
Submitted:
28 October 2025
Posted:
28 October 2025
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
To contrast the COVID-19 pandemic brought by the corona virus SARS-CoV-2, two mRNA-based anti-COVID-19 vaccines (by Pfizer-BioNTech and Moderna) were rela-tively soon made available and deployed worldwide based on an emergency approval. Being considered vulnerable and at risk of infection, cancer patients have been priori-tized for COVID-19 vaccination and vaccinated repeatedly because of the short-time protection provided by these vaccines. During the pandemic, because of the large number of infected patients requiring assistance, many hospitals opted for giving pri-ority to these patients while postponing the specialist treatment for other pathologies, including cancer. Recently, a surge in the incidence and rapid progression of cancers has been observed in many countries, which could (at least partially) represent those cancers undiagnosed or untreated during the pandemic. It has also been suggested that the SARS-CoV-2 itself or even the anti-COVID-19 mRNA vaccines could have contrib-uted to the recurrence and worse clinical outcome in cancer patients, given the high incidence of COVID-19 in hospitalized patients and that these patients have been vac-cinated with priority several times and in a short period. Although it appears ex-tremely unlikely that SARS-CoV-2 and anti-COVID-19 mRNA vaccines elicit genotoxic events and cause neo-cancerogenesis in a short time, they could still cause non-genotoxic pro-carcinogenic effects by triggering an exaggerated inflammatory re-action, compromising immune homeostasis, stimulating cell proliferation, and nega-tively affecting cellular stress response and damage repair machinery. This could re-sult in the promotion of regrowth of dormant micrometastases or relapse of stable minimal residual disease. Such a harmful outcome may likely result from a synergy between the virus and the vaccine, especially in multi-vaccinated and multi-infected individuals. Here, I bring the cell pathologist's point of view and discuss the multiple possible mechanisms by which the virus and the anti-COVID-19 mRNA vaccine might favor tumorigenesis. While a causal link cannot be established at this stage, knowledge of potential carcinogenic risks could help doctors and health policymakers take the best actions to protect vulnerable patients and convince the vaccine developer to de-sign a vaccine free from such harm.
Keywords:
COVID-19
; mRNA vaccine
; lipid nanoparticle
; autophagy
; tumor dormancy
; epigenetics
; tumor microenvironment
; cytokines
; tumor suppressors
1. Introduction
On March 2020 the WHO officially declared COVID-19, the infection brought by the Coronavirus SARS-CoV2, as a global pandemic. At that time, because of the large number of infected patients requiring assistance, many hospitals opted to give priority to these patients, converting specialist wards into COVID-dedicated wards, and this has inevitably postponed specialist treatment for other pathologies, including cancer [1,2]. As a result, diagnosis and medical care at earlier stages of disease were negated to a large population [3]. Few months later (i.e., December 2020 and January 2021), the administration of mRNA-based anti-COVID-19 vaccines (BNT162b2/Comirnaty from Pfizer-BioNTech and mRNA-1273/Spikevax from Moderna), manufactured with a novel technology and approved for emergency use, was prioritized for elders (over 60s) and “vulnerable” (so-called “frail”) patients with chronic diseases such as neuro-degenerative disorders, autoimmune diseases, and cancer particularly among others [4,5]. Soon after, vaccination was made mandatory for health-care workers and thereafter for the general population to be able to work in public settings. As of August 2023, a total of 84.8% of the European Union adult population had been vaccinated at least once against the virus (https://commission.europa.eu/strategy-and-policy/coronavirus-response/safe-covid-19-vaccines-europeans_en; accessed August 31, 2025). As of April 2023, almost 400 million doses of Pfizer-BioNTech and 250 million doses of Moderna vaccines have been administered in the United States (https://www.statista.com/statistics/1198516/covid-19-vaccinations-administered-us-by-company/; accessed on August 31st, 2025). Since then, the rate of vaccination has steadily declined in all western countries.
During the pandemic, almost 775,615,736 confirmed cases and over and 7,051,323 deaths have been attributed to COVID-19 by the World Health Organization (WHO) (https://covid19.who.int/; accessed on June 28th, 2024). According to a recent study, during the period December 2020 through March 2023 the anti-COVID-19 vaccines would have saved up to 1.6 million of lives among people aged >25 years old in European countries [6].
The COVID-19 pandemic was officially declared terminated on May 5th, 2023 [7], yet WHO warns the governments on the need to continue the anti-COVID-19 vaccination campaign as a preventive measure to reduce the hospitalization burden. Again, cancer patients are forced to vaccinate against COVID-19 as per the recommendation of the scientific societies of oncologists (ASCO, American Society of Clinical Oncology in US, and ESMO, European Society of Medical Oncology in Europe, and others similar). Comirnaty (Pfizer-BioNTech) and Spikevax (Moderna) remain the most widely used anti-COVID-19 vaccines.
Presently, the worst of the virus seems to be over. On the opposite, cancer is on the rise worldwide [8], with almost 20 million new cases and 9.7 million cancer-related deaths in 2022, and it is predicted to further increase up to 35 million new cases in 2050 (https://www.uicc.org/news/globocan-2022-latest-global-cancer-data-shows-rising-incidence-and-stark-inequities; accessed on December 30th, 2024). In the United States, cancer mortality rate decreased by 33% from 1991 through 2021 [9]. In 2025, it is estimated that two million new cases of cancer will be diagnosed and about 618,000 people will die from cancer in the United States (https://www.cancer.gov/about-cancer/understanding/statistics; accessed on August 31st, 2025). More worrisome is the recent projection showing an increased incidence of cancer at younger age (<50) for the generation born in the1965-1980 compared to the generation born before 1964 [10]. The style of life is blamed as the main culprit for such trend, though other factors including infections and medications that negatively impact on the immune system and metabolic homeostasis should not be neglected.
The case reports describing the sudden occurrence of rapidly progressing cancers diagnosed at advanced stage in otherwise healthy patients or the relapse and fast progression of cancers in cancer-bearing patients after anti-COVID-19 vaccination are increasing in the peer-reviewed literature, not considering the retracted one (see Table 1 and Table 2). Consequential- and time-correlations, which per se do not imply a causal correlation, between the vaccination campaign and such increased incidence of cancer has raised concerns about the possible causal link. However, establishing a causal link is challenging because the national cancer registries do not consider the newly diagnosed or the recurrent cancers as possibly linked to the vaccination status. On the other hand, prospective active pharmacovigilance that matches vaccinated and unvaccinated individuals, healthy or cancer carriers, has not been pursued in the last three years, and would in any case be impracticable because both populations of healthy individuals aged > 60 years (those more susceptible to developing cancer) and cancer patients have largely been vaccinated. Adding to the complexity, these patients likely have also been infected with SARS-CoV2 before and/or after vaccination. Now that five years have passed since the spread of the virus throughout the world and that the vaccine has been administered to a large population for three years, we can take stock and try to answer the fundamental questions: (i) is it plausible that the COVID-19 virus and the anti-COVID-19 mRNA vaccines may cause cancer? And, if yes, (ii) how much have they contributed and how much will they contribute in the future to the increase in cancer? When addressing these questions, we must consider that cancer intrinsically tends to evolve to the worse (despite the treatment), and that these patients have been vaccinated three or more times and, most likely, have also contracted COVID-19 [11,12,13]. Thus, it is objectively difficult to determine and weigh the factor(s) causing clinical worsening in cancer patients. Similarly, in the case of (apparently) healthy individuals who experience the sudden onset of cancer after vaccination we must consider possible previous SARS-CoV2 infections or other predisposing factors that could have favored carcinogenesis. In the latter case, the virus and the vaccine could still have had an add-on triggering role. But one thing must be clarified immediately: the concept of "turbocancer" developing in little more than two years has no scientific basis, even in the case of injecting carcinogenic chemicals into the bloodstream.
Here, I will not delve into the causality assessment, which would be intricate and challenging [14], and instead will present and discuss the potential mechanisms and pathways through which the SARS-CoV2 virus and the anti-COVID-19 genetic vaccines could contribute to carcinogenesis or worsening of pre-existing tumors. This knowledge is useful for informing policymakers and clinicians in choosing the best public health intervention to protect citizens and patients facing similar viral pandemics in the future. The objective is not to blame cancer on the virus or anti-COVID-19 vaccines but rather to instill doubts and stimulate reflection free from any prejudice, dogmas, and conflict of interest on the safety of these mRNA vaccines and on the best precautions to implement to protect patients at risk of viral infections.
2. The Virus, the Cancer, and the mRNA Vaccine: the Ugly, the Bad, and the Good?
Parodying the cult film “The Good, the Bad, and the Ugly”, we can say for sure that the "Ugly" is the virus and the "Bad" is the cancer, but are we sure that the vaccine is the "Good"?
2.1. The Cancer
Let’s start with the “Bad” guy. Cancer is not a single well-defined disease, rather it is a complex-multifaceted disease that evolves continuously and dynamically changes its features in response to local and systemic environmental signals [15,16]. At the time of cancer diagnosis, we face a mass that is constituted by many different malignant clones of cells that behave differently (in terms of proliferation, metabolism, survival, migration, and other characteristics) and that very likely have already spread in other body districts to form metastases, some of which may not be detectable by diagnostic imaging [17]. During this process, other cells in the same tissue may undergo transformation into cancer cells and start that very same process of clonal evolution and spreading. Thus, when the patient eventually calls upon a doctor because of the symptoms the body is likely to have many different cancers at various degrees of progression in one or more organ(s). According to the “somatic mutation theory” transformation into a cancer cell results from the accumulation of mutations in the functioning of several genes (belonging to the families of oncogenes, tumor suppressor genes, and DNA repair genes) that control cell proliferation, cell differentiation, cell death, cell migration, cell metabolism, and systems for repairing the DNA and protein/organelle damages [17]. Mutated functioning of these genes results from either genetic change in their DNA coding sequences or from epigenetic changes in their expression [18,19]. Unrepaired genetic and epigenetic mutations in so many genes accumulate over many years, and this explains why spontaneous (sporadic) cancers develop in decades and are in fact diagnosed more frequently in the 70s [9]. However, this can be anticipated in cases of chronic exposure to environmental mutagenic/epimutagenic factors (so-called “genotoxic and non-genotoxic carcinogens”) and/or malfunctioning (in some cases genetically inherited) of the machineries that keep under check the cellular damages and the abnormal cell behavior [20,21]. However, in some cancers massive genomic alterations have been shown to occur as a single catastrophic event in a short time [22].
Still, the presence of mutated oncogenes and tumor suppressor genes is not sufficient for the development of cancer because the surrounding microenvironment can build a barrier that contrasts the proliferation and spread of such transformed cells [23,24]. Even more intriguingly, tumors without genetic alterations have also been described, which calls into question the “somatic mutation theory” [35].
2.2. The Virus
What can we say about the “Ugly” guy? SARS-CoV2 was named so due to its high similarity with SARS-CoV, the Coronavirus that caused the very similar respiratory distress syndrome described by Carlo Urbani in 2003 [26]. COVID-19 may present with mild to severe flu-like symptoms, though in certain patients, particularly elders and with co-morbidities (such as cardiovascular, diabetes, obesity), the disease can rapidly progress and lead to death following respiratory distress and multiorgan failure arising from an hyperactivation of the inflammatory response associated with hyperproduction of cytokines and multiple thromboembolisms [27,28]. The spheric virion (approx. 100 nm in diameter) consists of an envelope made of a by-layer lipid membrane with inserted the structural proteins E (Envelope), M (Membrane) and S (Spike, a highly glycosylated protein that assembles as trimers) and containing a 29.9 kb single-stranded, positive-sense RNA filament complexed with the Nucleocapsid protein (N) [29]. The virus exploits the Spike (S) protein to infect the cells through binding to the angiotensin-converting enzyme 2 (ACE2) protein expressed on the membrane of endothelial and epithelial cells of various organs, particularly the lungs, intestine, and kidneys [29,30]. The SARS-CoV2 Spike protein presents the unique polybasic sequence (681PRRAR685) for the furin-mediated cleavage into the two subunits S1 (aa 1-685, that contains the ACE2 binding domain) and S2 (aa 686-1273, that mediates the virion envelope fusion with host cellular membrane), and this peculiar feature is believed to enhance the virus cellular infectivity and transmissibility [24]. Virus entry is also facilitated by the proteolytic cleavage of the S protein (at the furin site) by the recipient cell-surface serine protease TMPRSS2, which promotes the virion-cell fusion mediated by the S2 subunit.
Endocytosis and endosomal cysteine proteases cathepsins B and L can also contribute to the virus entry and thereafter release of the viral RNA in the cytoplasm. Once entered the cell, the viral RNA is freed in the cytoplasm and is copied as full length, for inclusion in the new virions, and as sub-genomic RNA fragments for directing the synthesis of the structural and accessory proteins. The whole genome codes for the four structural proteins (E, M, S and N), two polyproteins (ORF1a and ORF1b) and six accessory proteins (of unknown function) [29,30]. The polyproteins ORF1a and ORF1b are proteolyzed, respectively, by the Papain-like protease (PLpro=nsp3) and the 3-chymotrypsin-like protease (3CLpro, aka Main protease Mpro= nsp5) to generate 16 nonstructural proteins (NSP 1-11 and NSP12-16, respectively) necessary for viral replication and assembly [29,30]. Virus replication involves the formation of endoplasmic reticulum-derived double-membrane vesicles, that share similarities with autophagosomes, and the assembled virions then leave the cell by exocytosis passing through the Golgi complex or the secretory lysosome pathway [31,32]. The autophagy-lysosomal pathway plays a dual role in viral infection and replication: on one hand it can lead to lysosomal degradation of the whole virion yet on the opposite it can be manipulated by the virus to serve as membrane platform (the double membrane vesicles) for its replication and assembly [32,33]. The SARS-CoV2 can in fact be directed for lysosomal degradation once it enters via endocytosis or it is in the cytoplasm, yet certain viral proteins (namely NSP6) can impair the autophagosome-lysosome fusion and lysosomal degradation of the viral particles resulting in the accumulation of autophagosomes [32,34]. Autophagy also plays a role in innate and adaptive (MHC-dependent antigen presentation) immune responses [35]. Therefore, SARS-CoV2 disruption of autophagy in antigen presenting cells can compromise the anti-viral immune response. Since autophagy opposes to inflammasome biogenesis in immune and epithelial cells, the disruption of autophagy by SARS-CoV2 may lead to inflammasome-dependent pyroptosis in infected airway cells [36] and cytokine storm and thromboembolisms [37,38]. The question here is whether cancer patients are more likely to be infected by SARS-CoV2 and, in that case, what could be the biological explanation. It is a fact that COVID-19 is diagnosed with high frequency (and often with fatal outcome) in cancer patients, and the simplest explanations are the hospitalization (where the probability of encountering the virus is high) and immunodepression arising from the cancer itself and the treatments [39]. Yet, it has been argued that, paradoxically, immunodepression could protect the SARS-CoV2-infected cancer patients from the fatal risk of hyper-cytokinemia [40].
As for factors that may favor infectivity in cancer patients, the most obvious ones to consider are the cell surface ACE2 (the SARS-CoV2 receptor) and TMPRSS2 (the enzyme that processes the Spike protein to facilitate the cellular entry of SARS-CoV2). High levels of ACE2 and of TMPRSS2 were found, respectively, in renal, colorectal and gastric carcinomas and in prostate, colorectal and esophagogastric carcinomas [41]. It has been hypothesized that patients bearing such cancers are more prone to be infected and to exhibit the severe form of COVID-19. In prostate cancer cells, TMPRSS2 has been shown to activate the protease-activated receptor-2, which triggers downstream signaling pathways associated with inflammation, metastasis, and invasion (see below). Interestingly, androgen deprivation therapy decreases the expression of TMPRSS2 and reduces the risk of SARS-CoV2 infection in prostate cancer patients [42]. In addition to ACE2, other membrane proteins may act as Spike-receptors (or co-receptors) for SARS-CoV2 among which we only mention AXL [43], Neuropilin-1 [44], and CD147 [45] that are highly expressed on cancer cells and might explain the increased susceptibility of cancer patients to infection [46,47,48].
There is also the possibility that susceptibility to SARS-CoV2 infection and to cancer development shares some genetic and epigenetic factors. Genomic and epigenomic studies have delineated the host genetic determinants of COVID-19 susceptibility and of the clinical outcome [49,50,51,52]. In a study where the gene and protein expression of seventeen COVID-19 susceptibility genes were correlated with lung cancer prognosis, it was found that the hyper-expression of FYCO1, CXCR6, XCR1, and TAC4 in cancer cells was protective while that of TMEM65 and OAS1 represented a risk factor for SARS-CoV2 infection [53]. Another study found that the genetic predisposition to colorectal or to lung cancer was causally associated with a decreased or increased susceptibility to COVID-19 severity, respectively, and this association pointed to the LZTFL1, CCR9, FYCO1, CXCR6, XCR1, and ABO genes [54]. To be noted, gene mutations altering the tertiary structure of the FYCO1 protein were suggestive of increased viral replication and spreading via enhanced exocytosis, which could explain the severity of COVID-19 [55].
2.3. The Vaccine
Finally, in 2021 it entered the scene what was hailed as the “Good” guy, i.e. the vaccine. The two mostly used anti-COVID-19 mRNA vaccines, manufactured by Pfizer-BioNTech and Moderna, have been approved under emergency circumstances by drug regulatory agencies (FDA in US and EMA in EU) for the prevention of COVID-19 disease in individuals 16 years of age and older. Approval was based on a 3-month trial demonstrating greater than 94% (relative) efficacy in preventing infection and severity of outcomes and showing only mild to moderate reactions in the 2 months after the second dose [56,57]. It is to be stressed that cancer patients were not included in these clinical trials.
Both these vaccines are made of lipid nanoparticles (LNPs) containing the coding mRNA for the Spike protein. In this sense, they do not act like traditional protein-based vaccines in that the immunogenic protein is synthesized by the host, which makes this product more like an “immunomodulatory genetic pro-drug” (for the sake of simplicity, hereafter I will call it “mRNA pro-vaccine”). It has also been noticed that these “mRNA pro-vaccines” do not contrast SARS-CoV2 infection, instead they induce the synthesis of neutralizing IgG that can limit the reproduction and organ spread of the virus thus attenuating the clinical symptoms of the disease [58], and for this they are better known as anti-COVID-19 vaccines. This is also due to the inability of intramuscular injection of this mRNA pro-vaccine to trigger the production of anti-Spike mucosal IgA even after three doses [59]. More of concern is the fact that the multiple vaccinations with these products shift the immune response to a tolerant response where the inert subclass IgG4 are predominantly produced [60], particularly in patients that are infected after the vaccination [61]. The question here is: “How much it revealed to be useful and safe for cancer patients to be vaccinated for COVID-19 with these mRNA pro-vaccines?”. Cancer patients are generally immunosuppressed, as a side effect of both the treatments (many chemotherapeutics are myelosuppressive) and the disease itself, and this makes them more vulnerable to infections, and hospitalization itself increases the risk of exposure to bacteria and viruses. Not surprisingly, patients with solid or hematologic cancers, and particularly those with metastases, were shown to be more susceptible to contract the severe form of COVID-19 [62,63].
Anti-influenza (traditionally made) vaccines are routinely administered to onco-hematologic and solid cancer-bearing patients, based on the assumption that immunization will protect these vulnerable patients from severe outcomes with no side effects. Based on these considerations and on the results of the clinical trials claiming a relative (not absolute) 95% efficacy [56,57], the anti-COVID vaccination has been prioritized for cancer patients [62,64], neglecting the fact that those clinical trials did not include such typology of patients [56,57]. Several studies have previously shown that vaccination against influenza viruses fails to elicit the expected protection in patients with solid cancer or hematological malignancies [65,66]. It has also been emphasized that cancer patients should not be vaccinated while under radio- or chemotherapy because of the inefficacious immune response [67]. ESMO press release on 20 Sept 2021 emphasized the data reported at the ESMO Congress 2021 proving the safety and the protective efficacy of two or better three vaccine doses in cancer patients (https://www.esmo.org/newsroom/press-and-media-hub/esmo-media-releases/the-evidence-is-in-covid-vaccines-do-protect-patients-with-cancer; accessed on June 27th, 2024). A multicenter study in a cohort of 84 non-vaccinated and 49 vaccinated (majority with mRNA-based vaccine; one-third with three doses) cancer patients who tested positive for SARS-CoV2 reported that in the latter group COVID-19 was milder and the vaccine better protected from COVID-19-related death [68]. These reports were based on a relatively short period of observation and from a few cohorts. Few months later (in June 2022), the first real-world data analysis showed that breakthrough infections, even with severe outcomes, may occur in cancer patients vaccinated with mRNA-based anti-COVID-19 vaccines (the rate was lower for Moderna vaccine respect to the Pfizer vaccine) [69]. Compared to healthy controls, cancer patients receiving three doses of the Comirnaty (Pfizer-BioNTech) mRNA vaccine showed lower cell-mediated response and lower anti-Spike antibody titers, indicating the need for additional boosters to provide protection [70,71].
Other studies confirmed that vaccinated cancer patients can contract the SARS-CoV2 infection and that those under treatment, particularly the hematologic patients receiving anti-CD20 therapy, have an increased risk for severe COVID-19 [72,73,74,75].
Patients bearing solid cancers under treatment showed sub-optimal seroconversion in response to anti-COVID-19 mRNA pro-vaccine [76] and may develop serious immune-related adverse effects [77] Compared to matched healthy patients, the humoral immune response to BNT162b2 (Pfizer-BioNTech Comirnaty) mRNA pro-vaccine was markedly lower in B chronic lymphocytic leukemias patients under treatment with Bruton Kinase inhibitors or venetoclax ± anti-CD20 [78].
Corticosteroids are routinely administered to cancer patients as co-medication and, given their immune-suppressive activity, one may expect a low vaccination efficacy in these patients [79]. Similarly, since PD-1 blockade impairs the CD8 response to antigenic stimulation [80], it is likely that therapies with immune-checkpoint inhibitors in solid cancer patients would abrogate the T-cell specific response to COVID-19 vaccination. While chemotherapy has been reported to interfere with seroconversion, immunotherapy appears not compromising the humoral response to mRNA-anti-COVID-19 vaccination in cancer patients, yet in these patients the production of autoantibodies has been reported, raising concerns about the risk of developing autoimmune diseases [81]. Autoimmune-related adverse effects have been reported after the third dose of COVID-19 mRNA pro-vaccine (Comirnaty) in cancer patients under treatment with immune-checkpoint inhibitors [82]. These observations caution that introducing mRNA-based immunostimulants along with immunosuppressive therapies in the context of a dysregulated immune system (such as in cancer patients) may have unpredictable consequences. The mRNA anti-COVID-19 vaccination exacerbated pro-inflammatory Th17 immune response along with neutrophilia in oncologic patients, particularly in those recovered from COVID-19 [83]. Since this condition poses the risk of triggering cytokine storm, it requires caution in vaccinating cancer patients with previous SARS-CoV2 infection, and this is of particular concern when multiple boosters are administered. A recent literature review concluded that COVID-19 vaccination was generally well-tolerated, safe, and effective in cancer patients, with rare severe side effects including necrotizing myopathy, thromboembolisms and allergic reactions [84]. Yet, this same study also revealed that protection was moderate and limited in time, since despite the vaccination, cancer patients could get infected, with hospitalization and high risk of mortality, requiring continuous booster doses as preventive measure [84]. Indeed, T-cell response (which is the most important to combat viral infections) in cancer patients was weak even after the third dose [70]. In this regard, concerns have been raised about the possibility that multiple boosters could induce CD8+ T cell exhaustion along with increased expression of PD1 [85]. In line with the above, patients with chronic lymphocytic leukemia showed impaired CD4+ and CD8+ T cell memory response to viral Spike eight months after two doses of the Comirnaty (BNT162b2) mRNA vaccine [86]. Overall, these studies question the assumption that COVID-19 vaccination is beneficial, in terms of protection from COVID-19 infection and clinical outcome and instead raise important concerns about its safety for cancer patients, especially because short-term protection imposes continuous booster vaccination.
3. The Complex Interplay Between COVID-19, Anti-COVID-19 mRNA Pro-Vaccine and Cancer
We may not close this first introductory section without mentioning case reports suggesting a paradoxical effect of the SARS-CoV2 infection and of the anti-COVID-19 vaccination, in combination or alone, associated with a partial and transitory regression of cancer. In a small cohort of cancer patients undergoing checkpoint immunotherapy, an increase in the absolute number of circulating NK cells, not of T and B cells, occurred four weeks after the third dose of the Comirnaty vaccine, and these patients showed a reduced likelihood of disease progression within six months of vaccination [87]. To be noted, this is the same cohort in which one fifth of the high antibody responders to vaccination developed autoimmune thyroiditis [82].
A very recent study shows that certain patients bearing aggressive forms of skin and lung cancer who received COVID-19 mRNA vaccines within hundred days of their immune checkpoint therapy experienced a surge of type I interferon response that enhanced T-cell response and led to improved survival by a few months [88].
In their review, Meo et al describe the clinical cases of nine patients with hematological malignancies (including lymphomas, leukemias, and myelomas) and five patients with solid tumors (two renal tumors and three colorectal tumors) in whom a spontaneous temporary remission (the longest was up to 12 months) was observed following SARS-CoV2 infection [89]. The author’s explanations for this effect include a possible direct oncolytic effect of the virus in infected malignant lymphocytes and the stimulation of T-cytotoxic cells by pro-inflammatory cytokines within the solid tumor microenvironment, though no mechanistic studies were performed to confirm such biological and immunological activities.
In literature there are also three cases of partial cancer regression following the anti-COVID-19 vaccination. In one case, a patient diagnosed with left parotid myoepithelial carcinoma and possible metastatic nodules in the lung received two doses of the Moderna mRNA-1273 COVID-19 vaccine, to which experienced a severe adverse reaction that resolved within two weeks, and over the next 9 months showed a 73% reduction in tumor burden associated with a phenotypic shift in the tumor immune microenvironment from a pro-tumorigenic (characterized by M2 macrophages) to a pro-inflammatory anti-tumor phenotype enriched in CD8+ T cells [90]. In a patient affected by primary cutaneous anaplastic large-cell lymphoma (pcALCL) showing recurrence and multiple lung nodules (suspicious of metastases) after therapy, it was observed a marked regression of the cervical lymph node and lung lesions one week after administration of one dose of Comirnaty vaccine, suggestive of possible causal correlation [91]. The authors, however, correctly mention that pcALCL frequently undergoes spontaneous regression. A third case refers to a patient diagnosed with cutaneous Merkel Cell Carcinoma (MCC) who experienced the regression of an enlarged axillary metastatic lymph node after the third dose of Comirnaty vaccine [92]. As noticed by the Authors, despite being highly aggressive spontaneous regression of MCC is relatively frequent. The latter two cases may be explained with the intrinsic propensity of spontaneous regression of small metastatic lesions possibly favored by vaccine-induced immune stimulation. Finally, there is a case of a patient who recurred hepatocarcinoma four months after partial hepatectomy and six months later showed regression of the hepatic lesion after three doses of Moderna mRNA-1273 and SARS-CoV2 infection [93]. This latter case points to the complex interplay between the host immune response in cancer patients, COVID-19 and mRNA anti-COVID-19 vaccination. These few case reports remain anecdotal and presents important limitations for establishing any causal correlation or generalizability especially when considering the lack of clear mechanistic explanation, insufficient reproducibility, and the larger number of cases where such effect has not been reported or, instead, an opposite effect has been reported as it will be discussed in the next paragraphs.
4. Can SARS-CoV2 and/or Anti-COVID-19 mRNA Pro-Vaccine Cause Cancer? Putting the Puzzle Pieces Together
In this second part, we will address the question whether and how the SARS-CoV2 and the anti-COVID-19 mRNA pro-vaccine can cause cancer or worsen the prognosis of pre-existing cancers.
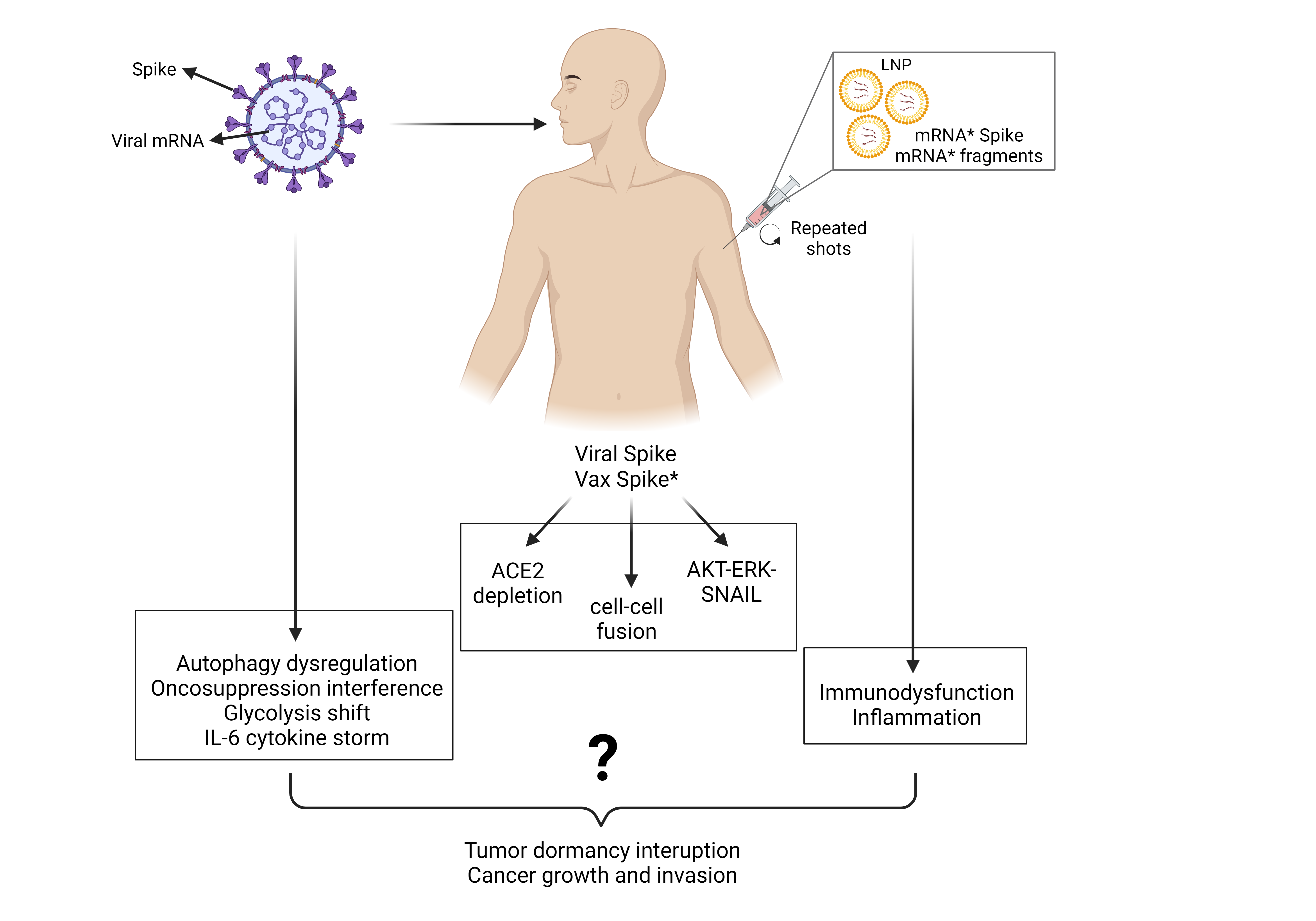
We have learned that cancer is a dynamically evolving proliferative and invasive disease arising from the accumulation of genetic and epigenetic changes in the parenchymal cells whose growth and spread is facilitated by microenvironmental factors. Preneoplastic nodules, micrometastases, and residual disease (after surgical debulking and anti-cancer therapy) may remain stable for decades in a dormant state due to insufficient blood supply (angiogenic dormancy), due to efficient immune-suppression (immune-mediated dormancy), and due to up-regulated autophagy (autophagy-mediated cancer cell dormancy) [94,95,96]. Tissue inflammation is the main cause of dormancy interruption and cancer outgrowth, by promoting neoangiogenesis and immune suppression, while inhibiting cellular autophagy [94,97,98,99]. The role of growth factors and hormones, neoangiogenesis, inflammation and immune-suppressive cells in the microenvironment in the growth of metastases was outlined by Stephen Paget (1889) in his “seed and soil” theory [100]. Another important feature of cancer cells is the alteration of the glucose and amino acids metabolisms so that glucose is preferentially glycolyzed with production of lactic acid while mitochondria utilize preferentially glutamine for the Kreb’s cycle [101].
Therefore, to induce or promote carcinogenesis the virus and the mRNA pro-vaccine must possess one or more of the following abilities: 1. Induce gene mutagenesis; 2. Induce epigenetic changes; 3. Interfere with the oncogenic and tumor suppressor pathways that control cell behavior and fate impinging on cell proliferation and cell migration, autophagy, cell survival and cell death, and energetic metabolism; 4. Induce inflammation, angiogenesis, and lymphopenia in the tissue microenvironment.
While the possibility that the viral genetic code or the reverse-transcribed pro-vaccine mRNA could integrate into the cellular genome and cause gene mutagenesis is deemed extremely unlikely, all other events have indeed been associated with SARS-CoV2 infection and anti-COVID-19 vaccination.
Although some events are uniquely associated with the peculiar individual physical-chemical structure of the virus or anti-COVID-19 mRNA pro-vaccine, we will see that both share the characteristics to trigger the very same events. In particular, the viral Spike protein and that produced by the vaccine mRNA, being structurally very similar, will likely trigger the same reactions. Another major cancer-promoting mechanism shared by SARS-CoV2 infection and anti-COVID mRNA pro-vaccines is inflammation.
5. The SARS-CoV2 Virus and the Cancer
Recent studies pointed out the possibility that SARS-CoV2 infection might create conditions for cancer progression [102,103]. A special and obvious suspect is IL-6 due to its role in COVID-19-associated inflammation [104]. However, as we will see in detail, many other factors and pathways can link SARS-CoV2 infection with cancer, with the Spike protein as the main trigger.
5.1. Oncogenic Potential of SARS-CoV2 Receptors ACE2 and AXL
In SARS-CoV2-infected patients, as well as in anti-COVID-19 mRNA pro-vaccinee, membrane bound and circulating ACE2 levels are decreased due to Spike attack, and this has been linked to inflammation, thrombosis, and hypertension [105,106,107]. Could the depletion of ACE2 have a role in carcinogenesis? ACE2 is a peptidase that can be found as membrane-bound on the surface of endothelial and epithelial cells and as soluble form in the circulation. It has a pivotal role in the Renin-Angiotensin-System, which controls cardiovascular functioning. In brief, the liver secretes in the blood angiotensinogen that is processed to angiotensin I (AngI) by Renin (secreted by the kidneys), and AngI is further processed by the enzyme ACE (angiotensin converting enzyme; expressed particularly, not exclusively, in the epithelial cells of the lungs) into the vasoconstrictor AngII, which can eventually be processed by ACE2 into the vasodilator peptide Ang1-7. Thus, while AngII favors hypertension, Ang1-7 contrasts hypertension, dampens inflammation, and prevents thromboembolisms [107]. More than that, AngII has mitogenic and angiogenic activities and inhibits cancer cell apoptosis, whereas Ang1-7 inhibits angiogenesis and cancer growth [108,109]. Consistently, ACE2 was reported to inhibit angiogenesis and to prevent metastasization in breast and lung cancer models [110,111,112]. ACE2 deficiency was shown to increase the risk of hepatocarcinogenesis and the resistance to anti-PD-L1 immunotherapy, while promoting a permissive tumor microenvironment associated with M2-like macrophages, angiogenesis, and immunosuppressive myeloid cells [113]. SARS-CoV2 promoted Epithelial-to-Mesenchymal Transition (EMT) of infected lung cancer cells, associated with high expression of ZEB1 and AXL and decreased expression of membrane ACE2 [114].
In benign mammary epithelial cells transgenically expressing ACE2, the challenge with SARS-CoV2 Spike protein induced the transcription of SNAIL and acquisition of a migratory and invasive mesenchymal phenotype [115]. Further, the hyperglycosylated Spike protein from SARS-CoV2 gamma variant was shown to induce SNAIL-mediated EMT and to promote in vivo metastasization of xenografted human breast cancer cells [116].
Thus, depletion of ACE2 results in the loss of an anti-cancer barrier against the growth and spread of pre-existing (micro)tumors, favoring metastasization [117,118,119]. In this context, it is worth mentioning that ACE2 can be targeted by MDM2 (mouse-double-minute 2) and thereafter ubiquitinated and degraded via proteasome [120]. To be noted, MDM2 is considered an oncogenic protein since it can direct the proteasome degradation of TP53, a major tumor suppressor protein (see 5.3). It is tempting to speculate that Spike-induced ACE2 depletion might leave MDM2 free to bind and direct the degradation of TP53 further increasing malignancy (more in 5.3).
Another receptor for SARS-CoV2 potentially linking COVID-19 to cancer is AXL (Anexelekto). This is a transmembrane receptor protein (its physiological ligand is GAS6) that plays an important role in cancer progression, as its activation promotes cell proliferation, EMT and metastasization [121]. It is worth noting that ACE2 and AXL are involved in other cancerogenic pathways, as will be explained in detail below.
5.2. SARS-CoV2 Spike Protein Can Trigger Oncogenic Signaling Pathways
Human lung carcinoma A549 cells (type II pneumocyte) incubated with SARS-CoV(1)-like particle or its isolated Spike showed the Casein Kinase II-mediate phosphorylation of ACE2 and the activation of a Ras-ERK (extracellular regulated kinase)-AP1 pathway [122]. More recently, the S1 subunit of SARS-CoV2 Spike protein was shown to trigger the ERK signaling in lung endothelial cells, and this effect was not mediated by the interaction with ACE2 [123]. In lung carcinoma A549 and in hepatocarcinoma Huh-7.5 cells, the SARS-CoV2 Spike activated the MAPK-NF-κB pathway and downstream induction of IL-6 synthesis [124]. In lung epithelial cells, SARS-CoV2 was shown to activate the Epidermal Growth Factor Receptor (EGFR)-AKT survival signaling pathway along with stimulation of mitochondrial ATP production [125]. This mechanism is believed to help the virus to sustain its replication by keeping alive and boosting the energy metabolism of the infected cell in the early phase of infection.
The (Ras-)ERK/MAPK and the AKT pathways drive transcription, protein synthesis, cell proliferation and cell survival, and are hyperactivated in cancer cells [126,127].
Additionally, the Spike proteins was shown to interact with the Estrogen receptor and induce ERα-dependent cell proliferation of breast cancer cells [128].
Finally, an in-silico study found that the viral Spike protein potentially interacts with and activates the EGFR and VEGFR pathways [129].
Whether the above signaling triggered by viral Spike could result in aberrant survival and stimulation of proliferation and migration of infected pre-neoplastic cells has not been investigated yet, and it cannot be excluded.
5.3. SARS-CoV2 Spike Protein Can Inactivate Tumor Suppressor Signaling Pathways
TP53 and BRCA1/2 are two major tumor suppressor proteins that play a major role in cancer progression and therapy resistance [130,131]. TP53 (p53) has nuclear and cytosolic functions: in the nucleus, as a homo-tetramer it binds the DNA to direct the transcription of genes that regulate cell cycle, DNA repair, apoptosis, autophagy and cell metabolism; in the cytoplasm, as a monomer, it directs the BAX oligomerization on the outer mitochondrial membrane and lysosome membrane for inducing cell death [132]. Certain p53 mutants unable to bind the DNA may act as “dominant negative” and impair apoptosis and autophagy [133].
It was previously found a complex interplay between p53 and the SARS-CoV: p53 was shown able to inhibit viral replication and, on the other hand, the virus can promote the ubiquitination and degradation of p53 [134]. Given the similarity between the domains involved between the SARS-CoV and SARS-CoV2 it is reasonable to hypothesize that also the latter have a similar relationship with p53.
In this context, an in-silico study found that the C- terminal domain of the heptic repeat-2 region of S2 subunit (which plays a role in membrane fusion) has the potential to bind p53, BRCA-1 and BRCA-2 proteins [135]. Should this interaction be confirmed it would open a dangerous scenario. In fact, the possible sequestration of these proteins by the S2 Spike protein would have catastrophic consequences in the cell because of the loss on the control of genome integrity and cell behavior.
Very recently, Zhang and El-Deiry [136] have tested this hypothesis in various cancer cell lines in which the SARS-CoV2 protein was transgenically expressed. Co-immunoprecipitation did not confirm the interaction between S2 and p53, likely because the two proteins reside in different compartments (cytosol and nucleus, respectively). However, these authors found that the exogenous expression of the Spike protein attenuated the transcriptional activity of p53, and this was not due to the MDM2-mediated degradation of p53 [136]. Of note, when the cancer cells were treated with the DNA damaging chemotherapeutic drug cis-platinum, the Spike-expressing cells could not transcribe p21 for blocking the cell cycle and to induce cell death.
When the AKT pathway is activated by growth factor receptors, p53 is degraded by the proteasome via MDM2 and this is more likely to occur when ACE2, an alternative substrate of MDM2, is less abundant in the cell. SARS-CoV2 has been shown to trigger the EGFR-AKT pathway and to deplete ACE2 (see 5.3 and 5.4), a combination that could favor p53 degradation.
5.4. Spike Protein Induces Cell-to-Cell Fusion: A Step Toward Cancer Transformation?
Cell-cell fusion is a well-known phenomenon characteristic of cancer leading to hybrid cells with mixed behavior due to the combined contribution of not only the nuclei but also of cytoplasmatic organelles, among which mitochondria and lysosomes may play a major role [137]. Within the tumor microenvironment, cancer cells may increase their malignant potential by fusing with mesenchymal stem cells, fibroblasts, and macrophages [138]. In addition, the formation of such syncytia within the tumor microenvironment could promote the immune evasion of tumor cells following the entrapment of T and NK cells (forming "cell-in-cell structures"), thus leading to lymphopenia [139]. Cell fusion is relatively rare in spontaneous tumors, yet this event could be promoted by SARS-CoV2 infection. The presence of the polybasic furin-sensitive site enhances the fusogenic property of the S2 subunit of the Spike protein, which can lead to the formation of syncytia [139]. This happens when a SARS-CoV2-infected cell exposes on the membrane the Spike protein that interacts with the ACE2-TMPRSS2 or ACE2-AXL-NRP1 proteins expressed on a neighboring cell [140,141]. This process is believed to enhance cell-to-cell infectivity.
Worthy of note, hydroxychloroquine, an inhibitor of endosomal-lysosomal acidification and of autolysosome formation, was found to inhibit this process [139], which gives credit to the use of this drug for the prophylaxis of COVID-19 [142].
Given that ACE2 and AXL are highly expressed in cancer epithelial cells and in stromal/mesenchymal cells, the ability of Spike protein to promote the fusion of neighboring cells expressing these proteins raises concerns about the possible formation of hybrid tumor cells with increased metastatic potential [143,144].
5.5. SARS-CoV2 Replication Dysregulates Autophagy: A Step Toward Carcinogenesis?
Autophagy is a lysosomal-driven degradation process that eliminates damaged and redundant subcellular structures and maintains tissue homeostasis, keeping under check cell proliferation and cell migration [145]. Autophagy is meant to entrap within the autophagosome (a double-membrane vesicle arising from the endoplasmic reticulum) any cytosolic protein agglomerate and organelle that perturbs cell homeostasis and direct their degradation by fusing with lysosomes, acidic organelles endowed with a wide range of hydrolytic enzymes [145]. Perturbation of the autophagy pathway has negative impacts on cell homeostasis and might favor the malignant phenotype [146]. To be noted, dysregulated autophagy in cancer cells may instead favor survival against anti-cancer therapies [147]. As we will see, dysregulation of autophagy occurs in SARS-CoV2-infected cells.
Autophagy is involved in coronavirus infection, replication, and viral spreading. SARS-CoV2, alike many other viruses, can exploit this vesicular process for its own replication and egression from the cell [148,149]. Particularly, the nonstructural proteins NSP15 and NSP6 can hijack the autophagy pathway so that the former induces the formation of autophagosomes while the latter alters the acidification of the lysosomes impairing their fusion with autophagosomes [148]. In so doing, the virus will escape the lysosomal degradation and instead will divert the nascent autophagosomes versus the formation of double-membrane vesicles for its assembly [34,148]. Another study found that SARS-CoV2 ORF10 localizes at the mitochondria where it binds to mitochondrial antiviral signaling protein and directs its degradation via mitophagy [150]. In SARS-CoV2 infection, autophagy plays an important role in protecting cells from death [151], for instance it protects from pyroptosis infected immune cells by degrading the inflammasome [36,37]. Finally, it is worth mentioning that FYCO1 (FYVE and coiled-coil domain autophagy adaptor 1), one of the SARS-CoV2 infection susceptibility genes [55], encodes a RAB7 adaptor involved in autolysosome formation, and it is considered a novel oncogene in that it promotes EMT and migration in breast and cervical cancer cells [152,153].
5.6. SARS-CoV2 Alters Mitochondrial Respiration and Induces Oxidative Stress
A link between glucose metabolism and SARS-CoV2 infection emerged with the observation that uncontrolled glycemia was a risk factor for COVID-19 [154]. An important feature of cancer cells is the altered metabolism of glucose known as the Warburg effect by which cancer cells avidly uptake glucose that is fully glycolyzed in the cytosol (to provide substrates for nucleoside synthesis) instead of being completely oxidized via mitochondrial respiration [155]. This divergence in the glucose metabolism is directed by the oncogenic transcription factor HIF-1α (Hypoxia-Inducible Factor-1α), that besides the genes of the glycolytic pathway, also transcribes, among others, genes involved in angiogenesis (VEGF, vascular endothelial growth factor) and cell motility (HGF, hepatocyte growth factor), inflammation, and tumor microenvironment remodeling [156].
It has been reported that SARS-CoV2 induces the glycolytic shift in infected lung macrophages [157]. Mechanistically, the SARS-CoV2 ORF3a induces mitochondrial ROS production that stabilizes HIF-1α, which then promotes glycolysis [158]. A similar glycolytic shift might occur also in SARS-CoV2-infected epithelial cells, and this would be an add on in the case of pre-neoplastic cells.
To close the circle, IL-6, which drives the cytokine storm in COVID-19, induces the glycolytic shift in cancer cells and promotes the phenoconversion of stromal fibroblasts into permissive cancer-associated fibroblasts through inhibition of autophagy [159,160].
In cancer cells, there is an interplay between the altered glucose metabolism and the mitochondrial respiration [161]. Dysfunctional mitochondria produce oxidative radicals (ROS) that can trigger the inflammasome with production and secretion of inflammatory cytokines. The role of ROS in cancer development and progression depends on how much they are produced: low to moderate levels trigger cell proliferation and migration, high levels of ROS damage the proteins, membranes, and DNA and induce cell death [162]. To be noted, ROS may induce or inhibit autophagy (and mitophagy, particularly) with opposite consequences in tumorigenesis and metastasization [163].
SARS-CoV2 infection has been shown to affect mitochondrial respiration. Disruption of mitochondrial morphology and functioning with over production of ROS in peripheral leukocytes and muscles is a common finding in COVID-19 patients [164]. Thus, SARS-CoV2 infection of pre-malignant or dormant tumor cells could elicit such effects triggering tumor growth and invasion.
5.7. SARS-CoV2 Triggers the Inflammatory Cytokine Storm and Induces Immune Cell Depletion Leading to a Microenvironment Permissive for Relapses and Metastasis
SARS-CoV2-infected monocytes and macrophages isolated from the lungs of COVID-19 patients produce huge amounts of pro-inflammatory cytokines such as IL-1β, TNF-α, IL-6, and IFN α, β, and λ, which preludes the so-called cytokine storm and meanwhile abates the T-cell immune response [157]. This production of cytokines was directed by HIF-1α in consequence of mitochondrial ROS production and could in fact be prevented by antioxidants such as N-Acetyl Cysteine [157]. To be noted, the secretion of SARS-CoV2-infected monocytes, and particularly IL-1β, inhibited the proliferation of CD4 and CD8 T cells and increased the surface expression of PD-1 in CD4 lymphocytes, indicative of immune cell exhaustion [157]. These findings are consistent with the reported association between the cytokine storm (particularly IFN-γ and TNF-α) and lymphopenia [165,166]. In infected patients, hyper-production of proinflammatory cytokines (particularly, IL-6, TNF-α, and IFN-γ) may follow the accumulation of undegraded angiotensin II and overactivation of its type 1 receptor (AT1R) because of Spike-induced downregulation of ACE2 [167].
Pro-inflammatory cytokines recruit myeloid-derived suppressor cells that create microenvironment permissive for tumor growth and development of metastases by inhibiting the T cell anti-tumor immune response [168].
Further concern is that the inflammatory response and immune system dysregulation associated with SARS-CoV2 could create the conditions for the awakening of dormant tumor cells [169,170]. Intriguingly, AXL, the receptor for Spike SARS-CoV2 highly expressed in cancer cells, has been shown essential for TGF-β2-induced dormancy of metastatic cancer cells [171]. Adding to the complexity, the GAS6-AXL axis induces autophagy in macrophages inhibiting the activation of the inflammasome and the release of IL-1β and IL-18, thus mitigating inflammation [172]. IL-6, the main culprit of cytokine storm [173], is a pro-tumorigenic cytokine as it promotes cancer cell proliferation and migration, interrupts cancer cell dormancy, and worsens the prognosis of cancer patients through inhibition of autophagy [97,174].
It is unknown whether these scenarios can occur in the tumor context of patients infected with the virus, an aspect that deserves to be investigated in depth. The observation that the risk of cancer-related mortality and lung metastasis is higher in SARS-CoV-2-infected patients and the demonstration that IL-6 associated with SARS-CoV2 infection can awaken dormant breast carcinoma cells metastasized in the lung of mice support this possibility [175].
6. The Anti-COVID-19 mRNA Pro-Vaccine and the Cancer
Both the anti-COVID-19 mRNA pro-vaccines BNT162b2 and mRNA-1273 consist of lipid nanoparticles containing the full-length mRNA (of 4284 and 4004 nucleotides, respectively) encoding the Spike protein; however, they differ in the 5′ and 3′ untranslated regions, the total amount of mRNA (30 µg/0.3 ml and 100 µg/0.5 ml, respectively), and the chemical composition of the lipid nanoparticles (the cationic ionizable lipid is ALC-0315 and SM102, respectively) [176]. To prevent the prompt degradation of the mRNA pro-vaccine within the transfected cells, all the uridines have been replaced by N1-methyl-pseudouridine, and a number of codons have been modified in their third nucleoside to optimize translation efficiency [176]. The vaccine Spike is identical to the viral protein (it has the furin-sensitive cleavage site for splitting into the S1 and S2 subunits) except for the substitutions of the amino acids 986K and 987V with two prolines to fix the protein in the pre-fusion form [176]. Though stabilized in the prefusion conformation, the vaccine Spike can bind to ACE2 and be cleaved by furin [177]. The above modifications may explain why the mRNA and the intact protein or fragments of the vaccinal Spike persist in the circulation of the vaccinee for long time and can be found in organs distant from the injection (deltoid) site [178,179,180,181]. Additionally, the vaccinal mRNA and Spike may travel throughout the body with the exosomes [182], which increases the risk of triggering epiphenomenal reactions associated with severe adverse effects in various organs [107,183,184]. The mRNA vaccine technology relies on the endogenous synthesis of the immunogen (in this case, the Spike protein) that is further processed by antigen processing cells to instruct the lymphocytes for producing neutralizing antibodies and to mount a T cell immune response [185].
The facts that the exogenous protein driven by a modified mRNA is synthesized within the host cells and that portions of it or of its fragments (since it can be cleaved by furin) can be exposed on the membrane of any cell (since the mRNA is delivered via lipid nanoparticles) increase the risk of deceiving the immune system, which adds on the tissue damages brought by the Spike-ACE2 interaction. The serious and sometimes (fortunately rare) fatal adverse effects associated with mRNA COVID-19 vaccination have been covered in other articles [107,183,184,186,187] and are not the subject of this article as here we focus on the potential pro-carcinogenic effect of the COVID-19 vaccination with such products.
There are several mechanisms and pathways that could link the mRNA anti-COVID-19 vaccination with an increased risk of cancer progression, some of which are common with those connected with SARS-CoV2 infection (the Spike; the inflammatory cytokines) and others that are unique to the mRNA pro-vaccine being associated with its peculiar composition (the presence of pseudouridine; the presence of impurities such as truncated mRNAs and traces of DNA; the presence of inflammatory cationic lipids) and with the vaccination schedule that comprises several shots in a too short time. The latter has implications as it exposes the vaccinated person to a greater risk of infection, thus facilitating exposure to the side effects of SARS-CoV2 described above.
Unlike the viral spike protein, the vaccine spike protein has not been associated with dysregulation of autophagy and energy metabolism. However, other factors may link the vaccine Spike protein to processes that potentially increase the risk of carcinogenesis as we will illustrate in the next paragraphs.
6.1. The Vaccinal Spike Displays Pro-Carcinogenic Properties Like Viral Spike
The vaccinal Spike shares very similar structural characteristics with viral Spike in terms of binding to surface receptors and thus triggering similar pathways.
In brief, recalling what has been documented for the viral counterpart, vaccinal Spike has the potential to: (i) deplete membrane-bound and soluble ACE2 (see 5.1); (ii) trigger the oncogenic ERK/MAPK, EGFR-AKT, AXL, and SNAIL-TGFβ pathways (see 5.1 and 5.2); (iii) interact with ERs in breast cancer cells (see 5.2), (iv) interfere with tumor suppressor TP53 stability and transcriptional activity (see 5.3); (v) induce the formation of syncytia (see 5.4). The consequent effects of these actions include the promotion of cell proliferation and cell migration, induction of EMT, and inhibition of cell death, as discussed in detail in paragraph 5.
Importantly, the brief protection induced by mRNA pro-vaccines against COVID-19 requires frequent and closely spaced vaccinations, resulting in a tolerogenic immune response and subsequent increased susceptibility to SARS-CoV-2 infection, which creates the conditions under which these non-genotoxic pro-carcinogenic pathways are likely to be activated.
6.2. Molecular, Biochemical, Genetic and Epigenetic Effects of the mRNA Pro-Vaccine: Hypothesizing the Unpredictable
Due to patent protection, data on the manufacturing technology and quality control of COVID-19 mRNA pro-vaccines are scarce [176]. What follows is therefore inevitably theoretical and based on the limited information available.
T7 RNA polymerase-directed in vitro transcription of a DNA template yields the desired RNA, but it also has some drawbacks such as the generation of unwanted RNA species, including double-strand RNA and a mixture of short abortive transcripts of various length. The presence in the market of different batches of vaccine with different composition due to non-standardized manufacturing and quality control remains controversial, denied by some studies and confirmed by other studies 176, 188. In this regard, some batches of BNT162b2 were found to contain on average only 50% of intact Spike-coding mRNA, the rest being fragments of various length [188,189]. These fragments could theoretically impair the synthesis of target proteins functioning as sponges for a variety of cellular mRNAs. However, the sequence of these fragments has never been disclosed, and therefore their possible interference on the translation of cellular mRNAs remains speculative.
The replacement of uridines with N1-methylpseudouridine deceives the reading machinery in the translation of the mRNA into protein, causing frameshifts in the reading of the codons with the consequence of synthesizing unintended proteins [190] that might have unpredicted consequences [191]. Quite reassuring, a synthetic mRNA resembling the BNT162b2 Spike-coding mRNA with 100% N1-methylpseudouridine was found to translate into intact Spike protein when expressed in HEK293 cells [192]. In vivo the story goes differently. N1-methylpseudouridine containing mRNAs do not efficiently stimulate dendritic cells, with reduced production of type I Interferon (which exerts anti-cancer functions) and decreased T cytotoxic activity, and this may be relevant for the antitumor immune response [193]. Consistently, BNT162b2 vaccination modulated the innate immune responses by increasing the production of inflammatory cytokines IL-1β and IL-6, while decreasing that of IFN-α [194]. Such immune suppressive and tumor permissive scenario was reported in an in vivo (OVA-expressing) melanoma model of cancer vaccination with N1-methylpseudouridine modified mRNA encoding the transgenic tumor antigen (OVA, ovalbumin) encapsulated in lipid nanoparticles [195]. While the unmodified mRNA OVA vaccine elicited antitumor effects characterized by robust infiltration of CD40+ DCs and OVA-specific IFN-γ secreting T cells, the vaccination with pseudo-uridine modified mRNA greatly decreased immunogenicity (decreased IFN-γ production and TNFα-producing CD8+ T cells) in spite of the highest translation efficiency, resulting in increased tumor growth and number of lung metastases [195].
RNA Adenosine-to-Inosine editing is a co-transcriptional process catalyzed by adenosine deaminase ADAR1 acting on double-stranded portions of immature RNA and potentially resulting in transcriptome and proteome changes, as Inosine is read as Guanosine. RNA editing may involve coding and non-coding regions and can affect stability, alternative splicing, and translation of mRNAs as well as the processing and specific targeting of non-coding RNAs. Up-regulation of ADAR1 expression and overall increase in RNA editing have been associated with the malignant phenotype [196]. A recent study showed that the expression of ADAR1 in the blood of vaccinee increases with the number anti-COVID-19 mRNA pro-vaccine doses [197]. While this observation is not sufficient to establish a possible cancer link, it is somewhat concerning that among the top three genes with significant A-to-I editing is Slingshot protein phosphatase (SSH), a cofilin phosphatase known to promote cancer invasiveness and metastasis [198].
Another concerning issue for the COVID mRNA pro-vaccine is that the optimization with enriched guanosine-cytosine (GC) and N1-methylpseudouridine may favor the formation of tetrads of guanine called G4 (G quadruplex) that are known to destabilize DNA and are frequently found in cancer [199,200]. G quadruplex is a preferential target of the Polycomb Repressor Complex II, that exerts epigenetic control of gene transcription [200]. However, to elicit possible damaging effect on the transcription and DNA repair machineries the G-rich mRNA (fragments) of the pro-vaccine should relocate in the nucleus of the cell. This eventuality seems very unlikely, although it cannot be excluded.
There is however another issue: following a modification in the manufacturing procedure (that now makes use of DNA plasmid instead of PCR to produce Spike mRNA) trace DNA impurities have been found in the BNT162b2 pro-vaccine mRNA [201], although the biological significance of this finding is still unknown and deserves further investigation.
6.3. Disruption of the Immune Surveillance and Induction of Inflammation: Creating the Condition for Awakening the Dormant Tumor
In chapter 5.7 we have discussed the molecular and cellular mechanisms through which SARS-CoV2 could interrupt tumor dormancy. With respect to SARS-CoV2 infection, the injection with LNP-mRNA pro-vaccine brings additional stress to tumor microenvironment for the following reasons: 1. repetitive vaccination shifts the immunogenic response toward a tolerogenic and pro-inflammatory response and overall suppression of the immune response; 2. the lipid component of the nanoparticle is strongly inflammatory.
Due to short term of protection by the anti-COVID-19 mRNA pro-vaccine, repeat vaccination has been recommended on average every 6 months. However, repetitive boosters with Spike mRNA pro-vaccines modulate the adaptive immune system determining a shift from immune to tolerogenic response. After three doses a class switch of immune reactive IgG1 and IgG3 versus the tolerogenic IgG4 was observed in almost half of the vaccinee [60]. A very recent study showed that in children the serum level of anti-Spike IgG4 continues to raise up to one year after the second dose of Comirnaty [202]. Experiments of vaccination with mRNA coding for the receptor binding domain of the SARS-CoV2 Spike in mice confirmed that repetitive boosters determine a condition of humoral and cellular immune tolerance [203]. Worthy of note, a literature search and meta-analysis found that high levels of IgG4 increased the risk of developing cancer, particularly pancreatic cancer and lymphoma [204]. Local concentration of IgG4, regardless of the antigen specificity, has been shown to drive immune evasion in the tumor microenvironment by inhibiting IgG1-mediated cancer cytotoxicity [205].
T-cell immunity plays a major role in anti-cancer response as well as in keeping dormant (immunogenic dormancy) micrometastases [206]. Unfortunately, multiple vaccinations with anti-COVID-19 mRNA pro-vaccines have been shown to cause T-cell exhaustion and increased expression of PD-1 [85]. A Phase II study reported a transient lymphopenia in some 50% of the vaccinee with one dose (30 or 100 µg) of BNT162b1, and in 33% of those who received the highest dose (100 µg) lymphopenia was of grade 3 [207]. Further contributing to interruption of the dormant-associated immune tumor microenvironment is that these mRNA pro-vaccines can trigger a strong inflammatory response with elevated levels of circulating IL-17 [208] and, particularly in cancer patients previously infected with SARS-CoV2, a shift of memory T-cell toward pro-inflammatory IL-17+ CD8 [83]. IL-17 is known to promote cancer cells proliferation besides impairing T-cell mediated anti-tumor response [209].
Increased levels of circulating cytokines (among which IL-6 and IL-17) and growth factors (among which VEGF and bFGF) may be detected in vaccinees up to one year after vaccination with anti-COVID-19 mRNA Comirnaty [210]. Hypothetically, these cytokines and growth factors could interrupt autophagy-mediated [97,174] and angiogenic-mediated [211] tumor dormancy.
Further contributing to an inflammatory potentially tumorigenic microenvironment is the LNP component, which is said to function as an immune stimulator adjuvant. The cationic LNP component of the mRNA pro-vaccine was shown to induce the release of inflammatory cytokines (mainly, IL-6, TNFα and IL-1β) by macrophages and to activate the serum complement via the alternative pathway [212]. This could explain the so-called “radiation recall phenomenon” shown to occur in cancer patients few days after the second dose of the BNT162b2 mRNA pro-vaccine [213].
Overall, continued vaccination with these COVID-19 mRNA pro-vaccines impairs the innate and adaptive immune system and sustains an elevated inflammatory state with IL-6 and IL-17 overproduction, along with inhibition of autophagy and stimulation of AXL and VEGFR pathways that altogether are conducive to awakening of dormant tumors and cancer progression.
7. Data from the Real World: Case Reports Linking Anti-COVID-19 mRNA Vaccination and Cancer
Although supported by the whelm of data in the literature, the mechanisms and pathways illustrated above are only indicative of the carcinogenic potential of COVID-19 mRNA-based pro-vaccines. There are no specific research focusing on cancer prognosis and fatal events in vaccinated cancer patients in relation to COVID-19, except one study reporting the causal correlation in two out of three patients [68].
How about data from the real world? Recently, a population-based retrospective study in a large cohort of unvaccinated (595,007) and vaccinated (2,380,028) individuals in Seoul (South Korea), where the cumulative incidences and corresponding Hazard Ratio of cancers were measured one year after COVID-19 vaccination, found an association between vaccination and increased risk of thyroid, gastric, colorectal, lung, breast and prostate cancer [214].
In absence of an active pharmacovigilance to collect data specifically addressing the possible correlation, if not causation, between COVID-19 mRNA vaccination and cancer, we rely on case reports in literature.
Table 1 and Table 2 summarize the known cases, respectively referring to the anti-COVID-19 mRNA pro-vaccines from Pfizer and Moderna (Table 1) and to other types of anti-COVID-19 genetic vaccines (Table 2). Although when taken individually the clinical cases can be dismissed as anecdotal facts, when considered altogether a series of reports converging on the same conclusion should trigger the suspicion and stimulate a discussion in the scientific community.

Table 1.
Case reports relating cancer to mRNA COVID-19 pro-vaccines.
| DISEASE (onset) |
CLINICAL FEATURES | HYSTOLOGICAL-BIOLOGICAL FEATURES |
TYPE OF VACCINE | Ref |
|---|---|---|---|---|
| Angio-immunoblastic T-lymphoma (onset 6 months after 2nd dose) | 66-y, man; presented with lymphadenopathies; increased number, size, and metabolic activity (of lymph nodes 8 days after 3rd dose) | Gene mutations: RHO, TET2, DNMT3A, IDH2 | BNT162b2 (March, April, Sept 2021) |
[215] |
| Recurrence of axillary lymphoproliferative disorder (2 days after 1st dose) |
79-y, man; in remission from a primary cutaneous anaplastic large-cell lymphoma cured two years before; presented with ulcerated tumor with surrounding erythema | CD-30 positive lymphoproliferative disorder; TCR gene rearrangement matching the previous 2019 clone | BNT162b2 | [216] |
| Nodal Marginal zone B-cell lymphoma (sudden appearance of temporal mass the day after 1st dose) | 80-y, woman; presented with multiple (n. 12) lymphadenopathies at week 6 from 1st dose (week 3 from 2nd dose); increased number (>22) and size (2.5x) in ten weeks | Lymphoid cells positive for CD20, CD79a, and BCL-2; negative for CD3, BCL6 | BNT162b2 (2 doses, 3 weeks apart) |
[217] |
| Diffuse large B-cell non-Hodgkin lymphoma (cervical mass appearance one week after 2nd dose) | 58-y woman; presented with tumor mass at the angle of the left parotid gland progressively growing from June to September with multiple reactive lymph nodes, and finally operated in October 2021 | Confirmed DLBC NHL positive for CD20, PAX5 and negative for CD30, AE1/AE3; 85% Ki-67 positivity | BNT162b2 22 May; 12 June 2021 |
[218] |
| Extranodal malignant non-HodgkinT/NK-cell lymphoma (ulcerative lesions appeared 3 days after 1st dose) |
53-y man; presented (December 2021) with multiple ulcerative oral lesions appeared shortly after the 1st dose which worsened after the 2nd dose | Tumor proliferation with T cells positive for CD3 and CD7, granzyme B, CD30; negative for CD4, CD8, and CD20 | BNT162b2 6 November; 28 November 2021 |
[218] |
| A) Acute lymphoblastic leukemia (two days after 1st dose of mRNA vaccine); B) Recurrence of B-Acute lymphoblastic leukemia (after 1st dose of mRNA vaccine); C) Recurrence of Acute myeloid leukemia (after the booster with BNT162b2) |
A) 49-y woman; presenting with petechiae and bicytopenia, diagnosed with B-ALL; B) 47-y woman; two years before diagnosed with B-cell lymphoma in remission in the last 14 months; C) 67-y woman; diagnosed with AML in 2007 and in remission in the last 14 years after bone marrow transplant. She had two doses of inactivated SARS-CoV2 vaccine in July 2021 and mRNA BNT162b2 in September 2021 |
A) B-ALL: bone marrow showed 20-30% stained with CD19 diffuse positive TdT in blastic cells; B) Bicytopenia and blasts; C) 90% blasts |
BNT162b2 | [219] |
| Four cases of acute myeloid leukemias, one of which extramedullary | A) 61-y man; 30 days after 3rd mRNA dose; B) 28-y woman; 2 weeks after 2nd dose; C) 72-y man; 5 weeks after the 5th dose; D) 60-y man; 1 month after the 4th dose |
A) 80% blastic infiltration; B) bicytopenia; blastic infiltration; C) pancytopenia; 70% blastic infiltration; D) occipital granulocytic sarcoma of CD34, CD123, and MPO positive immature cells; 30% myeloid blasts |
BNT162b2 | [220] |
| Diffuse large B-cell lymphoma (lymphadenopathy was observed one day after the 1st dose) | 67-y man; presented with 6 cm subcutaneous lymphadenopathies mass in the left axilla 2 weeks after the 2nd BNT162b2 vaccination | Large, atypical lymphocytes were positive for CD20, BCL2 and MUM-1/IRF4; negative for CD3; over 80% Ki-67 positivity | BNT162b2 (2 doses) |
[221] |
| Diffuse large B-cell lymphoma (lymphadenopathy was observed two days after the 1st dose) | 80-y woman; presented with enlarged 4.1 cm axillary nodule that developed 1 day after the 2nd dose; two months later the nodule increased to 6 cm and additional lesions appeared in the mesentery and the left cavernous sinus | Germinal center B-cell DLBC lymphoma positive for CD20, BCL6, BCL2; negative for CD3 and MUM-1/IRF4; over 90% Ki-67 positivity | BNT162b2 (2 doses) |
[221] |
| Primary cutaneous anaplastic large cell lymphoma (10 days after the 3rd dose) | 76-y man; presented a fast-growing lesion at the site of the injection 10 days after the 3rd dose. A large erythematous tumor of 6 cm diameter was diagnosed 1 month later. Spontaneous regression after 6 weeks | Anaplastic large cell lymphoma T1bN0M0; positive for CD30, CD4, CD2, CD5, MUM1, and negative for CD20, CD8, TIA1, ALK, EMA, CD56, CD123 and CD68 | BNT162b2 (1st and 2nd dose) Moderna mRNA-1273 (3rd dose) |
[222] |
| High grade sarcoma | 73-y, woman; history of angiomyolipoma in 2019; presented with swelling 2-4 days after 2nd dose developed in 6 cm diameter soft mass in the right upper arm | Grade 3, stage IIIA undifferentiated, pleomorphic high-grade sarcoma | Moderna mRNA-1273 (2 doses) |
[223] |
| Primary cutaneous lymphoproliferative disorders | Series of 14 cases, of which 6 classified as relapse and 8 as primary lesions; complete and partial remission within the 19 months follow-up | N.A. | BNT162b2 | [224] |
| Non-Hodgkin lymphoma (few weeks after the 3rd dose) | 66-y, man; presented with right axillary lymphadenopathy developed 10 days after the 3rd dose, which grew up to 7 cm in the following 3 months | Stage-II anaplastic large-cell lymphoma, ALK negative and CD30 positive, over 90% Ki-67 positivity | BNT162b2 (January, February, October 2021) |
[225] |
| Conjunctival classic Kaposi sarcoma (few weeks after vaccine booster) | 75-y, woman; with complex ophthalmologic history that includes, among others, uveitic glaucoma OU, epiretinal membrane OU, and cystoid macular degeneration OS, presented with irritated conjunctival area |
Conjunctival epithelium shows early squamous metaplasia and positive immunostaining with HHV8 within the CD34 positive vascular proliferation | BNT162b2 (three doses) |
[226] |
| Basaloid carcinoma, wrongly cured as Bell’s palsy for almost 8 months (symptoms appeared 4 days after 1st dose) | 56-y man; no previous health problems; presented with a massive and aggressively infiltrating basaloid-featured cancer in the right side of his face that rapidly progressed and led the patient to death. CT scan (11 months after vaccination) revealed the presence of infiltrating tumor masses in the parotid gland, likely of cutaneous origin | D-dimer value was 1523 ng/ml (normal range is < 500 ng/ml). Biopsy confirmed the diagnosis of basal cell carcinoma |
BNT162b2 (one dose) |
[227] |
| Philadelphia-positive B-cell acute lymphoblastic leukemia (five days after the booster vaccination with bi-valent mRNA vaccine) | 43-y woman; insignificant previous medical history; presented with splenomegaly, severe anemia and thrombocytopenia along with leukocytosis (1.0% neutrophil, 9.0% lymphocyte, 0% monocyte, eosinophil and basophil, and 90.0% blast) | Bone marrow shows 68% blastic infiltration; cells were positive for CD34 and TdT, negative for CD117 and MPO. The p190 BCR-ABL1 gene rearrangement was identified by RT-PCR | Five vaccinations as follow: two doses of Oxford/AstraZeneca (4 June and 31 August 2021); half-dose of Moderna mRNA-1273 (15 January 2022), NovaVax (15 July 2022), and booster dose of the bivalent (Omicron BA.4/BA.5–containing) mRNA-1273 COVID-19 vaccine (January 2023) plus SARS-CoV-2 infection on 19 August 2021. | [228] |
| Epstein–Barr virus-positive marginal zone lymphoma (EBV + MZL) at autopsy (17 days after 1st vaccination) | 71-y woman; with history of methotrexate-treated rheumatoid arthritis; died due thrombosis and multi-organ failure 17 days after vaccination. The autopsy revealed systemic lymphadenopathy comprising atypical lymphocytes and scattered Hodgkin/Reed–Sternberg (H/RS)-like cells | Atypical lymphocytes were positive for CD79a, CD19, EBV-encoded small RNA and MUM-1 and negative for CD3, CD5, CD10, BCL6. H/RS-like cells were positive for CD3 | Unspecified the type of anti-COVID-19 vaccine | [229] |
| Intravascular large B-cell lymphoma at autopsy (105 days after the second dose) | 61-y woman affected by systemic lupus erythematosus recovered 1 month after vaccination for joint pain, clonic spasms, left-sided paralysis, and fever | Diagnosis of hemophagocytic lymphohistiocytosis with intra- and perivascular infiltration of CD20-positive atypical B lymphocytes in spleen, liver, and lungs | Pfizer BNT162b2 mRNA vaccine (2 doses one month apart) | [230] |
| Longitudinal melanonychia that progressed into subungual melanoma | 53-y woman affected by longitudinal melanonychia with no known risk factors for melanoma development | Malignant transformation into acral lentiginous melanoma within 2 years from vaccination | Pfizer BNT162b2 mRNA vaccine (3 doses) | [231] |
| Breast cancer skin metastasis that manifested 1 month after the 6th dose of mRNA vaccination | 85-y woman affected by breast cancer that was successfully removed by partial mastectomy with clear margins 2 years before | Metastatic cancer cells in the dermis and epidermis showed pagetoid atypical cells with ample cytoplasm features and were positive for spike protein, but not for nucleocapsid protein of SARS-CoV-2 | Pfizer-BioNtech BNT162b2 (six doses in 2 years) | [232] |
Table 2.
Case reports relating cancer to anti-COVID-19 genetic vaccines other than mRNA.
| DISEASE | CLINICAL FEATURES | TYPE OF VACCINE | Ref |
|---|---|---|---|
| Pheochromocytoma | 63-y man; pheochromocytoma (very rare benign tumor) of 7 cm developed few days after the vaccination | Johnson and Johnson COVID-19 vaccine | [233] |
| Recurrence of cutaneous T-cell lymphoma | T-cell lymphoma has been reported in two patients, who were in remission since many years, after the 2nd | Vaxzevria (Oxford/AstraZeneca) | [234] |
| EBV–positive, diffuse large B-cell lymphoma | 51-y man; rapidly growing diffuse large B-cell lymphoma was reported in a heart post-transplanted (under immunosuppressant therapy since many years) 7 days after receiving the 1st dose | ChAdOx1 nCoV-19 vaccine | [235] |
| Primary cutaneous T-cell lymphoma | 28-y woman; primary cutaneous T-cell lymphoma (CD31, CD71, CD81 positive) mimicking a panniculites has been reported in a few days after 1st vaccination | COVID-19 Janssen vaccine | [236] |
| Chronic myelomonocytic leukemia | 74-y woman; chronic myelomonocytic leukemia and scleroderma were diagnosed, with first signs manifesting two days after receiving the 1st dose, which then progressed to acute myeloid leukemia, severe anemia, and thrombocytopenia, and eventually died due to COVID-19-associated respiratory failure | Johnson and Johnson COVID-19 vaccine | [237] |
| Classic Kaposi sarcoma manifested 7 days after the 3rd dose of ChAdOx1 vaccine |
73-y man with a skin nodule of 2×3x1 cm HIV negative, positive for CD34 and HHV-8 | ChAdOx1 nCoV-19 vaccine | [238] |
8. Discussion and Concluding Remarks
Anti-COVID-19 vaccination has helped, at least in the early phase of their deployment, to manage COVID-19 by reducing the hospitalizations of the vaccinee and thus relieving the workload of health care workers [239,240], although their real efficacy in protecting from death of hospitalized patients has been recently questioned [241,242]. However, the immune protection provided by these mRNA pro-vaccines was found to last for a few months, necessitating additional shots to maintain anti-Spike IgG levels. Vaccines are generally considered safe with respect to potential carcinogenicity, and therefore their approval normally does not require experimental proof of non-mutagenicity unless the injectable product contains a component never tested in humans and for which it is reasonable to suspect potential mutagenic activity. In the case of the anti-COVID-19 mRNA pro-vaccines it was deemed that the mRNA coding for the Spike protein and the LNP would not have such mutagenic activity. I am of the same opinion, and personally, I believe that these “vaccines” may not have such activity.
Cancer develops after several decades from exposure to mutagenic substances, yet cancerogenesis might be anticipated in individuals with familial predisposition because of inherited mutations in tumor suppressor genes or DNA repair system genes (see 2.1 and 5). Thus, even if conducted in animals, the period of observation (generally 6-24 months) would not be sufficient to show up the potential carcinogenicity of the vaccines in “healthy” animals (with no cancer predisposing genetic defects) maintained in cages under standard conditions with no extra inflammatory hits.
Still, a series of clinical case reports points to a temporal correlation between vaccination with genetically based anti-COVID-19 vaccines and newly diagnosed cancer and cancer progression.
Cancer eventually emerges clinically after a variety of endogenous, exogenous and circumstantial events have altered the structure and composition of the parenchyma and the stroma. Besides genetic mutations, epigenetic dysregulation, inflammation (and angiogenesis), immune suppression, dysregulation of autophagy, impairment of DNA damage repair, activation of signaling for proliferation and migration, inhibition of signaling for cell death, increased energetic metabolism, all these events contribute to cancer development and progression, and to the awakening of dormant tumors leading to cancer relapse. A thorough review of the current literature shows that the SARS-CoV2 infection and (multiple) LNP-mRNA vaccinations could elicit a cancer-promoting effect through several mechanisms, including disruption of the immunosurveillance and induction of inflammation in the tumor microenvironment, disruption of autophagy control, disruption of tumor suppressor pathways, and activation of kinase receptors involved in cell proliferation, cell migration, and EMT. A major player in these events is the Spike proteins, which can lead to down-regulation of protective ACE2 and concomitant activation of the AXL pathway.
These events could combine and be redundantly activated in patients who have been vaccinated and have contracted the infection several times, and in a relatively short time. This unfortunate situation (cocktail effect) would determine a synergism of the damages and alterations caused by the virus and the mRNA pro-vaccine, which can lead to a "catastrophic" effect: cancer. This scenario would be more probable in oncologic patients and in individuals with undiagnosed cancer, and even more in individuals susceptible to cancer because of predisposing genetic defects. An alerting, albeit ignored, signal was reported in a multicenter study where cancer progression and death were reported in some vaccinated patients [68]. As for non-cancer patients, an emblematic case is that of a 43-year-old woman with no significant clinical history who was diagnosed with Ph-positive ALL a few days after vaccination with the double mRNA-1273 vaccine administered in addition to four previous vaccinations with different anti-COVID-19 vaccines plus SARS-CoV2 infections [228].
It is to be noted that the clinical cases discussed here do not establish a causal relationship between the vaccine and the cancer. Such an assessment would require an “ad hoc” investigation [14].
In the practical impossibility to demonstrate a causal link, the biological plausibility of the link between the SARS-CoV2 virus and the anti-COVID-19 mRNA pro-vaccine with cancer must suggest caution in using these type of vaccines and meanwhile adopt appropriate measure to protect the patient at risk (particularly, cancer patients) from the infection, while waiting for vaccine developers to take into account what is expressed here to design safer and more effective vaccines. The present observations call for extra caution when using this type of vaccines, taking into consideration the potential risk of triggering the awakening of dormant cancers or of facilitating the development of cancer in individuals with a genetic predisposition to cancer. First and foremost, it is imperative to elucidate the mechanisms underlying the complex interplay between the virus and vaccination, on the one hand, and oncogenic pathways and the immune system, on the other. This knowledge would also help better stratify patients who truly need vaccination and could inform how to prevent and treat unwanted side effects. In other words, it is advisable to perform a personalized assessment of the real need to vaccinate patients at risk, guided by the principles of vaccinomics and adversomics [51,243]. In the meantime, the adoption of other strategies to protect cancer patients, for instance, by using monoclonal antibodies [244] and convalescent hyperimmune plasma [245], is advised.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The author declares no conflicts of interest. All claims expressed in this article are solely those of the author and do not necessarily represent those of his affiliated organizations, or those of the publisher, the editors, and the reviewers.
References
- Gremke N, Griewing S, Bausch E, Alymova S, Wagner U, Kostev K, Kalder M. Therapy delay due to COVID-19 pandemic among European women with breast cancer: prevalence and associated factors. J Cancer Res Clin Oncol. 2023 Jul 5. Epub ahead of print. PMID: 37405476. [CrossRef]
- Roderburg C, Loosen SH, Leyh C, Joerdens MS, Mohr R, Luedde T, Alymova S, Klein I, Kostev K. Prevalence of and factors associated with a treatment delay due to the COVID-19 pandemic in patients with gastrointestinal cancer in Europe. J Cancer Res Clin Oncol. 2023 Jul 6. Epub ahead of print. PMID: 37414990. [CrossRef]
- Burus T, Lei F, Huang B, Christian WJ, Hull PC, Ellis AR, Slavova S, Tucker TC, Lang Kuhs KA. Undiagnosed Cancer Cases in the US During the First 10 Months of the COVID-19 Pandemic. JAMA Oncol. 2024 Apr 1;10(4):500-507. PMID: 38386344; PMCID: PMC10884945. [CrossRef]
- Castelo-Branco L, Lee R, Brandão M, Cortellini A, Freitas A, Garassino M, Geukens T, Grivas P, Halabi S, Oliveira J, Pinato DJ, Ribeiro J, Peters S, Pentheroudakis G, Warner JL, Romano E. Learning lessons from the COVID-19 pandemic for real-world evidence research in oncology-shared perspectives from international consortia. ESMO Open. 2023 Jun 19;8(4):101596. Epub ahead of print. PMID: 37418836; PMCID: PMC10277850 . [CrossRef]
- Pinato DJ, Scotti L, Gennari A, Colomba-Blameble E, Dolly S, Loizidou A, Chester J, Mukherjee U, Zambelli A, Aguilar-Company J, Bower M, Galazi M, Salazar R, Bertuzzi A, Brunet J, Mesia R, Sita-Lumsden A, Colomba J, Pommeret F, Seguí E, Biello F, Generali D, Grisanti S, Rizzo G, Libertini M, Moss C, Evans JS, Russell B, Wuerstlein R, Vincenzi B, Bertulli R, Ottaviani D, Liñan R, Marrari A, Carmona-García MC, Sng CCT, Tondini C, Mirallas O, Tovazzi V, Fotia V, Cruz CA, Saoudi-Gonzalez N, Felip E, R Lloveras A, Lee AJX, Newsom-Davis T, Sharkey R, Chung C, García-Illescas D, Reyes R, Sophia Wong YN, Ferrante D, Marco-Hernández J, Ruiz-Camps I, Gaidano G, Patriarca A, Sureda A, Martinez-Vila C, Sanchez de Torre A, Rimassa L, Chiudinelli L, Franchi M, Krengli M, Santoro A, Prat A, Tabernero J, V Hemelrijck M, Diamantis N, Cortellini A; OnCovid study group. Determinants of enhanced vulnerability to coronavirus disease 2019 in UK patients with cancer: a European study. Eur J Cancer. 2021 Jun;150:190-202. Epub 2021 Apr 6. PMID: 33932726; PMCID: PMC8023206. [CrossRef]
- Meslé MMI, Brown J, Mook P, Katz MA, Hagan J, Pastore R, Benka B, Redlberger-Fritz M, Bossuyt N, Stouten V, Vernemmen C, Constantinou E, Maly M, Kynčl J, Sanca O, Krause TG, Vestergaard LS, Leino T, Poukka E, Gkolfinopoulou K, Mellou K, Tsintziloni M, Molnár Z, Aspelund G, Thordardottir M, Domegan L, Kelly E, O'Donell J, Urdiales AM, Riccardo F, Sacco C, Bumšteinas V, Liausediene R, Mossong J, Vergison A, Borg ML, Melillo T, Kocinski D, Pollozhani E, Meijerink H, Costa D, Gomes JP, Leite PP, Druc A, Gutu V, Mita V, Lazar M, Popescu R, Popovici O, Musilová M, Mrzel M, Socan M, Učakar V, Limia A, Mazagatos C, Olmedo C, Dabrera G, Kall M, Sinnathamby M, McGowan G, McMenamin J, Morrison K, Nitzan D, Widdowson MA, Smallwood C, Pebody R; WHO European Respiratory Surveillance Network. Estimated number of lives directly saved by COVID-19 vaccination programmes in the WHO European Region from December, 2020, to March, 2023: a retrospective surveillance study. Lancet Respir Med. 2024 Sep;12(9):714-727. Epub 2024 Aug 7. PMID: 39127051. [CrossRef]
- Sarker R, Roknuzzaman ASM, Nazmunnahar, Shahriar M, Hossain MJ, Islam MR. The WHO has declared the end of pandemic phase of COVID-19: Way to come back in the normal life. Health Sci Rep. 2023 Sep 5;6(9):e1544. PMID: 37674622; PMCID: PMC10478644. [CrossRef]
- Ugai T, Sasamoto N, Lee HY, Ando M, Song M, Tamimi RM, Kawachi I, Campbell PT, Giovannucci EL, Weiderpass E, Rebbeck TR, Ogino S. Is early-onset cancer an emerging global epidemic? Current evidence and future implications. Nat Rev Clin Oncol. 2022 Oct;19(10):656-673. Epub 2022 Sep 6. PMID: 36068272; PMCID: PMC9509459. [CrossRef]
- Siegel RL, Giaquinto AN, Jemal A. Cancer statistics, 2024. CA Cancer J Clin. 2024 Jan-Feb;74(1):12-49. Epub 2024 Jan 17. Erratum in: CA Cancer J Clin. 2024 Mar-Apr;74(2):203. PMID: 38230766. doi: 10.3322/caac.21830. [CrossRef]
- Rosenberg PS, Miranda-Filho A. Cancer Incidence Trends in Successive Social Generations in the US. JAMA Netw Open. 2024 Jun 3;7(6):e2415731. PMID: 38857048; PMCID: PMC11165384. [CrossRef]
- Seegers V, Rousseau G, Zhou K, Blanc-Lapierre A, Bigot F, Mahammedi H, Lambert A, Moreau-Bachelard C, Campone M, Conroy T, Penault-Llorca F, Bellanger MM, Raoul JL. COVID-19 Infection despite Previous Vaccination in Cancer Patients and Healthcare Workers: Results from a French Prospective Multicenter Cohort (PAPESCO-19). Cancers (Basel). 2023 Sep 28;15(19):4777. PMID: 37835471; PMCID: PMC10571737. [CrossRef]
- Schmidt AL, Labaki C, Hsu CY, Bakouny Z, Balanchivadze N, Berg SA, Blau S, Daher A, El Zarif T, Friese CR, Griffiths EA, Hawley JE, Hayes-Lattin B, Karivedu V, Latif T, Mavromatis BH, McKay RR, Nagaraj G, Nguyen RH, Panagiotou OA, Portuguese AJ, Puc M, Santos Dutra M, Schroeder BA, Thakkar A, Wulff-Burchfield EM, Mishra S, Farmakiotis D, Shyr Y, Warner JL, Choueiri TK; COVID-19 and Cancer Consortium. COVID-19 vaccination and breakthrough infections in patients with cancer. Ann Oncol. 2022 Mar;33(3):340-346. Epub 2021 Dec 24. PMID: 34958894; PMCID: PMC8704021. [CrossRef]
- Anand ST, Vo AD, La J, Do NV, Fillmore NR, Brophy M, Branch-Elliman W, Monach PA. Severe COVID-19 in Vaccinated Adults with Hematologic Cancers in the Veterans Health Administration. JAMA Netw Open. 2024 Feb 5;7(2):e240288. PMID: 38393725; PMCID: PMC10891464. [CrossRef]
- Bellavite P, Donzelli A, Isidoro C. The WHO Algorithm for Causality Assessment of Adverse Effects Following Immunization with Genetic-Based Anti-COVID-19 Vaccines: Pitfalls and Suggestions for Improvement. J Clin Med. 2024 Nov 30;13(23):7291. PMID: 39685749; PMCID: PMC11642405. [CrossRef]
- Ayuso JM, Ochoa Garrido I. The Importance of the Tumor Microenvironment to Understand Tumor Origin, Evolution, and Treatment Response. Cancers (Basel). 2022 Apr 14;14(8):1983. PMID: 35454888; PMCID: PMC9030088. [CrossRef]
- Beatrice Garavaglia, Letizia Vallino, Alessandra Ferraresi, Danny N. Dhanasekaran, Ciro Isidoro, Chapter 12 - Tumor evolution during chemotherapy (pages 285-305) in Peritoneal Tumor Microenvironment of Cancers on Cancer Hallmarks (Academic Press, 2024; ISBN 9780128240403), Yong Sang Song, Danny N. Dhanasekaran, Benjamin K. Tsang, Johji Inazawa, Massoud Mirshahi, Marc Pocard, Ciro Isidoro Editors. (https://www.sciencedirect.com/science/article/pii/B9780128240403000121). [CrossRef]
- Hanahan D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022 Jan;12(1):31-46. PMID: 35022204. [CrossRef]
- Sinkala M. Mutational landscape of cancer-driver genes across human cancers. Sci Rep. 2023 Aug 7;13(1):12742. PMID: 37550388; PMCID: PMC10406856. [CrossRef]
- Recillas-Targa F. Cancer Epigenetics: An Overview. Arch Med Res. 2022 Dec;53(8):732-740. Epub 2022 Nov 18. PMID: 36411173. [CrossRef]
- Nohmi T. Thresholds of Genotoxic and Non-Genotoxic Carcinogens. Toxicol Res. 2018 Oct;34(4):281-290. Epub 2018 Oct 15. PMID: 30370002; PMCID: PMC6195886. [CrossRef]
- Imyanitov EN, Kuligina ES, Sokolenko AP, Suspitsin EN, Yanus GA, Iyevleva AG, Ivantsov AO, Aleksakhina SN. Hereditary cancer syndromes. World J Clin Oncol. 2023 Feb 24;14(2):40-68. PMID: 36908677; PMCID: PMC9993141. [CrossRef]
- Stephens PJ, Greenman CD, Fu B, Yang F, Bignell GR, Mudie LJ, Pleasance ED, Lau KW, Beare D, Stebbings LA, McLaren S, Lin ML, McBride DJ, Varela I, Nik-Zainal S, Leroy C, Jia M, Menzies A, Butler AP, Teague JW, Quail MA, Burton J, Swerdlow H, Carter NP, Morsberger LA, Iacobuzio-Donahue C, Follows GA, Green AR, Flanagan AM, Stratton MR, Futreal PA, Campbell PJ. Massive genomic rearrangement acquired in a single catastrophic event during cancer development. Cell. 2011 Jan 7;144(1):27-40. PMID: 21215367; PMCID: PMC3065307. [CrossRef]
- Adashek JJ, Kato S, Lippman SM, Kurzrock R. The paradox of cancer genes in non-malignant conditions: implications for precision medicine. Genome Med. 2020 Feb 17;12(1):16. PMID: 32066498; PMCID: PMC7027240. [CrossRef]
- Fiala, C., Diamandis, E.P. Mutations in normal tissues—some diagnostic and clinical implications. BMC Med 18, 283 (2020). [CrossRef]
- Baker SG. A cancer theory kerfuffle can lead to new lines of research. J Natl Cancer Inst. 2014 Dec 20;107(2):dju405. PMID: 25528755; PMCID: PMC4326310. [CrossRef]
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2. Nat Microbiol. 2020 Apr;5(4):536-544. Epub 2020 Mar 2. PMID: 32123347; PMCID: PMC7095448. [CrossRef]
- Nazerian Y, Ghasemi M, Yassaghi Y, Nazerian A, Hashemi SM. Role of SARS-CoV-2-induced cytokine storm in multi-organ failure: Molecular pathways and potential therapeutic options. Int Immunopharmacol. 2022 Dec;113(Pt B):109428. Epub 2022 Nov 7. PMID: 36379152; PMCID: PMC9637536. [CrossRef]
- Singh, M., Pushpakumar, S., Zheng, Y. et al. Novel mechanism of the COVID-19 associated coagulopathy (CAC) and vascular thromboembolism. npj Viruses 1, 3 (2023). [CrossRef]
- Brant AC, Tian W, Majerciak V, Yang W, Zheng ZM. SARS-CoV-2: from its discovery to genome structure, transcription, and replication. Cell Biosci. 2021 Jul 19;11(1):136. PMID: 34281608; PMCID: PMC8287290. [CrossRef]
- V'kovski P, Kratzel A, Steiner S, Stalder H, Thiel V. Coronavirus biology and replication: implications for SARS-CoV-2. Nat Rev Microbiol. 2021 Mar;19(3):155-170. Epub 2020 Oct 28. PMID: 33116300; PMCID: PMC7592455. [CrossRef]
- Ghosh S, Dellibovi-Ragheb TA, Kerviel A, et al. b-Coronaviruses use lysosomes for egress instead of the biosynthetic secretory pathway. Cell. 2020;183(6):1520e1535. e14 . [CrossRef]
- Vidoni C, Fuzimoto A, Ferraresi A, Isidoro C. Targeting autophagy with natural products to prevent SARS-CoV-2 infection. J Tradit Complement Med. 2022 Jan;12(1):55-68. Epub 2021 Oct 14. PMID: 34664025; PMCID: PMC8516241. [CrossRef]
- Choi, Y., Bowman, J.W. & Jung, J.U. Autophagy during viral infection — a double-edged sword. Nat Rev Microbiol 16, 341–354 (2018). [CrossRef]
- Zhang Y, Sun H, Pei R, et al. The SARS-CoV-2 protein ORF3a inhibits fusion of autophagosomes with lysosomes. Cell Discov. 2021;7(1):31. [CrossRef]
- Tao S, Drexler I. Targeting Autophagy in Innate Immune Cells: Angel or Demon During Infection and Vaccination? Front Immunol. 2020 Mar 19;11:460. PMID: 32265919; PMCID: PMC7096474. [CrossRef]
- Sun X, Yu J, Wong SH, Chan MTV, Zhang L, Wu WKK. SARS-CoV-2 targets the lysosome to mediate airway inflammatory cell death. Autophagy. 2022 Sep;18(9):2246-2248. Epub 2022 Jan 22. PMID: 35067165; PMCID: PMC9397439. [CrossRef]
- Yong YY, Zhang L, Hu YJ, Wu JM, Yan L, Pan YR, Tang Y, Yu L, Law BY, Yu CL, Zhou J, Li M, Qin DL, Zhou XG, Wu AG. Targeting autophagy regulation in NLRP3 inflammasome-mediated lung inflammation in COVID-19. Clin Immunol. 2022 Nov;244:109093. Epub 2022 Aug 6. PMID: 35944881; PMCID: PMC9356669. [CrossRef]
- Resnik R, Lopez Mingorance F, Rivera F, Mitchell F, Gonzalez CD, Vaccaro MI. Autophagy in Inflammatory Response against SARS-CoV-2. Int J Mol Sci. 2023 Mar 3;24(5):4928. PMID: 36902354; PMCID: PMC10002778. [CrossRef]
- Afshar ZM, Dayani M, Naderi M, Ghanbarveisi F, Shiri S, Rajati F. Fatality rate of COVID-19 in patients with malignancies: a systematic review and meta-analysis. J Infect. 2020 Aug;81(2):e114-e116. Epub 2020 May 28. PMID: 32474042; PMCID: PMC7255731. [CrossRef]
- Gallo O, Locatello LG, Orlando P, Martelli F, Piccica M, Lagi F, Trotta M. Cancer population may be paradoxically protected from severe manifestations of COVID-19. J Infect. 2020 Aug;81(2):e156-e158. Epub 2020 Jun 10. PMID: 32534002; PMCID: PMC7286269. [CrossRef]
- Ravaioli S, Tebaldi M, Fonzi E, Angeli D, Mazza M, Nicolini F, Lucchesi A, Fanini F, Pirini F, Tumedei MM, Cerchione C, Viale P, Sambri V, Martinelli G, Bravaccini S. ACE2 and TMPRSS2 Potential Involvement in Genetic Susceptibility to SARS-COV-2 in Cancer Patients. Cell Transplant. 2020 Jan-Dec;29:963689720968749. PMID: 33108902; PMCID: PMC7593730. [CrossRef]
- Montopoli M, Zumerle S, Vettor R, Rugge M, Zorzi M, Catapano CV, Carbone GM, Cavalli A, Pagano F, Ragazzi E, Prayer-Galetti T, Alimonti A. Androgen-deprivation therapies for prostate cancer and risk of infection by SARS-CoV-2: a population-based study (N = 4532). Ann Oncol. 2020 Aug;31(8):1040-1045. Epub 2020 May 6. PMID: 32387456; PMCID: PMC7202813. [CrossRef]
- Wang S, Qiu Z, Hou Y, Deng X, Xu W, Zheng T, Wu P, Xie S, Bian W, Zhang C, Sun Z, Liu K, Shan C, Lin A, Jiang S, Xie Y, Zhou Q, Lu L, Huang J, Li X. AXL is a candidate receptor for SARS-CoV-2 that promotes infection of pulmonary and bronchial epithelial cells. Cell Res. 2021 Feb;31(2):126-140. [CrossRef]
- Cantuti-Castelvetri L, Ojha R, Pedro LD, Djannatian M, Franz J, Kuivanen S, van der Meer F, Kallio K, Kaya T, Anastasina M, Smura T, Levanov L, Szirovicza L, Tobi A, Kallio-Kokko H, Österlund P, Joensuu M, Meunier FA, Butcher SJ, Winkler MS, Mollenhauer B, Helenius A, Gokce O, Teesalu T, Hepojoki J, Vapalahti O, Stadelmann C, Balistreri G, Simons M. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science. 2020 Nov 13;370(6518):856-860. [CrossRef]
- Wang, K., Chen, W., Zhang, Z. et al. CD147-spike protein is a novel route for SARS-CoV-2 infection to host cells. Sig Transduct Target Ther 5, 283 (2020). [CrossRef]
- You J, Huang R, Zhong R, Shen J, Huang S, Chen J, Chen F, Kang Y, Chen L. Serum AXL is a potential molecular marker for predicting COVID-19 progression. Front Immunol. 2024 May 10;15:1394429. [CrossRef]
- Huang Y, Wang Y, Xu D, Xiao L, Qin W, Liu B, Yuan X. Characterization of the SARS-CoV-2 co-receptor NRP1 expression profiles in healthy people and cancer patients: Implication for susceptibility to COVID-19 disease and potential therapeutic strategy. Front Genet. 2022 Oct 19;13:995736. [CrossRef]
- Xia P, Dubrovska A. Tumor markers as an entry for SARS-CoV-2 infection? FEBS J. 2020 Sep;287(17):3677-3680. [CrossRef]
- Degenhardt F, Ellinghaus D, Juzenas S, Lerga-Jaso J, Wendorff M, Maya-Miles D, Uellendahl-Werth F, ElAbd H, Rühlemann MC, Arora J, Özer O, Lenning OB, Myhre R, Vadla MS, Wacker EM, Wienbrandt L, Blandino Ortiz A, de Salazar A, Garrido Chercoles A, Palom A, Ruiz A, Garcia-Fernandez AE, Blanco-Grau A, Mantovani A, Zanella A, Holten AR, Mayer A, Bandera A, Cherubini A, Protti A, Aghemo A, Gerussi A, Ramirez A, Braun A, Nebel A, Barreira A, Lleo A, Teles A, Kildal AB, Biondi A, Caballero-Garralda A, Ganna A, Gori A, Glück A, Lind A, Tanck A, Hinney A, Carreras Nolla A, Fracanzani AL, Peschuck A, Cavallero A, Dyrhol-Riise AM, Ruello A, Julià A, Muscatello A, Pesenti A, Voza A, Rando-Segura A, Solier A, Schmidt A, Cortes B, Mateos B, Nafria-Jimenez B, Schaefer B, Jensen B, Bellinghausen C, Maj C, Ferrando C, de la Horra C, Quereda C, Skurk C, Thibeault C, Scollo C, Herr C, Spinner CD, Gassner C, Lange C, Hu C, Paccapelo C, Lehmann C, Angelini C, Cappadona C, Azuure C; COVICAT study group, Aachen Study (COVAS); Bianco C, Cea C, Sancho C, Hoff DAL, Galimberti D, Prati D, Haschka D, Jiménez D, Pestaña D, Toapanta D, Muñiz-Diaz E, Azzolini E, Sandoval E, Binatti E, Scarpini E, Helbig ET, Casalone E, Urrechaga E, Paraboschi EM, Pontali E, Reverter E, Calderón EJ, Navas E, Solligård E, Contro E, Arana-Arri E, Aziz F, Garcia F, García Sánchez F, Ceriotti F, Martinelli-Boneschi F, Peyvandi F, Kurth F, Blasi F, Malvestiti F, Medrano FJ, Mesonero F, Rodriguez-Frias F, Hanses F, Müller F, Hemmrich-Stanisak G, Bellani G, Grasselli G, Pezzoli G, Costantino G, Albano G, Cardamone G, Bellelli G, Citerio G, Foti G, Lamorte G, Matullo G, Baselli G, Kurihara H, Neb H, My I, Kurth I, Hernández I, Pink I, de Rojas I, Galván-Femenia I, Holter JC, Afset JE, Heyckendorf J, Kässens J, Damås JK, Rybniker J, Altmüller J, Ampuero J, Martín J, Erdmann J, Banales JM, Badia JR, Dopazo J, Schneider J, Bergan J, Barretina J, Walter J, Hernández Quero J, Goikoetxea J, Delgado J, Guerrero JM, Fazaal J, Kraft J, Schröder J, Risnes K, Banasik K, Müller KE, Gaede KI, Garcia-Etxebarria K, Tonby K, Heggelund L, Izquierdo-Sanchez L, Bettini LR, Sumoy L, Sander LE, Lippert LJ, Terranova L, Nkambule L, Knopp L, Gustad LT, Garbarino L, Santoro L, Téllez L, Roade L, Ostadreza M, Intxausti M, Kogevinas M, Riveiro-Barciela M, Berger MM, Schaefer M, Niemi MEK, Gutiérrez-Stampa MA, Carrabba M, Figuera Basso ME, Valsecchi MG, Hernandez-Tejero M, Vehreschild MJGT, Manunta M, Acosta-Herrera M, D'Angiò M, Baldini M, Cazzaniga M, Grimsrud MM, Cornberg M, Nöthen MM, Marquié M, Castoldi M, Cordioli M, Cecconi M, D'Amato M, Augustin M, Tomasi M, Boada M, Dreher M, Seilmaier MJ, Joannidis M, Wittig M, Mazzocco M, Ciccarelli M, Rodríguez-Gandía M, Bocciolone M, Miozzo M, Imaz Ayo N, Blay N, Chueca N, Montano N, Braun N, Ludwig N, Marx N, Martínez N; Norwegian SARS-CoV-2 Study group; Cornely OA, Witzke O, Palmieri O; Pa Study Group; Faverio P, Preatoni P, Bonfanti P, Omodei P, Tentorio P, Castro P, Rodrigues PM, España PP, Hoffmann P, Rosenstiel P, Schommers P, Suwalski P, de Pablo R, Ferrer R, Bals R, Gualtierotti R, Gallego-Durán R, Nieto R, Carpani R, Morilla R, Badalamenti S, Haider S, Ciesek S, May S, Bombace S, Marsal S, Pigazzini S, Klein S, Pelusi S, Wilfling S, Bosari S, Volland S, Brunak S, Raychaudhuri S, Schreiber S, Heilmann-Heimbach S, Aliberti S, Ripke S, Dudman S, Wesse T, Zheng T; STORM Study group, The Humanitas Task Force, The Humanitas Gavazzeni Task Force; Bahmer T, Eggermann T, Illig T, Brenner T, Pumarola T, Feldt T, Folseraas T, Gonzalez Cejudo T, Landmesser U, Protzer U, Hehr U, Rimoldi V, Monzani V, Skogen V, Keitel V, Kopfnagel V, Friaza V, Andrade V, Moreno V, Albrecht W, Peter W, Poller W, Farre X, Yi X, Wang X, Khodamoradi Y, Karadeniz Z, Latiano A, Goerg S, Bacher P, Koehler P, Tran F, Zoller H, Schulte EC, Heidecker B, Ludwig KU, Fernández J, Romero-Gómez M, Albillos A, Invernizzi P, Buti M, Duga S, Bujanda L, Hov JR, Lenz TL, Asselta R, de Cid R, Valenti L, Karlsen TH, Cáceres M, Franke A. Detailed stratified GWAS analysis for severe COVID-19 in four European populations. Hum Mol Genet. 2022 Nov 28;31(23):3945-3966. PMID: 35848942; PMCID: PMC9703941. [CrossRef]
- Corley MJ, Pang APS, Dody K, Mudd PA, Patterson BK, Seethamraju H, Bram Y, Peluso MJ, Torres L, Iyer NS, Premeaux TA, Yeung ST, Chandar V, Borczuk A, Schwartz RE, Henrich TJ, Deeks SG, Sacha JB, Ndhlovu LC. Genome-wide DNA methylation profiling of peripheral blood reveals an epigenetic signature associated with severe COVID-19. J Leukoc Biol. 2021 Jul;110(1):21-26. Epub 2021 Jan 19. PMID: 33464637; PMCID: PMC8013321. [CrossRef]
- Ferraresi A, Isidoro C. Will Omics Biotechnologies Save Us from Future Pandemics? Lessons from COVID-19 for Vaccinomics and Adversomics. Biomedicines. 2022 Dec 26;11(1):52. PMID: 36672560; PMCID: PMC9855897. [CrossRef]
- Ghosh M, Kumar S, Ganguly K, Ghosh P, Tabassum S, Basu B, Basu M. COVID-19 and cancer: insights into their association and influence on genetic and epigenetic landscape. Epigenomics. 2023 Feb;15(4):227–48. Epub 2023 May 2. PMCID: PMC10155782.
- Qiu S, Hu Y. Are COVID-19 susceptibility genes related to lung cancer? J Infect. 2021 Nov;83(5):607-635. Epub 2021 Aug 23. PMID: 34437926; PMCID: PMC8381633. https://doi.org/10.2217/epi-2023-0052. [CrossRef]
- Zhang Y, Mao Q, Li Y, Cheng J, Xia Q, Chen G, Chen P, Jin S, Li D, Zhong C, Yang J, Fan X, Liang Y, Lin H. Cancer and COVID-19 Susceptibility and Severity: A Two-Sample Mendelian Randomization and Bioinformatic Analysis. Front Cell Dev Biol. 2022 Jan 24;9:759257. PMID: 35141230; PMCID: PMC8818950. [CrossRef]
- Gusakova MS, Ivanov MV, Kashtanova DA, Taraskina AN, Erema VV, Mikova VM, Loshkarev RI, Ignatyeva OA, Akinshina AI, Mitrofanov SI, Snigir EA, Yudin VS, Makarov VV, Keskinov AA, Yudin SM. GWAS reveals genetic basis of a predisposition to severe COVID-19 through in silico modeling of the FYCO1 protein. Front Med (Lausanne). 2023 Jul 20;10:1178939. PMID: 37547597; PMCID: PMC10399629. [CrossRef]
- Polack FP, Thomas SJ, Kitchin N, Absalon J, Gurtman A, Lockhart S, Perez JL, Pérez Marc G, Moreira ED, Zerbini C, Bailey R, Swanson KA, Roychoudhury S, Koury K, Li P, Kalina WV, Cooper D, Frenck RW Jr, Hammitt LL, Türeci Ö, Nell H, Schaefer A, Ünal S, Tresnan DB, Mather S, Dormitzer PR, Şahin U, Jansen KU, Gruber WC; C4591001 Clinical Trial Group. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N Engl J Med. 2020 Dec 31;383(27):2603-2615. [CrossRef]
- Baden LR, El Sahly HM, Essink B, Kotloff K, Frey S, Novak R, Diemert D, Spector SA, Rouphael N, Creech CB, McGettigan J, Khetan S, Segall N, Solis J, Brosz A, Fierro C, Schwartz H, Neuzil K, Corey L, Gilbert P, Janes H, Follmann D, Marovich M, Mascola J, Polakowski L, Ledgerwood J, Graham BS, Bennett H, Pajon R, Knightly C, Leav B, Deng W, Zhou H, Han S, Ivarsson M, Miller J, Zaks T; COVE Study Group. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N Engl J Med. 2021 Feb 4;384(5):403-416. [CrossRef]
- Beukenhorst AL, Koch CM, Hadjichrysanthou C, Alter G, de Wolf F, Anderson RM, Goudsmit J. SARS-CoV-2 elicits non-sterilizing immunity and evades vaccine-induced immunity: implications for future vaccination strategies. Eur J Epidemiol. 2023 Mar;38(3):237-242. Epub 2023 Feb 4. PMID: 36738380; PMCID: PMC9898703. [CrossRef]
- Yoshimura M, Sakamoto A, Ozuru R, Kurihara Y, Itoh R, Ishii K, Shimizu A, Chou B, Sechi Y, Fujikane A, Nabeshima S, Hiromatsu K. Insufficient anti-spike RBD IgA responses after triple vaccination with intramuscular mRNA BNT162b2 vaccine against SARS-CoV-2. Heliyon. 2023 Dec 13;10(1):e23595. PMID: 38187240; PMCID: PMC10770545. [CrossRef]
- Irrgang P, Gerling J, Kocher K, Lapuente D, Steininger P, Habenicht K, Wytopil M, Beileke S, Schäfer S, Zhong J, Ssebyatika G, Krey T, Falcone V, Schülein C, Peter AS, Nganou-Makamdop K, Hengel H, Held J, Bogdan C, Überla K, Schober K, Winkler TH, Tenbusch M. Class switch toward noninflammatory, spike-specific IgG4 antibodies after repeated SARS-CoV-2 mRNA vaccination. Sci Immunol. 2023 Jan 27;8(79):eade2798. Epub 2023 Jan 27. PMID: 36548397; PMCID: PMC9847566. [CrossRef]
- Kiszel P, Sík P, Miklós J, Kajdácsi E, Sinkovits G, Cervenak L, Prohászka Z. Class switch towards spike protein-specific IgG4 antibodies after SARS-CoV-2 mRNA vaccination depends on prior infection history. Sci Rep. 2023 Aug 13;13(1):13166. PMID: 37574522; PMCID: PMC10423719. [CrossRef]
- Hwang JK, Zhang T, Wang AZ, et al: COVID-19 vaccines for patients with cancer: Benefits likely outweigh risks. J Hematol Oncol 14:38, 2021.
- Dai M, Liu D, Liu M, et al. Patients with cancer appear more vulnerable to SARS-COV-2: a multi-center study during the COVID-19 outbreak. Cancer Discov 2020:CD-20-0422.
- Corti C, Crimini E, Tarantino P, Pravettoni G, Eggermont AMM, Delaloge S, Curigliano G. SARS-CoV-2 vaccines for cancer patients: a call to action. Eur J Cancer. 2021 May;148:316-327. [CrossRef]
- Ljungman P, Nahi H, Linde A. Vaccination of patients with haematological malignancies with one or two doses of influenza vaccine: a randomised study. Br J Haematol 2005;130:96–8.
- Mackay HJ, McGee J, Villa D, et al. Evaluation of pandemic H1N1 (2009) influenza vaccine in adults with solid tumor and hematological malignancies on active systemic treatment. Journal of Clinical Virology 2011;50:212–6.
- Sommer AL, Wachel BK, Smith JA. Evaluation of vaccine dosing in patients with solid tumors receiving myelosuppressive chemotherapy. J Oncol Pharm Pract 2006;12:143–54.
- Shumilov E, Aperdannier L, Schmidt N, Szuszies C, Neesse A, Hoffknecht P, Khandanpour C, Mikesch JH, Stelljes M, Boeckel GR, Tepasse PR, Reitnauer L, Koch R, Hasenkamp J, Bacher U, Scheithauer S, Trümper L, Schmitz N, Wulf G, Kerkhoff A, Lenz G, Krekeler C, Bleckmann A. Clinical Post-SARS-CoV-2 Infection Scenarios in Vaccinated and Non-Vaccinated Cancer Patients in Three German Cancer Centers: A Retrospective Analysis. Cancers (Basel). 2022 Jul 31;14(15):3746. [CrossRef]
- Song Q, Bates B, Shao YR, Hsu FC, Liu F, Madhira V, Mitra AK, Bergquist T, Kavuluru R, Li X, Sharafeldin N, Su J, Topaloglu U. Risk and Outcome of Breakthrough COVID-19 Infections in Vaccinated Patients With Cancer: Real-World Evidence From the National COVID Cohort Collaborative. J Clin Oncol. 2022 May 1;40(13):1414-1427. Epub 2022 Mar 14. PMID: 35286152; PMCID: PMC9061155. [CrossRef]
- Campagna R, Dominelli F, Zingaropoli MA, Ciurluini F, Grilli G, Amoroso A, De Domenico A, Amatore D, Lia MS, Cortesi E, Picone V, Mastroianni CM, Ciardi MR, De Santis R, Lista F, Antonelli G, Turriziani O. COVID-19 vaccination in cancer patients: Immune responses one year after the third dose. Vaccine. 2024 Apr 11;42(10):2687-2694. Epub 2024 Mar 17. PMID: 38499458. [CrossRef]
- Hua T, Fan R, Fan Y, Chen F. Immune response of COVID-19 vaccines in solid cancer patients: A meta-analysis. Hum Vaccin Immunother. 2024 Dec 31;20(1):2357424. Epub 2024 May 24. PMID: 38785118; PMCID: PMC11135846. [CrossRef]
- Gong IY, Vijenthira A, Powis M, Calzavara A, Patrikar A, Sutradhar R, Hicks LK, Wilton D, Singh S, Krzyzanowska MK, Cheung MC. Association of COVID-19 Vaccination With Breakthrough Infections and Complications in Patients With Cancer. JAMA Oncol. 2023 Mar 1;9(3):386-394. PMID: 36580318; PMCID: PMC10020872. [CrossRef]
- Wang L, Kaelber DC, Xu R, Berger NA. COVID-19 breakthrough infections, hospitalizations and mortality in fully vaccinated patients with hematologic malignancies: A clarion call for maintaining mitigation and ramping-up research. Blood Rev. 2022 Jul;54:100931. Epub 2022 Jan 31. PMID: 35120771; PMCID: PMC8802493. [CrossRef]
- Seegers V, Rousseau G, Zhou K, Blanc-Lapierre A, Bigot F, Mahammedi H, Lambert A, Moreau-Bachelard C, Campone M, Conroy T, Penault-Llorca F, Bellanger MM, Raoul JL. COVID-19 Infection despite Previous Vaccination in Cancer Patients and Healthcare Workers: Results from a French Prospective Multicenter Cohort (PAPESCO-19). Cancers (Basel). 2023 Sep 28;15(19):4777. PMID: 37835471; PMCID: PMC10571737. [CrossRef]
- Anand ST, Vo AD, La J, Do NV, Fillmore NR, Brophy M, Branch-Elliman W, Monach PA. Severe COVID-19 in Vaccinated Adults with Hematologic Cancers in the Veterans Health Administration. JAMA Netw Open. 2024 Feb 5;7(2):e240288. PMID: 38393725; PMCID: PMC10891464. [CrossRef]
- Amatu A, Pani A, Patelli G, Gagliardi OM, Loparco M, Piscazzi D, Cassingena A, Tosi F, Ghezzi S, Campisi D, Grifantini R, Abrignani S, Siena S, Scaglione F, Sartore-Bianchi A. Impaired seroconversion after SARS-CoV-2 mRNA vaccines in patients with solid tumours receiving anticancer treatment. Eur J Cancer. 2022 Mar;163:16-25. Epub 2021 Dec 22. PMID: 35032813; PMCID: PMC8692068. [CrossRef]
- Buttiron Webber T, Provinciali N, Musso M, Ugolini M, Boitano M, Clavarezza M, D'Amico M, Defferrari C, Gozza A, Briata IM, Magnani M, Paciolla F, Menghini N, Marcenaro E, De Palma R, Sacchi N, Innocenti L, Siri G, D'Ecclesiis O, Cevasco I, Gandini S, DeCensi A. Predictors of poor seroconversion and adverse events to SARS-CoV-2 mRNA BNT162b2 vaccine in cancer patients on active treatment. Eur J Cancer. 2021 Dec;159:105-112. Epub 2021 Oct 11. PMID: 34742157; PMCID: PMC8502731. [CrossRef]
- Herishanu Y, Avivi I, Aharon A, et al. Efficacy of the BNT162b2 mRNA COVID-19 vaccine in patients with chronic lymphocytic leukemia. Blood (2021) 137 (23): 3165–3173. [CrossRef]
- Vormehr M, Lehar S, Kranz LM, et al. Dexamethasone premedication suppresses vaccine-induced immune responses against cancer. Oncoimmunology 2020;9:1758004.
- Verma V, Shrimali RK, Ahmad S, et al. Pd-1 blockade in subprimed CD8 cells induces dysfunctional PD-1+CD38hi cells and anti-PD-1 resistance. Nat Immunol 2019;20:1231–43.
- Spiliopoulou P, Janse van Rensburg HJ, Avery L, Kulasingam V, Razak A, Bedard P, Hansen A, Chruscinski A, Wang B, Kulikova M, Chen R, Speers V, Nguyen A, Lee J, Coburn B, Spreafico A, Siu LL. Longitudinal efficacy and toxicity of SARS-CoV-2 vaccination in cancer patients treated with immunotherapy. Cell Death Dis. 2023 Jan 20;14(1):49. PMID: 36670100; PMCID: PMC9853486. [CrossRef]
- Nelli, F., Giannarelli, D., Fabbri, A. et al. Immune-related adverse events and disease outcomes after the third dose of SARS-CoV-2 mRNA-BNT162b2 vaccine in cancer patients receiving immune checkpoint inhibitors. Cancer Immunol Immunother (2023). [CrossRef]
- Echaide M, Labiano I, Delgado M, Fernández de Lascoiti A, Ochoa P, Garnica M, Ramos P, Chocarro L, Fernández L, Arasanz H, Bocanegra A, Blanco E, Piñeiro-Hermida S, Morente P, Vera R, Alsina M, Escors D, Kochan G. Immune Profiling Uncovers Memory T-Cell Responses with a Th17 Signature in Cancer Patients with Previous SARS-CoV-2 Infection Followed by mRNA Vaccination. Cancers (Basel). 2022 Sep 14;14(18):4464. PMID: 36139625; PMCID: PMC9496802. [CrossRef]
- Alshehri S, Almutawif YA, Khan NU. Impact of COVID-19 vaccination on cancer patients: safety, efficacy, and long-term effects. Support Care Cancer. 2025 Aug 4;33(8):753. PMID: 40760361. [CrossRef]
- Benitez Fuentes JD, Mohamed Mohamed K, de Luna Aguilar A, Jiménez García C, Guevara-Hoyer K, Fernandez-Arquero M, Rodríguez de la Peña MA, Garciía Bravo L, Jiménez Ortega AF, Flores Navarro P, Bartolome Arcilla J, Alonso Arenilla B, Baos Muñoz E, Delgado-Iribarren García-Campero A, Montealegre Sanz M, Sanchez-Ramon S, Perez Segura P. Evidence of exhausted lymphocytes after the third anti-SARS-CoV-2 vaccine dose in cancer patients. Front Oncol. 2022 Dec 20;12:975980. PMID: 36605446; PMCID: PMC9808030. [CrossRef]
- Raineri D, Mazzucca CB, Moia R, Bruna R, Kustrimovic N, Cappellano G, Bellan M, Perazzi M, Gaidano G, Chiocchetti A. Impairment of the T cell memory response in chronic lymphocytic leukemia patients after SARS-CoV-2 vaccination. Vaccine. 2025 Feb 27;48:126723. Epub 2025 Jan 18. PMID: 39827600. [CrossRef]
- Nelli F, Signorelli C, Fabbri A, Giannarelli D, Virtuoso A, Giron Berrios JR, Marrucci E, Fiore C, Schirripa M, Chilelli MG, Primi F, Panichi V, Topini G, Silvestri MA, Ruggeri EM. Changes in Peripheral Immune Cells after the Third Dose of SARS-CoV-2 mRNA-BNT162b2 Vaccine and Disease Outcomes in Cancer Patients Receiving Immune Checkpoint Inhibitors: A Prospective Analysis of the Vax-on-Third-Profile Study. Cancers (Basel). 2023 Jul 14;15(14):3625. PMID: 37509286; PMCID: PMC10377319. [CrossRef]
- Grippin AJ, Marconi C, Copling S, Li N, Braun C, Woody C, Young E, Gupta P, Wang M, Wu A, Jeong SD, Soni D, Weidert F, Xie C, Goldenberg E, Kim A, Zhao C, DeVries A, Castillo P, Lohray R, Rooney MK, Schrank BR, Wang Y, Ma Y, Chang E, Kouzy R, Dyson K, Jafarnia J, Nariman N, Gladish G, New J, Argueta A, Amaya D, Thomas N, Doty A, Chen J, Copling N, Alatrash G, Simon J, Davies AB, Dennis W, Liang R, Lewis J, Wei X, Rinsurongkawong W, Vaporciyan AA, Johns A; D3CODE Team; Lee J, Lee JH, Sun R, Sharma P, Tran H, Zhang J, Gibbons DL, Wargo J, Kim BYS, Heymach JV, Mendez-Gomez HR, Jiang W, Sayour EJ, Lin SH. SARS-CoV-2 mRNA vaccines sensitize tumours to immune checkpoint blockade. Nature. 2025 Oct 22. Epub ahead of print. PMID: 41125896. [CrossRef]
- Meo C, Palma G, Bruzzese F, Budillon A, Napoli C, de Nigris F. Spontaneous cancer remission after COVID-19: insights from the pandemic and their relevance for cancer treatment. J Transl Med. 2023 Apr 21;21(1):273. PMID: 37085802; PMCID: PMC10119533. [CrossRef]
- Sousa LG, McGrail DJ, Li K, Marques-Piubelli ML, Gonzalez C, Dai H, Ferri-Borgogno S, Godoy M, Burks J, Lin SY, Bell D, Ferrarotto R. Spontaneous tumor regression following COVID-19 vaccination. J Immunother Cancer. 2022 Mar;10(3):e004371. PMID: 35241495; PMCID: PMC8896046. [CrossRef]
- Gambichler T, Boms S, Hessam S, Tischoff I, Tannapfel A, Lüttringhaus T, Beckman J, Stranzenbach R. Primary cutaneous anaplastic large-cell lymphoma with marked spontaneous regression of organ manifestation after SARS-CoV-2 vaccination. Br J Dermatol. 2021 Dec;185(6):1259-1262. Epub 2021 Oct 3. PMID: 34228815; PMCID: PMC8444914. [CrossRef]
- Wesselmann, UlrichKvasnicka, Hans M.Bozkurt, AhmetWieland, UlrikeHofmann, Silke C. et al. Long lasting complete regression of a metastatic polyomavirus-positive Merkel cell carcinoma after COVID-19 booster vaccination. EJC Skin Cancer, Volume 2, 100275.
- Eslinger C, Uson PLS Jr, Nagalo BM, Borad MJ. Spontaneous regression of advanced hepatocellular carcinoma following COVID-19 infection and vaccination: a case report and review of literature. J Gastrointest Oncol. 2024 Aug 31;15(4):1933-1938. Epub 2024 Jul 22. PMID: 39279952; PMCID: PMC11399873. [CrossRef]
- Neophytou CM, Kyriakou TC, Papageorgis P. Mechanisms of Metastatic Tumor Dormancy and Implications for Cancer Therapy. Int J Mol Sci. 2019 Dec 6;20(24):6158. PMID: 31817646; PMCID: PMC6940943. [CrossRef]
- Wang Y, Wang L, Wei Y, Wei C, Yang H, Chen Q, Zhang R, Shen H. Advances in the molecular regulation mechanism of tumor dormancy and its therapeutic strategy. Discov Oncol. 2024 May 25;15(1):184. [CrossRef]
- Gunes D, Ustal A, Ertem YE, Akkoc Y, Gozuacik D. Autophagy in the regulation of cancer dormancy. FEBS Lett. 2025 Aug;599(16):2272-2300. Epub 2025 Aug 13. PMID: 40804788. [CrossRef]
- Esposito A, Ferraresi A, Salwa A, Vidoni C, Dhanasekaran DN, Isidoro C. Resveratrol Contrasts IL-6 Pro-Growth Effects and Promotes Autophagy-Mediated Cancer Cell Dormancy in 3D Ovarian Cancer: Role of miR-1305 and of Its Target ARH-I. Cancers (Basel). 2022 Apr 25;14(9):2142. PMID: 35565270; PMCID: PMC9101105. [CrossRef]
- Manjili SH, Isbell M, Ghochaghi N, Perkinson T, Manjili MH. Multifaceted functions of chronic inflammation in regulating tumor dormancy and relapse. Semin Cancer Biol. 2022 Jan;78:17-22. Epub 2021 Mar 27. PMID: 33785450; PMCID: PMC8473586. [CrossRef]
- Ferraresi A, Girone C, Esposito A, Vidoni C, Vallino L, Secomandi E, Dhanasekaran DN, Isidoro C. How Autophagy Shapes the Tumor Microenvironment in Ovarian Cancer. Front Oncol. 2020 Dec 7;10:599915. PMID: 33364196; PMCID: PMC7753622. [CrossRef]
- Langley RR, Fidler IJ. The seed and soil hypothesis revisited--the role of tumor-stroma interactions in metastasis to different organs. Int J Cancer. 2011 Jun 1;128(11):2527-35. [CrossRef]
- Unterlass JE, Curtin NJ. Warburg and Krebs and related effects in cancer. Expert Rev Mol Med. 2019 Sep 27;21:e4. PMID: 31558177. [CrossRef]
- Costanzo, M.; De Giglio, M.A.R.; Roviello, G.N. Deciphering the Relationship between SARS-CoV-2 and Cancer. Int. J. Mol. Sci. 2023, 24, 7803.
- Rapti, V.; Tsaganos, T.; Vathiotis, I.A.; Syrigos, N.K.; Li, P.; Poulakou, G. New Insights into SARS-CoV-2 and Cancer Cross-Talk:Does a Novel Oncogenesis Driver Emerge? Vaccines 2022, 10, 1607..
- Venuti A, Donzelli S, Nisticò P, Blandino G, Ciliberto G. Does Interleukin-6 Bridge SARS-CoV-2 With Virus-Associated Cancers? J Immunother Precis Oncol. 2021 Jan 29;4(2):79-85. PMID: 35663529; PMCID: PMC9153257. [CrossRef]
- Vieira C, Nery L, Martins L, Jabour L, Dias R, Simões E Silva AC. Downregulation of Membrane-bound Angiotensin Converting Enzyme 2 (ACE2) Receptor has a Pivotal Role in COVID-19 Immunopathology. Curr Drug Targets. 2021;22(3):254-281. PMID: 33081670. [CrossRef]
- Lei Y, Zhang J, Schiavon CR, He M, Chen L, Shen H, Zhang Y, Yin Q, Cho Y, Andrade L, Shadel GS, Hepokoski M, Lei T, Wang H, Zhang J, Yuan JX, Malhotra A, Manor U, Wang S, Yuan ZY, Shyy JY. SARS-CoV-2 Spike Protein Impairs Endothelial Function via Downregulation of ACE 2. Circ Res. 2021 Apr 30;128(9):1323-1326. Epub 2021 Mar 31. PMID: 33784827; PMCID: PMC8091897. [CrossRef]
- Bellavite P, Ferraresi A, Isidoro C. Immune Response and Molecular Mechanisms of Cardiovascular Adverse Effects of Spike Proteins from SARS-CoV-2 and mRNA Vaccines. Biomedicines. 2023 Feb 3;11(2):451. PMID: 36830987; PMCID: PMC9953067. [CrossRef]
- Hassani B, Attar Z, Firouzabadi N. The renin-angiotensin-aldosterone system (RAAS) signaling pathways and cancer: foes versus allies. Cancer Cell Int. 2023 Oct 27;23(1):254. PMID: 37891636; PMCID: PMC10604988. [CrossRef]
- Menon J, Soto-Pantoja DR, Callahan MF, Cline JM, Ferrario CM, Tallant EA, Gallagher PE. Angiotensin-(1-7) inhibits growth of human lung adenocarcinoma xenografts in nude mice through a reduction in cyclooxygenase-2. Cancer Res. 2007 Mar 15;67(6):2809-15. PMID: 17363603. [CrossRef]
- Soto-Pantoja DR, Menon J, Gallagher PE, Tallant EA. Angiotensin-(1-7) inhibits tumor angiogenesis in human lung cancer xenografts with a reduction in vascular endothelial growth factor. Mol Cancer Ther. 2009 Jun;8(6):1676-83. Epub 2009 Jun 9. PMID: 19509262; PMCID: PMC3314264. [CrossRef]
- Zhang Q, Lu S, Li T, Yu L, Zhang Y, Zeng H, Qian X, Bi J, Lin Y. ACE2 inhibits breast cancer angiogenesis via suppressing the VEGFa/VEGFR2/ERK pathway. J Exp Clin Cancer Res. 2019 Apr 25;38(1):173. PMID: 31023337; PMCID: PMC6482513. [CrossRef]
- Feng Y, Wan H, Liu J, Zhang R, Ma Q, Han B, Xiang Y, Che J, Cao H, Fei X, Qiu W. The angiotensin-converting enzyme 2 in tumor growth and tumor-associated angiogenesis in non-small cell lung cancer. Oncol Rep. 2010 Apr;23(4):941-8. PMID: 20204277. [CrossRef]
- Xie P, Guo L, Yu Q, Zhao Y, Yu M, Wang H, Wu M, Xu W, Xu M, Zhu XD, Xu Y, Xiao YS, Huang C, Zhou J, Fan J, Hung MC, Sun H, Ye QH, Zhang B, Li H. ACE2 Enhances Sensitivity to PD-L1 Blockade by Inhibiting Macrophage-Induced Immunosuppression and Angiogenesis. Cancer Res. 2025 Jan 15;85(2):299-313. PMID: 39495239. [CrossRef]
- Stewart CA, Gay CM, Ramkumar K, Cargill KR, Cardnell RJ, Nilsson MB, Heeke S, Park EM, Kundu ST, Diao L, Wang Q, Shen L, Xi Y, Zhang B, Della Corte CM, Fan Y, Kundu K, Gao B, Avila K, Pickering CR, Johnson FM, Zhang J, Kadara H, Minna JD, Gibbons DL, Wang J, Heymach JV, Byers LA. Lung Cancer Models Reveal Severe Acute Respiratory Syndrome Coronavirus 2-Induced Epithelial-to-Mesenchymal Transition Contributes to Coronavirus Disease 2019 Pathophysiology. J Thorac Oncol. 2021 Nov;16(11):1821-1839. Epub 2021 Jul 16. PMID: 34274504; PMCID: PMC8282443. [CrossRef]
- Lai YJ, Chao CH, Liao CC, Lee TA, Hsu JM, Chou WC, Wang J, Huang HC, Chang SJ, Lin YL, Li CW. Epithelial-mesenchymal transition induced by SARS-CoV-2 required transcriptional upregulation of Snail. Am J Cancer Res. 2021 May 15;11(5):2278-2290. PMID: 34094684; PMCID: PMC8167694.
- Huang HC, Liao CC, Wang SH, Lee IJ, Lee TA, Hsu JM, Kuo CT, Wang J, Hsieh WC, Chang SJ, Chen SY, Tao MH, Lin YL, Lai YJ, Li CW. Hyperglycosylated spike of SARS-CoV-2 gamma variant induces breast cancer metastasis. Am J Cancer Res. 2021 Oct 15;11(10):4994-5005.
- Yu C, Tang W, Wang Y, Shen Q, Wang B, Cai C, Meng X, Zou F. Downregulation of ACE2/Ang-(1-7)/Mas axis promotes breast cancer metastasis by enhancing store-operated calcium entry. Cancer Lett. 2016 Jul 1;376(2):268-77. Epub 2016 Apr 7. PMID: 27063099. [CrossRef]
- Catarata MJ, Ribeiro R, Oliveira MJ, Robalo Cordeiro C, Medeiros R. Renin-Angiotensin System in Lung Tumor and Microenvironment Interactions. Cancers (Basel). 2020 Jun 3;12(6):1457. PMID: 32503281; PMCID: PMC7352181. [CrossRef]
- Wu Z, Yao T, Wang Z, Liu B, Wu N, Lu M, Shen N. Association between angiotensin-converting enzyme inhibitors and the risk of lung cancer: a systematic review and meta-analysis. Br J Cancer. 2023 Jan;128(2):168-176. Epub 2022 Nov 17. PMID: 36396817; PMCID: PMC9670057. [CrossRef]
- Emslander Q, Krey K, Hamad S, Maidl S, Oubraham L, Hesse J, Henrici A, Austen K, Mergner J, Grass V, Pichlmair A. MDM2 Influences ACE2 Stability and SARS-CoV-2 Uptake. Viruses. 2023 Aug 18;15(8):1763. PMID: 37632105; PMCID: PMC10459000. [CrossRef]
- Goyette MA, Duhamel S, Aubert L, Pelletier A, Savage P, Thibault MP, Johnson RM, Carmeliet P, Basik M, Gaboury L, Muller WJ, Park M, Roux PP, Gratton JP, Côté JF. The Receptor Tyrosine Kinase AXL Is Required at Multiple Steps of the Metastatic Cascade during HER2-Positive Breast Cancer Progression. Cell Rep. 2018 May 1;23(5):1476-1490. doi: 10.1016/j.celrep.2018.04.019. Erratum in: Cell Rep. 2023 Dec 26;42(12):113604. [CrossRef]
- Chen IY, Chang SC, Wu HY, Yu TC, Wei WC, Lin S, Chien CL, Chang MF. Upregulation of the chemokine (C-C motif) ligand 2 via a severe acute respiratory syndrome coronavirus spike-ACE2 signaling pathway. J Virol. 2010 Aug;84(15):7703-12. Epub 2010 May 19. PMID: 20484496; PMCID: PMC2897593. [CrossRef]
- Suzuki YJ, Nikolaienko SI, Dibrova VA, Dibrova YV, Vasylyk VM, Novikov MY, Shults NV, Gychka SG. SARS-CoV-2 spike protein-mediated cell signaling in lung vascular cells. Vascul Pharmacol. 2021 Apr;137:106823. Epub 2020 Nov 21. PMID: 33232769; PMCID: PMC7680014. [CrossRef]
- Patra T, Meyer K, Geerling L, Isbell TS, Hoft DF, Brien J, Pinto AK, Ray RB, Ray R. SARS-CoV-2 spike protein promotes IL-6 trans-signaling by activation of angiotensin II receptor signaling in epithelial cells. PLoS Pathog. 2020 Dec 7;16(12):e1009128. PMID: 33284859; PMCID: PMC7746263. [CrossRef]
- Shin HJ, Lee W, Ku KB, Yoon GY, Moon HW, Kim C, Kim MH, Yi YS, Jun S, Kim BT, Oh JW, Siddiqui A, Kim SJ. SARS-CoV-2 aberrantly elevates mitochondrial bioenergetics to induce robust virus propagation. Signal Transduct Target Ther. 2024 May 11;9(1):125. PMID: 38734691; PMCID: PMC11088672. [CrossRef]
- McCubrey JA, Steelman LS, Abrams SL, Lee JT, Chang F, Bertrand FE, Navolanic PM, Terrian DM, Franklin RA, D'Assoro AB, Salisbury JL, Mazzarino MC, Stivala F, Libra M. Roles of the RAF/MEK/ERK and PI3K/PTEN/AKT pathways in malignant transformation and drug resistance. Adv Enzyme Regul. 2006;46:249-79. Epub 2006 Jul 18. PMID: 16854453. [CrossRef]
- Guo YJ, Pan WW, Liu SB, Shen ZF, Xu Y, Hu LL. ERK/MAPK signalling pathway and tumorigenesis. Exp Ther Med. 2020 Mar;19(3):1997-2007. Epub 2020 Jan 15. PMID: 32104259; PMCID: PMC7027163. [CrossRef]
- Solis O, Beccari AR, Iaconis D, Talarico C, Ruiz-Bedoya CA, Nwachukwu JC, Cimini A, Castelli V, Bertini R, Montopoli M, Cocetta V, Borocci S, Prandi IG, Flavahan K, Bahr M, Napiorkowski A, Chillemi G, Ooka M, Yang X, Zhang S, Xia M, Zheng W, Bonaventura J, Pomper MG, Hooper JE, Morales M, Rosenberg AZ, Nettles KW, Jain SK, Allegretti M, Michaelides M. The SARS-CoV-2 spike protein binds and modulates estrogen receptors. Sci Adv. 2022 Dec 2;8(48):eadd4150. [CrossRef]
- Khan I, Hatiboglu MA. Can COVID-19 induce glioma tumorogenesis through binding cell receptors? Med Hypotheses. 2020 Nov;144:110009. Epub 2020 Jun 19. PMID: 32758869; PMCID: PMC7303027. [CrossRef]
- Raimundo L, Ramos H, Loureiro JB, Calheiros J, Saraiva L. BRCA1/P53: Two strengths in cancer chemoprevention. Biochim Biophys Acta Rev Cancer. 2020 Jan;1873(1):188339. Epub 2020 Jan 7. PMID: 31917206. [CrossRef]
- Salwa A, Ferraresi A, Chinthakindi M, Vallino L, Vidoni C, Dhanasekaran DN, Isidoro C. BECN1 and BRCA1 Deficiency Sensitizes Ovarian Cancer to Platinum Therapy and Confers Better Prognosis. Biomedicines. 2021 Feb 18;9(2):207. PMID: 33670664; PMCID: PMC7922320. [CrossRef]
- Kastenhuber ER, Lowe SW. Putting p53 in Context. Cell. 2017; 170:1062–78.
- Shim D, Duan L, Maki CG. P53-regulated autophagy and its impact on drug resistance and cell fate. Cancer Drug Resist. 2021;4(1):85-95. Epub 2021 Mar 19. PMID: 34532654; PMCID: PMC8443158. [CrossRef]
- Ma-Lauer Y, Carbajo-Lozoya J, Hein MY, Müller MA, Deng W, Lei J, Meyer B, Kusov Y, von Brunn B, Bairad DR, Hünten S, Drosten C, Hermeking H, Leonhardt H, Mann M, Hilgenfeld R, von Brunn A. p53 down-regulates SARS coronavirus replication and is targeted by the SARS-unique domain and PLpro via E3 ubiquitin ligase RCHY1. Proc Natl Acad Sci U S A. 2016 Aug 30;113(35):E5192-201. Epub 2016 Aug 12. PMID: 27519799; PMCID: PMC5024628. [CrossRef]
- Singh N, Bharara Singh A. S2 subunit of SARS-nCoV-2 interacts with tumor suppressor protein p53 and BRCA: an in silico study. Transl Oncol. 2020 Oct;13(10):100814. Epub 2020 Jun 30. PMID: 32619819; PMCID: PMC7324311. [CrossRef]
- Zhang S, El-Deiry WS. Transfected SARS-CoV-2 spike DNA for mammalian cell expression inhibits p53 activation of p21(WAF1), TRAIL Death Receptor DR5 and MDM2 proteins in cancer cells and increases cancer cell viability after chemotherapy exposure. Oncotarget. 2024 May 3;15:275-284. PMID: 38709242; PMCID: PMC11073320. [CrossRef]
- Wang HF, Xiang W, Xue BZ, Wang YH, Yi DY, Jiang XB, Zhao HY, Fu P. Cell fusion in cancer hallmarks: Current research status and future indications. Oncol Lett. 2021 Jul;22(1):530. Epub 2021 May 16. PMID: 34055095; PMCID: PMC8138896. [CrossRef]
- Shultes PV, Weaver DT, Tadele DS, Barker-Clarke RJ, Scott JG. Cell-cell fusion in cancer: The next cancer hallmark? Int J Biochem Cell Biol. 2024 Oct;175:106649. Epub 2024 Aug 24. PMID: 39186970; PMCID: PMC11752790. [CrossRef]
- Zhang Z, Zheng Y, Niu Z, Zhang B, Wang C, Yao X, Peng H, Franca DN, Wang Y, Zhu Y, Su Y, Tang M, Jiang X, Ren H, He M, Wang Y, Gao L, Zhao P, Shi H, Chen Z, Wang X, Piacentini M, Bian X, Melino G, Liu L, Huang H, Sun Q. SARS-CoV-2 spike protein dictates syncytium-mediated lymphocyte elimination. Cell Death Differ. 2021 Sep;28(9):2765-2777. Epub 2021 Apr 20. PMID: 33879858; PMCID: PMC8056997. [CrossRef]
- Rajah MM, Hubert M, Bishop E, Saunders N, Robinot R, Grzelak L, Planas D, Dufloo J, Gellenoncourt S, Bongers A, Zivaljic M, Planchais C, Guivel-Benhassine F, Porrot F, Mouquet H, Chakrabarti LA, Buchrieser J, Schwartz O. SARS-CoV-2 Alpha, Beta, and Delta variants display enhanced Spike-mediated syncytia formation. EMBO J. 2021 Dec 15;40(24):e108944. Epub 2021 Oct 25. PMID: 34601723; PMCID: PMC8646911. [CrossRef]
- Fang Q, He X, Zheng X, Fu Y, Fu T, Luo J, Du Y, Lan J, Yang J, Luo Y, Chen X, Zhou N, Wang Z, Lyu J, Chen L. Verifying AXL and putative proteins as SARS-CoV-2 receptors by DnaE intein-based rapid cell-cell fusion assay. J Med Virol. 2023 Jul;95(7):e28953. PMID: 37461287. [CrossRef]
- Schilling WHK, Mukaka M, Callery JJ, Llewelyn MJ, Cruz CV, Dhorda M, Ngernseng T, Waithira N, Ekkapongpisit M, Watson JA, Chandna A, Nelwan EJ, Hamers RL, Etyang A, Beg MA, Sow S, Yavo W, Allabi AC, Basnyat B, Sharma SK, Amofa-Sekyi M, Yonga P, Adler A, Yuentrakul P, Cope T, Thaipadungpanit J, Rienpradub P, Imwong M, Abdad MY, Blacksell SD, Tarning J, Goudjo FF, Dossou AD, Konaté-Touré A, Assi SB, Ouffoué K, Nasronudin N, Rachman BE, Romadhon PZ, Dewanto DD, Heryana MO, Novi T, Pasaribu AP, Mutiara M, Nasution MPR, Khairunnisa K, Dalimunthe FA, Airlangga E, Fahrezzy A, Subronto Y, Ananda NR, Rahardjani M, Rimainar A, Lucinde RK, Timbwa M, Onyango OE, Agutu C, Akech S, Hamaluba M, Kipyego J, Ngachi O, Haidara FC, Traoré OY, Diarra F, Khanal B, Dahal P, Shrestha S, Rijal S, Kabore Y, Adehossi E, Guindo O, Qamar FN, Kazi AM, Woodrow CJ, Laird S, Cheeba M, Ayles H, Cheah PY, Taylor WRJ, Batty EM, Chotivanich K, Pukrittayakamee S, Phumratanaprapin W, von Seidlein L, Dondorp A, Day NPJ, White NJ; COPCOV Collaborative Group. Evaluation of hydroxychloroquine or chloroquine for the prevention of COVID-19 (COPCOV): A double-blind, randomised, placebo-controlled trial. PLoS Med. 2024 Sep 12;21(9):e1004428. [CrossRef]
- Tretyakova MS, Subbalakshmi AR, Menyailo ME, Jolly MK, Denisov EV. Tumor Hybrid Cells: Nature and Biological Significance. Front Cell Dev Biol. 2022 Feb 15;10:814714. PMID: 35242760; PMCID: PMC8886020. [CrossRef]
- Melzer C, Ohe JV, Hass R. Altered Tumor Plasticity after Different Cancer Cell Fusions with MSC. Int J Mol Sci. 2020 Nov 6;21(21):8347. PMID: 33172211; PMCID: PMC7664391. [CrossRef]
- Dikic I, Elazar Z. Mechanism and medical implications of mammalian autophagy. Nat. Rev. Mol. Cell Biol. 2018;19:349–364. [CrossRef]
- Debnath J, Gammoh N, Ryan KM. Autophagy and autophagy-related pathways in cancer. Nat Rev Mol Cell Biol. 2023 Aug;24(8):560-575. Epub 2023 Mar 2. PMID: 36864290; PMCID: PMC9980873. [CrossRef]
- Ferraresi A, Esposito A, Girone C, Vallino L, Salwa A, Ghezzi I, Thongchot S, Vidoni C, Dhanasekaran DN, Isidoro C. Resveratrol Contrasts LPA-Induced Ovarian Cancer Cell Migration and Platinum Resistance by Rescuing Hedgehog-Mediated Autophagy. Cells. 2021 Nov 17;10(11):3213. PMID: 34831435; PMCID: PMC8625920. [CrossRef]
- Koepke L, et al. Manipulation of autophagy by SARS-CoV-2 proteins. Autophagy. 2021;17:2659–2661. [CrossRef]
- Chen D, et al. ORF3a of SARS-CoV-2 promotes lysosomal exocytosis-mediated viral egress. Dev. Cell. 2021;56:3250–3263 e3255. [CrossRef]
- Li, X., Hou, P., Ma, W. et al. SARS-CoV-2 ORF10 suppresses the antiviral innate immune response by degrading MAVS through mitophagy. Cell Mol Immunol 19, 67–78 (2022). [CrossRef]
- Li X, Zhang Z, Wang Z, Gutiérrez-Castrellón P, Shi H. Cell deaths: Involvement in the pathogenesis and intervention therapy of COVID-19. Signal Transduct Target Ther. 2022 Jun 13;7(1):186. PMID: 35697684; PMCID: PMC9189267. [CrossRef]
- Eissa S, Matboli M, Awad N, Kotb Y. Identification and validation of a novel autophagy gene expression signature for human bladder cancer patients. Tumour Biol. 2017 Apr;39(4):1010428317698360. PMID: 28381171. [CrossRef]
- Sun X, Zhou L, Wang X, Li Y, Liu X, Chen Y, Zhong Z, Chen J. FYCO1 regulates migration, invasion, and invadopodia formation in HeLa cells through CDC42/N-WASP/Arp2/3 signaling pathway. Biochem Cell Biol. 2022 Dec 1;100(6):458-472. Epub 2022 Nov 7. PMID: 36342046. [CrossRef]
- Zhu L., She Z.G., Cheng X., Qin J.J., Zhang X.J., Cai J., Lei F., Wang H., Xie J., Wang W. Association of blood glucose control and outcomes in patients with COVID-19 and pre-existing type 2 diabetes. Cell Metab. 2020;31:1068–1077.e3.
- Liberti MV, Locasale JW. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem Sci. 2016 Mar;41(3):211-218. doi: 10.1016/j.tibs.2015.12.001. Epub 2016 Jan 5. Erratum in: Trends Biochem Sci. 2016 Mar;41(3):287. Erratum in: Trends Biochem Sci. 2016 Mar;41(3):287. doi: 10.1016/j.tibs.2016.01.004. PMID: 26778478; PMCID: PMC4783224.
- Balamurugan K. HIF-1 at the crossroads of hypoxia, inflammation, and cancer. Int J Cancer. 2016 Mar 1;138(5):1058-66. Epub 2015 Apr 7. PMID: 25784597; PMCID: PMC4573780. [CrossRef]
- Codo AC, Davanzo GG, Monteiro LB, de Souza GF, Muraro SP, Virgilio-da-Silva JV, Prodonoff JS, Carregari VC, de Biagi Junior CAO, Crunfli F, Jimenez Restrepo JL, Vendramini PH, Reis-de-Oliveira G, Bispo Dos Santos K, Toledo-Teixeira DA, Parise PL, Martini MC, Marques RE, Carmo HR, Borin A, Coimbra LD, Boldrini VO, Brunetti NS, Vieira AS, Mansour E, Ulaf RG, Bernardes AF, Nunes TA, Ribeiro LC, Palma AC, Agrela MV, Moretti ML, Sposito AC, Pereira FB, Velloso LA, Vinolo MAR, Damasio A, Proença-Módena JL, Carvalho RF, Mori MA, Martins-de-Souza D, Nakaya HI, Farias AS, Moraes-Vieira PM. Elevated Glucose Levels Favor SARS-CoV-2 Infection and Monocyte Response through a HIF-1α/Glycolysis-Dependent Axis. Cell Metab. 2020 Sep 1;32(3):437-446.e5. doi: 10.1016/j.cmet.2020.07.007. Epub 2020 Jul 17. Erratum in: Cell Metab. 2020 Sep 1;32(3):498-499. doi: 10.1016/j.cmet.2020.07.015. PMID: 32697943; PMCID: PMC7367032.
- Tian M, Liu W, Li X, Zhao P, Shereen MA, Zhu C, Huang S, Liu S, Yu X, Yue M, Pan P, Wang W, Li Y, Chen X, Wu K, Luo Z, Zhang Q, Wu J. HIF-1α promotes SARS-CoV-2 infection and aggravates inflammatory responses to COVID-19. Signal Transduct Target Ther. 2021 Aug 18;6(1):308. PMID: 34408131; PMCID: PMC8371950. [CrossRef]
- Vidoni C, Ferraresi A, Vallino L, Salwa A, Ha JH, Seca C, Garavaglia B, Dhanasekaran DN, Isidoro C. Glycolysis Inhibition of Autophagy Drives Malignancy in Ovarian Cancer: Exacerbation by IL-6 and Attenuation by Resveratrol. Int J Mol Sci. 2023 Jan 15;24(2):1723. PMID: 36675246; PMCID: PMC9866176. [CrossRef]
- Ferraresi A, Girone C, Maheshwari C, Vallino L, Dhanasekaran DN, Isidoro C. Ovarian Cancer Cell-Conditioning Medium Induces Cancer-Associated Fibroblast Phenoconversion through Glucose-Dependent Inhibition of Autophagy. Int J Mol Sci. 2024 May 23;25(11):5691. PMID: 38891879; PMCID: PMC11171902. [CrossRef]
- Unterlass JE, Curtin NJ. Warburg and Krebs and related effects in cancer. Expert Rev Mol Med. 2019 Sep 27;21:e4. PMID: 31558177. [CrossRef]
- Nakamura H, Takada K. Reactive oxygen species in cancer: Current findings and future directions. Cancer Sci. 2021 Oct;112(10):3945-3952. Epub 2021 Aug 2. PMID: 34286881; PMCID: PMC8486193. [CrossRef]
- Hasan A, Rizvi SF, Parveen S, Pathak N, Nazir A, Mir SS. Crosstalk Between ROS and Autophagy in Tumorigenesis: Understanding the Multifaceted Paradox. Front Oncol. 2022 Mar 10;12:852424. PMID: 35359388; PMCID: PMC8960719. [CrossRef]
- Molnar T, Lehoczki A, Fekete M, Varnai R, Zavori L, Erdo-Bonyar S, Simon D, Berki T, Csecsei P, Ezer E. Mitochondrial dysfunction in long COVID: mechanisms, consequences, and potential therapeutic approaches. Geroscience. 2024 Apr 26. Epub ahead of print. PMID: 38668888. [CrossRef]
- Karki R, Sharma BR, Tuladhar S, et al. Synergism of TNF-α and IFN-γ triggers inflammatory cell death, tissue damage, and mortality in SARS-CoV-2 infection and cytokine shock syndromes. Cell. 2021;184:149–168.
- Zheng M., Gao Y., Wang G., Song G., Liu S., Sun D., Xu Y., Tian Z. Functional exhaustion of antiviral lymphocytes in COVID-19 patients. Cell. Mol. Immunol. 2020;17:533–535.
- Iwasaki M, Saito J, Zhao H, Sakamoto A, Hirota K, Ma D. Inflammation triggered by SARS-CoV-2 and ACE2 augment drives multiple organ failure of severe COVID-19: molecular mechanisms and implications. Inflammation. 2021;44:13–34.
- Li, K., Shi, H., Zhang, B. et al. Myeloid-derived suppressor cells as immunosuppressive regulators and therapeutic targets in cancer. Sig Transduct Target Ther 6, 362 (2021). [CrossRef]
- Francescangeli F, De Angelis ML, Baiocchi M, Rossi R, Biffoni M, Zeuner A. COVID-19-Induced Modifications in the Tumor Microenvironment: Do They Affect Cancer Reawakening and Metastatic Relapse? Front Oncol. 2020 Oct 26;10:592891. PMID: 33194755; PMCID: PMC7649335. [CrossRef]
- Francescangeli F, De Angelis ML, Zeuner A. COVID-19: a potential driver of immune-mediated breast cancer recurrence? Breast Cancer Res. 2020 Oct 30;22(1):117. PMID: 33126915; PMCID: PMC7598231. [CrossRef]
- Yumoto K, Eber MR, Wang J, Cackowski FC, Decker AM, Lee E, Nobre AR, Aguirre-Ghiso JA, Jung Y, Taichman RS. Axl is required for TGF-β2-induced dormancy of prostate cancer cells in the bone marrow. Sci Rep. 2016 Nov 7;6:36520. [CrossRef]
- Han J, Bae J, Choi CY, Choi SP, Kang HS, Jo EK, Park J, Lee YS, Moon HS, Park CG, Lee MS, Chun T. Autophagy induced by AXL receptor tyrosine kinase alleviates acute liver injury via inhibition of NLRP3 inflammasome activation in mice. Autophagy. 2016 Dec;12(12):2326-2343. [CrossRef]
- Copaescu A, et al. The role of IL-6 and other mediators in the cytokine storm associated with SARS-CoV-2 infection. J. Allergy Clin. Immunol. 2020;146:518–534 e511. [CrossRef]
- Thongchot S, Vidoni C, Ferraresi A, Loilome W, Khuntikeo N, Sangkhamanon S, Titapun A, Isidoro C, Namwat N. Cancer-Associated Fibroblast-Derived IL-6 Determines Unfavorable Prognosis in Cholangiocarcinoma by Affecting Autophagy-Associated Chemoresponse. Cancers (Basel). 2021 Apr 28;13(9):2134. PMID: 33925189; PMCID: PMC8124468. [CrossRef]
- Chia SB, Johnson BJ, Hu J, Valença-Pereira F, Chadeau-Hyam M, Guntoro F, Montgomery H, Boorgula MP, Sreekanth V, Goodspeed A, Davenport B, De Dominici M, Zaberezhnyy V, Schleicher WE, Gao D, Cadar AN, Petriz-Otaño L, Papanicolaou M, Beheshti A, Baylin SB, Guarnieri JW, Wallace DC, Costello JC, Bartley JM, Morrison TE, Vermeulen R, Aguirre-Ghiso JA, Rincon M, DeGregori J. Respiratory viral infections awaken metastatic breast cancer cells in lungs. Nature. 2025 Sep;645(8080):496-506. Epub 2025 Jul 30. PMID: 40739350; PMCID: PMC12422975. [CrossRef]
- Xia X. Detailed Dissection and Critical Evaluation of the Pfizer/BioNTech and Moderna mRNA Vaccines. Vaccines (Basel). 2021 Jul 3;9(7):734. PMID: 34358150; PMCID: PMC8310186. [CrossRef]
- Riley TP, Chou HT, Hu R, et al. Enhancing the prefusion conformational stability of SARS-CoV-2 spike protein through structure-guided design. Front Immunol. 2021;12:660198.
- Ogata AF, Cheng CA, Desjardins M, Senussi Y, Sherman AC, Powell M, Novack L, Von S, Li X, Baden LR, Walt DR. Circulating Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Vaccine Antigen Detected in the Plasma of mRNA-1273 Vaccine Recipients. Clin Infect Dis. 2022 Mar 1;74(4):715-718. PMID: 34015087; PMCID: PMC8241425. [CrossRef]
- Brogna C, Cristoni S, Marino G, Montano L, Viduto V, Fabrowski M, Lettieri G, Piscopo M. Detection of recombinant Spike protein in the blood of individuals vaccinated against SARS-CoV-2: Possible molecular mechanisms. Proteomics Clin Appl. 2023 Nov;17(6):e2300048. Epub 2023 Aug 31. PMID: 37650258. [CrossRef]
- Castruita JAS, Schneider UV, Mollerup S, Leineweber TD, Weis N, Bukh J, Pedersen MS, Westh H. SARS-CoV-2 spike mRNA vaccine sequences circulate in blood up to 28 days after COVID-19 vaccination. APMIS. 2023 Mar;131(3):128-132. Epub 2023 Jan 29. PMID: 36647776; PMCID: PMC10107710. [CrossRef]
- Boros LG, Kyriakopoulos AM, Brogna C, Piscopo M, McCullough PA, Seneff S. Long-lasting, biochemically modified mRNA, and its frameshifted recombinant spike proteins in human tissues and circulation after COVID-19 vaccination. Pharmacol Res Perspect. 2024 Jun;12(3):e1218. PMID: 38867495; PMCID: PMC11169277. [CrossRef]
- Bansal S, Perincheri S, Fleming T, Poulson C, Tiffany B, Bremner RM, Mohanakumar T. Cutting Edge: Circulating Exosomes with COVID Spike Protein Are Induced by BNT162b2 (Pfizer-BioNTech) Vaccination prior to Development of Antibodies: A Novel Mechanism for Immune Activation by mRNA Vaccines. J Immunol. 2021 Nov 15;207(10):2405-2410. Epub 2021 Oct 15. PMID: 34654691; PMCID: PMC11073804. [CrossRef]
- Yasmin F, Najeeb H, Naeem U, Moeed A, Atif AR, Asghar MS, Nimri N, Saleem M, Bandyopadhyay D, Krittanawong C, Fadelallah Eljack MM, Tahir MJ, Waqar F. Adverse events following COVID-19 mRNA vaccines: A systematic review of cardiovascular complication, thrombosis, and thrombocytopenia. Immun Inflamm Dis. 2023 Mar;11(3):e807. PMID: 36988252; PMCID: PMC10022421. [CrossRef]
- Choi JY, Lee Y, Park NG, Kim MS, Rhie SJ. Serious Safety Signals and Prediction Features Following COVID-19 mRNA Vaccines Using the Vaccine Adverse Event Reporting System. Pharmaceuticals (Basel). 2024 Mar 10;17(3):356. PMID: 38543142; PMCID: PMC10974993. [CrossRef]
- Zhang Z, Shen Q, Chang H. Vaccines for COVID-19: A Systematic Review of Immunogenicity, Current Development, and Future Prospects. Front Immunol. 2022 Apr 27;13:843928. PMID: 35572592; PMCID: PMC9092649. [CrossRef]
- Hajjo R, Sabbah DA, Tropsha A. Analyzing the Systems Biology Effects of COVID-19 mRNA Vaccines to Assess Their Safety and Putative Side Effects. Pathogens. 2022 Jun 29;11(7):743. PMID: 35889989; PMCID: PMC9320269. [CrossRef]
- Faksova K, Walsh D, Jiang Y, Griffin J, Phillips A, Gentile A, Kwong JC, Macartney K, Naus M, Grange Z, Escolano S, Sepulveda G, Shetty A, Pillsbury A, Sullivan C, Naveed Z, Janjua NZ, Giglio N, Perälä J, Nasreen S, Gidding H, Hovi P, Vo T, Cui F, Deng L, Cullen L, Artama M, Lu H, Clothier HJ, Batty K, Paynter J, Petousis-Harris H, Buttery J, Black S, Hviid A. COVID-19 vaccines and adverse events of special interest: A multinational Global Vaccine Data Network (GVDN) cohort study of 99 million vaccinated individuals. Vaccine. 2024 Apr 2;42(9):2200-2211. Epub 2024 Feb 12. PMID: 38350768. [CrossRef]
- Bruce Yu Y, Taraban MB, Briggs KT. All vials are not the same: Potential role of vaccine quality in vaccine adverse reactions. Vaccine. 2021 Oct 29;39(45):6565-6569. Epub 2021 Oct 6. PMID: 34625289; PMCID: PMC8492451. [CrossRef]
- Tinari S, The EMA covid-19 data leak, and what it tells us about mRNA instability BMJ 2021; 372:n627. (Published 10 March 2021). [CrossRef]
- Mulroney TE, Pöyry T, Yam-Puc JC, Rust M, Harvey RF, Kalmar L, Horner E, Booth L, Ferreira AP, Stoneley M, Sawarkar R, Mentzer AJ, Lilley KS, Smales CM, von der Haar T, Turtle L, Dunachie S, Klenerman P, Thaventhiran JED, Willis AE. N1-methylpseudouridylation of mRNA causes +1 ribosomal frameshifting. Nature. 2024 Jan;625(7993):189-194. Epub 2023 Dec 6. PMID: 38057663; PMCID: PMC10764286. [CrossRef]
- Guillen-Angel M, Roignant JY. Exploring pseudouridylation: dysregulation in disease and therapeutic potential. Curr Opin Genet Dev. 2024 Jun 3;87:102210. Epub ahead of print. PMID: 38833893. [CrossRef]
- Kim KQ, Burgute BD, Tzeng SC, Jing C, Jungers C, Zhang J, Yan LL, Vierstra RD, Djuranovic S, Evans BS, Zaher HS. N1-methylpseudouridine found within COVID-19 mRNA vaccines produces faithful protein products. Cell Rep. 2022 Aug 30;40(9):111300. Epub 2022 Aug 15. PMID: 35988540; PMCID: PMC9376333. [CrossRef]
- Rubio-Casillas A, Cowley D, Raszek M, Uversky VN, Redwan EM. Review: N1-methyl-pseudouridine (m1Ψ): Friend or foe of cancer? Int J Biol Macromol. 2024 May;267(Pt 1):131427. doi: 10.1016/j.ijbiomac.2024.131427. Epub 2024 Apr 5. Erratum in: Int J Biol Macromol. 2024 Jun;270(Pt 2):132447. doi: 10.1016/j.ijbiomac.2024.132447. PMID: 38583833.
- Föhse K, Geckin B, Zoodsma M, Kilic G, Liu Z, Röring RJ, Overheul GJ, van de Maat J, Bulut O, Hoogerwerf JJ, Ten Oever J, Simonetti E, Schaal H, Adams O, Müller L, Ostermann PN, van de Veerdonk FL, Joosten LAB, Haagmans BL, van Crevel R, van Rij RP, GeurtsvanKessel C, de Jonge MI, Li Y, Domínguez-Andrés J, Netea MG. The impact of BNT162b2 mRNA vaccine on adaptive and innate immune responses. Clin Immunol. 2023 Oct;255:109762. Epub 2023 Sep 6. PMID: 37673225. [CrossRef]
- Sittplangkoon C, Alameh MG, Weissman D, Lin PJC, Tam YK, Prompetchara E, Palaga T. mRNA vaccine with unmodified uridine induces robust type I interferon-dependent anti-tumor immunity in a melanoma model. Front Immunol. 2022 Oct 14;13:983000. [CrossRef]
- Cheng H, Yu J, Wong CC. Adenosine-to-inosine RNA editing in cancer: molecular mechanisms and downstream targets. Protein Cell. 2025 Jun 20;16(6):391-417. doi: 10.1093/procel/pwae039. Erratum in: Protein Cell. 2024 Nov 08:pwae062. doi: 10.1093/procel/pwae062. PMID: 39126156; PMCID: PMC12187073.
- Jin YY, Liang YP, Pan JQ, Huang WH, Feng YM, Sui WJ, Yu H, Tang XD, Zhu L, Chen JH. RNA editing in response to COVID-19 vaccines: unveiling dynamic epigenetic regulation of host immunity. Front Immunol. 2024 Sep 6;15:1413704. PMID: 39308856; PMCID: PMC11413487. [CrossRef]
- Wang Y, Kuramitsu Y, Kitagawa T, Baron B, Yoshino S, Maehara S, Maehara Y, Oka M, Nakamura K. Cofilin-phosphatase slingshot-1L (SSH1L) is over-expressed in pancreatic cancer (PC) and contributes to tumor cell migration. Cancer Lett. 2015 May 1;360(2):171-6. Epub 2015 Feb 12. PMID: 25684665. [CrossRef]
- Seneff S, Nigh G, Kyriakopoulos AM, McCullough PA. Innate immune suppression by SARS-CoV-2 mRNA vaccinations: The role of G-quadruplexes, exosomes, and MicroRNAs. Food Chem Toxicol. 2022 Jun;164:113008. Epub 2022 Apr 15. PMID: 35436552; PMCID: PMC9012513. [CrossRef]
- Miglietta G, Russo M, Capranico G. G-quadruplex-R-loop interactions and the mechanism of anticancer G-quadruplex binders. Nucleic Acids Res. 2020 Dec 2;48(21):11942-11957. doi: 10.1093/nar/gkaa944. Erratum in: Nucleic Acids Res. 2021 Jan 11;49(1):595. doi: 10.1093/nar/gkaa1206. Erratum in: Nucleic Acids Res. 2021 Jun 4;49(10):6000-6001. doi: 10.1093/nar/gkab483. PMID: 33137181; PMCID: PMC7708042.
- König B, Kirchner JO. Methodological Considerations Regarding the Quantification of DNA Impurities in the COVID-19 mRNA Vaccine Comirnaty®. Methods Protoc. 2024 May 8;7(3):41. PMID: 38804335; PMCID: PMC11130870. [CrossRef]
- Kobbe R, Rau C, Schulze-Sturm U, Stahl F, Fonseca-Brito L, Diemert A, Lütgehetmann M, Addo MM, Arck P, Weskamm LM. Delayed Induction of Noninflammatory SARS-CoV-2 Spike-Specific IgG4 Antibodies Detected 1 Year After BNT162b2 Vaccination in Children. Pediatr Infect Dis J. 2024 Jul 30;43(12):1200–3. [CrossRef]
- Gao FX, Wu RX, Shen MY, Huang JJ, Li TT, Hu C, Luo FY, Song SY, Mu S, Hao YN, Han XJ, Wang YM, Li L, Li SL, Chen Q, Wang W, Jin AS. Extended SARS-CoV-2 RBD booster vaccination induces humoral and cellular immune tolerance in mice. iScience. 2022 Dec 22;25(12):105479. Epub 2022 Nov 2. PMID: 36338436; PMCID: PMC9625849. [CrossRef]
- Yu T, Wu Y, Liu J, Zhuang Y, Jin X, Wang L. The risk of malignancy in patients with IgG4-related disease: a systematic review and meta-analysis. Arthritis Res Ther. 2022 Jan 5;24(1):14. PMID: 34986892; PMCID: PMC8728936. [CrossRef]
- Wang H, Xu Q, Zhao C, Zhu Z, Zhu X, Zhou J, Zhang S, Yang T, Zhang B, Li J, Yan M, Liu R, Ma C, Quan Y, Zhang Y, Zhang W, Geng Y, Chen C, Chen S, Liu D, Chen Y, Tian D, Su M, Chen X, Gu J. An immune evasion mechanism with IgG4 playing an essential role in cancer and implication for immunotherapy. J Immunother Cancer. 2020 Aug;8(2):e000661. [CrossRef]
- Shrestha P, Ghoreyshi ZS, George JT. How modulation of the tumor microenvironment drives cancer immune escape dynamics. Sci Rep. 2025 Mar 1;15(1):7308. PMID: 40025156; PMCID: PMC11873109. [CrossRef]
- Mulligan, M.J., Lyke, K.E., Kitchin, N. et al. Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature 586, 589–593 (2020). [CrossRef]
- Gandolfo, C.; Anichini, G.; Mugnaini, M.; Bocchia, M.; Terrosi, C.; Sicuranza, A.; Gori Savellini, G.; Gozzetti, A.; Franchi, F.; Cusi, M.G. Overview of Anti-SARS-CoV-2 Immune Response Six Months after BNT162b2 mRNA Vaccine. Vaccines 2022, 10, 171.
- Zhang X, Li B, Lan T, Chiari C, Ye X, Wang K, Chen J. The role of interleukin-17 in inflammation-related cancers. Front Immunol. 2025 Jan 21;15:1479505. PMID: 39906741; PMCID: PMC11790576. [CrossRef]
- Alghamdi A, Hussain SD, Wani K, Sabico S, Alnaami AM, Amer OE, Al-Daghri NM. Altered Circulating Cytokine Profile Among mRNA-Vaccinated Young Adults: A Year-Long Follow-Up Study. Immun Inflamm Dis. 2025 Apr;13(4):e70194. PMID: 40202571; PMCID: PMC11980434. [CrossRef]
- Indraccolo S, Stievano L, Minuzzo S, Tosello V, Esposito G, Piovan E, Zamarchi R, Chieco-Bianchi L, Amadori A. Interruption of tumor dormancy by a transient angiogenic burst within the tumor microenvironment. Proc Natl Acad Sci U S A. 2006 Mar 14;103(11):4216-21. Epub 2006 Mar 6. PMID: 16537511; PMCID: PMC1449673. [CrossRef]
- Bakos T, Mészáros T, Kozma GT, Berényi P, Facskó R, Farkas H, Dézsi L, Heirman C, de Koker S, Schiffelers R, Glatter KA, Radovits T, Szénási G, Szebeni J. mRNA-LNP COVID-19 Vaccine Lipids Induce Complement Activation and Production of Proinflammatory Cytokines: Mechanisms, Effects of Complement Inhibitors, and Relevance to Adverse Reactions. Int J Mol Sci. 2024 Mar 22;25(7):3595. [CrossRef]
- Soyfer V, Gutfeld O, Shamai S, et al. COVID-19 vaccine-induced radiation recall phenomenon. Int J Radiat Oncol Biol Phys 2021. Volume 110, issue 4, P957-961. [CrossRef]
- Kim HJ, Kim MH, Choi MG, Chun EM. 1-year risks of cancers associated with COVID-19 vaccination: a large population-based cohort study in South Korea. Biomark Res. 2025 Sep 26;13(1):114. PMID: 41013858; PMCID: PMC12465339. [CrossRef]
- Goldman S, Bron D, Tousseyn T, Vierasu I, Dewispelaere L, Heimann P, Cogan E, Goldman M. Rapid Progression of Angioimmunoblastic T Cell Lymphoma Following BNT162b2 mRNA Vaccine Booster Shot: A Case Report. Front Med (Lausanne). 2021 Nov 25;8:798095. PMID: 34901098; PMCID: PMC8656165. [CrossRef]
- Brumfiel CM, Patel MH, DiCaudo DJ, Rosenthal AC, Pittelkow MR, Mangold AR. Recurrence of primary cutaneous CD30-positive lymphoproliferative disorder following COVID-19 vaccination. Leuk Lymphoma. 2021 Oct;62(10):2554-2555. [CrossRef]
- Sekizawa A, Hashimoto K, Kobayashi S, Kozono S, Kobayashi T, Kawamura Y, Kimata M, Fujita N, Ono Y, Obuchi Y, Tanaka Y. Rapid progression of marginal zone B-cell lymphoma after COVID-19 vaccination (BNT162b2): A case report. Front Med (Lausanne). 2022 Aug 1;9:963393. PMID: 35979213. [CrossRef]
- Zamfir MA, Moraru L, Dobrea C, Scheau AE, Iacob S, Moldovan C, Scheau C, Caruntu C, Caruntu A. Hematologic Malignancies Diagnosed in the Context of the mRNA COVID-19 Vaccination Campaign: A Report of Two Cases. Medicina (Kaunas). 2022 Jun 30;58(7):874. PMID: 35888593; PMCID: PMC9316988. [CrossRef]
- Batuhan Erdogdu, Olgu Erkin Cinar, Umit Yavuz Malkan, Salih Aksu, Haluk Demiroglu, Yahya Buyukasik, Hakan Goker, Nilgun Sayinalp,Ibrahim Celalettin Haznedarogluolgu Erkin. Hematopoietic Adverse Events Associated with BNT162b2 mRNA Covid-19 Vaccine. International Journal of Hematology and Oncology UHOD Number: 1 Volume: 32 Year: 2022;. [CrossRef]
- Çınar OE, Erdoğdu B, Karadeniz M, Ünal S, Malkan ÜY, Göker H, Haznedaroğlu İC. Comment on Zamfir et al. Hematologic Malignancies Diagnosed in the Context of the mRNA COVID-19 Vaccination Campaign: A Report of Two Cases. Medicina 2022, 58, 874. Medicina (Kaunas). 2022 Nov 1;58(11):1575. PMID: 36363532; PMCID: PMC9696869. [CrossRef]
- Mizutani M, Mitsui H, Amano T, Ogawa Y, Deguchi N, Shimada S, Miwa A, Kawamura T, Ogido Y. Two cases of axillary lymphadenopathy diagnosed as diffuse large B-cell lymphoma developed shortly after BNT162b2 COVID-19 vaccination. J Eur Acad Dermatol Venereol. 2022 Aug;36(8):e613-e615. Epub 2022 Apr 18. PMID: 35398921; PMCID: PMC9114986. [CrossRef]
- Revenga-Porcel L, Peñate Y, Granados-Pacheco F. Anaplastic large cell lymphoma at the SARS-CoV2 vaccine injection site. J Eur Acad Dermatol Venereol. 2023 Jan;37(1):e32-e34. Epub 2022 Oct 8. PMID: 36166359. [CrossRef]
- Bae E, Bae S, Vaysblat M, Abdelwahed M, Sarkar K, Bae S. Development of High-Grade Sarcoma After Second Dose of Moderna Vaccine. Cureus. 2023 Apr 15;15(4):e37612. PMID: 37197108; PMCID: PMC10184721. [CrossRef]
- Avallone G, Maronese CA, Conforti C, Fava P, Gargiulo L, Marzano AV, Massone C, Mastorino L, Paradisi A, Pileri A, Quaglino P, Rizzo N, Ribero S, Roccuzzo G, Tavoletti G, Vignoli CA, Zalaudek I, Berti E, Alberti-Violetti S. Real-world data on primary cutaneous lymphoproliferative disorders following SARS-CoV-2 vaccination: A multicentre experience from tertiary referral hospitals. J Eur Acad Dermatol Venereol. 2023 Apr;37(4):e451-e455. Epub 2022 Dec 19. PMID: 36469002; PMCID: PMC9877594. [CrossRef]
- Cavanna L, Grassi SO, Ruffini L, Michieletti E, Carella E, Palli D, Zangrandi A, Inzerilli N, Bernuzzi P, Di Nunzio C, Citterio C. Non-Hodgkin Lymphoma Developed Shortly after mRNA COVID-19 Vaccination: Report of a Case and Review of the Literature. Medicina (Kaunas). 2023 Jan 12;59(1):157. PMID: 36676781; PMCID: PMC9861188. [CrossRef]
- White E, Fazio N, Tourmouzis K, Ryu S, Finger PT, Sassoon J, Keresztes R, Chou T, Kaplowitz K, Honkanen R. Unilateral conjunctival Classic Kaposi Sarcoma following a COVID 19 booster. Am J Ophthalmol Case Rep. 2023 Dec 26;34:101986. PMID: 38601193; PMCID: PMC11004082. [CrossRef]
- Kyriakopoulos AM, Nigh G, McCullough PA, Olivier MD, Seneff S. Bell's palsy or an aggressive infiltrating basaloid carcinoma post-mRNA vaccination for COVID-19? A case report and review of the literature. EXCLI J. 2023 Sep 11;22:992-1011. PMID: 37927346; PMCID: PMC10620857. [CrossRef]
- Ang SY, Huang YF, Chang CT. Ph-Positive B-Cell Acute Lymphoblastic Leukemia Occurring after Receipt of Bivalent SARS-CoV-2 mRNA Vaccine Booster: A Case Report. Medicina (Kaunas). 2023 Mar 21;59(3):627. PMID: 36984629; PMCID: PMC10054610. [CrossRef]
- Wang Z, Muto R, Miyoshi H, Aoki M, Uesugi N, Murayama H, Masutani K, Hamasaki M. The first autopsy case of Epstein-Barr virus-positive marginal zone lymphoma that deteriorated after COVID-19 vaccination. Pathol Int. 2024 Feb;74(2):87-92. Epub 2023 Dec 20. PMID: 38116849. [CrossRef]
- Ueda Y, Sakai T, Yamada K, Arita K, Ishige Y, Hoshi D, Yanagisawa H, Iwao-Kawanami H, Kawanami T, Mizuta S, Fukushima T, Yamada S, Yachie A, Masaki Y. Fatal hemophagocytic lymphohistiocytosis with intravascular large B-cell lymphoma following coronavirus disease 2019 vaccination in a patient with systemic lupus erythematosus: an intertwined case. Immunol Med. 2024 Sep;47(3):192-199. Epub 2024 Apr 15. PMID: 38619098. [CrossRef]
- Farhat M, Zouein J, Abou Khater J, Sarkis AS, Helou J. A Case of Rapid Transformation of a Nail Matrix Nevi to Melanoma After Messenger RNA COVID-19 Vaccine: A Cause or a Coincidence? Cureus. 2024 Dec 24;16(12):e76312. PMID: 39850193; PMCID: PMC11756429. [CrossRef]
- Sano S. A case of metastatic breast carcinoma to the skin expressing SARS-CoV-2 spike protein possibly derived from mRNA vaccine. J Dermatol Sci. 2025 Oct 3:S0923-1811(25)00180-X. Epub ahead of print. PMID: 41076388. [CrossRef]
- Haji N Jr, Ali S, Wahashi EA, Khalid M, Ramamurthi K. Johnson and Johnson COVID-19 Vaccination Triggering Pheochromocytoma Multisystem Crisis. Cureus. 2021 Sep 22;13(9):e18196. PMID: 34707965; PMCID: PMC8536310. [CrossRef]
- Panou E, Nikolaou V, Marinos L, Kallambou S, Sidiropoulou P, Gerochristou M, Stratigos A. Recurrence of cutaneous T-cell lymphoma post viral vector COVID-19 vaccination. J Eur Acad Dermatol Venereol. 2022 Feb;36(2):e91-e93. Epub 2021 Oct 31. PMID: 34628691; PMCID: PMC9088536. [CrossRef]
- Tang WR, Hsu CW, Lee CC, Huang WL, Lin CY, Hsu YT, Chang C, Tsai MT, Hu YN, Hsu CH, Chen PL, Chow NH, Roan JN. A Case Report of Posttransplant Lymphoproliferative Disorder After AstraZeneca Coronavirus Disease 2019 Vaccine in a Heart Transplant Recipient. Transplant Proc. 2022 Jul-Aug;54(6):1575-1578. Epub 2021 Sep 30. PMID: 34702598; PMCID: PMC8481090. [CrossRef]
- Kreher MA, Ahn J, Werbel T, Motaparthi K. Subcutaneous panniculitis-like T-cell lymphoma after COVID-19 vaccination. JAAD Case Rep. 2022 Oct;28:18-20. Epub 2022 Aug 10. PMID: 35966352; PMCID: PMC9364717. [CrossRef]
- Veeraballi S, Patel A, Are G, Ramahi A, Chittamuri S, Shaaban H. A Case of Chronic Myelomonocytic Leukemia Unmasked After Receiving J&J COVID-19 Vaccine. Cureus. 2022 Jun 18;14(6):e26070. PMID: 35865440; PMCID: PMC9292133. [CrossRef]
- Martínez-Ortega JI, Ramirez Cibrian AG, Martinez-Jaramillo E, García Silva MDC. Sporadic Kaposi Sarcoma Following a COVID-19 Vaccine: Mere Coincidence or Something More? Cureus. 2024 Feb 9;16(2):e53925. PMID: 38465101; PMCID: PMC10924926. [CrossRef]
- Tenforde MW, Self WH, Adams K, Gaglani M, Ginde AA, McNeal T, Ghamande S, Douin DJ, Talbot HK, Casey JD, Mohr NM, Zepeski A, Shapiro NI, Gibbs KW, Files DC, Hager DN, Shehu A, Prekker ME, Erickson HL, Exline MC, Gong MN, Mohamed A, Henning DJ, Steingrub JS, Peltan ID, Brown SM, Martin ET, Monto AS, Khan A, Hough CL, Busse LW, Ten Lohuis CC, Duggal A, Wilson JG, Gordon AJ, Qadir N, Chang SY, Mallow C, Rivas C, Babcock HM, Kwon JH, Halasa N, Chappell JD, Lauring AS, Grijalva CG, Rice TW, Jones ID, Stubblefield WB, Baughman A, Womack KN, Rhoads JP, Lindsell CJ, Hart KW, Zhu Y, Olson SM, Kobayashi M, Verani JR, Patel MM; Influenza and Other Viruses in the Acutely Ill (IVY) Network. Association Between mRNA Vaccination and COVID-19 Hospitalization and Disease Severity. JAMA. 2021 Nov 23;326(20):2043-2054. PMID: 34734975; PMCID: PMC8569602. [CrossRef]
- Andrews N, Stowe J, Kirsebom F, Toffa S, Sachdeva R, Gower C, Ramsay M, Lopez Bernal J. Effectiveness of COVID-19 booster vaccines against COVID-19-related symptoms, hospitalization and death in England. Nat Med. 2022 Apr;28(4):831-837. Epub 2022 Jan 14. PMID: 35045566; PMCID: PMC9018410. [CrossRef]
- Adhikari B, Bednash JS, Horowitz JC, Rubinstein MP, Vlasova AN. Brief research report: impact of vaccination on antibody responses and mortality from severe COVID-19. Front Immunol. 2024 Feb 7;15:1325243. doi: 10.3389/fimmu.2024.1325243. Erratum in: Front Immunol. 2024 Feb 28;15:1384209. doi: 10.3389/fimmu.2024.1384209. PMID: 38390335; PMCID: PMC10883056.
- Berrino F, Donzelli A, Bellavite P, Malatesta G. COVID-19 vaccination and all-cause and non-COVID-19 mortality. A revaluation of a study carried out in an Italian Province. Epidemiol Prev. 2023 Nov-Dec;47(6):374-378. English. PMID: 38314545. [CrossRef]
- Teodori L, Osimani B, Isidoro C, Ramakrishna S. Mass versus personalized medicine against COVID-19 in the "system sciences" era. Cytometry A. 2022 Dec;101(12):995-999. Epub 2022 Jun 18. PMID: 35716116. [CrossRef]
- Boeckel GR, Hölscher SD, Bürger C, Jacob T, Krekeler C, Shumilov E, Reicherts C, Bleckmann A, Lenz G, Vollenberg R, Tepasse PR. Comprehensive Treatment of Hematological Patients with SARS-CoV-2 Infection Including Anti-SARS-CoV-2 Monoclonal Antibodies: A Single-Center Experience Case Series. Curr Oncol. 2022 Mar 26;29(4):2312-2325. [CrossRef]
- Janssen M, Leo A, Wolf C, Stenzinger M, Bartenschlager M, Brandt J, Sauer S, Schmitt M, Dreger P, Schlenk RF, Denkinger CM, Müller-Tidow C. Treatment of chronic COVID-19 with convalescent/postvaccination plasma in patients with hematologic malignancies. Int J Cancer. 2024 Aug 15;155(4):618-626. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



