
Submitted:
30 October 2025
Posted:
03 November 2025
You are already at the latest version
Abstract
Traumatic brain injury (TBI) is a leading cause of death and disability with broad heterogeneity in recovery outcomes particularly noted in pediatric patients. Children post-TBI are vulnerable to aberrant neurodevelopment specifically in structural and functional neural networks as they correlate with cognitive, behavioral, and motor function outcomes. Consideration for sex as a biological variable which innately influences neuroanatomy, neurodevelopment, and functional organization may elucidate risk factors for negative outcomes in pediatric TBI. For example, TBI damage in sexually dimorphic neural structures and networks may explain deficits in social cognition, working memory, as well as internalizing and externalizing behaviors, which differentially impact the quality of life in male versus female TBI patients. However, characterization of sex in conjunction with developmental patterns in normal and injured pediatric populations, is limited due to small sample sizes, low representation of females, lack of longitudinal data, and the utilization of analyses which are not sensitive enough to detect subtle differences in TBI pathologies and recovery between the sexes. This review aims to analyze and synthesize the existing evidence regarding the influence of sex on the developmental trajectories of neuroanatomical structures, white and gray matter compartments, and the network disruptions that align with sex-specific functional recovery outcomes following pediatric TBI. Delineation of these sex influences will facilitate better precision-based medicine approaches to improve patient outcomes.
Keywords:
traumatic brain injury
; pediatric
; sex
; neuroanatomy
; neurodevelopment
; white matter
; gray matter
; connectivity
1. Introduction
Pediatric traumatic brain injury (TBI) is well established as a leading cause of mortality and disability, causing an immense public health burden with >500,000 emergency department (ED) visits and 2,000 deaths annually in the United States [1,2]. Despite declining incidence of severe pediatric TBI, likely due to measures taken to improve safety and education, TBI-related ED visits have continued to rise, due in part to increasing incidence and reporting of mild pediatric TBI cases [1]. Pediatric risk for TBI is of particular concern due to coalignment with critical neurodevelopmental periods and disruption of brain development (e.g., delayed brain growth, abnormalities in white matter microstructure) which leads to negative functional outcomes (e.g., learning, memory, motor function) [3]. Given the heterogeneity in TBI pathophysiology and recovery, research efforts have focused on patient- and injury-specific delineation of risk factors to coordinate precision medical approaches and potential therapeutic targets. Sex has recently been identified as a key biological variable in TBI pathophysiology and a mediator of recovery outcomes.
Human and preclinical TBI studies have shown sex-related differences in cognitive, behavioral, and motor dysfunction, in addition to somatic symptom presentation and persistence in adults [4,5,6]. Neuroprotective effects of estrogen and progesterone may partially explain these sex-dependent recovery outcomes, but studies have been unable to draw consistent conclusions, as therapeutic administration of sex hormones have failed in clinical trials [7]. Additionally, hormonal profiles fail to account for observed sex effects in pediatric populations where sex hormone production is limited, thus suggesting the role of sex to be more complex than previously determined.
Age is well defined as an important predictor for functional recovery post-TBI [8,9]. Age and sex interactions further influence TBI recovery in children as sex differences in developmental patterns can influence vulnerability to chronic perturbations in response to damage. Consideration of sex in pediatric TBI pathology and recovery has been detrimentally understudied, often due to low sample sizes and underrepresented patient populations. Convergent evidence suggests delineation of sex effects could have important implications for TBI pathophysiology and treatment development. This review aims to analyze and synthesize the existing evidence regarding the influence of sex on the developmental trajectories of neuroanatomical structures, white and gray matter compartments, and the network disruptions that align with sex-specific functional recovery outcomes following pediatric TBI.
Search Strategy and Selection Criteria
The articles in this review were retrieved using the following electronic databases: PubMed, MEDLINE, Google Scholar, and Web of Science. The search was limited to articles published between 1986 and 2025. Search terms were identified in the title, abstract, and key words using the following search terms: traumatic brain injury, TBI, pediatric/juvenile/children, sex, gender, gray matter, white matter, basal ganglia, limbic system, cognition, behavior, motor function, social cognition, attention, memory, working memory, atrophy, neurodegeneration, alcohol, putamen, globus pallidus/pallidum, hypothalamus, pituitary gland, thalamus, hippocampus, cerebellum, diffuse axonal injury, structural connectivity, diffusion tensor imaging, microstructure, cortical thickness, total gray matter volume, gray matter density, symmetry, functional connectivity, intrinsic connectivity, extrinsic connectivity, resting-state fMRI/rs-fMRI, task-based fMRI/tb-fMRI, default mode network/DMN, sensorimotor network, and attentional network.
2. Sexual Dimorphisms in Specific Brain Structures
Recent evidence suggests that fundamental sex differences in brain anatomy and neurodevelopment may contribute to the observed differences in TBI severity and consequent prognosis between male and female children [10,11,12,13]. Even when controlling for the larger brain volumes of males, these sex differences are maintained in multiple gray and white matter structures throughout the brain from pediatrics to adulthood [14,15,16]. Specifically, sex effects within the basal ganglia, limbic system, and cerebellum, among other brain structures, may influence the complex nature of functional outcomes post-TBI (Figure 1). Considering the heterogeneity of TBI recovery, fundamental sex differences in cortical and subcortical structure size and development may influence patient recovery more than previously thought and thus remains a critical topic of intensive discussion.
2.1. The Basal Ganglia
The basal ganglia consist of a group of connected subcortical structures located at the base of the forebrain. These structures play critical roles in decision making, reward-based learning, motor function, addiction, and contribute to unique and specialized deficits following damage from neural injury. In 2024, Backhausen et al. found significantly larger volumes in adolescent and young adult males versus females across subcortical regions including the caudate nucleus, putamen, globus pallidus, amygdala, and nucleus accumbens [17]. This research expanded upon previous studies in which sex-specific structural differences were observed in the putamen and globus pallidus in children and adults [15,16]. Interestingly, sex differences in age-related trajectories were also observed in these basal ganglia structures with normal healthy adult males demonstrating a higher degree of progressive atrophy in basal ganglia structures as they age relative to females [14,18]. Therefore, TBI-induced damage and tissue atrophy in the basal ganglia in youth may disturb this natural age-related trajectory, thus potentially resulting in disproportionally greater cognitive and emotional deficits in males as they age through adulthood [17,19,20,21,22].
In addition to differing psychopathology between males and females, disturbance of the basal ganglia and its development following TBI has been linked to increased addiction-related behaviors [23,24]. Seminal animal studies have demonstrated marked sex differences in reward processing and reinforcement learning, both of which are implicated in drug addiction [25,26,27]. Recent human functional MRI (fMRI) investigations supported these findings as reward prediction error-related brain activity was enhanced in women compared to men and to an even greater extent when estradiol levels were elevated in both sexes. This is likely due to the effects of estrogen on the mesocorticolimbic dopaminergic pathway as it passes through the basal ganglion structures [28,29]. Because substance use disorders and alcohol addictions are common long-term outcomes post-TBI, researchers believe this may be due to tissue damage within the basal ganglia. One study found that neural activity in the fronto-basal ganglia network of TBI patients 9-10 years old predicted increased odds of sipping alcohol by ages 11-13, with males reporting significantly greater alcohol sipping than females [30]. In a study of students enrolled in grades 7-12, Ilie and colleagues found that males with a history of TBI were also more likely to report daily nicotine use than females [23].
It is important to note that future studies evaluating post-TBI addiction should also consider alternative addiction-related behaviors such as internet addiction, social media addiction, gaming disorders, and mobile phone addiction, all of which are more accessible to children and more socially acceptable for females. For example, an assessment of childhood TBI patients aged 0-17 years old, found that males and females displayed significant differences in internalizing versus externalizing behaviors such that males were more likely to report substance abuse and females were more likely to report anxiety and depression [31]. Young females with depressive symptoms had significantly higher risks of internet addiction relative to males [32], while also demonstrating a higher risk of mobile phone addiction [33,34]. These differences in psychosocial norms and how they relate to reporting symptoms of physical and mental health among females versus males may therefore influence the documentation of addiction disorder in TBI patient populations.
2.1.1. The Putamen
The putamen is involved in everyday functions including speech and language, reward processing, cognition, and motor control. During normal brain development, the putamen has both age-related volume decreases as well as sex-related volumetric differences, with larger putamen volumes noted in male children beginning around 8 years of age [35,36,37,38]. Females and males also have different putamen development trajectories, where putamen volumes in female children decrease until approximately 20 years of age. After 20 years old, the rate of volumetric atrophy decreases in females. Comparatively, putamen volumes in male children have a slower decrease until approximately 16 years of age. After 16 years old, the rate of volumetric atrophy increases in males [17,39,40]. In moderate to severe TBI patients, volume loss in the putamen can contribute to sex-related differences in cognitive function, attention, emotion control, motor function, and behavior [20,41,42]. Therefore, the age of the TBI patient and the corresponding point in which the putamen is undergoing inherent volumetric decreases, exacerbated putamen loss post-TBI may explain the higher prevalence of functional deficits [43] and cognitive and emotional dysfunction [14,44,45] in males.
Putamen volume may also serve as a structural neural marker for many neuropsychiatric and neurodegenerative conditions [46]. Comparable neurological studies have found that putamen volume is also significantly reduced in patients diagnosed with Huntington’s disease [47], dementia with Lewy bodies [48], Alzheimer’s disease [49], multiple sclerosis [50], attention deficit hyperactivity disorder (ADHD) [51], and major depression [52]. In a study investigating subcortical gray matter volume loss and its relation to executive dysfunction following TBI, loss of putamen volume was significantly associated with poorer task switching performance [53]. Lower functional independence scores were also found to be correlated with putamen atrophy in an injury and sex differential manner [42]. In a 5 year longitudinal study, Simeone and colleagues found TBI induced progressive volumetric atrophy in subcortical gray matter structures, including the putamen, to occur at a similar rate to aggressive neurodegenerative diseases and was associated with lowered episodic memory and executive function compared to healthy controls [54]. In mice, males were shown to be more susceptible to neurodegeneration following TBI in a Parkinson’s Disease model [55]. Given the findings of Jenkins et al on distinct striatal dopamine abnormalities in TBI and Parkinson’s Disease patients, disrupted putaminal dopamine signaling should be considered as a potential mechanism for TBI. This could also indicate TBI is a premorbid risk factor for Parkinson’s [56]. Therefore, a patient’s biological sex, age, and associated putamen volume may be a premorbid risk factor that predisposes patients, particularly males, to worse outcomes post-TBI.
2.1.2. The Globus Pallidus
The globus pallidus, also known as the pallidum when considering both the internus and externus halves, controls conscious and proprioceptive movement and projects into all the other basal ganglia structures. Since the globus pallidus is a critical structure in basal ganglia circuits, characterization of its developmental trajectory is essential to gaining a better understanding of motor and non-motor dysfunctions as a consequence of TBI. This gray matter structure possesses both age and sex-dependent volumetric differences as males have a greater increase in volume until 16 years of age, followed by a sharp decline later in life. This contrasts with female neurodevelopment, as the globus pallidus volume has been found to steadily increase until 24 years of age [14,17]. Understanding how these trajectories influence motor behavior deficits following TBI has become a major research objective, as recent evidence suggests that volume loss in the globus pallidus due to diffuse injury from TBI is significantly correlated with functional deficits [57,58]. Furthermore, Gooijers et al. demonstrated an association in adults between globus pallidus volumes and alternate finger tapping, thus demonstrating worse bimanual behavioral deficits in the TBI group, which were correlated with reduced volume in the globus pallidus [43]. This was recently supported, as United States military service members with a history of mild TBI demonstrated an association between significantly smaller globus pallidus volumes and reductions in processing speed performance involving motor output when compared to controls [59]. Studies such as these suggest volumetric measures of the globus pallidus may serve as valuable prognostic markers of chronic neural pathology, which complicates motor control rehabilitation in TBI patients. Interestingly, Niemann et al. demonstrated in older adults that motor fitness positively correlated with the volume of the globus pallidus and that coordination training increased globus pallidus volumes leading to improved patient recovery [60]. This suggests that the globus pallidus may serve as a novel therapeutic target for more individualized rehabilitation programs and/or treatment interventions such as deep brain stimulation.
2.2. The Limbic System
The limbic system is also susceptible to damage following TBI which can result in chronic negative recovery outcomes in cognition, behavior, and autonomic nervous system regulation [20,61]. Sexually divergent limbic structures include the hypothalamus, thalamus, and hippocampus. The hypothalamic-pituitary axis is also critically important for the endocrine system, which influences many aspects of neurodevelopment including homeostatic function.
2.2.1. The Hypothalamus and Pituitary Gland
Recent studies involving the limbic system have revealed MRI-based differences in male and female hypothalamic and pituitary gland volumes with 7-9 year old males exhibiting significantly larger hypothalamus and pituitary gland volumes, compared to age-matched females [16,62,63,64]. Interestingly, this trend is reversed as 14-17 year old females possess significantly larger pituitary gland volumes, compared to age-matched males [64]. Hypothalamic development rates also differ between males and females, with males reaching maximum hypothalamus volumes at age 18, compared to females who reach maximum hypothalamus volumes earlier at 15-16 years old [65]. Given the role of the hypothalamus in maintaining homeostasis and regulating the endocrine system via hormonal signaling to the pituitary gland, the structural vulnerability of both the hypothalamus and pituitary—coupled with sex-specific differences in brain volume and developmental trajectories—may partially account for the diverse hormonal deficiencies observed in TBI patients.
A comprehensive review of the literature spanning 22 years found TBI-related hormonal deficits ranged from 22.5-86% and multiple hormonal deficits ranged from 5.9-50% in the studied pediatric population [66]. Additional studies found hypopituitarism incidence ranged from 5-70% in TBI patients, likely due to the lack of consistency in research methodologies; therefore, making hypopituitarism one of the most challenging morbidities to correctly identify and treat [67,68]. Hypopituitarism has been found to delay puberty in pediatric TBI populations and can cause physical and psychological dysfunctions that last well into adulthood [20,69,70]. Additional studies by Gray et al. and Prodam et al. go on to report a clear association between hypopituitarism and adverse cognitive outcomes such as memory, attention, language, physical conditioning, and mood disorders post-TBI [71,72]. In addition to hypopituitarism outcomes, specific growth hormones and gonadotropins—which are produced by the pituitary gland and regulate sexual development, reproduction, and growth—are commonly deficient in TBI patients [20,67,73]. This may help to explain sex-related differences in TBI recovery outcomes between male and female children [74,75]. Niederland et al. reported 42% of all TBI children showed insufficient growth hormone response and 73% of these cases were males [76]. Agha et al. determined decreased sex hormones affected primarily males post-TBI with lasting effects on fertility, psychosexual function, and general well-being. It was also found to affect the level of energy and motivational status, and not least, the mortality rate of male patients secondary to cardiovascular disease [77]. Studies investigating the intricate relationship between hypothalamic and pituitary gland volumes and development in collaboration with TBI location and severity are important to identify potential endocrine complications, which could be life-threatening and impact patients’ overall quality of life.
2.2.2. The Thalamus
The thalamus primarily functions to receive, process, and relay sensory information to the cerebral cortex, thereby contributing to states of consciousness, attention, cognition, and memory. Recent MRI-based studies revealed sex-based differences in thalamic volumes and developmental trajectories from childhood through adulthood [14,15,16]. Although there is some discrepancy in the literature on which sex has greater thalamic volumes, it is likely that the age ranges and sample sizes of these studies prevent cohesive results. For example, Xie et al. and Sowell et al. demonstrated sex affected the size of the thalamus, with female children and adolescents having larger thalami relative to gray matter and brain volumes [36,78]. More recently, Wagner and colleagues saw lateralization effects in pediatric populations where the females’ left thalamus volume increased by 17.3% and peaked at 16-17 years old, and the right thalamus volume increased by 11.4% and peaked at 11-12 years old [65]. Comparatively in males, the left thalamus volume increased by 23.2% and peaked at 15-16 years old, and the right thalamus volume increased by 20.7% and peaked at 17-18 years old [65]. This illustrates the delicate balance between sex and age on thalamic development. Given these differences, it is important to consider that the impact of TBI on the thalamus may vary depending on the child's age at the time of injury due to ongoing brain development. The thalamus is particularly vulnerable to shearing effects during TBI and consequent interruptions to thalamic development and volume loss have been shown to reduce cognitive and motor performance [20,43]. Unfortunately, there is limited research on symptoms and functional recovery directly related to sex differences in thalamic damage in pediatric TBI populations.
Reduced cortical thickness and deformation in key thalamic subregions that operate as part of the dorsolateral prefrontal cortex circuitry were significantly correlated with decreased executive functioning in pediatric patients [79]. Although not directly examined, understanding the thalamus' role in executive functions—such as planning and emotional regulation—adds nuance to the findings reported by Keenan and colleagues. Their study indicates that males aged 6-11 years old exhibit poorer executive function outcomes, particularly in emotional control, with additional evidence suggesting that male children demonstrate greater deficits in planning and organizational tests [80]. Another study found male children alone were predisposed to new onset compulsions which were significantly associated with thalamic lesions induced by TBI [81]. Given the different volumes and developmental trajectories of the thalamus in males and females, TBI-induced tissue damage could exacerbate atrophy trajectories.
2.2.3. The Hippocampus
The hippocampus is an integral structure of the limbic system and is primarily associated with declarative and spatial memory. More specifically, the hippocampus is responsible for autobiographical memory and the conversion of short-term memory to long-term memory [82]. During pediatric development, the hippocampus undergoes increased growth trajectories from 8-10 years old and then peaks in volume at around 16 years old [65]. Although sex differences in volumetrics have been widely reported, some researchers state female children possess greater hippocampal volumes, which were maintained throughout adolescence [16,82], while others reported larger volumes in males [83]. It is important to note that in order to avoid such discrepancies, future investigations should not only account for differences in total brain volumes between males and females, but also avoid voxel-based morphometry technologies which have been found to be too ambiguous to consider sex-based volumetric differences and age trajectories [16,82,83]. With these analytics considered, the hippocampus was found to be significantly smaller in males compared to females at 8-15 years old while controlling for both total brain volume and body size [84,85,86].
Sex-related differences in hippocampal development trajectories also remains a hotly debated topic. Early studies reported smaller hippocampi volumes in males were accompanied by decreased hippocampal development rates when compared to females [87]. Meanwhile, follow-up longitudinal studies have not found sex-related differences in developmental trajectories [88,89]. Observational neuroimaging data suggest that differences in hippocampal development may also be sensitive to sex steroid and chromosome effects. In a sample of 8-15 year old males and females, Neufang et al. found a larger hippocampus volume was related to increased levels of testosterone in both sexes [86]. Peper et al. showed changes in sex steroid availability during puberty and adolescence triggered structural reorganization of the hippocampus in the developing brain, as early pubertal females had less gray matter density in the hippocampus compared to non-pubertal females [90,91,92]. Collectively, variations in hippocampal segmentation methods may contribute to observed differences in developmental trajectories [93] and it is thereby recommended that conclusions regarding sex-biases in volumetrics and the timing of hippocampal maturation must include functional data analysis tools (e.g., multi-atlas automated subcortical segmentation, semi-parametric spline-based methods, and surface area change trajectory models) of milestone timing for multiple brain regions [94].
TBI-induced tissue damage to this critical brain region also demonstrates sex-bias. Research suggests inherent differences in hippocampal volume between male and female children results in differing hippocampal function and recovery. Because a loss of >20-30% of the dorsal hippocampus and >39-52% of the ventral hippocampus is required to observe hippocampal-dependent learning impairments [95], inherently smaller hippocampi volumes in male children may help explain why males are more susceptible to cognitive deficits including greater impairments in learning and declarative memory when compared to females [82,96,97]. Conversely, increased hippocampi volumes in females may help explain differences in verbal and non-verbal memory test performance post-TBI with male children performing significantly worse [98]. Furthermore, it has been hypothesized that inherently larger hippocampi volumes in females may contribute to improved encoding of long-term memories as well as spatial processing and navigation [99,100]. In the context of hippocampal involvement in the social brain network, male mice have demonstrated greater impairments in adult psychosocial behaviors—particularly in social recognition—following pediatric TBI. These deficits have been linked to alterations in neuronal morphology within the hippocampus and medial prefrontal cortex. [101]. Davila-Valencia and colleagues further found altered neurogenesis within the hippocampus to have a trending sex effect, with females showing reduced proliferation and short-term survival of newborn cells, which was further linked to observed sex effects in behavioral testing [102]. Collectively, these studies highlight the need to consider differences in hippocampal volume, developmental rates, and atrophy trajectories between males and females while assessing hippocampal tissue damage post-TBI. This could help explain variabilities in learning and memory deficits and improve prognostic measurements.
2.3. The Cerebellum
The cerebellum is responsible for a diverse number of functions ranging from motor control to cognition and emotion. Highly connected to the rest of the brain, the cerebellum influences specific aspects of cognitive development including executive function, cognitive planning, personality, and learning [103,104,105]. Recent evidence has found that cerebellar gray matter is closely related to cognitive function and volumetric analysis can be reliably used as a predictor of cognitive performance in children [104]. Furthermore, both sex and age have been identified as key mediators of cerebellar development [103,104,105]. Although male children demonstrated greater absolute gray matter volumes than females, distinct cerebellar lobules have been found to be larger in females when compared to males [15,103,106]. Female cerebellar volume has been found to peak around 12 years of age, as opposed to males who peak closer to 16 years of age, thus providing further evidence of the interplay between age and sex [83,103]. Additionally, the cerebellum develops in an anterior-to-posterior gradient in 6-10 year old children, where the posterior gray matter volume is larger than that of the anterior [105]. This white-gray matter ratio adapts from a negative to a positive relationship with cognitive functioning in adolescence, thus indicative of cognitive function acquisition during childhood [103,105].
Evidence for sex-based differences in cerebellum-related functional recovery following TBI also exists at the genetic level. For example, cerebellar damage is linked to circadian rhythm disruption, including sleep wake phase disorders, through altered clock gene expression. Sgro and colleagues demonstrated sex differences in expression of Clock and rev-erb-α genes in response to mild TBI in adolescent rats, wherein females showed reduced circadian time 0 expression, with sex-specific patterns for the following 24 hours [107]. Furthermore, knockout of the RBM5 gene in mice was associated with more visual deficits in males following TBI than females, which the authors thought to be related to sexually dimorphic effects of knockout on regulation of the RIMS2 gene [108]. Understanding sex differences in disrupted gene expression in the cerebellum following TBI offers unique insight to functional recovery challenges and offers therapeutic targets to overcome them. This is important as cerebellar regions can be specifically tied to cognitive functions and disruption can induce neuropsychiatric dysfunction [103,106]. Collectively, these cerebellar differences and their consequential impact on executive function and cognition, add a level of nuance to the understanding of how cortical atrophy of the cerebellum influences functional outcomes post-TBI.
3. Sexual Dimorphisms in Specific White and Gray Matter Compartments
White and gray matter tissues have different physical, metabolic, and cytoarchitectural properties which make them uniquely vulnerable to TBI effects. These unique variables influence mechanical transmission of force and result in distinct pathophysiology and neurodegeneration. White matter is composed of axons that originate and terminate in gray matter which can be myelinated or unmyelinated depending on developmental stage and specific function. White matter is also comprised of oligodendrocytes, which function to myelinate the axons, and fibrous astrocytes, which serve a wide range of functions including modulation of inflammation [109,110]. White and gray matter compartments have specialized roles and effects on cognitive function, including processing speed, short- and long-term memory, and attention, as well as behavioral functions, such as mood regulation. Damage to white and gray matter compartments caused by TBI are intrinsically related, as neurodegeneration of one frequently results in progressive loss of the other. Both white and gray matter have demonstrated sex-mediated differences, which in the context of TBI, could further affect the resiliency of tissue to damage.
3.1. White Matter
White matter is a highly organized tissue with its own cytoarchitecture and developmental trajectories that play an essential role in brain function. White matter function is centered around communication, wherein signals sent between gray matter regions are propagated by the myelinated axon tracts [111]. White matter microstructure integrity across the complex network of connections is integral to brain function and damage is becoming increasingly associated with cognitive dysfunction and psychiatric disorders [111]. However, there are a broad range of influencing factors, including sex, genetics, and environmental cues, that affect white matter development and resiliency to damage [111,112,113]. The interplay of these factors in combination with white matter damage could result in increased vulnerability to dysfunction, as signals are no longer sent or received at their normal rate. While sex differences in adult white matter have been firmly established in the literature, pediatric white matter sex differences have been investigated to a lesser extent [112,114]. Previous studies suggest sex differences in white matter are dependent on gonadal hormones and consequently appear during pubertal development [115,116]. However, Lawrence and colleagues found that sex influences in microstructure likely begin before the age of 10 [112]. These discrepancies are thought to be the result of insufficient diffusion tensor imaging (DTI) sensitivities and small study sizes with wide and variable age ranges [112]. Furthermore, as global white matter increases from childhood to adolescence, sex is seen to play a greater role in microstructure organization as indicated by diffusion metrics [115].
Shearing forces sustained during TBI cause diffuse axonal injury which is correlated with white matter microstructure damage. Given the essential role of white matter in cortical communication, disruptions due to injury have the potential to induce long-term dysfunction. Growing evidence suggests that even mild TBI-induced white matter damage can result in chronic deficits in cognition and behavior [117,118]. Of particular concern is pediatric TBI patients due to their unique alignment of a high incidence demographic with important neurodevelopmental periods. Following pediatric TBI, regional decreases in fractional anisotropy (FA), a key DTI-based measure of white matter integrity, were found to correlate with 12-month post-injury IQ scores. In accordance with the progression of abnormal FA values seen at chronic timepoints, neural repair and reorganization appeared to occur in a regional pattern [117]. White matter microstructure damage, as evaluated by FA and diffusivity measures, has been shown to be predictive of chronic recovery outcomes [118]. Long-term impacts of TBI, which are correlated with white matter microstructure and integrity, include deficits in cognitive functioning, such as social cognition and attention, as well as mood disorders [119,120,121]. Fundamental sex differences in regional white matter metrics and development in pediatric TBI may mediate recovery outcomes. Sex differences in FA measures localized to the uncinate fasciculus following TBI have been demonstrated to be predictive of time to symptom resolution in adults [122]. Regionally specific sex differences in white matter microstructure, as indicated by FA and diffusion metrics, are likely to be implicated in TBI recovery and may influence progression of negative cognitive and behavior outcomes as a function of predisposition in males or females.
Support also exists for sex-related differences in mood disorders, including depression and bipolar disorder [123], whereas TBI has also been documented as a risk factor for these mood disorders. This begs for further investigation of sex differences in the neuroanatomical foundations of these disorders and how damage caused by TBI can result in parallel conditions. Sex was found to influence FA values in white matter microstructure connecting the prefrontal cortex and limbic system, which were associated with the differing relationships adult males and females have with stress reactivity [124]. Novel Conduct Disorder and Oppositional Defiant Disorder were observed in approximately 14% of moderate to severe pediatric TBI patients with a significant association with frontal white matter lesions [125]. These behavioral disorders were further mediated by sex, where male children were more likely to develop comorbid ADHD, as opposed to increased predisposition to comorbid depression in females [125]. Nishat and colleagues found a correlation between disrupted superficial white matter maturation and increased internalizing behavior, independent of premorbid conditions, in female children following TBI [126]. This research builds upon previous findings in which increased premature white matter maturation in deep and superficial white matter in females following TBI was further associated with worse performance on cognitive assessments including the Picture Vocabulary Test and Pattern Comparison Processing Speed Test of the NIH Toolbox [127]. Alternatively, adult male athletes with sports-related concussion were found to have increased white matter microstructure impairment compared to females with similar injuries and reported worse symptoms [128]. This research may indicate important age and sex interactions which influence TBI recovery. White matter in female children may be less resilient to TBI compared to male children, thus leading to less cognitive recovery and increased vulnerability to internalizing behaviors. However, adult males appear to show a different response, which further delineates a unique pathophysiology that is dependent on both sex and the developmental period.
Sex differences in the morphometry of white matter tracts connecting the orbitofrontal cortex and nucleus accumbens, which are implicated in reward and punishment behaviors, were identified and linked to pubertal status in male, but not female, adolescents [129]. Additionally, the maturation of the microstructure was linked to sex differences in punishment sensitivity, with male adolescents with more fiber density and cross-section in the tract reporting less punishment sensitivity [129]. Similarly, a cross-sectional study found FA in the accumbofrontal tract peaked higher and earlier in male adolescents compared to female adolescents (13.9 vs. 18.6 years of age, respectively) [130]. These findings describe neural underpinnings which may be associated with increased risk-taking behavior in male adolescents, and indeed converge with the results of a longitudinal study that found sensation-seeking relative to impulse control trajectories were higher, with longer windows of increased disparity, in male compared to female adolescents [131]. Given the involvement of the reward and punishment system, and its interplay with the cognitive control system, addiction and conduct disorders may exhibit some shared neural network correlations, thus lending further context to sex-influenced behavior outcomes post-TBI. Disruptions in white matter have been limitedly characterized for alcohol abuse in adults, showing reduced FA in major white matter structures such as the corpus callosum, with greater reductions for males [132,133]. These findings remain to be confirmed in youth populations as investigation into white matter microstructure damage following TBI and its correlation with subsequent addiction outcomes overall is lacking. Given the evidence for innate sex differences in the white matter tracts associated with addiction pathology and the sensitivity of this tissue to TBI-related damage, future studies should consider sex as an underlying variable in propensity for addiction outcomes.
3.2. Gray Matter
Total gray matter volume is generally accepted to decline after it peaks around 2 years of age in humans [134]. However, it has been demonstrated that while gray matter volume decreases, gray matter density continues to increase with strong age and sex associations [134]. For example, two brain regions may have the same gray matter volume, but one could have a higher gray matter density if the neurons are more tightly packed within that region. While females have lower total gray matter volume, their gray matter density is higher than that of males [134]. Evidence has also shown the variability of gray matter volume throughout the brain to be associated with sex from pediatric to geriatric demographics [135]. Additionally, gray matter symmetry, which is thought to be established during neurodevelopment, has been shown to be influenced by sex both in total gray matter volume as well as within specific structures and regions [136,137]. A study spanning pediatric to adult data found female brains were more symmetrical globally, with some specific structures and regions showing greater symmetry in males [136]. Comparatively, a pediatric-only study did not find a global sex effect on symmetry, but did observe that female children exhibited more regions of asymmetry, while male children showed a greater magnitude of asymmetry within specific regions [137]. There are inconsistencies in the literature in regards to the sex effect on total symmetry as well as in the pallidus, putamen, and Hesch’s Gyrus [136,137]. However, differences in sample size, region of interest, and analysis methods must be considered as contributors for these discrepancies. In a large pediatric study, Kurth and colleagues noted significant increased asymmetry in the rostral anterior cingulate, thalamus, and nucleus accumbens in females, and significant increased asymmetry in the superior temporal gyrus and inferior parietal cortex in males with no relation to age [137]. Whole brain and regional gray matter volumes are associated with cognitive ability and function, which are closely related to behavior [134,135]. Moreover, lateralization of total and regional gray matter, while typical under normal developmental conditions in healthy individuals, was associated with predisposition to cortical diseases [136,138]. As such, sex differences in total and regional gray matter volumes, combined with asymmetry effects, may correlate to sex differences in behavior and cognition, especially in cases of dysfunction patterns related to tissue injury.
It is well established that TBI induces gray matter neurodegeneration that is not only confined to the acute window, but can persist for years after injury and alter functional outcomes [43,58,139,140]. TBI has been shown to cause premature atrophy trajectories associated with aging and predispose patients to disease states associated with neurodegeneration [139,140,141,142]. In adult cases of mild TBI, 80% of the gray matter structures were found to exhibit higher trajectories of decline compared to controls, resulting in neural atrophy patterns that do not reflect patients’ chronological age [142]. Belchev and colleagues observed that neurodegeneration following severe adult TBI does not attenuate over time, indicating persistently higher rates of premature atrophy [139]. However, even in children, the heightened rate of brain atrophy and corresponding increased brain age due to progressive neurodegeneration following TBI predisposes individuals to neurodegenerative diseases in addition to dysfunctions sustained from the original injury [140]. TBI sustained at an early age increases the risk factor of disease due to the introduction of premature neurodegeneration during periods of critical growth and neurodevelopment. Sex differences in total and regional gray matter volume may influence recovery trajectories as having more or less gray matter prior to injury could mediate threshold effects in volume loss—beyond which deficits in cognition, motor function, and behavioral processing may emerge. Additionally, sex-influenced asymmetries in gray matter will further exacerbate or protect impacted regions and influence long-term outcomes. Further investigation of the influence of sex on total and regional gray matter volumes and asymmetries using imaging and other non-invasive approaches are needed to elucidate key factors in recovery variability and in patterns of neurodegenerative disease incidence as it relates to TBI.
4. Sexual Dimorphisms in Functional Connectivity
Functional brain networks undergo profound changes during childhood and adolescence. Developmental changes in networks lead to increased network integration, efficiency, organization, and specialization [143]. These structural and functional network developmental changes are essential to normal maturation of function. Unfortunately, these networks possess limited compensatory potential, and damage often results in persistent neurological and functional deficits. Emerging evidence has found the default mode, dorsal attention, salience network, social brain, and sensorimotor networks are particularly implicated in pediatric TBI with notable differences between males and females (Figure 2) [144,145,146,147].
4.1. Resting-State Functional Connectivity
Resting-state functional connectivity occurs in the absence of an external stimulus and is reflected by resting state networks which are involved in cognitive, behavioral, and sensory functions to name a few [148]. Altered connectivity of resting-state networks has been noted in neurological and psychiatric conditions, and disruptions caused by TBI have been correlated with worse outcomes with sex emerging as a particular variable of interest [147,148].
Sex differences in functional connectivity have been demonstrated to emerge pre-puberty and persist well into adulthood [149]. For example, females were found to exhibit more within-module connectivity in a convergent pattern across network scales (i.e., connections to individual network nodes to entire networks), whereas males exhibited greater between-module connectivity (i.e., higher participation coefficient) [149]. The authors observed that, among individuals in the study, the extent to which their brain connectivity patterns aligned with those typically associated with males or females was significantly positively associated with sex-specific patterns of cognition; specifically, those exhibiting more male-typical connectivity patterns demonstrated superior performance on assessments traditionally favoring males [149]. For example, individuals with a higher level of between module connectivity (i.e., male-typical pattern) showed superior performance on spatial accuracy, language reasoning accuracy, and speed in a finger tapping test of motor function, thus reflective of a more male cognitive profile. However, individuals showing a higher level of within module connectivity (i.e., female-typical pattern) were correlated with better performance on emotion identification tests, a female dominated assessment [149]. This pattern of connectivity further suggests that female brain connectivity exhibits greater functional segregation considering accepted hierarchical organization of networks [150]. Sex specificity in network organization and patterns of cognition could help explain why some individuals are more resilient towards functional deficits. For example, better female performance and faster recovery on tests of facial emotion recognition following pediatric TBI could potentially be explained by this pattern of female-typical connectivity being more resistant to impairment [151].
Another important functional connectivity consideration is lateralization (Figure 2). The degree to which component structures of networks show hemispheric lateralization and activation specificity is dynamic, but specific to structures. Agcaoglu et al. reported both an age and sex interaction regarding lateralization of resting state networks in 6-10 year old children [152]. Notably, children exhibited both increases and decreases in lateralization of various networks at different stages of development, in contrast to adults, where laterality decreased with age [152]. Furthermore, males were found to have greater lateralization in activation of the left medial occipital gyrus, right middle frontal gyrus, right precentral gyrus, and right middle temporal gyrus components of the executive control network (i.e., cognitive control network) [152]. Females showed greater lateralization in the left superior parietal lobe of the sensorimotor network and the right superior frontal gyrus of the default mode network [152]. The greater degree of lateralization of the executive control network in the right middle frontal gyrus in male children may be related to male prevalence in development of secondary ADHD post-TBI. Higher cerebellar functional connectivity values in the right and left middle frontal gyrus were identified in children with ADHD [153]. Following damage due to TBI, male prevalence in comorbid ADHD development could arise in part due to compensation strategies increasing connectivity in the contralateral middle frontal gyrus. Alternatively, female right lateralization of the right superior frontal gyrus of the default mode network is believed to be an important underpinning for superior social cognition. The default mode network is predominantly bilateral and plays important roles in social cognition through episodic memory, self-referential processing, including that of social interactions and feelings. The right superior frontal gyrus is involved in a few different networks and thought to contribute to self-awareness. Greater lateralization in females could underlie superior self-awareness which could correlate with improved social cognition. Furthermore, Johnson et al. believes this characteristic could also help explain greater reporting of post-concussive emotional symptom load in female children [154]. Comparatively, a small study by Tuerk and colleagues noted increased resting state connectivity between the left superior frontal gyrus and the left fusiform gyrus seeds of the social brain network in adolescent TBI patients with social dysfunction, though they were not able to make significant brain-behavior associations [146]. Perhaps this node, being utilized in multiple complementary networks which are impacted by TBI, should be considered a key hub for mediation of social function following injury. Understanding innate lateralization effects may help researchers to find sex-specific patterns within the network organization complexities of normal and aberrant social processes.
TBI can disrupt the functional connectivity of the brain through direct impact, diffuse injury, and secondary injury which can damage functional nodes and connections. Resting-state connectivity disruption has been demonstrated to impact the social brain, default mode, sensorimotor, somatosensory, and executive control networks in addition to others, which contribute to cognitive, behavioral, and motor dysfunction outcomes in both children and adults [144,145,146,155,156]. The social brain network, composed of connectivity between the superior temporal sulcus, temporal pole, and orbitofrontal cortex, among other anatomical regions, undergoes change during developmental periods and is responsible for social cognition and behavior [146]. Alterations of this network following TBI have been linked to impairments in social functioning across age groups, but pediatric populations are likely to be predisposed to worse outcomes due to coinciding developmental disruption [146]. The default mode network plays a critical role in both acute and chronic outcomes following TBI. Better preservation of default mode network integrity has been correlated with recovery of consciousness in the acute stage of TBI, as well as improved behavioral and functional outcomes at chronic stages in both children and adults [145,157]. In pediatric TBI cases, one study found increased connectivity between the default mode network and right dorsal premotor cortex correlated with worse motor function [144]. The dorsal attention network, showed similarly increased connectivity with the sensorimotor cortex following TBI, which was correlated with better motor performance, indicating what the authors believed to be a compensatory mechanism to restore motor function following damage [144]. The motor network is implicated in TBI outcomes as a neural underpinning for disruption of response inhibition, which is commonly seen in children following TBI [158]. The sensorimotor network—as a whole—shows altered patterns of connectivity post-TBI, wherein patients with attention deficits had better performance in cases where there was stronger functional connectivity, possibly indicating another compensation mechanism in adults [156]. Taken together, these studies show that some compensatory changes in functional connectivity post-TBI can be maladaptive, while others restore function. One common strategy for compensatory function appears to be recruitment of networks which can aid in top-down control of the damaged function. It is important to note the sparse nature of literature that is specific to pediatric responses following TBI and further studies are needed to robustly account for influences due to age and the effects of longitudinal aberrant development of these networks.
Investigation into sexually influenced patterns of resting state functional connectivity disruption following TBI has been limited. However, one study found adult male TBI patients have greater connectivity in the motor, executive function, and cerebellum networks compared to female adults [159]. Alternatively, in a similar study done in a pediatric population, more alterations in resting-state functional connectivity patterns were found in female children compared to males, which persisted for at least a month [147]. Networks which displayed sex-specific alterations in connectivity included components of the salience and default mode networks [147]. It is important to note that both studies involved cohorts of mild TBI only, suggesting the observed sex differences in network connectivity pattern alteration following TBI is a sensitive phenomenon. Furthermore, it is hypothesized by both studies that challenges in delineating TBI symptom relationships with resting state fMRI disturbance may be better elucidated with the consideration of sex as a biological variable [147,159]. However, given the differences in age and consequent development between the two studies, despite similarities in objective and injury severity, the sex influence on the implicated networks is profoundly different. As such, the link to sex differences in symptom presentation may be more subtle than previously described.
4.2. Task-Based Functional Connectivity
Some networks activated as a result of external stimulation, referenced by task-based functional connectivity, have demonstrated sex effects in adults and, consequently, may also be impacted in children [160,161,162,163]. For example, there is support for the influence of sex on cerebrovascular reactivity that underlies fMRI signaling as males have been found to have higher reactivity values compared to females [164]. Other studies have found age and sex differences in task-based functional connectivity measures involving episodic and spatial memory tasks (Figure 2) [160,161]. The implication of sex differences in episodic memory tasks and memory-related disorders are suggested to be correlated with age-related declines and differences in functional connectivity patterns of episodic encoding and retrieval of spatial context processes [161]. Specifically, females showed greater activity in the lateral frontoparietal-related cognitive control and parahippocampal gyrus processes, which showed alterations in patterning as a function of advancing age. This may explain sex differences in memory decline [161]. Furthermore, age-related deficits in spatiotemporal context episodic memory are related to greater between-network integration in females and increased dorsal attention network-default mode network connectivity in males [160]. Sex differences in the task-based functional connectivity of the sensorimotor network have also been observed where females had lower volumes of brain activation and lower patterns of variability compared to males in motor tasks (Figure 2) [165]. It is important to note that these studies were conducted in adult populations and therefore do not account for the critical influence of age, as previously discussed. In addition to more research on sex differences in different task-based functional connectivity patterns throughout the brain, specific investigation is needed on when these sex differences in task-based connectivity emerge and how neurodevelopment changes network activation patterns in pediatric populations.
In pediatric TBI, disruption of task-based functional connectivity which results in impaired attention can further exacerbate functional deficits [162]. Pediatric patients who developed ADHD following TBI experienced worse behavioral and cognitive recovery at a ~7-year follow up [166]. Attention and executive function deficits were observed 7 years post-injury even in patients without diagnosed secondary ADHD [167]. Attention-related deficits have been significantly associated with abnormal topology of gyri in the parietal and temporal regions, to the extent a deep learning model was able to use alterations of these structures to identify pediatric TBI and predict future attention dysfunction [162,168]. Strazzer and colleagues provided evidence that when performing a sustained attention task following acquired brain injury, altered activation of structures within the attention network led to a decreased ability to recruit additional components of the attention network as task difficulty increased [169]. This altered activation is likely indicative of a default compensatory strategy of the attention network, in which increased load and strain, in addition to its own damage, may result in attentional disorders. Furthermore, increased activation in the frontotemporal regions—in addition to other components of working memory circuitry—have been associated with working memory impairments following pediatric TBI [162,163]. In the subacute period of TBI, Manelis and colleagues found pediatric patients were unable to activate the left inferior frontal gyrus during more difficult working memory tasks and recruit the left orbitofrontal cortex 6 months post-injury [170]. Westfall and his team assessed 1-year post-TBI patients and found increased activation of the frontotemporal regions and more recruitment in comparison to healthy controls during difficult working memory tasks [163]. Similarly, motor task impairments were correlated with alterations in parietal functional connectivity, while decreased activation in the motor cortex and anterior cingulate were associated with impaired inhibitory control processing [162,171]. Impairments of the primary motor cortex, and to a greater extent, the system level functional connectivity of the left parietal cortex, were correlated with worse regional efficiency and inattention in children, which further correlated with decreased performance in attention, working memory, and motor function tasks following TBI [162]. Further research on sexually influenced patterns of connectivity, specifically in the context of compensation and reorganization patterns following pediatric TBI, may help develop therapeutic targets for better functional recovery.
5. Considerations and Limitations
Assessment of sex influences on pediatric neurodevelopment is challenging due to the complex interplay between sex and other key biological variables, a high degree of variability in injury-specific parameters, and research study limitations (e.g., small sample sizes). Sex influences on TBI in both structural and functional vertices (Figure 1 and 2) have been frequently noted to have a dynamic relationship with age and neurodevelopment, in addition to the unique influences of distinct genetic backgrounds and environmental cues leading to variable responses. Furthermore, when evaluating human databases there is a high degree of variability in injury mechanisms which further influences patterns of damage. For example, combined rotational and focal injuries and differing degrees of cortical and sub-cortical tissue damage that are not commonly observed in controlled animal models. Studies with larger sample sizes can help to identify common elements across these variables, leading to more robust conclusions. However, larger studies with more patients have been limited. Most studies assessed in this review looked at smaller populations and were unable to complete longitudinal evaluations. This limited capacity to assess neuropathology from acute to chronic stages restricts our ability to understand the effect of sex on recovery. Additionally, the rapid growth and development of children means that 6–10-year-old children are not directly comparable to 0-4- or 14–17-year-old children, despite many studies analyzing all combinations of ages without defined comparable periods. As such, studies could be missing age-specific biomarkers of pathology which could help explain differing trajectories of recovery between younger and older children.
Additionally, most of the research evaluated in this review focused on MRI-based analytical approaches, which can be quite heterogenous due to differences in MRI scanners, coils, collection protocols, post-processing pipelines, atlases, and other key analytics. Finally, a considerable limitation in assessing functional connectivity included discrepancies in structures involved within individual networks. For example, the superior frontal gyrus is considered part of the default mode network in some studies, but not all. This makes comparison of networks across studies more difficult as the activation of a given structure can give rise to very different results or interpretations. Evaluation of sex differences in the context of pediatric TBI should therefore be characterized in the context of relationships with other critical variables (e.g., developmental stage) and utilized analytical techniques.
6. Conclusions
Pediatric TBI patients have been previously considered by the field to have greater likelihood of recovery due to neuroplasticity. However, disruption of the dynamic, tightly controlled, ongoing development of the pediatric brain can instead result in worse outcomes across all injury severities. Patient-specific factors should be considered with the same degree of interest as injury-specific ones, but knowledge gaps in TBI pathophysiology continue to prevent development of treatments. Sex as a biological variable in pediatric TBI pathology is a critically understudied component of structural and functional neural connections of recovery. Sex influences in gray matter structural volumes and developmental trajectories may affect resiliency to threshold volume loss deficits and can induce premature atrophy in the developing brain. Gray matter compartmental volume and density and white matter microstructural integrity highlight innate sex effects which translate to greater internalizing behavioral deficits in female children, compared to greater externalizing behavioral deficits in male children. Sex influences in functional connectivity may further affect compensatory strategies following damage and underlie cognitive, motor, and behavioral sequelae. Sex effects also show a strong interplay with age, thus illustrating nuanced differences in recovery throughout childhood and adolescence. Further elucidation of how sex influences neurodevelopment and functional recovery following pediatric TBI is an important step in addressing knowledge gaps to provide more informed care for this vulnerable patient population.
Transparency: Rigor, and Reproducibility Statement
The analysis of this review article was pre-specified by the lead and corresponding author prior to the literature search. The resources and software used in this review are clearly identified and openly available. This article will be published under a Creative Commons Open Access license, and upon publication will be freely available.
Author Contributions
All listed authors made a substantial contribution to the concept and interpretation of the literature; drafted the review article; revised it critically for important intellectual content; and approved the version to be published.
Funding
This work was supported by the National Institutes of Health grant 1R21NS131526-01. The authors would also like to thank the Franklin Foundation Fellowship program and the ARCS Foundation for their financial contributions.
Acknowledgments
The authors would like to thank Sydney Sneed for her valuable editorial critiques of this review. Figures created with BioRender https://BioRender.com/hlv5kpn license.
Conflicts of Interest
We have no conflicts of interest, financial or otherwise, to declare. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| TBI | Traumatic brain injury |
| ED | Emergency department |
| MRI | Magnetic resonance imaging |
| fMRI | Functional magnetic resonance imaging |
| ADHD | Attention deficit/hyperactivity disorder |
| DTI | Diffusion tensor imaging |
| FA | Fractional anisotropy |
References
- Chen, C.; Peng, J.; Sribnick, E.A.; Zhu, M.; Xiang, H. Trend of Age-Adjusted Rates of Pediatric Traumatic Brain Injury in U.S. Emergency Departments from 2006 to 2013. Int. J. Environ. Res. Public Health 2018, 15, 1171. [Google Scholar] [CrossRef]
- Taylor, C.A.; Bell, J.M.; Breiding, M.J.; Xu, L. Traumatic Brain Injury–Related Emergency Department Visits, Hospitalizations, and Deaths—United States, 2007 and 2013. MMWR Surveill. Summ. 2017, 66, 1–16. [Google Scholar] [CrossRef]
- Ignacio, D.A.; Babikian, T.; Dennis, E.L.; Bickart, K.C.; Choe, M.; Snyder, A.R.; Brown, A.; Giza, C.C.; Asarnow, R.F. The neurocognitive correlates of DTI indicators of white matter disorganization in pediatric moderate-to-severe traumatic brain injury. Front. Hum. Neurosci. 2024, 18, 1470710. [Google Scholar] [CrossRef]
- Levin, H.S.; Temkin, N.R.; Barber, J.; Nelson, L.D.; Robertson, C.; Brennan, J.; Stein, M.B.; Yue, J.K.; Giacino, J.T.; McCrea, M.A.; et al. Association of Sex and Age With Mild Traumatic Brain Injury–Related Symptoms: A TRACK-TBI Study. JAMA Netw. Open 2021, 4, e213046–e213046. [Google Scholar] [CrossRef] [PubMed]
- Free, K.E.; Greene, A.M.; Bondi, C.O.; Lajud, N.; de la Tremblaye, P.B.; Kline, A.E. Comparable impediment of cognitive function in female and male rats subsequent to daily administration of haloperidol after traumatic brain injury. Exp. Neurol. 2017, 296, 62–68. [Google Scholar] [CrossRef]
- Kowalski, R.G.; Hammond, F.M.; Weintraub, A.H.; Nakase-Richardson, R.; Zafonte, R.D.; Whyte, J.; Giacino, J.T. Recovery of Consciousness and Functional Outcome in Moderate and Severe Traumatic Brain Injury. JAMA Neurol. 2021, 78, 548–557. [Google Scholar] [CrossRef]
- Berry, C.; Ley, E.J.; Tillou, A.; Cryer, G.; Margulies, D.R.; Salim, A. The Effect of Gender on Patients With Moderate to Severe Head Injuries. J. Trauma: Inj. Infect. Crit. Care 2009, 67, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Fraser, E.E.; Downing, M.G.; Biernacki, K.; McKenzie, D.P.; Ponsford, J.L. Cognitive Reserve and Age Predict Cognitive Recovery after Mild to Severe Traumatic Brain Injury. J. Neurotrauma 2019, 36, 2753–2761. [Google Scholar] [CrossRef]
- Keenan, H.T.; Clark, A.E.; Holubkov, R.; Cox, C.S.; Ewing-Cobbs, L. Psychosocial and Executive Function Recovery Trajectories One Year after Pediatric Traumatic Brain Injury: The Influence of Age and Injury Severity. J. Neurotrauma 2018, 35, 286–296. [Google Scholar] [CrossRef]
- McCarthy, M.M.; Arnold, A.P.; Ball, G.F.; Blaustein, J.D.; De Vries, G.J. Sex Differences in the Brain: The Not So Inconvenient Truth. J. Neurosci. 2012, 32, 2241–2247. [Google Scholar] [CrossRef]
- Greenberger, P.E.; Knab, S. News from the Society for Women's Health Research: Subgroup Analysis in Clinical Trials: Detecting Sex Differences. J. Women's Heal. Gender-Based Med. 2002, 11, 7–9. [Google Scholar] [CrossRef]
- McCarthy, M.M.; Pickett, L.A.; VanRyzin, J.W.; Kight, K.E. Surprising origins of sex differences in the brain. Horm. Behav. 2015, 76, 3–10. [Google Scholar] [CrossRef]
- Hirschberg, R.; Weiss, D.; Zafonte, R. Traumatic Brain Injury and Gender: What is Known and What is Not. Futur. Neurol. 2008, 3, 483–489. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Q.; Luo, J.; Hu, M.; Zuo, C. Effects of Age and Sex on Subcortical Volumes. Front. Aging Neurosci. 2019, 11, 259. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liang, M.; Qin, W.; Wan, B.; Yu, C.; Ming, D. Gender Differences Are Encoded Differently in the Structure and Function of the Human Brain Revealed by Multimodal MRI. Front. Hum. Neurosci. 2020, 14, 244. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; van Tol, M.-J.; Li, M.; Miao, W.; Jiao, Y.; Heinze, H.-J.; Bogerts, B.; He, H.; Walter, M. Regional specificity of sex effects on subcortical volumes across the lifespan in healthy aging. Hum. Brain Mapp. 2014, 35, 238–247. [Google Scholar] [CrossRef]
- Backhausen, L.L.; Fröhner, J.H.; Lemaître, H.; Artiges, E.; Martinot, M.P.; Herting, M.M.; Sticca, F.; Banaschewski, T.; Barker, G.J.; Bokde, A.L.W.; et al. Adolescent to young adult longitudinal development of subcortical volumes in two European sites with four waves. Hum. Brain Mapp. 2024, 45, e26574. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Kobayashi, S.; Yamaguchi, S.; Iijima, K.-I.; Okada, K.; Yamashita, K. Gender Effects on Age-Related Changes in Brain Structure. AJNR Am J Neuroradiol 2000, 21, 112–118. [Google Scholar]
- Bartnik-Olson, B.L.; Holshouser, B.; Ghosh, N.; Oyoyo, U.E.; Nichols, J.G.; Pivonka-Jones, J.; Tong, K.; Ashwal, S. Evolving White Matter Injury following Pediatric Traumatic Brain Injury. J. Neurotrauma 2021, 38, 111–121. [Google Scholar] [CrossRef]
- Dennis, E.L.; Faskowitz, J.; Rashid, F.; Babikian, T.; Mink, R.; Babbitt, C.; Johnson, J.; Giza, C.C.; Jahanshad, N.; Thompson, P.M.; et al. Diverging volumetric trajectories following pediatric traumatic brain injury. NeuroImage: Clin. 2017, 15, 125–135. [Google Scholar] [CrossRef]
- A Ashman, T.; A Spielman, L.; Hibbard, M.R.; Silver, J.M.; Chandna, T.; A Gordon, W. Psychiatric challenges in the first 6 years after traumatic brain injury: cross-sequential analyses of axis I disorders11No commercial party having a direct financial interest in the results of the research supporting this article has or will confer a benefit upon the authors(s) or upon any organization with which the author(s) is/are associated. Arch. Phys. Med. Rehabilitation 2004, 85, 36–42. [Google Scholar] [CrossRef]
- Neumann, D.; Zupan, B. Sex Differences in Emotional Insight After Traumatic Brain Injury. Arch. Phys. Med. Rehabilitation 2020, 101, 1922–1928. [Google Scholar] [CrossRef]
- Ilie, G.; E Mann, R.; Boak, A.; Adlaf, E.M.; Hamilton, H.; Asbridge, M.; Rehm, J.; Cusimano, M.D. Cross-sectional examination of the association of co-occurring alcohol misuse and traumatic brain injury on mental health and conduct problems in adolescents in Ontario, Canada. BMJ Open 2016, 6, e011824. [Google Scholar] [CrossRef]
- Kennedy, E.; Heron, J.; Munafò, M. Substance use, criminal behaviour and psychiatric symptoms following childhood traumatic brain injury: findings from the ALSPAC cohort. Eur. Child Adolesc. Psychiatry 2017, 26, 1197–1206. [Google Scholar] [CrossRef]
- Cummings, J.A.; Jagannathan, L.; Jackson, L.R.; Becker, J.B. Sex differences in the effects of estradiol in the nucleus accumbens and striatum on the response to cocaine: Neurochemistry and behavior. Drug Alcohol Depend. 2014, 135, 22–28. [Google Scholar] [CrossRef]
- Zhang, D.; Yang, S.; Yang, C.; Jin, G.; Zhen, X. Estrogen regulates responses of dopamine neurons in the ventral tegmental area to cocaine. Psychopharmacology 2008, 199, 625–635. [Google Scholar] [CrossRef]
- Becker, J.B.; Chartoff, E. Sex differences in neural mechanisms mediating reward and addiction. Neuropsychopharmacology 2019, 44, 166–183. [Google Scholar] [CrossRef] [PubMed]
- Viviani, R.; Dommes, L.; Bosch, J.; Steffens, M.; Paul, A.; Schneider, K.L.; Stingl, J.C.; Beschoner, P. Signals of anticipation of reward and of mean reward rates in the human brain. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Joue, G.; Chakroun, K.; Bayer, J.; Gläscher, J.; Zhang, L.; Fuss, J.; Hennies, N.; Sommer, T. Sex Differences and Exogenous Estrogen Influence Learning and Brain Responses to Prediction Errors. Cereb. Cortex 2021, 32, 2022–2036. [Google Scholar] [CrossRef]
- Delfel, E.L.; Aguinaldo, L.; Correa, K.; Courtney, K.E.; Max, J.E.; Tapert, S.F.; Jacobus, J. Traumatic brain injury, working memory-related neural processing, and alcohol experimentation behaviors in youth from the ABCD cohort. Dev. Cogn. Neurosci. 2024, 66, 101344. [Google Scholar] [CrossRef] [PubMed]
- Scott, C.; McKinlay, A.; McLellan, T.; Britt, E.; Grace, R.; MacFarlane, M. A comparison of adult outcomes for males compared to females following pediatric traumatic brain injury. Neuropsychology 2015, 29, 501–508. [Google Scholar] [CrossRef]
- Dong, X.; Zhang, R.; Zhornitsky, S.; Le, T.M.; Wang, W.; Li, C.-S.R.; Zhang, S. Depression Mediates the Relationship between Childhood Trauma and Internet Addiction in Female but Not Male Chinese Adolescents and Young Adults. J. Clin. Med. 2021, 10, 5015. [Google Scholar] [CrossRef]
- Walsh, S.P.; White, K.M.; Cox, S.; Young, R.M. Keeping in constant touch: The predictors of young Australians’ mobile phone involvement. Comput. Hum. Behav. 2011, 27, 333–342. [Google Scholar] [CrossRef]
- Hong, F.-Y.; Chiu, S.-I.; Huang, D.-H. A model of the relationship between psychological characteristics, mobile phone addiction and use of mobile phones by Taiwanese university female students. Comput. Hum. Behav. 2012, 28, 2152–2159. [Google Scholar] [CrossRef]
- Østby, Y.; Tamnes, C.K.; Fjell, A.M.; Westlye, L.T.; Due-Tønnessen, P.; Walhovd, K.B. Heterogeneity in Subcortical Brain Development: A Structural Magnetic Resonance Imaging Study of Brain Maturation from 8 to 30 Years. J. Neurosci. 2009, 29, 11772–11782. [Google Scholar] [CrossRef] [PubMed]
- Sowell, E.R.; et al. Development of cortical and subcortical brain structures in childhood and adolescence: a structural MRI study. Dev Med Child Neurol 2002, 44, 4–16. [Google Scholar] [CrossRef]
- Koolschijn, P.C.M.; Crone, E.A. Sex differences and structural brain maturation from childhood to early adulthood. Dev. Cogn. Neurosci. 2013, 5, 106–118. [Google Scholar] [CrossRef]
- Rijpkema, M.; Everaerd, D.; van der Pol, C.; Franke, B.; Tendolkar, I.; Fernández, G. Normal sexual dimorphism in the human basal ganglia. Hum. Brain Mapp. 2012, 33, 1246–1252. [Google Scholar] [CrossRef] [PubMed]
- Abedelahi, A.; Hasanzadeh, H.; Hadizadeh, H.; Joghataie, M.T. Morphometric and volumetric study of caudate and putamen nuclei in normal individuals by MRI: Effect of normal aging, gender and hemispheric differences. Pol. J. Radiol. 2013, 78, 7–14. [Google Scholar] [CrossRef] [PubMed]
- McDonald, W.M.; Husain, M.; Doraiswamy, P.M.; Figiel, G.; Boyko, O.; Krishnan, K.R.R. A magnetic resonance image study of age-related changes in human putamen nuclei. NeuroReport 1991, 2, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Bourke, N.J.; Demarchi, C.; De Simoni, S.; Samra, R.; Patel, M.C.; Kuczynski, A.; Mok, Q.; Wimalasundera, N.; Vargha-Khadem, F.; Sharp, D.J. Brain volume abnormalities and clinical outcomes following paediatric traumatic brain injury. Brain 2022, 145, 2920–2934. [Google Scholar] [CrossRef] [PubMed]
- Molteni, E.; Pagani, E.; Strazzer, S.; Arrigoni, F.; Beretta, E.; Boffa, G.; Galbiati, S.; Filippi, M.; Rocca, M.A. Fronto-temporal vulnerability to disconnection in paediatric moderate and severe traumatic brain injury. Eur. J. Neurol. 2019, 26, 1183–1190. [Google Scholar] [CrossRef]
- Gooijers, J.; Chalavi, S.; Beeckmans, K.; Michiels, K.; Lafosse, C.; Sunaert, S.; Swinnen, S.P. Subcortical Volume Loss in the Thalamus, Putamen, and Pallidum, Induced by Traumatic Brain Injury, Is Associated With Motor Performance Deficits. Neurorehabilit. Neural Repair 2016, 30, 603–614. [Google Scholar] [CrossRef] [PubMed]
- E Alexander, G.; DeLong, M.R.; Strick, P.L. Parallel Organization of Functionally Segregated Circuits Linking Basal Ganglia and Cortex. Annu. Rev. Neurosci. 1986, 9, 357–381. [Google Scholar] [CrossRef]
- Garcia, D.; Hungerford, G.M.; Bagner, D.M. Topical Review: Negative Behavioral and Cognitive Outcomes Following Traumatic Brain Injury in Early Childhood. J. Pediatr. Psychol. 2015, 40, 391–397. [Google Scholar] [CrossRef]
- Luo, X.; Mao, Q.; Shi, J.; Wang, X.; Li, C.-S.R. Putamen gray matter volumes in neuropsychiatric and neurodegenerative disorders. . 2019, 3. [Google Scholar]
- Harris, G.J.; Pearlson, G.D.; Peyser, C.E.; Aylward, E.H.; Roberts, J.; Barta, P.E.; Chase, G.A.; Folstein, S.E. Putamen volume reduction on magnetic resonance imaging exceeds caudate changes in mild Huntington's disease. Ann. Neurol. 1992, 31, 69–75. [Google Scholar] [CrossRef]
- Cousins, D.A.; et al. Atrophy of the putamen in dementia with Lewy bodies but not Alzheimer's disease: an MRI study. Neurology 2003, 61, 1191–1195. [Google Scholar] [CrossRef]
- de Jong, L.W.; van der Hiele, K.; Veer, I.M.; Houwing, J.J.; Westendorp, R.G.J.; Bollen, E.L.E.M.; de Bruin, P.W.; Middelkoop, H.A.M.; van Buchem, M.A.; van der Grond, J. Strongly reduced volumes of putamen and thalamus in Alzheimer's disease: an MRI study. Brain 2008, 131 Pt 12 Pt 12, 3277–3285. [Google Scholar] [CrossRef] [PubMed]
- Krämer, J.; Meuth, S.G.; Tenberge, J.-G.; Schiffler, P.; Wiendl, H.; Deppe, M. Early and Degressive Putamen Atrophy in Multiple Sclerosis. Int. J. Mol. Sci. 2015, 16, 23195–23209. [Google Scholar] [CrossRef]
- Wellington, T.M.; et al. Magnetic resonance imaging volumetric analysis of the putamen in children with ADHD: combined type versus control. J Atten Disord 2006, 10, 171–80. [Google Scholar] [CrossRef]
- Husain, M.M.; et al. A magnetic resonance imaging study of putamen nuclei in major depression. Psychiatry Res 1991, 40, 95–9. [Google Scholar]
- Leunissen, I.; Coxon, J.P.; Caeyenberghs, K.; Michiels, K.; Sunaert, S.; Swinnen, S.P. Subcortical volume analysis in traumatic brain injury: The importance of the fronto-striato-thalamic circuit in task switching. Cortex 2014, 51, 67–81. [Google Scholar] [CrossRef]
- Simeone, P.; Auzias, G.; Lefevre, J.; Takerkart, S.; Coulon, O.; Lesimple, B.; Torkomian, G.; Battisti, V.; Jacquens, A.; Couret, D.; et al. Long-term follow-up of neurodegenerative phenomenon in severe traumatic brain injury using MRI. Ann. Phys. Rehabilitation Med. 2022, 65, 101599. [Google Scholar] [CrossRef]
- Davis, C.K.; Bathula, S.; Jeong, S.; Arruri, V.; Choi, J.; Subramanian, S.; Ostrom, C.M.; Vemuganti, R. An antioxidant and anti-ER stress combination therapy elevates phosphorylation of α-Syn at serine 129 and alleviates post-TBI PD-like pathology in a sex-specific manner in mice. Exp. Neurol. 2024, 377, 114795. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, P.O.; Roussakis, A.-A.; De Simoni, S.; Bourke, N.; Fleminger, J.; Cole, J.; Piccini, P.; Sharp, D. Distinct dopaminergic abnormalities in traumatic brain injury and Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 2020, 91, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Pettemeridou, E.; Constantinidou, F. The cortical and subcortical substrates of quality of life through substrates of self-awareness and executive functions, in chronic moderate-to-severe TBI. Brain Inj. 2022, 36, 110–120. [Google Scholar] [CrossRef]
- Drijkoningen, D.; Chalavi, S.; Sunaert, S.; Duysens, J.; Swinnen, S.P.; Caeyenberghs, K. Regional Gray Matter Volume Loss Is Associated with Gait Impairments in Young Brain-Injured Individuals. J. Neurotrauma 2017, 34, 1022–1034. [Google Scholar] [CrossRef]
- Dennis, E.L. , et al. Differences in Brain Volume in Military Service Members and Veterans After Blast-Related Mild TBI: A LIMBIC-CENC Study. JAMA Netw Open 2024, 7, e2443416. [Google Scholar] [CrossRef] [PubMed]
- Niemann, C.; Godde, B.; Staudinger, U.; Voelcker-Rehage, C. Exercise-induced changes in basal ganglia volume and cognition in older adults. Neuroscience 2014, 281, 147–163. [Google Scholar] [CrossRef]
- Takahashi, C.; Hinson, H.E.; Baguley, I.J. Autonomic dysfunction syndromes after acute brain injury. Clin Neurol 2015, 128, 539–551. [Google Scholar] [CrossRef]
- Giedd, J.N.; Rapoport, J.L. Structural MRI of Pediatric Brain Development: What Have We Learned and Where Are We Going? Neuron 2010, 67, 728–734. [Google Scholar] [CrossRef]
- Pfefferbaum, A.; Rohlfing, T.; Rosenbloom, M.J.; Chu, W.; Colrain, I.M.; Sullivan, E.V. Variation in longitudinal trajectories of regional brain volumes of healthy men and women (ages 10 to 85years) measured with atlas-based parcellation of MRI. NeuroImage 2013, 65, 176–193. [Google Scholar] [CrossRef]
- MacMaster, F.P.; Keshavan, M.; Mirza, Y.; Carrey, N.; Upadhyaya, A.R.; El-Sheikh, R.; Buhagiar, C.J.; Taormina, S.P.; Boyd, C.; Lynch, M.; et al. Development and sexual dimorphism of the pituitary gland. Life Sci. 2007, 80, 940–944. [Google Scholar] [CrossRef]
- Wagner, M.W.; Rafful, P.P.; Vidarsson, L.; Ertl-Wagner, B.B. MRI Volumetric Analysis of the Hypothalamus and Limbic System across the Pediatric Age Span. Children 2023, 10, 477. [Google Scholar] [CrossRef] [PubMed]
- Vlad, R.M.; Albu, A.I.; Nicolaescu, I.D.; Dobritoiu, R.; Carsote, M.; Sandru, F.; Albu, D.; Păcurar, D. An Approach to Traumatic Brain Injury-Related Hypopituitarism: Overcoming the Pediatric Challenges. Diagnostics 2023, 13, 212. [Google Scholar] [CrossRef]
- Silva, P.P.; Bhatnagar, S.; Herman, S.D.; Zafonte, R.; Klibanski, A.; Miller, K.K.; Tritos, N.A. Predictors of Hypopituitarism in Patients with Traumatic Brain Injury. J. Neurotrauma 2015, 32, 1789–1795. [Google Scholar] [CrossRef] [PubMed]
- Personnier, C.; Crosnier, H.; Meyer, P.; Chevignard, M.; Flechtner, I.; Boddaert, N.; Breton, S.; Mignot, C.; Dassa, Y.; Souberbielle, J.-C.; et al. Prevalence of Pituitary Dysfunction After Severe Traumatic Brain Injury in Children and Adolescents: A Large Prospective Study. J. Clin. Endocrinol. Metab. 2014, 99, 2052–2060. [Google Scholar] [CrossRef]
- Claessen, L.Ó.E.; Kristjánsdóttir, H.; Jónsdóttir, M.K.; Lund, S.H.; Kristensen, I.U.; Sigurjónsdóttir, H.Á. Pituitary dysfunction following mild traumatic brain injury in female athletes. Endocr. Connect. 2024, 13. [Google Scholar] [CrossRef]
- Dassa, Y. , et al. Pituitary deficiency and precocious puberty after childhood severe traumatic brain injury: a long-term follow-up prospective study. Eur J Endocrinol 2019, 180, 281–290. [Google Scholar] [CrossRef]
- Gray, S. , et al. Hypopituitarism After Traumatic Brain Injury. Cureus 2019, 11, e4163. [Google Scholar]
- Prodam, F.; Gasco, V.; Caputo, M.; Zavattaro, M.; Pagano, L.; Marzullo, P.; Belcastro, S.; Busti, A.; Perino, C.; Grottoli, S.; et al. Metabolic alterations in patients who develop traumatic brain injury (TBI)-induced hypopituitarism. Growth Horm. IGF Res. 2013, 23, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Nemes, O.; Kovacs, N.; Czeiter, E.; Kenyeres, P.; Tarjanyi, Z.; Bajnok, L.; Buki, A.; Doczi, T.; Mezosi, E. Predictors of post-traumatic pituitary failure during long-term follow-up. Hormones 2015, 14, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Acerini, C.L.; Tasker, R.C. Traumatic brain injury induced hypothalamic-pituitary dysfunction: a paediatric perspective. Pituitary 2007, 10, 373–380. [Google Scholar] [CrossRef]
- Su-Ching, L.; Zasler, N.D.; Kreutzer, J.S. Male pituitary-gonadal dysfunction following severe traumatic brain injury. Brain Inj. 1994, 8, 571–577. [Google Scholar] [CrossRef]
- Niederland, T.; Makovi, H.; Gál, V.; Andréka, B.; Ábrahám, C.S.; Kovács, J. Abnormalities of Pituitary Function after Traumatic Brain Injury in Children. J. Neurotrauma 2007, 24, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Agha, A.; Rogers, B.; Sherlock, M.; O’kelly, P.; Tormey, W.; Phillips, J.; Thompson, C.J. Anterior Pituitary Dysfunction in Survivors of Traumatic Brain Injury. J. Clin. Endocrinol. Metab. 2004, 89, 4929–4936. [Google Scholar] [CrossRef]
- Xie, Y.; Chen, Y.A.; De Bellis, M.D. The Relationship of Age, Gender, and IQ With the Brainstem and Thalamus in Healthy Children and Adolescents: A Magnetic Resonance Imaging Volumetric Study. J. Child Neurol. 2012, 27, 325–331. [Google Scholar] [CrossRef]
- Greer, K.M.; Snyder, A.; Junge, C.; Reading, M.; Jarvis, S.; Squires, C.; Bigler, E.D.; Popuri, K.; Beg, M.F.; Taylor, H.G.; et al. Surface-based abnormalities of the executive frontostriatial circuit in pediatric TBI. NeuroImage: Clin. 2022, 35, 103136. [Google Scholar] [CrossRef]
- Keenan, H.T.; Clark, A.E.; Holubkov, R.; Cox, C.S.; Ewing-Cobbs, L. Trajectories of Children's Executive Function After Traumatic Brain Injury. JAMA Netw. Open 2021, 4, e212624–e212624. [Google Scholar] [CrossRef]
- Grados, M.A.; Vasa, R.A.; Riddle, M.A.; Slomine, B.S.; Salorio, C.; Christensen, J.; Gerring, J. New onset obsessive-compulsive symptoms in children and adolescents with severe traumatic brain injury. Depression Anxiety 2008, 25, 398–407. [Google Scholar] [CrossRef]
- Kim, T.-H.; Choi, E.; Kim, H.; Kim, S.-Y.; Kim, Y.; Kim, B.-N.; Park, S.; Jung, K.-I.; Park, B.; Park, M.-H. The Association Between Hippocampal Volume and Level of Attention in Children and Adolescents. Front. Syst. Neurosci. 2021, 15. [Google Scholar] [CrossRef] [PubMed]
- Wierenga, L.; Langen, M.; Ambrosino, S.; van Dijk, S.; Oranje, B.; Durston, S. Typical development of basal ganglia, hippocampus, amygdala and cerebellum from age 7 to 24. NeuroImage 2014, 96, 67–72. [Google Scholar] [CrossRef]
- Raz, N.; Gunning-Dixon, F.; Head, D.; Rodrigue, K.M.; Williamson, A.; Acker, J.D. Aging, sexual dimorphism, and hemispheric asymmetry of the cerebral cortex: replicability of regional differences in volume. Neurobiol. Aging 2004, 25, 377–396. [Google Scholar] [CrossRef]
- Pruessner, J.C.; Collins, D.L.; Pruessner, M.; Evans, A.C. Age and Gender Predict Volume Decline in the Anterior and Posterior Hippocampus in Early Adulthood. J. Neurosci. 2001, 21, 194–200. [Google Scholar] [CrossRef]
- Neufang, S.; Specht, K.; Hausmann, M.; Güntürkün, O.; Herpertz-Dahlmann, B.; Fink, G.R.; Konrad, K. Sex Differences and the Impact of Steroid Hormones on the Developing Human Brain. Cereb. Cortex 2009, 19, 464–473. [Google Scholar] [CrossRef]
- Giedd, J.N.; Vaituzis, A.C.; Hamburger, S.D.; Lange, N.; Rajapakse, J.C.; Kaysen, D.; Vauss, Y.C.; Rapoport, J.L. Quantitative MRI of the temporal lobe, amygdala, and hippocampus in normal human development: Ages 4-18 years. J. Comp. Neurol. 1996, 366, 223–230. [Google Scholar] [CrossRef]
- Dennison, M.; Whittle, S.; Yücel, M.; Vijayakumar, N.; Kline, A.; Simmons, J.; Allen, N.B. Mapping subcortical brain maturation during adolescence: evidence of hemisphere- and sex-specific longitudinal changes. Dev. Sci. 2013, 16, 772–791. [Google Scholar] [CrossRef] [PubMed]
- Tamnes, C.K.; Bos, M.G.; van de Kamp, F.C.; Peters, S.; Crone, E.A. Longitudinal development of hippocampal subregions from childhood to adulthood. Dev. Cogn. Neurosci. 2018, 30, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Peper, J.S.; Brouwer, R.M.; Schnack, H.G.; van Baal, G.C.; van Leeuwen, M.; Berg, S.M.v.D.; de Waal, H.A.D.-V.; Boomsma, D.I.; Kahn, R.S.; Pol, H.E.H. Sex steroids and brain structure in pubertal boys and girls. Psychoneuroendocrinology 2009, 34, 332–342. [Google Scholar] [CrossRef]
- Peper, J.; Pol, H.H.; Crone, E.; van Honk, J. Sex steroids and brain structure in pubertal boys and girls: a mini-review of neuroimaging studies. Neuroscience 2011, 191, 28–37. [Google Scholar] [CrossRef]
- Peper, J.S.; Schnack, H.G.; Brouwer, R.M.; Van Baal, G.C.M.V.; Pjetri, E.; Székely, E.; van Leeuwen, M.; Van Den Berg, S.M.; Collins, D.L.; Evans, A.C.; et al. Heritability of regional and global brain structure at the onset of puberty: A magnetic resonance imaging study in 9-year-old twin pairs. Hum. Brain Mapp. 2009, 30, 2184–2196. [Google Scholar] [CrossRef]
- Mu, S.H.; Yuan, B.K.; Tan, L.H. Effect of Gender on Development of Hippocampal Subregions From Childhood to Adulthood. Front. Hum. Neurosci. 2020, 14. [Google Scholar] [CrossRef]
- Fish, A.M.; Nadig, A.; Seidlitz, J.; Reardon, P.K.; Mankiw, C.; McDermott, C.L.; Blumenthal, J.D.; Clasen, L.S.; Lalonde, F.; Lerch, J.P.; et al. Sex-biased trajectories of amygdalo-hippocampal morphology change over human development. NeuroImage 2020, 204, 116122–116122. [Google Scholar] [CrossRef] [PubMed]
- Moser, E.; Moser, M.; Andersen, P. Spatial learning impairment parallels the magnitude of dorsal hippocampal lesions, but is hardly present following ventral lesions. J. Neurosci. 1993, 13, 3916–3925. [Google Scholar] [CrossRef] [PubMed]
- Bear, M.F., B. W. Connors, and M.A. Paradiso, Neuroscience : exploring the brain. 3rd ed. 2007, Philadelphia, PA: Lippincott Williams & Wilkins. xxxviii, 857 p.
- Lundy-Ekman, L. , Neuroscience : fundamentals for rehabilitation. 4th ed. 2013, St. Louis, Mo.: Elsevier. xi, 539 p.
- Bazarian, J.J.; Blyth, B.; Mookerjee, S.; He, H.; McDermott, M.P. Sex Differences in Outcome after Mild Traumatic Brain Injury. J. Neurotrauma 2010, 27, 527–539. [Google Scholar] [CrossRef]
- Bird, C.M.; Burgess, N. The hippocampus and memory: insights from spatial processing. Nat. Rev. Neurosci. 2008, 9, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Farace, E.; Alves, W.M. Do women fare worse: a metaanalysis of gender differences in traumatic brain injury outcome. J. Neurosurg. 2000, 93, 539–545. [Google Scholar] [CrossRef]
- Semple, B.D.; Dixit, S.; Shultz, S.R.; Boon, W.C.; O’bRien, T.J. Sex-dependent changes in neuronal morphology and psychosocial behaviors after pediatric brain injury. Behav. Brain Res. 2017, 319, 48–62. [Google Scholar] [CrossRef]
- Davila-Valencia, I.; Saad, M.; Olthoff, G.; Faulkner, M.; Charara, M.; Farnum, A.; Dysko, R.C.; Zhang, Z. Sex specific effects of buprenorphine on adult hippocampal neurogenesis and behavioral outcomes during the acute phase after pediatric traumatic brain injury in mice. Neuropharmacology 2024, 245, 109829. [Google Scholar] [CrossRef]
- Rice, L.C.; Rochowiak, R.N.; Plotkin, M.R.; Rosch, K.S.; Mostofsky, S.H.; Crocetti, D. Sex Differences and Behavioral Associations with Typically Developing Pediatric Regional Cerebellar Gray Matter Volume. Cerebellum 2024, 23, 589–600. [Google Scholar] [CrossRef]
- Moore, D.M.; D’mEllo, A.M.; McGrath, L.M.; Stoodley, C.J. The developmental relationship between specific cognitive domains and grey matter in the cerebellum. Dev. Cogn. Neurosci. 2017, 24, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kipping, J.A.; Xie, Y.; Qiu, A. Cerebellar development and its mediation role in cognitive planning in childhood. Hum. Brain Mapp. 2018, 39, 5074–5084. [Google Scholar] [CrossRef] [PubMed]
- Steele, C.J.; Chakravarty, M.M. Gray-matter structural variability in the human cerebellum: Lobule-specific differences across sex and hemisphere. NeuroImage 2018, 170, 164–173. [Google Scholar] [CrossRef]
- Sgro, M.; Ellens, S.; Kodila, Z.N.; Christensen, J.; Li, C.; Mychasiuk, R.; Yamakawa, G.R. Repetitive mild traumatic brain injury alters central and peripheral clock gene expression in the adolescent rat. Neurobiol. Sleep Circadian Rhythm. 2023, 14, 100090. [Google Scholar] [CrossRef]
- Snyder, K.; Dixon, C.E.; Henchir, J.; Gorse, K.; Vagni, V.A.; Janesko-Feldman, K.; Kochanek, P.M.; Jackson, T.C. Gene knockout of RNA binding motif 5 in the brain alters RIMS2 protein homeostasis in the cerebellum and Hippocampus and exacerbates behavioral deficits after a TBI in mice. Exp. Neurol. 2024, 374, 114690–114690. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, K.; Rodriguez-Rodriguez, R.; Gaebler, A.; Casals, N.; Scheller, A.; Kuerschner, L. Astrocytes and oligodendrocytes in grey and white matter regions of the brain metabolize fatty acids. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Mazumder, A.G.; Julé, A.M.; Cullen, P.F.; Sun, D. Astrocyte heterogeneity within white matter tracts and a unique subpopulation of optic nerve head astrocytes. iScience 2022, 25, 105568. [Google Scholar] [CrossRef]
- Zhao, B.; Li, T.; Yang, Y.; Wang, X.; Luo, T.; Shan, Y.; Zhu, Z.; Xiong, D.; Hauberg, M.E.; Bendl, J.; et al. Common genetic variation influencing human white matter microstructure. Science 2021, 372, 1304. [Google Scholar] [CrossRef]
- Lawrence, K.E.; Abaryan, Z.; Laltoo, E.; Hernandez, L.M.; Gandal, M.J.; McCracken, J.T.; Thompson, P.M. White matter microstructure shows sex differences in late childhood: Evidence from 6797 children. Hum. Brain Mapp. 2023, 44, 535–548. [Google Scholar] [CrossRef]
- Ursache, A.; Noble, K.G. ; the Pediatric Imaging, Neurocognition and Genetics Study Socioeconomic status, white matter, and executive function in children. Brain Behav. 2016, 6, e00531. [Google Scholar] [CrossRef]
- Salminen, L.E.; Tubi, M.A.; Bright, J.; Thomopoulos, S.I.; Wieand, A.; Thompson, P.M. Sex is a defining feature of neuroimaging phenotypes in major brain disorders. Hum. Brain Mapp. 2022, 43, 500–542. [Google Scholar] [CrossRef]
- Ho, T.C.; Colich, N.L.; Sisk, L.M.; Oskirko, K.; Jo, B.; Gotlib, I.H. Sex differences in the effects of gonadal hormones on white matter microstructure development in adolescence. Dev. Cogn. Neurosci. 2020, 42, 100773. [Google Scholar] [CrossRef] [PubMed]
- van Hemmen, J.; Saris, I.M.J.; Cohen-Kettenis, P.T.; Veltman, D.J.; Pouwels, P.J.W.; Bakker, J. Sex Differences in White Matter Microstructure in the Human Brain Predominantly Reflect Differences in Sex Hormone Exposure. Cereb. Cortex 2017, 27, bhw156–3001. [Google Scholar] [CrossRef] [PubMed]
- Bartnik-Olson, B.L.; Holshouser, B.; Ghosh, N.; Oyoyo, U.E.; Nichols, J.G.; Pivonka-Jones, J.; Tong, K.; Ashwal, S. Evolving White Matter Injury following Pediatric Traumatic Brain Injury. J. Neurotrauma 2021, 38, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Palacios, E.M.; Yuh, E.L.; Mac Donald, C.L.; Bourla, I.; Wren-Jarvis, J.; Sun, X.; Vassar, M.J.; Diaz-Arrastia, R.; Giacino, J.T.; Okonkwo, D.O.; et al. Diffusion Tensor Imaging Reveals Elevated Diffusivity of White Matter Microstructure that Is Independently Associated with Long-Term Outcome after Mild Traumatic Brain Injury: A TRACK-TBI Study. J. Neurotrauma 2022, 39, 1318–1328. [Google Scholar] [CrossRef]
- Owens, J.A.; Spitz, G.; Ponsford, J.L.; Dymowski, A.R.; Willmott, C. An investigation of white matter integrity and attention deficits following traumatic brain injury. Brain Inj. 2018, 32, 776–783. [Google Scholar] [CrossRef]
- Ryan, N.P.; Genc, S.; Beauchamp, M.H.; Yeates, K.O.; Hearps, S.; Catroppa, C.; Anderson, V.A.; Silk, T.J. White matter microstructure predicts longitudinal social cognitive outcomes after paediatric traumatic brain injury: a diffusion tensor imaging study. Psychol. Med. 2018, 48, 679–691. [Google Scholar] [CrossRef]
- Spitz, G.; Alway, Y.; Gould, K.R.; Ponsford, J.L. Disrupted White Matter Microstructure and Mood Disorders after Traumatic Brain Injury. J. Neurotrauma 2017, 34, 807–815. [Google Scholar] [CrossRef]
- Fakhran, S.; Yaeger, K.; Collins, M.; Alhilali, L. Sex Differences in White Matter Abnormalities after Mild Traumatic Brain Injury: Localization and Correlation with Outcome. Radiology 2014, 272, 815–823. [Google Scholar] [CrossRef]
- Hermens, D.F.; Jamieson, D.; Fitzpatrick, L.; Sacks, D.D.; Iorfino, F.; Crouse, J.J.; Guastella, A.J.; Scott, E.M.; Hickie, I.B.; Lagopoulos, J. Sex differences in fronto-limbic white matter tracts in youth with mood disorders. Psychiatry Clin. Neurosci. 2022, 76, 481–489. [Google Scholar] [CrossRef]
- Wheelock, M.D.; Goodman, A.; Harnett, N.; Wood, K.; Mrug, S.; Granger, D.; Knight, D. Sex-related Differences in Stress Reactivity and Cingulum White Matter. Neuroscience 2021, 459, 118–128. [Google Scholar] [CrossRef]
- Lowet, D.S.; Vaida, F.; Hesselink, J.R.; Ewing-Cobbs, L.; Schachar, R.J.; Chapman, S.B.; Bigler, E.D.; Wilde, E.A.; Saunders, A.E.; Yang, T.T.; et al. Novel Oppositional Defiant Disorder or Conduct Disorder 24 Months After Traumatic Brain Injury in Children and Adolescents. J. Neuropsychiatry 2024, 36, 53–62. [Google Scholar] [CrossRef]
- Nishat, E.; Scratch, S.E.; Ameis, S.H.; Wheeler, A.L. Disrupted Maturation of White Matter Microstructure After Concussion Is Associated With Internalizing Behavior Scores in Female Children. Biol. Psychiatry 2024, 96, 300–308. [Google Scholar] [CrossRef]
- Nishat, E.; Stojanovski, S.; Scratch, S.E.; Ameis, S.H.; Wheeler, A.L. Premature white matter microstructure in female children with a history of concussion. Dev. Cogn. Neurosci. 2023, 62, 101275. [Google Scholar] [CrossRef]
- Wright, D.K.; Symons, G.F.; O’bRien, W.T.; McDonald, S.J.; Zamani, A.; Major, B.; Chen, Z.; Costello, D.; Brady, R.D.; Sun, M.; et al. Diffusion Imaging Reveals Sex Differences in the White Matter Following Sports-Related Concussion. Cereb. Cortex 2021, 31, 4411–4419. [Google Scholar] [CrossRef]
- Chahal, R.; Delevich, K.; Kirshenbaum, J.S.; Borchers, L.R.; Ho, T.C.; Gotlib, I.H. Sex differences in pubertal associations with fronto-accumbal white matter morphometry: Implications for understanding sensitivity to reward and punishment. NeuroImage 2021, 226, 117598–117598. [Google Scholar] [CrossRef]
- Karlsgodt, K.H.; John, M.; Ikuta, T.; Rigoard, P.; Peters, B.D.; Derosse, P.; Malhotra, A.K.; Szeszko, P.R. The accumbofrontal tract: Diffusion tensor imaging characterization and developmental change from childhood to adulthood. Hum. Brain Mapp. 2015, 36, 4954–4963. [Google Scholar] [CrossRef] [PubMed]
- Shulman, E.P.; Harden, K.P.; Chein, J.M.; Steinberg, L. Sex Differences in the Developmental Trajectories of Impulse Control and Sensation-Seeking from Early Adolescence to Early Adulthood. J. Youth Adolesc. 2015, 44, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Roediger, D.J.; Griffin, C.; Marin, F.V.; Verdoorn, H.; Fiecas, M.; A Mueller, B.; O Lim, K.; Camchong, J. Relating white matter microstructure in theoretically defined addiction networks to relapse in alcohol use disorder. Cereb. Cortex 2023, 33, 9756–9763. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, K.S.; Maleki, N.; Papadimitriou, G.; Makris, N.; Oscar-Berman, M.; Harris, G.J. Cerebral white matter sex dimorphism in alcoholism: a diffusion tensor imaging study. Neuropsychopharmacology 2018, 43, 1876–1883. [Google Scholar] [CrossRef]
- Gennatas, E.D.; Avants, B.B.; Wolf, D.H.; Satterthwaite, T.D.; Ruparel, K.; Ciric, R.; Hakonarson, H.; Gur, R.E.; Gur, R.C. Age-Related Effects and Sex Differences in Gray Matter Density, Volume, Mass, and Cortical Thickness from Childhood to Young Adulthood. J. Neurosci. 2017, 37, 5065–5073. [Google Scholar] [CrossRef] [PubMed]
- Lotze, M.; Domin, M.; Gerlach, F.H.; Gaser, C.; Lueders, E.; Schmidt, C.O.; Neumann, N. Novel findings from 2,838 Adult Brains on Sex Differences in Gray Matter Brain Volume. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Guadalupe, T.; Mathias, S.R.; Vanerp, T.G.M.; Whelan, C.D.; Zwiers, M.P.; Abe, Y.; Abramovic, L.; Agartz, I.; Andreassen, O.A.; Arias-Vásquez, A.; et al. Human subcortical brain asymmetries in 15,847 people worldwide reveal effects of age and sex. Brain Imaging Behav. 2017, 11, 1497–1514. [Google Scholar] [CrossRef]
- Kurth, F.; Schijven, D.; Heuvel, O.A.v.D.; Hoogman, M.; van Rooij, D.; Stein, D.J.; Buitelaar, J.K.; Bölte, S.; Auzias, G.; Kushki, A.; et al. Large-scale analysis of structural brain asymmetries during neurodevelopment: Associations with age and sex in 4265 children and adolescents. Hum. Brain Mapp. 2024, 45, e26754. [Google Scholar] [CrossRef]
- Núñez, C.; Theofanopoulou, C.; Senior, C.; Cambra, M.R.; Usall, J.; Stephan-Otto, C.; Brébion, G. A large-scale study on the effects of sex on gray matter asymmetry. Anat. Embryol. 2018, 223, 183–193. [Google Scholar] [CrossRef]
- Belchev, Z.; Gilboa, A.; Binns, M.; Colella, B.M.; Glazer, J.M.; Mikulis, D.J.M.; Green, R.E. Progressive Neurodegeneration Across Chronic Stages of Severe Traumatic Brain Injury. J. Head Trauma Rehabilitation 2022, 37, E144–E156. [Google Scholar] [CrossRef]
- Poudel, G.R.; D, J.F.D.; Verhelst, H.; Linden, C.V.; Deblaere, K.; Jones, D.K.; Cerin, E.; Vingerhoets, G.; Caeyenberghs, K. Network diffusion modeling predicts neurodegeneration in traumatic brain injury. Ann. Clin. Transl. Neurol. 2020, 7, 270–279. [Google Scholar] [CrossRef]
- Mayer, A.R.; Meier, T.B.; Dodd, A.B.; Stephenson, D.D.; Robertson-Benta, C.R.; Ling, J.M.; Reddy, S.P.; Zotev, V.; Vakamudi, K.; Campbell, R.A.; et al. Prospective Study of Gray Matter Atrophy Following Pediatric Mild Traumatic Brain Injury. Neurology 2023, 100, e516–e527. [Google Scholar] [CrossRef]
- Shida, A.F.; Massett, R.J.; Imms, P.; Vegesna, R.V.; Amgalan, A.; Irimia, A. Significant Acceleration of Regional Brain Aging and Atrophy After Mild Traumatic Brain Injury. Journals Gerontol. Ser. A 2023, 78, 1328–1338. [Google Scholar] [CrossRef]
- Ren, H.; Li, J.; Lin, Z.; Xu, Y.; Bi, H.-Y.; Xu, M.; Yang, Y. Developmental changes in brain activation and functional connectivity during Chinese handwriting. NeuroImage 2025, 317, 121330. [Google Scholar] [CrossRef] [PubMed]
- Risen, S.; Barber, A.; Mostofsky, S.; Suskauer, S. Altered functional connectivity in children with mild to moderate TBI relates to motor control. J. Pediatr. Rehabilitation Med. 2015, 8, 309–319. [Google Scholar] [CrossRef]
- Stephens, J.A.; Salorio, C.F.; Barber, A.D.; Risen, S.R.; Mostofsky, S.H.; Suskauer, S.J. Preliminary findings of altered functional connectivity of the default mode network linked to functional outcomes one year after pediatric traumatic brain injury. Dev. Neurorehabilit. 2018, 21, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Tuerk, C.; Dégeilh, F.; Catroppa, C.; Dooley, J.J.; Kean, M.; Anderson, V.; Beauchamp, M.H. Altered resting-state functional connectivity within the developing social brain after pediatric traumatic brain injury. Hum. Brain Mapp. 2020, 41, 561–576. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.; Nowikow, C.; DeMatteo, C.; Noseworthy, M.D.; Timmons, B.W. Sex-specific differences in resting-state functional brain activity in pediatric concussion. Sci. Rep. 2023, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Muetzel, R.L. , et al. Resting-state networks in 6-to-10 year old children. Hum Brain Mapp 2016, 37, 4286–4300. [Google Scholar] [CrossRef]
- Satterthwaite, T.D.; Wolf, D.H.; Roalf, D.R.; Ruparel, K.; Erus, G.; Vandekar, S.; Gennatas, E.D.; Elliott, M.A.; Smith, A.; Hakonarson, H.; et al. Linked Sex Differences in Cognition and Functional Connectivity in Youth. Cereb. Cortex 2015, 25, 2383–2394. [Google Scholar] [CrossRef]
- Supekar, K.; Musen, M.; Menon, V. Development of Large-Scale Functional Brain Networks in Children. PLOS Biol. 2009, 7, e1000157. [Google Scholar] [CrossRef]
- Schmidt, A.T.; Hanten, G.R.; Li, X.; Orsten, K.D.; Levin, H.S. Emotion recognition following pediatric traumatic brain injury: Longitudinal analysis of emotional prosody and facial emotion recognition. Neuropsychologia 2010, 48, 2869–2877. [Google Scholar] [CrossRef]
- Agcaoglu, O.; Muetzel, R.L.; Rashid, B.; White, T.; Tiemeier, H.; Calhoun, V.D. Lateralization of Resting-State Networks in Children: Association with Age, Sex, Handedness, Intelligence Quotient, and Behavior. Brain Connect. 2022, 12, 246–259. [Google Scholar] [CrossRef]
- Ding, L.; Pang, G. Identification of Brain Regions with Enhanced Functional Connectivity with the Cerebellum Region in Children with Attention Deficit Hyperactivity Disorder: A Resting-State fMRI Study. Int. J. Gen. Med. 2021, 14, 2109–2115. [Google Scholar] [CrossRef]
- Johnson, G.M.; Wild, J.T.; Burgess, J.K.; McCracken, K.; Malekian, S.; Turner, J.A.; King, K.; Kwon, S.; Carl, R.L.; LaBella, C.R. Assessment of post-concussion emotional symptom load using PCSS and PROMIS instruments in pediatric patients. Physician Sportsmed. 2024, 52, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Rigon, A.; Duff, M.C.; McAuley, E.; Kramer, A.F.; Voss, M.W. Is Traumatic Brain Injury Associated with Reduced Inter-Hemispheric Functional Connectivity? A Study of Large-Scale Resting State Networks following Traumatic Brain Injury. J. Neurotrauma 2016, 33, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Shumskaya, E.; van Gerven, M.A.J.; Norris, D.G.; Vos, P.E.; Kessels, R.P.C. Abnormal connectivity in the sensorimotor network predicts attention deficits in traumatic brain injury. Exp. Brain Res. 2017, 235, 799–807. [Google Scholar] [CrossRef]
- Threlkeld, Z.D.; Bodien, Y.G.; Rosenthal, E.S.; Giacino, J.T.; Nieto-Castanon, A.; Wu, O.; Whitfield-Gabrieli, S.; Edlow, B.L. Functional networks reemerge during recovery of consciousness after acute severe traumatic brain injury. Cortex 2018, 106, 299–308. [Google Scholar] [CrossRef]
- Stephens, J.A.; Salorio, C.E.; Gomes, J.P.; Nebel, M.B.; Mostofsky, S.H.; Suskauer, S.J. Response Inhibition Deficits and Altered Motor Network Connectivity in the Chronic Phase of Pediatric Traumatic Brain Injury. J. Neurotrauma 2017, 34, 3117–3123. [Google Scholar] [CrossRef]
- Wang, S.; Hu, L.; Cao, J.; Huang, W.; Sun, C.; Zheng, D.; Wang, Z.; Gan, S.; Niu, X.; Gu, C.; et al. Sex Differences in Abnormal Intrinsic Functional Connectivity After Acute Mild Traumatic Brain Injury. Front. Neural Circuits 2018, 12, 107. [Google Scholar] [CrossRef]
- Subramaniapillai, S. , et al. Age- and Episodic Memory-related Differences in Task-based Functional Connectivity in Women and Men. J Cogn Neurosci 2022, 34, 1500–1520. [Google Scholar] [CrossRef] [PubMed]
- Subramaniapillai, S.; Rajagopal, S.; Elshiekh, A.; Pasvanis, S.; Ankudowich, E.; Rajah, M.N. Sex Differences in the Neural Correlates of Spatial Context Memory Decline in Healthy Aging. J. Cogn. Neurosci. 2019, 31, 1895–1916. [Google Scholar] [CrossRef]
- Cao, M.; Halperin, J.M.; Li, X. Abnormal Functional Network Topology and Its Dynamics during Sustained Attention Processing Significantly Implicate Post-TBI Attention Deficits in Children. Brain Sci. 2021, 11, 1348. [Google Scholar] [CrossRef]
- Westfall, D.R.; West, J.D.; Bailey, J.N.; Arnold, T.W.; Kersey, P.A.; Saykin, A.J.; McDonald, B.C. Increased brain activation during working memory processing after pediatric mild traumatic brain injury (mTBI). J. Pediatr. Rehabil. Med. 2015, 8, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.Y.; Di, X.; Yu, X.; Biswal, B.B. The significance and limited influence of cerebrovascular reactivity on age and sex effects in task- and resting-state brain activity. Cereb. Cortex 2024, 34. [Google Scholar] [CrossRef]
- Andrushko, J.W.; Rinat, S.; Kirby, E.D.; Dahlby, J.; Ekstrand, C.; Boyd, L.A. Females exhibit smaller volumes of brain activation and lower inter-subject variability during motor tasks. Sci. Rep. 2023, 13, 1–12. [Google Scholar] [CrossRef]
- Narad, M.E.; Riemersma, J.; Wade, S.L.; Smith-Paine, J.; Morrison, P.; Taylor, H.G.; Yeates, K.O.; Kurowski, B.G. Impact of Secondary ADHD on Long-Term Outcomes After Early Childhood Traumatic Brain Injury. J. Head Trauma Rehabil. 2020, 35, E271–E279. [Google Scholar] [CrossRef]
- Le Fur, C.; Câmara-Costa, H.; Francillette, L.; Opatowski, M.; Toure, H.; Brugel, D.; Laurent-Vannier, A.; Meyer, P.; Watier, L.; Dellatolas, G.; et al. Executive functions and attention 7 years after severe childhood traumatic brain injury: Results of the Traumatisme Grave de l’Enfant (TGE) cohort. Ann. Phys. Rehabil. Med. 2020, 63, 270–279. [Google Scholar] [CrossRef]
- Cao, M.; Wu, K.; Halperin, J.M.; Li, X. Abnormal structural and functional network topological properties associated with left prefrontal, parietal, and occipital cortices significantly predict childhood TBI-related attention deficits: A semi-supervised deep learning study. Front. Neurosci. 2023, 17, 1128646. [Google Scholar] [CrossRef]
- Strazzer, S.; Rocca, M.A.; Molteni, E.; De Meo, E.; Recla, M.; Valsasina, P.; Arrigoni, F.; Galbiati, S.; Bardoni, A.; Filippi, M. Altered Recruitment of the Attention Network Is Associated with Disability and Cognitive Impairment in Pediatric Patients with Acquired Brain Injury. Neural Plast. 2015, 2015, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Manelis, A.; Santos, J.P.L.; Suss, S.J.; Holland, C.L.; Perry, C.A.; Hickey, R.W.; Collins, M.W.; Kontos, A.P.; Versace, A. Working Memory Recovery in Adolescents with Concussion: Longitudinal fMRI Study. J. Clin. Med. 2024, 13, 3585. [Google Scholar] [CrossRef] [PubMed]
- Tlustos, S.J.; Chiu, C.P.; Walz, N.C.; Wade, S.L. Neural substrates of inhibitory and emotional processing in adolescents with traumatic brain injury. J. Pediatr. Rehabil. Med. 2015, 8, 321–333. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sex Differences in Neuroanatomy, Neurodevelopment, and Functional Outcomes. Variations in anatomical volume and development of critical brain structures including the basal ganglia, hypothalamus, pituitary gland, thalamus, hippocampus, and cerebellum may contribute to differences in TBI severity and recovery outcomes commonly observed between male and female children.
Figure 1.
Sex Differences in Neuroanatomy, Neurodevelopment, and Functional Outcomes. Variations in anatomical volume and development of critical brain structures including the basal ganglia, hypothalamus, pituitary gland, thalamus, hippocampus, and cerebellum may contribute to differences in TBI severity and recovery outcomes commonly observed between male and female children.
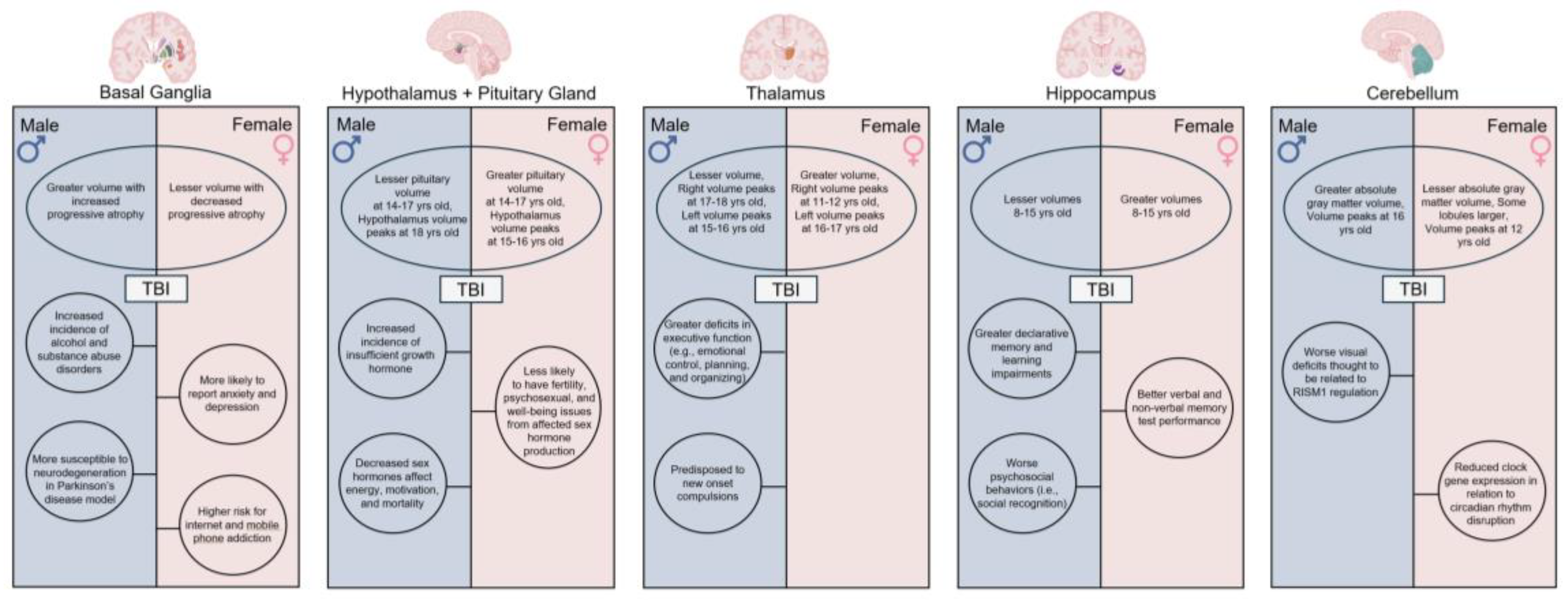
Figure 2.
Sex Differences in Resting State and Task Based Functional Connectivity. Resting state and task based functional connectivity analyses demonstrate unique differences in network organization, lateralization, and patterns of connectivity that are dependent upon both the age and sex of TBI patients.
Figure 2.
Sex Differences in Resting State and Task Based Functional Connectivity. Resting state and task based functional connectivity analyses demonstrate unique differences in network organization, lateralization, and patterns of connectivity that are dependent upon both the age and sex of TBI patients.
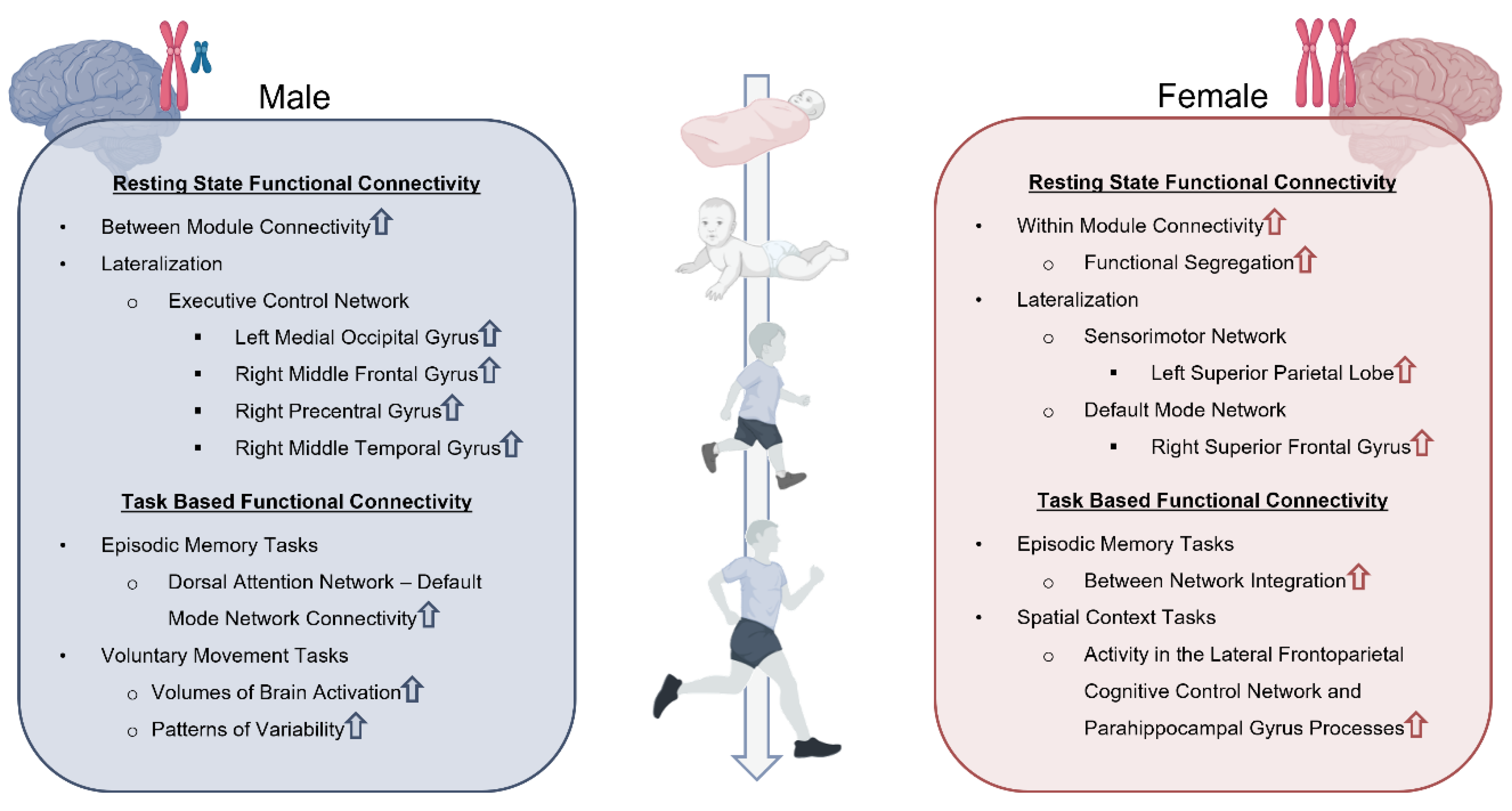
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



