
Submitted:
28 October 2025
Posted:
29 October 2025
You are already at the latest version
Abstract
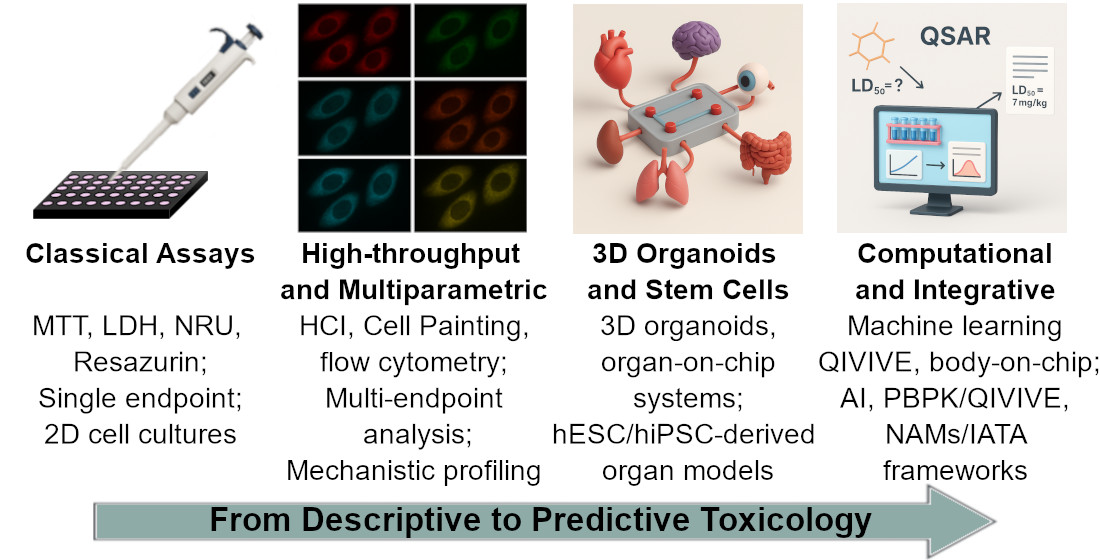
Cytotoxicity testing remains a cornerstone of modern toxicology, providing essential insight into how chemicals and drugs affect cell viability and function. Classical colourimetric assays such as MTT, LDH release, and neutral red uptake established the methodological basis of in vitro toxicology and continue to serve as regulatory benchmarks. However, their limited mechanistic depth and physiological relevance have prompted the field to evolve towards more predictive and human-centred approaches.
Recent advances in high-content imaging, flow cytometry, and real-time impedance analysis have transformed cytotoxicity testing into a multiparametric discipline capable of detecting subtle, adaptive, and sub-lethal cellular responses. Large-scale initiatives such as Tox21 and ToxCast, supported by computational pipelines like tcpl, have standardised data interpretation and improved reproducibility. In parallel, three-dimensional organoid cultures, organ-on-chip platforms, and bioprinted constructs now replicate the architec-ture, perfusion, and metabolic activity of human tissues, enhancing translational accuracy.
Stem cell–based models using human embryonic and induced pluripotent stem cells provide ethically sustainable systems for organ-specific and developmental toxicity testing, while in silico tools - QSAR, machine learning, and physiologically based pharmacokinetic (PBPK) modelling - enable quantitative in vitro–in vivo extrapolation (QIVIVE). Together, these developments underpin New Approach Methodologies (NAMs) and Integrated Approaches to Testing and Assessment (IATA), marking the transition from descriptive assays to predictive, mechanism-anchored frameworks that advance both biomedical research and regulatory science.
Keywords:
cytotoxicity assays
; toxicity testing
; 3D cell culture
; organ-on-a-chip
; high-content imaging
; in silico models
; integrated testing strategies (IATA)
1. Introduction
Cytotoxicity testing remains a central pillar of modern toxicology, linking cellular responses to hazard identification and risk evaluation in biomedical research, pharmaceutical development, and chemical safety assessment. By quantifying changes in cell viability and function, these assays provide critical data for understanding toxic mechanisms and supporting regulatory decisions. Agencies such as the United States Food and Drug Administration (FDA), the European Medicines Agency (EMA), and the Organisation for Economic Co-operation and Development (OECD) require cytotoxicity data before compounds advance to preclinical or clinical stages. In parallel, the ethical principles of the 3Rs - Replacement, Reduction, and Refinement - have accelerated the adoption of non-animal testing strategies [1,2]. The influential report Toxicity Testing in the 21st Century further highlighted the need for human-relevant, mechanism-based approaches [3].
Historically, cytotoxicity was evaluated using two-dimensional (2D) cell cultures combined with simple colourimetric or fluorometric readouts. The MTT assay, which measures mitochondrial reduction of tetrazolium salts, became a long-standing standard [4,5]. Complementary methods such as lactate dehydrogenase (LDH) release, reflecting plasma membrane integrity [6], and neutral red uptake (NRU), which probes lysosomal activity [6,7], provided alternative functional perspectives. Although inexpensive and reproducible, these classical assays offer only limited mechanistic insight and show variable correlation with in vivo outcomes [8,9]. Nonetheless, they established the methodological foundation of in vitro toxicology and remain useful as reference points in regulatory practice [10,11].
As toxicology advanced, the limitations of single-endpoint assays became increasingly evident. High false-positive rates in classical genotoxicity tests - especially in p53-deficient rodent cell lines - emphasised the need for more predictive, human-based models [12]. The emergence of large-scale high-throughput initiatives such as Tox21 [13] and ToxCast [14] enabled systematic profiling of thousands of compounds across multiple cellular pathways [15]. At the same time, stem-cell-derived systems, including human embryonic stem cells (hESCs) and induced pluripotent stem cells (hiPSCs), expanded the ability to study developmental and organ-specific toxicity in human-relevant settings [16,17]. The rapid growth of nanotechnology introduced new challenges and gave rise to nanotoxicology - a distinct field requiring dedicated in vitro methods to address oxidative stress and nanoparticle-specific mechanisms [18,19,20]. Meanwhile, the transition from 2D monolayers to three-dimensional (3D) culture systems improved physiological relevance and predictive accuracy [21].
Recent technological progress has transformed the scope and ambition of cytotoxicity testing. Organoids derived from pluripotent stem cells now reproduce the complex architecture and functionality of native tissues, improving the prediction of hepatotoxicity, nephrotoxicity, and ocular or dermal injury [22,23,24]. Microfluidic organ-on-chip systems allow dynamic investigation of absorption, distribution, metabolism, and excretion (ADME), and provide mechanistic insight into tissue–tissue interactions and systemic responses [25,26,27]. Alongside these in vitro innovations, computational tools - such as quantitative structure–activity relationship (QSAR) modelling, machine learning, and physiologically based pharmacokinetic (PBPK) or quantitative in vitro–in vivo extrapolation (QIVIVE) models - now enable quantitative translation of laboratory data into realistic human exposure scenarios [28,29,30,31].
Together, these advances form the foundation of New Approach Methodologies (NAMs) and Integrated Approaches to Testing and Assessment (IATA), which are now being incorporated into OECD, FDA, and EMA regulatory frameworks [32,33,34]. Validated 3D skin and corneal models have already replaced traditional animal-based assays such as the Draize test [32,35,36].
Overall, the evolution of cytotoxicity testing reflects a clear shift from simple viability measurements to integrated, mechanistic, and human-relevant systems. These new platforms bridge experimental biology, computational modelling, and regulatory science, advancing both the predictive power and ethical sustainability of toxicological assessment. The present review provides a comprehensive overview of this transition - from classical in vitro assays to high-throughput, stem-cell-based, in silico, and organ-on-chip technologies - and discusses their growing significance for biomedical research and regulatory decision-making.
2. Classical Cytotoxicity Assays: Foundations of In Vitro Toxicology
Classical cytotoxicity assays remain a fundamental part of in vitro toxicology. They provide accessible, reproducible, and straightforward means to evaluate cell viability and detect toxic responses. Although their limitations are well recognised, these methods continue to serve as essential reference points in both research and regulatory testing. Among the most widely applied assays are tetrazolium reduction (MTT), LDH release, NRU, resazurin reduction, and total protein or biomass quantification. Together, they form the practical foundation of routine cytotoxicity assessment, offering a balance between simplicity and reliability.
To ensure data quality and comparability, careful attention to assay design, appropriate controls, and transparent data reporting is critical. Key recommendations for good experimental practice - including validation steps, interference checks, and result normalisation - are summarised in Box 1.
- Box 1. Best practices for classical cytotoxicity assays
- Assay design and execution
- Verify signal linearity with cell density (5×10³–2×10⁴ cells/well in 96-well plates);
- Optimize dye incubation times (e.g., 2–4 h MTT; 3 h NRU) and report conditions;
- Control for LDH background in serum; use serum-free or heat-inactivated controls.
- Controls and interference checks
- Screen test compounds for intrinsic fluorescence or colour; include “no-cell” blanks;
- Assess dye adsorption by nanomaterials and confirm with independent endpoints;
- Use appropriate positive and negative controls (e.g., Triton X-100, staurosporine) to verify responsiveness.
- Data processing and normalization
- Subtract background from blank wells;
- Normalise viability to untreated controls (100 %) and maximal lysis (0 %);
- Report raw data, at least three biological replicates with technical triplicates, and variability metrics.
- Reporting transparency
- Specify seeding density, passage number, medium composition, incubation time, dye concentration, and detection settings;
- Describe curve fitting and statistical methods clearly;
- Note any deviations from OECD or ISO guidelines.
Tetrazolium-based assays such as MTT estimate cellular metabolic activity by monitoring the enzymatic reduction of tetrazolium salts into insoluble formazan crystals through mitochondrial and cytosolic dehydrogenases. Long regarded as the “gold standard” for assessing cytotoxicity, the MTT assay is now known to be susceptible to several artefacts, including non-specific reduction by test compounds or medium components, non-linear responses to cell number, and difficulties in formazan solubilisation [8,37]. As a result, data derived from MTT assays should be interpreted with caution and ideally confirmed using independent endpoints.
LDH release assays offer a more direct measure of plasma membrane integrity by quantifying extracellular enzyme activity through coupled colourimetric reactions. Their simplicity makes them attractive for routine use, yet accuracy can be affected by serum background, spontaneous leakage from stressed cells, or chemical interference. Combining LDH measurements with complementary readouts - such as proliferation or cell-cycle analyses - enhances the mechanistic interpretation of cytotoxic effects [6,38,39].
The NRU assay evaluates lysosomal function by measuring the accumulation of neutral red dye in viable cells. Compared with MTT, NRU is often more sensitive to early lysosomal stress, but results can be influenced by pH, incubation time, or lysosomal stability [7,40].
Resazurin (alamarBlue) reduction provides a non-destructive metabolic endpoint that allows repeated, same-well measurements and long-term monitoring. It is typically more sensitive and less variable than MTT or LDH, although very high metabolic activity can cause premature signal saturation [41,42].
Protein- and biomass-based assays such as sulforhodamine B (SRB) or Bradford staining quantify total cellular mass independently of metabolism. When used alongside metabolic assays, they help differentiate cytostatic from cytotoxic effects and reduce the risk of misinterpretation [43,44].
Comparative analyses consistently show that no single assay provides universally reliable results. NRU and resazurin methods often detect early toxicity more effectively, whereas MTT and LDH may underestimate subtle effects or yield higher variability [41]. Experimental artefacts - such as medium composition influencing dye uptake or nanoparticle adsorption of chromogenic reagents - can further confound outcomes [45,46,47]. Consequently, multiparametric strategies combining at least two independent endpoints are now considered best practice [43,48]. Common sources of error and assay-specific pitfalls are summarised in Box 2.
Beyond their technical advantages, classical cytotoxicity assays continue to play a key role in regulatory and screening contexts. In large-scale initiatives such as ToxCast, non-specific cytotoxicity signals often dominate and can obscure mechanistic responses [14]. Viability endpoints therefore remain indispensable for normalising data and guiding interpretation [39]. Even in advanced 3D organoid and spheroid models, commercially available cytotoxicity kits frequently show inter-donor or inter-platform variability, highlighting both the ongoing relevance of these classical assays and the need for multiparametric calibration [49]. Within tiered testing frameworks, they typically represent the first step of evaluation, preceding mechanistic, high-content, or omics-based analyses [50].
- Box 2. Common pitfalls in classical cytotoxicity assays
- MTT: non-specific reduction by compounds or medium; insoluble formazan crystals; metabolic stimulation mistaken for viability [37];
- NRU: dependence on pH or lysosomal health; false cytotoxicity when lysosomes are targeted [40];
- Resazurin: over-reduction in highly active cells; fluorescence quenching by test compounds [42];
- Protein/biomass assays: variability in fixation or staining; insensitivity to metabolic suppression without cell loss [44];
3. Transition from Viability Endpoints to Mechanistic Approaches
The limitations of single-endpoint viability assays have become increasingly apparent in contemporary toxicology. For many years, methods such as MTT, LDH, and NRU provided reliable first-tier screening tools, yet their reductionist perspective captured only a narrow view of cellular injury. As research and regulation have moved towards more human-relevant, mechanistic, and predictive approaches, cytotoxicity testing has undergone a marked transformation. The discipline now embraces high-throughput, multiparametric, and physiologically relevant models that provide richer and more quantitative insight into cellular responses. This evolution represents more than a technical improvement - it signifies a conceptual shift in how toxicity is understood, measured, and integrated into risk assessment.
3.1. From Viabifitnesslity to High-Throughput Screening
A decisive step forward came with the introduction of quantitative high-throughput screening (qHTS) technologies, developed through large-scale programmes such as Tox21 [15] and ToxCast [51]. These initiatives screened thousands of industrial chemicals, pharmaceuticals, and environmental contaminants in miniaturised 1536-well formats, producing extensive datasets that captured activity across nuclear receptor, stress-response, and enzyme-inhibition pathways [52,53]. One clear lesson emerged from these studies: broad cytotoxicity effects often dominated assay outcomes, obscuring genuine pathway-specific responses and complicating interpretation. For instance, in oestrogen-receptor reporter assays, apparent activity was frequently driven by non-monotonic dose–response curves or secondary stress effects rather than by true receptor engagement [54].
To prevent such misinterpretation, viability assays were incorporated into Tox21 [15] and ToxCast [51] not as primary endpoints but as parallel counterscreens. In practice, this meant that compounds showing apparent mechanistic activity were re-examined for their impact on cell viability: if the signal loss coincided with reduced viability, the response was classified as artefactual [52,55]. Mechanistic studies using dendrimers provided a concrete example - demonstrating that elevated cytotoxicity was accompanied by apoptotic signalling and TRAIL-mediated cell death in chronic lymphocytic leukaemia models [56].
Standardised data-analysis pipelines also improved reproducibility. The ToxCast Pipeline for Curve Fitting (tcpl) and curated curve-classification schemes [51,57] enabled consistent handling of replicates and background correction. Collectively, these developments transformed high-throughput screening from a largely descriptive exercise into a mechanistically anchored, data-driven paradigm - laying the foundation for truly multiparametric approaches.
3.2. Multiparametric and High-Content Imaging Approaches
A second wave of innovation in cytotoxicity testing was driven by the rise of high-content imaging (HCI) and phenotypic profiling technologies, which were designed to move beyond binary viability readouts. HCI integrates automated microscopy with quantitative image analysis, capturing a wide range of cellular features - nuclear and mitochondrial morphology, lysosomal integrity, cytoskeletal organisation, and more - across thousands of individual cells [58,59]. These multiparametric datasets reveal early, adaptive, or sub-lethal effects that conventional endpoint assays often miss. Among the most influential frameworks, the Cell Painting assay combines multiplexed fluorescent labelling of key organelles with computational feature extraction to generate morphological fingerprints that cluster structurally diverse compounds by their mechanisms of action [60,61].
When paired with transcriptomic or metabolomic profiling, these morphological signatures enable predictive modelling of toxicity pathways. Early comparative studies demonstrated that imaging-based cytotoxicity assays combined with proliferation markers were more sensitive than classical metabolic assays such as MTT or LDH [58]. Specialised variants have since emerged, including the BlueScreen HC genotoxicity assay, which incorporates DNA-damage reporters [62], and microglia-focused imaging platforms that quantify phagocytosis and cell health [58]. Crucially, these systems can detect adaptive stress responses well below overt cytotoxic thresholds, offering a window into the early stages of cell injury.
Phenotypic profiling has therefore transformed cytotoxicity assessment from a descriptive measure of cell death into a mechanistic discipline at the interface of cell biology, cheminformatics, and predictive toxicology [63].
Beyond imaging, impedance-based systems such as the xCELLigence Real-Time Cell Analysis (RTCA) platform provide continuous, label-free monitoring of proliferation, adhesion, and cell death. By recording time-resolved fluctuations in cell index, RTCA distinguishes transient stress from irreversible damage, capturing subtle morphological dynamics that static assays like MTT cannot resolve [64,65,66].
Flow cytometry represents another cornerstone of multiparametric cytotoxicity analysis, allowing simultaneous detection of apoptosis, mitochondrial depolarisation, and oxidative stress at the single-cell level [67]. Fluorescence-based measurements of Annexin V/PI staining, reactive oxygen species (ROS), and caspase activation provide detailed mechanistic information [68,69]. Because fluorescence detection is largely unaffected by nanoparticle interference, flow cytometry remains particularly valuable where optical artefacts compromise colourimetric assays [70,71].
Confocal microscopy complements these approaches by visualising intracellular compound distribution, including fluorescent nanocarriers, and identifying organelle-specific accumulation linked to oxidative stress or membrane disruption [70,71]. Integrated live-cell confocal imaging adds a spatial dimension, confirming the localisation of toxicity pathways and supporting mechanistic classification of compound-induced injury.
Together, these multiparametric and real-time analytical methods have redefined the purpose of cytotoxicity testing. Rather than serving merely to identify toxic hits, they now function as mechanistic tools that illuminate the pathways of cellular perturbation, bridging discovery toxicology with systems biology.
3.3. Refining Genotoxicity Assays to Reduce False Outcomes
Despite decades of routine use, in vitro genotoxicity assays have long struggled with issues of reproducibility and specificity. High false-positive rates - particularly in p53-deficient rodent cell lines - have frequently resulted in costly and unnecessary follow-up investigations [12]. Over the past decade, however, a series of methodological refinements has greatly improved both the human relevance and reliability of these assays.
One of the most effective improvements has been the use of p53-competent human cell lines, which significantly reduces spurious DNA-damage responses [72]. In addition, the introduction of refined viability parameters such as Relative Population Doubling (RPD) and Relative Increase in Cell Counts (RICC) has enhanced the ability to distinguish between general cytotoxicity and genuine genotoxicity [73]. Comparative studies consistently show that human cell lines respond more predictably to known genotoxicants than rodent systems, further supporting their use in human-relevant testing strategies [74].
In parallel, orthogonal high-throughput assays such as the CometChip - a microarray adaptation of the classical comet assay - have enabled the simultaneous quantification of DNA strand breaks across hundreds of samples [75]. By combining scalability with mechanistic endpoints, platforms like CometChip bridge the gap between traditional low-throughput tests and regulatory applications, reducing both false-positive and false-negative outcomes.
Together, these advances illustrate how improved methodological precision and mechanistic understanding can resolve many of the historical weaknesses of in vitro genotoxicity testing, strengthening its role in modern toxicological assessment
3.4. Bridging to Three-Dimensional Cultures and Organoids
A major turning point in cytotoxicity testing has been the emergence of 3D cultures and organoids, which more accurately reproduce tissue architecture, cell–cell communication, and physiological gradients absent from traditional 2D monolayers. By better reflecting the structural and functional complexity of native tissues, 3D systems offer markedly improved translational relevance and predictive power.
Co-culture spheroid models that integrate tumour and immune cells now enable real-time monitoring of immune-mediated cytotoxicity using luminescence-based killing assays and multicolour flow cytometry [76,77]. Comparable approaches have been applied to evaluate CAR-T cell–induced cytotoxicity in high-throughput settings, providing reproducible platforms for immunotoxicity research [78]. Microfluidic systems such as CACI-IMPACT further enhance these models by allowing continuous perfusion and kinetic imaging of cytotoxic responses under dynamic flow conditions [79].
Beyond oncology, the use of 3D neuronal spheroid cultures, such as LUHMES-derived models, facilitates mechanistic neurotoxicity screening [80], while liver organoids and liver-on-chip platforms surpass conventional 2D hepatocyte assays in predicting drug-induced liver injury [81,82]. Comparative proteomic and transcriptomic analyses consistently demonstrate distinct mechanistic signatures between 2D and 3D models, highlighting the superior physiological fidelity of the latter [83,84,85].
Together, these innovations bridge classical in vitro testing with in vivo relevance and align closely with the principles of NAMs and IATA [86]. Within the IATA framework, 3D cultures and organoids are increasingly integrated with in silico models, microphysiological systems, and omics datasets to provide human-relevant, non-animal evidence for toxicity evaluation [87]. Defined Approaches (DAs) combine outcomes from 3D assays with QSAR or read-across models, while PBPK and QIVIVE modelling translate in vitro activity into human-equivalent doses [88,89].
Practical recommendations for experimental design and quality control during this transition are summarised in Box 3.
- Box 3. Practical guidance for modern in vitro cytotoxicity studies.
4. Stem Cell-Based Models in Cytotoxicity Testing
Pluripotent stem cell–based models have become indispensable to modern cytotoxicity testing, providing human-relevant systems that overcome many of the limitations associated with immortalised tumour cell lines. Both hESCs and hiPSCs possess the remarkable capacity to differentiate into virtually any cell type, offering a unique platform to study xenobiotic effects across different developmental stages and organ systems [92,93,94]. By enabling controlled differentiation into functionally mature cells, these models allow researchers to investigate cellular mechanisms underlying toxicity within a physiologically meaningful human context.
Together, hESC- and hiPSC-derived systems have transformed toxicology from largely descriptive testing into a mechanistic science - one capable of linking cellular perturbations to adverse outcomes with unprecedented relevance for human health.
4.1. Human Embryonic Stem Cells (hESCs)
hESCs are derived from the inner cell mass of pre-implantation blastocysts, typically obtained from surplus embryos generated during in vitro fertilisation and donated with informed consent [95]. Their inherent pluripotency and virtually unlimited capacity for self-renewal make them a powerful platform for generating physiologically relevant human cell types in vitro. Over the past decade, refined differentiation protocols have enabled the establishment of diverse hESC-based models suited for organ-specific cytotoxicity testing [92,94].
Among these, hESC-derived cardiomyocytes have become benchmark systems for detecting drug-induced cardiotoxicity and contractility disturbances [96], while hepatocyte-like cells are increasingly employed to evaluate metabolism-dependent hepatotoxicity and drug-induced liver injury [97,98]. Neural and neuronal progenitor lineages derived from hESCs are widely used in developmental neurotoxicity studies [99,100], and epithelial derivatives from intestinal or pulmonary differentiation pathways now serve as human-relevant models for assessing mucosal and respiratory toxicity [101,102].
At a mechanistic level, transcriptomic and epigenetic profiling of differentiating hESCs exposed to environmental or pharmaceutical toxicants has revealed characteristic developmental hazard signatures [92]. These reproducible molecular fingerprints - defined by changes in gene expression and chromatin regulation - emerge when early developmental processes are perturbed. Model teratogens such as methylmercury, valproic acid, and all-trans retinoic acid elicit distinct transcriptional responses associated with neurogenesis, morphogenesis, and retinoid signalling. Such molecular fingerprints provide mechanistic evidence of early embryotoxicity and enable quantitative discrimination between toxic and non-toxic compounds. This level of mechanistic resolution demonstrates how hESC-based assays bridge molecular perturbations with adverse developmental outcomes, advancing predictive toxicology beyond descriptive endpoints.
Despite their scientific advantages, the use of hESC-based systems remains ethically and legally constrained in many regions. Access to approved lines is limited, and differentiated derivatives can exhibit partial immunogenicity [103]. Moreover, the persistence of residual undifferentiated cells presents a teratoma-formation risk, requiring rigorous purification and quality-control measures [104,105].
Overall, hESCs have laid the conceptual and methodological foundation for human developmental toxicity testing. Yet, their ethical and technical constraints have spurred the development of reprogrammed alternatives such as hiPSCs, which now represent a versatile and ethically sustainable next step in predictive toxicology.
4.2. Induced Pluripotent Stem Cells (hiPSCs)
HiPSCs are generated by reprogramming adult somatic cells - most commonly fibroblasts, peripheral blood mononuclear cells, or urinary epithelial cells - through enforced expression of the Yamanaka factors OCT4, SOX2, KLF4, and c-MYC [17,93,106,107]. This discovery revolutionised stem-cell biology and provided an ethically acceptable, patient-specific alternative to embryonic material.
Since their introduction, hiPSC-based models have become integral to contemporary toxicology. Among their most established applications are hiPSC-derived cardiomyocytes (hiPSC-CMs), which faithfully reproduce human electrophysiological and contractile properties and have proven highly effective for predicting chemotherapy- and drug-induced cardiotoxicity [96,108,109]. Hepatocyte-like cells and liver organoids derived from hiPSCs recapitulate key metabolic and cholestatic functions, enabling high-content and mechanistic evaluation of drug-induced liver injury [110,111]. Similarly, neuron-rich cultures and cerebral organoids have become essential tools for developmental neurotoxicity studies, capturing molecular and structural alterations caused by teratogens such as thalidomide [112] and environmental pollutants like perfluorooctanoic acid [113].
Beyond these core applications, hiPSCs enable the development of renal and ocular toxicity models - including nephron-like constructs [114] and retina-on-a-chip systems [115] - as well as complex multi-lineage organoids for studying cross-tissue interactions, such as combined cardiac–hepatic or tumour–microenvironment responses [116]. Because they retain the genetic background of the donor, hiPSC-based systems also support the emerging field of precision toxicology, allowing inter-individual susceptibility to xenobiotics to be explored directly in vitro.
Despite these advantages, challenges remain. Inter-line variability, incomplete cellular maturation, and batch-dependent differences in differentiation efficiency continue to limit reproducibility and regulatory acceptance [117]. Addressing these issues will be essential for the routine implementation of hiPSC-based assays in safety assessment pipelines.
In summary, hiPSCs combine ethical acceptability with patient specificity, providing a versatile bridge between mechanistic toxicology and personalised medicine.
4.3. Applications in Developmental and Organ-Specific Toxicity
The introduction of pluripotent stem cell–based assays has reshaped developmental toxicity testing, driving a shift from animal teratogenicity models to human-relevant, mechanistically anchored systems. Integrated platforms such as PluriLum and ReproTracker combine controlled PSC differentiation with transcriptomic, proteomic, and imaging readouts to quantitatively map teratogenic signatures. When integrated with PBPK modelling, these assays enable QIVIVE, substantially improving the prediction of human developmental hazards [118,119].
Organ-specific applications have expanded across multiple tissue types, bringing a level of physiological depth previously unattainable in in vitro cytotoxicity testing. In the cardiovascular field, hiPSC-CMs and 3D cardiac organoids have become cornerstone systems for assessing electrophysiological and structural cardiotoxicity. These models reproduce key human myocardial properties - including action potential dynamics, calcium handling, and contractility - allowing quantitative evaluation of pro-arrhythmic and cardio-depressive effects often missed in animal studies [108,120]. The integration of microelectrode array and optical mapping technologies further enhances predictive power by linking electrophysiological readouts to mitochondrial and molecular stress responses.
In hepatotoxicity research, hPSC-derived hepatocytes and liver organoids now serve as advanced platforms for evaluating xenobiotic metabolism and drug-induced liver injury. Their metabolic competence - including cytochrome P450 activity - together with 3D tissue organisation enables the detection of both acute hepatocellular damage and delayed cholestatic responses [110,111]. The presence of structured bile canaliculi and polarised hepatocytes allows realistic modelling of bile secretion and transporter-mediated toxicity, bridging the long-standing gap between in vitro and in vivo hepatic physiology.
Within the nervous system, PSC-derived brain organoids and neuron-enriched spheroids have become pivotal tools for studying developmental neurotoxicity and neurodegenerative mechanisms. These self-organising 3D cultures reproduce regional brain patterning, cortical layering, and synaptic maturation, making them uniquely suited to analyse neurodevelopmental disruption caused by teratogens or environmental chemicals [121,122]. Exposure studies in these models reveal alterations in neuronal differentiation, synaptogenesis, and glial–neuronal communication - mechanistic endpoints that remain inaccessible to traditional 2D assays.
Renal models derived from hiPSCs have also progressed rapidly. Nephron-like organoids and bioprinted kidney constructs reproduce key aspects of renal filtration and tubular transport, allowing detailed analysis of nephrotoxicity and tubular injury [114,122]. These constructs express segment-specific markers and support compound accumulation and transport studies, offering a more physiologically relevant alternative to immortalised renal cell lines.
Taken together, pluripotent stem cell–derived organoids and organ-on-chip systems mark a decisive step towards integrated, multi-tissue toxicology. By connecting metabolic, electrophysiological, and developmental endpoints, these platforms enhance the predictive accuracy of in vitro assays and bring toxicology closer than ever to truly human-relevant models.
4.4. Ethical and Technical Considerations
Research involving human stem cells continues to raise both ethical and technical challenges. Work with hESCs, derived from early-stage embryos, remains one of the most tightly regulated areas of biomedical science. In countries where hESC research is permitted, stringent oversight mechanisms govern every stage of the process, ensuring donor consent, traceability, and compliance with recognised bioethical standards. Foundational documents such as the Declaration of Helsinki [123] and the EU Directive 2004/23/EC on Human Tissues and Cells [124] outline donor protection principles across the European Union, while the National Institutes of Health (NIH) Stem Cell Registry [125] defines which hESC lines qualify for federally funded research in the United States. On a global level, the International Society for Stem Cell Research (ISSCR) Guidelines for Stem Cell Research and Clinical Translation (2021) provide a comprehensive ethical framework that addresses informed consent, genomic-data protection, and the prohibition of reproductive cloning [126,127].
Although hiPSCs avoid embryo-related ethical concerns, they introduce a different set of issues. Donor privacy, consent for genomic-data use, and the potential misuse of reprogramming technologies for reproductive purposes have all become areas of ethical scrutiny [126,128,129,130,131,132]. The expanding number of patient-specific hiPSC lines underscores the need for robust governance in biobanking, data sharing, and genomic security [128].
From a technical perspective, pluripotent stem cell–based models still face several challenges, particularly regarding reproducibility, inter-line variability, and incomplete maturation of differentiated derivatives. These factors can affect predictive accuracy and hinder regulatory validation. Substantial progress has been made through the development of automated culture systems, lineage-specific fluorescent reporters, and high-content phenotyping pipelines that help reduce variability and improve throughput [133,134]. Nevertheless, rigorous quality control remains essential: residual undifferentiated cells must be excluded, as both hESC- and hiPSC-derived products carry an inherent risk of teratoma formation [105,135].
As ethical frameworks become increasingly harmonised and technical refinements continue to improve reliability, PSC-based models are gaining broader recognition as both scientifically robust and ethically sound platforms for regulatory toxicology.
For clarity, the principal ethical frameworks and procedural requirements for stem-cell research are summarised in Box 4.
- Box 4. Ethical frameworks for stem cell toxicity models
- Declaration of Helsinki (2013) – Universal ethical principles for research involving human-derived material; mandates informed consent and independent ethical review [123];
- EU Directive 2004/23/EC – Standards for donor consent, traceability, and supervision across EU member states [124];
- NIH Stem Cell Registry (United States) – Specifies approved hESC lines for federally funded research in the US [125];
- ISSCR Guidelines for Stem Cell Research and Clinical Translation (2021) – Global reference for hESC/hiPSC research; emphasises informed consent, data protection, and prohibition of reproductive cloning [126];
- National and Institutional Oversight Committees – Ensure compliance with local ethical regulations [126].
- Practical requirements:
- Documented donor consent (in vitro fertilisation (IVF) or somatic cell source);
- Registration of cell lines in recognised repositories;
- Institutional ethics board approval and adherence to ISSCR guidance.
4.5. Adult Stem Cell Models (HSCs and MSCs)
In addition to pluripotent stem cell systems, adult stem cells provide valuable complementary tools for cytotoxicity testing, particularly when tissue-specific or immunological endpoints are of interest. Among these, haematopoietic stem cells (HSCs) and mesenchymal stromal cells (MSCs) derived from bone marrow are the most widely applied [136].
Human MSCs have also been employed to investigate the cytotoxicity and biocompatibility of photobiomodulation procedures. Measurements of viability, calcium signalling, and oxidative balance revealed a pronounced sensitivity of MSCs to irradiation parameters (Pasternak-Mnich et al., 2019). Interestingly, subsequent studies showed that sub-lethal photobiomodulation doses could enhance proliferation and maintain stemness, suggesting that stem cell–based toxicity models are capable of detecting adaptive, even beneficial, stress responses [137].
HSCs, on the other hand, have long served as highly sensitive indicators of bone marrow toxicity. Their intrinsic ability to form distinct haematopoietic colonies in vitro forms the basis of the colony-forming unit (CFU) assay, which quantifies progenitor survival and differentiation following exposure to xenobiotics or chemotherapeutic agents [138,139]. These assays provide direct mechanistic insight into myelotoxic and immunosuppressive effects that often mirror the clinical manifestations of haematopoietic injury [140].
MSCs isolated from bone marrow, adipose tissue, or umbilical cord have also gained prominence in evaluating the cytotoxicity and biocompatibility of biomaterials, nanomaterials, and regenerative scaffolds [141,142]. Their multipotent capacity to differentiate into osteogenic, chondrogenic, and adipogenic lineages enables mechanistic assessment of toxicant-induced alterations in bone, cartilage, and connective tissue physiology [143]. Notably, MSC-based assays have become essential for evaluating the safety of medical implants and nanoparticles, effectively linking toxicology with biomaterials science and regenerative medicine [144].
Compared with pluripotent stem cells, adult stem cell–based models are more accessible and ethically straightforward, making them particularly well suited for targeted, tissue-specific investigations. However, their limited differentiation potential and donor-dependent variability restrict their broader application in mechanistic or high-throughput toxicology [145]. They therefore occupy a complementary niche - providing valuable insight into immunotoxicity, myelotoxicity, and biomaterial compatibility - while hESC- and hiPSC-based systems remain the principal human-relevant platforms for comprehensive cytotoxicity evaluation.
The comparative features of pluripotent and adult stem-cell models, including their sources, differentiation potential, applications, and ethical considerations, are summarised in Table 1.
In concert, pluripotent and adult stem cell platforms unite ethical and scientific strengths, advancing human-relevant, mechanistically informed, and personalised toxicity testing for next-generation safety assessment.
5. Nanotoxicology and Specialized In Vitro Models
Building on the mechanistic insights gained from stem cell–based systems, nanotoxicology applies similar principles to examine how nanoscale materials interact with biological environments. Modern nanotoxicology increasingly prioritises mechanistic and physiologically relevant assessment over traditional, purely descriptive viability testing. Across diverse classes of engineered nanomaterials - including metallic, oxide, and polymeric types - oxidative stress has emerged as a central initiating mechanism driving downstream inflammatory and cytotoxic responses [146,147,148,149].
Recent refinements of classical cytotoxicity assays, coupled with the integration of 3D cultures, microfluidic platforms, and stem cell–based systems, have established a more predictive and human-relevant framework for nanotoxicology. These advances link the physicochemical properties of nanoparticles to molecular and cellular perturbations, bridging nanoscale structure with biological function and enabling more accurate assessment of human health risks [150,151,152,153].
5.1. Cytotoxicity of Nanomaterials: Mechanistic Basis of Oxidative Stress
Engineered nanomaterials interact with biological systems through distinctive physicochemical properties that can disrupt redox balance and trigger oxidative stress. Among the most consistent mechanisms of nanoparticle-induced cytotoxicity is the excessive generation of ROS, which impairs mitochondrial function, damages DNA, and activates pro-inflammatory signalling pathways. Metal oxides such as ZnO, TiO₂, Fe₂O₃, and CeO₂ readily catalyse ROS formation through surface redox reactions and electron transfer, leading to lipid peroxidation, mitochondrial dysfunction, and apoptotic cell death [146,147,148,154]. The magnitude of oxidative injury depends strongly on particle size, surface charge, aggregation behaviour, and the composition of the surrounding protein corona [155,156,157].
Similar redox-driven mechanisms have been observed with polymeric nanostructures such as dendrimers, where a high surface charge density can induce mitochondrial depolarisation, caspase activation, and oxidative DNA damage [149,158]. Surface functionalisation with neutral or carbohydrate groups markedly reduces ROS generation and helps restore cellular redox homeostasis [155,159]. Collectively, these findings support a unifying model in which nanoparticle toxicity arises primarily from physicochemical interactions that overwhelm endogenous antioxidant defences, rather than from the intrinsic chemical composition of the material.
Beyond direct oxidative damage, nanomaterial exposure can also provoke a spectrum of secondary stress responses - including inflammasome activation, mitophagy, and stress granule formation - largely driven by ROS-dependent signalling [160,161]. Together, these interconnected processes establish oxidative stress as a central initiating event that links nanoparticle physicochemistry to downstream pathways of apoptosis, inflammation, and adaptive stress responses.
5.2. Adaptation of Classical Cytotoxicity Assays to Nanomaterials
Traditional viability assays - such as MTT, LDH release, NRU, and resazurin reduction - were originally developed for testing soluble compounds and often yield unreliable results when applied to nanomaterials. Nanoparticles can adsorb assay dyes, scatter incident light, or catalyse redox reactions, resulting in false-positive or false-negative outcomes [70,162]. Comparative studies have consistently shown that no single assay endpoint can reliably capture nanoparticle-induced toxicity, and that optical interference remains a major source of experimental variability [71,162].
To address these limitations, current best practices emphasise careful control of experimental design, including the use of nanoparticle-only controls, orthogonal readouts, and thorough characterisation of dispersion state, serum content, and incubation time [163]. Studies using cationic polymer nanocarriers have demonstrated that many apparent cytotoxic effects stem from interactions with assay reagents rather than genuine cellular injury. In such cases, alternative assays - focusing on membrane integrity or haemolytic activity - can offer more reliable indicators of nanoparticle-induced damage [164,165].
Overall, these refinements highlight that classical viability assays alone are insufficient for accurate nanotoxicity assessment. Incorporating mechanistic endpoints - such as ROS quantification, mitochondrial membrane potential, or caspase activation - provides a more robust and interpretable evaluation of nanoparticle-induced cellular effects.
5.3. Specialized In Vitro Models and Specific Endpoints
Advances in in vitro methodology have transformed nanotoxicology from a largely descriptive discipline into one grounded in mechanistic understanding and physiological relevance. 3D cultures, organoids, and microfluidic “organ-on-chip” platforms now replicate native tissue architecture and biochemical gradients, substantially improving the predictive power of in vitro testing. For example, liver spheroids and organoid models preserve metabolic competence and reveal delayed hepatotoxic effects that remain undetectable in conventional monolayer cultures [91,152]. The pioneering lung-on-a-chip system introduced dynamic cyclic strain and epithelial–endothelial co-cultures to mimic nanoparticle deposition at the air–liquid interface [153], while subsequent intestine- and skin-on-chip designs have enabled real-time monitoring of barrier integrity and inflammatory mediator release [151,166].
Stem cell–derived and bioprinted organoids have further expanded nanosafety assessment into developmental and regenerative contexts, supporting long-term studies of sublethal toxicity and adaptive stress responses [150,167]. Mechanistic endpoints - such as ROS production, mitochondrial membrane potential, NF-κB activation, and cytokine release - are now routinely measured alongside classical viability assays, yielding a multidimensional picture of nanoparticle–cell interactions [159,160,168].
Together, these technological advances integrate detailed physicochemical characterisation with systems-level biology, establishing a comprehensive, mechanistically anchored, and human-relevant framework for nanosafety evaluation.
Key mechanistic principles and methodological standards of modern nanotoxicology are summarized in Box 5.
- Box 5. Practical and mechanistic insights into nanotoxicology.
- Assay adaptation: Nanoparticles interfere with colourimetric and fluorometric assays by adsorbing dyes or catalysing redox reactions. Reliable assessment therefore requires nanoparticle-only controls and confirmation using orthogonal endpoints such as ATP quantification or impedance-based measurements [70,71,162,165].
6. Advanced 3D Models: Organoids, Organ-on-Chip, and Bioprinting
Recent advances in bioengineering and microphysiological systems (MPSs) have profoundly reshaped in vitro toxicology, providing human-relevant models that reproduce organ-level functions and systemic pharmacokinetics. 3D organoid cultures, microfluidic organ-on-chip devices, and bioprinted tissues now capture essential features of native tissue organisation - such as perfusion, polarisation, and intercellular communication - that were long unattainable in conventional monolayers. By generating quantitative, mechanism-based data that link cellular perturbations to tissue- and organism-level outcomes, these systems effectively bridge the gap between molecular assays and clinical toxicology.
The incorporation of dynamic flow, multi-organ coupling, and computational integration within these platforms represents a decisive step towards predictive toxicology that aligns with emerging regulatory paradigms, including NAMs and QIVIVE [11,170,171].
6.1. Organoids: Tissue-Specific and Immune-Competent Models
Human organoids are self-organising, multicellular constructs that recapitulate key aspects of tissue morphology and function. Among the most advanced examples, hepatic organoids derived from pluripotent stem cells have become indispensable tools for the mechanistic evaluation of drug-induced liver injury (DILI) and metabolic safety. These high-fidelity systems reproduce clinical patterns of hepatotoxicity and enable quantitative risk assessment [81,110]. Integrating liver organoids into microfluidic chips further enhances throughput and precision by introducing physiological flow and nutrient exchange [172]. Moreover, multicellular liver constructs designed to model non-alcoholic fatty liver disease (NAFLD) now capture chronic toxicity phenotypes associated with metabolic disorders [173].
Kidney organoids and tubuloids have achieved similar progress, reproducing nephron-like structures, transporter expression profiles, and tubular polarity that enable the study of drug-induced nephrotoxicity and renal clearance. Functional proximal-tubule systems allow the investigation of infection, filtration, and injury under near-physiological conditions [174], while quantitative optical imaging enables real-time scoring of renal injury [175]. Comparative analyses highlight their potential as human-relevant alternatives to traditional animal kidney assays [176].
Intestinal organoids provide a complementary model that links absorption, barrier integrity, and microbiota interactions - critical factors in oral pharmacokinetics and first-pass metabolism. Bioengineered intestinal constructs now reproduce epithelial–mesenchymal–neuronal complexity [177], and human enteroid monolayers have been validated as robust and reproducible models of epithelial barrier function and transport [178]. Their application in in vitro–in vivo extrapolation of oral drug disposition has recently been reviewed in detail [179].
Other epithelial organoids extend this approach to barrier tissues such as the cornea, retina, and skin. Retina-on-chip platforms combine retinal organoids with microfluidic circuits to reconstruct neurovascular coupling and photoreceptor–glia interactions [115], while skin organoids have become standardised models for studying irritation, sensitisation, and microbial infection [180,181].
Incorporating immune components within organoid systems adds another layer of complexity. Co-cultures that integrate lymphoid or myeloid cells enable the study of tumour–immune interactions and immunomod
6.2. Microfluidics: Organ-on-Chip and Body-on-Chip Systems
MPSs embed human cells within perfused microenvironments that maintain long-term tissue viability, intercellular communication, and physiologically relevant pharmacokinetic gradients. Liver- and kidney-on-chip platforms reproduce key metabolic and excretory functions, generating quantitative endpoints for evaluating DILI and nephrotoxicity [172,186].
Coupled organ circuits - such as gut–liver or liver–kidney configurations - extend this approach by enabling the investigation of metabolite-driven cross-organ effects and systemic clearance [170,187]. Intestinal chips containing self-organising epithelial, mesenchymal, and neuronal components reproduce luminal flow and enteric regulation, providing physiologically coherent models for ADME studies [188].
At the frontier of bioengineering, multi-organ “body-on-chip” and digital-twin systems integrate multiple organ modules within a single circuit to emulate whole-body pharmacokinetics and complex physiological phenomena, including maternal–foetal exchange [189]. Collectively, these innovations represent a decisive shift from static cell culture toward dynamic, systems-level modelling of human biology, offering a powerful bridge between in vitro experimentation and clinical pharmacology.
6.3. Three-Dimensional Bioprinting: Standardisation and Reproducibility
3D bioprinting enables the precise, layer-by-layer fabrication of living tissues using bioinks composed of cells and extracellular matrix components. This technology enhances reproducibility, scalability, and architectural fidelity - features that are essential for the regulatory acceptance of organoid-based assays. Extrusion bioprinting has markedly improved the morphological uniformity of kidney constructs [171], while biofabricated hepatic models have demonstrated consistent performance in inter-laboratory toxicity screening studies [190].
Recent technological advances have further expanded the scope of this field. Miniaturised spinning bioreactors now accelerate epithelial organoid production [191], and bioprinted interstitial fibrosis models enable controlled investigation of chronic injury and drug-induced fibrogenesis under defined mechanical conditions [192].
Together, these developments form the foundation for standardised pipelines in 3D tissue fabrication, aligning with Good Cell and Tissue Culture Practice and supporting data harmonisation initiatives that will facilitate broader regulatory adoption.
6.4. Translational ADME–Tox Prediction and In Vivo Extrapolation
The convergence of organoid, microfluidic, and bioprinting technologies has ushered in a new era of predictive, mechanism-anchored approaches to human pharmacokinetics and toxicology. When combined with computational modelling and toxicogenomic profiling, these advanced in vitro systems enable robust extrapolation from cellular responses to clinical outcomes.
Recent genomic research has identified polygenic determinants of susceptibility to DILI [193] and mapped molecular response networks through large-scale toxicogenomic studies [194], providing a mechanistic foundation for risk assessment. Liver and intestinal organoid platforms are now routinely employed for ADME profiling and QIVIVE [81,179].
Microphysiological liver systems have demonstrated strong concordance between pharmacokinetic behaviour and dynamic toxicity endpoints [172,195], while multi-organ robotic chips now support automated QIVIVE workflows and integrated data analytics [196]. Collectively, these advances mark a transition from descriptive cytotoxicity testing to predictive, human-centred toxicology grounded in quantitative mechanistic evidence.
Key recommendations for experimental design, model integration, and regulatory alignment are summarized in Box 6.
- Box 6. Practical Guidance for Model Design and Integration
- Combine static and dynamic systems: Use organoids as foundational tissue modules and integrate them into microfluidic circuits to capture physiological flow, nutrient gradients, and metabolite exchange.
- Standardise culture conditions: Define media composition, extracellular matrix parameters, and bioprinting settings to minimise batch variation and improve reproducibility across laboratories.
- Benchmark with reference compounds: Validate functional readouts (e.g., albumin, urea, γ-GT, transporter activity) using well-characterised hepatotoxins or nephrotoxins before introducing novel agents.
- Implement multi-organ connectivity: Couple intestinal, hepatic, and renal modules to assess systemic ADME and metabolite-driven toxicity, supporting quantitative IVIVE modelling.
- Integrate computational tools: Apply PBPK and QIVIVE frameworks to translate microphysiological outputs into clinically relevant exposure predictions.
- Ensure regulatory alignment: Follow OECD and FDA recommendations on Good Cell and Tissue Culture Practice and NAMs to support data acceptance and cross-sector harmonisation.
7. In Silico Approaches and Computational Toxicology
Computational modelling has become an essential part of modern toxicology. By complementing in vitro systems, it allows prediction of biological effects across large chemical spaces, clarifies underlying mechanisms, and supports quantitative risk assessment [197,198]. The field is increasingly shaped by high-quality, shareable datasets and transparent modelling workflows [199,200] that connect molecular perturbations with physiological responses through pharmacokinetic modelling and quantitative extrapolation frameworks [89,201,202].
Contemporary in silico toxicology is structured around four methodological pillars:
All these approaches now operate within harmonised FAIR-data and model-validation frameworks. However, they share common challenges - particularly uncertainties in metabolic clearance and tissue distribution - that can distort extrapolations if not explicitly tested [202].
An overview of the four computational pillars is provided in Table 2, while a concise, practical workflow from data to regulatory decision-making is summarised in Box 7.
- Box 7. Practical workflow for computational toxicology (from data to decision).
- Define the question and endpoint. Select a suitable modelling family (QSAR or ML) and the kinetic coupling (PBPK or QIVIVE) appropriate to the context.
- FAIR data curation. Standardise identifiers, harmonise units, remove duplicates and outliers, and record provenance and data partitions [199].
7.1. Quantitative Structure–Activity Relationships (QSAR), Read-Across, and Cheminformatics
QSAR models mathematically link chemical structure with biological activity or toxicity using statistical and machine-learning methods. Structural information is translated into molecular descriptors to infer activity patterns, allowing prediction for untested compounds, prioritisation of further testing, and reduction of animal use under the NAM and IATA frameworks [199,200].
When built from high-quality datasets and applied within a clearly defined domain of applicability, QSAR models can perform reliably for systemic and safety-critical endpoints such as human carcinogenicity or cardiotoxicity [203,206].
Modern guidance stresses adherence to the FAIR principles - findability, accessibility, interoperability, and reusability - to ensure transparency and auditability. Key practical aspects include:
- careful descriptor selection and redundancy control,
- transparent separation of training and validation sets,
- Y-randomisation to exclude chance correlations, and
Applications range from chronic oral carcinogenicity prediction to cardiac-safety screening, where QSAR approaches efficiently flag potential liabilities before costly laboratory testing [203,206]. Comparative analyses consistently show that QSAR delivers the most value when combined with in vitro and preclinical evidence rather than used in isolation [207].
7.2. Machine Learning and AI for Cytotoxicity Prediction
Machine learning (ML) extends beyond QSAR by capturing non-linear structure–activity relationships and handling complex, multi-endpoint datasets that include omics and imaging features. Publicly accessible platforms such as ProTox 3.0 now provide continuously updated toxicity models with user-friendly interfaces suitable for both academia and industry [204].
Interpretability remains central to regulatory acceptance. Mechanism-aware and model-agnostic techniques - such as feature-importance mapping and attention-based visualisation - allow predictions to be linked to specific chemical substructures or biological pathways, thereby increasing confidence in model outputs [198].
Guidelines from recent studies clarify when to favour classical learners (e.g., random forests) versus deep neural networks, and emphasise the importance of rigorous external validation to prevent overfitting and ensure reproducibility across datasets [197].
End-to-end ML pipelines now automate data curation, descriptor generation, training, and deployment, reducing manual bias and improving reproducibility [208]. Explainable ML models for dermal toxicity achieve accuracy comparable to traditional baselines while providing clear insight into mechanistic drivers [209]. In cardiac safety assessment, curated datasets of hERG channel inhibition highlight that model reliability depends heavily on data quality, curation of negatives, and threshold optimisation [210].
Overall, these advances show that ML delivers greatest value when trained on well-annotated, purpose-specific datasets, implemented through reproducible pipelines, and accompanied by interpretable outputs. This union of transparency and mechanistic insight has moved ML from an exploratory tool to a credible, fit-for-purpose element of regulatory toxicology [208,209,210].
7.3. Physiologically Based Pharmacokinetic (PBPK) Modelling
PBPK models quantitatively describe how chemicals are absorbed, distributed, metabolised, and excreted by representing human physiology - blood flow, tissue partitioning, and metabolism - in a mechanistic framework. They link in vitro activity thresholds to predicted tissue concentrations in human populations, accounting for variability due to age, comorbidities, and drug–drug interactions [201].
From both industrial and regulatory perspectives, three key pillars define robust PBPK practice:
- Population relevance: evaluation of specific subgroups such as paediatrics, pregnancy, or hepatic/renal impairment.
- Uncertainty management: systematic sensitivity analysis of physiological and chemical parameters to assess influence on predictions.
- Model qualification: benchmarking against reliable clinical reference data [201].
Within model-informed drug development (MIDD), PBPK supports first-in-human dosing, drug–drug interaction prediction, and extrapolation to sensitive populations where direct data are limited [89]. Recent extensions include models accounting for obesity-related changes in organ perfusion and clearance [211] and AI-assisted PBPK frameworks for nanoparticle pharmacokinetics [212]. Specialised modules have been introduced for nanoparticle dynamics - addressing protein corona formation and mononuclear-phagocyte uptake [213].
Importantly, reproducibility requires external validation. Multi-centre comparisons, such as PBPK qualification in pregnancy, demonstrate that harmonised workflows can yield reliable maternal–foetal exposure predictions [202].
7.4. Quantitative In Vitro–In Vivo Extrapolation (QIVIVE)
QIVIVE integrates in vitro concentration–response data with PBPK or physiologically based toxicokinetic (PBTK) models to estimate human-relevant exposure levels [87,88].
The process typically involves four key elements:
- (i)
- correction of in vitro concentrations for plastic and protein binding;
- (ii)
- determination of binding fractions in blood and tissues;
- (iii)
- measurement of metabolic and excretory clearance; and
- (iv)
- definition of the relevant exposure metric - Cmax, AUC, or steady state - with quantified uncertainty.
High-throughput PBTK workflows now enable QIVIVE for thousands of compounds, integrating internal-dose predictions with bioactivity profiles for screening-level risk ranking [88]. Mechanistic key events can be embedded directly within the PBPK–QIVIVE chain - for instance, using epigenetic markers to refine risk estimates for PFAS [180], or achieving cross-species concordance for acetylcholinesterase inhibition when kinetic parameters are well characterised [214].
Proper diagnostics and uncertainty analyses are essential: inaccurate assumptions regarding clearance or partitioning can distort outcomes, so sensitivity and probabilistic error propagation should always be conducted [202]. Beyond single viability endpoints, phenotypic profiling - for example, the Cell Painting assay - offers mechanistic fingerprints to anchor the in vitro point of departure and strengthen translational validity [61].
In practice, reproducible QIVIVE requires simulation of tissue concentration–time curves within a PBPK framework, selection of the appropriate exposure metric, and execution of global or Monte Carlo uncertainty analyses, all implemented in transparent, script-based pipelines for traceability [87,88]. Properly applied, QIVIVE transforms in vitro findings into quantitative human risk estimates, positioning in silico toxicology as an integrative partner - rather than a replacement - for experimental systems.
Key references, practical guidance, and step-by-step implementation details are provided in Box 8 and Box 9.
- Box 8. Key resources for QIVIVE and PBPK modeling
- Reviews and Methods
- Practical roadmaps for QIVIVE and integration into IATA [87]
- High-throughput PBTK for IVIVE at scale [88]
- PBPK for decision-making and uncertainty analysis [201]
- Model-informed development for special populations [89]
- Linking phenotypic profiling with QIVIVE (Cell Painting) [61]
- Case Studies
- PFAS: epigenetic key event integration within PBPK [215]
- AChE inhibition: kinetic cross-species concordance [214]
- How-To Sources
- Box 9. Practical workflow for QIVIVE and PBPK implementation.
- (i)
- (ii)
- (iii)
- (iv)
8. Integrated Approaches and Regulatory Perspectives
The global transition from descriptive, animal-based testing to predictive and mechanistically anchored toxicology has been propelled by the emergence of IATA and NAMs. These frameworks weave together in vitro, in silico, and in chemico data with existing knowledge to support regulatory decisions aligned with the 3Rs principle [33].
Rather than denoting single assays, IATA represent structured, evidence-based strategies that combine data from validated experimental systems, computational models, and expert judgement to establish hazard or potency. Their architecture is deliberately flexible and transparent, designed to be fit-for-purpose and adaptable across different chemical domains and regulatory settings [218].
8.1. From Concept to Practice: Building Confidence in NAMs
Scientific and regulatory confidence in NAMs has grown steadily over the past decade under the coordinated guidance of organisations such as the OECD, EURL ECVAM (EU Reference Laboratory for Alternatives to Animal Testing), and U.S. agencies including the Environmental Protection Agency (EPA), FDA, and NIH. Consensus has emerged that the credibility of NAMs does not rely on a one-to-one substitution of animal studies but rather on the demonstration of reproducibility, mechanistic coherence, and well-defined domains of applicability [242; van der Zalm et al., 2022].
NAMs now span a broad technological spectrum - from high-content imaging and transcriptomic profiling to organ-on-chip microphysiological systems and computational modelling. Their regulatory acceptance is evaluated through context-of-use validation, whereby a method’s reliability is assessed within the specific decision framework for which the data are intended [86,219].
Selected examples of this principle are summarised in Box 10, highlighting validated NAMs accepted within defined regulatory contexts.
-
Box 10. Context-of-use validation: how NAMs gain regulatory credibilityRegulatory confidence in NAMs is achieved through context-of-use validation, which establishes a method’s reliability for a defined regulatory purpose rather than as a universal replacement for animal testing.
- Examples:
- Skin sensitization – The DA (OECD TG 497) is validated for identifying sensitising chemicals but not for potency ranking or quantitative risk assessment [220].
- Skin irritation – Reconstructed human epidermis models (OECD TG 439) are accepted for classification and labelling but not for chronic or systemic toxicity testing [221].
-
Key principle:Confidence in a NAM depends on demonstrated reliability within its regulatory context - each method is accepted only for what it has been proven to do.
Collaborative initiatives such as APCRA (Accelerating the Pace of Chemical Risk Assessment) and PARC (Partnership for the Assessment of Risks from Chemicals) have further strengthened harmonisation through shared case studies and alignment of interpretive criteria [223]. More recently, expert groups have called for unified validation principles and standardised reporting to support cross-jurisdictional acceptance of NAM-derived data [224]. Ultimately, successful implementation depends not only on technological maturity but also on the preparedness of the regulatory ecosystem - its training, resources, and institutional culture - which determine how effectively innovation is embedded in practice [225].
8.2. Case Studies and Regulatory Uptake
Validated IATA and DAs have progressed from theoretical constructs to operational tools within several OECD Test Guidelines, providing practical alternatives to animal testing.
In the field of skin corrosion and irritation, reconstructed human epidermis models such as EpiDerm™, SkinEthic™, and epiCS - when combined with in chemico assays like the Direct Peptide Reactivity Assay (DPRA) - are formally recognised under OECD TG 431 and TG 439, marking a mature application of IATA principles in regulatory toxicology [221,226,227]. Similarly, eye irritation testing has been revolutionised by in vitro corneal models such as EpiOcular™ and SkinEthic™ HCE, which underpin OECD TG 492 and effectively replace the classical Draize rabbit eye test [35,228,229,230]. Real-time impedance-based monitoring now enhances these models, allowing quantitative distinction between reversible and irreversible injury.
Advances are also evident in developmental and reproductive toxicity - traditionally a major barrier to non-animal assessment. Modern IATA integrate human pluripotent stem-cell differentiation assays with multi-omics and PBPK/QIVIVE modelling to connect early mechanistic perturbations with adverse developmental outcomes [231,232].
New-generation assay platforms such as ReproTracker, PluriLum, and the UKN4DNT framework now make it possible to assess embryotoxicity in a structured and mechanistically informed way. By analysing transcriptomic and proteomic patterns, these systems reveal how toxicants disturb key developmental processes - including neurogenesis, cardiogenesis, and morphogenesis - within human stem-cell–based models (see Box 11).
- Box 11. Emerging Platforms for Developmental and Reproductive Toxicity Testing
- ReproTracker – Tracks differentiation of human pluripotent stem cells into germ layers to detect embryotoxic and teratogenic effects through gene-expression markers [231].
- PluriLum Test – Combines stem-cell differentiation with high-content imaging and transcriptomics, generating mechanistic fingerprints of disrupted morphogenesis [232].
When linked with pharmacokinetic modelling, these data support quantitative points of departure and safety margins without recourse to animal studies [87,88,233,234]. A notable example of next-generation risk assessment (NGRA) is the daidzein read-across case, in which exposure modelling, in vitro assays, and computational predictions were integrated into a tiered workflow yielding regulatory-quality safety conclusions [235]. Collectively, these examples illustrate how IATA and NAMs are transforming mechanistic concepts into practical regulatory instruments.
8.3 Global Regulatory Perspectives
The OECD has formalised principles underpinning IATA and NAMs in its Guidance Document No. 255 on the reporting of DAs (2017) and Guidance Document No. 311 on Weight-of-Evidence evaluation (2019), which together establish standards for data integration, uncertainty analysis, and transparent reporting [236,237]. These documents support the DA concept exemplified in OECD TG 497 on skin sensitisation [220].
Parallel developments at the FDA and EMA demonstrate similar intent. The FDA’s Roadmap to Reducing Animal Testing (2025) encourages inclusion of NAM-generated data in preclinical submissions [233], while the EMA’s Horizon Scanning Report on NAMs (2025) highlights their centrality to future regulatory science [234]. Within the EU, EURL ECVAM continues to track progress through its annual Status Reports [238,239,240], ensuring transparency in validation and acceptance. Notably, recent OECD updates introduced an IATA for Phototoxicity [241] and revised Test Guideline 442D to include EpiSensA [242], extending non-animal strategies for skin sensitisation.
Although regional differences persist in validation criteria and reviewer expertise, these collective actions demonstrate a strong and coordinated global momentum toward harmonised, mechanistically grounded regulation.
Representative examples of IATA/NAM implementation across regulatory endpoints are shown in Table 3.
8.4. Outlook and Emerging Trends
Looking ahead, the convergence of AI, multi-omics, and MPS networks is poised to enhance the predictive and translational scope of NAMs. Rapid progress in QIVIVE is enabling quantitative linkage between in vitro dose–response relationships and human exposure scenarios [87,214]. In parallel, computational innovations - such as machine-learning-optimised PBPK models and genetic-algorithm parameter estimation - are refining model representativeness and reproducibility [212,243].
On the policy side, the CHANGE Initiative (2024–2026) (Collaboration to Harmonize the Assessment of Next Generation Evidence) seeks to accelerate NAM adoption through coordinated action across governance, funding, and education [225]. Harmonised IATA and DAs are now increasingly acknowledged as formal decision-making frameworks across regulatory systems [86]. Beyond human health, new IATA/NAM applications are emerging in ecotoxicology and mixture assessment, extending mechanistic and ethical principles to environmental safety [244,245].
Together, these developments demonstrate that IATA and NAMs now form the operational backbone of 21st-century toxicology - uniting computational modelling, AI-assisted data interpretation, and human-relevant biology into a predictive, quantitative, and ethically sustainable science..
9. Conclusions and Future Perspectives
Cytotoxicity testing has progressed from simple viability measurements to integrated, mechanistic, and human-relevant models that connect cellular biology with systems toxicology and computational prediction. The classical colourimetric assays - MTT, LDH, and Neutral Red - laid the groundwork for in vitro toxicology, offering the first reliable means of assessing cell injury and establishing reference points still recognised in regulatory practice [4,6,7,8]. Yet, their simplicity could not capture the complexity of toxic mechanisms or predict systemic responses [9].
Over the past two decades, these limitations have inspired a complete rethinking of how cellular toxicity is studied. High-throughput and high-content screening have enabled large-scale mapping of cellular perturbations [15], while stem-cell–based systems have introduced genuine human developmental and organ-level relevance [16,17]. The rise of organoids, organ-on-chip devices, and 3D bioprinted tissues now allows researchers to recreate physiological microenvironments and tissue–tissue interactions, offering unprecedented insight into pharmacokinetics, metabolism, and adaptive stress responses [21,23,27]. Together, these advances have transformed in vitro toxicology from a descriptive exercise into a mechanistically grounded science that actively supports risk assessment, drug discovery, and regulatory evaluation while stem-cell–based systems introduced human developmental and organ-specific relevance [2,11].
The influence of this shift reaches far beyond experimental innovation. In biomedicine, advanced cytotoxicity platforms enable earlier and more accurate safety screening, reducing costly late-stage failures in drug development [1,26]. In regulatory science, they provide the scientific basis for implementing NAMs and IATA, reducing dependence on animal models while improving human relevance [32,33]. In public health, these methods expand our ability to assess chemical mixtures, nanomaterials, and complex exposure scenarios with greater ethical confidence and translational accuracy [18,19,20]. New kinetic and multiparametric approaches - such as real-time impedance assays (xCELLigence) and flow cytometry - further refine our understanding by distinguishing short-term adaptive stress from irreversible cell death [44,70].
Looking to the future, progress in cytotoxicity testing will depend on the integration of artificial intelligence, multi-organ systems, and personalised toxicology. AI-driven models, combined with omics-based signatures and QIVIVE frameworks, are paving the way for predictive simulations of toxicity across scales - from molecular interactions to organism-level outcomes [87,197,198]. Body-on-chip systems will provide dynamic, human-physiology analogues for studying systemic toxicity, metabolism, and inter-organ communication [26,195,196]. Meanwhile, patient-specific hiPSC-derived models will bring toxicology closer to precision medicine, enabling individualised assessment of chemical susceptibility [96,108,113].
In essence, cytotoxicity testing is evolving from a static endpoint measure into a predictive and integrative discipline - one that underpins safer therapeutics, more sustainable chemical design, and evidence-based regulation in the 21st century[2,3,11]. A concise overview of the historical milestones and emerging directions in cytotoxicity testing is provided in Table 4.
Author Contributions
The author confirms being the sole contributor to this work and has approved it for publication.
Funding
There is no specific funding to be disclosed.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
During the preparation of this manuscript, the authors used ChatGPT (OpenAI GPT-5, 2025 version) for the purposes of analyzing the drafted text, generating tables and Box summaries, and assisting with language refinement. The authors have reviewed and edited all AI-generated content and take full responsibility for the final version of the manuscript.
Conflicts of Interest
The author declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| 2D / 3D | Two-/Three-Dimensional Cell Culture |
| 3Rs | Replacement, Reduction, and Refinement |
| ADME | Absorption, Distribution, Metabolism, and Excretion |
| AI | Artificial Intelligence |
| CFU | Colony-Forming Unit |
| DA | Defined Approach |
| DDI | Drug–drug interaction |
| D(E)T | Developmental (Embryo) Toxicity |
| DILI | Drug-Induced Liver Injury |
| DNT | Developmental Neurotoxicity |
| EMA | European Medicines Agency |
| ER | Estrogen Receptor |
| FDA | Food and Drug Administration |
| HCI | High-Content Imaging |
| HCS | High-Content Screening |
| hESC | Human Embryonic Stem Cell |
| hiPSC / iPSC | (Human) Induced Pluripotent Stem Cell |
| hiPSC-CM | hiPSC-Derived Cardiomyocyte |
| HSC | Hematopoietic Stem Cell |
| HTS / qHTS | (Quantitative) High-Throughput Screening |
| IATA | Integrated Approaches to Testing and Assessment |
| ISSCR | International Society for Stem Cell Research |
| LDH | Lactate Dehydrogenase |
| MCS | Mesenchymal Stromal Cell |
| MEA | Microelectrode Array |
| ML | Machine Learning |
| MPS | Microphysiological System |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| NAMs | New Approach Methodologies |
| NIH | National Institutes of Health |
| NRU | Neutral Red Uptake |
| OECD | Organisation for Economic Co-operation and Development |
| PBPK | Physiologically Based Pharmacokinetic (Modeling) |
| PFAS | polyfluoroalkyl substances |
| PI | Propidium Iodide |
| PSC | Pluripotent Stem Cell |
| QIVIVE | Quantitative In Vitro–In Vivo Extrapolation |
| QSAR | Quantitative Structure–Activity Relationship |
| ROS | Reactive Oxygen Species |
| RTCA | Real-Time Cell Analysis |
| SRB | Sulforhodamine B |
| TCPL | ToxCast Pipeline for Curve Fitting |
| Tox21 / ToxCast | U.S. Toxicology Data Programs for High-Throughput Screening |
| TT21C | Toxicity Testing in the 21st Century |
| WoE | Weight-of-Evidence |
References
- Taylor, K. Ten Years of REACH - An Animal Protection Perspective. Altern Lab Anim 2018, 46, 347–373. [CrossRef]
- Jennings, P. “The Future of in Vitro Toxicology.” Toxicology in Vitro 2015, 29, 1217–1221. [CrossRef]
- Bhattacharya, S.; Zhang, Q.; Carmichael, P.L.; Boekelheide, K.; Andersen, M.E. Toxicity Testing in the 21 Century: Defining New Risk Assessment Approaches Based on Perturbation of Intracellular Toxicity Pathways. PLoS One 2011, 6. [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J Immunol Methods 1983, 65, 55–63. [CrossRef]
- Gerlier, D.; Thomasset, N. Use of MTT Colorimetric Assay to Measure Cell Activation. J Immunol Methods 1986, 94, 57–63. [CrossRef]
- Decker, T.; Lohmann-Matthes, M.L. A Quick and Simple Method for the Quantitation of Lactate Dehydrogenase Release in Measurements of Cellular Cytotoxicity and Tumor Necrosis Factor (TNF) Activity. J Immunol Methods 1988, 115, 61–69. [CrossRef]
- Borenfreund, E.; Puerner, J.A. Toxicity Determined in Vitro by Morphological Alterations and Neutral Red Absorption. Toxicol Lett 1985, 24, 119–124. [CrossRef]
- Fotakis, G.; Timbrell, J.A. In Vitro Cytotoxicity Assays: Comparison of LDH, Neutral Red, MTT and Protein Assay in Hepatoma Cell Lines Following Exposure to Cadmium Chloride. Toxicol Lett 2006, 160, 171–177. [CrossRef]
- Eisenbrand, G.; Pool-Zobel, B.; Baker, V.; Balls, M.; Blaauboer, B.J.; Boobis, A.; Carere, A.; Kevekordes, S.; Lhuguenot, J.C.; Pieters, R.; et al. Methods of in Vitro Toxicology. Food and Chemical Toxicology 2002, 40, 193–236. [CrossRef]
- Sipes, N.S.; Padilla, S.; Knudsen, T.B. Zebrafish: As an Integrative Model for Twenty-First Century Toxicity Testing. Birth Defects Res C Embryo Today 2011, 93, 256–267. [CrossRef]
- Hartung, T. Toxicology for the Twenty-First Century. Nature 2009, 460, 208–212. [CrossRef]
- Dearfield, K.L.; Thybaud, V.; Cimino, M.C.; Custer, L.; Czich, A.; Harvey, J.S.; Hester, S.; Kim, J.H.; Kirkland, D.; Levy, D.D.; et al. Follow-up Actions from Positive Results of in Vitro Genetic Toxicity Testing. Environ Mol Mutagen 2011, 52, 177–204. [CrossRef]
- Toxicology Testing in the 21st Century (Tox21) | US EPA Available online: https://www.epa.gov/chemical-research/toxicology-testing-21st-century-tox21 (accessed on 14 October 2025).
- Toxicity Forecasting (ToxCast) | US EPA Available online: https://www.epa.gov/comptox-tools/toxicity-forecasting-toxcast (accessed on 14 October 2025).
- Shukla, S.J.; Huang, R.; Austin, C.P.; Xia, M. The Future of Toxicity Testing: A Focus on in Vitro Methods Using a Quantitative High-Throughput Screening Platform. Drug Discov Today 2010, 15, 997–1007. [CrossRef]
- Scott, C.W.; Peters, M.F.; Dragan, Y.P. Human Induced Pluripotent Stem Cells and Their Use in Drug Discovery for Toxicity Testing. Toxicol Lett 2013, 219, 49–58. [CrossRef]
- Shinde, V.; Sureshkumar, P.; Sotiriadou, I.; Hescheler, J.; Sachinidis, A. Human Embryonic and Induced Pluripotent Stem Cell Based Toxicity Testing Models: Future Applications in New Drug Discovery. Curr Med Chem 2016, 23, 3495–3509. [CrossRef]
- Schrand, A.M.; Dai, L.; Schlager, J.J.; Hussain, S.M. Toxicity Testing of Nanomaterials. Adv Exp Med Biol 2012, 745, 58–75. [CrossRef]
- Arora, S.; Rajwade, J.M.; Paknikar, K.M. Nanotoxicology and in Vitro Studies: The Need of the Hour. Toxicol Appl Pharmacol 2012, 258, 151–165. [CrossRef]
- Hillegass, J.M.; Shukla, A.; Lathrop, S.A.; MacPherson, M.B.; Fukagawa, N.K.; Mossman, B.T. Assessing Nanotoxicity in Cells in Vitro. Wiley Interdiscip Rev Nanomed Nanobiotechnol 2010, 2, 219–231. [CrossRef]
- Pampaloni, F.; Stelzer, E.H.K. Three-Dimensional Cell Cultures in Toxicology. Biotechnol Genet Eng Rev 2010, 26, 117–138. [CrossRef]
- Lu, H.; Zuo, X.; Yuan, J.; Xie, Z.; Yin, L.; Pu, Y.; Chen, Z.; Zhang, J. Research Progress in the Development of 3D Skin Models and Their Application to in Vitro Skin Irritation Testing. J Appl Toxicol 2024, 44, 1302–1316. [CrossRef]
- Augustyniak, J.; Bertero, A.; Coccini, T.; Baderna, D.; Buzanska, L.; Caloni, F. Organoids Are Promising Tools for Species-Specific in Vitro Toxicological Studies. J Appl Toxicol 2019, 39, 1610–1622. [CrossRef]
- Fair, K.L.; Colquhoun, J.; Hannan, N.R.F. Intestinal Organoids for Modelling Intestinal Development and Disease. Philos Trans R Soc Lond B Biol Sci 2018, 373. [CrossRef]
- Van Midwoud, P.M.; Verpoorte, E.; Groothuis, G.M.M. Microfluidic Devices for in Vitro Studies on Liver Drug Metabolism and Toxicity. Integr Biol (Camb) 2011, 3, 509–521. [CrossRef]
- Mahler, G.J.; Esch, M.B.; Stokol, T.; Hickman, J.J.; Shuler, M.L. Body-on-a-Chip Systems for Animal-Free Toxicity Testing. Altern Lab Anim 2016, 44, 469–478. [CrossRef]
- Ishida, S. Organs-on-a-Chip: Current Applications and Consideration Points for in Vitro ADME-Tox Studies. Drug Metab Pharmacokinet 2018, 33, 49–54. [CrossRef]
- Groothuis, F.A.; Heringa, M.B.; Nicol, B.; Hermens, J.L.M.; Blaauboer, B.J.; Kramer, N.I. Dose Metric Considerations in in Vitro Assays to Improve Quantitative in Vitro-in Vivo Dose Extrapolations. Toxicology 2015, 332, 30–40. [CrossRef]
- Meek, M.E.B.; Lipscomb, J.C. Gaining Acceptance for the Use of in Vitro Toxicity Assays and QIVIVE in Regulatory Risk Assessment. Toxicology 2015, 332, 112–123. [CrossRef]
- Patlewicz, G.; Fitzpatrick, J.M. Current and Future Perspectives on the Development, Evaluation, and Application of in Silico Approaches for Predicting Toxicity. Chem Res Toxicol 2016, 29, 438–451. [CrossRef]
- Combes, R.D. In Silico Methods for Toxicity Prediction. Adv Exp Med Biol 2012, 745, 96–116. [CrossRef]
- Serafini, M.M.; Sepehri, S.; Midali, M.; Stinckens, M.; Biesiekierska, M.; Wolniakowska, A.; Gatzios, A.; Rundén-Pran, E.; Reszka, E.; Marinovich, M.; et al. Recent Advances and Current Challenges of New Approach Methodologies in Developmental and Adult Neurotoxicity Testing. Arch Toxicol 2024, 98, 1271–1295. [CrossRef]
- Worth, A.P.; Patlewicz, G. Integrated Approaches to Testing and Assessment. Adv Exp Med Biol 2016, 856, 317–342. [CrossRef]
- Creton, S.; Dewhurst, I.C.; Earl, L.K.; Gehen, S.C.; Guest, R.L.; Hotchkiss, J.A.; Indans, I.; Woolhiser, M.R.; Billington, R. Acute Toxicity Testing of Chemicals-Opportunities to Avoid Redundant Testing and Use Alternative Approaches. Crit Rev Toxicol 2010, 40, 50–83. [CrossRef]
- Wilhelmus, K.R. The Draize Eye Test. Surv Ophthalmol 2001, 45, 493–515. [CrossRef]
- Wilson, S.L.; Ahearne, M.; Hopkinson, A. An Overview of Current Techniques for Ocular Toxicity Testing. Toxicology 2015, 327, 32–46. [CrossRef]
- Van Tonder, A.; Joubert, A.M.; Cromarty, A.D. Limitations of the 3-(4,5-Dimethylthiazol-2-Yl)-2,5-Diphenyl-2H-Tetrazolium Bromide (MTT) Assay When Compared to Three Commonly Used Cell Enumeration Assays. BMC Res Notes 2015, 8, 47. [CrossRef]
- Specian, A.F.L.; Serpeloni, J.M.; Tuttis, K.; Ribeiro, D.L.; Cilião, H.L.; Varanda, E.A.; Sannomiya, M.; Martinez-Lopez, W.; Vilegas, W.; Cólus, I.M.S. LDH, Proliferation Curves and Cell Cycle Analysis Are the Most Suitable Assays to Identify and Characterize New Phytotherapeutic Compounds. Cytotechnology 2016, 68, 2729. [CrossRef]
- Niles, A.L.; Moravec, R.A.; Riss, T.L. In Vitro Viability and Cytotoxicity Testing and Same-Well Multi-Parametric Combinations for High Throughput Screening. Curr Chem Genomics 2009, 3, 33. [CrossRef]
- Bacanli, M.; Anlar, H.G.; Başaran, A.A.; Başaran, N. Assessment of Cytotoxicity Profiles of Different Phytochemicals: Comparison of Neutral Red and MTT Assays in Different Cells in Different Time Periods. Turk J Pharm Sci 2017, 14, 95. [CrossRef]
- Bopp, S.K.; Lettieri, T. Comparison of Four Different Colorimetric and Fluorometric Cytotoxicity Assays in a Zebrafish Liver Cell Line. BMC Pharmacol 2008, 8, 8. [CrossRef]
- Gong, X.; Liang, Z.; Yang, Y.; Liu, H.; Ji, J.; Fan, Y. A Resazurin-Based, Nondestructive Assay for Monitoring Cell Proliferation during a Scaffold-Based 3D Culture Process. Regen Biomater 2020, 7, 271. [CrossRef]
- Żyro, D.; Radko, L.; Śliwińska, A.; Chęcińska, L.; Kusz, J.; Korona-Głowniak, I.; Przekora, A.; Wójcik, M.; Posyniak, A.; Ochocki, J. Multifunctional Silver(I) Complexes with Metronidazole Drug Reveal Antimicrobial Properties and Antitumor Activity against Human Hepatoma and Colorectal Adenocarcinoma Cells. Cancers (Basel) 2022, 14, 900. [CrossRef]
- Limame, R.; Wouters, A.; Pauwels, B.; Fransen, E.; Peeters, M.; Lardon, F.; de Wever, O.; Pauwels, P. Comparative Analysis of Dynamic Cell Viability, Migration and Invasion Assessments by Novel Real-Time Technology and Classic Endpoint Assays. PLoS One 2012, 7, e46536. [CrossRef]
- Figueiró, L.R.; Comerlato, L.C.; Da Silva, M.V.; Zuanazzi, J.Â.S.; Von Poser, G.L.; Ziulkoski, A.L. Toxicity of Glandularia Selloi (Spreng.) Tronc. Leave Extract by MTT and Neutral Red Assays: Influence of the Test Medium Procedure. Interdiscip Toxicol 2017, 9, 25. [CrossRef]
- Kourkopoulos, A.; Sijm, D.T.H.M.; Geerken, J.; Vrolijk, M.F. Compatibility and Interference of Food Simulants and Organic Solvents with the in Vitro Toxicological Assessment of Food Contact Materials. J Food Sci 2025, 90, e17659. [CrossRef]
- Oostingh, G.J.; Casals, E.; Italiani, P.; Colognato, R.; Stritzinger, R.; Ponti, J.; Pfaller, T.; Kohl, Y.; Ooms, D.; Favilli, F.; et al. Problems and Challenges in the Development and Validation of Human Cell-Based Assays to Determine Nanoparticle-Induced Immunomodulatory Effects. Part Fibre Toxicol 2011, 8, 8. [CrossRef]
- de Oliveira, L.C.B.; Nunes, H.L.; Ribeiro, D.L.; do Nascimento, J.R.; da Rocha, C.Q.; de Syllos Cólus, I.M.; Serpeloni, J.M. Aglycone Flavonoid Brachydin A Shows Selective Cytotoxicity and Antitumoral Activity in Human Metastatic Prostate (DU145) Cancer Cells. Cytotechnology 2021, 73, 761. [CrossRef]
- Lewis, M.A.; Patil, K.; Ettayebi, K.; Estes, M.K.; Atmar, R.L.; Ramani, S. Divergent Responses of Human Intestinal Organoid Monolayers Using Commercial in Vitro Cytotoxicity Assays. PLoS One 2024, 19, e0304526. [CrossRef]
- Tabernilla, A.; Rodrigues, B.D.S.; Pieters, A.; Caufriez, A.; Leroy, K.; Campenhout, R. Van; Cooreman, A.; Gomes, A.R.; Arnesdotter, E.; Gijbels, E.; et al. In Vitro Liver Toxicity Testing of Chemicals: A Pragmatic Approach. Int J Mol Sci 2021, 22, 5038. [CrossRef]
- Filer, D.L.; Kothiya, P.; Woodrow Setzer, R.; Judson, R.S.; Martin, M.T. Tcpl: The ToxCast Pipeline for High-Throughput Screening Data. Bioinformatics 2017, 33, 618–620. [CrossRef]
- Wang, J.; Hallinger, D.R.; Murr, A.S.; Buckalew, A.R.; Lougee, R.R.; Richard, A.M.; Laws, S.C.; Stoker, T.E. High-Throughput Screening and Chemotype-Enrichment Analysis of ToxCast Phase II Chemicals Evaluated for Human Sodium-Iodide Symporter (NIS) Inhibition. Environ Int 2019, 126, 377–386. [CrossRef]
- Chen, S.; Hsieh, J.H.; Huang, R.; Sakamuru, S.; Hsin, L.Y.; Xia, M.; Shockley, K.R.; Auerbach, S.; Kanaya, N.; Lu, H.; et al. Cell-Based High-Throughput Screening for Aromatase Inhibitors in the Tox21 10K Library. Toxicol Sci 2015, 147, 446–457. [CrossRef]
- Shi, Z.; Xia, M.; Xiao, S.; Zhang, Q. Identification of Nonmonotonic Concentration-Responses in Tox21 High-Throughput Screening Estrogen Receptor Assays. Toxicol Appl Pharmacol 2022, 452. [CrossRef]
- Stoker, T.E.; Wang, J.; Murr, A.S.; Bailey, J.R.; Buckalew, A.R. High-Throughput Screening of ToxCast PFAS Chemical Library for Potential Inhibitors of the Human Sodium Iodide Symporter. Chem Res Toxicol 2023, 36, 380–389. [CrossRef]
- Franiak-Pietryga, I.; Ostrowska, K.; Maciejewski, H.; Appelhans, D.; Misiewicz, M.; Ziemba, B.; Bednarek, M.; Bryszewska, M.; Borowiec, M. PPI-G4 Glycodendrimers Upregulate TRAIL-Induced Apoptosis in Chronic Lymphocytic Leukemia Cells. Macromol Biosci 2017, 17. [CrossRef]
- Huang, R. A Quantitative High-Throughput Screening Data Analysis Pipeline for Activity Profiling. Methods Mol Biol 2022, 2474, 133–145. [CrossRef]
- O’Brien, P.J. High-Content Analysis in Toxicology: Screening Substances for Human Toxicity Potential, Elucidating Subcellular Mechanisms and in Vivo Use as Translational Safety Biomarkers. Basic Clin Pharmacol Toxicol 2014, 115, 4–17. [CrossRef]
- Martin, H.L.; Adams, M.; Higgins, J.; Bond, J.; Morrison, E.E.; Bell, S.M.; Warriner, S.; Nelson, A.; Tomlinson, D.C. High-Content, High-Throughput Screening for the Identification of Cytotoxic Compounds Based on Cell Morphology and Cell Proliferation Markers. PLoS One 2014, 9. [CrossRef]
- Bray, M.A.; Singh, S.; Han, H.; Davis, C.T.; Borgeson, B.; Hartland, C.; Kost-Alimova, M.; Gustafsdottir, S.M.; Gibson, C.C.; Carpenter, A.E. Cell Painting, a High-Content Image-Based Assay for Morphological Profiling Using Multiplexed Fluorescent Dyes. Nat Protoc 2016, 11, 1757. [CrossRef]
- Nyffeler, J.; Willis, C.; Harris, F.R.; Foster, M.J.; Chambers, B.; Culbreth, M.; Brockway, R.E.; Davidson-Fritz, S.; Dawson, D.; Shah, I.; et al. Application of Cell Painting for Chemical Hazard Evaluation in Support of Screening-Level Chemical Assessments. Toxicol Appl Pharmacol 2023, 468. [CrossRef]
- Thakkar, Y.; Joshi, K.; Hickey, C.; Wahler, J.; Wall, B.; Etter, S.; Smith, B.; Griem, P.; Tate, M.; Jones, F.; et al. The BlueScreen HC Assay to Predict the Genotoxic Potential of Fragrance Materials. Mutagenesis 2022, 37, 13–23. [CrossRef]
- Moffat, J.G.; Rudolph, J.; Bailey, D. Phenotypic Screening in Cancer Drug Discovery - Past, Present and Future. Nat Rev Drug Discov 2014, 13, 588–602. [CrossRef]
- Bernardo, L.; Corallo, L.; Caterini, J.; Su, J.; Gisonni-Lex, L.; Gajewska, B. Application of XCELLigence Real-Time Cell Analysis to the Microplate Assay for Pertussis Toxin Induced Clustering in CHO Cells. PLoS One 2021, 16. [CrossRef]
- Martinez-Serra, J.; Ggutierrez, A.; Muñoz-Ccapó, S.; Nnavarro-Palou, M.; Rros, T.; Aamat, J.C.; Lopez, B.; Marcus, T.F.; Fueyo, L.; Suquia, A.G.; et al. XCELLigence System for Real-Time Label-Free Monitoring of Growth and Viability of Cell Lines from Hematological Malignancies. Onco Targets Ther 2014, 7, 985–994. [CrossRef]
- Ramis, G.; Martínez-Alarcón, L.; Quereda, J.J.; Mendonça, L.; Majado, M.J.; Gomez-Coelho, K.; Mrowiec, A.; Herrero-Medrano, J.M.; Abellaneda, J.M.; Pallares, F.J.; et al. Optimization of Cytotoxicity Assay by Real-Time, Impedance-Based Cell Analysis. Biomed Microdevices 2013, 15, 985–995. [CrossRef]
- Bonnevier, J.; Hammerbeck, C.; Goetz, C. Flow Cytometry: Definition, History, and Uses in Biological Research. 2018, 1–11. [CrossRef]
- Pant, A.B.; Khare, P.; Pandey, A.K. Flow Cytometry: Applications in Cellular and Molecular Toxicology. Flow Cytometry: Applications in Cellular and Molecular Toxicology 2025, 1–304. [CrossRef]
- Franiak-Pietryga, I.; Ziółkowska, E.; Ziemba, B.; Appelhans, D.; Maciejewski, H.; Voit, B.; Kaczmarek, A.; Robak, T.; Klajnert-Maculewicz, B.; Cebula-Obrzut, B.; et al. Glycodendrimer PPI as a Potential Drug in Chronic Lymphocytic Leukaemia. The Influence of Glycodendrimer on Apoptosis in In Vitro B-CLL Cells Defined by Microarrays. Anticancer Agents Med Chem 2022, 17, 102–114. [CrossRef]
- Holder, A.L.; Goth-Goldstein, R.; Lucas, D.; Koshland, C.P. Particle-Induced Artifacts in the MTT and LDH Viability Assays. Chem Res Toxicol 2012, 25, 1885–1892. [CrossRef]
- de la Fuente-Jiménez, J.L.; Rodríguez-Rivas, C.I.; Mitre-Aguilar, I.B.; Torres-Copado, A.; García-López, E.A.; Herrera-Celis, J.; Arvizu-Espinosa, M.G.; Garza-Navarro, M.A.; Arriaga, L.G.; García, J.L.; et al. A Comparative and Critical Analysis for In Vitro Cytotoxic Evaluation of Magneto-Crystalline Zinc Ferrite Nanoparticles Using MTT, Crystal Violet, LDH, and Apoptosis Assay. Int J Mol Sci 2023, 24, 12860. [CrossRef]
- Fowler, P.; Smith, K.; Young, J.; Jeffrey, L.; Kirkland, D.; Pfuhler, S.; Carmichael, P. Reduction of Misleading (“ False”) Positive Results in Mammalian Cell Genotoxicity Assays. I. Choice of Cell Type. Mutat Res Genet Toxicol Environ Mutagen 2012, 742, 11–25. [CrossRef]
- Fowler, P.; Smith, R.; Smith, K.; Young, J.; Jeffrey, L.; Kirkland, D.; Pfuhler, S.; Carmichael, P. Reduction of Misleading (“ False” ) Positive Results in Mammalian Cell Genotoxicity Assays. II. Importance of Accurate Toxicity Measurement. Mutat Res Genet Toxicol Environ Mutagen 2012, 747, 104–117. [CrossRef]
- Fowler, P.; Smith, R.; Smith, K.; Young, J.; Jeffrey, L.; Carmichael, P.; Kirkland, D.; Pfuhler, S. Reduction of Misleading (“false”) Positive Results in Mammalian Cell Genotoxicity Assays. III: Sensitivity of Human Cell Types to Known Genotoxic Agents. Mutat Res Genet Toxicol Environ Mutagen 2014, 767, 28–36. [CrossRef]
- Watson, C.; Ge, J.; Cohen, J.; Pyrgiotakis, G.; Engelward, B.P.; Demokritou, P. High-Throughput Screening Platform for Engineered Nanoparticle-Mediated Genotoxicity Using CometChip Technology. ACS Nano 2014, 8, 2118–2133. [CrossRef]
- Zhao, R.; Yanamandra, A.K.; Qu, B. A High-Throughput 3D Kinetic Killing Assay. Eur J Immunol 2023, 53. [CrossRef]
- Olivo Pimentel, V.; Yaromina, A.; Marcus, D.; Dubois, L.J.; Lambin, P. A Novel Co-Culture Assay to Assess Anti-Tumor CD8+ T Cell Cytotoxicity via Luminescence and Multicolor Flow Cytometry. J Immunol Methods 2020, 487. [CrossRef]
- Chen, Z.; Han, S.; Sanny, A.; Chan, D.L.K.; van Noort, D.; Lim, W.; Tan, A.H.M.; Park, S. 3D Hanging Spheroid Plate for High-Throughput CAR T Cell Cytotoxicity Assay. J Nanobiotechnology 2022, 20. [CrossRef]
- Park, D.; Son, K.; Hwang, Y.; Ko, J.; Lee, Y.; Doh, J.; Jeon, N.L. High-Throughput Microfluidic 3D Cytotoxicity Assay for Cancer Immunotherapy (CACI-IMPACT Platform). Front Immunol 2019, 10. [CrossRef]
- Tong, Z. Bin; Huang, R.; Braisted, J.; Chu, P.H.; Simeonov, A.; Gerhold, D.L. 3D-Suspension Culture Platform for High Throughput Screening of Neurotoxic Chemicals Using LUHMES Dopaminergic Neurons. SLAS Discovery 2024, 29. [CrossRef]
- Zhang, C.J.; Meyer, S.R.; O’Meara, M.J.; Huang, S.; Capeling, M.M.; Ferrer-Torres, D.; Childs, C.J.; Spence, J.R.; Fontana, R.J.; Sexton, J.Z. A Human Liver Organoid Screening Platform for DILI Risk Prediction. J Hepatol 2023, 78, 998–1006. [CrossRef]
- Sun, B.; Liang, Z.; Wang, Y.; Yu, Y.; Zhou, X.; Geng, X.; Li, B. A 3D Spheroid Model of Quadruple Cell Co-Culture with Improved Liver Functions for Hepatotoxicity Prediction. Toxicology 2024, 505. [CrossRef]
- Karassina, N.; Hofsteen, P.; Cali, J.J.; Vidugiriene, J. Time- and Dose-Dependent Toxicity Studies in 3D Cultures Using a Luminescent Lactate Dehydrogenase Assay. Methods Mol Biol 2021, 2255, 77–86. [CrossRef]
- Dominijanni, A.J.; Devarasetty, M.; Forsythe, S.D.; Votanopoulos, K.I.; Soker, S. Cell Viability Assays in Three-Dimensional Hydrogels: A Comparative Study of Accuracy. Tissue Eng Part C Methods 2021, 27, 401–410. [CrossRef]
- Hurrell, T.; Lilley, K.S.; Cromarty, A.D. Proteomic Responses of HepG2 Cell Monolayers and 3D Spheroids to Selected Hepatotoxins. Toxicol Lett 2019, 300, 40–50. [CrossRef]
- Schmeisser, S.; Miccoli, A.; von Bergen, M.; Berggren, E.; Braeuning, A.; Busch, W.; Desaintes, C.; Gourmelon, A.; Grafström, R.; Harrill, J.; et al. New Approach Methodologies in Human Regulatory Toxicology – Not If, but How and When! Environ Int 2023, 178. [CrossRef]
- Chang, X.; Tan, Y.M.; Allen, D.G.; Bell, S.; Brown, P.C.; Browning, L.; Ceger, P.; Gearhart, J.; Hakkinen, P.J.; Kabadi, S. V.; et al. IVIVE: Facilitating the Use of In Vitro Toxicity Data in Risk Assessment and Decision Making. Toxics 2022, 10. [CrossRef]
- Breen, M.; Ring, C.L.; Kreutz, A.; Goldsmith, M.R.; Wambaugh, J.F. High-Throughput PBTK Models for in Vitro to in Vivo Extrapolation. Expert Opin Drug Metab Toxicol 2021, 17, 903–921. [CrossRef]
- Arya, V.; Venkatakrishnan, K. Role of Physiologically Based Pharmacokinetic Modeling and Simulation in Enabling Model-Informed Development of Drugs and Biotherapeutics. J Clin Pharmacol 2020, 60 Suppl 1, S7–S11. [CrossRef]
- Lee, O.W.; Austin, S.; Gamma, M.; Cheff, D.M.; Lee, T.D.; Wilson, K.M.; Johnson, J.; Travers, J.; Braisted, J.C.; Guha, R.; et al. Cytotoxic Profiling of Annotated and Diverse Chemical Libraries Using Quantitative High-Throughput Screening. SLAS Discovery 2020, 25, 9–20. [CrossRef]
- Sambale, F.; Lavrentieva, A.; Stahl, F.; Blume, C.; Stiesch, M.; Kasper, C.; Bahnemann, D.; Scheper, T. Three Dimensional Spheroid Cell Culture for Nanoparticle Safety Testing. J Biotechnol 2015, 205, 120–129. [CrossRef]
- Krug, A.K.; Kolde, R.; Gaspar, J.A.; Rempel, E.; Balmer, N. V.; Meganathan, K.; Vojnits, K.; Baquié, M.; Waldmann, T.; Ensenat-Waser, R.; et al. Human Embryonic Stem Cell-Derived Test Systems for Developmental Neurotoxicity: A Transcriptomics Approach. Arch Toxicol 2013, 87, 123–143. [CrossRef]
- Gunaseeli, I.; Doss, M.; Antzelevitch, C.; Hescheler, J.; Sachinidis, A. Induced Pluripotent Stem Cells as a Model for Accelerated Patient- and Disease-Specific Drug Discovery. Curr Med Chem 2010, 17, 759–766. [CrossRef]
- Shinde, V.; Klima, S.; Sureshkumar, P.S.; Meganathan, K.; Jagtap, S.; Rempel, E.; Rahnenführer, J.; Hengstler, J.G.; Waldmann, T.; Hescheler, J.; et al. Human Pluripotent Stem Cell Based Developmental Toxicity Assays for Chemical Safety Screening and Systems Biology Data Generation. J Vis Exp 2015, 2015. [CrossRef]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic Stem Cell Lines Derived from Human Blastocysts. Science (1979) 1998, 282, 1145–1147. [CrossRef]
- Magdy, T.; Schuldt, A.J.T.; Wu, J.C.; Bernstein, D.; Burridge, P.W. Human Induced Pluripotent Stem Cell (HiPSC)-Derived Cells to Assess Drug Cardiotoxicity: Opportunities and Problems. Annu Rev Pharmacol Toxicol 2018, 58, 83–103. [CrossRef]
- Varghese, D.S.; Alawathugoda, T.T.; Ansari, S.A. Fine Tuning of Hepatocyte Differentiation from Human Embryonic Stem Cells: Growth Factor vs. Small Molecule-Based Approaches. Stem Cells Int 2019, 2019. [CrossRef]
- Baxter, M.; Withey, S.; Harrison, S.; Segeritz, C.P.; Zhang, F.; Atkinson-Dell, R.; Rowe, C.; Gerrard, D.T.; Sison-Young, R.; Jenkins, R.; et al. Phenotypic and Functional Analyses Show Stem Cell-Derived Hepatocyte-like Cells Better Mimic Fetal Rather than Adult Hepatocytes. J Hepatol 2015, 62, 581–589. [CrossRef]
- Hoelting, L.; Scheinhardt, B.; Bondarenko, O.; Schildknecht, S.; Kapitza, M.; Tanavde, V.; Tan, B.; Lee, Q.Y.; Mecking, S.; Leist, M.; et al. A 3-Dimensional Human Embryonic Stem Cell (HESC)-Derived Model to Detect Developmental Neurotoxicity of Nanoparticles. Arch Toxicol 2013, 87, 721–733. [CrossRef]
- Yeo, Y.; Tan, J.B.L.; Lim, L.W.; Tan, K.O.; Heng, B.C.; Lim, W.L. Human Embryonic Stem Cell-Derived Neural Lineages as In Vitro Models for Screening the Neuroprotective Properties of Lignosus Rhinocerus (Cooke) Ryvarden. Biomed Res Int 2019, 2019. [CrossRef]
- Kotasová, H.; Capandová, M.; Pelková, V.; Dumková, J.; Koledová, Z.; Remšík, J.; Souček, K.; Garlíková, Z.; Sedláková, V.; Rabata, A.; et al. Expandable Lung Epithelium Differentiated from Human Embryonic Stem Cells. Tissue Eng Regen Med 2022, 19, 1033–1050. [CrossRef]
- Ryu, B.; Son, M.Y.; Jung, K.B.; Kim, U.; Kim, J.; Kwon, O.; Son, Y.S.; Jung, C.R.; Park, J.H.; Kim, C.Y. Next-Generation Intestinal Toxicity Model of Human Embryonic Stem Cell-Derived Enterocyte-Like Cells. Front Vet Sci 2021, 8. [CrossRef]
- van der Torren, C.R.; Zaldumbide, A.; Duinkerken, G.; Brand-Schaaf, S.H.; Peakman, M.; Stangé, G.; Martinson, L.; Kroon, E.; Brandon, E.P.; Pipeleers, D.; et al. Immunogenicity of Human Embryonic Stem Cell-Derived Beta Cells. Diabetologia 2017, 60, 126–133. [CrossRef]
- Tan, H.L.; Tan, B.Z.; Goh, W.X.T.; Cua, S.; Choo, A. In Vivo Surveillance and Elimination of Teratoma-Forming Human Embryonic Stem Cells with Monoclonal Antibody 2448 Targeting Annexin A2. Biotechnol Bioeng 2019, 116, 2996–3005. [CrossRef]
- Lin, Y.T.; Wang, C.K.; Yang, S.C.; Hsu, S.C.; Lin, H.; Chang, F.P.; Kuo, T.C.; Shen, C.N.; Chiang, P.M.; Hsiao, M.; et al. Elimination of Undifferentiated Human Embryonic Stem Cells by Cardiac Glycosides. Sci Rep 2017, 7. [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [CrossRef]
- Hansen, J.; Xiong, Y.; Siddiq, M.M.; Dhanan, P.; Hu, B.; Shewale, B.; Yadaw, A.S.; Jayaraman, G.; Tolentino, R.E.; Chen, Y.; et al. Multiscale Mapping of Transcriptomic Signatures for Cardiotoxic Drugs. Nat Commun 2024, 15. [CrossRef]
- Walker, L.M.; Sparks, N.R.L.; Puig-Sanvicens, V.; Rodrigues, B.; Zur Nieden, N.I. An Evaluation of Human Induced Pluripotent Stem Cells to Test for Cardiac Developmental Toxicity. Int J Mol Sci 2021, 22. [CrossRef]
- Shinozawa, T.; Kimura, M.; Cai, Y.; Saiki, N.; Yoneyama, Y.; Ouchi, R.; Koike, H.; Maezawa, M.; Zhang, R.R.; Dunn, A.; et al. High-Fidelity Drug-Induced Liver Injury Screen Using Human Pluripotent Stem Cell–Derived Organoids. Gastroenterology 2021, 160, 831-846.e10. [CrossRef]
- Carberry, C.K.; Ferguson, S.S.; Beltran, A.S.; Fry, R.C.; Rager, J.E. Using Liver Models Generated from Human-Induced Pluripotent Stem Cells (IPSCs) for Evaluating Chemical-Induced Modifications and Disease across Liver Developmental Stages. Toxicol In Vitro 2022, 83. [CrossRef]
- Kato, Y.; Inaba, T.; Shinke, K.; Hiramatsu, N.; Horie, T.; Sakamoto, T.; Hata, Y.; Sugihara, E.; Takimoto, T.; Nagai, N.; et al. Comprehensive Search for Genes Involved in Thalidomide Teratogenicity Using Early Differentiation Models of Human Induced Pluripotent Stem Cells: Potential Applications in Reproductive and Developmental Toxicity Testing. Cells 2025, 14. [CrossRef]
- Wu, S.; Xie, J.; Zhao, H.; Zhao, X.; Sánchez, O.F.; Rochet, J.C.; Freeman, J.L.; Yuan, C. Developmental Neurotoxicity of PFOA Exposure on HiPSC-Derived Cortical Neurons. Environ Int 2024, 190. [CrossRef]
- Kandasamy, K.; Chuah, J.K.C.; Su, R.; Huang, P.; Eng, K.G.; Xiong, S.; Li, Y.; Chia, C.S.; Loo, L.H.; Zink, D. Prediction of Drug-Induced Nephrotoxicity and Injury Mechanisms with Human Induced Pluripotent Stem Cell-Derived Cells and Machine Learning Methods. Sci Rep 2015, 5. [CrossRef]
- Achberger, K.; Probst, C.; Haderspeck, J.C.; Bolz, S.; Rogal, J.; Chuchuy, J.; Nikolova, M.; Cora, V.; Antkowiak, L.; Haq, W.; et al. Merging Organoid and Organ-on-a-Chip Technology to Generate Complex Multi-Layer Tissue Models in a Human Retina-on-a-Chip Platform. Elife 2019, 8. [CrossRef]
- Weng, K.C.; Kurokawa, Y.K.; Hajek, B.S.; Paladin, J.A.; Shirure, V.S.; George, S.C. Human Induced Pluripotent Stem-Cardiac-Endothelial-Tumor-on-a-Chip to Assess Anticancer Efficacy and Cardiotoxicity. Tissue Eng Part C Methods 2020, 26, 44–55. [CrossRef]
- Lauschke, K.; Rosenmai, A.K.; Meiser, I.; Neubauer, J.C.; Schmidt, K.; Rasmussen, M.A.; Holst, B.; Taxvig, C.; Emnéus, J.K.; Vinggaard, A.M. A Novel Human Pluripotent Stem Cell-Based Assay to Predict Developmental Toxicity. Arch Toxicol 2020, 94, 3831–3846. [CrossRef]
- Moreau, M.; Antonijevic, T.; Hall, J.C.; Fisher, J. In Vitro and Computational Approaches to Predict Developmental Toxicity: Integrating PBPK Models with Cell-Based Assays. Toxicol Appl Pharmacol 2025, 502. [CrossRef]
- Galanjuk, S.; Zühr, E.; Dönmez, A.; Bartsch, D.; Kurian, L.; Tigges, J.; Fritsche, E. The Human Induced Pluripotent Stem Cell Test as an Alternative Method for Embryotoxicity Testing. Int J Mol Sci 2022, 23. [CrossRef]
- Yang, J.; Lei, W.; Xiao, Y.; Tan, S.; Yang, J.; Lin, Y.; Yang, Z.; Zhao, D.; Zhang, C.; Shen, Z.; et al. Generation of Human Vascularized and Chambered Cardiac Organoids for Cardiac Disease Modelling and Drug Evaluation. Cell Prolif 2024, 57. [CrossRef]
- Tidball, A.M.; Niu, W.; Ma, Q.; Takla, T.N.; Walker, J.C.; Margolis, J.L.; Mojica-Perez, S.P.; Sudyk, R.; Deng, L.; Moore, S.J.; et al. Deriving Early Single-Rosette Brain Organoids from Human Pluripotent Stem Cells. Stem Cell Reports 2023, 18, 2498–2514. [CrossRef]
- Mariani, A.; Comolli, D.; Fanelli, R.; Forloni, G.; De Paola, M. Neonicotinoid Pesticides Affect Developing Neurons in Experimental Mouse Models and in Human Induced Pluripotent Stem Cell (IPSC)-Derived Neural Cultures and Organoids. Cells 2024, 13. [CrossRef]
- Association, W.M. World Medical Association Declaration of Helsinki: Ethical Principles for Medical Research Involving Human Subjects. JAMA 2013, 310, 2191–2194. [CrossRef]
- Directive - 2004/23 - EN - EU Tissue Directive - EUR-Lex Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:32004L0023 (accessed on 6 October 2025).
- HESCRegistry - Public Lines | STEM Cell Information Available online: https://stemcells.nih.gov/registry/eligible-to-use-lines (accessed on 6 October 2025).
- Lovell-Badge, R.; Anthony, E.; Barker, R.A.; Bubela, T.; Brivanlou, A.H.; Carpenter, M.; Charo, R.A.; Clark, A.; Clayton, E.; Cong, Y.; et al. ISSCR Guidelines for Stem Cell Research and Clinical Translation: The 2021 Update. Stem Cell Reports 2021, 16, 1398–1408. [CrossRef]
- Weltner, J.; Lanner, F. Refined Transcriptional Blueprint of Human Preimplantation Embryos. Cell Stem Cell 2021, 28, 1503–1504. [CrossRef]
- Schuster, P. Some Mechanistic Requirements for Major Transitions. Philosophical Transactions of the Royal Society B: Biological Sciences 2016, 371. [CrossRef]
- Aalto-Setälä, K.; Conklin, B.R.; Lo, B. Obtaining Consent for Future Research with Induced Pluripotent Cells: Opportunities and Challenges. PLoS Biol 2009, 7, e1000042. [CrossRef]
- Lo, B.; Parham, L. Ethical Issues in Stem Cell Research. Endocr Rev 2009, 30, 204–213. [CrossRef]
- Orzechowski, M.; Schochow, M.; Kühl, M.; Steger, F. Content and Method of Information for Participants in Clinical Studies With Induced Pluripotent Stem Cells (IPSCs). Front Cell Dev Biol 2021, 9, 627816. [CrossRef]
- Kropf, M. Ethical Aspects of Human Induced Pluripotent Stem Cells and Alzheimer’s Disease: Potentials and Challenges of a Seemingly Harmless Method. J Alzheimers Dis Rep 2023, 7, 993–1006. [CrossRef]
- Witt, G.; Keminer, O.; Leu, J.; Tandon, R.; Meiser, I.; Willing, A.; Winschel, I.; Abt, J.C.; Brändl, B.; Sébastien, I.; et al. An Automated and High-Throughput-Screening Compatible Pluripotent Stem Cell-Based Test Platform for Developmental and Reproductive Toxicity Assessment of Small Molecule Compounds. Cell Biol Toxicol 2021, 37, 229–243. [CrossRef]
- Lauschke, K.; Treschow, A.F.; Rasmussen, M.A.; Davidsen, N.; Holst, B.; Emnéus, J.; Taxvig, C.; Vinggaard, A.M. Creating a Human-Induced Pluripotent Stem Cell-Based NKX2.5 Reporter Gene Assay for Developmental Toxicity Testing. Arch Toxicol 2021, 95, 1659–1670. [CrossRef]
- Han, I.C.; Bohrer, L.R.; Gibson-Corley, K.N.; Wiley, L.A.; Shrestha, A.; Harman, B.E.; Jiao, C.; Sohn, E.H.; Wendland, R.; Allen, B.N.; et al. Biocompatibility of Human Induced Pluripotent Stem Cell-Derived Retinal Progenitor Cell Grafts in Immunocompromised Rats. Cell Transplant 2022, 31. [CrossRef]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal Stem Cell Perspective: Cell Biology to Clinical Progress. NPJ Regen Med 2019, 4, 1–15. [CrossRef]
- Pasternak-Mnich, K.; Szwed-Georgiou, A.; Ziemba, B.; Pieszyński, I.; Bryszewska, M.; Kujawa, J. Effect of Photobiomodulation Therapy on the Morphology, Intracellular Calcium Concentration, Free Radical Generation, Apoptosis and Necrosis of Human Mesenchymal Stem Cells-an in Vitro Study. Lasers Med Sci 2024, 39. [CrossRef]
- Ezeh, P.C.; Xu, H.; Wang, S.C.; Medina, S.; Burchiel, S.W. CURRENT PROTOCOLS IN TOXICOLOGY: Evaluation of Toxicity in Mouse Bone Marrow Progenitor Cells. Current protocols in toxicology / editorial board, Mahin D. Maines (editor-in-chief) ... [et al.] 2016, 67, 18.9.1. [CrossRef]
- Yadav, N.K.; Shukla, P.; Omer, A.; Singh, R.K. In-Vitro Hematological Toxicity Prediction by Colony-Forming Cell Assays. Toxicol Environ Health Sci 2013, 5, 169–176. [CrossRef]
- Parent-Massin, D.; Hymery, N.; Sibiril, Y. Stem Cells in Myelotoxicity. Toxicology 2010, 267, 112–117. [CrossRef]
- Liu, X.; Yang, Z.; Sun, J.; Ma, T.; Hua, F.; Shen, Z. A Brief Review of Cytotoxicity of Nanoparticles on Mesenchymal Stem Cells in Regenerative Medicine. Int J Nanomedicine 2019, 14, 3875. [CrossRef]
- Christodoulou, I.; Goulielmaki, M.; Kritikos, A.; Zoumpourlis, P.; Koliakos, G.; Zoumpourlis, V. Suitability of Human Mesenchymal Stem Cells Derived from Fetal Umbilical Cord (Wharton’s Jelly) as an Alternative In Vitro Model for Acute Drug Toxicity Screening. Cells 2022, 11, 1102. [CrossRef]
- Sharma, S.; Venkatesan, V.; Prakhya, B.M.; Bhonde, R. Human Mesenchymal Stem Cells as a Novel Platform for Simultaneous Evaluation of Cytotoxicity and Genotoxicity of Pharmaceuticals. Mutagenesis 2015, 30, 391–399. [CrossRef]
- Hu, B.; Cheng, Z.; Liang, S. Advantages and Prospects of Stem Cells in Nanotoxicology. Chemosphere 2022, 291, 132861. [CrossRef]
- Tayebi, B.; Babaahmadi, M.; Pakzad, M.; Hajinasrollah, M.; Mostafaei, F.; Jahangiri, S.; Kamali, A.; Baharvand, H.; Baghaban Eslaminejad, M.; Hassani, S.N.; et al. Standard Toxicity Study of Clinical-Grade Allogeneic Human Bone Marrow-Derived Clonal Mesenchymal Stromal Cells. Stem Cell Res Ther 2022, 13, 1–18. [CrossRef]
- Gong, Y.; Ji, Y.; Liu, F.; Li, J.; Cao, Y. Cytotoxicity, Oxidative Stress and Inflammation Induced by ZnO Nanoparticles in Endothelial Cells: Interaction with Palmitate or Lipopolysaccharide. J Appl Toxicol 2017, 37, 895–901. [CrossRef]
- Ghosh, M.; Sinha, S.; Jothiramajayam, M.; Jana, A.; Nag, A.; Mukherjee, A. Cyto-Genotoxicity and Oxidative Stress Induced by Zinc Oxide Nanoparticle in Human Lymphocyte Cells in Vitro and Swiss Albino Male Mice in Vivo. Food and Chemical Toxicology 2016, 97, 286–296. [CrossRef]
- Zhang, L.; Haddouti, E.M.; Beckert, H.; Biehl, R.; Pariyar, S.; Rüwald, J.M.; Li, X.; Jaenisch, M.; Burger, C.; Wirtz, D.C.; et al. Investigation of Cytotoxicity, Oxidative Stress, and Inflammatory Responses of Tantalum Nanoparticles in THP-1-Derived Macrophages. Mediators Inflamm 2020, 2020. [CrossRef]
- Ziemba, B.; Franiak-Pietryga, I.; Pion, M.; Appelhans, D.; Muñoz-Fernández, M.Á.; Voit, B.; Bryszewska, M.; Klajnert-Maculewicz, B. Toxicity and Proapoptotic Activity of Poly(Propylene Imine) Glycodendrimers in Vitro: Considering Their Contrary Potential as Biocompatible Entity and Drug Molecule in Cancer. Int J Pharm 2014, 461, 391–402. [CrossRef]
- Busch, M.; Brouwer, H.; Aalderink, G.; Bredeck, G.; Kämpfer, A.A.M.; Schins, R.P.F.; Bouwmeester, H. Investigating Nanoplastics Toxicity Using Advanced Stem Cell-Based Intestinal and Lung in Vitro Models. Frontiers in toxicology 2023, 5. [CrossRef]
- Costa, S.; Vilas-Boas, V.; Lebre, F.; Granjeiro, J.M.; Catarino, C.M.; Moreira Teixeira, L.; Loskill, P.; Alfaro-Moreno, E.; Ribeiro, A.R. Microfluidic-Based Skin-on-Chip Systems for Safety Assessment of Nanomaterials. Trends Biotechnol 2023, 41, 1282–1298. [CrossRef]
- Dubiak-Szepietowska, M.; Karczmarczyk, A.; Jönsson-Niedziółka, M.; Winckler, T.; Feller, K.H. Development of Complex-Shaped Liver Multicellular Spheroids as a Human-Based Model for Nanoparticle Toxicity Assessment in Vitro. Toxicol Appl Pharmacol 2016, 294, 78–85. [CrossRef]
- Huh, D.; Matthews, B.D.; Mammoto, A.; Montoya-Zavala, M.; Yuan Hsin, H.; Ingber, D.E. Reconstituting Organ-Level Lung Functions on a Chip. Science 2010, 328, 1662–1668. [CrossRef]
- Benameur, L.; Auffan, M.; Cassien, M.; Liu, W.; Culcasi, M.; Rahmouni, H.; Stocker, P.; Tassistro, V.; Bottero, J.Y.; Rose, J.; et al. DNA Damage and Oxidative Stress Induced by CeO2 Nanoparticles in Human Dermal Fibroblasts: Evidence of a Clastogenic Effect as a Mechanism of Genotoxicity. Nanotoxicology 2015, 9, 696–705. [CrossRef]
- Khalid, H.; Mukherjee, S.P.; O’Neill, L.; Byrne, H.J. Structural Dependence of in Vitro Cytotoxicity, Oxidative Stress and Uptake Mechanisms of Poly(Propylene Imine) Dendritic Nanoparticles. J Appl Toxicol 2016, 36, 464–473. [CrossRef]
- Magdolenova, Z.; Drlickova, M.; Henjum, K.; Rundén-Pran, E.; Tulinska, J.; Bilanicova, D.; Pojana, G.; Kazimirova, A.; Barancokova, M.; Kuricova, M.; et al. Coating-Dependent Induction of Cytotoxicity and Genotoxicity of Iron Oxide Nanoparticles. Nanotoxicology 2015, 9 Suppl 1, 44–56. [CrossRef]
- Buerki-Thurnherr, T.; Xiao, L.; Diener, L.; Arslan, O.; Hirsch, C.; Maeder-Althaus, X.; Grieder, K.; Wampfler, B.; Mathur, S.; Wick, P.; et al. In Vitro Mechanistic Study towards a Better Understanding of ZnO Nanoparticle Toxicity. Nanotoxicology 2013, 7, 402–416. [CrossRef]
- Franiak-Pietryga, I.; Ziemba, B.; Sikorska, H.; Jander, M.; Appelhans, D.; Bryszewska, M.; Borowiec, M. Neurotoxicity of Poly(Propylene Imine) Glycodendrimers. Drug Chem Toxicol 2022, 45, 1484–1492. [CrossRef]
- Gorzkiewicz, M.; Sztandera, K.; Jatczak-Pawlik, I.; Zinke, R.; Appelhans, D.; Klajnert-Maculewicz, B.; Pulaski, Ł. Terminal Sugar Moiety Determines Immunomodulatory Properties of Poly(Propyleneimine) Glycodendrimers. Biomacromolecules 2018, 19, 1562–1572. [CrossRef]
- Illarionova, N.B.; Morozova, K.N.; Petrovskii, D. V.; Sharapova, M.B.; Romashchenko, A. V.; Troitskii, S.Y.; Kiseleva, E.; Moshkin, Y.M.; Moshkin, M.P. “Trojan-Horse” Stress-Granule Formation Mediated by Manganese Oxide Nanoparticles. Nanotoxicology 2020, 14, 1432–1444. [CrossRef]
- Liu, N.; Liang, Y.; Wei, T.; Huang, X.; Zhang, T.; Tang, M. ROS-Mediated NRF2/p-ERK1/2 Signaling-Involved Mitophagy Contributes to Macrophages Activation Induced by CdTe Quantum Dots. Toxicology 2024, 505. [CrossRef]
- Almutary, A.; Sanderson, B.J.S. The MTT and Crystal Violet Assays: Potential Confounders in Nanoparticle Toxicity Testing. Int J Toxicol 2016, 35, 454–462. [CrossRef]
- Eder, K.M.; Marzi, A.; Wågbø, A.M.; Vermeulen, J.P.; de la Fonteyne-Blankestijn, L.J.J.; Rösslein, M.; Ossig, R.; Klinkenberg, G.; Vandebriel, R.J.; Schnekenburger, J. Standardization of an in Vitro Assay Matrix to Assess Cytotoxicity of Organic Nanocarriers: A Pilot Interlaboratory Comparison. Drug Deliv Transl Res 2022, 12, 2187–2206. [CrossRef]
- Ziemba, B.; Matuszko, G.; Bryszewska, M.; Klajnert, B. Influence of Dendrimers on Red Blood Cells. Cell Mol Biol Lett 2012, 17, 21–35. [CrossRef]
- Wrobel, D.; Marcinkowska, M.; Janaszewska, A.; Appelhans, D.; Voit, B.; Klajnert-Maculewicz, B.; Bryszewska, M.; Štofik, M.; Herma, R.; Duchnowicz, P.; et al. Influence of Core and Maltose Surface Modification of PEIs on Their Interaction with Plasma Proteins—Human Serum Albumin and Lysozyme. Colloids Surf B Biointerfaces 2017, 152, 18–28. [CrossRef]
- Kämpfer, A.A.M.; Busch, M.; Schins, R.P.F. Advanced In Vitro Testing Strategies and Models of the Intestine for Nanosafety Research. Chem Res Toxicol 2020, 33, 1163–1178. [CrossRef]
- Baek, A.; Kwon, I.H.; Lee, D.H.; Choi, W.H.; Lee, S.W.; Yoo, J.; Heo, M.B.; Lee, T.G. Novel Organoid Culture System for Improved Safety Assessment of Nanomaterials. Nano Lett 2024, 24, 805–813. [CrossRef]
- Franiak-Pietryga, I.; Ostrowska, K.; Maciejewski, H.; Ziemba, B.; Appelhans, D.; Voit, B.; Jander, M.; Treliński, J.; Bryszewska, M.; Borowiec, M. Affecting NF-ΚB Cell Signaling Pathway in Chronic Lymphocytic Leukemia by Dendrimers-Based Nanoparticles. Toxicol Appl Pharmacol 2018, 357, 33–38. [CrossRef]
- Nikonorova, V.G.; Chrishtop, V. V.; Mironov, V.A.; Prilepskii, A.Y. Advantages and Potential Benefits of Using Organoids in Nanotoxicology. Cells 2023, 12. [CrossRef]
- Lucchetti, M.; Aina, K.O.; Grandmougin, L.; Jäger, C.; Pérez Escriva, P.; Letellier, E.; Mosig, A.S.; Wilmes, P. An Organ-on-Chip Platform for Simulating Drug Metabolism Along the Gut-Liver Axis. Adv Healthc Mater 2024, 13. [CrossRef]
- Lawlor, K.T.; Vanslambrouck, J.M.; Higgins, J.W.; Chambon, A.; Bishard, K.; Arndt, D.; Er, P.X.; Wilson, S.B.; Howden, S.E.; Tan, K.S.; et al. Cellular Extrusion Bioprinting Improves Kidney Organoid Reproducibility and Conformation. Nat Mater 2021, 20, 260–271. [CrossRef]
- Meyer, S.R.; Zhang, C.J.; Garcia, M.A.; Procario, M.C.; Yoo, S.; Jolly, A.L.; Kim, S.; Kim, J.; Baek, K.; Kersten, R.D.; et al. A High-Throughput Microphysiological Liver Chip System to Model Drug-Induced Liver Injury Using Human Liver Organoids. Gastro Hep Advances 2024, 3, 1045–1053. [CrossRef]
- Bronsard, J.; Savary, C.; Massart, J.; Viel, R.; Moutaux, L.; Catheline, D.; Rioux, V.; Clement, B.; Corlu, A.; Fromenty, B.; et al. 3D Multi-Cell-Type Liver Organoids: A New Model of Non-Alcoholic Fatty Liver Disease for Drug Safety Assessments. Toxicology in Vitro 2024, 94. [CrossRef]
- Vanslambrouck, J.M.; Wilson, S.B.; Tan, K.S.; Groenewegen, E.; Rudraraju, R.; Neil, J.; Lawlor, K.T.; Mah, S.; Scurr, M.; Howden, S.E.; et al. Enhanced Metanephric Specification to Functional Proximal Tubule Enables Toxicity Screening and Infectious Disease Modelling in Kidney Organoids. Nat Commun 2022, 13. [CrossRef]
- Dilz, J.; Auge, I.; Groeneveld, K.; Reuter, S.; Mrowka, R. A Proof-of-Concept Assay for Quantitative and Optical Assessment of Drug-Induced Toxicity in Renal Organoids. Sci Rep 2023, 13. [CrossRef]
- Yousef Yengej, F.A.; Jansen, J.; Rookmaaker, M.B.; Verhaar, M.C.; Clevers, H. Kidney Organoids and Tubuloids. Cells 2020, 9. [CrossRef]
- Mitrofanova, O.; Nikolaev, M.; Xu, Q.; Broguiere, N.; Cubela, I.; Camp, J.G.; Bscheider, M.; Lutolf, M.P. Bioengineered Human Colon Organoids with in Vivo-like Cellular Complexity and Function. Cell Stem Cell 2024, 31, 1175-1186.e7. [CrossRef]
- Jelinsky, S.A.; Derksen, M.; Bauman, E.; Verissimo, C.S.; van Dooremalen, W.T.M.; Roos, J.L.; Barón, C.H.; Caballero-Franco, C.; Johnson, B.G.; Rooks, M.G.; et al. Molecular and Functional Characterization of Human Intestinal Organoids and Monolayers for Modeling Epithelial Barrier. Inflamm Bowel Dis 2023, 29, 195–206. [CrossRef]
- O’Mahony, E.T.; Arian, C.M.; Aryeh, K.S.; Wang, K.; Thummel, K.E.; Kelly, E.J. Human Intestinal Enteroids: Nonclinical Applications for Predicting Oral Drug Disposition, Toxicity, and Efficacy. Pharmacol Ther 2025, 273. [CrossRef]
- Li, J.; Ma, J.; Cao, R.; Zhang, Q.; Li, M.; Wang, W.; Wang, Y.; Li, W.; Zhu, Y.; Leng, L. A Skin Organoid-Based Infection Platform Identifies an Inhibitor Specific for HFMD. Nat Commun 2025, 16. [CrossRef]
- Lee, S.; Rim, Y.A.; Kim, J.; Lee, S.H.; Park, H.J.; Kim, H.; Ahn, S.J.; Ju, J.H. Guidelines for Manufacturing and Application of Organoids: Skin. Int J Stem Cells 2024, 17, 182–193. [CrossRef]
- Neal, J.T.; Li, X.; Zhu, J.; Giangarra, V.; Grzeskowiak, C.L.; Ju, J.; Liu, I.H.; Chiou, S.H.; Salahudeen, A.A.; Smith, A.R.; et al. Organoid Modeling of the Tumor Immune Microenvironment. Cell 2018, 175, 1972-1988.e16. [CrossRef]
- Zhou, G.; Lieshout, R.; van Tienderen, G.S.; de Ruiter, V.; van Royen, M.E.; Boor, P.P.C.; Magré, L.; Desai, J.; Köten, K.; Kan, Y.Y.; et al. Modelling Immune Cytotoxicity for Cholangiocarcinoma with Tumour-Derived Organoids and Effector T Cells. Br J Cancer 2022, 127, 649–660. [CrossRef]
- Avnet, S.; Pompo, G. Di; Borciani, G.; Fischetti, T.; Graziani, G.; Baldini, N. Advantages and Limitations of Using Cell Viability Assays for 3D Bioprinted Constructs. Biomed Mater 2024, 19. [CrossRef]
- Cortesi, M.; Giordano, E. Non-Destructive Monitoring of 3D Cell Cultures: New Technologies and Applications. PeerJ 2022, 10, e13338. [CrossRef]
- Sakolish, C.; Moyer, H.L.; Tsai, H.H.D.; Ford, L.C.; Dickey, A.N.; Wright, F.A.; Han, G.; Bajaj, P.; Baltazar, M.T.; Carmichael, P.L.; et al. Analysis of Reproducibility and Robustness of a Renal Proximal Tubule Microphysiological System OrganoPlate 3-Lane 40 for in Vitro Studies of Drug Transport and Toxicity. Toxicol Sci 2023, 196, 52–70. [CrossRef]
- Theobald, J.; Ghanem, A.; Wallisch, P.; Banaeiyan, A.A.; Andrade-Navarro, M.A.; Taškova, K.; Haltmeier, M.; Kurtz, A.; Becker, H.; Reuter, S.; et al. Liver-Kidney-on-Chip To Study Toxicity of Drug Metabolites. ACS Biomater Sci Eng 2018, 4, 78–89. [CrossRef]
- Moerkens, R.; Mooiweer, J.; Ramírez-Sánchez, A.D.; Oelen, R.; Franke, L.; Wijmenga, C.; Barrett, R.J.; Jonkers, I.H.; Withoff, S. An IPSC-Derived Small Intestine-on-Chip with Self-Organizing Epithelial, Mesenchymal, and Neural Cells. Cell Rep 2024, 43. [CrossRef]
- Graf, K.; Murrieta-Coxca, J.M.; Vogt, T.; Besser, S.; Geilen, D.; Kaden, T.; Bothe, A.K.; Morales-Prieto, D.M.; Amiri, B.; Schaller, S.; et al. Digital Twin-Enhanced Three-Organ Microphysiological System for Studying Drug Pharmacokinetics in Pregnant Women. Front Pharmacol 2025, 16. [CrossRef]
- Bouwmeester, M.C.; Bernal, P.N.; Oosterhoff, L.A.; van Wolferen, M.E.; Lehmann, V.; Vermaas, M.; Buchholz, M.B.; Peiffer, Q.C.; Malda, J.; van der Laan, L.J.W.; et al. Bioprinting of Human Liver-Derived Epithelial Organoids for Toxicity Studies. Macromol Biosci 2021, 21. [CrossRef]
- Ye, S.; Marsee, A.; van Tienderen, G.S.; Rezaeimoghaddam, M.; Sheikh, H.; Samsom, R.A.; de Koning, E.J.P.; Fuchs, S.; Verstegen, M.M.A.; van der Laan, L.J.W.; et al. Accelerated Production of Human Epithelial Organoids in a Miniaturized Spinning Bioreactor. Cell Reports Methods 2024, 4. [CrossRef]
- Addario, G.; Moroni, L.; Mota, C. Kidney Fibrosis In Vitro and In Vivo Models: Path Toward Physiologically Relevant Humanized Models. Adv Healthc Mater 2025, 14. [CrossRef]
- Koido, M.; Kawakami, E.; Fukumura, J.; Noguchi, Y.; Ohori, M.; Nio, Y.; Nicoletti, P.; Aithal, G.P.; Daly, A.K.; Watkins, P.B.; et al. Polygenic Architecture Informs Potential Vulnerability to Drug-Induced Liver Injury. Nat Med 2020, 26, 1541–1548. [CrossRef]
- Lauschke, V.M. Toxicogenomics of Drug Induced Liver Injury - from Mechanistic Understanding to Early Prediction. Drug Metab Rev 2021, 53, 245–252. [CrossRef]
- Hermann, N.G.; Ficek, R.A.; Markov, D.A.; McCawley, L.J.; Hutson, M.S. Toxicokinetics for Organ-on-Chip Devices. Lab Chip 2025, 25, 2017–2029. [CrossRef]
- Novak, R.; Ingram, M.; Marquez, S.; Das, D.; Delahanty, A.; Herland, A.; Maoz, B.M.; Jeanty, S.S.F.; Somayaji, M.R.; Burt, M.; et al. Robotic Fluidic Coupling and Interrogation of Multiple Vascularized Organ Chips. Nat Biomed Eng 2020, 4, 407–420. [CrossRef]
- Lin, Z.; Chou, W.C. Machine Learning and Artificial Intelligence in Toxicological Sciences. Toxicol Sci 2022, 189, 7–19. [CrossRef]
- Jia, X.; Wang, T.; Zhu, H. Advancing Computational Toxicology by Interpretable Machine Learning. Environ Sci Technol 2023, 57, 17690–17706. [CrossRef]
- Cronin, M.T.D.; Belfield, S.J.; Briggs, K.A.; Enoch, S.J.; Firman, J.W.; Frericks, M.; Garrard, C.; Maccallum, P.H.; Madden, J.C.; Pastor, M.; et al. Making in Silico Predictive Models for Toxicology FAIR. Regulatory Toxicology and Pharmacology 2023, 140. [CrossRef]
- Gini, G. QSAR Methods. Methods Mol Biol 2022, 2425, 1–26. [CrossRef]
- Heimbach, T.; Chen, Y.; Chen, J.; Dixit, V.; Parrott, N.; Peters, S.A.; Poggesi, I.; Sharma, P.; Snoeys, J.; Shebley, M.; et al. Physiologically-Based Pharmacokinetic Modeling in Renal and Hepatic Impairment Populations: A Pharmaceutical Industry Perspective. Clin Pharmacol Ther 2021, 110, 297–310. [CrossRef]
- Silva, L.L.; Silvola, R.M.; Haas, D.M.; Quinney, S.K. Physiologically Based Pharmacokinetic Modelling in Pregnancy: Model Reproducibility and External Validation. Br J Clin Pharmacol 2022, 88, 1441–1451. [CrossRef]
- Chung, E.; Russo, D.P.; Ciallella, H.L.; Wang, Y.T.; Wu, M.; Aleksunes, L.M.; Zhu, H. Data-Driven Quantitative Structure-Activity Relationship Modeling for Human Carcinogenicity by Chronic Oral Exposure. Environ Sci Technol 2023, 57, 6573–6588. [CrossRef]
- Banerjee, P.; Kemmler, E.; Dunkel, M.; Preissner, R. ProTox 3.0: A Webserver for the Prediction of Toxicity of Chemicals. Nucleic Acids Res 2024, 52, W513–W520. [CrossRef]
- Sedykh, A.Y.; Shah, R.R.; Kleinstreuer, N.C.; Auerbach, S.S.; Gombar, V.K. Saagar-A New, Extensible Set of Molecular Substructures for QSAR/QSPR and Read-Across Predictions. Chem Res Toxicol 2021, 34, 634–640. [CrossRef]
- Mou, Z.; Volarath, P.; Racz, R.; Cross, K.P.; Girireddy, M.; Chakravarti, S.; Stavitskaya, L. Quantitative Structure-Activity Relationship Models to Predict Cardiac Adverse Effects. Chem Res Toxicol 2024, 37, 1924–1933. [CrossRef]
- Price, E.; Kalvass, J.C.; Degoey, D.; Hosmane, B.; Doktor, S.; Desino, K. Global Analysis of Models for Predicting Human Absorption: QSAR, In Vitro, and Preclinical Models. J Med Chem 2021, 64, 9389–9403. [CrossRef]
- Romano, J.D.; Hao, Y.; Moore, J.H.; Penning, T.M. Automating Predictive Toxicology Using ComptoxAI. Chem Res Toxicol 2022, 35, 1370–1382. [CrossRef]
- Lou, S.; Yu, Z.; Huang, Z.; Wang, H.; Pan, F.; Li, W.; Liu, G.; Tang, Y. In Silico Prediction of Chemical Acute Dermal Toxicity Using Explainable Machine Learning Methods. Chem Res Toxicol 2024, 37, 513–524. [CrossRef]
- Chen, Y.; Yu, X.; Li, W.; Tang, Y.; Liu, G. In Silico Prediction of HERG Blockers Using Machine Learning and Deep Learning Approaches. J Appl Toxicol 2023, 43, 1462–1475. [CrossRef]
- Berton, M.; Bettonte, S.; Stader, F.; Battegay, M.; Marzolini, C. Physiologically Based Pharmacokinetic Modelling to Identify Physiological and Drug Parameters Driving Pharmacokinetics in Obese Individuals. Clin Pharmacokinet 2023, 62, 277–295. [CrossRef]
- Chou, W.C.; Chen, Q.; Yuan, L.; Cheng, Y.H.; He, C.; Monteiro-Riviere, N.A.; Riviere, J.E.; Lin, Z. An Artificial Intelligence-Assisted Physiologically-Based Pharmacokinetic Model to Predict Nanoparticle Delivery to Tumors in Mice. Journal of Controlled Release 2023, 361, 53–63. [CrossRef]
- Ozbek, O.; Genc, D.E.; O. Ulgen, K. Advances in Physiologically Based Pharmacokinetic (PBPK) Modeling of Nanomaterials. ACS Pharmacol Transl Sci 2024, 7, 2251–2279. [CrossRef]
- Chen, J.; Zhao, S.; Wesseling, S.; Kramer, N.I.; Rietjens, I.M.C.M.; Bouwmeester, H. Acetylcholinesterase Inhibition in Rats and Humans Following Acute Fenitrothion Exposure Predicted by Physiologically Based Kinetic Modeling-Facilitated Quantitative In Vitro to In Vivo Extrapolation. Environ Sci Technol 2023, 57, 20521–20531. [CrossRef]
- Li, C.; Jiang, L.; Zhang, D.; Qi, Y.; Wang, X.; Jin, Y.; Liu, X.; Lin, Y.; Luo, J.; Xu, L.; et al. Human Health Risk Assessment of 6:2 Cl-PFESA through Quantitative in Vitro to in Vivo Extrapolation by Integrating Cell-Based Assays, an Epigenetic Key Event, and Physiologically Based Pharmacokinetic Modeling. Environ Int 2023, 173. [CrossRef]
- Madden, J.C.; Thompson, C. V. Pharmacokinetic Tools and Applications. Methods Mol Biol 2022, 2425, 57–83. [CrossRef]
- Tess, D.; Chang, G.C.; Keefer, C.; Carlo, A.; Jones, R.; Di, L. In Vitro-In Vivo Extrapolation and Scaling Factors for Clearance of Human and Preclinical Species with Liver Microsomes and Hepatocytes. AAPS J 2023, 25. [CrossRef]
- Casati, S. Integrated Approaches to Testing and Assessment. Basic Clin Pharmacol Toxicol 2018, 123 Suppl 5, 51–55. [CrossRef]
- Sewell, F.; Alexander-White, C.; Brescia, S.; Currie, R.A.; Roberts, R.; Roper, C.; Vickers, C.; Westmoreland, C.; Kimber, I. New Approach Methodologies (NAMs): Identifying and Overcoming Hurdles to Accelerated Adoption. Toxicol Res (Camb) 2024, 13. [CrossRef]
- OECD Guideline No. 497: Defined Approaches on Skin Sensitisation; OECD Guidelines for the Testing of Chemicals, Section 4; OECD Publishing, 2025;
- OECD Test No. 439: In Vitro Skin Irritation: Reconstructed Human Epidermis Test Method; OECD Guidelines for the Testing of Chemicals, Section 4; OECD Publishing: Paris, 2025;
- van der Zalm, A.J.; Barroso, J.; Browne, P.; Casey, W.; Gordon, J.; Henry, T.R.; Kleinstreuer, N.C.; Lowit, A.B.; Perron, M.; Clippinger, A.J. A Framework for Establishing Scientific Confidence in New Approach Methodologies. Arch Toxicol 2022, 96, 2865–2879. [CrossRef]
- Stucki, A.O.; Barton-Maclaren, T.S.; Bhuller, Y.; Henriquez, J.E.; Henry, T.R.; Hirn, C.; Miller-Holt, J.; Nagy, E.G.; Perron, M.M.; Ratzlaff, D.E.; et al. Use of New Approach Methodologies (NAMs) to Meet Regulatory Requirements for the Assessment of Industrial Chemicals and Pesticides for Effects on Human Health. Frontiers in toxicology 2022, 4. [CrossRef]
- Ouedraogo, G.; Alépée, N.; Tan, B.; Roper, C.S. A Call to Action: Advancing New Approach Methodologies (NAMs) in Regulatory Toxicology through a Unified Framework for Validation and Acceptance. Regulatory Toxicology and Pharmacology 2025, 162. [CrossRef]
- Mathisen, G.H.; Bearth, A.; Jones, L.B.; Hoffmann, S.; Vist, G.E.; Ames, H.M.; Husøy, T.; Svendsen, C.; Tsaioun, K.; Ashikaga, T.; et al. Time for CHANGE: System-Level Interventions for Bringing Forward the Date of Effective Use of NAMs in Regulatory Toxicology. Arch Toxicol 2024, 98, 2299–2308. [CrossRef]
- OECD Test No. 431: In Vitro Skin Corrosion: Reconstructed Human Epidermis (RHE) Test Method; OECD Guidelines for the Testing of Chemicals, Section 4; OECD Publishing, 2025;
- Braakhuis, H.M.; Murphy, F.; Ma-Hock, L.; Dekkers, S.; Keller, J.; Oomen, A.G.; Stone, V. An Integrated Approach to Testing and Assessment to Support Grouping and Read-Across of Nanomaterials After Inhalation Exposure. Appl In Vitro Toxicol 2021, 7, 112–128. [CrossRef]
- OECD Test No. 492: Reconstructed Human Cornea-like Epithelium (RhCE) Test Method for Identifying Chemicals Not Requiring Classification and Labelling for Eye Irritation or Serious Eye Damage; OECD Guidelines for the Testing of Chemicals, Section 4; OECD Publishing, 2025;
- Knetzger, N.; Ertych, N.; Burgdorf, T.; Beranek, J.; Oelgeschläger, M.; Wächter, J.; Horchler, A.; Gier, S.; Windbergs, M.; Fayyaz, S.; et al. Non-Invasive in Vitro NAM for the Detection of Reversible and Irreversible Eye Damage after Chemical Exposure for GHS Classification Purposes (ImAi). Arch Toxicol 2025, 99, 1011–1028. [CrossRef]
- Weissinger, H.; Knetzger, N.; Cleve, C.; Lotz, C. Impedance-Based in Vitro Eye Irritation Testing Enables the Categorization of Diluted Chemicals. Sci Rep 2024, 14, 1–9. [CrossRef]
- Tal, T.; Myhre, O.; Fritsche, E.; Rüegg, J.; Craenen, K.; Aiello-Holden, K.; Agrillo, C.; Babin, P.J.; Escher, B.I.; Dirven, H.; et al. New Approach Methods to Assess Developmental and Adult Neurotoxicity for Regulatory Use: A PARC Work Package 5 Project. Frontiers in toxicology 2024, 6. [CrossRef]
- Kreutz, A.; Oyetade, O.B.; Chang, X.; Hsieh, J.H.; Behl, M.; Allen, D.G.; Kleinstreuer, N.C.; Hogberg, H.T. Integrated Approach for Testing and Assessment for Developmental Neurotoxicity (DNT) to Prioritize Aromatic Organophosphorus Flame Retardants. Toxics 2024, 12. [CrossRef]
- FDA Roadmap to Reducing Animal Testing in Preclinical Safety Studies [PDF]; https://www.fda.gov/files/newsroom/published/roadmap_to_reducing_animal_testing_in_preclinical_safety_studies.pdf, 2025;
- European Medicines Agency New Approach Methodologies – EU Horizon Scanning Report.; Amsterdam, The Netherlands, 2025;
- Najjar, A.; Kühnl, J.; Lange, D.; Géniès, C.; Jacques, C.; Fabian, E.; Zifle, A.; Hewitt, N.J.; Schepky, A. Next-Generation Risk Assessment Read-across Case Study: Application of a 10-Step Framework to Derive a Safe Concentration of Daidzein in a Body Lotion. Front Pharmacol 2024, 15. [CrossRef]
- OECD Guidance Document on the Reporting of Defined Approaches to Be Used Within Integrated Approaches to Testing and Assessment; OECD Series on Testing and Assessment, No. 255; OECD, 2017;
- OECD Guiding Principles and Key Elements for Establishing a Weight of Evidence for Chemical Assessment; OECD Series on Testing and Assessment, No. 311; OECD: Paris, 2019;
- Zuang, V.; Baccaro, M.; Barroso, J.; Berggren, E.; Bopp, S.; Bridio, S.; Capeloa, T.; Carpi, D.; Casati, S.; Chinchio, E.; et al. Non-Animal Methods in Science and Regulation. In Proceedings of the Publications Office of the European Union; Publications Office of the European Union: Luxembourg, 2025.
- Zuang, V.; Barroso, J.; Berggren, E.; Bopp, S.; Bridio, S.; Casati, S.; Corvi, R.; Deceuninck, P.; Franco, A.; Gastaldello, A.; et al. Non-Animal Methods in Science and Regulation – EURL ECVAM Status Report 2023; Publications Office of the European Union: Luxembourg, 2024;
- Zuang, V..; Daskalopoulos, E.P..; Worth, A..; Corvi, R..; Munn, S..; Deceuninck, P..; Barroso, J..; Batista Leite, Sofia.; Berggren, Elisabeth.; Bopp, Stephanie.; et al. Non-Animal Methods in Science and Regulation – EURL ECVAM Report 2022; Publications Office of the European Union: Luxembourg, 2023;
- OECD Guidance Document on Integrated Approaches to Testing and Assessment (IATA) for Phototoxicity Testing; OECD Series on Testing and Assessment, No. 397; OECD: Paris, 2024;
- OECD Performance Standards for the Assessment of Proposed Similar or Modified In Vitro Epidermal Sensitisation Assay (EpiSensA) Test Methods; OECD Series on Testing and Assessment, No. 396; OECD: Paris, 2024;
- Tsiros, P.; Minadakis, V.; Li, D.; Sarimveis, H. Parameter Grouping and Co-Estimation in Physiologically Based Kinetic Models Using Genetic Algorithms. Toxicol Sci 2024, 200, 31–46. [CrossRef]
- Langan, L.M.; Paparella, M.; Burden, N.; Constantine, L.; Margiotta-Casaluci, L.; Miller, T.H.; Moe, S.J.; Owen, S.F.; Schaffert, A.; Sikanen, T. Big Question to Developing Solutions: A Decade of Progress in the Development of Aquatic New Approach Methodologies from 2012 to 2022. Environ Toxicol Chem 2024, 43, 559–574. [CrossRef]
- Schultz, M.; Krause, S.; Brinkmann, M. Validation of Methods for in Vitro- in Vivo Extrapolation Using Hepatic Clearance Measurements in Isolated Perfused Fish Livers. Environ Sci Technol 2022, 56, 12416–12423. [CrossRef]

Table 1.
Comparison of hESC-, hiPSC-, and adult stem cell–based models in cytotoxicity testing.
| Feature | hESCs | hiPSCs | Adult stem cells (HSCs, MSCs) |
|---|---|---|---|
| Source | Inner cell mass of human blastocysts (IVF surplus embryos) | Reprogrammed adult somatic cells (fibroblasts, blood, urine) using Yamanaka factors | Bone marrow, peripheral blood (HSCs), adipose or umbilical cord tissue (MSCs) |
| Potency | Pluripotent (all germ layers) | Pluripotent (patient-specific, variable) | Multipotent (restricted to specific tissue lineages) |
| Applications | Developmental toxicity; cardiac, hepatic, neuronal, epithelial, ocular models [96,102] | Cardiotoxicity, hepatotoxicity, developmental neurotoxicity, renal and ocular assays, precision toxicology [108,111,113] |
Immunotoxicity, myelotoxicity, biomaterial and nanomaterial cytotoxicity [139,140,142,144] |
| Advantages | Natural pluripotency; reproducible protocols; validated differentiation | Ethically acceptable; scalable; patient-specific |
Easy access; ethically uncontroversial; tissue-relevant |
| Limitations | Ethical controversy; limited access; teratoma risk | Variability; incomplete maturation; donor heterogeneity | Limited potency; donor variability; senescence |
| Ethical/Legal | Strict oversight (NIH Registry, EU Directive 2004/23/EC, ISSCR) | Informed consent; data protection (ISSCR 2021) | Standard medical consent; minimal restrictions |
Table 2.
Summary of In silico Pillars for Cytotoxicity/Health Endpoints.
| Method | Primary Inputs | Typical Outputs | Strengths | Common Pitfalls | Use Cases | Key Refs |
|---|---|---|---|---|---|---|
| QSAR/ read-across |
Molecular structu-res, curated labels | Class or continuous risk | Fast, interpretable | Limited do-main, data leakage | Early hazard identification | [199,200,203] |
| ML/AI | Structures + omics/phenotypes | Multi-endpoint predictions | Handles non-linear, multi-task data | Interpretability drift | Portfolio triage, prioritisation | [197,198,204] |
| PBPK | Physiology, ADME parametres | Tissue concentra-tion–time (C(t)) | Human- relevance |
Parameter uncertainty | Populations, DDI, exposure assess-ment | [89,201,202] |
| QIVIVE | In vitro ECx + PBPK | Human- equivalent dose | Translational, mechanistic | Mis-specified clearance | Screening-level risk, potency estimation | [87,88] |
Table 3.
Examples of IATA/NAM implementation across regulatory endpoints.
| Endpoint | Primary NAM(s)/DA | OECD TG/ Guidance | Regulatory Scope | Status / Notes |
Key Refs |
|---|---|---|---|---|---|
| Skin sensitisation |
DPRA + KeratinoSens™ + h-CLAT (DA) |
OECD TG 497 (2025) | Classification & labeling | Fully accepted | [220,222] |
| Skin irritation |
Reconstructed epidermis (EpiDerm™, SkinEthic™, epiCS) |
OECD TG 439 (2025) | Classification & labeling | Fully accepted | [218,221] |
| Eye irritation |
Reconstructed corneal epithelium (EpiOcular™, SkinEthic™ HCE) |
OECD TG 492 (2025) | Classification & labeling | Accepted; replaces Draize test | [228,229,230] |
| Phototoxicity | IATA for Phototoxicity | OECD Guidance No. 397 (2024) | Screening / Hazard ID |
Recently introduced | [241] |
| Nanomaterial inhalation |
Grouping / Read-Across Approach |
– | Occupational risk assessment | Emerging application | [227] |
| Developmental toxicity | PluriLum / ReproTracker + PBPK/QIVIVE | – | Developmental & Reproductive | Under validation | [231,232] |
Table 4.
Milestones and emerging directions in cytotoxicity testing.
| Stage / Era | Key Advances | Representative Methods / Systems | Main Impact |
|---|---|---|---|
| Classical (1980s–2000s) |
Colorimetric and metabolic viability assays | MTT, LDH, Neutral Red, Resazurin | Foundation of in vitro toxicology; standardized endpoints; regulatory benchmarks |
| Mechanistic (2000s–2010s) | High-throughput and high-content screening; mechanistic readouts | HCI, Cell Painting, flow cytometry, xCELLigence | Multiparametric mechanistic insight; reduction of false positives/negatives |
| Human-relevant (2010s–2020s) | Stem-cell–based and 3D models | hPSC/hiPSC assays, organoids, organ-on-chip | Human-specific predictive systems; translation to tissue- and organ-level toxicity |
| Computational and Integrative (2020s–present) | AI, PBPK/QIVIVE, NAMs/IATA frameworks | Machine learning, IVIVE, body-on-chip | Mechanistic–quantitative risk assessment; regulatory adoption of non-animal evidence |
| Emerging (Future) |
Personalized, multi-organ, and AI-driven toxicology | Patient-derived hiPSC models, multi-MPS networks, digital twins | Predictive, individualized safety assessment; convergence of toxicology and precision medicine |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



