche, T.; Lauteri, M.; Villani, F. Identification of QTLs Affecting Adaptive Traits in Castanea Sativa Mill. Plant Cell Environ 2004, 27, 1088–1101. [CrossRef]
- Casasoli, M.; Derory, J.; Morera-Dutrey, C.; Brendel, O.; Porth, I.; Guehl, J.M.; Villani, F.; Kremer, A. Comparison of Quantitative Trait Loci for Adaptive Traits Between Oak and Chestnut Based on an Expressed Sequence Tag Consensus Map. Genetics 2006, 172, 533–546. [CrossRef]
- Barreneche, T.; Casasoli, M.; Russell, K.; Akkak, A.; Meddour, H.; Plomion, C.; Villani, F.; Kremer, A. Comparative Mapping between Quercus and Castanea Using Simple-Sequence Repeats (SSRs). Theoretical and Applied Genetics 2004, 108, 558–566. [CrossRef]
- Durand, J.; Bodénès, C.; Chancerel, E.; Frigerio, J.-M.; Vendramin, G.; Sebastiani, F.; Buonamici, A.; Gailing, O.; Koelewijn, H.-P.; Villani, F.; et al. A Fast and Cost-Effective Approach to Develop and Map EST-SSR Markers: Oak as a Case Study. BMC Genomics 2010, 11, 570. [CrossRef]
- Fernandes, P.; Pimentel, D.; Ramiro, R.S.; Silva, M. do C.; Fevereiro, P.; Costa, R.L. Dual Transcriptomic Analysis Reveals Early Induced Castanea Defense-Related Genes and Phytophthora cinnamomi Effectors. Front Plant Sci 2024, 15. [CrossRef]
- Serrazina, S.; Santos, C.; Machado, H.; Pesquita, C.; Vicentini, R.; Pais, M.S.; Sebastiana, M.; Costa, R. Castanea Root Transcriptome in Response to Phytophthora cinnamomi Challenge. Tree Genet Genomes 2015, 11, 6. [CrossRef]
- Santos, C.; Duarte, S.; Tedesco, S.; Fevereiro, P.; Costa, R.L. Expression Profiling of Castanea Genes during Resistant and Susceptible Interactions with the Oomycete Pathogen Phytophthora cinnamomi Reveal Possible Mechanisms of Immunity. Front Plant Sci 2017, 8, 515. [CrossRef]
- Camisón, Á.; Monteiro, P.; Dorado, F.J.; Sánchez-Bel, P.; Leitão, F.; Meijón, M.; Pinto, G. Choosing the Right Signaling Pathway: Hormone Responses to Phytophthora cinnamomi during Compatible and Incompatible Interactions with Chestnut (Castanea Spp.). Tree Physiol 2025, 45. [CrossRef]
- Fernandes, P.; Machado, H.; Silva, M. do C.; Costa, R.L. A Histopathological Study Reveals New Insights Into Responses of Chestnut (Castanea Spp.) to Root Infection by Phytophthora cinnamomi. Phytopathology 2021, 111, 345–355. [CrossRef]
- Miyakawa, T.; Sawano, Y.; Miyazono, K.I.; Hatano, K.I.; Tanokura, M. Crystallization and Preliminary X-Ray Analysis of Ginkbilobin-2 from Ginkgo Biloba Seeds: A Novel Antifungal Protein with Homology to the Extracellular Domain of Plant Cysteine-Rich Receptor-like Kinases. Acta Crystallogr Sect F Struct Biol Cryst Commun 2007, 63, 737–739. [CrossRef]
- Gao, N.; Wadhwani, P.; Mühlhäuser, P.; Liu, Q.; Riemann, M.; Ulrich, A.S.; Nick, P. An Antifungal Protein from Ginkgo Biloba Binds Actin and Can Trigger Cell Death. Protoplasma 2016, 253, 1159–1174. [CrossRef]
- Miyakawa, T.; Hatano, K.I.; Miyauchi, Y.; Suwa, Y.I.; Sawano, Y.; Tanokura, M. A Secreted Protein with Plant-Specific Cysteine-Rich Motif Functions as a Mannose-Binding Lectin That Exhibits Antifungal Activity. Plant Physiol 2014, 166, 766–778. [CrossRef]
- Serrazina, S.; Martínez, M.T.; Fernandes, P.; Colavolpe, B.; Dias, F.; Conde, P.; Malhó, R.; Corredoira, E.; Costa, R.L. Castanea crenata Ginkbilobin2-like as a Resistance Gene to Phytophthora cinnamomi Infection. Acta Hortic 2024, 1, 77–87. [CrossRef]
- Serrazina, S.; Martínez, M.T.; Cano, V.; Malhó, R.; Costa, R.L.; Corredoira, E. Genetic Transformation of Quercus ilex Somatic Embryos with a Gnk2-like Protein That Reveals a Putative Anti-Oomycete Action. Plants 2022, 11, 304. [CrossRef]
- Serrazina, S.; Martínez, M.; Soudani, S.; Candeias, G.; Berrocal-Lobo, M.; Piñeiro, P.; Malhó, R.; Costa, R.L.; Corredoira, E. Overexpression of Ginkbilobin-2 Homologous Domain Gene Improves Tolerance to Phytophthora cinnamomi in Somatic Embryos of Quercus Suber. Sci Rep 2024, 14, 19357. [CrossRef]
- McGuigan, L.; Fernandes, P.; Oakes, A.; Stewart, K.; Powell, W. Transformation of American Chestnut (Castanea dentata (Marsh.) Borkh) Using Rita® Temporary Immersion Bioreactors and We Vitro Containers. Forests 2020, 11, 1–15. [CrossRef]
- Saiz-Fernández, I.; Milenković, I.; Berka, M.; Černý, M.; Tomšovský, M.; Brzobohatý, B.; Kerchev, P. Integrated Proteomic and Metabolomic Profiling of Phytophthora cinnamomi Attack on Sweet Chestnut (Castanea sativa) Reveals Distinct Molecular Reprogramming Proximal to the Infection Site and Away from It. Int J Mol Sci 2020, 21, 8525. [CrossRef]
- Tian, H.; Xu, L.; Li, X.; Zhang, Y. Salicylic Acid: The Roles in Plant Immunity and Crosstalk with Other Hormones. J Integr Plant Biol 2025, 67, 773–785. [CrossRef]
- Pavese, V.; Moglia, A.; Gonthier, P.; Torello Marinoni, D.; Cavalet-Giorsa, E.; Botta, R. Identification of Susceptibility Genes in Castanea sativa and Their Transcription Dynamics Following Pathogen Infection. Plants 2021, 10, 913. [CrossRef]
- Serrazina, S.; Machado, H.; Costa, R.L.; Duque, P.; Malhó, R. Expression of Castanea crenata Allene Oxide Synthase in Arabidopsis Improves the Defense to Phytophthora cinnamomi. Front Plant Sci 2021, 12, 149. [CrossRef]
- Langin, G.; González-Fuente, M.; Üstün, S. The Plant Ubiquitin-Proteasome System as a Target for Microbial Manipulation. Annu Rev Phytopathol 2023, 61, 351–375. [CrossRef]
- Santos, C.; Zhebentyayeva, T.; Serrazina, S.; Nelson, C.D.; Costa, R. Development and Characterization of EST-SSR Markers for Mapping Reaction to Phytophthora cinnamomi in Castanea Spp. Sci Hortic 2015, 194, 181–187. [CrossRef]
- Santos, C.; Nelson, C.D.; Zhebentyayeva, T.; Machado, H.; Gomes-Laranjo, J.; Costa, R.L. First Interspecific Genetic Linkage Map for Castanea sativa x Castanea crenata Revealed QTLs for Resistance to Phytophthora cinnamomi. PLoS One 2017, 12, e0184381. [CrossRef]
- Zhebentyayeva, T.N.; Sisco, P.H.; Georgi, L.L.; Jeffers, S.N.; Perkins, M.T.; James, J.B.; Hebard, F. V.; Saski, C.; Nelson, C.D.; Abbott, A.G. Dissecting Resistance to Phytophthora cinnamomi in Interspecific Hybrid Chestnut Crosses Using Sequence-Based Genotyping and QTL Mapping. Phytopathology 2019, 109, 1594–1604. [CrossRef]
- Lovat, C.A.; Donnelly, D.J. Mechanisms and Metabolomics of the Host–Pathogen Interactions between Chestnut (Castanea Species) and Chestnut Blight (Cryphonectria parasitica). For Pathol 2019, 49, e12562. [CrossRef]
- Antipova, T. V.; Zhelifonova, V.P.; Litovka, Y.A.; Baskunov, B.P.; Noskov, A.E.; Timofeyev, A.A.; Pavlov, I.N. Secondary Metabolites of Phytopathogenic Cryphonectria parasitica (Murrill) and Their in Vitro Phytotoxicity. Journal of Plant Pathology 2025, 1–13. [CrossRef]
- Ježić, M.; Nuskern, L.; Peranić, K.; Popović, M.; Ćurković-Perica, M.; Mendaš, O.; Škegro, I.; Poljak, I.; Vidaković, A.; Idžojtić, M. Regional Variability of Chestnut (Castanea sativa) Tolerance Toward Blight Disease. Plants 2024, 13, 3060. [CrossRef]
- Kovács, G.E.; Szőke, L.; Tóth, B.; Kovács, B.; Bojtor, C.; Illés, Á.; Radócz, L.; Moloi, M.J.; Radócz, L. The Physiological and Biochemical Responses of European Chestnut (Castanea sativa L.) to Blight Fungus (Cryphonectria parasitica (Murill) Barr). Plants 2021, Vol. 10, Page 2136 2021, 10, 2136. [CrossRef]
- Kazmierczak, P.; Pfeiffer, P.; Zhang, L.; Van Alfen, N.K. Transcriptional Repression of Specific Host Genes by the Mycovirus Cryphonectria Hypovirus 1. J Virol 1996, 70, 1137–1142. [CrossRef]
- Heiniger, U.; Rigling, D. BIOLOGICAL CONTROL OF CHESTNUT BLIGHT IN EUROPE. Annu Rev Phytopathol 1994, 32, 581–599. [CrossRef]
- Bolvanský, M.; Adamčíková, K.; Kobza, M. Screening Resistance to Chestnut Blight in Young Chestnut Trees Derived from Castanea sativa × C. crenata Hybrids. FOLIA OECOLOGICA 2014, 41, 1–7.
- Chira, D.; Teodorescu, R.; Mantale, C.; Chira, F.; Isaia, G.; Achim, G.; Scutelnicu, A.; Botu, M. Testing Chestnut Hybrids for Resistance to Cryphonectria parasitica. Acta Hortic 2018, 1220, 113–120. [CrossRef]
- van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of Inducible Defense-Related Proteins in Infected Plants. Annu Rev Phytopathol 2006, 44, 135–162. [CrossRef]
- Schafleitner, R.; Wilhelm, E. Effect of Virulent and Hypovirulent Cryphonectria parasitica (Murr.) Barr on the Intercellular Pathogen Related Proteins and on Total Protein Pattern of Chestnut (Castanea sativa Mill.). Physiol Mol Plant Pathol 1997, 51, 323–332. [CrossRef]
- Schafleitner, R.; Wilhelm, E. CHESTNUT BLIGHT: MONITORING THE HOST RESPONSE WITH HETEROLOGOUS CDNA PROBES. Acta Hortic 1999, 494, 481–488. [CrossRef]
- Vannini, A.; Caruso, C.; Leonardi, L.; Rugini, E.; Chiarot, E.; Caporale, C.; Buonocore, V. Antifungal Properties of Chitinases from Castanea sativa against Hypovirulent and Virulent Strains of the Chestnut Blight Fungus Cryphonectria parasitica. Physiol Mol Plant Pathol 1999, 55, 29–35. [CrossRef]
- Allona, I.; Collada, C.; Casado, R.; Paz-Ares, J.; Aragoncillo, C. Bacterial Expression of an Active Class Ib Chitinase from Castanea sativa Cotyledons. Plant Mol Biol 1996, 32, 1171–1176. [CrossRef]
- Corredoira, E.; San José, M.C.; Vieitez, A.M.; Allona, I.; Aragoncillo, C.; Ballester, A. Agrobacterium-Mediated Transformation of European Chestnut Somatic Embryos with a Castanea sativa (Mill.) Endochitinase Gene. New For (Dordr) 2016, 47, 669–684. [CrossRef]
- Pandey, D.; Rajendran, S.R.C.K.; Gaur, M.; Sajeesh, P.K.; Kumar, A. Plant Defense Signaling and Responses Against Necrotrophic Fungal Pathogens. J Plant Growth Regul 2016, 35, 1159–1174. [CrossRef]
- Barakat, A.; Staton, M.; Cheng, C.-H.; Park, J.; Yassin, N.B.M.; Ficklin, S.; Yeh, C.-C.; Hebard, F.; Baier, K.; Powell, W.; et al. Chestnut Resistance to the Blight Disease: Insights from Transcriptome Analysis. BMC Plant Biol 2012, 12. [CrossRef]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.J. Signaling Crosstalk between Salicylic Acid and Ethylene/Jasmonate in Plant Defense: Do We Understand What They Are Whispering? International Journal of Molecular Sciences 2019, Vol. 20, Page 671 2019, 20, 671. [CrossRef]
- Staton, M.; Addo-Quaye, C.; Cannon, N.; Yu, J.; Zhebentyayeva, T.; Huff, M.; Islam-Faridi, N.; Fan, S.; Georgi, L.L.; Nelson, C.D.; et al. “A Reference Genome Assembly and Adaptive Trait Analysis of Castanea mollissima ‘Vanuxem,’ a Source of Resistance to Chestnut Blight in Restoration Breeding.” Tree Genet Genomes 2020, 16, 57. [CrossRef]
- Zulfiqar, S.; Aslam, M.M.; Ditta, A.; Iqbal, R.; Mustafa, A.E.Z.M.A.; Elshikh, M.S.; Uzair, M.; Aghayeva, S.; Qasim, M.; Ercisli, S.; et al. Evaluation of Genetic Diversity and Population Structure of the Chinese Chestnut (Castanea mollissima) by Using NR-SSR Markers. Genet Resour Crop Evol 2025, 72, 2445–2457. [CrossRef]
- Cheng, L.; Huang, W.; Lan, Y.; Cao, Q.; Su, S.; Zhou, Z.; Wang, J.; Liu, J.; Hu, G. The Complete Chloroplast Genome Sequence of the Wild Chinese Chestnut (Castanea Mollissima). Conserv Genet Resour 2018, 10, 291–294. [CrossRef]
- Kubisiak, T.L.; Hebard, F. V.; Nelson, C.D.; Zhang, J.; Bernatzky, R.; Huang, H.; Anagnostakis, S.L.; Doudrick, R.L. Molecular Mapping of Resistance to Blight in an Interspecific Cross in the Genus Castanea. Phytopathology 1997, 87, 751–759. [CrossRef]
- Fan, S.; Georgi, L.L.; Hebard, F. V.; Zhebentyayeva, T.; Yu, J.; Sisco, P.H.; Fitzsimmons, S.F.; Staton, M.E.; Abbott, A.G.; Nelson, C.D. Mapping QTLs for Blight Resistance and Morpho-Phenological Traits in Inter-Species Hybrid Families of Chestnut (Castanea Spp.). Front Plant Sci 2024, 15, 1365951. [CrossRef]
- Westbrook, J.W.; Malukiewicz, J.; Sreedasyam, A.; Jenkins, J.W.; Zhang, Q.; Lakoba, V.; Fitzsimmons, S.F.; Van Clief, J.; Collins, K.; Hoy, S.; et al. Improving American Chestnut Resistance to Two Invasive Pathogens through Genome-Enabled Breeding. bioRxiv 2025, 2025.01.30.635736.
- Powell, W.A.; Newhouse, A.E.; Coffey, V. Developing Blight-Tolerant American Chestnut Trees. Cold Spring Harb Perspect Biol 2019, 11, a034587. [CrossRef]
- Newhouse, A.; Breda, K.; McGuigan, L.; Coffey, V.; Oakes, A.; Matthews, D.; Drake, J.; Pilkey, H.; Satchwell, S.; Carlson, E.; et al. Petition for Determination of Nonregulated Status for Blight-Tolerant Darling 54 American Chestnut (Castanea Dentata) Available online: https://www.regulations.gov/document/APHIS-2020-0030-17585 (accessed on 22 October 2025).
- Zhang, B.; Oakes, A.D.; Newhouse, A.E.; Baier, K.M.; Maynard, C.A.; Powell, W.A. A Threshold Level of Oxalate Oxidase Transgene Expression Reduces Cryphonectria parasitica-Induced Necrosis in a Transgenic American Chestnut (Castanea dentata) Leaf Bioassay. Transgenic Res 2013, 22, 973–982. [CrossRef]
- USDA-APHIS. Draft Environmental Impact Statement: The State University of New York College of Environmental Science and Forestry Petition (19-309-01p) for Determination of Nonregulated Status for Blight-Tolerant Darling 54 American Chestnut (Castanea dentata) Available online: https://www.regulations.gov/document/APHIS-2020-0030-17583 (accessed on 28 October 2025).
- Barakat, A.; Diloreto, D.S.; Zhang, Y.; Smith, C.; Baier, K.; Powell, W.A.; Wheeler, N.; Sederoff, R. Comparison of the Transcriptomes of American Chestnut (Castanea dentata) and Chinese Chestnut (Castanea mollissima) in Response to the Chestnut Blight Infection. BMC Plant Biol 2009, 9. [CrossRef]
- Ghozlan, M.H.; EL-Argawy, E.; Tokgöz, S.; Lakshman, D.K.; Mitra, A.; Ghozlan, M.H.; EL-Argawy, E.; Tokgöz, S.; Lakshman, D.K.; Mitra, A. Plant Defense against Necrotrophic Pathogens. Am J Plant Sci 2020, 11, 2122–2138. [CrossRef]
- Künstler, A.; Bacsó, R.; Gullner, G.; Hafez, Y.M.; Király, L. Staying Alive – Is Cell Death Dispensable for Plant Disease Resistance during the Hypersensitive Response? Physiol Mol Plant Pathol 2016, 93, 75–84. [CrossRef]
- Almagro, L.; Gómez Ros, L. V.; Belchi-Navarro, S.; Bru, R.; Ros Barceló, A.; Pedreño, M.A. Class III Peroxidases in Plant Defence Reactions. J Exp Bot 2009, 60, 377–390. [CrossRef]
- Passardi, F.; Cosio, C.; Penel, C.; Dunand, C. Peroxidases Have More Functions than a Swiss Army Knife. Plant Cell Rep 2005, 24, 255–265. [CrossRef]
- Havir, E.A.; Anagnostakis, S.L. Seasonal Variation of Peroxidase Activity in Chestnut Trees. Phytochemistry 1998, 48, 41–47. [CrossRef]
- Dane, F.; Wang, Z.; Goertzen, L. Analysis of the Complete Chloroplast Genome of Castanea Pumila Var. Pumila, the Allegheny Chinkapin. Tree Genet Genomes 2015, 11, 1–6. [CrossRef]
- Nie, X.; Zhao, S.; Hao, Y.; Gu, S.; Zhang, Y.; Qi, B.; Xing, Y.; Qin, L. Transcriptome Analysis Reveals Key Genes Involved in the Resistance to Cryphonectria parasitica during Early Disease Development in Chinese Chestnut. BMC Plant Biol 2023, 23, 1–15. [CrossRef]
- Khatib, M.; Campo, M.; Bellumori, M.; Cecchi, L.; Vignolini, P.; Innocenti, M.; Mulinacci, N. Tannins from Different Parts of the Chestnut Trunk (Castanea sativa Mill.): A Green and Effective Extraction Method and Their Profiling by High-Performance Liquid Chromatography-Diode Array Detector-Mass Spectrometry. ACS Food Science & Technology 2023, 3, 1903–1912. [CrossRef]
- Anagnostakis, S.L. Chestnut Bark Tannin Assays and Growth of Chestnut Blight Fungus on Extracted Tannin. J Chem Ecol 1992, 18, 1365–1373. [CrossRef]
- Elkins, J.R.; Pate, W.; Hicks, S. Evidence for a Role of Hamamelitannin in the Pathogenicity of Endothia parasitica. Phytopathology 1979, 69, 1027–1028.
- Hebard, F. V.; Griffin, G.J.; Elkins, J.R. Developmental Histopathology of Cankers Incited by Hypovirulent and Virulent Isolates of Endothia Parasitica on Susceptible and Resistant Chestnut Trees. Phytopathology 1984, 74, 140. [CrossRef]
- Cooper, W.R.; Rieske, L.K. Differential Responses in American (Castanea dentata Marshall) and Chinese (C. mollissima Blume) Chestnut (Falales: Fagaceae) to Foliar Application of Jasmonic Acid. Chemoecology 2008, 18, 121–127. [CrossRef]
- Gao, S.; Shain, L. Activity of Polygalacturonase Produced by Cryphonectria parasitica in Chestnut Bark and Its Inhibition by Extracts from American and Chinese Chestnut. Physiol Mol Plant Pathol 1995, 46, 199–213. [CrossRef]
- Ferracini, C.; Ferrari, E.; Pontini, M.; Saladini, M.A.; Alma, A. Effectiveness of Torymus sinensis: A Successful Long-Term Control of the Asian Chestnut Gall Wasp in Italy. J Pest Sci (2004) 2019, 92, 353–359. [CrossRef]
- Quacchia, A.; Moriya, S.; Bosio, G.; Scapin, I.; Alma, A. Rearing, Release and Settlement Prospect in Italy of Torymus sinensis, the Biological Control Agent of the Chestnut Gall Wasp Dryocosmus kuriphilus. BioControl 2008, 53, 829–839. [CrossRef]
- Sartor, C.; Botta, R.; Mellano, M.G.; Beccaro, G.L.; Bounous, G.; Torello Marinoni, D.; Quacchia, A.; Alma, A. Evaluation of Susceptibility to Dryocosmus kuriphilus Yasumatsu (Hymenoptera: Cynipidae) in Castanea sativa Miller and in Hybrid Cultivars. Acta Hortic 2009, 815, 289–298. [CrossRef]
- Dini, F.; Sartor, C.; Botta, R. Detection of a Hypersensitive Reaction in the Chestnut Hybrid ‘Bouche de Bétizac’ Infested by Dryocosmus kuriphilus Yasumatsu. Plant Physiology and Biochemistry 2012, 60, 67–73. [CrossRef]
- Botta, R.; Sartor, C.; Marinoni, D.T.; Quacchia, A.; Alma, A. Differential Gene Expression in Chestnut Buds Following Infestation by Gall-Wasp (Dryocosmus kuriphilus Yasumatsu, Hymenoptera: Cynipidae). Acta Hortic 2009, 844, 405–410. [CrossRef]
- Acquadro, A.; Torello Marinoni, D.; Sartor, C.; Dini, F.; Macchio, M.; Botta, R. Transcriptome Characterization and Expression Profiling in Chestnut Cultivars Resistant or Susceptible to the Gall Wasp Dryocosmus kuriphilus. Molecular Genetics and Genomics 2020, 295, 107–120. [CrossRef]
- Zhu, C.; Wang, W.; Chen, Y.; Zhao, Y.; Zhang, S.; Shi, F.; Khalil-Ur-Rehman, M.; Nieuwenhuizen, N.J. Transcriptomics and Antioxidant Analysis of Two Chinese Chestnut (Castanea mollissima BL.) Varieties Provides New Insights Into the Mechanisms of Resistance to Gall Wasp Dryocosmus kuriphilus Infestation. Front Plant Sci 2022, 13, 874434. [CrossRef]
- Liu, B.; Ren, Y.; Su, C.; Wang, X.; Zeng, Y.; Zhu, D. Chromosome-Level Genome Assembly of Oriental Chestnut Gall Wasp (Dryocosmus kuriphilus). Sci Data 2024, 11, 963. [CrossRef]
- Bianco, L.; Fontana, P.; Marchesini, A.; Torre, S.; Moser, M.; Piazza, S.; Alessandri, S.; Pavese, V.; Pollegioni, P.; Vernesi, C.; et al. The de Novo, Chromosome-Level Genome Assembly of the Sweet Chestnut (Castanea Sativa Mill.) Cv. Marrone Di Chiusa Pesio. BMC Genom Data 2024, 25, 64. [CrossRef]
- Uncu, A.O.; Cetin, D.; Srivastava, V.; Uncu, A.T.; Akbudak, M.A. A Genome Sequence Resource for the European Chestnut (Castanea Sativa Mill.) and the Development of Genic Microsatellite Markers. Genet Resour Crop Evol 2025, 72, 3621–3636. [CrossRef]
- Shirasawa, K.; Nishio, S.; Terakami, S.; Botta, R.; Marinoni, D.T.; Isobe, S. Chromosome-Level Genome Assembly of Japanese Chestnut (Castanea crenata Sieb. et Zucc.) Reveals Conserved Chromosomal Segments in Woody Rosids. DNA Research 2021, 28. [CrossRef]
- Sandercock, A.M.; Westbrook, J.W.; Zhang, Q.; Holliday, J.A. A Genome-Guided Strategy for Climate Resilience in American Chestnut Restoration Populations. Proceedings of the National Academy of Sciences 2024, 121, e2403505121. [CrossRef]
- Pavese, V.; Moglia, A.; Abbà, S.; Milani, A.M.; Marinoni, D.T.; Corredoira, E.; Martinez, M.T.; Botta, R. First Report on Genome Editing via Ribonucleoprotein (RNP) in Castanea sativa Mill. Int J Mol Sci 2022, 23. [CrossRef]
- Vieitez, A.M.; Vieitez, M.L.; Vieitez, E. Chestnut Spp. . In Biotechnology in Agriculture and Forestry, Trees I; Bajaj, Y.P.S., Ed.; Springer Verlag: Berlin, 1986; pp. 393–414.
- Ballester, A.; Sánchez, M.C.; Vieitez, A.M. Etiolation as a Pretreatment for in Vitro Establishment and Multiplication of Mature Chestnut. Physiol Plant 1989, 77, 395–400. [CrossRef]
- Sánchez, M.; Ballester, A.; Vieitez, A. Reinvigoration Treatments for the Micropropagation of Mature Chestnut Trees. Annales des Sciences Forestières 1997, 54, 359–370. [CrossRef]
- Giovannelli, A.; Giannini, R. Reinvigoration of Mature Chestnut (Castanea sativa) by Repeated Graftings and Micropropagation. Tree Physiol 2000, 20, 1243–1248. [CrossRef]
- Corredoira, E.; Martínez, M.; Cernadas, M.; San José, M. Application of Biotechnology in the Conservation of the Genus Castanea. Forests 2017, 8, 394. [CrossRef]
- Fernandes, P.; Tedesco, S.; da Silva, I.V.; Santos, C.; Machado, H.; Costa, R.L. A New Clonal Propagation Protocol Develops Quality Root Systems in Chestnut. Forests 2020, 11, 826. [CrossRef]
- Merkle, S.A.; Viéitez, F.J.; Corredoira, E.; Carlson, J.E. Castanea Spp. Chestnut. In Biotechnology of fruit and nut crops; CAB International: UK, 2020; pp. 206–237.
- Pavese, V.; Ruffa, P.; Abbà, S.; Costa, R.L.; Corredoira, E.; Silvestri, C.; Torello Marinoni, D.; Botta, R. An In Vitro Protocol for Propagating Castanea Sativa Italian Cultivars. Plants 2022, 11, 3308. [CrossRef]
- Song, G.; Chen, Q.; Callow, P.; Mandujano, M.; Han, X.; Cuenca, B.; Bonito, G.; Medina-Mora, C.; Fulbright, D.W.; Guyer, D.E. Efficient Micropropagation of Chestnut Hybrids (Castanea Spp.) Using Modified Woody Plant Medium and Zeatin Riboside. Hortic Plant J 2021, 7, 174–180. [CrossRef]
- Vieitez, A.M.; Sänchez, M.C.; García-Nimo, M.L.; Ballester, A. Protocol for Micropropagation of Castanea sativa. In Protocols for Micropropagation of Woody Trees and Fruits; Springer Netherlands: Dordrecht, 2007; pp. 299–312.
- Marino, L.A.; Ruffa, P.; Mozzanini, E.; Patono, D.L.; Sereno, A.; Pavese, V. LEDs in Plant Tissue Culture: Boosting Micropropagation of Castanea sativa Cultivars. J Plant Growth Regul 2025, 44, 6046–6060. [CrossRef]
- Vieitez, F.J.; San Jose, M.C.; Ballester, A.; Vieitez, A.M. Somatic Embryogenesis in Cultured Immature Zygotic Embryos in Chestnut. J Plant Physiol 1990, 136, 253–256. [CrossRef]
- Corredoira, E.; Ballester, A.; Vieitez, F.J.; Vieitez, A.M. Somatic Embryogenesis in Chestnut. In Somatic Embryogenesis; Mujib, S., Samaj, J., Eds.; Springer-Verlag: Berlin/Heidelberg, 2005; pp. 177–199.
- Sauer, U.; Wilhelm, E. Somatic Embryogenesis from Ovaries, Developing Ovules and Immature Zygotic Embryos, and Improved Embryo Development of Castanea Sativa. Biol Plant 2005, 49, 1–6. [CrossRef]
- Sezgin, M.; Dumanoğlu, H. Somatic Embryogenesis and Plant Regeneration from Immature Cotyledons of European Chestnut (Castanea sativa Mill.). In Vitro Cellular & Developmental Biology - Plant 2014, 50, 58–68. [CrossRef]
- Sezgin, M.; Dumanoğlu, H. In Vitro Propagation Potential via Somatic Embryogenesis of the Two Maturing Early Cultivars of European Chestnut (Castanea sativa Mill.). Am J Plant Sci 2016, 07, 1001–1012. [CrossRef]
- Vieitez, F.J. Somatic Embryogenesis in Chestnut. 1995, 375–407. [CrossRef]
- Corredoira, E.; Ballester, A.; Vieitez, A.M. Proliferation , Maturation and Germination of Castanea sativa Mill Somatic Embryos Originated from Leaf Explants. Ann Bot 2003, 92, 129–136. [CrossRef]
- Corredoira, E.; Vieitez, A.M.; San José, M.C.; Vieitez, F.J.; Ballester, A. Advances in Somatic Embryogenesis and Genetic Transformation of European Chestnut: Development of Transgenic Resistance to Ink and Blight Disease. In Vegetative Propagation of Forest Trees; Park, Y.S., Bonga, J.M., Moon, H.-K., Eds.; National Institute of Forest Science (NIFoS): Seoul, Korea, 2016; pp. 279–301.
- Vieitez, F.J. Mass Balance of a Long-Term Somatic Embryo Cultures of Chestnut. In Proceedings of the Application of Biotechnology to Forest Genetics, BIOFOR-99; Espinel, S., Ritter, E., Eds.; Vitoria-Gasteiz, Spain, 1999; pp. 199–211.
- Corredoira, E.; Valladares, S.; Vieitez, A.M.; Ballester, A. Improved Germination of Somatic Embryos and Plant Recovery of European Chestnut Improved Germination of Somatic Embryos and Plant Recovery of European Chestnut. In Vitro Cellular & Developmental Biology - Plant 2008. [CrossRef]
- Seabra, R.C.; Pais, M.S. Genetic Transformation of European Chestnut. Plant Cell Rep 1998, 17, 177–182.
- Corredoira, E.; Montenegro, D.; San-José, M.C.; Vieitez, A.M.; Ballester, A. Agrobacterium-Mediated Transformation of European Chestnut Embryogenic Cultures. Plant Cell Rep 2004, 23, 311–318. [CrossRef]
- Corredoira, E.; San-José, M.C.; Vieitez, A.M.; Ballester, A. Improving Genetic Transformation of European Chestnut and Cryopreservation of Transgenic Lines. Plant Cell Tissue Organ Cult 2007, 91, 281–288. [CrossRef]
- Corredoira, E.; San-josé, M.C.; Ballester, A.; Vieitez, A.M. Genetic Transformation of Castanea sativa Mill. by Agrobacterium Tumefaciens. Acta Hortic 2005, 693, 387–393. [CrossRef]
- Yeung, E.C. Structural and Developmental Patterns in Somatic Embryogenesis. In In Vitro Embryogenesis in PlantsCurrent Plant Science and Biotechnology in Agriculture; Thorpe, T.A., Ed.; Springer, Dordrecht, 1995; Vol. 20, pp. 205–247 ISBN 978-94-011-0485-2.
- Garcia-Casado, G.; Collada, C.; Allona, I.; Soto, A.; Casado, R.; Rodriguez-Cerezo, E.; Gomez, L.; Aragoncillo, C. Characterization of an Apoplastic Basic Thaumatin-like Protein from Recalcitrant Chestnut Seeds. Physiol Plant 2000, 110, 172–180. [CrossRef]
- Corredoira, E.; Valladares, S.; Allona, I.; Aragoncillo, C.; Vieitez, A.M.; Ballester, A. Genetic Transformation of European Chestnut Somatic Embryos with a Native Thaumatin-like Protein (CsTL1) Gene Isolated from Castanea sativa Seeds. Tree Physiol 2012, 00, 1–14. [CrossRef]
- Serrazina, S.; Martínez, M.T.; Valladares, S.; Del Castillo-González, L.; Francisco, M.; Berrocal-Lobo, M.; Piñas, E.; Piñeiro, P.; Malhó, R.; Costa, R.L.; et al. Overexpression of Ginkbilobin-2 Homologous Domain Gene to Enhance the Tolerance to Phytophthora cinnamomi in Plants of European Chestnut. BMC Genomics 2025. (in press).
- Colavolpe, M.B.; Vaz Dias, F.; Serrazina, S.; Malhó, R.; Lourenço Costa, R. Castanea crenata Ginkbilobin-2-like Recombinant Protein Reveals Potential as an Antimicrobial against Phytophthora cinnamomi, the Causal Agent of Ink Disease in European Chestnut. Forests 2023, Vol. 14, Page 785 2023, 14, 785. [CrossRef]
- Corredoira, E.; Valladares, S.; Vieitez, A.M.; Ballester, A. Chestnut, European (Castanea sativa). In Agrobacterium Protocols; Wang, K., Ed.; 2015; pp. 163–176.
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas Genome Editing and Precision Plant Breeding in Agriculture. Annu Rev Plant Biol 2019, 70, 667–697. [CrossRef]
- Walawage, S.L.; Zaini, P.A.; Mubarik, M.S.; Martinelli, F.; Balan, B.; Caruso, T.; Leslie, C.A.; Dandekar, A.M. Deploying Genome Editing Tools for Dissecting the Biology of Nut Trees. Front Sustain Food Syst 2019, 3, 491199. [CrossRef]
- Liao, H.; Wu, J.; VanDusen, N.J.; Li, Y.; Zheng, Y. CRISPR-Cas9-Mediated Homology-Directed Repair for Precise Gene Editing. Mol Ther Nucleic Acids 2024, 35, 102344. [CrossRef]
- Bewg, W.P.; Ci, D.; Tsai, C.-J. Genome Editing in Trees: From Multiple Repair Pathways to Long-Term Stability. Front Plant Sci 2018, 9, 412688. [CrossRef]
- Woo, J.W.; Kim, J.; Kwon, S. Il; Corvalán, C.; Cho, S.W.; Kim, H.; Kim, S.-G.; Kim, S.-T.; Choe, S.; Kim, J.-S. DNA-Free Genome Editing in Plants with Preassembled CRISPR-Cas9 Ribonucleoproteins. Nat Biotechnol 2015, 33, 1162–1164. [CrossRef]
- Santillán Martínez, M.I.; Bracuto, V.; Koseoglou, E.; Appiano, M.; Jacobsen, E.; Visser, R.G.F.; Wolters, A.-M.A.; Bai, Y. CRISPR/Cas9-Targeted Mutagenesis of the Tomato Susceptibility Gene PMR4 for Resistance against Powdery Mildew. BMC Plant Biol 2020, 20, 284. [CrossRef]
- Pavese, V.; Moglia, A.; Corredoira, E.; Martínez, M.T.; Torello Marinoni, D.; Botta, R. First Report of CRISPR/Cas9 Gene Editing in Castanea sativa Mill. Front Plant Sci 2021, 12, 1752. [CrossRef]
- Ballesteros, D.; Martínez, M.T.; Sánchez-Romero, C.; Montalbán, I.A.; Sales, E.; Moncaleán, P.; Arrillaga, I.; Corredoira, E. Current Status of the Cryopreservation of Embryogenic Material of Woody Species. Front Plant Sci 2024, 14, 1337152. [CrossRef]
- Corredoira, E.; San-José, M.C.; Ballester, A.; Vieitez, A.M. Cryopreservation of Zygotic Embryo Axes and Somatic Embryos of European Chestnut. Cryo Letters 2004, 25, 33–42.
- Gaidamashvili, M.; Khurtsidze, E.; Kutchava, T.; Lambardi, M.; Benelli, C. Efficient Protocol for Improving the Development of Cryopreserved Embryonic Axes of Chestnut (Castanea sativa Mill.) by Encapsulation–Vitrification. Plants 2021, Vol. 10, Page 231 2021, 10, 231. [CrossRef]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Cryopreservation of Nucellar Cells of Navel Orange (Citrus sinensis Osb. Var. Brasiliensis Tanaka) by Vitrification. Plant Cell Rep 1990, 9, 30–33. [CrossRef]
- Vidal, N.; Sánchez, C.; Jorquera, L.; Ballester, A.; Vieitez, A.M. Cryopreservation of Chestnut by Vitrification of in Vitro-Grown Shoot Tips. In Vitro Cellular & Developmental Biology - Plant 2005, 41, 63–68. [CrossRef]
- Vidal, N.; Vieitez, A.M.; Fernández, M.R.; Cuenca, B.; Ballester, A. Establishment of Cryopreserved Gene Banks of European Chestnut and Cork Oak. Eur J For Res 2010, 129, 635–643. [CrossRef]
- Carneros, E.; Berenguer, E.; Pérez-Pérez, Y.; Pandey, S.; Welsch, R.; Palme, K.; Gil, C.; Martínez, A.; Testillano, P.S. Small Molecule Inhibitors of Human LRRK2 Enhance in Vitro Embryogenesis and Microcallus Formation for Plant Regeneration of Crop and Model Species. J Plant Physiol 2024, 303, 154334. [CrossRef]
Figure 2.
Schematic representation of the “lab-to-field” pipeline illustrating key strategies for developing improved European chestnut genotypes with enhanced resistance to pathogens, pests, and climate change. The diagram highlights integrative approaches, including molecular breeding, genomic selection, gene editing, phenotypic screening, and micropropagation.
Figure 2.
Schematic representation of the “lab-to-field” pipeline illustrating key strategies for developing improved European chestnut genotypes with enhanced resistance to pathogens, pests, and climate change. The diagram highlights integrative approaches, including molecular breeding, genomic selection, gene editing, phenotypic screening, and micropropagation.
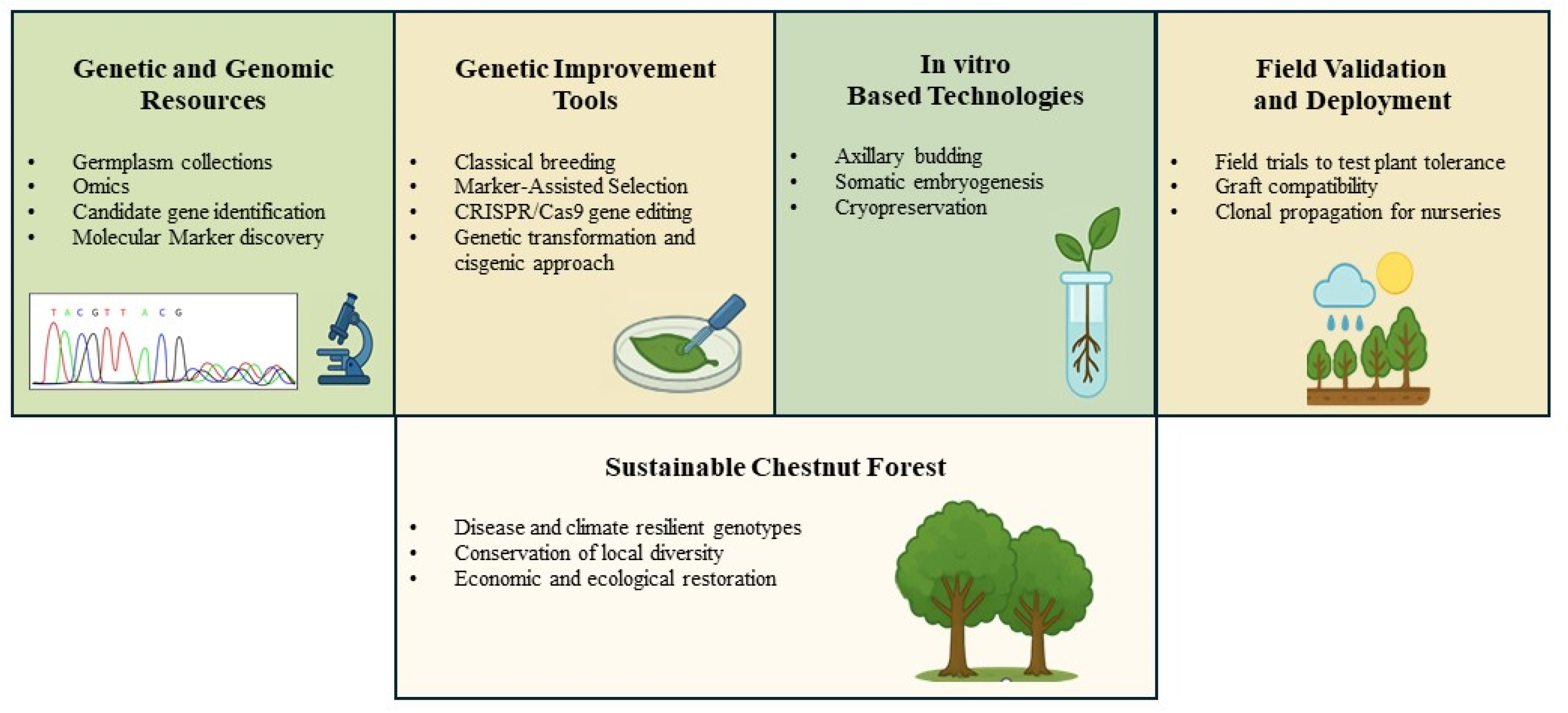
Table 2.
Biotechnological tools to study chestnut’s resistance to Cryphonectria parasitica, focused on C. sativa (tolerant/susceptible), C. dentata (susceptible), and C. mollissima (resistant).
Table 2.
Biotechnological tools to study chestnut’s resistance to Cryphonectria parasitica, focused on C. sativa (tolerant/susceptible), C. dentata (susceptible), and C. mollissima (resistant).
| Methodology | Species / Genotypes | Results / Findings | References |
|---|---|---|---|
|
Physiological and biochemical responses |
C. sativa | Reduced photosynthetic pigments; Increased APX, POD, SOD; Accumulation of proline and MDA |
[116] |
|
Biological control CHV1 hypovirus |
C. sativa | Mitigation of disease severity via hypovirulent strains | [117] |
|
Chitinase and β-1,3-glucanase expression |
C. sativa | Systemic induction; Higher activity with hypovirulent strains; Antifungal activity of Ch3 protein |
[122,123,124,125] |
|
Susceptibility gene expression profiling |
C. sativa | Upregulation of pmr4 and dmr6; Suppression of SA-mediated responses |
[107] |
|
SA accumulation studies (metabolite and transcriptome analysis) |
C. sativa C. dentata C. mollissima |
Higher SA levels with hypovirulent strains; SA-related gene expression in canker tissue |
[123,128] |
| Genomic and transcriptomic studies |
C. mollissima ‘Vanuxem’ |
Identification of resistance genes; Rapid wound response and cell wall lignification |
[130] |
|
Chloroplast genome sequencing |
Wild C. mollissima | 131 genes involved in stress responses and metabolic regulation | [132] |
| Genetic mapping and GWAS | C. dentata × C. mollissima | Resistance loci on all chromosomes; Candidate resistance and susceptibility genes identified |
[63,134,135] |
|
Transgenic OxOexpression |
C. dentata | Oxalate oxidase degrades oxalic acid from pathogen; field trials and regulatory review ongoing | [136,137] |
| Transcriptome comparison via pyrosequencing |
C. dentata vs. C. mollissima |
Differential expression of defense genes; Stronger defense response in C. mollissima |
[128,140] |
| Transcriptomic profiling | Wild C. mollissima ‘HBY-1’ |
283 DEGs in metabolism and defense pathways; Early JA pathway activation |
[147] |
|
Tannin profiling (metabolite analysis) |
C. mollissima C. dentata C. sativa |
Prevalence of hamamelitannin in C. sativa and C. dentata; Higher vescalagin and castalagin in C. mollissima - inhibition of fungal enzymes |
[151,152,153] [113](and references within) |
APX: Ascorbate peroxidase; ch3: Chitinase; CHV1: Cryphonectria hypovirus 1; DEGs: Differentially expressed genes; dmr6: Downy Mildew Resistant 6; GWAS: genome-wide association studies; JA: Jasmonic acid; MDA: Malondialdehyde; OxO: Oxalate oxidase; pmr4: Powdery Mildew Resistant 4; POD: Guaiacol peroxidase; SA: Salicylic acid; SOD: Superoxide dismutase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



