Submitted:
28 October 2025
Posted:
29 October 2025
You are already at the latest version
Abstract
The transcription factor Pax6 acts as a molecular switch regulating various phases of the cell cycle during the early stages of embryonic neurogenesis and serves as a molecular mediator integrating multiple signaling pathways involved in the development of pathophysiological conditions. In this regard, Pax6 is considered an attractive molecular target for developing new drugs aimed at combating neurodegeneration, oncology, and aniridia. The aim of this review is to examine the range of studies describing various Pax6-dependent molecular pathways to identify similarities and differences in the signaling cascades activated by Pax6 in health and disease. These insights may provide a basis for developing new therapeutic strategies targeting Pax6 as a molecular regulator. This review summarizes the data indicating the leading regulatory role of Pax6 in the neuronal cell cycle in health and pathology. It is possible that Pax6 may act as a therapeutic target in certain pathophysiological conditions; however, the effectiveness of such a strategy will depend on the substrate chain in the signaling pathway, its branching, and the redundancy of mediators involved.
Keywords:
Pax6
; cell cycle
; proliferation
; neurogenesis
; aniridia
; neurodegeneration
; oncology
1. Introduction
Pax6 is a pleiotropic factor [1] and is considered a master gene in the control of eye morphogenesis and evolution [2,3]. However, its function is not limited to vision. Pax6 is known to play a role in the development of the central nervous system (CNS) [4], nose, pancreas and pituitary gland [5,6]. Pax6 controls neurogenesis and gliogenesis [7,8], also plays a role in establishing boundaries between developing areas of the forebrain [9]. The Pax6 gene acts as a molecular switch that controls the specification and differentiation of different cell types. The production of the Pax6 gene protein is regulated at the pre-mRNA and post-transcriptional levels by microRNAs [10,11]. The role of Pax6 as a dual transcriptional activator and repressor is known [12,13,14]. During embryonic development, Pax6 protein activates several target genes using two DNA-binding domains and a proline-serine-threonine transcriptional activation domain [15,16,17]. The regulatory role of Pax6 is associated with gene expression by recruiting chromatin remodeling enzymes and alleviating heterochromatin repression [18,19]. Pax6 has been shown to influence the morphology and electrophysiological properties of neurons in the prethalamic neuroepithelium [20]. The aim of this review was to examine the range of studies describing various molecular signaling pathways involving Pax6, to find similarities and differences in the signaling cascades that Pax6 can activate in health and disease, which will allow these data to be used to search for new therapeutic strategies targeting Pax6 as a molecular target.
2. Regulatory Role of Pax6 in Embryogenesis
2.1. Molecular Organization of the Pax6 Gene, Its Isoforms and Functions
The Pax6 gene encodes a tissue-specific transcriptional factor located on the short arm of chromosome 11 (locus 11p13) [21]. The Pax6 gene has been shown to be highly conserved among vertebrate and invertebrate species. The pared box (pax) gene family was discovered in Drosophila as segmentation genes such as paired (prd), gooseberry-distal (gsb-d), and gooseberry-proximal (gsb-p), then in other animal species. Based on sequence homology to the Drosophila pared box gsb-d, nine pax genes have been identified in mouse (pax1-9) and humans (Pax1-9). All Pax genes contain a 128-amino acid paired N-terminal box that can be present alone (Pax1 and Pax9) or with a full-length (Pax3, Pax4, Pax6, and Pax7) or truncated (Pax2, Pax5, and Pax8) paired homeodomain [22].
The structure of the gene and its encoded protein have been studied in detail and described in a number of works [1,23,24,25,26]. Pax6 consists of 14 exons, its expression is controlled by three independent promoters (P0, P1, and Pα) [27], which produce a number of alternative transcripts [28]. Genomic DNA sequencing showed that the Pax6 promoter has a TATA-like box (ATATTTT) at position -26 base pairs (bp), two CCAAT boxes are located at positions -70 and -100 bp, and a 38 bp poly(CA) sequence is located 992 bp upstream of the initiation site [22].
Pax6 expression is restricted to specific regions of the forebrain, optic cup, hindbrain, and spinal cord, as well as the lens placode and nasal epithelium [22]. The Pax6 gene is necessary for the development of the CNS, plays a key role in the morphogenesis of the eye, participates in secondary inductive interaction during the formation of the lens [22] and is also critical for the development and function of endocrine cells [29]. Pax6 is also required for the maintenance of the progenitor cell pool in the cerebral cortex and spinal cord, and the multipotency of progenitor cells in the retina [10]. Heterozygous mutations in the Pax6 gene result in eye abnormalities in humans (aniridia), mice (small eyes), and Drosophila melanogaster (absent eyes) [22].
Nowadays, more than 700 pathogenic variants of the Pax6 gene have been identified. A significant portion of these variants result in premature stop codons due to nonsense codons, frameshifts, or splicing errors, which results in haploinsufficiency of the Pax6 protein. Missense variants are predominantly localized in the paired and, less commonly, homeobox domain of Pax6. According to recent studies, the number of pathogenic splicing variants is up to 30% of all pathogenic variants of the Pax6 gene. Most of them were identified in the terminal AG-GT dinucleotides of the canonical acceptor (3′SS) and donor (5′SS) splice sites. Variants affecting splicing have also been identified in non-canonical splice site nucleotides, regulatory elements, deep intronic sequences, and 5′- and 3′-untranslated regions (UTRs) of the Pax6 gene [28].
The Pax6 gene produces the Pax6 protein, the expression level of which is tightly regulated at the pre-mRNA, post-transcriptional, and protein levels [30]. Too low or too high levels of Pax6 may have profound effects on tissue development and maintenance. However, the molecular mechanisms regulating the expression level of Pax6 are still unclear. Post-transcriptional regulation of Pax6 by microRNAs may represent an important mechanism for maintaining Pax6 expression levels [10,30].
The mammalian PAX6 gene encodes three isoforms: the “canonical” Pax6, Pax6(5a), and Pax6(ΔPD). The canonical Pax6 protein consists of 422 amino acids and contains a DNA-binding paired domain (PD) near the N-terminus (128 amino acid residues), a paired homeodomain (HD) (61 amino acid residues) separated by a flexible linker, and a proline-serine-threonine-rich domain (PST) (152 amino acid residues) at the C-terminus [28]. Both PD and HD have DNA binding activity, whereas the PST domain has transactivation function [16,31,32,33]. The paired domain starts at the N-terminus of the protein and consists of two subdomains (PAI and RED), each of which has a conserved helix-turn-helix (HTH) structure that binds to DNA. The homeobox domain is formed by three α-helices and has its own pattern of interaction with DNA binding sites. The paired box and the homeobox are connected by a 72-amino acid linker that is involved in interaction with the minor groove of DNA [26]. At the C-terminus of the protein is the PST domain sequence (proline, serine and threonine-rich region) containing 152 amino acids that interact with transcriptional coregulators [1]. The 40 amino acids of the PST domain form a highly conserved C-terminal portion of the protein that links the homeodomain to target sequences [23].
The Pax6(5a) isoform is formed by alternative splicing preserving exon 5a and contains a 14-residue insertion in the PD N-terminal subdomain, which alters the specificity of the DNA binding [17,32]. Pax6 and Pax6(5a) mRNA are expressed in the telencephalon, diencephalon and rhombencephalon with expression levels 6-10-times higher than that of Pax6(5a) during early neurogenesis [34]. The vertebrate Pax6 gene also expresses a third isoform, Pax6ΔPD, which lacks PD and is produced by an internal Pα promoter (located between exons 4 and 5) and an alternative translation start codon in exon 7 [35,36]. Despite its unknown physiological role, Pax6ΔPD functions differently from PD-containing Pax6 isoforms [28]. The over expression of Pax6(PD) induces a severe microphthalmic phenotype, which is caused by apoptotic cell death in the lens during embryonic development [36,37]. Pax6 paired domain is necessary for the regulation of neurogenesis, cell proliferation and is crucial for normal brain development [27,38]. The expression of the Pax6 locus is regulated by three promoters (P0, P1 and Pα) [39]. Pax6 levels are regulated by positive and negative autoregulation in vitro [40]. In the mouse optic vesicle, Pax6 overexpression suppresses neuroepithelial cell proliferation [41].
At the molecular level, the Pax6 protein regulates transcription of target genes involved in embryonic development and organogenesis [13,42,43,44]. It is also noted that Pax6 is involved in maintaining the multipotent state of several progenitor cell lines, such as the neuronal retina, retinal pigment epithelium, the iris [45,46,47], specific subpopulations of granular and periglomerular neurons of the olfactory bulbs [48] as well as the cortex and subcortical structures of the brain [49,50]. The presence of various functional domains in the Pax6 protein molecule ensures its regulatory function not only in cell proliferation processes [51,52] but also in differentiation [29]. Protein Pax6 competes with Pax4 in binding to the common element in the glucagon, insulin and somatostatin promoters. Pax4 acts primarily as a transcriptional repressor, whereas Pax6 transactivates the glucagon, insulin, and somatostatin genes [53].
In the adults, the Pax6 protein is involved in the formation of new neurons in the dentate gyrus of the hippocampus and the subventricular zone of the lateral ventricle [54], also determines the phenotype of forming dopaminergic neurons [55] and neurons of the olfactory bulb [56,57]. In the adult eye, Pax6 is involved in maintaining the structure and function of the eye by regulating the proliferation and differentiation of iris cells during lens regeneration [58]. In the pancreas, Pax6 is involved in its development and the formation of various cell phenotypes, while in the adult organism its regulatory function is associated with the maintenance of endocrine function and glucose homeostasis [59].
Thus, Pax6 is an early universal regulator of morphogenesis, ensuring the correct formation of the eyes, neurons in the brain and pancreatic cells. Its ability to regulate gene expression at various levels makes it a key factor that coordinates complex developmental processes at the stage of embryogenesis, as well as an important factor in maintaining tissue homeostasis in the adults.
2.2. Pax6 Expression and Pathways in CNS Development
Pax6 is expressed in distinct spatiotemporal patterns during mammalian brain development. It is thought to be upstream of gene networks involved in brain patterning, neuronal migration and neural chains formation [60,61]. At the neural plate stage, which consists of proliferating neuroepithelial cells, early Pax6 expression was detected at embryonic day 8 in mice and day 10 in rats. Pax6 is expressed in the forebrain and spinal cord [62], in the germinal zone of the lateral ventricular wall, and in a small number of neurons in the marginal zone, near the brain surface on the 10th day of embryonic development [63]. The thickness of the germinal layer increases up to the 12th day of embryonic development during which time a sufficient number of glutamatergic pyramidal neurons are formed. Inhibitory interneurons from the ventral telencephalon are formed which produce gamma-aminobutyric acid in the cortical rudiment [64,65]. Pax6 expression persists in the germinal layer of the dorsal telencephalon and the border region between the dorsal and ventral telencephalon at the stage of neuronal embryogenesis [66]. Subsequently, the thickness of the germinal layer decreases and the ependymal layer is formed that expresses Pax6 [54]. The level of Pax6 expression maintains the balance between proliferation and differentiation of neuronal precursors in the cerebral cortex [67].
Pax6 has been shown to target a large number of promoters in neuronal progenitor cells. Pax6 has a dual role in which it activates neuronal (ectodermal) genes while simultaneously suppressing meso- and endodermal genes, thereby providing unidirectional lines to neuronal differentiation ensuring a unidirectional lineage toward neuronal differentiation [42]. Pax6 plays a regulatory role at the earliest stages of cortical neurogenesis and is involved in the development of the forebrain. In the works [42,49,68,69,70,71,72] on the model of mice with the mutation of the Pax6 gene (Pax6-/-) the disturbance in the development of the cortex, the decrease in the number of neurons, which leads to the formation of a thin cortical plate was proved. The mechanism of this phenomenon was studied in the work [73], according to the results, a decrease in the production of neurons in the cortex of mutant mice is associated with the early depletion of the pool of precursors through the exit of dividing cells from the cell cycle. In the paper [72] an analysis of the kinetics of the cell cycle was performed, and it was proven that the absence of Pax6 accelerates cortical neurogenesis in vivo. In rodents with homozygous mutations of Pax6 (Pax6Sey/Sey) a decrease in the thickness of the forebrain and cortex was observed [69]. Humans with Pax6 haploinsufficiency reduction frontal-parietal cortex thickness has been shown [74]. Patients with heterozygous Pax6 mutations have been shown to have reduced frontoparietal cortex thickness, which has also been observed in mice with small eyes.
In patients with heterozygous Pax6 mutations, a reduction in the thickness of the frontal-parietal cortex was found, which was also observed in mice with small eyes. In the study [52], it was found that the overexpression of Pax6 increased the thickness of the cortex, especially in the intermediate zone. The researchers associate this phenomenon with the retention of neurons in the intermediate zone and their proliferation. It was found that the effect of Pax6 overexpression on the cortical thickness and neuronal migration was temporary and is explained by a change in the ratio of Pax6 isoforms through autoregulation. The researchers do not exclude that Pax6 regulates neuronal migration and cell proliferation through interaction with Wnt3.
In the development of the cortex researchers attach importance to the synergistic interaction of Pax6 with downstream target genes such as FABP7, Ngn2, p271, and Wnt7b1 [14,75,76]. In the lens, the neurogenesis is regulated by Pax6 signaling pathway involving Wnt—Dkk1, Sfrp1, and Sfrp2 [77]. In the rat embryo hindbrain, a regulatory role of Pax6 in the expression of Cdh7 and Wnt5a has been demonstrated [78]. It is known, that expression of the neural stem cell marker Fabp7/BLBP gene encoding a protein of brain-type fatty acids is suppressed in the forebrain and hindbrain of homozygous Pax6 mutant rats [79].
Pax6 plays a key role in the development of the eye [79]. The absence of Pax6 causes neonatal lethality in mice as well as the eye loss and cortical malformations [80]. Heterozygous mutations of Pax6 cause aniridia in humans and a small eye phenotype in mice [81,82]. In a study of the development of the retina, which expresses Pax6 and is considered to be part of the forebrain, premature neuronal differentiation have shown with Mash1 upregulation in Pax6-/- mutant mouse embryos [83]. Heterozygous loss of Pax6 function (haploinsufficiency) in humans causes various structural brain abnormalities, absence or hypoplasia of the pineal gland, which regulates circadian rhythm and decreased function of the olfactory bulb, unilateral polymicrogyria [84,85,86].
One of the key signaling pathways triggered by neurogenesis involving Pax6 is the Wnt/β-catenin pathway. This pathway is the most important regulator of stem cells and plays a monumental role in neuroepithelial cells at the early stages of embryogenesis. It has been established that β-catenin binds to the Pax6 promoter and activates its transcription. In β-catenin-deficient neural stem cells increased Pax6 expression partially restores impaired proliferation and neurogenesis. Therefore, Pax6 is a downstream target of the Wnt/β-catenin pathway and β-catenin/Pax6 signaling regulates self-renewal and neurogenesis of radial glia/neural stem cells during neocortical development [87].
In early neural tube development as well as in late retinal cell development Pax6 is involved in the fibroblast growth factor (FGF) signaling pathway. Pax6 activation has been shown to occur during somitogenesis in the spinal cord. Presomitic mesoderm can inhibit Pax6 expression via the FGF signaling pathway. In presomitic mesoderm decreased Pax6 activation was observed in response to an FGF8 gradient. On the contrary, disruption of the FGF signal resulted in premature activation of Pax6 in the neural plate. The authors conclude that the decrease of FGF8 activity acts as a transition from the early (posterior) to the later (advanced) stage of neural epithelium development [88]. The fibroblast growth factor signaling pathway is mediated by Pax6 in neuronal differentiation of the spinal cord. In the small-eyed mouse model (Sey/Sey) a role for Pax6 in the specification of ventral spinal cord neuron subtypes has been demonstrated. Pax6 controls the expansion of the dorsal neural tube [89,90] and changes in motor neuron specification [91]. Activation of Pax6 is controlled by signaling pathways that regulate neuronal differentiation of neurons. In the caudal neural tube, where stem cells are concentrated Pax6 is absent [92] as its activity is suppressed by fibroblast growth factor (FGF) to prevent premature neuronal differentiation [88,93]. Then, Pax6 is activated in the closing neural tube by retinoic acid signaling concomitant with activation of neural determinants [94,95]. The level of Pax6 protein is critical for cell cycle exit and initiation of neuronal differentiation in spinal cord. Once neural progenitors are recruited into the neural program the Pax6 protein should be turned off to provide neuronal differentiation. Moreover, loss-of-function experiments in chick or mouse embryos show that the absence of Pax6 results in premature differentiation of neurons. Direct regulation of the proneuronal gene Ngn2 depending on the level of the Pax6 protein has been established. It has been proven that the suppression of Pax6 expression is controlled by high levels of Ngn2 [96]. The fibroblast growth factor signaling pathway is also involved in the development of the retinal epithelium. According to the work [46], two main targets of the Pax6/Mitf/Tfec signaling pathway were identified: Fgf15 that promotes the development of retinal cells, and Dkk3, which together with FGF inhibits the Wnt signaling pathway and stimulates the development of retinal pigment cells. Pax6 was shown to suppress the expression of Fgf15 and Dkk3. Pax6 has been reported to control cell adhesion properties, the neuronal polarity, the migration and the axon formation via a signaling pathway involving δ-catenin. The cell adhesion is impaired in the optic vesicle of embryos with a homophilic mutation of Pax6 [97]. Abnormalities in neuronal polarity in radial glia and granule cells of the cerebellum have been described in Pax6 mutants [98,99,100]. In paper [101], δ-catenin was identified as a target of Pax6 in the CNS. Coexpression of the δ-catenin protein transcript with Pax6 was proven in the optic vesicle, the neocortex and developing cerebellum. However, δ-catenin expression levels were significantly reduced in the CNS of Pax6 mutants. Pax6 can specifically bind a DNA sequence in the conserved δ-catenin promoter. The authors of the work indicate that other transcription factors can also control δ-catenin expression in the CNS since its expression was preserved in regions outside the Pax6 expression domain.
In general, converging molecular pathways are involved in the regulation of neuronal cytoskeletal dynamics during neurogenesis. A study [99] proposed a link between Pax6 and the Rho GTPase and RhoA pathways in neuronal cytoskeletal dynamics. Inhibition of RhoA stimulates neurite outgrowth while mutation of Pax6 leads to neurite shortening. A number of studies have also shown that R-cadherin expression is reduced in the forebrain with a Pax6 mutation and Pax6-mediated R-cadherin expression can control pioneer axon growth [9,100]. δ-catenin maintains the balance between neurite elongation and branching by acting at the interface between cortactin and Rho [102]. A stimulatory effect of neurite elongation under the action of δ-catenin, which at the same time suppresses the activity of RhoA during neurite branching was shown in experiments on cell cultures [102]. δ-catenin regulates cadherin turnover and links cortactin, RhoA, cadherins in the CNS [103].
Pax6 is involved in gliogenesis through regulation of glutamate transporter expression. Glutamate signaling is critical in neurogenesis because it regulates the migration of astrocyte progenitor cells. The presence of glutamate transporter 1 (GLT1) at the astrocyte plasma membrane ensures low levels of extracellular glutamate and buffers synaptically released glutamate. In addition, GLT1 can function as a signal transduction molecule [104]. The work [105] presents evidence that astrocyte precursors migrate to the cerebral cortex postnatally and their migration is regulated by glutamate signaling. The transcription factor Pax6 induces the expression of the Na+-dependent glutamate transporter GLT1 (EAAT2) in astrocytes by interacting with its conserved distal enhancer element [106]. There is evidence that the GLT1/EAAT2 transporter mediates glutamate uptake in vivo by astroglial cells. An increase in extracellular glutamate concentration and excitotoxic cell death were observed in a decrease in GLT1 expression [107]. Premature death with spontaneous seizures in mice with genetic deletion of GLT1 was demonstrated [108]. Co-culture of astrocytes with neurons induces GLT1 expression in astrocytes. This effect is mediated by both direct cell-cell contact and neuronal soluble factors [109], including cAMP [110] and epidermal growth factor [111]. The astrocytes from mice lacking Pax6 exhibited increased proliferation and higher migration potential [112]. Exogenous Pax6 increased the expression of GLT1, which is a marker of astroglial maturation [106].
Thus, Pax6 is a key transcription factor that is involved in the regulation of various stages of neurogenesis. This regulation can be carried out through direct or indirect interaction with genes or proteins of various signaling pathways controlling the proliferation, differentiation and migration of neurons. Pax6 is a critical and vital factor in the development of individual brain regions such as the retina, olfactory bulb as well as development of various glial cell subtypes in the spinal cord and brain.
3. Pax6 Involvement in the Development of Metabolic Disorders in Patients with Aniridia
The main mechanism underlying the development of aniridia is the haploinsufficiency of the Pax6 gene. This disease is characterized by damage to the cornea, iris, foveolar hypoplasia, glaucoma [113]. The severity of the disease varies depending on which genes are involved in the development of the disease [114]. Milder forms are characterized by changes in the architecture of the iris, good vision and normal foveolar structure. Complex forms are accompanied by changes in the cornea, glaucoma, cataracts, strabismus, optic nerve hypoplasia, decreased visual acuity and progressive loss of vision [115]. It has been reported that the majority of cases of congenital aniridia are attended by metabolic disorders such as thyroid dysfunction, obesity, hypertension and diabetes mellitus [116]. In addition, neurological disorders such as autism, epilepsy, structural brain abnormalities, auditory processing disorders and sleep-wake cycle disturbances in patients with aniridia syndrome are increasingly observed [85].
Currently, there are increasing research results indicating neuronal and endocrine abnormalities in the majority of patients with aniridia. Dysfunction of Pax6 in the regulation of pancreatic function and energy metabolism is recognized as one of the main causes of obesity [117,118], insulin resistance and increased body mass index [119,120], diabetes [120,121] in patients with aniridia. Carriers of the Pax6 mutation to have abnormal glucose tolerance were shown. Oral glucose tolerance tests have shown impaired insulin secretion in all patients with the Pax6 gene mutation [118]. PAX6 may be involved in insulin production and regulation of the conversion of proinsulin to insulin. The heterozygous mutations of the gene are associated with glucose intolerance [120]. The molecular mechanism underlying abnormal glucose metabolism involves a deficiency of the enzyme prohormone convertase PC (1/3), which is necessary for the conversion of proinsulin to insulin [120,122]. This deficiency is caused by the mutations in the PAX6 gene [122].
The study [123] presents a comprehensive metabolomic analysis of plasma markers in patients with congential aniridia. This study identified disturbances in oxidative stress regulation, based on the following parameters:
- an increase in glutathione synthesis metabolites—such as cysteine, α-ketobutyrate, and cysteine-S-sulfate—along with a reduction in metabolites such as cysteine-glutathione disulfide and γ-glutamyl amino acids;
- decreased taurine synthesis, which is associated with neurological disorders (depression, autism spectrum disorders, schizophrenia), hypertension, obesity and hypothyroidism [124];
- a decrease in the level of plasmalogens, which are necessary for myelin formation in the brain and lens, and their levels are known to decrease with age [125].
Deficiency of Pax6 is associated with the formation of cataracts. According to the work [126], 85% of patients with aniridia have early cataracts and lens opacities that correlate with the level of plasmalogens;
- reduced levels of total lysophospholipids, which were identified as negative predictors of type 2 diabetes progression [127];
- elevated levels of dicarboxylic and hydroxy fatty acids in plasma, which indicate disturbances in mitochondrial β-oxidation processes.
The researchers also point to increased levels of ketone bodies and acetoacetate in the plasma of patients with aniridia [123]. It is possible that ketoacidosis may be associated with insulin deficiency and abnormal glucose metabolism. In an experiment on mice with the Pax6 gene deleted from pancreatic β-cells, a decrease in insulin levels, hyperglycemia and ketoacidosis were observed [128]. Investigators note increasing insulin resistance in patients with aniridia which is associated with increased levels of bile acids that regulate lipid and carbohydrate metabolism. Taurolithcholate-3 sulfate incrase in patients with aniridia by almost 2-fold was reported [123]. This compound was proven to induce insulin resistance in rat hepatocytes in vitro [129].
Metabolic defects in patients with aniridia also are accompanied by deregulation of the superfamily of ligand-activated transcription factors (peroxisome proliferator-activated receptors, PPARs). It is known, PPARγ interacts with Pax6 in the regulation of the glucagon gene [130,131]. PAX6 is required for normal proglucagon gene expression in both the pancreas and intestine [132]. Thiazolidinediones inhibit glucagon gene transcription by binding to the PPARγ-activated receptor and inhibiting PAX6 transcriptional activity [130]. PPARγ expression is reduced in limbal stem cells from aniridia patients [133] and in Pax6-edited cells [134].
In general, patients with aniridia are characterized by established metabolic disorders and changes in carbohydrate metabolism, such as insulin resistance, dyslipidemia, abdominal obesity, and the risk of developing type 2 diabetes mellitus.
4. Role of Pax6 in the Development of the Neurodegenerative Diseases
Pax6 plays an important regulatory role not only in neurogenesis during embryonic development but also in pathogenic signaling pathways associated with the development of neurodegenerative diseases. Pax6 was established to tegulate the expression of neurodegenerative genes through all gene cascades involved in the growth, differentiation and maturation of neurons and glia. In the Pax6 gene knockdown state neurodegenerative markers S100β, GFAP, BDNF, NGN2, p73α, p73δ were suppressed. Pax6 may be directly associated with p53 and TGFβ mediated pathways and indirectly with the regulation of redox-sensitive pathways [135].
Alzheimer’s disease. A study [136] showed that Pax6 is an important molecular mediator between amyloid-β hyperphosphorylation and tau protein. Pax6 expression is increased in the brains of APP transgenic mice and patients with Alzheimer’s disease. Researchers observed an increase in the expression of Pax6 and E2F1 protein that regulate cell cycle phases and proliferation in neurons of the entorhinal cortex of patients with mid-stage Alzheimer’s disease [137]. Gradual increase in Pax6 expression as Alzheimer’s disease progresses by 1.2 times in the initial stage and by 1.8 times in severe cases have shown in studies [138]. The mechanism by which amyloid-β activates cellular signaling pathways triggering the transcription factors c-Myb and PAX6 and ultimately leading to hyperphosphorylation of the tau protein is presented in paper [136]. The authors identified a potential target gene for Pax6. This is the kinase GSK3B (GSK-3b), which phosphorylates tau protein and leads to the formation of neurofibrillary tangles. Amyloid-β neurotoxicity causes Pax6 to transactivate GSK-3b. Increased expression of GSK-3b has been described in several models of Alzheimer’s disease as well as in postmortem brain samples from people with Alzheimer’s disease [139,140]. The molecular model proposed by the authors in work [137] presents a pathway for activating expression of nuclear transcription factors through initiation of amyloid-β cell death signals CD1 and CDK4/6, resulting in hyperphosphorylation of the pRB factor. This starts the transcription factor E2F1, which in the nucleus triggers transcription and translation of the Pax6 and c-Myb genes. In addition to this mechanism activated E2F1 and c-Myb also transactivate Pax6. Total tau protein was reduced after Pax6 knockdown at both RNA and protein levels. Pax6 silencing in neurons significantly reduced the expression levels of kinases such as Cdk5 and Mapk1 which phosphorylate tau protein. Therefore, the authors conclude that there are multiple downstream targets of Pax6 involved in the pathogenesis of Alzheimer’s disease and propose the established E2F1/c-Myb/Pax6 pathway as a new target for therapeutic intervention [137].
Parkinson’s disease. In the adult human brain Pax6 may play a role in the development of Parkinson’s disease. Increased survival of the SH-SY5Y cell line, decreased markers of apoptosis and oxidative stress with excessive expression of Pax6 was found in the study [141]. In a cell culture model of the SH-SY5Y line, a relevant model of Parkinson’s disease, expression of the Pax6 gene was induced using tetracycline. Then, the cells were neurotoxicized with rotenone and 1 methyl-4 phenyl-1,2,3,6-tetrahydropyridine. It was found that Pax6 induction improved cell survival, reduced apoptosis and decreased the effect of oxidative damage on mitochondria by increasing cell resistance to programmed cell death. Researchers note that during the development of Parkinson’s disease, the compact and reticular parts of the substantia nigra are differently vulnerable as the pathology progresses. The greatest degree of degeneration is observed in the compact part of the substantia nigra. The reticular part degenerates later as the disease progresses. Pax6 expression is observed in the reticular part of the substantia nigra. It has been proven that Pax6 has a protective function in dopamine-producing cells [142]. We didn’t find in literature describing direct interaction of alpha-synuclein with Pax6. However, there are indications that pathological alpha-synuclein itself is capable of regulating the cell cycle. A study conducted on PC-12 cells engineered to conditionally induce alpha-synuclein expression found an increase in the proliferation rate and the number of cells in the S phase of the cell cycle as well as an increase in mitotic markers and a decrease in the regulation of tumor suppressor markers [143]. Scientists believe that early intracellular accumulation of alpha-synuclein in postmitotic neurons of the human midbrain is accompanied by changes in the expression of microRNA, which leads to the activation of cell cycle genes [144].
Amyotrophic lateral sclerosis. Amyotrophic lateral sclerosis is a neurodegenerative disease that occurs with progressive muscle paralysis, degeneration of upper and lower motor neurons in the cortex, brainstem and spinal cord [145]. The main contribution to the pathogenesis of this disease is made by immune imbalance, which develops as the disease progresses [146]. Progression of amyotrophic lateral sclerosis is regulated by a shift in the balance towards activation of proinflammatory immune factors [147]. Pax6 to be involved in the regulation of inflammatory genes and oxidative stress in neuropathy has been reported. Hydrogen peroxide levels to affect extracellular expression of Pax6 in cultured cells derived from the eye (cornea) and pancreas have been shown [148]. Pax6 has an immunomodulatory function in the brain by activating microglia either through direct interaction with the ionized binding protein 1 (Iba1) of microglia or indirectly through inflammation associated with neurodegeneration [149]. Iba1 is a calcium binding protein that is involved in actin assembly, cell migration and phagocytosis in activated microglia [150]. The protein is triggered by the proinflammatory cytokine IFNγ and forms complexes with L-fimbrin and small GTPases RAC of activated microglia [151].
Thus, the regulatory role of Pax6 in the development of neurodegenerative processes is associated with age-related changes in the brain and its effect on the neuronal cell cycle, as well as intracellular signaling pathways. These pathways are realized either through phosphorylation, as seen in Alzheimer’s disease, or through mechanisms involving apoptosis and oxidative stress, as in Parkinson’s disease. In addition, in the aging brain, Pax6 performs an immunoregulatory function, which may be a decisive factor in changing the balance towards proinflammatory factors in the development of amyotrophic lateral sclerosis.
5. Molecular Mechanisms in the Development of Tumor Degeneration of Tissues with the Participation of Pax6
Glioblastomas. In gliomas Pax6 acts as a tumor suppressor reducing tumor growth [152]. Pax6 expression decreases with increasing glioma malignancy [153,154]. Pax6 has been used as a prognostic marker for malignant astrocytic gliomas where low levels of Pax6 expression in anaplastic astrocytomas and glioblastomas correlate with poor patient outcomes [154]. Glioblastomas express lower levels of Pax6 compared to adjacent healthy tissue. Anaplastic astrocytomas have three-fold higher Pax6 expression compared to glioblastomas [154]. Pax6 mutations in gliomas have not been identified, and the lower expression is thought to be caused by epigenetic changes as the tumors evolve [155]. The regulation role microRNA (miR)-355 in expression of Pax6 was found in tumor cells [156]. It was demonstated that microRNA (miR)-335 is significantly increased in glioma. Inhibition of miR-335 increased the expression of Pax6 protein, whereas upregulation of miR-335 promoted proliferation of U251 cell line, colony formation and invasion, which was suppressed by overexpression of Pax6. In addition, the effect of miR-335 on U251 cell invasion was mediated by modulating the expression of matrix metalloproteinase (MMP)-2 and MMP-9 by targeting Pax6. The authors concluded that Pax6 is a target of miR-335 and has an anti-oncogenic function in glioma development.
It has been shown that the removal of Pax6 in U251 glioblastoma cells alters cell morphology, promotes their increased proliferation, migration and colony formation ability, and enhances their resistance to oxidative stress [157]. Glioblastoma cell proliferation and colony formation are increased when miR-335 causes an approximately 2-fold downregulation of Pax6. Others studies have shown that Pax6 overexpression in U251 cells reduced colony formation ability in soft agar assays but did not affect cell doubling time in U251 [153]. In vivo animal experiments have demonstrated suppression of cell growth by increasing their number in G1 and decreasing them in S phase with temporary overexpression of PAX6 via adenovirus. Later, overexpression Pas 6 induced the dramatic levels of cell death. Repeated subcutaneous and intracranial implantation experiments in nude mice using Pax6-stable transfectants provided compelling evidence that Pax6 suppresses tumor growth in vivo and significantly increases mouse survival.
Prostate cancer. Prostate cancer is associated with abnormal neuroendocrine cell differentiation. Androgens and androgen receptors are involved in the normal development and function of the prostate gland. Abnormal androgen signaling plays a key role in the growth and progression of prostate cancer [158]. A model to explain the origin of androgen-resistant neuroendocrine cells originating from a small stem cell niche has been proposed. The main strategy is androgen-protective therapy, which directs stem cells to differentiate into androgen-independent neuroendocrine cells [159]. Pax6 acts as a prostate cancer repressor by inhibiting androgen action. As demonstrated by immunohistochemical data [160], Pax6 expression is higher in the normal epithelial cells adjacent to the cancer cells than in cancer cells. Pax6 expression was different in three different prostate cancer lines and the lowest in the androgen-sensitive cell line LNCaP. The authors of the work demonstrated the suppressive function of Pax6 by silencing the transcriptional activity of androgen receptors through its direct interaction with these receptors.
Subsequent studies have demonstrated the coactivating function of the transcriptional coregulator SPBP in relation to androgen receptors. SPBP stimulates transcription of the probasin promoter and expression of the androgen receptor target gene PSA in LNCaP cells. SPBP is expressed in most cell lines but its highest expression is found in the androgen-dependent cancer cell line LNCaP. Pax6 inhibits SPBP-mediated stimulation of androgen receptor activity and the association of SPBP with the androgen receptor promoter had been established [161].
In addition to its suppressive role in the development of prostate cancer, Pax6 has been implicated in the transformation of the tumor from an androgen-dependent phenotype to an aggressive, resistant form known as neuroendocrine prostate cancer (NEPC) [162]. NEPC is characterized by loss of androgen receptor expression, elevated neuronal markers such as synaptophysin (SYP), chromogranin A (CGHA), neuron specific enolase (NSE). NEPC is highly aggressive and hasn’t effective therapeutic treatments [163,164,165]. NEPC differentiation reflects a cell lineage transition to neuronal phenotypes that mimics the process of neuronal differentiation during embryogenesis. Given its role in neurogenesis, Pax6 is considered to be one of the molecular switches in signal transduction that reprograms chromatin accessibility via the MET/STAT5a pathway, thereby enhancing lineage plasticity. STAT5a inhibits the expression of methyltransferases KMT5c and SMYD5, which downregulate the DNA regulatory region of histone 4 (H4K20me3-trimethylation of lysine 20 of histone 4), H4K20me3 is involved in maintaining genomic stability and regulating gene expression. H4K20me3 downregulation alters the cell line phenotype and confers a transcriptional profile of neuroendocrine cancer [162].
Breast tumor. The oncogenic role of the Pax6 gene has been established in breast cancer. The specific micro RNAs in breast cancer can inhibit cell proliferation and invasion, which ultimately suppresses its overexpression in tumor tissues [166,167]. Increased Pax6 expression is associated with unfavorable prognosis in patients with breast cancer [168], and Pax6 methylation is linked to cell metastasis [169]. A study [166] demonstrated Pax6 overexpression stimulates breast cancer cell migration and metastasis. Pax6 activates the TGF-β signaling pathway and downstream signaling through SMAD by mediating epithelial-mesenchymal transition transcription factors in the nucleus. TGF-β initiates the interaction of TGF-β cytokine with TGF-βR II receptors which recruits TGF-β I for downstream signaling through SMAD [170,171]. MicroRNA-135b has been shown to inhibit TGF-β signaling [172]. Studies have demonstrated activation of the TGF-β/SMAD signaling pathway in breast tumor metastasis [173,174,175]. Epithelial-mesenchymal transformation plays an important role in breast cancer metastasis. Overexpression of Pax6 in MCF7 and MDA-MB-231 breast cancer cell lines resulted in decreased expression of E-cadherin while mesenchymal markers N-cadherin, vimentin and fibronectin were significantly increased [166].
Liver cancer. Pax6 acts as a tumor suppressor in the development of hepatocellular carcinoma. Overexpression of the Pax6 protein suppressed independent growth and invasion of hepatocellular carcinoma cells due to upregulation of E-cadherin and suppression of thrombospodlin-1 was shown in a study [176]. Sequencing of liver cancer cells was performed and it was found that the Pax6 gene promoter is highly methylated [177]. The regulatory role of Pax6 in liver carcinoma cell lines through natural killer cells has been proven. Overexpression of Pax6 inhibits the growth of hepatocellular carcinoma cells by suppressing the expression of metalloproteinase and reducing the secretion levels of the oncoproteins sMICA and sULBP2 [178].
Pancreatic cancer. Overexpression of the MET tyrosine kinase receptor and the Pax6 transcription factor has been established in pancreatic cancer. The Pax6(5a) isoform is more highly expressed in pancreatic carcinoma cells than the canonical Pax6 protein. Both of these protein forms bind to an enhancer element in the MET promoter and activate expression of the MET gene [179]. The MET gene produces the c-MET protein, which is a receptor tyrosine kinase present on the surface of pancreatic cells. The ligand of this protein is the hepatocyte growth factor (HGF) produced by pancreatic stellate cells [180]. The HGF/c-MET interaction triggers uncontrolled cell proliferation, evasion of apoptosis and increased tumor invasion via the MAP/ERK signaling pathway [181]. In addition, the phosphorylated region of c-MET is a docking site for the p85 subunit of phosphatylinositol 3-kinase (PI3K). This binding subsequently leads to a series of biochemical reactions that inhibit tumor suppressors and increase tumor cell survival through a signaling pathway [182].
Lung cancer. Pax6 acts as an oncogene, promoting tumor growth and the formation of its chemoresistance in lung cancer. Pax6 promotes cell proliferation and cell cycle progression in the development of small cell lung cancer. Pax6 expression increases tumor cell resistance to apoptosis induced by cisplatin and etoposide. Pax6 in small cell cancer tumor lines suppresses the activation of the NOTCH signaling pathway enhancing Nanog expression thereby triggering cell proliferation, protection from apoptosis and resistance to treatment. Suppression of Pax6 expression increases the sensitivity of cells to chemotherapy [183]. In small cell lung cancer the crucial role of Pax6 in regulating the properties of stromal stem cells has been proven. The NOTCH-Nanog signaling pathway is involved in the reprogramming of stromal stem cells into tumor cells in small cell lung cancer [183]. Pax6 has the ability to bind to the promoter region of the pluripotent genes Oct4 and Nanog, which leads to the suppression of the expression of pluripotent genes and the transition of cells from the embryonic stem state [184].
A high degree of expression of Pax6 in tumor cells and a significant correlation with a decrease in the overall life expectancy of patients were demonstrated in the study of non-small cell lung cancer. Pax6 promotes cell migration and invasion as well as the development of epithelial-mesenchymal transition in non-small cell lung cancer. Pax6 regulates the transcriptional activity of the zinc finger of E-box-binding protein 2 (ZEB2) and binds to its promoter region. PAX6-ZEB2 signaling pathway promotes metastasis by mediating E-cadherin downregulation via the PI3K/AKT pathway in non-small cell lung cancer. Collectively, researches results suggest that PAX6 may be a novel therapeutic target to overcome metastasis and cisplatin resistance in non-small cell lung cancer [185]. Other studies have shown that Pax6 activates the tumor suppressor PTEN as well as SFRP2, which is an inhibitor of the WNT pathway. Pax6 inhibits the WNT pathway through regulation of its suppressor SFRP2 and suppression of its effector WNTB2 [186].
Another study established the proliferative role of Pax6 in the development of non-small cell lung cancer. Pax6 activates the transition of cells in the G-S phases of the life cycle by triggering the MAPK signaling pathway. Pax6 mRNA was expressed more often in lung cancer tissue compared to surrounding tissues. Regulation in cell cycle phases is associated with the fact that Pax6 enhances cyclin D expression and cell growth, thus promoting the transition from the G1 to S phase. The authors of the work indicate that Pax6 regulates the cell cycle through mitogen-induced signaling through phosphorylation of ERK1/2 and p38 proteins [187].
Retinoblastoma. Retinoblastoma is one of the most common tumors in newborns babies. Researchers attribute the leading role in the development of retinoblastoma to the inactivation of the Rb-1 suppressor gene [188]. Pax6 plays a dual role and can act as both a promoter and a tumor suppressor in the development of retinoblastoma. There is evidence that overexpression of Pax6 regulates the growth and apoptosis of human retinoblastoma cells [189,190]. Overexpression of Pax6 mediated by a lentiviral vector in human retinoblastoma cells is associated with increased cell proliferation against the background of a decrease in the rate of caspase-3-dependent apoptosis, as well as with a change in the cell cycle regulated by p53 [190]. It was found that suppression of the Pax6 gene using microRNA leads to inhibition of retinoblastoma cell growth and to apoptotic death of tumor cells. These changes were accompanied by increased regulation of P21 and P27 proteins, suppression of cdc2 protein [189]. Inhibition of Pax6 in human retinoblastoma cell lines resulted in a low rate of apoptosis, as evidenced by a decrease in the levels of the pro-apoptotic proteins Bcl-2 and p21 [191]. There is evidence that Pax6 is a key regulator of retinal development and correlates with the development and progression of retinoblastoma [189]. Pax6 expression was shown to be regulated by micro RNA-365b-3p (miR-365b-3p) [192]. Reduced expression of miR-365b-3p was found in retinoblastoma tissues. However, miR-365b-3p directly inhibits Pax6 expression and acts as a tumor suppressor, causing cell cycle arrest and cell apoptosis.
Thus, the Pax6 gene can act as either a tumor suppressor or an oncogene, depending on the tissue and the molecular signals from the extracellular microenvironment.
6. Conclusions
The current level of scientific work demonstrates a whole spectrum of functions and signal regulatory pathways in which the Pax6 gene and its protein are involved. However, despite the diverse findings, a consistent role of Pax6 in the control of the cell cycle can be traced across both embryonic and adult stages. Various signaling pathways involving Pax6, as reviewed here, ultimately converge to regulate cell cycle duration in certain cell types. This process is critical during morphogenesis and in the development of pathophysiological conditions. Based on the results of the analysis, we propose a generalized scheme describing the participation of Pax6 in implementation of various intracellular signaling pathways (Figure 1).
The most important signaling pathway involving Pax6 in the cell cycle of neuroepithelial cells at the early stages of embryogenesis is the Wnt/β-catenin pathway (Figure 1A). β-catenin, binding to the Pax6 promoter, activates its transcription and ultimately triggers the MAP-kinase signaling pathway, which through the activation of cyclin D regulates cell cycle proliferation and the transition of cells from the G1 to S phase. At the stage of late neurogenesis during neuronal differentiation Pax6 controls the properties of cell adhesion, neuronal polarity and migration through the δ-catenin signaling pathway. Regulation of Pax6 expression plays an important role in the formation of the spinal cord. Pax6 activation is important at the stage of somitogenesis. Later, when the presomitic mesoderm is formed, it inhibits Pax6 expression via the fibroblast growth factor (FGF8) signaling pathway.
The level of Pax6 protein is critical for the completion of proliferation and the exit of spinal cord neurons from the cell cycle to the differentiation stage. At this stage the level of neurogenin 2 (Ngn2) plays a decisive role suppressing Pax6 expression and thereby triggering the process of differentiation of spinal cord neurons. Pax6 plays an important role in glial differentiation during gliogenesis (Figure 1B). Pax6 carries out its regulatory role via glutamate signaling through the induction of expression of the glutamate transporter GLT1 on the plasma membrane of astrocytes.
Pax6 can perform both a suppressive and oncogenic function during tumor development. In glioblastomas Pax6 acts as a tumor growth suppressor (Figure 1C). Its expression is regulated by microRNA (miR)-355. An increase in the concentration of miR-355 causes an approximately 2-fold decrease in the Pax6 level as a result of which the cells actively proliferate and do not enter the differentiation pathway. In breast cancer, Pax6 acts as a promoter of the tumor process (Figure 1D). Activation of TGF-β/SMAD has been proven during breast tumor metastasis while overexpression of Pax-6 suppresses the expression of the cell adhesion protein E-cadherin that further enhances the invasive potential of the cells.
Pax6 plays a dual role acting as both a promoter and a tumor suppressor in the development of retinoblastoma (Figure 1E). The promoter function of Pax6 is associated with the inhibition of apoptosis by reducing the levels of caspase-3, Bcl-2 and Bax proteins. Pax6 is associated with cell proliferation and the cell cycle through the cdc25A/CDK2 and p53/p21/CDK1 pathways. The suppressor function of Pax6 in the development of breast tumors is associated with the inhibitory effect of microRNA mir-365b-3p on Pax6, which causes cell cycle arrest and apoptosis of tumor cells.
Pax6 plays a certain regulatory role in the development of neurodegenerative processes. In Parkinson’s disease, Pax6 improves cell survival in the dopamine formation by inhibiting oxidative stress and reducing neuronal apoptosis (Figure 1F). Pax6 acts as an important molecular mediator between amyloid-β hyperphosphorylation and tau protein in the development of Alzheimer’s disease (Figure 1G). At the same time, the main signaling pathways that are launched in this pathological process enhance the expression of Pax6. The main signaling pathway that is involved in the development of Alzheimer’s disease is E2F1, c-Myb, Pax6.
The molecular mechanisms through which Pax6 regulates cell cycle phases are diverse and redundant in both health and disease. This functional versatility depends on the organ and microenvironmental signals. However, a few studies have suggested that Pax6 may regulate levels of cyclin/Cdk complexes [193,194]. The effectiveness of molecular therapy at the level of the transcription factor Pax6 as a therapeutic target for various types of pathologies such as neurodegeneration and oncology is currently quite difficult to assess since everything depends on the context of the disease and multiple signaling pathways that can sometimes be realized in the same organ. In this regard, Pax6 can be a molecular switch of cell cycle phases towards promotion or suppression. It is possible that Pax6 serves as a therapeutic target in certain pathophysiological conditions. However, the effectiveness of such a strategy will depend on the substrate chain in the signaling pathway, its branching, and redundancy of mediators involved.
Author Contributions
Writing—original draft preparation, I.M., M.S. and T.K.; writing—review and editing, I.M., M.S. and T.K.; editing the final version of the manuscript, T.K., M.S., I.M., I.P. and N.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by governmental assignment of the Ministry of Health of Russian Federation No. 056-03-2025-071.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AKT | Protein Kinase B |
| APP | Amyloid Precursor Protein |
| BDNF | Brain-Derived Neurotrophic Factor |
| BLBP | Brain Lipid-Binding Protein |
| cAMP | Cyclic Adenosine Monophosphate |
| CD1 | Cluster of Differentiation 1 |
| cdc2 | Cell Division Cycle 2 |
| Cdh7 | Cadherin 7 |
| CDK | Cyclin-Dependent Kinase |
| CGHA | Chromogranin A |
| CMYD5 | SET and MYND Domain-Containing Protein 5 |
| CNS | Central Nervous System |
| DKK | Dickkopf-related protein |
| E2F1 | E2F Transcription Factor 1 |
| EAAT2 | Excitatory Amino Acid Transporter 2 |
| ERK | Extracellular Signal-Regulated Kinase |
| FABP7 | Fatty Acid-Binding Protein 7 |
| FGF | Fibroblast Growth Factor |
| GFAP | Glial Fibrillary Acidic Protein |
| GLT1 | Glutamate Transporter 1 |
| GSK3B | Glycogen Synthase Kinase 3 Beta |
| GTPase | Guanosine Triphosphatase |
| HGF | Hepatocyte Growth Factor |
| Iba1 | Ionized Calcium-Binding Adapter Molecule 1 |
| IFN | Interferon |
| KMT5c | Lysine Methyltransferase 5C |
| MET | Mesenchymal-Epithelial Transition Factor |
| Mitf | Microphthalmia-associated transcription factor |
| MMP | Matrix Metalloproteinase |
| c-Myb | Cellular Myb |
| NEPC | Neuroendocrine Prostate Cancer |
| Ngn2 | Neurogenin 2 |
| NSE | Neuron-Specific Enolase |
| Oct4 | Octamer-Binding Transcription Factor 4 |
| PC | Proprotein Convertase |
| PD | Paired Domain |
| PI3K | Phosphatidylinositol 3-Kinase |
| PPAR | Peroxisome Proliferator-Activated Receptor |
| SFRP | Secreted Frizzled-Related Protein |
| sMICA | Soluble MHC Class I Chain-Related Protein A |
| SPBP | Stromelysin-1 PDGF-responsive element Binding Protein |
| STAT | Signal Transducer and Activator of Transcription |
| sULBP2 | Soluble UL16-Binding Protein 2 |
| SYP | Synaptophysin |
| Tfec | Transcription Factor EC |
| TGF | Transforming Growth Factor |
| Wnt | Wingless-related integration site |
| WNTB2 | Wnt Family Member B2 |
| ZEB2 | Zinc Finger E-Box Binding Homeobox 2 |
References
- Cvekl, A.; Callaerts, P. PAX6: 25th Anniversary and More to Learn. Exp. Eye Res. 2017, 156, 10–21. [Google Scholar] [CrossRef]
- Kozmik, Z. Pax Genes in Eye Development and Evolution. Curr. Opin. Genet. Dev. 2005, 15, 430–438. [Google Scholar] [CrossRef]
- Hanson, I.; Van Heyningen, V. Pax6: More than Meets the Eye. Trends Genet. 1995, 11, 268–272. [Google Scholar] [CrossRef]
- Osumi, N. The Role of Pax6 in Brain Patterning. Tohoku J. Exp. Med. 2001, 193, 163–174. [Google Scholar] [CrossRef]
- Kioussi, C.; O’Connell, S.; St-Onge, L.; Treier, M.; Gleiberman, A.S.; Gruss, P.; Rosenfeld, M.G. Pax6 Is Essential for Establishing Ventral-Dorsal Cell Boundaries in Pituitary Gland Development. Proc. Natl. Acad. Sci. USA 1999, 96, 14378–14382. [Google Scholar] [CrossRef]
- Dohrmann, C.; Gruss, P.; Lemaire, L. Pax Genes and the Differentiation of Hormone-Producing Endocrine Cells in the Pancreas. Mech. Dev. 2000, 92, 47–54. [Google Scholar] [CrossRef]
- Manuel, M.N.; Mi, D.; Mason, J.O.; Price, D.J. Regulation of Cerebral Cortical Neurogenesis by the Pax6 Transcription Factor. Front. Cell. Neurosci. 2015, 9, 70. [Google Scholar] [CrossRef] [PubMed]
- Ypsilanti, A.R.; Rubenstein, J.L.R. Transcriptional and Epigenetic Mechanisms of Early Cortical Development: An Examination of How Pax6 Coordinates Cortical Development. J. Comp. Neurol. 2016, 524, 609–629. [Google Scholar] [CrossRef] [PubMed]
- Stoykova, A.; Götz, M.; Gruss, P.; Price, J. Pax6- Dependent Regulation of Adhesive Patterning, R-Cadherin Expression and Boundary Formation in Developing Forebrain. Development 1997, 124, 3765–3777. [Google Scholar] [CrossRef] [PubMed]
- Ryan, B.C.; Lowe, K.; Hanson, L.; Gil, T.; Braun, L.; Howard, P.L.; Chow, R.L. Mapping the Pax6 3’ Untranslated Region microRNA Regulatory Landscape. BMC Genom. 2018, 19, 820. [Google Scholar] [CrossRef]
- Abbasi, M.; Amini, M.; Moustardas, P.; Gutsmiedl, Q.; Javidjam, D.; Suiwal, S.; Seitz, B.; Fries, F.N.; Dashti, A.; Rautavaara, Y.; et al. Effects of miR-204-5p Modulation on PAX6 Regulation and Corneal Inflammation. Sci. Rep. 2024, 14, 26436. [Google Scholar] [CrossRef]
- Huang, J.; Rajagopal, R.; Liu, Y.; Dattilo, L.K.; Shaham, O.; Ashery-Padan, R.; Beebe, D.C. The Mechanism of Lens Placode Formation: A Case of Matrix-Mediated Morphogenesis. Dev. Biol. 2011, 355, 32–42. [Google Scholar] [CrossRef]
- Shaham, O.; Gueta, K.; Mor, E.; Oren-Giladi, P.; Grinberg, D.; Xie, Q.; Cvekl, A.; Shomron, N.; Davis, N.; Keydar-Prizant, M.; et al. Pax6 Regulates Gene Expression in the Vertebrate Lens through miR-204. PLoS Genet. 2013, 9, e1003357. [Google Scholar] [CrossRef] [PubMed]
- Walcher, T.; Xie, Q.; Sun, J.; Irmler, M.; Beckers, J.; Öztürk, T.; Niessing, D.; Stoykova, A.; Cvekl, A.; Ninkovic, J.; et al. Functional Dissection of the Paired Domain of Pax6 Reveals Molecular Mechanisms of Coordinating Neurogenesis and Proliferation. Development 2013, 140, 1123–1136. [Google Scholar] [CrossRef] [PubMed]
- Cvekl, A.; Yang, Y.; Chauhan, B.K.; Cveklova, K. Regulation of Gene Expression by Pax6 in Ocular Cells: A Case of Tissue-Preferred Expression of Crystallins in Lens. Int. J. Dev. Biol. 2004, 48, 829–844. [Google Scholar] [CrossRef] [PubMed]
- Czerny, T.; Busslinger, M. DNA-Binding and Transactivation Properties of Pax-6: Three Amino Acids in the Paired Domain Are Responsible for the Different Sequence Recognition of Pax-6 and BSAP (Pax-5). Mol. Cell. Biol. 1995, 15, 2858–2871. [Google Scholar] [CrossRef]
- Epstein, J.A.; Glaser, T.; Cai, J.; Jepeal, L.; Walton, D.S.; Maas, R.L. Two Independent and Interactive DNA-Binding Subdomains of the Pax6 Paired Domain Are Regulated by Alternative Splicing. Genes Dev. 1994, 8, 2022–2034. [Google Scholar] [CrossRef]
- He, S.; Pirity, M.K.; Wang, W.-L.; Wolf, L.; Chauhan, B.K.; Cveklova, K.; Tamm, E.R.; Ashery-Padan, R.; Metzger, D.; Nakai, A.; et al. Chromatin Remodeling Enzyme Brg1 Is Required for Mouse Lens Fiber Cell Terminal Differentiation and Its Denucleation. Epigenetics Chromatin 2010, 3, 21. [Google Scholar] [CrossRef]
- Yang, Y.; Stopka, T.; Golestaneh, N.; Wang, Y.; Wu, K.; Li, A.; Chauhan, B.K.; Gao, C.Y.; Cveklová, K.; Duncan, M.K.; et al. Regulation of αA-Crystallin via Pax6, c-Maf, CREB and a Broad Domain of Lens-Specific Chromatin. EMBO J. 2006, 25, 2107–2118. [Google Scholar] [CrossRef]
- Tian, T.; Quintana-Urzainqui, I.; Kozić, Z.; Pratt, T.; Price, D.J. Pax6 Loss Alters the Morphological and Electrophysiological Development of Mouse Prethalamic Neurons. Development 2022, 149, dev200052. [Google Scholar] [CrossRef]
- Ton, C.C.; Hirvonen, H.; Miwa, H.; Weil, M.M.; Monaghan, P.; Jordan, T.; van Heyningen, V.; Hastie, N.D.; Meijers-Heijboer, H.; Drechsler, M. Positional Cloning and Characterization of a Paired Box- and Homeobox-Containing Gene from the Aniridia Region. Cell 1991, 67, 1059–1074. [Google Scholar] [CrossRef]
- Xu, Z.P.; Saunders, G.F. Transcriptional Regulation of the Human PAX6 Gene Promoter. J. Biol. Chem. 1997, 272, 3430–3436. [Google Scholar] [CrossRef]
- Vasilyeva, T.A.; Voskresenskaya, A.A.; Pozdeyeva, N.A.; Marakhonov, A.V.; Zinchenko, R.A. PAX6 Gene Characteristic and Causative Role of PAX6 Mutations in Inherited Eye Pathologies. Russ. J. Genet. 2018, 54, 995–1002. [Google Scholar] [CrossRef]
- Onuma, Y.; Takahashi, S.; Asashima, M.; Kurata, S.; Gehring, W.J. Conservation of Pax6 Function and Upstream Activation by Notch Signaling in Eye Development of Frogs and Flies. Proc. Natl. Acad. Sci. USA 2002, 99, 2020–2025. [Google Scholar] [CrossRef]
- Halder, G.; Callaerts, P.; Gehring, W.J. Induction of Ectopic Eyes by Targeted Expression of the Eyeless Gene in Drosophila. Science 1995, 267, 1788–1792. [Google Scholar] [CrossRef]
- Xu, H.E.; Rould, M.A.; Xu, W.; Epstein, J.A.; Maas, R.L.; Pabo, C.O. Crystal Structure of the Human Pax6 Paired Domain-DNA Complex Reveals Specific Roles for the Linker Region and Carboxy-Terminal Subdomain in DNA Binding. Genes. Dev. 1999, 13, 1263–1275. [Google Scholar] [CrossRef] [PubMed]
- Haubst, N.; Berger, J.; Radjendirane, V.; Graw, J.; Favor, J.; Saunders, G.F.; Stoykova, A.; Götz, M. Molecular Dissection of Pax6 Function: The Specific Roles of the Paired Domain and Homeodomain in Brain Development. Development 2004, 131, 6131–6140. [Google Scholar] [CrossRef] [PubMed]
- Davydenko, K.; Filatova, A.; Skoblov, M. Assessing Splicing Variants in the PAX6 Gene: A Comprehensive Minigene Approach. J. Cell. Mol. Med. 2025, 29, e70459. [Google Scholar] [CrossRef] [PubMed]
- Gosmain, Y.; Marthinet, E.; Cheyssac, C.; Guérardel, A.; Mamin, A.; Katz, L.S.; Bouzakri, K.; Philippe, J. Pax6 Controls the Expression of Critical Genes Involved in Pancreatic α Cell Differentiation and Function. J. Biol. Chem. 2010, 285, 33381–33393. [Google Scholar] [CrossRef]
- Amini, M.; Stachon, T.; Hsu, S.-L.; Li, Z.; Chai, N.; Fries, F.N.; Seitz, B.; Kundu, S.; Suiwal, S.; Szentmáry, N. Effect of MiRNA 204-5P Mimics and Lipopolysaccharide-Induced Inflammation on Transcription Factor Levels, Cell Maintenance, and Retinoic Acid Signaling in Primary Limbal Epithelial Cells. Int. J. Mol. Sci. 2025, 26, 3809. [Google Scholar] [CrossRef]
- Epstein, J.; Cai, J.; Glaser, T.; Jepeal, L.; Maas, R. Identification of a Pax Paired Domain Recognition Sequence and Evidence for DNA-Dependent Conformational Changes. J. Biol. Chem. 1994, 269, 8355–8361. [Google Scholar] [CrossRef]
- Jun, S.; Desplan, C. Cooperative Interactions between Paired Domain and Homeodomain. Development 1996, 122, 2639–2650. [Google Scholar] [CrossRef]
- Tang, H.K.; Singh, S.; Saunders, G.F. Dissection of the Transactivation Function of the Transcription Factor Encoded by the Eye Developmental Gene PAX6. J. Biol. Chem. 1998, 273, 7210–7221. [Google Scholar] [CrossRef] [PubMed]
- Pinson, J.; Mason, J.O.; Simpson, T.I.; Price, D.J. Regulation of the Pax6: Pax6(5a) mRNA Ratio in the Developing Mammalian Brain. BMC Dev. Biol. 2005, 5, 13. [Google Scholar] [CrossRef]
- Kammandel, B.; Chowdhury, K.; Stoykova, A.; Aparicio, S.; Brenner, S.; Gruss, P. Distinct Cis-Essential Modules Direct the Time-Space Pattern of the Pax6 Gene Activity. Dev. Biol. 1999, 205, 79–97. [Google Scholar] [CrossRef]
- Kim, J.; Lauderdale, J.D. Analysis of Pax6 Expression Using a BAC Transgene Reveals the Presence of a Paired-Less Isoform of Pax6 in the Eye and Olfactory Bulb. Dev. Biol. 2006, 292, 486–505. [Google Scholar] [CrossRef]
- Kim, J.; Lauderdale, J.D. Overexpression of Pairedless Pax6 in the Retina Disrupts Corneal Development and Affects Lens Cell Survival. Dev. Biol. 2008, 313, 434–454. [Google Scholar] [CrossRef] [PubMed]
- Kozmik, Z.; Kozmikova, I. Ancestral Role of Pax6 in Chordate Brain Regionalization. Front. Cell Dev. Biol. 2024, 12, 1431337. [Google Scholar] [CrossRef] [PubMed]
- Aota, S.; Nakajima, N.; Sakamoto, R.; Watanabe, S.; Ibaraki, N.; Okazaki, K. Pax6 Autoregulation Mediated by Direct Interaction of Pax6 Protein with the Head Surface Ectoderm-Specific Enhancer of the Mouse Pax6 Gene. Dev. Biol. 2003, 257, 1–13. [Google Scholar] [CrossRef]
- Manuel, M.; Georgala, P.A.; Carr, C.B.; Chanas, S.; Kleinjan, D.A.; Martynoga, B.; Mason, J.O.; Molinek, M.; Pinson, J.; Pratt, T.; et al. Controlled Overexpression of Pax6 in Vivo Negatively Autoregulates the Pax6 Locus, Causing Cell-Autonomous Defects of Late Cortical Progenitor Proliferation with Little Effect on Cortical Arealization. Development 2007, 134, 545–555. [Google Scholar] [CrossRef]
- Duparc, R.-H.; Abdouh, M.; David, J.; Lépine, M.; Tétreault, N.; Bernier, G. Pax6 Controls the Proliferation Rate of Neuroepithelial Progenitors from the Mouse Optic Vesicle. Dev. Biol. 2007, 301, 374–387. [Google Scholar] [CrossRef]
- Thakurela, S.; Tiwari, N.; Schick, S.; Garding, A.; Ivanek, R.; Berninger, B.; Tiwari, V.K. Mapping Gene Regulatory Circuitry of Pax6 during Neurogenesis. Cell Discov. 2016, 2, 15045. [Google Scholar] [CrossRef] [PubMed]
- Buckle, A.; Nozawa, R.; Kleinjan, D.A.; Gilbert, N. Functional Characteristics of Novel Pancreatic Pax6 Regulatory Elements. Hum. Mol. Genet. 2018, 27, 3434–3448. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Rockowitz, S.; Xie, Q.; Ashery-Padan, R.; Zheng, D.; Cvekl, A. Identification of in Vivo DNA-Binding Mechanisms of Pax6 and Reconstruction of Pax6-Dependent Gene Regulatory Networks during Forebrain and Lens Development. Nucleic Acids Res. 2015, 43, 6827–6846. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, T.; Ashery-Padan, R.; Andrejewski, N.; Scardigli, R.; Guillemot, F.; Gruss, P. Pax6 Is Required for the Multipotent State of Retinal Progenitor Cells. Cell 2001, 105, 43–55. [Google Scholar] [CrossRef]
- Bharti, K.; Gasper, M.; Ou, J.; Brucato, M.; Clore-Gronenborn, K.; Pickel, J.; Arnheiter, H. A Regulatory Loop Involving PAX6, MITF, and WNT Signaling Controls Retinal Pigment Epithelium Development. PLoS Genet. 2012, 8, e1002757. [Google Scholar] [CrossRef]
- Davis, N.; Yoffe, C.; Raviv, S.; Antes, R.; Berger, J.; Holzmann, S.; Stoykova, A.; Overbeek, P.A.; Tamm, E.R.; Ashery-Padan, R. Pax6 Dosage Requirements in Iris and Ciliary Body Differentiation. Dev. Biol. 2009, 333, 132–142. [Google Scholar] [CrossRef]
- Kohwi, M.; Osumi, N.; Rubenstein, J.L.R.; Alvarez-Buylla, A. Pax6 Is Required for Making Specific Subpopulations of Granule and Periglomerular Neurons in the Olfactory Bulb. J. Neurosci. 2005, 25, 6997–7003. [Google Scholar] [CrossRef]
- Warren, N.; Caric, D.; Pratt, T.; Clausen, J.A.; Asavaritikrai, P.; Mason, J.O.; Hill, R.E.; Price, D.J. The Transcription Factor, Pax6, Is Required for Cell Proliferation and Differentiation in the Developing Cerebral Cortex. Cereb. Cortex 1999, 9, 627–635. [Google Scholar] [CrossRef]
- Georgala, P.A.; Carr, C.B.; Price, D.J. The Role of Pax6 in Forebrain Development. Dev. Neurobiol. 2011, 71, 690–709. [Google Scholar] [CrossRef]
- Hsieh, Y.-W.; Yang, X.-J. Dynamic Pax6 Expression during the Neurogenic Cell Cycle Influences Proliferation and Cell Fate Choices of Retinal Progenitors. Neural Dev. 2009, 4, 32. [Google Scholar] [CrossRef]
- Zhang, B.; Hou, M.; Huang, J.; Liu, Y.; Yang, C.; Lin, J. Pax6 Regulates Neuronal Migration and Cell Proliferation via Interacting with Wnt3a during Cortical Development. Sci. Rep. 2025, 15, 4726. [Google Scholar] [CrossRef]
- Ritz-Laser, B.; Estreicher, A.; Gauthier, B.; Philippe, J. The Paired Homeodomain Transcription Factor Pax-2 Is Expressed in the Endocrine Pancreas and Transactivates the Glucagon Gene Promoter. J. Biol. Chem. 2000, 275, 32708–32715. [Google Scholar] [CrossRef]
- Osumi, N.; Shinohara, H.; Numayama-Tsuruta, K.; Maekawa, M. Concise Review: Pax6 Transcription Factor Contributes to Both Embryonic and Adult Neurogenesis as a Multifunctional Regulator. Stem Cells 2008, 26, 1663–1672. [Google Scholar] [CrossRef]
- Ninkovic, J.; Pinto, L.; Petricca, S.; Lepier, A.; Sun, J.; Rieger, M.A.; Schroeder, T.; Cvekl, A.; Favor, J.; Götz, M. The Transcription Factor Pax6 Regulates Survival of Dopaminergic Olfactory Bulb Neurons via Crystallin αA. Neuron 2010, 68, 682–694. [Google Scholar] [CrossRef]
- Brill, M.S.; Ninkovic, J.; Winpenny, E.; Hodge, R.D.; Ozen, I.; Yang, R.; Lepier, A.; Gascón, S.; Erdelyi, F.; Szabo, G.; et al. Adult Generation of Glutamatergic Olfactory Bulb Interneurons. Nat. Neurosci. 2009, 12, 1524–1533. [Google Scholar] [CrossRef]
- Hack, M.A.; Saghatelyan, A.; De Chevigny, A.; Pfeifer, A.; Ashery-Padan, R.; Lledo, P.-M.; Götz, M. Neuronal Fate Determinants of Adult Olfactory Bulb Neurogenesis. Nat. Neurosci. 2005, 8, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, M.; Haynes, T.L.; Frisch, N.C.; Call, M.K.; Minich, C.M.; Tsonis, P.A.; Del Rio-Tsonis, K. The Role of Pax-6 in Lens Regeneration. Proc. Natl. Acad. Sci. USA 2006, 103, 14848–14853. [Google Scholar] [CrossRef]
- Hart, A.W.; Mella, S.; Mendrychowski, J.; Van Heyningen, V.; Kleinjan, D.A. The Developmental Regulator Pax6 Is Essential for Maintenance of Islet Cell Function in the Adult Mouse Pancreas. PLoS ONE 2013, 8, e54173. [Google Scholar] [CrossRef] [PubMed]
- Manuel, M.; Price, D.J. Role of Pax6 in Forebrain Regionalization. Brain Res. Bull. 2005, 66, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Osumi-Yamashita, N.; Noji, S.; Ohuchi, H.; Koyama, E.; Myokai, F.; Matsuo, N.; Taniguchi, S.; Doi, H.; Iseki, S.; et al. A Mutation in the Pax-6 Gene in Rat Small Eye Is Associated with Impaired Migration of Midbrain Crest Cells. Nat. Genet. 1993, 3, 299–304. [Google Scholar] [CrossRef]
- Inoue, T.; Nakamura, S.; Osumi, N. Fate Mapping of the Mouse Prosencephalic Neural Plate. Dev. Biol. 2000, 219, 373–383. [Google Scholar] [CrossRef]
- Soriano, E.; Del Río, J.A. The Cells of Cajal-Retzius: Still a Mystery One Century After. Neuron 2005, 46, 389–394. [Google Scholar] [CrossRef]
- Marín, O.; Rubenstein, J.L. A Long, Remarkable Journey: Tangential Migration in the Telencephalon. Nat. Rev. Neurosci. 2001, 2, 780–790. [Google Scholar] [CrossRef]
- Flames, N.; Marín, O. Developmental Mechanisms Underlying the Generation of Cortical Interneuron Diversity. Neuron 2005, 46, 377–381. [Google Scholar] [CrossRef]
- Hirata, T.; Nomura, T.; Takagi, Y.; Sato, Y.; Tomioka, N.; Fujisawa, H.; Osumi, N. Mosaic Development of the Olfactory Cortex with Pax6-Dependent and -Independent Components. Dev. Brain Res. 2002, 136, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Gómez-López, S.; Wiskow, O.; Favaro, R.; Nicolis, S.K.; Price, D.J.; Pollard, S.M.; Smith, A. Sox2 and Pax6 Maintain the Proliferative and Developmental Potential of Gliogenic Neural Stem Cells In Vitro. Glia 2011, 59, 1588–1599. [Google Scholar] [CrossRef] [PubMed]
- Schmahl, W.; Knoedlseder, M.; Favor, J.; Davidson, D. Defects of Neuronal Migration and the Pathogenesis of Cortical Malformations Are Associated with Small Eye (Sey) in the Mouse, a Point Mutation at the Pax-6-Locus. Acta Neuropathol. 1993, 86, 126–135. [Google Scholar] [CrossRef]
- Stoykova, A.; Fritsch, R.; Walther, C.; Gruss, P. Forebrain Patterning Defects in Small Eye Mutant Mice. Development 1996, 122, 3453–3465. [Google Scholar] [CrossRef] [PubMed]
- Carić, D.; Gooday, D.; Hill, R.E.; McConnell, S.K.; Price, D.J. Determination of the Migratory Capacity of Embryonic Cortical Cells Lacking the Transcription Factor Pax-6. Development 1997, 124, 5087–5096. [Google Scholar] [CrossRef]
- Fukuda, T.; Kawano, H.; Osumi, N.; Eto, K.; Kawamura, K. Histogenesis of the Cerebral Cortex in Rat Fetuses with a Mutation in the Pax-6 Gene. Dev. Brain Res. 2000, 120, 65–75. [Google Scholar] [CrossRef]
- Estivill-Torrus, G.; Pearson, H.; Van Heyningen, V.; Price, D.J.; Rashbass, P. Pax6 Is Required to Regulate the Cell Cycle and the Rate of Progression from Symmetrical to Asymmetrical Division in Mammalian Cortical Progenitors. Development 2002, 129, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Heins, N.; Malatesta, P.; Cecconi, F.; Nakafuku, M.; Tucker, K.L.; Hack, M.A.; Chapouton, P.; Barde, Y.-A.; Götz, M. Glial Cells Generate Neurons: The Role of the Transcription Factor Pax6. Nat. Neurosci. 2002, 5, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Yogarajah, M.; Matarin, M.; Vollmar, C.; Thompson, P.J.; Duncan, J.S.; Symms, M.; Moore, A.T.; Liu, J.; Thom, M.; Van Heyningen, V.; et al. PAX6, Brain Structure and Function in Human Adults: Advanced MRI in Aniridia. Ann. Clin. Transl. Neurol. 2016, 3, 314–330. [Google Scholar] [CrossRef]
- Xie, Q.; Yang, Y.; Huang, J.; Ninkovic, J.; Walcher, T.; Wolf, L.; Vitenzon, A.; Zheng, D.; Götz, M.; Beebe, D.C.; et al. Pax6 Interactions with Chromatin and Identification of Its Novel Direct Target Genes in Lens and Forebrain. PLoS ONE 2013, 8, e54507. [Google Scholar] [CrossRef]
- Kikkawa, T.; Obayashi, T.; Takahashi, M.; Fukuzaki-Dohi, U.; Numayama-Tsuruta, K.; Osumi, N. Dmrta1 Regulates Proneural Gene Expression Downstream of P Ax6 in the Mammalian Telencephalon. Genes. Cells 2013, 18, 636–649. [Google Scholar] [CrossRef]
- Machon, O.; Kreslova, J.; Ruzickova, J.; Vacik, T.; Klimova, L.; Fujimura, N.; Lachova, J.; Kozmik, Z. Lens Morphogenesis Is Dependent on Pax6-mediated Inhibition of the Canonical Wnt/Beta-catenin Signaling in the Lens Surface Ectoderm. Genesis 2010, 48, 86–95. [Google Scholar] [CrossRef]
- Numayama-Tsuruta, K.; Arai, Y.; Takahashi, M.; Sasaki-Hoshino, M.; Funatsu, N.; Nakamura, S.; Osumi, N. Downstream Genes of Pax6 Revealed by Comprehensive Transcriptome Profiling in the Developing Rat Hindbrain. BMC Dev. Biol. 2010, 10, 6. [Google Scholar] [CrossRef]
- Simpson, T.I.; Price, D.J. Pax6; A Pleiotropic Player in Development. BioEssays 2002, 24, 1041–1051. [Google Scholar] [CrossRef]
- Tyas, D.A.; Pearson, H.; Rashbass, P.; Price, D.J. Pax6 Regulates Cell Adhesion during Cortical Development. Cereb. Cortex 2003, 13, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Van Der Meer-de Jong, R.; Dickinson, M.E.; Woychik, R.P.; Stubbs, L.; Hetherington, C.; Hogan, B.L.M. Location of the Gene Involving the Small Eye Mutation on Mouse Chromosome 2 Suggests Homology with Human Aniridia 2 (AN2). Genomics 1990, 7, 270–275. [Google Scholar] [CrossRef]
- Abdolkarimi, D.; Cunha, D.L.; Lahne1, M.; Moosajee, M. PAX6 Disease Models for Aniridia. Indian J. Ophthalmol. 2022, 70, 4119–4129. [Google Scholar] [CrossRef]
- Philips, G.T.; Stair, C.N.; Young Lee, H.; Wroblewski, E.; Berberoglu, M.A.; Brown, N.L.; Mastick, G.S. Precocious Retinal Neurons: Pax6 Controls Timing of Differentiation and Determination of Cell Type. Dev. Biol. 2005, 279, 308–321. [Google Scholar] [CrossRef]
- Grant, M.K.; Bobilev, A.M.; Pierce, J.E.; DeWitte, J.; Lauderdale, J.D. Structural Brain Abnormalities in 12 Persons with Aniridia. F1000Research 2017, 6, 255. [Google Scholar] [CrossRef] [PubMed]
- Hanish, A.E.; Butman, J.A.; Thomas, F.; Yao, J.; Han, J.C. Pineal Hypoplasia, Reduced Melatonin and Sleep Disturbance in Patients with PAX6 Haploinsufficiency. J. Sleep Res. 2016, 25, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, T.N.; Free, S.L.; Williamson, K.A.; Stevens, J.M.; Churchill, A.J.; Hanson, I.M.; Shorvon, S.D.; Moore, A.T.; Van Heyningen, V.; Sisodiya, S.M. Polymicrogyria and Absence of Pineal Gland Due to PAX6 Mutation. Ann. Neurol. 2003, 53, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Gan, Q.; Lee, A.; Suzuki, R.; Yamagami, T.; Stokes, A.; Nguyen, B.C.; Pleasure, D.; Wang, J.; Chen, H.-W.; Zhou, C.J. Pax6 Mediates SS-Catenin Signaling for Self-Renewal and Neurogenesis by Neocortical Radial Glial Stem Cells. Stem Cells 2014, 32, 45–58. [Google Scholar] [CrossRef]
- Bertrand, N.; Médevielle, F.; Pituello, F. FGF Signalling Controls the Timing of Pax6 Activation in the Neural Tube. Development 2000, 127, 4837–4843. [Google Scholar] [CrossRef]
- Briscoe, J.; Pierani, A.; Jessell, T.M.; Ericson, J. A Homeodomain Protein Code Specifies Progenitor Cell Identity and Neuronal Fate in the Ventral Neural Tube. Cell 2000, 101, 435–445. [Google Scholar] [CrossRef]
- Lei, Q.; Jeong, Y.; Misra, K.; Li, S.; Zelman, A.K.; Epstein, D.J.; Matise, M.P. Wnt Signaling Inhibitors Regulate the Transcriptional Response to Morphogenetic Shh-Gli Signaling in the Neural Tube. Dev. Cell 2006, 11, 325–337. [Google Scholar] [CrossRef]
- Ericson, J.; Rashbass, P.; Schedl, A.; Brenner-Morton, S.; Kawakami, A.; Van Heyningen, V.; Jessell, T.M.; Briscoe, J. Pax6 Controls Progenitor Cell Identity and Neuronal Fate in Response to Graded Shh Signaling. Cell 1997, 90, 169–180. [Google Scholar] [CrossRef]
- Akai, J.; Halley, P.A.; Storey, K.G. FGF-Dependent Notch Signaling Maintains the Spinal Cord Stem Zone. Genes Dev. 2005, 19, 2877–2887. [Google Scholar] [CrossRef] [PubMed]
- Lobjois, V.; Benazeraf, B.; Bertrand, N.; Medevielle, F.; Pituello, F. Specific Regulation of Cyclins D1 and D2 by FGF and Shh Signaling Coordinates Cell Cycle Progression, Patterning, and Differentiation during Early Steps of Spinal Cord Development. Dev. Biol. 2004, 273, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Del Corral, R.D.; Olivera-Martinez, I.; Goriely, A.; Gale, E.; Maden, M.; Storey, K. Opposing FGF and Retinoid Pathways Control Ventral Neural Pattern, Neuronal Differentiation, and Segmentation during Body Axis Extension. Neuron 2003, 40, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Novitch, B.G.; Wichterle, H.; Jessell, T.M.; Sockanathan, S. A Requirement for Retinoic Acid-Mediated Transcriptional Activation in Ventral Neural Patterning and Motor Neuron Specification. Neuron 2003, 40, 81–95. [Google Scholar] [CrossRef]
- Bel-Vialar, S.; Medevielle, F.; Pituello, F. The on/off of Pax6 Controls the Tempo of Neuronal Differentiation in the Developing Spinal Cord. Dev. Biol. 2007, 305, 659–673. [Google Scholar] [CrossRef]
- Collinson, J.M.; Hill, R.E.; West, J.D. Different Roles for Pax6 in the Optic Vesicle and Facial Epithelium Mediate Early Morphogenesis of the Murine Eye. Development 2000, 127, 945–956. [Google Scholar] [CrossRef]
- Engelkamp, D.; Rashbass, P.; Seawright, A.; Heyningen, V.V. Role of Pax6 in Development of the Cerebellar System. Development 1999, 126, 3585–3596. [Google Scholar] [CrossRef]
- Yamasaki, T.; Kawaji, K.; Ono, K.; Bito, H.; Hirano, T.; Osumi, N.; Kengaku, M. Pax6 Regulates Granule Cell Polarization during Parallel Fiber Formation in the Developing Cerebellum. Development 2001, 128, 3133–3144. [Google Scholar] [CrossRef]
- Andrews, G.L.; Mastick, G.S. R-Cadherin Is a Pax6-Regulated, Growth-Promoting Cue for Pioneer Axons. J. Neurosci. 2003, 23, 9873–9880. [Google Scholar] [CrossRef]
- Duparc, R.-H.; Boutemmine, D.; Champagne, M.-P.; Tétreault, N.; Bernier, G. Pax6 Is Required for Delta-Catenin/Neurojugin Expression during Retinal, Cerebellar and Cortical Development in Mice. Dev. Biol. 2006, 300, 647–655. [Google Scholar] [CrossRef]
- Martinez, M.C.; Ochiishi, T.; Majewski, M.; Kosik, K.S. Dual Regulation of Neuronal Morphogenesis by a δ-Catenin–Cortactin Complex and Rho. J. Cell Biol. 2003, 162, 99–111. [Google Scholar] [CrossRef]
- Davis, M.A.; Ireton, R.C.; Reynolds, A.B. A Core Function for P120-Catenin in Cadherin Turnover. J. Cell Biol. 2003, 163, 525–534. [Google Scholar] [CrossRef]
- Martinez-Lozada, Z.; Ortega, A. Milestone Review: Excitatory Amino Acid Transporters—Beyond Their Expected Function. J. Neurochem. 2023, 165, 457–466. [Google Scholar] [CrossRef]
- Martinez-Lozada, Z.; Guillem, A.M.; Song, I.; Gonzalez, M.V.; Takano, H.; Parikh, E.; Rothstein, J.D.; Putt, M.E.; Robinson, M.B. Identification of a Subpopulation of Astrocyte Progenitor Cells in the Neonatal Subventricular Zone: Evidence That Migration Is Regulated by Glutamate Signaling. Neurochem. Res. 2025, 50, 77. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Lane, M.; Krizman, E.; Sattler, R.; Rothstein, J.D.; Robinson, M.B. The Transcription Factor Pax6 Contributes to the Induction of GLT-1 Expression in Astrocytes through an Interaction with a Distal Enhancer Element. J. Neurochem. 2016, 136, 262–275. [Google Scholar] [CrossRef] [PubMed]
- Rothstein, J.D.; Dykes-Hoberg, M.; Pardo, C.A.; Bristol, L.A.; Jin, L.; Kuncl, R.W.; Kanai, Y.; Hediger, M.A.; Wang, Y.; Schielke, J.P.; et al. Knockout of Glutamate Transporters Reveals a Major Role for Astroglial Transport in Excitotoxicity and Clearance of Glutamate. Neuron 1996, 16, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Watase, K.; Manabe, T.; Yamada, K.; Watanabe, M.; Takahashi, K.; Iwama, H.; Nishikawa, T.; Ichihara, N.; Kikuchi, T.; et al. Epilepsy and Exacerbation of Brain Injury in Mice Lacking the Glutamate Transporter GLT1. Science 1997, 276, 1699–1702. [Google Scholar] [CrossRef]
- Gegelashvili, G.; Danbolt, N.C.; Schousboe, A. Neuronal Soluble Factors Differentially Regulate the Expression of the GLT1 and GLAST Glutamate Transporters in Cultured Astroglia. J. Neurochem. 1997, 69, 2612–2615. [Google Scholar] [CrossRef]
- Schlag, B.D.; Vondrasek, J.R.; Munir, M.; Kalandadze, A.; Zelenaia, O.A.; Rothstein, J.D.; Robinson, M.B. Regulation of the Glial Na+-Dependent Glutamate Transporters by Cyclic AMP Analogs and Neurons. Mol. Pharmacol. 1998, 53, 355–369. [Google Scholar] [CrossRef]
- Zelenaia, O.; Schlag, B.D.; Gochenauer, G.E.; Ganel, R.; Song, W.; Beesley, J.S.; Grinspan, J.B.; Rothstein, J.D.; Robinson, M.B. Epidermal Growth Factor Receptor Agonists Increase Expression of Glutamate Transporter GLT1 in Astrocytes through Pathways Dependent on Phosphatidylinositol 3-Kinase and Transcription Factor NF-κB. Mol. Pharmacol. 2000, 57, 667–678. [Google Scholar] [CrossRef]
- Sakurai, K.; Osumi, N. The Neurogenesis-Controlling Factor, Pax6, Inhibits Proliferation and Promotes Maturation in Murine Astrocytes. J. Neurosci. 2008, 28, 4604–4612. [Google Scholar] [CrossRef] [PubMed]
- Moosajee, M.; Hingorani, M.; Moore, A.T. PAX6-Related Aniridia. In GeneReviews®; Adam, M.P., Feldman, J., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Amemiya, A., Eds.; University of Washington, Seattle: Seattle, WA, USA, 1993. [Google Scholar]
- Tripathy, K.; Salini, B. Aniridia. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2025. [Google Scholar]
- Hingorani, M.; Williamson, K.A.; Moore, A.T.; Van Heyningen, V. Detailed Ophthalmologic Evaluation of 43 Individuals with PAX6 Mutations. Investig. Opthalmology Vis. Sci. 2009, 50, 2581. [Google Scholar] [CrossRef] [PubMed]
- Obst, J.; Fries, F.N.; Amini, M.; Náray, A.; Munteanu, C.; Stachon, T.; Suiwal, S.; Lagali, N.; Seitz, B.; Käsmann-Kellner, B.; et al. Systemic Diseases in Patients with Congenital Aniridia: A Report from the Homburg Registry for Congenital Aniridia. Ophthalmol. Ther. 2025, 14, 433–445. [Google Scholar] [CrossRef]
- Boese, E.A.; Tollefson, M.R.; Schnieders, M.J.; Darbro, B.W.; Alward, W.L.M.; Fingert, J.H. Novel Intragenic PAX6 Deletion in a Pedigree with Aniridia, Morbid Obesity, and Diabetes. Curr. Eye Res. 2020, 45, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, T.; Kajimoto, Y.; Fujitani, Y.; Watada, H.; Yamamoto, S.; Watarai, T.; Umayahara, Y.; Matsuhisa, M.; Gorogawa, S.; Kuwayama, Y.; et al. PAX6 Mutation as a Genetic Factor Common to Aniridia and Glucose Intolerance. Diabetes 2002, 51, 224–230. [Google Scholar] [CrossRef]
- Motoda, S.; Fujita, S.; Kozawa, J.; Kimura, T.; Fukui, K.; Ikuno, Y.; Imagawa, A.; Iwahashi, H.; Shimomura, I. Case of a Novel PAX6 Mutation with Aniridia and Insulin-dependent Diabetes Mellitus. J. Diabetes Investig. 2019, 10, 552–553. [Google Scholar] [CrossRef]
- Tian, W.; Zhu, X.; Qiao, C.; Ma, Y.; Yang, F.; Zhou, Z.; Feng, J.; Sun, R.; Xie, R.; Lu, J.; et al. Heterozygous PAX6 Mutations May Lead to Hyper-proinsulinaemia and Glucose Intolerance: A Case–Control Study in Families with Congenital Aniridia. Diabet. Med. 2021, 38, e14456. [Google Scholar] [CrossRef]
- Macdonald, G.C.; Hesselson, S.E.; Chan, J.Y.; Jenkins, A.B.; Laybutt, D.R.; Hesselson, D.; Campbell, L.V. Deletion Distal to the PAX6 Coding Region Reveals a Novel Basis for Familial Cosegregation of Aniridia and Diabetes Mellitus. Diabetes Res. Clin. Pract. 2019, 148, 64–71. [Google Scholar] [CrossRef]
- Wen, J.H.; Chen, Y.Y.; Song, S.J.; Ding, J.; Gao, Y.; Hu, Q.K.; Feng, R.P.; Liu, Y.Z.; Ren, G.C.; Zhang, C.Y.; et al. Paired Box 6 (PAX6) Regulates Glucose Metabolism via Proinsulin Processing Mediated by Prohormone Convertase 1/3 (PC1/3). Diabetologia 2009, 52, 504–513. [Google Scholar] [CrossRef]
- Cunha, D.L.; Kit, V.; Skinner, J.; Welch, A.A.; Moosajee, M. Metabolic and Neuroactivity Imbalances in Plasma from Aniridia Patients with PAX6 Haploinsufficiency. bioRxiv 2024, ahead of print. [Google Scholar]
- Jakaria, M.; Azam, S.; Haque, M.E.; Jo, S.-H.; Uddin, M.S.; Kim, I.-S.; Choi, D.-K. Taurine and Its Analogs in Neurological Disorders: Focus on Therapeutic Potential and Molecular Mechanisms. Redox Biol. 2019, 24, 101223. [Google Scholar] [CrossRef] [PubMed]
- Braverman, N.E.; Moser, A.B. Functions of Plasmalogen Lipids in Health and Disease. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2012, 1822, 1442–1452. [Google Scholar] [CrossRef]
- Lima Cunha, D.; Arno, G.; Corton, M.; Moosajee, M. The Spectrum of PAX6 Mutations and Genotype-Phenotype Correlations in the Eye. Genes 2019, 10, 1050. [Google Scholar] [CrossRef] [PubMed]
- Gall, W.E.; Beebe, K.; Lawton, K.A.; Adam, K.-P.; Mitchell, M.W.; Nakhle, P.J.; Ryals, J.A.; Milburn, M.V.; Nannipieri, M.; Camastra, S.; et al. α-Hydroxybutyrate Is an Early Biomarker of Insulin Resistance and Glucose Intolerance in a Nondiabetic Population. PLoS ONE 2010, 5, e10883. [Google Scholar] [CrossRef] [PubMed]
- Swisa, A.; Avrahami, D.; Eden, N.; Zhang, J.; Feleke, E.; Dahan, T.; Cohen-Tayar, Y.; Stolovich-Rain, M.; Kaestner, K.H.; Glaser, B.; et al. PAX6 Maintains β Cell Identity by Repressing Genes of Alternative Islet Cell Types. J. Clin. Investig. 2016, 127, 230–243. [Google Scholar] [CrossRef]
- Mannack, G.; Graf, D.; Donner, M.M.; Richter, L.; Görg, B.; Vom Dahl, S.; Häussinger, D.; Schliess, F. Taurolithocholic Acid-3 Sulfate Impairs Insulin Signaling in Cultured Rat Hepatocytes and Perfused Rat Liver. Cell. Physiol. Biochem. 2008, 21, 137–150. [Google Scholar] [CrossRef]
- Krätzner, R.; Fröhlich, F.; Lepler, K.; Schröder, M.; Röher, K.; Dickel, C.; Tzvetkov, M.V.; Quentin, T.; Oetjen, E.; Knepel, W. A Peroxisome Proliferator-Activated Receptor γ-Retinoid X Receptor Heterodimer Physically Interacts with the Transcriptional Activator PAX6 to Inhibit Glucagon Gene Transcription. Mol. Pharmacol. 2008, 73, 509–517. [Google Scholar] [CrossRef]
- Schinner, S.; Dellas, C.; Schröder, M.; Heinlein, C.A.; Chang, C.; Fischer, J.; Knepel, W. Repression of Glucagon Gene Transcription by Peroxisome Proliferator-Activated Receptor γ through Inhibition of Pax6 Transcriptional Activity. J. Biol. Chem. 2002, 277, 1941–1948. [Google Scholar] [CrossRef]
- Schonhoff, S.E.; Giel-Moloney, M.; Leiter, A.B. Minireview: Development and Differentiation of Gut Endocrine Cells. Endocrinology 2004, 145, 2639–2644. [Google Scholar] [CrossRef]
- Latta, L.; Ludwig, N.; Krammes, L.; Stachon, T.; Fries, F.N.; Mukwaya, A.; Szentmáry, N.; Seitz, B.; Wowra, B.; Kahraman, M.; et al. Abnormal Neovascular and Proliferative Conjunctival Phenotype in Limbal Stem Cell Deficiency Is Associated with Altered microRNA and Gene Expression Modulated by PAX6 Mutational Status in Congenital Aniridia. Ocul. Surf. 2021, 19, 115–127. [Google Scholar] [CrossRef]
- Roux, L.N.; Petit, I.; Domart, R.; Concordet, J.-P.; Qu, J.; Zhou, H.; Joliot, A.; Ferrigno, O.; Aberdam, D. Modeling of Aniridia-Related Keratopathy by CRISPR/Cas9 Genome Editing of Human Limbal Epithelial Cells and Rescue by Recombinant PAX6 Protein. Stem Cells 2018, 36, 1421–1429. [Google Scholar] [CrossRef]
- Mishra, S.; Maurya, S.K.; Srivastava, K.; Shukla, S.; Mishra, R. Pax6 Influences Expression Patterns of Genes Involved in Neurodegeneration. Ann. Neurosci. 2015, 22, 226–231. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Aman, Y.; Ng, C.T.; Chau, W.-H.; Zhang, Z.; Yue, M.; Bohm, C.; Jia, Y.; Li, S.; et al. Amyloid-β Toxicity Modulates Tau Phosphorylation through the PAX6 Signalling Pathway. Brain 2021, 144, 2759–2770. [Google Scholar] [CrossRef]
- Dunckley, T.; Beach, T.G.; Ramsey, K.E.; Grover, A.; Mastroeni, D.; Walker, D.G.; LaFleur, B.J.; Coon, K.D.; Brown, K.M.; Caselli, R.; et al. Gene Expression Correlates of Neurofibrillary Tangles in Alzheimer’s Disease. Neurobiol. Aging 2006, 27, 1359–1371. [Google Scholar] [CrossRef] [PubMed]
- Blalock, E.M.; Geddes, J.W.; Chen, K.C.; Porter, N.M.; Markesbery, W.R.; Landfield, P.W. Incipient Alzheimer’s Disease: Microarray Correlation Analyses Reveal Major Transcriptional and Tumor Suppressor Responses. Proc. Natl. Acad. Sci. USA 2004, 101, 2173–2178. [Google Scholar] [CrossRef]
- Hernandez, F.; Lucas, J.J.; Avila, J. GSK3 and Tau: Two Convergence Points in Alzheimer’s Disease. J. Alzheimer’s Dis. 2012, 33, S141–S144. [Google Scholar] [CrossRef]
- Schaffer, B.A.J.; Bertram, L.; Miller, B.L.; Mullin, K.; Weintraub, S.; Johnson, N.; Bigio, E.H.; Mesulam, M.; Wiedau-Pazos, M.; Jackson, G.R.; et al. Association of GSK3B With Alzheimer Disease and Frontotemporal Dementia. Arch. Neurol. 2008, 65, 1368–1374. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.G.; Welch, C.; Stone, L.; Allan, P.; Barker, R.A.; White, R.B. PAX6 Expression May Be Protective against Dopaminergic Cell Loss in Parkinson’s Disease. CNS Neurol. Disord.—Drug Targets 2016, 15, 73–79. [Google Scholar] [CrossRef] [PubMed]
- White, R.; Thomas, M. Developmental Transcription Factors in Age-Related CNS Disease: A Phoenix Rising from the Ashes? Neural Regen. Res. 2016, 11, 64. [Google Scholar] [CrossRef]
- Bonsberger, J.; Richter, F.; Stanojlovic, M. Impact of Alpha-Synuclein Pathology on Adult Neurogenesis: Evidence for Multilayered Mechanisms. Neural Regen. Res. 2022, 17, 1491–1492. [Google Scholar] [CrossRef]
- Findeiss, E.; Schwarz, S.C.; Evsyukov, V.; Rösler, T.W.; Höllerhage, M.; Chakroun, T.; Nykänen, N.-P.; Shen, Y.; Wurst, W.; Kohl, M.; et al. Comprehensive miRNome-Wide Profiling in a Neuronal Cell Model of Synucleinopathy Implies Involvement of Cell Cycle Genes. Front. Cell Dev. Biol. 2021, 9, 561086. [Google Scholar] [CrossRef] [PubMed]
- Bonafede, R.; Mariotti, R. ALS Pathogenesis and Therapeutic Approaches: The Role of Mesenchymal Stem Cells and Extracellular Vesicles. Front. Cell. Neurosci. 2017, 11, 80. [Google Scholar] [CrossRef]
- Hovden, H.; Frederiksen, J.L.; Pedersen, S.W. Immune System Alterations in Amyotrophic Lateral Sclerosis. Acta Neurol. Scand. 2013, 128, 287–296. [Google Scholar] [CrossRef]
- Thonhoff, J.R.; Simpson, E.P.; Appel, S.H. Neuroinflammatory Mechanisms in Amyotrophic Lateral Sclerosis Pathogenesis. Curr. Opin. Neurol. 2018, 31, 635–639. [Google Scholar] [CrossRef]
- Shukla, S.; Mishra, R. Level of Hydrogen Peroxide Affects Expression and Sub-Cellular Localization of Pax6. Mol. Biol. Rep. 2018, 45, 533–540. [Google Scholar] [CrossRef]
- Maurya, S.K.; Mishra, R. Pax6 Interacts with Iba1 and Shows Age-Associated Alterations in Brain of Aging Mice. J. Chem. Neuroanat. 2017, 82, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Ohsawa, K.; Imai, Y.; Sasaki, Y.; Kohsaka, S. Microglia/Macrophage-specific Protein Iba1 Binds to Fimbrin and Enhances Its Actin-bundling Activity. J. Neurochem. 2004, 88, 844–856. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, H.; Ohsawa, K.; Sasaki, Y.; Kohsaka, S.; Imai, Y. Macrophage/Microglia-Specific Protein Iba1 Enhances Membrane Ruffling and Rac Activation via Phospholipase C-γ-Dependent Pathway. J. Biol. Chem. 2002, 277, 20026–20032. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, X.; Wang, J.; Liang, T.; Gu, Y.; Yang, D. Down-Regulation of PAX6 by Promoter Methylation Is Associated with Poor Prognosis in Non Small Cell Lung Cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 11452–11457. [Google Scholar]
- Zhou, Y.-H.; Wu, X.; Tan, F.; Shi, Y.-X.; Glass, T.; Liu, T.J.; Wathen, K.; Hess, K.R.; Gumin, J.; Lang, F.; et al. PAX6 Suppresses Growth of Human Glioblastoma Cells. J. Neuro-Oncol. 2005, 71, 223–229. [Google Scholar] [CrossRef]
- Zhou, Y.-H.; Tan, F.; Hess, K.R.; Yung, W.K.A. The Expression of PAX6, PTEN, Vascular Endothelial Growth Factor, and Epidermal Growth Factor Receptor in Gliomas: Relationship to Tumor Grade and Survival. Clin. Cancer Res. 2003, 9, 3369–3375. [Google Scholar] [PubMed]
- Pinto, G.R.; Clara, C.A.; Santos, M.J.; Almeida, J.R.W.; Burbano, R.R.; Rey, J.A.; Casartelli, C. Mutation Analysis of Gene PAX6 in Human Gliomas. Genet. Mol. Res. 2007, 6, 1019–1025. [Google Scholar]
- Shu, M.; Zhou, Y.; Zhu, W.; Zhang, H.; Wu, S.; Chen, J.; Yan, G. MicroRNA 335 Is Required for Differentiation of Malignant Glioma Cells Induced by Activation of cAMP/Protein Kinase A Pathway. Mol. Pharmacol. 2012, 81, 292–298. [Google Scholar] [CrossRef]
- Hegge, B.; Sjøttem, E.; Mikkola, I. Generation of a PAX6 Knockout Glioblastoma Cell Line with Changes in Cell Cycle Distribution and Sensitivity to Oxidative Stress. BMC Cancer 2018, 18, 496. [Google Scholar] [CrossRef]
- Roy-Burman, P.; Tindall, D.J.; Robins, D.M.; Greenberg, N.M.; Hendrix, M.J.C.; Mohla, S.; Getzenberg, R.H.; Isaacs, J.T.; Pienta, K.J. Letter to the Editor: Androgens and Prostate Cancer: Are the Descriptors Valid? Cancer Biol. Ther. 2005, 4, 4–5. [Google Scholar] [CrossRef]
- Bonkhoff, H.; Remberger, K. Differentiation Pathways and Histogenetic Aspects of Normal and Abnormal Prostatic Growth: A Stem Cell Model. Prostate 1996, 28, 98–106. [Google Scholar] [CrossRef]
- Shyr, C.; Tsai, M.; Yeh, S.; Kang, H.; Chang, Y.; Wong, P.; Huang, C.; Huang, K.; Chang, C. Tumor Suppressor PAX6 Functions as Androgen Receptor Co-repressor to Inhibit Prostate Cancer Growth. Prostate 2010, 70, 190–199. [Google Scholar] [CrossRef]
- Elvenes, J.; Thomassen, E.I.S.; Johnsen, S.S.; Kaino, K.; Sjøttem, E.; Johansen, T. Pax6 Represses Androgen Receptor-Mediated Transactivation by Inhibiting Recruitment of the Coactivator SPBP. PLoS ONE 2011, 6, e24659. [Google Scholar] [CrossRef] [PubMed]
- Jing, N.; Du, X.; Liang, Y.; Tao, Z.; Bao, S.; Xiao, H.; Dong, B.; Gao, W.-Q.; Fang, Y.-X. PAX6 Promotes Neuroendocrine Phenotypes of Prostate Cancer via Enhancing MET/STAT5A-Mediated Chromatin Accessibility. J. Exp. Clin. Cancer Res. 2024, 43, 144. [Google Scholar] [CrossRef] [PubMed]
- Rickman, D.S.; Beltran, H.; Demichelis, F.; Rubin, M.A. Biology and Evolution of Poorly Differentiated Neuroendocrine Tumors. Nat. Med. 2017, 23, 664–673. [Google Scholar] [CrossRef]
- Beltran, H.; Rickman, D.S.; Park, K.; Chae, S.S.; Sboner, A.; MacDonald, T.Y.; Wang, Y.; Sheikh, K.L.; Terry, S.; Tagawa, S.T.; et al. Molecular Characterization of Neuroendocrine Prostate Cancer and Identification of New Drug Targets. Cancer Discov. 2011, 1, 487–495. [Google Scholar] [CrossRef]
- De Wit, R.; De Bono, J.; Sternberg, C.N.; Fizazi, K.; Tombal, B.; Wülfing, C.; Kramer, G.; Eymard, J.-C.; Bamias, A.; Carles, J.; et al. Cabazitaxel versus Abiraterone or Enzalutamide in Metastatic Prostate Cancer. N. Engl. J. Med. 2019, 381, 2506–2518. [Google Scholar] [CrossRef]
- Jin, M.; Gao, D.; Wang, R.; Sik, A.; Liu, K. Possible Involvement of TGF-β-SMAD-mediated Epithelial-mesenchymal Transition in Pro-metastatic Property of PAX6. Oncol. Rep. 2020, 44, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.R.; Chen, H. MicroRNAs in Pathogenesis of Breast Cancer: Implications in Diagnosis and Treatment. World J. Clin. Oncol. 2014, 5, 48–60. [Google Scholar] [CrossRef]
- Xia, X.; Yin, W.; Zhang, X.; Yu, X.; Wang, C.; Xu, S.; Feng, W.; Yang, H. PAX6 Overexpression Is Associated with the Poor Prognosis of Invasive Ductal Breast Cancer. Oncol. Lett. 2015, 10, 1501–1506. [Google Scholar] [CrossRef]
- Urrutia, G.; Laurito, S.; Campoy, E.; Nasif, D.; Branham, M.T.; Roqué, M. PAX6 Promoter Methylation Correlates with MDA-MB-231 Cell Migration, and Expression of MMP2 and MMP9. Asian Pac. J. Cancer Prev. 2018, 19, 2859–2866. [Google Scholar] [CrossRef] [PubMed]
- Buck, M.B.; Knabbe, C. TGF-Beta Signaling in Breast Cancer. Ann. New York Acad. Sci. 2006, 1089, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.A.; Lee, Y.-H.; Schiemann, W.P. Role of TGF-β and the Tumor Microenvironment During Mammary Tumorigenesis. Gene Expr. 2011, 15, 117–132. [Google Scholar] [CrossRef]
- Bhinge, A.; Poschmann, J.; Namboori, S.C.; Tian, X.; Loh, S.J.H.; Traczyk, A.; Prabhakar, S.; Stanton, L.W. Mi R -135b Is a Direct PAX6 Target and Specifies Human Neuroectoderm by Inhibiting TGF-β/ BMP Signaling. EMBO J. 2014, 33, 1271–1283. [Google Scholar] [CrossRef]
- Khoshakhlagh, M.; Soleimani, A.; Binabaj, M.M.; Avan, A.; Ferns, G.A.; Khazaei, M.; Hassanian, S.M. Therapeutic Potential of Pharmacological TGF-β Signaling Pathway Inhibitors in the Pathogenesis of Breast Cancer. Biochem. Pharmacol. 2019, 164, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Ling, L.; Van Dam, H.; Zhou, F.; Zhang, L. TGF-β Signaling in Cancer Metastasis. Acta Biochim. Biophys. Sin. 2018, 50, 121–132. [Google Scholar] [CrossRef]
- Valastyan, S.; Weinberg, R.A. Tumor Metastasis: Molecular Insights and Evolving Paradigms. Cell 2011, 147, 275–292. [Google Scholar] [CrossRef]
- Yeh, C.-H.; Chen, R.-Y.; Wu, T.-H.; Chang, S.-Y.; Hsieh, T.-Y.; Shih, Y.-L.; Lin, Y.-W. Promoter Hypermethylation-Mediated Downregulation of PAX6 Promotes Tumor Growth and Metastasis during the Progression of Liver Cancer. Clin. Epigenetics 2024, 16, 174. [Google Scholar] [CrossRef]
- Shih, Y.-L.; Kuo, C.-C.; Yan, M.-D.; Lin, Y.-W.; Hsieh, C.-B.; Hsieh, T.-Y. Quantitative Methylation Analysis Reveals Distinct Association between PAX6 Methylation and Clinical Characteristics with Different Viral Infections in Hepatocellular Carcinoma. Clin. Epigenetics 2016, 8, 41. [Google Scholar] [CrossRef]
- Zhu, Q.; Huang, B.; Wu, L.; Luo, Q. Mechanism of PAX6 Overexpression in Inhibiting the Growth of Hepatocellular Carcinoma Cells and Promoting the Killing Ability of the Natural Killer Cells. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2023, 48, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Mascarenhas, J.B.; Young, K.P.; Littlejohn, E.L.; Yoo, B.K.; Salgia, R.; Lang, D. PAX6 Is Expressed in Pancreatic Cancer and Actively Participates in Cancer Progression through Activation of the MET Tyrosine Kinase Receptor Gene. J. Biol. Chem. 2009, 284, 27524–27532. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, T.S.; Lee, M.H.; Cho, I.R.; Ryu, J.K.; Kim, Y.-T.; Lee, S.H.; Paik, W.H. Pancreatic Cancer Treatment Targeting the HGF/c-MET Pathway: The MEK Inhibitor Trametinib. Cancers 2024, 16, 1056. [Google Scholar] [CrossRef] [PubMed]
- Comoglio, P.M.; Trusolino, L.; Boccaccio, C. Known and Novel Roles of the MET Oncogene in Cancer: A Coherent Approach to Targeted Therapy. Nat. Rev. Cancer 2018, 18, 341–358. [Google Scholar] [CrossRef] [PubMed]
- Shaw, R.J.; Cantley, L.C. Ras, PI(3)K and mTOR Signalling Controls Tumour Cell Growth. Nature 2006, 441, 424–430. [Google Scholar] [CrossRef]
- Guo, X.-F.; Wang, L.-L.; Zheng, F.-M.; Li, H.-P. PAX6 Enhances Nanog Expression by Inhibiting NOTCH Signaling to Promote Malignant Properties in Small Cell Lung Cancer Cells. Heliyon 2025, 11, e41795. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, C.T.; Chen, J.; Pankratz, M.T.; Xi, J.; Li, J.; Yang, Y.; Lavaute, T.M.; Li, X.-J.; Ayala, M.; et al. Pax6 Is a Human Neuroectoderm Cell Fate Determinant. Cell Stem Cell 2010, 7, 90–100. [Google Scholar] [CrossRef]
- Wu, D.; Zhang, T.; Liu, Y.; Deng, S.; Han, R.; Liu, T.; Li, J.; Xu, Y. The PAX6-ZEB2 Axis Promotes Metastasis and Cisplatin Resistance in Non-Small Cell Lung Cancer through PI3K/AKT Signaling. Cell Death Dis. 2019, 10, 349. [Google Scholar] [CrossRef]
- Kiselev, Y.; Andersen, S.; Johannessen, C.; Fjukstad, B.; Standahl Olsen, K.; Stenvold, H.; Al-Saad, S.; Donnem, T.; Richardsen, E.; Bremnes, R.M.; et al. Transcription Factor PAX6 as a Novel Prognostic Factor and Putative Tumour Suppressor in Non-Small Cell Lung Cancer. Sci. Rep. 2018, 8, 5059. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yue, W.; Zhang, L.; Ma, L.; Jia, W.; Qian, Z.; Zhang, C.; Wang, Y. Downregulation of PAX6 by shRNA Inhibits Proliferation and Cell Cycle Progression of Human Non-Small Cell Lung Cancer Cell Lines. PLoS ONE 2014, 9, e85738. [Google Scholar] [CrossRef]
- MacCarthy, A.; Draper, G.J.; Steliarova-Foucher, E.; Kingston, J.E. Retinoblastoma Incidence and Survival in European Children (1978–1997). Report from the Automated Childhood Cancer Information System Project. Eur. J. Cancer 2006, 42, 2092–2102. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Li, B.; Zhang, H.; Jonas, J.B.; Zhao, B.; Shen, L.; Wang, Y. Pax6 Regulates Proliferation and Apoptosis of Human Retinoblastoma Cells. Investig. Opthalmology Vis. Sci. 2011, 52, 4560. [Google Scholar] [CrossRef]
- Li, L.; Li, B.; Zhang, H.; Bai, S.; Wang, Y.; Zhao, B.; Jonas, J.B. Lentiviral Vector-Mediated PAX6 Overexpression Promotes Growth and Inhibits Apoptosis of Human Retinoblastoma Cells. Investig. Opthalmology Vis. Sci. 2011, 52, 8393–8400. [Google Scholar] [CrossRef]
- Meng, B.; Wang, Y.; Li, B. Suppression of PAX6 Promotes Cell Proliferation and Inhibits Apoptosis in Human Retinoblastoma Cells. Int. J. Mol. Med. 2014, 34, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Golabchi, K.; Soleimani-Jelodar, R.; Aghadoost, N.; Momeni, F.; Moridikia, A.; Nahand, J.S.; Masoudifar, A.; Razmjoo, H.; Mirzaei, H. MicroRNAs in Retinoblastoma: Potential Diagnostic and Therapeutic Biomarkers. J. Cell. Physiol. 2018, 233, 3016–3023. [Google Scholar] [CrossRef]
- Mi, D.; Carr, C.B.; Georgala, P.A.; Huang, Y.-T.; Manuel, M.N.; Jeanes, E.; Niisato, E.; Sansom, S.N.; Livesey, F.J.; Theil, T.; et al. Pax6 Exerts Regional Control of Cortical Progenitor Proliferation via Direct Repression of Cdk6 and Hypophosphorylation of pRb. Neuron 2013, 78, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Sudou, N.; Nomura-Komoike, K.; Iida, T.; Fujieda, H. Age- and Cell Cycle-Related Expression Patterns of Transcription Factors and Cell Cycle Regulators in Müller Glia. Sci. Rep. 2022, 12, 19584. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Participation of Pax6 in the implementation of intracellular signaling pathways: A—neurogenesis: green arrows show the signaling pathways of activation of Pax6 expression under the influence of the Wnt/β-catenin signaling pathway at the early stages of neuroepithelial proliferation as well as with the participation of δ-catenin at the stage of differentiation, adhesion and migration of neurons, red lines indicate the inhibitory effect of the fibroblast growth factor (FGF) signaling pathway during the differentiation of spinal cord neurons; B—gliogenesis: activation of the expression of the glutamate transporter GLT1 under the influence of Pax6 on the astrocyte membrane during maturation and differentiation is shown; C—glioblastoma: Pax6 is a downstream target of (miR)-355. In glioblastomas, expression of this microRNA inhibits Pax6 activity, preventing cellular differentiation and maintaining cells in the proliferative phase of the cell cycle; D—breast tumor: green arrows show the activation of the TGF-β/SMAD signaling pathway that triggers the proliferation and metastasis of breast tumor cells; E—retinoblastoma: the promoter role of Pax6 in breast tumor development is demonstrated through suppression of apoptotic proteins. Tumor suppression is associated with a decrease in Pax6 expression under the influence of miRNA-356b-3p; F—Parkinson’s disease: the inhibitory effect of Pax6 overexpression on oxidative stress and neuronal apoptosis is demonstrated; G—Alzheimer’s disease: Pax6 activates the kinase GSK-3b, which in turn hyperphosphorylates the pRB factor via CD1 and CDK 4/6. pRB activates the transcription factor E2F1, which triggers transcription and translation of the Pax6 and cMyb genes in the nucleus. Pax6 expression enhances the activity of Cdk5 and Mapk1 kinases, which phosphorylate the tau protein. Phosphorylated tau can, in turn, independently activate the expression of the Pax6 gene in the nucleus.
Figure 1.
Participation of Pax6 in the implementation of intracellular signaling pathways: A—neurogenesis: green arrows show the signaling pathways of activation of Pax6 expression under the influence of the Wnt/β-catenin signaling pathway at the early stages of neuroepithelial proliferation as well as with the participation of δ-catenin at the stage of differentiation, adhesion and migration of neurons, red lines indicate the inhibitory effect of the fibroblast growth factor (FGF) signaling pathway during the differentiation of spinal cord neurons; B—gliogenesis: activation of the expression of the glutamate transporter GLT1 under the influence of Pax6 on the astrocyte membrane during maturation and differentiation is shown; C—glioblastoma: Pax6 is a downstream target of (miR)-355. In glioblastomas, expression of this microRNA inhibits Pax6 activity, preventing cellular differentiation and maintaining cells in the proliferative phase of the cell cycle; D—breast tumor: green arrows show the activation of the TGF-β/SMAD signaling pathway that triggers the proliferation and metastasis of breast tumor cells; E—retinoblastoma: the promoter role of Pax6 in breast tumor development is demonstrated through suppression of apoptotic proteins. Tumor suppression is associated with a decrease in Pax6 expression under the influence of miRNA-356b-3p; F—Parkinson’s disease: the inhibitory effect of Pax6 overexpression on oxidative stress and neuronal apoptosis is demonstrated; G—Alzheimer’s disease: Pax6 activates the kinase GSK-3b, which in turn hyperphosphorylates the pRB factor via CD1 and CDK 4/6. pRB activates the transcription factor E2F1, which triggers transcription and translation of the Pax6 and cMyb genes in the nucleus. Pax6 expression enhances the activity of Cdk5 and Mapk1 kinases, which phosphorylate the tau protein. Phosphorylated tau can, in turn, independently activate the expression of the Pax6 gene in the nucleus.
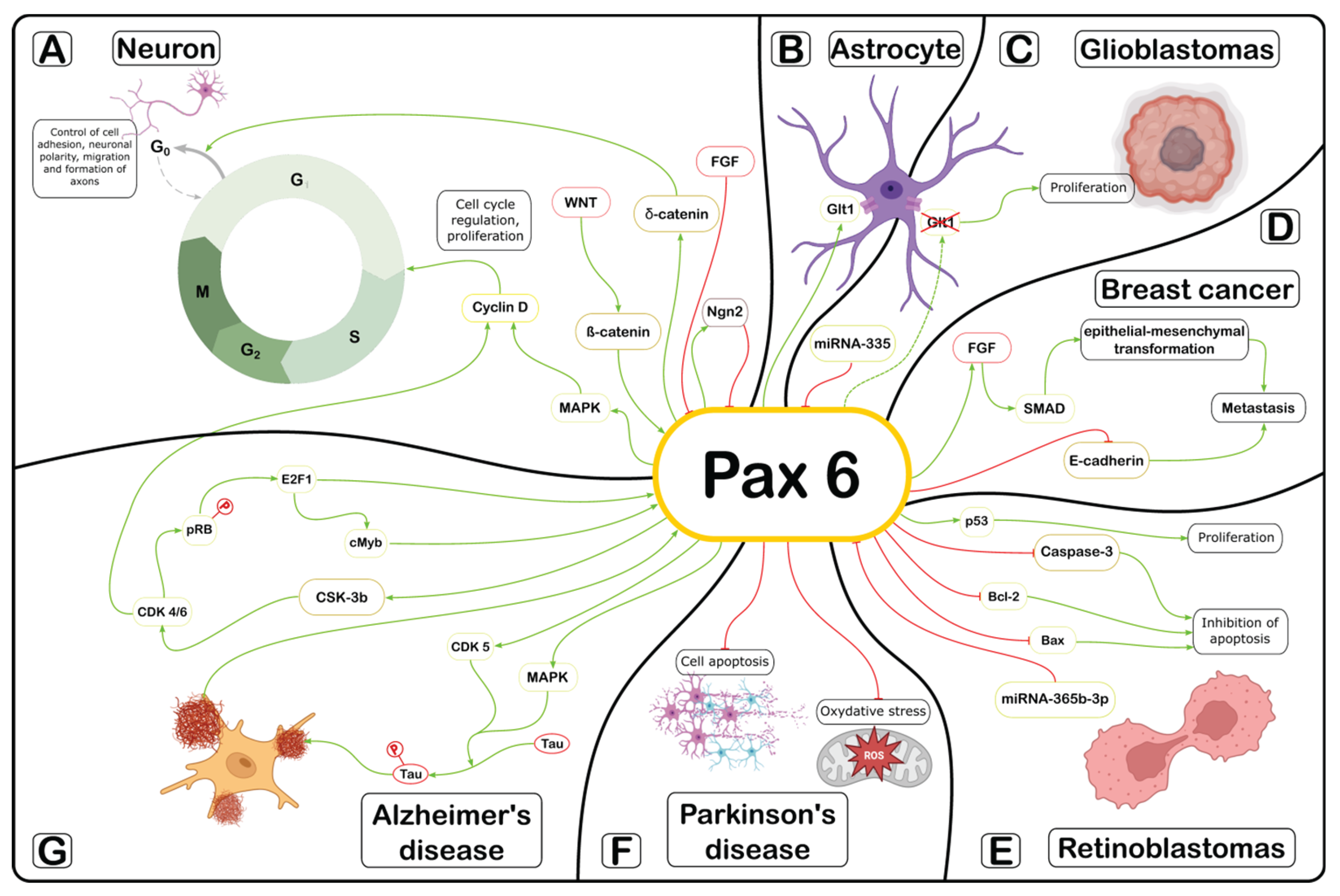
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



